
Epitope Mapping of Cytochrome P450cam (CYP101
9
ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS Vol. 341, No. 2, May 15, pp. 229–237, 1997 Article No. BB979934 Epitope Mapping of Cytochrome P450cam (CYP101) Ekaterina F. Kolesanova,* ,1 Sergey A. Kozin,² Andrey B. Rumyantsev,* Christiane Jung,‡ Gaston Hui Bon Hoa,² and Alexander I. Archakov* *Institute of Biomedical Chemistry, Russian Academy of Medical Sciences, Moscow, Russia; ²Institut de Biologie Physico-Chimique, INSERM-U310, Paris, France; and ‡Max-Delbruck-Centrum fur Molekulare Medizin, Berlin-Buch, Federal Republic of Germany Received December 5, 1996 terminations have led to the classification and creation Eighteen linear antigenically active sites were re- of a systematic nomenclature of cytochromes P450 (1, vealed in cytochrome P450 from Pseudomonas putida 2, 5). Despite rather great sequence variability between (P450cam) by hexapeptide scanning. These sites oc- different members of the P450 2 superfamily, their sec- cupy about 31% of the protein sequence. Hexapeptide ondary and tertiary structures are assumed to be quite epitope sequences of P450cam are not found in other similar. This assumption is based on their common cat- cytochromes P450. However, several cytochromes alytic mechanism, existence of several conservative se- P450 contain shorter fragments of P450cam epitope quence regions, and results of secondary structure pre- sequences which may cause weak immune cross-reac- dictions (5 – 11). The immunochemical studies with the tions. P450cam antigenic determinants are located help of monoclonal and polyclonal antibodies to P450s generally at the boundaries of secondary structure ele- provide an important approach to the investigation of ments. Mapping of P450cam antigenic determinants on both different and common features of these enzymes the three-dimensional structure of this protein reveals including structural relatedness and peculiarities of 14 highly water-accessible antigenic sites and only 1 the common catalytic mechanism (7, 12– 17) and also site (No. 322 – 327, QMLSGL) which is inaccessible to provide an opportunity for the development of specific water. Several functionally important sites and amino diagnostic tests (15, 18–20). However, despite the nu- acid residues of P450cam are localized within revealed merous immunochemical studies there is little knowl- linear epitopes or very close to them. These sites in- edge about antigenic determinants of cytochromes clude substrate-binding regions, residues responsible P450 (7, 13, 14, 20, 21). Antigenic mapping of different for the putidaredoxin interaction (Arg72, Arg112, P450s would undoubtedly provide an important tool for Lys314, and Arg364), heme binding (Gln108, Arg112, their structural comparison including sequence homol- Asp297, Arg299, and Cys357), and proton translocation ogy, spatial structure, and orientation of enzyme mole- (Lys178, Arg186, and Glu366). q 1997 Academic Press cules with respect to cellular structures. At present, Key Words: cytochrome P450; epitope mapping; pep- tide scanning. P450 2B4 is the only cytochrome P450 whose antigenic map has been revealed (21) by PEPSCAN approach (22, 23). In addition, cytochrome P4502D6 epitopes which induce the formation of autoantibodies have been Cytochromes P450 comprise the large superfamily of mapped by the expression of cDNA constructs (20). enzymes (1, 2) which play a crucial role in the oxidative However, since the 2B4 and 2D6 secondary and ter- metabolism of physiological substrates and xenobiotics tiary structures are unknown and functional sites are (3, 4). These enzymes exist in various living organisms not precisely determined, antigenic maps of these ranging from bacteria and yeast to plants and higher animals. Analyses of amino acid sequences derived 2 Abbreviations used: ABTS, 2,2 *-azino-bis(3-ethylbenzthiazoline- from gene sequences and direct primary structure de- 6-sulfonic acid); BSA, bovine serum albumin; CPD, cytochrome P450 database; ELISA, enzyme-linked immunosorbent assay; Fmoc, 9- fluorenylmethyloxycarbonyl; OPhF5 , pentafluorophenyl ester; P450, 1 To whom correspondence should be addressed at Institute of Bio- cytochrome P450; P450cam, cytochrome P450 from Pseudomonas putida (CYP101); NaCl/Pi , phosphate-buffered saline; PEPSCAN, medical Chemistry, Russian Academy of Medical Sciences, Pogodin- skaya ul., 10, 119832, Moscow, Russia. Fax: 7(095)245-08-57. E-mail: peptide scanning. One-letter symbols are used for amino acid resi- dues. Enzyme:cytochrome P450 (EC 1.14.14.1). [email protected]. 229 0003-9861/97 $25.00 Copyright q 1997 by Academic Press All rights of reproduction in any form reserved.
Transcript of Epitope Mapping of Cytochrome P450cam (CYP101

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS
Vol. 341, No. 2, May 15, pp. 229–237, 1997Article No. BB979934
Epitope Mapping of Cytochrome P450cam (CYP101)
Ekaterina F. Kolesanova,*,1 Sergey A. Kozin,† Andrey B. Rumyantsev,* Christiane Jung,‡Gaston Hui Bon Hoa,† and Alexander I. Archakov**Institute of Biomedical Chemistry, Russian Academy of Medical Sciences, Moscow, Russia; †Institut de BiologiePhysico-Chimique, INSERM-U310, Paris, France; and ‡Max-Delbruck-Centrum fur Molekulare Medizin,Berlin-Buch, Federal Republic of Germany
Received December 5, 1996
terminations have led to the classification and creationEighteen linear antigenically active sites were re- of a systematic nomenclature of cytochromes P450 (1,
vealed in cytochrome P450 from Pseudomonas putida 2, 5). Despite rather great sequence variability between(P450cam) by hexapeptide scanning. These sites oc- different members of the P4502 superfamily, their sec-cupy about 31% of the protein sequence. Hexapeptide ondary and tertiary structures are assumed to be quiteepitope sequences of P450cam are not found in other similar. This assumption is based on their common cat-cytochromes P450. However, several cytochromes alytic mechanism, existence of several conservative se-P450 contain shorter fragments of P450cam epitope quence regions, and results of secondary structure pre-sequences which may cause weak immune cross-reac- dictions (5–11). The immunochemical studies with thetions. P450cam antigenic determinants are located help of monoclonal and polyclonal antibodies to P450sgenerally at the boundaries of secondary structure ele- provide an important approach to the investigation ofments. Mapping of P450cam antigenic determinants on both different and common features of these enzymesthe three-dimensional structure of this protein reveals including structural relatedness and peculiarities of14 highly water-accessible antigenic sites and only 1 the common catalytic mechanism (7, 12– 17) and alsosite (No. 322–327, QMLSGL) which is inaccessible to provide an opportunity for the development of specificwater. Several functionally important sites and amino
