
Epitope Repertoire of Human CD4+ T Cells on Tetanus Toxin
11
Epitope Repertoire of Human CD4+ T Cells on Tetanus Toxin: Identification of Immunodominant Sequence Segments Brenda M. Diethelm-Okita, Raghavanpillai Raju, David K. Okita, Bianca M. Contl-Fine Downloaded from https://academic.oup.com/jid/article/175/2/382/816687 by guest on 04 July 2022
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Epitope Repertoire of Human CD4+ T Cells on Tetanus Toxin

Epitope Repertoire of Human CD4+ T Cells on
Tetanus Toxin: Identification of Immunodominant
Sequence Segments
Brenda M. Diethelm-Okita, Raghavanpillai Raju, David K. Okita, Bianca M.
Contl-Fine
Dow
nloaded from https://academ
ic.oup.com/jid/article/175/2/382/816687 by guest on 04 July 2022

382
Epitope Repertoire of Human CD4+ T Cells on Tetanus Toxin: Identification ofImmunodominant Sequence Segments
Brenda M. Diethelm-Okita, Raghavanpillai Raju,*David K. Okita, and Bianca M. Contl-Fine"
Department ofBiochemistry, College ofBiological Sciences, Universityof Minnesota, St. Paul, and Department ofPharmacology, School of
Medicine, University ofMinnesota, Minneapolis, Minnesota
Sequence regionsoftetanus toxin-forming CD4+ cellepitopes in 8 HLA-disparatesubjectswereidentified. Overlapping synthetic peptides corresponding to the complete tetanus toxin sequencewere used to test, in a proliferationassay,unselected bloodCD4+ cellsor CD4+ celllines propagatedby stimulation with tetanus toxoid.The CD4+ cell lines recognized most peptides recognized by thebloodCD4+ cellsand they recognized additional peptides.Their responses werestronger than thoseof unselectedbloodCD4+ cells. Twopeptides wererecognized by all subjects: one largelyoverlappeda tetanus toxin sequence region previously identified as a "universal" T cellepitope.Thirteen otherpeptides elicited a CD4+ cell response in 6 or 7 of the 8 subjects, and another 10 elicited responsesin 5 subjects.
Tetanus, caused by the bacterium Clostridium tetani, is stilla major health problem in underdeveloped countries. Tetanusrelated mortality is due to the secreted tetanus toxin (TTX), apowerful neurotoxin that reaches by retroaxonal transport andtranscytosis the cytoplasm of spinal inhibitory intemeurons andblocks their ability to release neurotransmitters [1-3]. TTX isstructurally similar to botulinum toxin, which is secreted byClostridium botulinum and causes flaccid paralysis by blockingthe release of the neurotransmitter acetylcholine by the nerveterminals innervating the muscle [1-3]. Tetanus and botulinumtoxins are the most potent toxins known [1- 3].
TTX is synthesized as a precursor protein of ~ 150 kDa,which is cleaved by clostridial proteases into a light (L, ~50kDa) and a heavy (H, ~ 100 kDa) chain, linked by two disulfidebridges and noncovalent interactions [1-3]. The H chain anchors onto the surface receptors of neurons and mediates theentry of the L chain into the cytoplasm [2- 3]. The L chain isresponsible for the toxic activity of TTX, has a Zn2+ bindingdomain (HEXXH) [4-5]-a characteristic feature of Zn2+ endopeptidases [6]-and possesses zinc endopeptidase activityspecific for synaptobrevin, an integral membrane protein ofsmall synaptic vesicles [2].
Tetanus toxoid (TT), a partially denatured, nontoxic form ofTTX, is a key component of antitetanus vaccines [1] and isextensively used as a carrier in other vaccines [7-11].
Received 5 April 1996; revised 23 September 1996.Informed consent was obtained from all donors of blood used in these
studies. The guidelines for the use of human subjects in research used at theUniversity of Minnesota were strictly followed.
Grant support: NIH (NS-239l9 to B.M.C.-F.).Reprints or correspondence: Dr. Bianca M. Conti-Fine, Dept. of Biochemistry,CBS, University of Minnesota, 1479 Gortner Ave., St. Paul, MN 55108.
* Present affiliation: Department of Immunology, Mayo Clinic, Rochester,Minnesota.
t Previously known as Bianca M. Conti-Tronconi.
The Journal of Infectious Diseases 1997;175:382-91© 1997 by The University of Chicago. All rights reserved.0022~1899/9717502~OOI7$Ol.OO
Studies using synthetic sequences of protein antigens, suchas TTX, diphtheria toxin (DTX), and the nicotinic acetylcholinereceptor (the autoantigen in myasthenia gravis), have identifiedmany epitopes recognized by human CD4+ T helper cells onthese molecules [12-17]. Some sequence regions, immunodominant region sequences (IRSs), of these proteins were recognized by the CD4+ cells of all subjects tested, irrespectiveof their HLA class II haplotype (i.e., they can be restricted bymultiple class II alleles) [12-17]. A previous study on thehuman T cell response to TTX suggested that two sequenceregions corresponding to residues 830-843 and 947-966 ofthe precursor (i.e., residues 374-387 and 491-511 of the TTXH chain) might be recognized by all subjects [13]. Anotherstudy investigated the response ofhuman peripheral blood lymphocytes (PBL) to synthetic peptides spanning the TTX sequence and suggested that TTX contains several CD4+ cellepitopes, some of which were recognized by different subjects;the study did not confirm the existence of "universal" CD4+
cell epitopes [15].Because of the extensive use of TTX vaccines, it is worth
while to identify sequence regions that dominate the humanCD4+ cell response to this protein. Identification of such TTXsequences would allow for the design of synthetic or semisynthetic vaccines composed of or fortified with the TTX peptidescontaining universal CD4+ cell epitopes.
Also, the response to TTX can be used for studies on theCD4+ T cell response either as a control antigen to detect theoverall CD4+ cell function or as a test antigen to investigatethe mechanism of processing and recognition of exogenousantigens by CD4+ cells in humans [18]. A panel of commonlyrecognized, chemically well-defined synthetic TTX epitopewould be a useful tool for those studies.
Procedures allowing reliable identification of the sequenceregions of given pathogens that dominate the sensitization ofCD4+ cells would be of general interest. Direct testing ofunselected PBL is unlikely to have sufficient sensitivity. Antigenspecific CD4+ T cell lines, which can be propagated fromsubjects previously exposed to the vaccine or the pathogen l:!!ld
Dow
nloaded from https://academ
ic.oup.com/jid/article/175/2/382/816687 by guest on 04 July 2022

JID 1997; 175 (February) Human Th Epitopes on Tetanus Toxin 383
Table 1. Characteristics of subjects used for this study.
AgeSubject Sex (years) DR(B1) DR(B3-5) DQ
1 M 27 4 5(w12) w53 w52a w7(w3)2 M 26 4 6(w13) w53 w52b w8(w3) w6(w1)3 M 35 3(w17) 5(wll) w52 w52 w2 w7(w3)4 M 29 2(w16-Dw21) 5(wll) w51 w52 w5(w1) w7(w3)5 F 28 3(w17) 5(w11) w52 w52 w2 w7(w3)6 M 29 4(Dw10) 2(w15-Dw2) w53 w51 w8(w3) w6(w1)7 M 26 2(w16-Dw21) 7 w51 w53 w5(w1) w28 F 40 4 5(wll) w53 w52 w7(w3) w2
NOTE. DRlDQ haplotypes were determined by restriction fragment length polymorphism [20], oligonucleotidetyping [21], or both. Italicized letters indicate class II genes whose expressions were not directly tested but whichare generally associated withsubjects' particular DR hap1otypes.
which are frequently used to study normal and autoimmune
responses, might not faithfully represent the original CD4+
cell epitope repertoire because selective loss or preferential
propagation ofT cell clones may occur in culture, due to differ
ences in antigen processing in vivo and in vitro [19]. Also, the
high concentrations of antigens used in vitro might spawn a
higher degree of repertoire diversity than that existing in vivo.