diagnostic tests (15, 18–20). However, despite the nu-acid residues of P450cam are localized within revealedmerous immunochemical studies there is little knowl-linear epitopes or very close to them. These sites in-edge about antigenic determinants of cytochromesclude substrate-binding regions, residues responsibleP450 (7, 13, 14, 20, 21). Antigenic mapping of differentfor the putidaredoxin interaction (Arg72, Arg112,P450s would undoubtedly provide an important tool forLys314, and Arg364), heme binding (Gln108, Arg112,their structural comparison including sequence homol-Asp297, Arg299, and Cys357), and proton translocationogy, spatial structure, and orientation of enzyme mole-(Lys178, Arg186, and Glu366). q 1997 Academic Press
cules with respect to cellular structures. At present,Key Words: cytochrome P450; epitope mapping; pep-tide scanning. P450 2B4 is the only cytochrome P450 whose antigenic
map has been revealed (21) by PEPSCAN approach (22,23). In addition, cytochrome P4502D6 epitopes whichinduce the formation of autoantibodies have been
Cytochromes P450 comprise the large superfamily of mapped by the expression of cDNA constructs (20).enzymes (1, 2) which play a crucial role in the oxidative However, since the 2B4 and 2D6 secondary and ter-metabolism of physiological substrates and xenobiotics tiary structures are unknown and functional sites are(3, 4). These enzymes exist in various living organisms not precisely determined, antigenic maps of theseranging from bacteria and yeast to plants and higheranimals. Analyses of amino acid sequences derived 2 Abbreviations used: ABTS, 2,2 *-azino-bis(3-ethylbenzthiazoline-from gene sequences and direct primary structure de- 6-sulfonic acid); BSA, bovine serum albumin; CPD, cytochrome P450
database; ELISA, enzyme-linked immunosorbent assay; Fmoc, 9-fluorenylmethyloxycarbonyl; OPhF5, pentafluorophenyl ester; P450,
1 To whom correspondence should be addressed at Institute of Bio- cytochrome P450; P450cam, cytochrome P450 from Pseudomonasputida (CYP101); NaCl/Pi , phosphate-buffered saline; PEPSCAN,medical Chemistry, Russian Academy of Medical Sciences, Pogodin-
skaya ul., 10, 119832, Moscow, Russia. Fax: 7(095)245-08-57. E-mail: peptide scanning. One-letter symbols are used for amino acid resi-dues. Enzyme:cytochrome P450 (EC 1.14.14.1)[email protected].
2290003-9861/97 $25.00Copyright q 1997 by Academic PressAll rights of reproduction in any form reserved.
AID ABB 9934 / 6b32$$$341 04-14-97 09:45:59 arca

230 KOLESANOVA ET AL.
0.05% sodium azide) for 2 h at 377C. After washing as above, reagentP450s cannot be compared with their structural–func-antibodies [horseradish peroxidase-labeled anti(rabbit IgG)-immu-tional maps.noglobulins] diluted 1:1000 in the supercocktail without sodiumCytochrome P450 from Pseudomonas putida azide were added (200 ml per well) and allowed to stand for 2 h
(CYP101 or P450cam) is one of four members of the at 377C. Following the same washings with water and NaCl/Pi theadsorbed peroxidase label (i.e., the presence of antibodies toP450 superfamily whose three-dimensional structuresP450cam) was determined with 0.03% H2O2 and 0.5 mM ABTS (ahave been determined (24–27). Many functionally im-color-developing substrate) in 0.08 M sodium citrate buffer, pH 4.0portant residues of P450cam have been localized by (200 ml of freshly prepared substrate solution per well), and ab-
site-directed mutagenesis experiments (28). This pro- sorbance at 405 nm was read in a Multiscan Plus plate reader (Lab-tein and three other microbial cytochromes P450 with systems, Finland) after 20 min incubation at room temperature.
Synthesis of pin-attached peptides. Peptides attached to polyeth-known spatial structure can represent convenient mod-ylene pins (Cambridge Research Biochemicals Co.) were synthesizedels for investigating cytochrome P450 catalytic mecha-by the active esters coupling procedure using Fmoc–amino acid–nisms and protein–protein interactions in monooxy-OPhF5 esters in the presence of an equimolar amount of 1-hydroxy-genase complexes; they also may serve as frameworks benzotriazole according to the manufacturer’s instructions (Cam-
for eukaryotic P450 modeling (10, 25). Hence, antigenic bridge Research Biochemicals Co.) (21, 23, 30). Briefly, following thedeprotection of the anchoring Fmoc-protected amino group with 20%mapping of P450cam should not only provide the infor-piperidine in N,N-dimethylformamide for 30 min at 227C, the cou-mation about the structure of its linear antigenic deter-pling step was allowed to proceed for 18 h at 227C with 30 mM solu-minants, but also estimate the relationships of anti-tions of Fmoc–amino acid–OPhF5 esters and 1-hydroxybenzotriazole
genic determinants to various structural elements and in N,N-dimethylformamide. An additional 30-min washing with 15functionally important sites of this enzyme. mM 1-hydroxybenzotriazole in N,N-dimethylformamide for 30 min
at 227C was introduced prior to the coupling step to remove tracesof piperidine and, thus, to increase coupling efficiency. At the comple-
MATERIALS AND METHODS tion of the sixth coupling step, the pin-coupled peptides were finallydeprotected on the N-terminal a-amino group and then acetylatedMaterials. Pentafluorophenyl (OPhF5) esters of a-N-Fmoc–by acetic anhydride (dimethylformamide:acetic anhydride:triethyla-amino acids and blocks of derivatized polyethylene pins for solid-mine, 5:2:1 v/v/v) for 90 min at 227C to remove the N-terminal posi-phase synthesis of noncleavable polypeptides were from Cambridgetive charge. The peptide No. 1, which corresponded to the N-terminalResearch Biochemicals (Northwich, UK). 2,2 *-azino-bis(3-ethylbenz-hexapeptide of P450cam, was not acetylated. The side-chain depro-thiazoline-6-sulfonic acid) (ABTS), BSA, Freund’s complete adjuvant,tection was carried out by 4-h treatment of pins with a mixture ofand 1-hydroxybenzotriazole were from Calbiochem (Lucerne, Swit-trifluoroacetic acid:phenol:ethanedithiol (95:2.5:2.5 v/w/v) at roomzerland). N-Ethyl-diisopropylamine, 1,2-ethanedithiol, 2-mercapto-temperature followed by the neutralization with 5% N-ethyl-diiso-ethanol, phenol, and Tween 20 were from Merck (Darmstadt, FRG).propylamine in dichloromethane for 5 min. The P450cam-derivedGoat anti(rabbit IgG)-immunoglobulins labeled with horseradishhexapeptide synthesis schedule was created on an IBM-PC-AT 486peroxidase and sodium azide were from Dia-M (Moscow, Russia).according to the General Net (GNET) Approach of the CambridgeTrifluoroacetic acid, analytic grade, was from the Perm Division ofResearch Biochemicals Peptide Synthesis software. P450cam se-the State Institute of Applied Chemistry (Perm, Russia). Other re-quence (31) was taken from the cytochrome P450 database (CPD)agents were from Reachim (Moscow, Russia).created at the Institute of Biomedical Chemistry, Russian AcademyPolypropylene plates for the synthesis of pin-linked peptides, poly-of Medical Sciences (Moscow, Russia) (3).3 A hexapeptide framechlorovinyl EIA/RIA plates, and polystyrene Serocluster plates weremoved along the protein sequence with a step of one residue; hence,from Costar (Badhoevedorp, The Netherlands).the first hexapeptide to be synthesized represented the N-terminalCytochrome P450cam from Ps. putida was expressed in Esche-hexapeptide of P450cam, the second one contained amino acid resi-richia coli, strain TB1, and was isolated and purified to homogeneitydues from 2 to 7, and so on. Thus, each previous hexapeptide over-as described in (29).lapped five residues of the next one.Antiserum preparation. Antisera to the purified cytochrome
Six peptides (56–61, TRCNGG; 88–93, IPREAG; 141–146, SLR-P450cam from Ps. putida were prepared in two male Shinshilla rab-PQG; 187–192, PDGSMT; 305–310, YEFHGV; and 376–381bits. Animals received four injections of 200 mg purified P450cam: atRIPDFS) were synthesized in duplicate, three of the duplicate sam-Days 0 and 14, in 0.5 ml of phosphate-buffered saline (NaCl/Pi : 145ples (TRCNGG, IPREAG, and SLRPQG) were hydrolyzed with 5.7mM NaCl, 10 mM NaPi , pH 7.2) diluted 1:1 in complete Freund’sM HCl just after the completion of the synthesis procedure, and theiradjuvant, subcutaneously along the back; at Days 28 and 42, in 1amino acid analyses were made to check the amino acid compositionml of NaCl/Pi , intramuscularly. Rabbits were bled through an earand determine the amount of pin-attached peptides. In addition, twovein 7 days after the last immunization. Blood was left for coagula-control tetrapeptides, PLAQ and GLAQ, were synthesized on eachtion for 2 h at room temperature and then centrifuged 15 min at96-pin plate, as recommended by the Cambridge Research Biochemi-3000g. Sera were collected, combined in a 1:1 ratio after titer deter-cals Co., and analyzed for their interaction with monospecific anti-mination, and stored at 0207C. Control (preimmune) sera were re-peptide antibodies against PLAQ, as described below.ceived from the same animals 1 week before the immunizations.
To check the peptide integrity after repeated ELISA and washings,Antiserum testing by ELISA. Antisera to the cytochrome three peptides (187–192, PDGSMT; 305–310 YEFHGV; and 376–
P450cam were tested for the interaction with native P450cam and 381, RIPDFS) were subjected to hydrolysis with 5.7 M HCl and totheir titers were determined by ELISA (21). Briefly, 8 mg of cyto- further amino acid analysis after the completion of all 30 ELISAchrome P450cam in 200 ml of 10 mM sodium carbonate buffer, pH tests.9.4, was adsorbed on the surface of flat-bottom wells of polychlorovi- ELISA of pin-attached peptides. The interaction of the P450cam-nyl EIA/RIA plates during 18 h at 47C. After six washings with water derived pin-attached hexapeptides with the P450cam antiserum wasand six washings with NaCl/Pi/0.05% Tween 20, the adsorbedP450cam was allowed to interact with 200 ml of antisera or controlsera diluted 1:500, 1:1000, 1:2000, 1:4000, or 1:8000 in the blocking 3 To get more information about CPD please contact Academician
Professor A. I. Archakov.buffer (‘‘supercocktail’’) (NaCl/Pi with 0.1% Tween 20, 2% BSA, and
AID ABB 9934 / 6b32$$$341 04-14-97 09:45:59 arca

231EPITOPE MAPPING OF CYTOCHROME P450cam
determined by ELISA according to the manufacturer’s instructions bility or surface exposure was defined as the ability for physicalcontact between a sphere with van der Waals radius of a testing(21, 22, 30). Antisera from two rabbits were combined in a 1:1 ratio,
as were the control sera. Prior to the incubation with the antiserum atom and the water-sized probe sphere (r Å 0.14 nm) (22, 33). Thesurface of each testing atom (represented as a sphere) in the molecule(or control serum) the pins were agitated in the supercocktail (200
ml/pin) for 1 h at room temperature to block nonspecific binding. Then was divided into 642 equal segments and the number of segmentswhich were accessible to contact with the water-sized sphere wasthe pins were incubated overnight at 47C in the antiserum (175 ml/
pin) diluted 1:4000, 1:2000, 1:1000, 1:700, or 1:400 in the supercock- counted. Each atom or group of atoms was considered water-accessi-ble or exposed on the surface if more than 10% of its surface wastail or in the control serum diluted in the same way. After four 10-
min washings at room temperature with NaCl/Pi/0.05% Tween 20 the accessible to the water-sized probe sphere. Cytochrome P450camatom coordinates and the information about secondary structurespins were agitated with reagent antibodies [horseradish peroxidase-
labeled goat anti(rabbit IgG)-immunoglobulins] diluted 1:1000 in the were extracted from the Brookhaven Data Bank (34).A computer search for P450cam linear epitope sequences in varioussupercocktail without sodium azide (175 ml/pin) for 1 h at 257C. The
pins were then washed as described and the presence of antibodies members of the cytochrome P450 superfamily was performed withthe help of CPD.adsorbed to pin-linked peptides was detected by the reaction with
150 ml (per pin) of freshly prepared substrate solution (0.5 mM ABTS,0.03% H2O2, 0.1 M NaPi , and 0.08 M citric acid, pH 4.0). After 40 RESULTS AND DISCUSSIONmin incubation at room temperature the absorbance at 405 nm wasread in the Multiscan Plus plate reader. The antibody binding capaci- Linear peptide B-epitopes or antigenic determinantsties of peptides are designated in terms of A405 values. All ELISA are considered to contain three to six amino acid resi-experiments were made in triplicate. Testing of pin-linked peptides dues (22, 33, 35). According to this consideration a hex-with the antiserum was always followed in succession by their testing
apeptide frame moving along the molecule of the cyto-with the control serum, in order to ensure the removal of antibodiesfrom pins by washing procedures. chrome P450cam with a step of one amino acid residue
Testing of control tetrapeptides PLAQ and GLAQ (see above) with is sufficient for scanning this enzyme molecule in ordera monoclonal antibody against PLAQ was performed as described to reveal its linear epitopes with the help of pin-linkedabove. The reconstituted solution of PLAQ-derived monoclonal anti- peptides. The acetyl blocking group is too small in size,body (antibody preparation is supplied together with the Epitope
and the N-acryloyl-N*-b-alanyl-hexamethylenedia-Scanning Kit from Cambridge Research Biochemicals) was used in-stead of P450cam antiserum, and goat anti(mouse IgG)-immunoglob- mide linker group is too flexible and devoid of side-ulins labeledwith horseradish peroxidase (Calbiochem; working dilu- chain groups to interfere with hexapeptide binding totion 1:10,000) were applied as reagent antibodies. Control peptides antibodies. The noncleavable PEPSCAN procedure waswere tested in triplicate two times: at the first and at the last ELISA
chosen because of the ability of pin-linked peptides toexperiments.be used repeatedly in the ELISA tests. Composition ofAfter each ELISA procedure and after the completion of the syn-
thesis the pins were washed with the disruption buffer (0.1 M NaPi , three chosen P450cam hexapeptides (see Materials andpH 7.1, 1% SDS, 0.1% 2-mercaptoethanol) in a sonication bath at Methods) was checked by amino acid analysis just after607C for 30 min, twice with hot distilled water (55–607C), then with the synthesis procedure and proved to coincide withboiling methanol and finally air dried.