The CD4+ cell response to TT in humans is an excellent system
to investigate the extent to which the epitope repertoire of
antigen-specific CD4+ cell lines is representative of that of the
CD4+ T cells in vivo, because there is no shortage of persons
immunized against this antigen.
The goals of the present studies were to verify the existence
of IRSs on the TTX sequence and to investigate the extent
of similarity of the repertoire of the blood CD4+ T cells of
recently vaccinated subjects compared with that recognized
by TTX-specific CD4+ cell lines propagated in vitro from the
same subjects. Toward those aims, we used synthetic peptides
that were 20 residues long and overlapping by 5 residues,
spanning the complete sequences of the TTX Hand L chains,
to map epitopes recognized by CD4+ cells of 8 healthy sub
jects with different major histocompatibility complex class II
haplotypes. The peptides were used in a proliferation assay
to test unselected blood CD4+ cells and TT-specific CD4+
cell lines propagated from the same subjects by stimulation
of the PBL with TT.
Materials and Methods
Subjects. Table 1 shows characteristics ofthe 8 study subjects.All had been immunized with TT, and all but one (donor 5) hadbeen boosted within 6 months prior to the study. Donor 5 hadbeen boosted 3 years earlier.
Peptide synthesis. We synthesized by manual parallel synthesis [22] 87 peptides (20 residues long and overlapping each otherby 5 residues) corresponding to the complete sequences of theTTX L chain (456 aa residues) and H chain (858 residues) [4].The peptides are indicated with codes that included the letter L orH for the L or H chain and two numbers referring to the position
of the first and the last residue of the peptide on the Land H chainsequences.
The peptide length was chosen because it is comparable to thatof naturally processed peptides [23-24]. Although class II-restricted epitopes are only 9-14 residues long [23-24], the presenceof extra residues at either end of the epitope sequence should notaffect peptide attachment to the binding cleft of the DR molecules,which is open at both ends [25]. The use of peptides of a lengthwithin the range of naturally processed peptides could result inpresentation of the specific epitope(s) without the need for processing, thus avoiding failure to stimulate the epitope-specific Tcell due to inappropriate processing of the peptide [15]. The peptides we used overlapped by ~5 residues to reduce the risk ofmissing epitopes "broken" between peptides. The synthetic peptides thus designed seemed a reasonable compromise between theconflicting needs of reducing the risk ofmissing epitopes and usinga peptide panel of manageable size.
The synthetic method we used yields peptides that are 65%85% pure [17-18,22]. Contaminants are shorter analogues randomly missing a residue because of incomplete coupling. Theymight be able to interact with the restricting HLA class II moleculebut not with the specific T cell receptor, thus, compared withpure epitope peptides, shifting the dose dependence of the T cellresponse to the peptide preparation containing the analogues. Inthe experiments reported here, we used generous amounts of stimulatory peptides, so such a risk is negligible.
The amino acid composition of the peptides found to containIRSs (see below) and of several other randomly selected peptideswas verified by derivation of amino acid residues released by acidhydrolysis with phenylisothiocyanate, followed by separation byreverse-phase high-pressure liquid chromatography [26]. For allpeptides, the results of the composition analysis corresponded tothe expected theoretical values (data not shown).
CD8+ T cell depletion and proliferation assay. CD8 + T cellsmay inhibit the in vitro response of CD4 + cells to antigens [2728] and recognize TTX peptides, making the results of proliferation assays done with populations containing both CD4 + and CD8+cells difficult to interpret. To identify the TTX peptides recognizedby unselected blood CD4 + T cells, we used PBL depleted of CD8+cells by use of mouse anti-CD8 antibody (OKT8; Ortho, Raritan,NJ) and paramagnetic beads coated with goat antibody againstmouse immunoglobulin (Advanced Magnetics, Cambridge, MA)
Dow
nloaded from https://academ
ic.oup.com/jid/article/175/2/382/816687 by guest on 04 July 2022
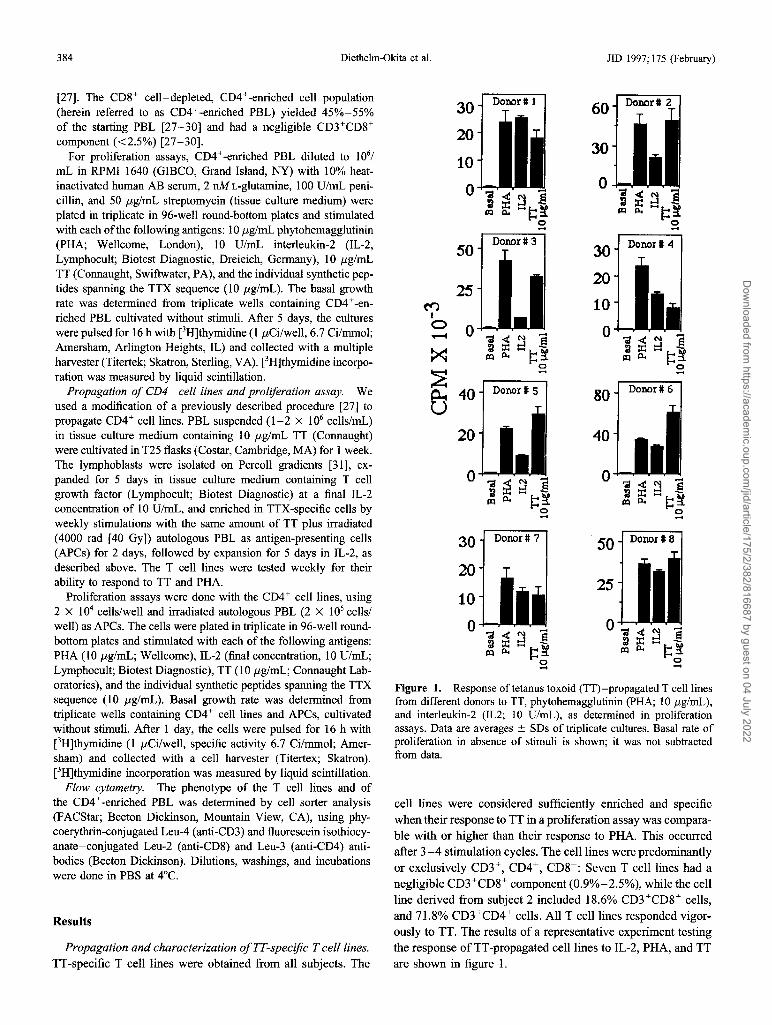
384 Diethelm-Okita et al. JID 1997;175 (February)
Donor I 6
Donor I 4
Donor # 8
~ -e ~ a~ g: - ~~
-
50
25
80
30
20
10
o
Donor I 7
40 Donor # 5
30
20
10
o
20
30
20
10
o fa -e N a~ g: dl::~
~
Figure 1. Responseof tetanus toxoid (TT)-propagated T cell1inesfrom different donors to TT, phytohemagglutinin (PHA; 10 j.lg/mL),and interleukin-2 (IL2; 10 U/mL), as determined in proliferationassays. Data are averages ± SDs of triplicate cultures. Basal rate ofproliferation in absence of stimuli is shown; it was not subtractedfrom data.
cell lines were considered sufficiently enriched and specificwhen their response to TT in a proliferation assay was comparable with or higher than their response to PHA. This occurredafter 3-4 stimulation cycles. The cell lines were predominantlyor exclusively CD3+, CD4+, CD8-: Seven T cell lines had anegligible CD3+CD8+ component (0.9%-2.5%), while the cellline derived from subject 2 included 18.6% CD3+CD8+ cells,and 71.8% CD3+CD4+ cells. All T cell lines responded vigorously to TT. The results of a representative experiment testingthe response of TT-propagated cell lines to IL-2, PHA, and TTare shown in figure I.
[27]. The CD8+ cell-depleted, CD4+-enriched cell population(herein referred to as CD4+-enriched PBL) yielded 45%-55%of the starting PBL [27-30] and had a negligible CD3+CD8+component «2.5%) [27-30].
For proliferation assays, CD4+-enriched PBL diluted to 1061
mL in RPMI 1640 (GIBCO, Grand Island, NY) with 10% heatinactivated human AB serum, 2 nML-glutamine, 100 U/mL penicillin, and 50 J.lg/mL streptomycin (tissue culture medium) wereplated in triplicate in 96-well round-bottom plates and stimulatedwith each ofthe following antigens: 10 J.lg/mL phytohemagglutinin(PHA; Wellcome, London), 10 U/mL interleukin-2 (IL-2,Lymphocult; Biotest Diagnostic, Dreieich, Germany), 10 J.lg/mLTT (Connaught, Swiftwater, PA), and the individual synthetic peptides spanning the TTX sequence (10 J.lg/mL). The basal growthrate was determined from triplicate wells containing CD4+-enriched PBL cultivated without stimuli. After 5 days, the cultureswere pulsed for 16 h with eH]thymidine (1 J.lCilwell, 6.7 Ci/mmol;Amersham, Arlington Heights, IL) and collected with a multipleharvester (Titertek; Skatron, Sterling, VA). eH]thymidine incorporation was measured by liquid scintillation.
Propagation of CD4+ cell lines and proliferation assay. Weused a modification of a previously described procedure [27] topropagate CD4+ cell lines. PBL suspended (1-2 X 106 cells/mL)in tissue culture medium containing 10 J.lg/mL TT (Connaught)were cultivated in T25 flasks (Costar, Cambridge, MA) for 1 week.The lymphoblasts were isolated on Percoll gradients [31], expanded for 5 days in tissue culture medium containing T cellgrowth factor (Lymphocult; Biotest Diagnostic) at a final IL-2concentration of 10 U/mL, and enriched in TTX-specific cells byweekly stimulations with the same amount of TT plus irradiated(4000 rad [40 GyD autologous PBL as antigen-presenting cells(APCs) for 2 days, followed by expansion for 5 days in IL-2, asdescribed above. The T cell lines were tested weekly for theirability to respond to TT and PHA.
Proliferation assays were done with the CD4+ cell lines, using2 X 104 cells/well and irradiated autologous PBL (2 X 105 cellslwell) as APCs. The cells were plated in triplicate in 96-well roundbottom plates and stimulated with each of the following antigens:PHA (10 J.lg/mL; Wellcome), IL-2 (final concentration, 10 U/mL;Lymphocult; Biotest Diagnostic), TT (10 J.lglmL; Connaught Laboratories), and the individual synthetic peptides spanning the TTXsequence (10 J.lg/mL). Basal growth rate was determined fromtriplicate wells containing CD4+ cell lines and APCs, cultivatedwithout stimuli. After 1 day, the cells were pulsed for 16 h with[3H]thymidine (I J.lCi/well, specific activity 6.7 Ci/mmol; Amersham) and collected with a cell harvester (Titertex; Skatron).eH]thymidine incorporation was measured by liquid scintillation.
Flow cytometry. The phenotype of the T cell lines and ofthe CD4+-enriched PBL was determined by cell sorter analysis(FACStar; Becton Dickinson, Mountain View, CA), using phycoerythrin-conjugated Leu-4 (anti-CD3) and fluorescein isothiocyanate-conjugated Leu-2 (anti-CD8) and Leu-3 (anti-CD4) antibodies (Becton Dickinson). Dilutions, washings, and incubationswere done in PBS at 4°C.
Results
Propagation and characterization ofTT-specific T cell lines.TT-specific T cell lines were obtained from all subjects. The
Dow
nloaded from https://academ
ic.oup.com/jid/article/175/2/382/816687 by guest on 04 July 2022
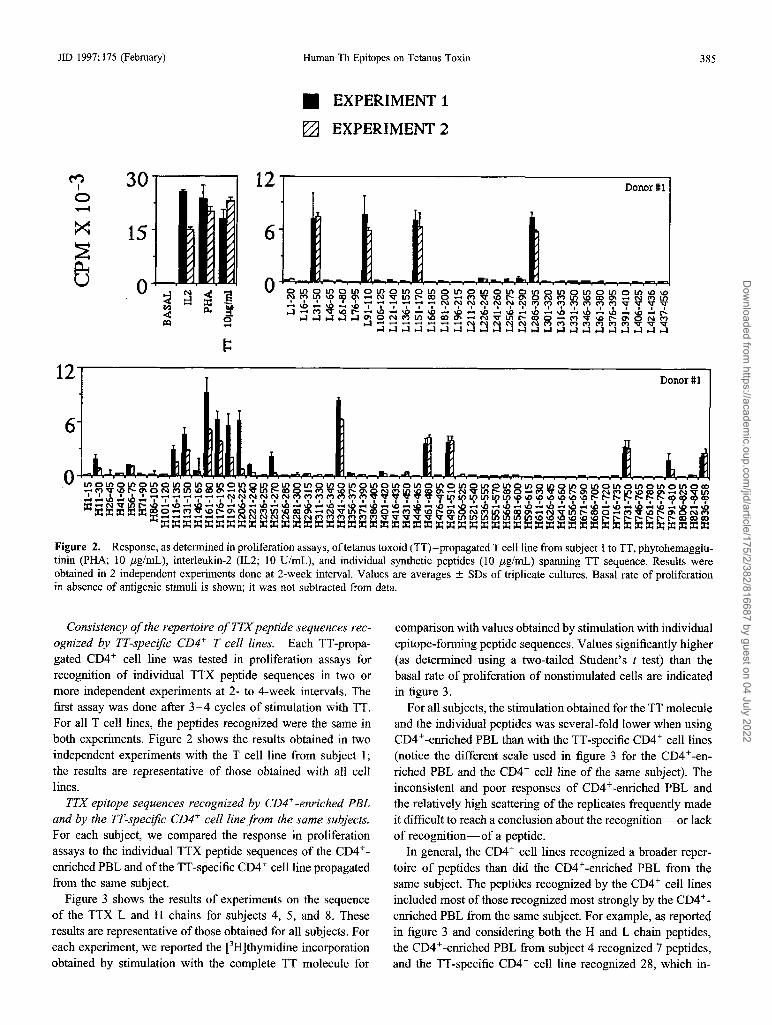
lID 1997;175 (February) Human Th Epitopes on Tetanus Toxin
II EXPERIMENT 1
~ EXPERIMENT 2
385
MIo--t
12
6
30
15
o
~~
~ ~~ ~ ~~ ~ ~
12
6
Donor#1
- - -- _ ...
Donor#1
Figure 2. Response, as determined in proliferation assays, oftetanus toxoid (TT)-propagated T cell line from subject 1 to TT, phytohemagglutinin (PHA; 10 j.lg/mL), interleukin-2 (IL2; 10 U/mL), and individual synthetic peptides (10 j.lg/mL) spanning TT sequence. Results wereobtained in 2 independent experiments done at 2-week interval. Values are averages ± SDs of triplicate cultures. Basal rate of proliferationin absence of antigenic stimuli is shown; it was not subtracted from data.