the given composition of these peptides. The amountsThe results obtained were plotted and processed with the helpof the peptides attached to the pins were about 45–50of SigmaPlot software on an IBM-PC-AT 486. The limit for cut-off
statistically nonsignificant values of antibody binding to the peptides nmol; contaminations did not excede 5%, according towas determined by the analysis of the A405 data distribution curve. the amino acid analysis data. Composition of the otherThe A405 data distribution curve consisted of a rather sharp as- three chosen P450cam hexapeptides was checked bycending part followed by a smooth descending part. Baseline A405 amino acid analysis after 30 cycles of ELISA. Loss ofvalues which form the ascending part and a portion of the descendingpart of the curve were considered to be normally distributed. Ac- less than 10% of peptide material from pins was de-cording to this assumption the ascending part of the data distribution tected; this result coincides with our previous data ob-curve was used to calculate the normal distribution parameters M tained with cytochrome P450 2B4- and 1A2-derivedand s with the help of SigmaPlot software (curve-fitting function). hexapeptides (36). ELISA tests of control tetrapeptidesThe limit value of (M / 4s) was applied for cutting off statistically
PLAQ and GLAQ with PLAQ-specific monoclonal anti-nonsignificant values of antibody binding. Such a limit correspondsto a 0.01% probability of dropping the baseline values into the group body were positive for PLAQ and negative for GLAQ,of statistically significant values of antibody binding. The described thus pointing to the accuracy of PLAQ synthesis. Thesedata processing procedure was applied to each ELISA experiment tests were also used as controls to assess color develop-because of considerable variations of A405 data levels (but not of the
ment.ratios of A405 values between the antigenic and nonantigenic pep-Prepared antisera samples from two rabbits had thetides) in the parallel experiments. The peptides were considered anti-
genically active if they showed statistically significant values of anti- same titer of 1:4000 with respect to the native P450cam;body binding in all parallel ELISA tests. they were combined in a 1:1 ratio for further experi-
Spatial localization of the linear B-epitopes in the P450cam mole- ments with the pin-linked peptides (this combined anti-cule and their water accessibility were examined with the help ofserum preparation is designated further as antiserum).ONIX software at the Institute of Biomedical Chemistry, RussianCombined antiserum was retested by ELISA four timesAcademy of Medical Sciences (Moscow, Russia) (32).4 Water accessi-in the period when ELISA of pin-linked peptides wasperformed; the titer value of 1:4000 was retained up tothe last experiment with the P450cam-derived hexapep-4 To get more information about ONIX software please contact
Professor A. S. Ivanov. tides. These repeated analyses were combined with
AID ABB 9934 / 6b32$$$342 04-14-97 09:45:59 arca
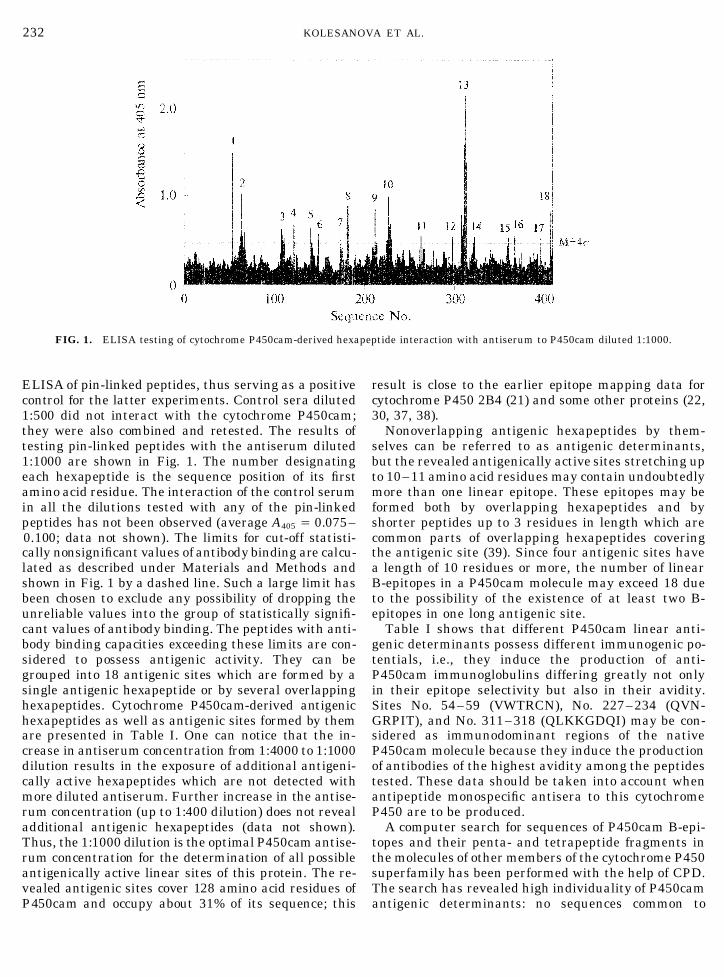
232 KOLESANOVA ET AL.
FIG. 1. ELISA testing of cytochrome P450cam-derived hexapeptide interaction with antiserum to P450cam diluted 1:1000.