Consistency ofthe repertoire ofTTXpeptide sequences recognized by TT-specific CD4+ T cell lines. Each TT-propagated CD4 + cell line was tested in proliferation assays forrecognition of individual TTX peptide sequences in two ormore independent experiments at 2- to 4-week intervals. Thefirst assay was done after 3-4 cycles of stimulation with TT.For all T cell lines, the peptides recognized were the same inboth experiments. Figure 2 shows the results obtained in twoindependent experiments with the T cell line from subject I;the results are representative of those obtained with all celllines.
TTX epitope sequences recognized by CD4+-enriched PBLand by the TT-specific CD4+ cell line from the same subjects.For each subject, we compared the response in proliferationassays to the individual TTX peptide sequences of the CD4 +
enriched PBL and of the TT-specific CD4 + cell line propagatedfrom the same subject.
Figure 3 shows the results of experiments on the sequenceof the TTX Land H chains for subjects 4, 5, and 8. Theseresults are representative of those obtained for all subjects. Foreach experiment, we reported the [3H]thymidine incorporationobtained by stimulation with the complete TT molecule for
comparison with values obtained by stimulation with individualepitope-forming peptide sequences. Values significantly higher(as determined using a two-tailed Student's t test) than thebasal rate of proliferation of nonstimulated cells are indicatedin figure 3.
For all subjects, the stimulation obtained for the TT moleculeand the individual peptides was several-fold lower when usingCD4 +-enriched PBL than with the TT-specific CD4 + cell lines(notice the different scale used in figure 3 for the CD4 +-enriched PBL and the CD4+ cell line of the same subject). Theinconsistent and poor responses of CD4 +-enriched PBL andthe relatively high scattering of the replicates frequently madeit difficult to reach a conclusion about the recognition-or lackof recognition-of a peptide.
In general, the CD4+ cell lines recognized a broader repertoire of peptides than did the CD4 +-enriched PBL from thesame subject. The peptides recognized by the CD4 + cell linesincluded most of those recognized most strongly by the CD4 +
enriched PBL from the same subject. For example, as reportedin figure 3 and considering both the H and L chain peptides,the CD4 +-enriched PBL from subject 4 recognized 7 peptides,and the TT-specific CD4 + cell line recognized 28, which in-
Dow
nloaded from https://academ
ic.oup.com/jid/article/175/2/382/816687 by guest on 04 July 2022

..-.....~
Z~
~
~~~~~~~~~~ffi~~~~~+vQU"'-'
~ a::E40~~~~~~~~~~~~u
f 2:~"z,xa~~~~~~....~~~
6 Donor # 4,.-......~CO 3c,
+v- aQ 8u<:»
('tj
4I
0'P"""'l
~,.-......
20@~ Donor # 4 "**#co Z~ .......
+ 10 ~v- +Q vU a ~~ 30 U
(It") "'-'I (""')0 I'P"""'l 15 0
~~
~~
~O ~~u
f
U
25 t0
Figure 3. Response, as determined in proliferation assays, of CD4 + cell- enriched peripheral blood lymphocytes (PBL) and TTspecific CD4 + cell lines from donors 4, 5, and 8 to tetanus toxoid (TT) and individual synthetic peptides (10 tLg/mL) spanning tetanustoxin sequence. Values are averages ± SDs of triplicate cultures. Basal rate of proliferation in absence of antigenic stimuli is shown;rate was subtracted from data. Symbols indicate significant increase of [3H]thymidine incorporation in presence of peptide comparedwith that obtained in absence of antigenic stimuli: For CD4 +-enriched PBL, P < # .05 and ## .01; for CD4 + cell lines, P < * .05 and** .01).
Dow
nloaded from https://academ
ic.oup.com/jid/article/175/2/382/816687 by guest on 04 July 2022

JID 1997; 175 (February)
Respo.nse intenSi~:i.··:·C:% of the response •toTT
o
Light Chain Peptides
Human Th Epitopes on Tetanus Toxin 387
Subject #.....,....,.81HlI~~76
~--t-3H-tI--FlI:::HI-HI-H~I-I-I-f-I~54-H-i::i-+:f~"""++~++++++-+-I3
Heavy Chain Peptides
Figure 4. Contour map summarizing response of tetanus toxoid (TT)-specific T cell lines propagated from 8 subjects (1 for each horizontalrow) to individual tetanus toxin peptide sequences. Response of T cell lines to TT, at extreme left, is followed by response to individualpeptides (1 peptide for each vertical line), starting with amino-terminal sequence of light chain and ending with carboxyl-terminal sequenceof heavy chain. Responses to peptide are represented relative to response to TT.
eluded 6 of the 7 peptides recognized by the CD4+-enrichedPBL. The CD4+-enriched PBL from subject 5 recognized 6peptides, and the TT-specific CD4+ cell line recognized 31,which included all 6 peptides recognized by the PBL. TheCD4+-enriched PBL from subject 8 recognized 62 peptides,and the TT-specific CD4+ cell line recognized 63, which included 51 peptides recognized by the CD4+-enriched PBL.
In most subjects, a few peptides recognized significantly bythe CD4+-enriched PBL did not elicit a response from the antiTT CD4+ cell line propagated from the same donor. They werein general, but not always (e.g., see peptide H79l-810 forsubject 4), peptides recognized by the CD4+ PBL to a verylow extent (figure 3).
When CD4+-enriched PBL were used, the response to TTwas always higher than that to any TTX peptide. When theTT-specific CD4+ cell lines were tested, some peptides induceda proliferative response comparable to that induced by TT.
On the basis of the results summarized above, we concludedthat the response to TTX sequences of the CD4+ cell lines wasmore consistent and more easily detectable than that of theCD4+-enriched PBL from the same subject and identified abroader epitope repertoire, although recognition of some epitopes marginally recognized by the CD4+-enriched PBL mighthave gone undetected when the CD4+ cell lines were used.
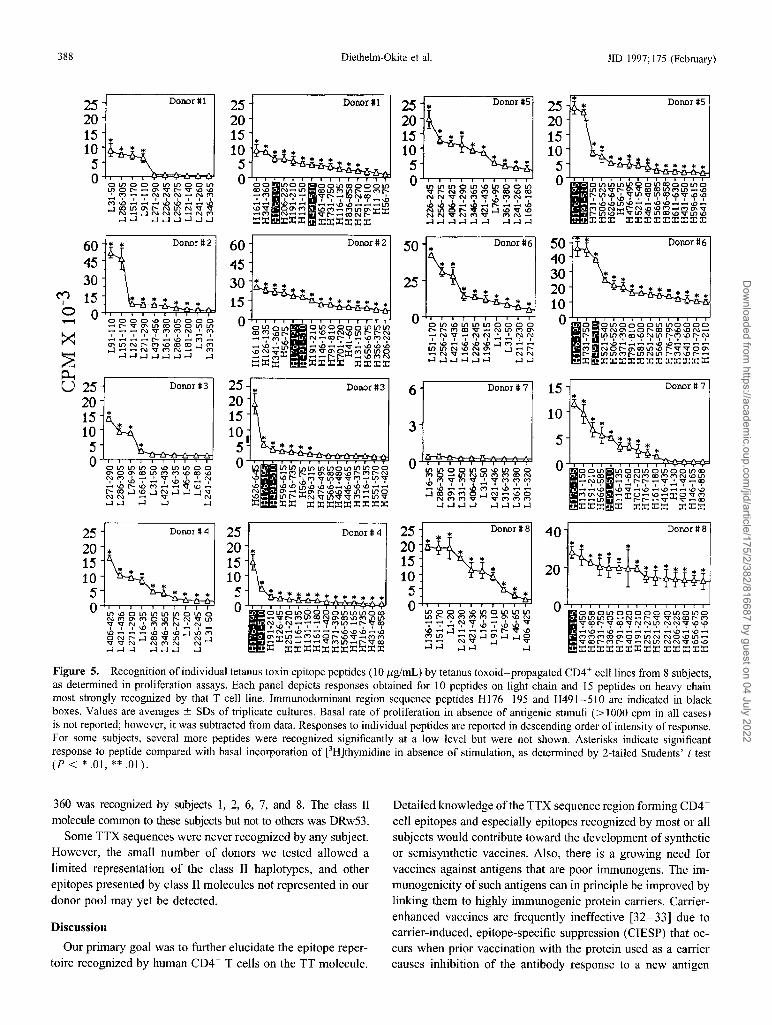
TTX epitope sequences recognized by the TT-specijic CD4+cell lines. Each subject recognized several peptides and hada characteristic pattern of peptide recognition. The response ofthe CD4+ cell lines to the individual TTX peptides is summarized in figure 4 as a contour map. Figure 5 shows the actualdata obtained for the peptides most strongly recognized byeach T cell line (10 peptides for the L chain and 15 for the Hchain). For some subjects, several peptides that were recognized significantly but at a low level are not included in figure5 but do appear in figure 4.
Most subjects recognized several peptides on both TTXchains. Donors 6 and 8 recognized a large number of peptides.At the opposite extreme, subjects 3 and 7 recognized very fewpeptides on the L chain, and subject 7 also recognized veryfew on the H chain.
Two peptides (H176-I95, IDKISDVSTIVPYIGPALNI,and H491-5IO, NNFTVSFWLRVPKVSASHLE) were recognized by the CD4+ cell lines of all subjects. The response ofthe T cell lines to these peptides was generally intense, togetheraccounting for 8%-39% of the overall response of the T celllines to the 87 TTX peptides (table 2). In addition, in mostcases, the CD4+-enriched PBL significantly recognized thesesequences (figure 3 and data not shown).
Several other peptides were recognized by most subjects.Thirteen peptides (L3I-50, L271-290, L286-305, H56-75,H1l6-135, H131-150, H161-180, HI9I-21O, H251-270,H431-450, H566-585, H73l-750 and H791-81O) elicited asignificant CD4+ cell response in 6 or 7 of the 8 subjects.In most cases, a significant response to these sequences wasobserved when using the TT-specific CD4+ cell lines and theCD4+-enriched PBL from the same subject. In subject 7, asignificant response to these sequences was frequently observedwhen using only the CD4+-enriched PBL (peptides L3I-50,H131-150, H25I-270, H431-450, H731-750, and H791810; data not shown). Nine peptides (L9I-llO, L151-170,L256-275, L361-380, L421-436, H146-I65, H341-360,H70I-720, and H836-858) were recognized by the CD4+cells of 5 subjects.
Some TTX peptides were uniquely recognized by 1 subject orby a groupof subjects expressing a commonHLAclassII molecule.Peptides LI8l-200 and H7I-90 were uniquely recognized bysubject2, who was the only donor expressing DR6(w13). PeptidesH206-225 and H221-240 were recognized by subjects 1, 2, 6,and 8, who all express the DR4/DQw3 molecules. PeptideH341-
Dow
nloaded from https://academ
ic.oup.com/jid/article/175/2/382/816687 by guest on 04 July 2022
388 Diethelm-Okita et al. lID 1997; 175 (February)
Donor #5
Donor # 7
Donor #6
OLnOOOOLnLnOOOOVNC1' .....Ol'"COC1'\D\DN .....l{'l{'C(op"P~u;>~f(\flr;--~......\D ...................... \O\D ...................NOl'"C1'COLn\Dl'"VVOC1'LnLnMl'"LnNLnl'"M\Ol'" .....
-:r::r: :r::r::r::r::r::r::r::r::r::r:
'"'"
'"
5040302010
oI!15
10
25201510
5 ~--r--T"""...,....-r-~:!;:!;.~~o
Donor #5
Donor # 76
3
25 '"201510
5o~T'"""T"--r-r-T"""'T"""'1---'-~
Donor #2
Donor #3
Donor #1
6045
3015
O-+-r-,..,...."'T"'"'"T-,--,..,......,....,--,-,"""'T"""".....
25201510
50..L,-......-T""-r--1"~r-r-"'T""'"T""""T"""1--r::y::....r,il
25 '"201510.
gl..-.-~~~~~~
Donor #2
Donor#3
Donor #1
...
... ...
OOOO\oOLnOOO..... l'"VC1'LnCOOOLnLn...... ~ ..... NVMMN'M
I I I I I I I I .... I......................... ['- ...... \0 ..... (\") ....C1'LnNl'"M\D CO CO.....lM.....l .......... NVMN ..... M
.....l.....l....:l....:l....:l....:l....:l ....:l
25201510
5o+r-"T""""""T"~~~)-O-oOJ
60453015O~~~~~
~
~~
U 252015 ...10
5O~~~~~¥:l
25 Donor # <:I 25 Donor # <:I 25 Donor # 8 40 Donor#8
20 '" 20 '"20
~15 15 15 '" '"20 ",,,, "'",,,,,,,,,,10 10 10
'"5 '" '" 5 5 '"'" '" '" '" ... "'''' ... ''' ...... '''''''''''''''''' '"0 0 0 0
IO~o~oooo~~~oro LnOOO\DLnOLnLnLn IOroo~oooooo~o~oLn\DOLnLnLnLnOLnO .....Vl'"MLnCONC1'CO\DMLnLn Lnl'"NMMM ..... C1'\ON LnLnLnO .....N .....l'"VVNCOl'"MNMC1'MO\Dl'"NVLn N'N ...............VMLn .....l'"VCO ......... I NV I ..... I I "'q'"y~r-;-'r~~~~U(~~~'f\fi>VVN'MMN'N' I \0 I I I I I I I I I I I I ........ '\OI\D\DI
I I I \0 I I I ..... I .... \D .... .....:1 ....................... r---V\D ..... \D .... \D ......................................\D ......\D ......\D .......... -\D\D\D....:l\DM .... N ........ \D ..................... \C)\/)\D ...... \lJ
~:r:~:::~~~q:;:;~3~~~MLn ..... N....:lC1'....:l....:l0 MMMCOC1'OC1'LnNNO\DLn .....
~~~....:l~~~ ~....:l .......... NV....:l V VCOl'"C"ll'"V .....NLnNNV\o\o
....:l....:l.....l ....:l....:l....:l ....:l .:r: :r::r::r::r::r::r::r::r::r::r::r: ....:l....:l....:l....:l ....:l :r::r::r::r::r::r::r::r::r::r::r::r::r::r:
Figure 5. Recognition of individual tetanus toxin epitope peptides (10 I-ig/mL) by tetanus toxoid-propagated CD4+ cell lines from 8 subjects,as determined in proliferation assays. Each panel depicts responses obtained for 10 peptides on light chain and 15 peptides on heavy chainmost strongly recognized by that T cell line. Immunodominant region sequence peptides H176-195 and H491-51O are indicated in blackboxes. Values are averages ± SDs of triplicate cultures. Basal rate of proliferation in absence of antigenic stimuli (> 1000 cpm in all cases)is not reported; however, it was subtracted from data. Responses to individual peptides are reported in descending order of intensity of response.For some subjects, several more peptides were recognized significantly at a low level but were not shown. Asterisks indicate significantresponse to peptide compared with basal incorporation of [3H]thymidine in absence of stimulation, as determined by 2-tailed Students' t test(P < * .01, ** .01).