ELISA of pin-linked peptides, thus serving as a positive result is close to the earlier epitope mapping data forcytochrome P450 2B4 (21) and some other proteins (22,control for the latter experiments. Control sera diluted
1:500 did not interact with the cytochrome P450cam; 30, 37, 38).Nonoverlapping antigenic hexapeptides by them-they were also combined and retested. The results of
testing pin-linked peptides with the antiserum diluted selves can be referred to as antigenic determinants,but the revealed antigenically active sites stretching up1:1000 are shown in Fig. 1. The number designating
each hexapeptide is the sequence position of its first to 10–11 amino acid residues may contain undoubtedlymore than one linear epitope. These epitopes may beamino acid residue. The interaction of the control serum
in all the dilutions tested with any of the pin-linked formed both by overlapping hexapeptides and byshorter peptides up to 3 residues in length which arepeptides has not been observed (average A405 Å 0.075–
0.100; data not shown). The limits for cut-off statisti- common parts of overlapping hexapeptides coveringthe antigenic site (39). Since four antigenic sites havecally nonsignificant values of antibody binding are calcu-
lated as described under Materials and Methods and a length of 10 residues or more, the number of linearB-epitopes in a P450cam molecule may exceed 18 dueshown in Fig. 1 by a dashed line. Such a large limit has
been chosen to exclude any possibility of dropping the to the possibility of the existence of at least two B-epitopes in one long antigenic site.unreliable values into the group of statistically signifi-
cant values of antibody binding. The peptides with anti- Table I shows that different P450cam linear anti-genic determinants possess different immunogenic po-body binding capacities exceeding these limits are con-
sidered to possess antigenic activity. They can be tentials, i.e., they induce the production of anti-P450cam immunoglobulins differing greatly not onlygrouped into 18 antigenic sites which are formed by a
single antigenic hexapeptide or by several overlapping in their epitope selectivity but also in their avidity.Sites No. 54–59 (VWTRCN), No. 227–234 (QVN-hexapeptides. Cytochrome P450cam-derived antigenic
hexapeptides as well as antigenic sites formed by them GRPIT), and No. 311–318 (QLKKGDQI) may be con-sidered as immunodominant regions of the nativeare presented in Table I. One can notice that the in-
crease in antiserum concentration from 1:4000 to 1:1000 P450cam molecule because they induce the productionof antibodies of the highest avidity among the peptidesdilution results in the exposure of additional antigeni-
cally active hexapeptides which are not detected with tested. These data should be taken into account whenantipeptide monospecific antisera to this cytochromemore diluted antiserum. Further increase in the antise-
rum concentration (up to 1:400 dilution) does not reveal P450 are to be produced.A computer search for sequences of P450cam B-epi-additional antigenic hexapeptides (data not shown).
Thus, the 1:1000 dilution is the optimal P450cam antise- topes and their penta- and tetrapeptide fragments inthe molecules of other members of the cytochrome P450rum concentration for the determination of all possible
antigenically active linear sites of this protein. The re- superfamily has been performed with the help of CPD.The search has revealed high individuality of P450camvealed antigenic sites cover 128 amino acid residues of
P450cam and occupy about 31% of its sequence; this antigenic determinants: no sequences common to
AID ABB 9934 / 6b32$$$343 04-14-97 09:45:59 arca
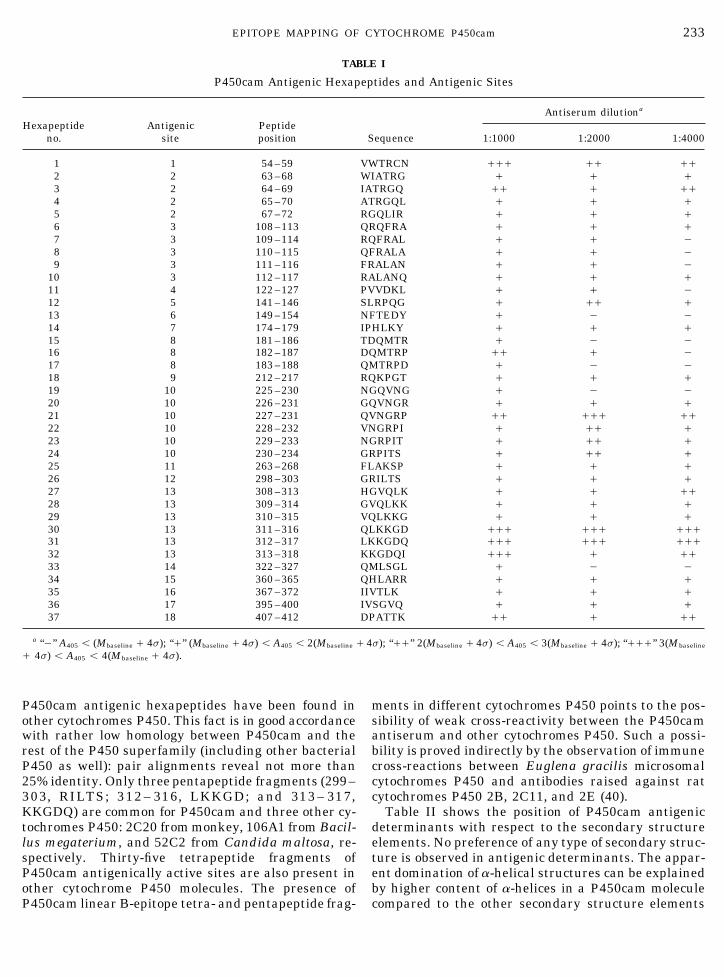
233EPITOPE MAPPING OF CYTOCHROME P450cam
TABLE I
P450cam Antigenic Hexapeptides and Antigenic Sites
Antiserum dilutiona
Hexapeptide Antigenic Peptideno. site position Sequence 1:1000 1:2000 1:4000
1 1 54 –59 VWTRCN /// // //2 2 63 –68 WIATRG / / /3 2 64 –69 IATRGQ // / //4 2 65 –70 ATRGQL / / /5 2 67 –72 RGQLIR / / /6 3 108 –113 QRQFRA / / /7 3 109 –114 RQFRAL / / 08 3 110 –115 QFRALA / / 09 3 111 –116 FRALAN / / 0
10 3 112 –117 RALANQ / / /11 4 122 –127 PVVDKL / / 012 5 141 –146 SLRPQG / // /13 6 149 –154 NFTEDY / 0 014 7 174 –179 IPHLKY / / /15 8 181 –186 TDQMTR / 0 016 8 182 –187 DQMTRP // / 017 8 183 –188 QMTRPD / 0 018 9 212 –217 RQKPGT / / /19 10 225 –230 NGQVNG / 0 020 10 226 –231 GQVNGR / / /21 10 227 –231 QVNGRP // /// //22 10 228 –232 VNGRPI / // /23 10 229 –233 NGRPIT / // /24 10 230 –234 GRPITS / // /25 11 263 –268 FLAKSP / / /26 12 298 –303 GRILTS / / /27 13 308 –313 HGVQLK / / //28 13 309 –314 GVQLKK / / /29 13 310 –315 VQLKKG / / /30 13 311 –316 QLKKGD /// /// ///31 13 312 –317 LKKGDQ /// /// ///32 13 313 –318 KKGDQI /// / //33 14 322 –327 QMLSGL / 0 034 15 360 –365 QHLARR / / /35 16 367 –372 IIVTLK / / /36 17 395 –400 IVSGVQ / / /37 18 407 –412 DPATTK // / //
a ‘‘0’’ A405 õ (Mbaseline / 4s); ‘‘/’’ (Mbaseline / 4s) õ A405 õ 2(Mbaseline / 4s); ‘‘//’’ 2(Mbaseline / 4s) õ A405 õ 3(Mbaseline / 4s); ‘‘///’’ 3(Mbaseline
/ 4s) õ A405 õ 4(Mbaseline / 4s).
P450cam antigenic hexapeptides have been found in ments in different cytochromes P450 points to the pos-sibility of weak cross-reactivity between the P450camother cytochromes P450. This fact is in good accordance
with rather low homology between P450cam and the antiserum and other cytochromes P450. Such a possi-bility is proved indirectly by the observation of immunerest of the P450 superfamily (including other bacterial
P450 as well): pair alignments reveal not more than cross-reactions between Euglena gracilis microsomalcytochromes P450 and antibodies raised against rat25% identity. Only three pentapeptide fragments (299–
303, RILTS; 312 – 316, LKKGD; and 313 – 317, cytochromes P450 2B, 2C11, and 2E (40).Table II shows the position of P450cam antigenicKKGDQ) are common for P450cam and three other cy-
tochromes P450: 2C20 from monkey, 106A1 from Bacil- determinants with respect to the secondary structureelements. No preference of any type of secondary struc-lus megaterium, and 52C2 from Candida maltosa, re-
spectively. Thirty-five tetrapeptide fragments of ture is observed in antigenic determinants. The appar-ent domination of a-helical structures can be explainedP450cam antigenically active sites are also present in
other cytochrome P450 molecules. The presence of by higher content of a-helices in a P450cam moleculecompared to the other secondary structure elementsP450cam linear B-epitope tetra- and pentapeptide frag-
AID ABB 9934 / 6b32$$$343 04-14-97 09:45:59 arca

234 KOLESANOVA ET AL.
TABLE II
Relationship between Epitope Sites and Secondary Structure Elements in P450cama
Position andNo. length Sequence Participation in secondary structure elements
1 54–59 (6) VWTRCN Part of antiparallel b b1 (52–66); CN, part of hairpin turn (58–61)2 63–72 (10) WIATRGQLIR WIAT, part of antiparallel b b1 (52–66); RGQLIR, part of a-helix B (67–77)3 108–117 (10) QRQFRALANQ Part of a-helix C (106–126); Q108, part of hairpin turn (104–108)4 122–127 (6) PVVDKL PVVDK, part of a-helix C; L, part of a-helix D (127–145)5 141–146 (6) SLRPQG SLRPQ, part of a-helix D; G, part of antiparallel b b5 (146–150, 382–405)6 149–154 (6) NFTEDY Beginning of a-helix E (149–169)7 174–179 (6) IPHLKY Part of a-helix F (173–185)8 181–188 (8) TDQMTRPD TDQMT, part of a-helix F; RPD, part of irregular surface loop (186–192)9 212–217 (6) RQKPGT RQK, part of a-helix G (192–214); PGT, part of a loop (215–218) connecting helices
G and H stabilized by D218 and R211 ion pair10 225–235 (11) NGQVNGRPITS N, the end of a-helix H (218–225); GQVNGRPI, antiparallel b b2 (226–233)
including hairpin turn VNGR (228–231)11 263–268 (6) FLAKSP Part of a-helix I (234–267) and the beginning of a-helix J (267–276)12 298–303 (6) GRILTS Antiparallel b b3 (295–301) and coil (302–304)13 308–318 (11) HGVQLKKGDQI HGVQL, part of antiparallel b b4 (305–312), with a part of hairpin turn (307–310);
KK, coil; GDQI, part of antiparallel b b3 (295–301, 315–323)14 322–327 (6) QMLSGL QM, part of antiparallel b b3; LSGL, part of 3/10 helix (324–328)15 360–365 (6) QHLARR Part of a-helix L (359–378)16 367–372 (6) IIVTLK Part of a-helix L17 395–400 (6) IVSGVQ Part of antiparallel b b5 (382–405)18 407–413 (6) DPATTK Coil
a Designation and localization of P450cam secondary structure elements are taken from (10, 24).
(10, 24). Nevertheless, a common feature of almost all (Nos. 4, 5, and 9) are among the most water-accessibleregions (water accessibility of more than 50%), whileP450cam antigenic determinants does exist. One can
easily notice that linear B-epitopes are located at the the least immunogenic site, No. 14, is fully water inac-cessible. It should be noted that this is common forboundaries of the secondary structure elements, in
loops (such as surface loop 186–192) and in the termi- the other antigenically mapped proteins with knowntertiary structures including myohemerythrine (22)nal regions with nonregular secondary structure (coiled
C-terminus). The same observation has been made and dihydrofolate reductase (38). While the majority oflinear antigenic determinants is preferably located onwith the other protein, dihydrofolate reductase (38).
Such a location of protein antigenic sites with respect the surface of the molecules, one or two B-epitopes ofeach protein are fully water inaccessible or containto the secondary structure elements may represent the
common feature of linear B-epitopes especially taking more than 50% water-inaccessible amino acid residues.Despite water inaccessibility, these sites are slightlyinto account the rather high segment mobility of these
regions. hydrophilic and may appear on the surface after a par-tial denaturation or hydrolysis of the native protein.The visualization of B-epitopes on the computer
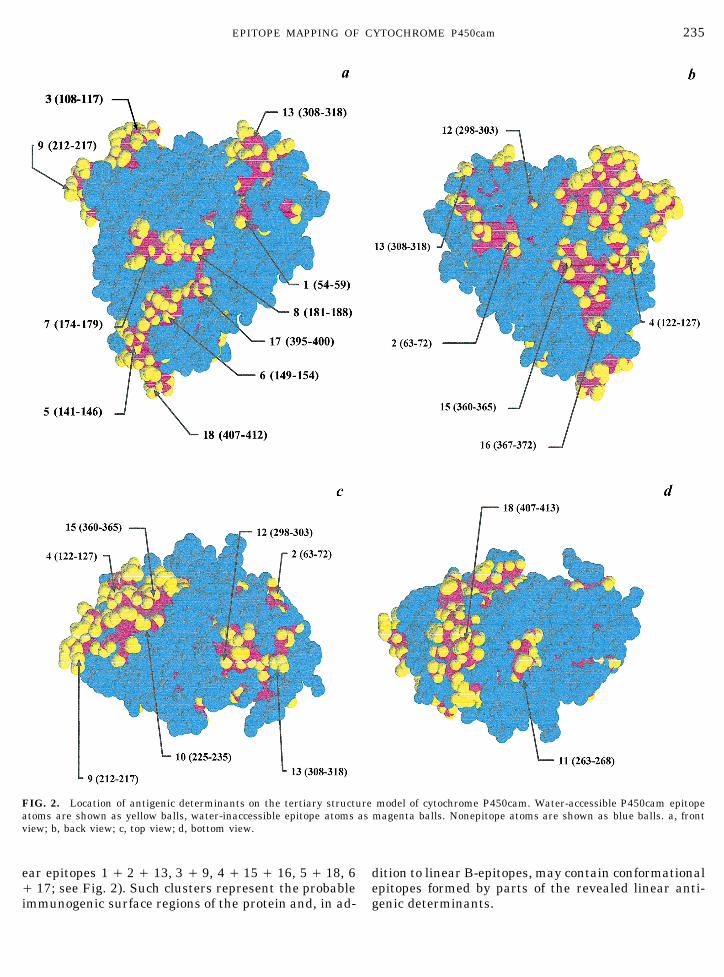
model of the three-dimensional structure of cytochrome Moreover, amino acid residues which are buried intothe protein globule may be more important for antibodyP450cam with the help of ONIX software (Figs. 2a–2d)
has revealed that almost all antigenic sites are oriented recognition than the surface-oriented residues, as hasbeen shown for the myohemerythrine linear hexapep-to the surface of the molecule and are fully or partly
accessible to water. According to the water-sized sphere tide epitope (41). Nevertheless, water inaccessibility ofpeptide Nos. 322– 327 may be the reason for its lowprobe (see Materials and Methods), three antigenic
sites (Nos. 4, 5, and 9) are fully accessible to water; 11 immunogenicity: this B-epitope may be inaccessible towater and immunoglobulins in the native P450camantigenic sites contain 50% or more water-accessible
amino acid residues. Amino acid residues of three other molecule and becomes accessible after a partial changein the protein native conformation.antigenic sites (Nos. 2, 16, and 17) are water accessible