360 was recognized by subjects 1, 2, 6, 7, and 8. The class IImoleculecommon to these subjects but not to others was DRw53.
Some TTX sequences were never recognized by any subject.However, the small number of donors we tested allowed alimited representation of the class II haplotypes, and otherepitopes presented by class II molecules not represented in ourdonor pool may yet be detected.
Discussion
Our primary goal was to further elucidate the epitope repertoire recognized by human CD4+ T cells on the TT molecule.
Detailed knowledge of the TTX sequence region forming CD4+
cell epitopes and especially epitopes recognized by most or allsubjects would contribute toward the development of syntheticor semisynthetic vaccines. Also, there is a growing need forvaccines against antigens that are poor immunogens. The immunogenicity of such antigens can in principle be improved bylinking them to highly immunogenic protein carriers. Carrierenhanced vaccines are frequently ineffective [32-33] due tocarrier-induced, epitope-specific suppression (CIESP) that occurs when prior vaccination with the protein used as a carriercauses inhibition of the antibody response to a new antigen
Dow
nloaded from https://academ
ic.oup.com/jid/article/175/2/382/816687 by guest on 04 July 2022

JID 1997; 175 (February) Human Th Epitopes on Tetanus Toxin 389
Table 2. Contribution of immunodominant region sequences (IRSs)
to the response to tetanus toxin (TTX) peptide sequences of anti
tetanus toxoid (TT) T cell lines.
% response to % response to % total response
Subject H176-l95 H49l-5l0 to both IRSs
1 9 6 15
2 12 12 24
3 19 8 27
4 10 7 17
5 20 19 39
6 16 14 30
7 17 7 24
8 6 2 8
NOTE. Fraction of total response to TTX peptide sequences due to response to IRSs was determined as follows: Each anti-TT line was challengedin triplicate proliferation assays with each individual peptide screening TTXsequence. Cell proliferation in absence of antigenic stimuli (blank) was determined in triplicate cultures containing T cell lines and antigen-presenting cellsonly. Average cpm values of nonstimulated cultures were subtracted fromthose obtained in presence of TTX peptides, and resulting values were added,to yield what was considered 100% stimulation by TTX peptides. Averagecpm obtained for each IRS minus blank was added, and fraction of totalresponse represented by this sum was calculated.
linked to that protein [34]. CIESP is due to a suppressiveimmune response to individual epitope sequences ofthe carrier.
TT has been used frequently as a carrier to increase theimmunogenicity of vaccines [7-11]. Because of the widespread use of antitetanus vaccination, the risk of CIESP raisesquestions about the suitability of TT as a vaccine enhancer.The universal CD4+ cell epitope sequences identified here andin other studies [13-15] are good candidates as alternativeeffective vaccine enhancers: If future studies indicate that thesetwo TTX sequences do not cause CIESP, their use would takeadvantage of the CD4+ cell priming resulting from prior immunization to TT, rather than being negatively affected by it [34].
TTX sequences forming CD4+ cell epitopes have beenshown to avoid CIESP and be effective vaccine enhancers: InBALB/c mice previously immunized against TTX, a TTX peptide recognized by T helper cells and linked with a peptidecontaining a B cell epitope, retained carrier but not suppressivefunction and enhanced the antibody response to the B cellepitope [34]. Bypass of CIESP by T cell epitope sequenceshas been demonstrated for a gonadotropin-releasing hormonebased vaccine: DTX-induced CIESP was overcome using aDTX T helper epitope sequence [35].
Other studies demonstrated that universal TTX T epitopesenhance the immune response to other antigens or haptens. ATTX sequence recognized by the CD4+ cells of most humans(residues 830-844 of the TTX precursor) induced, when usedas a carrier, an antibody response and overcame genetic nonresponsiveness to a synthetic sequence of Plasmodium falciparum in mice [36]. Constructs containing repetitive sequencesof P. falciparum and Plasmodium berghei and two universalTTX T cell epitopes (residues 830-843 and 947-967 of theTTX precursor) stimulated antibody production in mice to
Plasmodium epitopes and the in vitro response of human PBL[37]. Universal TTX epitopes strongly enhanced the immunogenicity in mice and rabbits of B cell epitope peptides forthe contraceptive vaccine candidate, lactate dehydrogenase C4[38]. The universal TTX CD4+ cell epitope peptide 830-843coupled to an anti-lymphoma idiotype antibody increased theeffectiveness of the antibody by facilitating cytotoxic lysis ofthe B cell lymphoma by specific CD4+ T cells [39].
The present results indicate that the antigen-propagated Tcell lines can be used with some confidence for studies on theepitope repertoire of CD4+ cells: The repertoire of peptidesrecognized by the CD4+ cell lines was frequently broader thanthat of CD4+ cell-enriched PBL, but it included most peptidesmore strongly recognized by the blood CD4+ cells (figure 3).The CD4+ cell line responses to individual epitopes were consistent and clear. On the other hand, antigen-specific CD4+cell lines may not include T cell clones recognizing epitopesequences less immunogenic or poorly processed in vitro.
Sequences H176-195 and H491-51O were vigorously recognized by the CD4+ cell lines from all subjects (figures 4and 5; table 2). Also, the CD4+ cell-enriched PBL frequentlyrecognized these peptides better than most other TTX sequences (figure 3). The strong response to those IRSs is the"tip" of a response to sequence regions that include peptidesequences on either side of the IRS (figure 4). Other TTXsequence regions strongly recognized by most subjects arethose flanking and including peptides L271-290 and H131150 (figure 4). The sequence region comprising the overlappingpeptides L406-425 and L421-436 was recognized by only 5subjects, and their response was very strong (figure 4). Whereasonly two TTX sequence regions were recognized by all thesubjects, the several other regions recognized by most subjects(figure 4 and Results) might also be viable candidates, as apool, for the design of synthetic vaccines and as an indicatorin serial measurements of the activity ofCD4+cells in immunosuppressed subjects.
IRSs have been found for the response of human CD4+ cellsto DTX [17] and to the muscle acetylcholine receptor [16].The molecular basis of the universal CD4+ cell recognition ispuzzling. Promiscuous binding to different class II moleculesis necessary but insufficient for a sequence to form an IRS [13,15, 40- 42]. Structural properties of protein antigens influencedetermination of IRSs in humans, as shown for DTX [17], inwhich all IRSs are flanked by relatively unstructured loopsfully exposed to the solvent on the DTX surface [43]. Thoseloops may be easy targets for the processing enzymes even inthe absence of substantial denaturation of DTX, and they givethe IRS an advantage during antigen processing. It is likelythat in the native TT molecule, the IRSs described here areflanked by or contain similar exposed loops.
Other studies identified TTX sequence-forming epitopes forhuman CD4+ cells [12-15]. The IRS H176-195 (residues632-651 of the TTX precursor) includes the sequence 640651 of the TTX precursor, which another study found to be aTTX sequence frequently recognized by human PBL [15]. IRS
Dow
nloaded from https://academ
ic.oup.com/jid/article/175/2/382/816687 by guest on 04 July 2022

390 Diethelm-Okita et al. JID 1997; 175 (February)
H49I-510 overlaps a previously described immunodominantTTX sequence (residues 947-967 of the TTX precursor, i.e.,residues H490-51O) [12, 13].
The sequence comprising residues 830-844 of the TTX precursor (residues H374-388) has also been identified as universally immunogenic in humans: Clones specific for that sequence were easily isolated from all TT-primed donors tested,irrespective of their DR type [13]. This sequence is includedin our peptide H37I-390, which was recognized by 3 donors(figure 4). Lack of recognition by the CD4+ cell lines derivedfrom the other donors is likely due to our use ofpolyclonal TTspecific T cell lines instead of clones: CD4+ cells recognizingepitopes within the sequence H37I-390 may have been lostduring propagation of the T cell lines due to competition byother clones. Also, in peptide H37I-390, residues 830-844are flanked at both the amino- and the carboxyl-terminal andby other residues, which may have exerted a negative influenceon its binding to class II molecules [44]. In support of thepossibility that we missed the universal recognition of thissequence for technical reasons, Panina-Bordignon et al. [13]demonstrated that sequence 830-844 was recognized in association with DR5(wII), DR6(w13), DR7, DRw52a, andDRw52b; all donors in our study whose T cell lines did notrecognize peptide H37I-390 expressed one or more of thoseDR molecules (table 1).
Demotz et al. [44] investigated the binding preference fortwo highly represented alleles ofDR5(w11)-DRB1*1101 andDRB1*1104, which differ only at residue DR,886-of twoimmunodominant TTX epitopes, residues 830-844 and 947967 of the TTX precursor (named peptides P2 and P30 in [44])and of other TTX sequences that can form epitopes recognizedby CD4+ cells of DR5(w11) donors. Two of the latter sequences, named P23 and P32, roughly corresponded to ourpeptides H626-645 and H7I6-735. In agreement with thatstudy, the peptides including the sequences listed above werefrequently recognized by our donors 3-5 and 8, who expressedthe DR5(wII) molecule (figure 4). Demotz et al. found that,while epitopes P2 and P32 could be recognized in associationwith both DRBI *1101 and DRBI *1104, P23 and P30 wererecognized in association with DRBI *1101 only. Peptide P30corresponds to residues H490-51O, which are included in ourIRS H49I-51O. Our finding that H49I-51O is recognized byall DR5(w11) subjects (we did not determine the specificDR5(w11) allele they expressed)-in fact, by all donors-isexplained by Panina-Bordignon et al. [13], who found that theDP molecules may also function as restriction elements ofpeptide 947-967 (i.e., P30). Thus, our peptide H491-51Omight be universally recognized by all donors tested by virtueof its ability to bind different class II isotypes and differentalleles of the same isotype [13].
In support of the promiscuous binding of the IRS peptidesHI76-195 and H491-51O are the results ofpreliminary experiments investigating whether both sets ofthe DR!DQ moleculescould present TTX epitopes. We challenged TT-specific T celllines from 5 subjects with individual TTX peptides, using APCs
from donors homozygous for each of the DR!DQ alleles of thesubject. The IRS peptides H176-195 and H491-51O werealways presented by APCs homozygous for either of the DR!DQ alleles of the donor, while other TTX epitope peptideswere frequently presented only by APCs homozygous for oneof the two DR!DQ alleles. Since in those experiments the DQmolecules expressed by the APCs used were unknown, theymight also have matched those of the donors of the T cell linesand, thus, restricted presentation of TTX epitope peptides [13].
Other TTX sequence regions frequently recognized by PBLin humans [15] corresponded to or included TTX peptides thatwe found strongly and frequently stimulatory for human CD4+cells. In excellent agreement with our results, a pool of overlapping peptides that roughly correspond to our peptides H356375 and H371- 390 was recognized in that study by 4 of 9donors [15]. A second region corresponded to residues L141171, which includes our peptide L151-170, which were recognized by 5 of our donors. A third region roughly correspondedto the sequence segment HI23-H207 [15], which overlaps fivepeptides that induced a response in 5 or 6 of our donors, andto the IRS peptide H176-I95.
We do not know which of the class II molecules expressedby our donors restrict presentation of the TTX epitopes described here. The donors expressed different DR/DQ molecules(and presumably different DP molecules as well), yet the CD4+cells of all of them recognized the IRS; therefore, the IRSpeptides must be promiscuous in their binding to class II molecules.
Some TTX peptide sequences were uniquely recognized bysubjects expressing a particular DR/DQ allele, suggesting thattheir recognition had more stringent demands for binding toclass II molecules. Peptides LI8I-200 and H71-90 were recognized only by subject 2, the only donor expressing theDR6(w13) molecule, which might restrict presentation of thosepeptides. Peptides H206-225 and H22I-240 were recognizedonly by subjects 1, 2, 6, and 8, who express the DR4/DQw3molecules. Peptide H341-360 was recognized by subjects 1,2, 6, 7, and 8. The DRw53 molecule, common to these 5subjects but which was not expressed by the other subjects,might be the restriction element used for presentation of thatpeptide.
References
1. Rappuoli R. New and improved vaccines against diphtheria and tetanus.In: Woodrow GC, Levine MM, eds. New generation vaccines. NewYork: Marcel Dekker, 1990:251-68.
2. Schiavo G, Benfenati F, Poulain B, et al. Tetanus and botulinum-B neurotoxins block neurotransmitter release by proteolytic cleavage of synaptobrevin. Nature 1992;359:832-5.
3. Montecucco C, Schiavo G. Tetanus and botulism neurotoxins: a new groupof zinc proteases. Trends Biochem Sci 1993;324-7.
4. Eisel D, Jarausch W, Goretzki K, et al. Tetanus toxin: primary structure,expression in E. coli, and homology with botulinum toxins. EMBO J1986;5:2495- 502.
5. Wright JF, Pemollet M, Reboul A, Aude C, Colomb MG. Identificationand partial characterization of a low affinity metal-binding site in thelight chain of tetanus toxin. J Biol Chern 1992;267:9053-8.
Dow
nloaded from https://academ
ic.oup.com/jid/article/175/2/382/816687 by guest on 04 July 2022

JID 1997;175 (February) Human Th Epitopes on Tetanus Toxin 391
6. Jongeneel CV, Bouvier J, Bairoch A. A unique signature identifies a familyof zinc-dependent metallopeptidases. FEBS Lett 1989;242:211-4.
7. Paoletti LC, Wessels MR, Michon F, DiFabio J, Jennings HJ, Kasper DlL.Group B Streptococcus type II polysaccharide-tetanus toxoid conjugate
vaccine. Infect Immun 1992;60:4009-14.8. Dame JB, Williams JL, McCutchan TF, et al. Structure of the gene encod
ing the immunodominant surface antigen on the sporozoite of the humanmalaria parasite Plasmodium falciparum. Science 1984;225:539-59.
9. Zavala F, Tam JP, Hollingdale MR, et al. Rationale for development of asynthetic vaccine against Plasmodium falciparum malaria. Science
1985;228:1436-40.10. Mukherjee J, Casadevall A, Scharff MD. Molecular characterization of
the humoral responses to Cryptococcus neoformans infection andglucuronoxylomannan- tetanus toxoid conjugate immunization. J Exp
Med 1993; 177:1105-16.11. Palker n, Riggs ER, Spragion DE, et al. Mapping of homologous, amino
terminal neutralizing regions of human T-cell Iymphotropic virus typeI and II gp46 envelope glycoproteins. J ViroI1992;66:5879-89.
12. Demotz S, Lanzavecchia A, Eisel U, Niemann H, Widmann C, CorradinG. Delineation of several DR-restricted tetanus toxin T cell epitopes. JImmunol 1989; 142:394-402.
13. Panina-Bordignon P, Tan A, Termijtelen A, Demotz S, Corradin G, Lanza
vecchia A. Universally immunogenic T cell epitopes: promiscuous binding to human MHC class II and promiscuous recognition by T cells.Eur J Immuno11989; 19:2237-42.