for 30–50%, and only one antigenic site, No. 14 (322– It should also be mentioned that the majority ofP450cam linear epitopes are not evenly distributed on327, QMLSGL), is fully inaccessible to water mole-
cules. Though the strict correlation between the immu- the surface of the molecule but form several clusterswhich include linear epitopes located in different sitesnogenicity and water accessibility is not observed, the
most immunogenic P450cam linear antigenic sites of the P450cam sequence (e.g., clusters formed by lin-
AID ABB 9934 / 6b32$$$343 04-14-97 09:45:59 arca
235EPITOPE MAPPING OF CYTOCHROME P450cam
FIG. 2. Location of antigenic determinants on the tertiary structure model of cytochrome P450cam. Water-accessible P450cam epitopeatoms are shown as yellow balls, water-inaccessible epitope atoms as magenta balls. Nonepitope atoms are shown as blue balls. a, frontview; b, back view; c, top view; d, bottom view.
ear epitopes 1 / 2 / 13, 3 / 9, 4 / 15 / 16, 5 / 18, 6 dition to linear B-epitopes, may contain conformationalepitopes formed by parts of the revealed linear anti-/ 17; see Fig. 2). Such clusters represent the probable
immunogenic surface regions of the protein and, in ad- genic determinants.
AID ABB 9934 / 6b32$$$343 04-14-97 09:45:59 arca

236 KOLESANOVA ET AL.
FIG. 3. Superposition of antigenic map over structural– functional map of P450cam.
Figure 3 demonstrates the superposition of the can be made. Eighteen linear antigenically active siteswith different immunogenic potential have been re-P450cam antigenic map over the structural–functional
map of this protein. Surface location of P450cam anti- vealed in the cytochrome P450cam with the help of PEP-SCAN. No one antigenic hexapeptide of P450cam is de-genic sites explains well why several functionally ac-
tive residues which are responsible for intermolecular tected in any other member of the P450 superfamily.P450cam linear antigenic determinants resideinteractions in the Ps. putida monooxygenase system
are positioned inside the found B-epitopes. Four basic mainly at the boundaries of secondary structure ele-ments. They are preferably located on the surface ofamino acid residues proven to be involved in putidare-
doxin and cytochrome b5 binding (Arg72, Arg112, the protein molecule and are accessible to water. Thesame is true for two other proteins with known tertiaryLys314, and Arg364) (9, 10, 28) are located inside
P450cam linear antigenic sites Nos. 2, 3, 13, and 15, structures, myohemerythrine (22) and dihydrofolatereductase (38). Thus, these data can be used to predictrespectively. Several probable substrate-binding sites
of P450cam (II, V, and VI) (10, 24, 28) partially or fully protein antigenic determinants. Vice versa, antigenicmapping results may be applied to some extent for cor-coincide with antigenic sites Nos. 8, 12, and 17, respec-
tively. Gln108, Arg112, and Arg299 which interact with recting protein spatial structure predictions.Several functionally active amino acid residues ofheme propionate groups (10, 24, 28) are also positioned
inside P450cam antigenic sites Nos. 3 and 12. Lys178 P450cam which participate in the inter- and intramo-lecular protein–ligand interactions in the Ps. putidaand Arg186 are the components of the proton-translo-
cation chain (37) and also of antigenic sites Nos. 7 and monooxygenase system are located inside or close tothe found B-epitopes. These results give an excellent8. In addition, there are several functionally important
residues which are located very close to the revealed opportunity to produce various monospecific antipep-tide antisera to P450cam which can be used in theantigenic sites. Val295 and Asp297 take part in sub-
strate binding, and Asp297 interacts with the heme studies of different types of interactions within themonooxygenase system.propionate residue (10, 24, 28); both Val295 and
Asp297 are also adjacent to the B-epitope No. 12. Theaxial heme ligand Cys357 (10, 24, 28) is located close
ACKNOWLEDGMENTSto the B-epitope No. 15. Glu366 which participates inthe formation of the inner proton-translocating channel We are indebted to Dr. O. G. Grebenshchikova for performingresides just between B-epitopes Nos. 15 and 16. Thus, amino acid analyses and to Dr. Yu. A. Lyulkin for help with the
ONIX software. Assistance of Mrs. J. G. Kiselar in preparing theP450cam sites which are considered to be responsibleP450cam antiserum is gratefully acknowledged. We are also gratefulfor the protein–protein or enzyme–substrate interac-to Mr. V. G. Kuznetsov for his excellent technical assistance. Thistions also possess immunogenic activities in the native work was supported by Grant 93-04-6227 from the Russian Founda-
P450cam molecule. tion for Basic Research, Russian Academy of Sciences, and GrantM1K000 from the International Science Foundation.To summarize our work, some principal conclusions
AID ABB 9934 / 6b32$$$343 04-14-97 09:45:59 arca

237EPITOPE MAPPING OF CYTOCHROME P450cam
22. Geysen, H. M., Tainer, J. A., Rodda, S. J., Mason, T. J., Alexan-REFERENCESder, H., Getzoff, E. D., and Lerner, R. A. (1987) Science 235,
1. Nebert, D. W., and Nelson, D. R. (1991) Methods Enzymol. 206, 1184–1190.3–11.
23. Worthington, J., and Morgan, K. (1994) in Peptide Antigens. A2. Nebert, D. W., Nelson, D. R., Conn, M. J., Estabrook, R. W., Fey- Practical Approach (Wisdom, G. B., Ed.), pp. 181–217, Oxford
ereisen, R., Fujii-Kuriyama, Y., Gonzalez, F. J., Guengerich, Univ. Press, Oxford, New York, Tokyo.F. P., Gunsalus, I. C., Johnson, E. F., Loper, J. C., Sato, R., Wa-
24. Poulos, T. L., Finzel, B. C., and Howard, A. J. (1987) J. Mol. Biol.terman, M. R., and Waxman, D. J. (1991) DNA Cell Biol. 10, 1–195, 687–700.14.