14. Ho PC, Mutch DA, Winkel KD, et al. Identification of two promiscuous
T cell epitopes from tetanus toxin. Eur J ImmunoI1990;20:477-83.15. Reece JC, Geysen HM, Rodda S1. Mapping the major human T helper
epitopes of tetanus toxin. The emerging picture. J Immunol1993; 151:
6175-84.16. Protti MP, Manfredi AA, Horton BM, Bellone M, Conti-Tronconi BM.
Myasthenia gravis: recognition of a human autoantigen at the molecularlevel. Immunol Today 1993; 14:363-8.
17. Raju R, Navaneethan D, Okita DK, Diethelm-Okita B, McCormick D,Conti-Fine BM. Epitopes for human CD4+ T cells on diphtheria toxin:structural features of sequence segments forming epitopes recognizedby most subjects. Eur J ImmunoI1995;25:3207-14.
18. Panina-Bordignon P, Demotz S, Corradin G, Lanzavecchia A. Study onthe immunogenicity of human class-II-restricted T-cell epitopes: processing constraints, degenerate binding, and promiscuous recognition.Cold Spring Harb Symp Quant Bioi 1989;54:445-5.
19. Raju R, Diethelm-Okita B, Okita DK, Conti-Fine BM. Epitope repertoireof human CD4 + lines propagated with tetanus toxoid or with synthetictetanus toxin sequences. J Autommun 1996; 9:79-88.
20. Noreen HJ, Davidson ML, McCullough J, Bach FH, Segall M. HLAclass II typing by restriction fragment length polymorphism (RFLP) inunrelated bone marrow transplant patients. Transplant Proc 1989; 21:2968-70.
21. Kimura A, Sasazuki T. Reference protocol for the HLA DNA-typing technique. In: Proceedings of the II th International Histocompatibility Workshop and Conference. Oxford, UK: Oxford University Press, 1992:397.
22. Houghten RA. General method for the rapid solid phase synthesis of largenumbers of peptides: specificity of antigen-antibody interaction at the levelof individual amino acids. Proc Natl Acad Sci USA 1985;82:7048-52.
23. Rudensky AY, Preston-Hurlburt P, Hong SC, Barlow A, Janeway CA Jr.Sequence analysis of peptide bound to MHC class II molecules. Nature1991; 353:622-7.
24. Hunt DF, Michel H, Dickinson TA, et al. Peptides presented to the immunesystem by the murine class II major histocompatibility complex molecule I_Ad. Science 1992;256:1817-20.
25. Stem LJ, Brown JH, Jardetzky TS, et al. Crystal structure of the humanclass II MHC protein HLA-DRI complexed with an influenza viruspeptide. Nature 1994;368:215-21.
26. Heinrickson S, Meredith B. Amino acid analysis by reverse-phase highperformance liquid chromatography: precolumn derivatization with phenylisothiocyanate. Anal Biochem 1983; 136:65-74.
27. Protti MP, Manfredi AA, Straub C, Howard J Jr, Conti-Tronconi BM.
CD4 + T cell response to human acetylcholine receptor a subunit corre
lates with myasthenia gravis severity. A study with synthetic peptides.
J Irnmunol 1990; 144:1276-81.
28. Manfredi AA, Protti MP, WuXD, Howard JF, Conti-Tronconi BM. CD4+
epitope repertoire on the human acetylcholine receptor a subunit in
severe myasthenia gravis. A study with synthetic peptides. Neurology
1992;42:1092-100.
29. Manfredi AA, Protti MP, Dalton MWM, Howard JF Jr, Conti-Tronconi
BM. T helper recognition of muscle acetylcholine receptor in myasthe
nia gravis. Epitopes on the 'Y and .6. subunits. J Clin Invest 1993; 92:
1055-67.
30. Moiola L, Karachunski P, Protti MP, Howard J Jr, Conti-Tronconi BM.
Epitopes on the f3 subunit of human muscle acetylcholine receptor
recognized by CD4 + cells of myasthenia gravis and healthy subjects. J
Clin Invest 1994;93:1020-8.
31. Protti MP, Manfredi AA, Straub C, Howard JF, Conti-Tronconi BM. Im
munodominant regions for T helper-cell sensitization on the human
nicotinic receptor a subunit in myasthenia gravis. Proc Natl Acad Sci
USA 1990; 87:7792-6.
32. Anderson P, Pichichero ME, Insel RA. Immunogens consisting of oligo
saccharides from the capsul of Haemophilus influenzae type b coupled
to diphtheria toxoid or the toxin protein CRM197. J Clin Invest 1985;
76:52-9.
33. Herrington DA, Clyde DE, Losonsky G, et al. Safety and immunogenicity
in man of a synthetic peptide malaria vaccine against Plasmodium fal
ciparum sporozoites. Nature 1987;328:257-9.
34. Etlinger HM, Gillessen D, Lahm HW, Matile H, Schonfeld HJ, Trzeciak
A. Use of prior vaccinations for the development of new vaccines.
Science 1990;249:423-5.
35. Sad S, Rao K, Arora R, Talwar GP, Raghupathy R. Bypass of carrier
induced epitope-specific suppression using aT-helper epitope. Immu
nology 1992; 76:599-603.
36. Kumar A, Arora R, Kaur P, Chauhan VS, Sharma P. "Universal" T helper
cell determinants enhance immunogenicity of a Plasmodium falciparum
merozoite surface antigen peptide. J Immuno11992; 148:1499-505.
37. Valmori D, Pessi A, Bianchi E, Corradin G. Use of human universally
antigenic tetanus toxin T cell epitopes as carriers for human vaccination.
J Immuno11992; 149:717 -21.
38. Kaurnaya PT, Kobs-Conrad S, Seo YH, et al. Peptide vaccines incorporat
ing a "promiscuous" T-cell epitope bypass certain haplotype restricted
immune responses and provide broad spectrum immunogenicity. J Mol
Recognit 1993;6:81-94.
39. Yu Z, Healy F, Valmori D, Escobar P, Corradin G, Mach JP. Peptide
antibody conjugates for tumour therapy: a MHC-class-II-restricted
tetanus toxin peptide coupled to an anti-Ig light chain antibody can
induce cytotoxic lysis of a human B-celllymphoma by specific CD4 T
cells. Int J Cancer 1994; 56:244-8.
40. O'Sullivan D, Arrhenius T, Sidney J, et al. On the interaction ofpromiscu
ous antigenic peptides with different DR alleles. J Immunol1991; 147:
2663-9.
41. Manfredi AA, Yuen MH, Moiola L, Protti MP, Conti-Tronconi BM. Hu
man acetylcholine receptor presentation in myasthenia gravis: DR re
striction of autoimmune T epitopes and binding of synthetic receptor
sequences to DR molecules. J Immuno11994; 152:4165-74.
42. Yuen MR, Macklin K, Conti-Fine BM. MHC class II presentation of
human acetylcholine receptor in myasthenia gravis. Binding of synthetic
'Y subunit sequences to purified DR molecules. J Autommun 1996; 9:
67-77.
43. Choe S, Bennett MJ, Fujii G, et al. The crystal structure of diphtheria
toxin. Nature 1992;357:216-22.
44. Demotz S, Barbey C, Corradin G, et al. The set of naturally processed
peptides displayed by DR molecules is tuned by polymorphism of res i
due 86. Eur J ImmunoI1993;23:425-32.
Dow
nloaded from https://academ
ic.oup.com/jid/article/175/2/382/816687 by guest on 04 July 2022




