25. Ravichandran, K. G., Boddupalli, S. S., Hasemann, C. A., Pe-3. Archakov, A. I., and Bachmanova, G. I. (1990) Cytochrome P- terson, J. A., and Deisenhofer, J. (1993) Science 261, 731–736.
450 and Active Oxygen, Taylor & Francis, London, New York,26. Hasemann, C. A., Ravichandran, K. G., Peterson, J. A., andPhiladelphia.
Deisenhofer, J. (1994) J. Mol. Biol. 236, 1169–1185.4. Parke, D. V., Ioannides, C., and Lewis, D. F. V. (1991) Can. J.27. Cupp-Vickery, J.-R., and Poulos, T. L. (1995) Nature Struct. Biol.Physiol. Pharmacol. 69, 537–549.
2, 144–153.5. Degtyarenko, K. N., and Archakov, A. I. (1993) Biochem. Mol.28. Sligar, S. G., Filipovic, D., and Stayton, P. S. (1991) MethodsBiol. Int. 31, 1071–1080.
Enzymol. 206, 31–50.6. Edwards, R. J., Murray, B. P., Boobis, A. R., and Davies, D. S.29. Jung, C., Hui Bon Hoa, G., Schroeder, K.-L., Simon, M., and(1989) Biochemistry 28, 3762–3770.
Doucet, J. P. (1992) Biochemistry 31, 12855–12862.7. Edwards, R. J., Murray, B. P., and Boobis, A. R. (1991) Methods30. Das, M. K., and Lindstrom, J. (1991) Biochemistry 30, 2470–Enzymol. 206, 220 –233.
2477.8. Gotoh, O. (1992) in Cytochrome P450: Biochemistry and Bio-31. Gunsalus, I. C., Sligar, S. G., and Unger, B. P. (1986) J. Biol.physics (Archakov, A. I., and Bachmanova, G. I., Eds.), pp. 686–
Chem. 261, 1158–1163.691, INCO-TNC, Moscow.32. Ivanov, A. S., Rumjantsev, A. B., Skvortsov, V. S., and Archakov,9. Korzekwa, K. R., and Jones, J. P. (1993) Pharmacogenetics 3, 1–
A. I. (1996) J. Chem. Inf. Comput. Sci. 36, 660–663.18.33. Novotny, J., Handschmacher, M., Haber, E., Bruccoleri, R. E.,10. Poulos, T. L. (1991) Methods Enzymol. 206, 11–30.
Carlson, W. B., Fanning, D. W., Smith, J. A., and Rose, G. D.11. Tretiakov, V. E., Degtyarenko, K. N., Uvarov, V. Yu., and Archa- (1986) Proc. Natl. Acad. Sci. USA 83, 226 –230.
kov, A. I. (1989) Arch. Biochem. Biophys. 275, 429–439.34. Abola, E. E., Berstein, F. C., Bryant, S. H., Koetzle, T. F., and
12. De Lemos-Chiarandini, C., Frey, A. B., Sabatini, D. D., and Krei- Weng, J. (1987) in Data Commission of the International Unionbich, G. (1987) J. Cell Biol. 104, 209–219. of Crystallography (Allen, F. H., Bergerhoff, G., and Sievers, R.,
13. Edwards, R. J., Murray, B. P., Murray, S., Singleton, A. M., Da- Eds.), pp. 107 –132, Bonn-Cambridge-Chester.vies, D. S., and Boobis, A. R. (1993) Biochim. Biophys. Acta 1161, 35. Novotny, J., Bruccoleri, R. E., and Saul, F. A. (1989) Biochemis-38–46. try 28, 4735–4742.
14. Kronbach, T., and Johnson, E. F. (1991) J. Biol. Chem. 266, 36. Kolesanova, E. F., Kiselar, J. G., Jung, C., Kozin, S. A., Hui Bon6215–6220. Hoa, G., and Archakov, A. I. (1996) Biochimie 78.
15. Myers, M. J., Liu, G., Miller, H., Gelboin, H. V., Robinson, R. C., 37. Shimado, H., Makino, R., Unno, M., Hariuchi, T., and Tshimura,and Friedman, F. K. (1990) Biochem. Biophys. Res. Commun. Y. (1994) in Cytochrome P-450 Biochemistry, Biophysics and169, 171–176. Molecular Biology (Lechner, M. C., Ed.), pp. 299–306, John Lib-
16. Reik, L. M., Levin, W., Ryan, D. E., and Thomas, P. E. (1982) J. bey Eurotext, Paris.Biol. Chem. 257, 3950–3957. 38. Tan, X., Ratnam, M., Huang, S., Smith, P. L., and Freisheim,
17. Ryan, D. E., Thomas, P. E., Levin, W., Maines, S. L., Bandiera, J. H. (1990) J. Biol. Chem. 265, 8022–8026.S., and Reik, L. (1993) Arch. Biochem. Biophys. 301, 282 –293. 39. Ching, W.-M., Wychowski, C., Beach, M. J., Wang, H., Davies,
18. Edwards, R. J., Murray, B. P., Singleton, A. M., Davies, D. S., C. L., Carl, M., Bradley, D. W., Alter, H. J., Feinstone, S. M., andand Boobis, A. R. (1993) Biochem. Pharmacol. 46, 213–220. Shih, J. W.-K. (1992) Proc. Natl. Acad. Sci. USA 89, 3190–3194.
19. Thomas, P. E., Reidy, J., Reik, L. M., Ryan, D. E., Koop, D. R., 40. Briand, J., Julistiono, H., Beaune, P., Flinois, J. P., Dewaziers,and Levin, W. (1984) Arch. Biochem. Biophys. 235, 239–253. I., and Leroux, J. P. (1993) Biochim. Biophys. Acta 1203, 199–
204.20. Yamamoto, A. M., Cresteil, D., Boniface, O., Clerc, F. F., andAlvarez, F. (1993) Eur. J. Immunol. 23, 1105–1111. 41. Alexander, H., Alexander, S., Getzoff, E. D., Tainer, J. A.,
Geysen, H. M., and Lerner, R. A. (1992) Proc. Natl. Acad. Sci.21. Kolesanova, E. F., Kozin, S. A., Lemeshko, A. O., and Archakov,A. I. (1994) Biochem. Mol. Biol. Int. 32, 465–473. USA 89, 3352–3356.
AID ABB 9934 / 6b32$$$344 04-14-97 09:45:59 arca




















