
Neuronal Targeting of Cardiotrophin-1 by Coupling with Tetanus Toxin C Fragment
13
Neuronal Targeting of Cardiotrophin-1 by Coupling with Tetanus Toxin C Fragment T. Bordet,* L. Castelnau-Ptakhine,* F. Fauchereau,* G. Friocourt,* A. Kahn,* and G. Haase ²,1 *INSERM U.129, Institut Cochin de Ge ´ne ´tique Mole ´culaire, 24, Rue du Faubourg St Jacques, 75014 Paris, France; and ² INSERM U.382, Institut de Biologie du De ´veloppement de Marseille, Campus de Luminy, Case 907, 13288 Marseille, France Cardiotrophin-1 (CT-1) is a potent neurotrophic factor for motoneurons but its clinical use in motor neuron diseases is precluded by side effects on the heart and liver. We explored the possibility of targeting CT-1 to neurons by coupling with the tetanus toxin fragment TTC. Genetic fusion proteins between CT-1 or GFP and TTC were pro- duced in Escherichia coli and assayed in vitro. In contrast to uncoupled CT-1 or GFP, TTC-coupled proteins bound with high affinity to cerebral neurons and spinal cord motoneurons and were rapidly internalized. Glia, hepato- cytes, or cardiomyocytes did not show detectable binding or uptake of TTC-coupled proteins. Similar to CT-1, TTC- coupled CT-1 induced IL-6 secretion by KB cells, acti- vated Reg-2 gene expression, and promoted motoneuron survival in a dose-dependent manner. In vivo studies will test whether TTC-coupled CT-1 might be targeted to de- generating spinal cord or brain-stem motoneurons and migrate trans-synaptically to cortical motoneurons, which are also affected in amyotrophic lateral sclerosis. INTRODUCTION The IL-6-related cytokines leukemia inhibitory factor (LIF), ciliary neurotrophic factor (CNTF), and cardiotro- phin-1 (CT-1) increase motoneuron survival in primary culture and after peripheral nerve axotomy in neonates (Arakawa et al., 1990; Martinou et al., 1992; Pennica et al., 1996; Sendtner et al., 1990). Their therapeutic use in motor neuron disease treatment has been proposed and they have been tested in murine models of motor neu- ron disease. LIF and CNTF have protective effects in wobbler mice (Mitsumoto et al., 1994; Ikeda et al., 1995), and we and others have shown that CNTF and CT-1, when delivered by cellular or viral gene transfer, have strong neuroprotective effects in the pmn (progressive motor neuronopathy) mouse (Sendtner et al., 1992; Sagot et al., 1995; Bordet et al., 1999; Haase et al., 1999). However, the action of cytokines is not restricted to neurons. Indeed, these proteins signal via the ubiqui- tous LIFRb/gp130 receptor complex and the Jak/STAT signaling pathway (for review see Heinrich et al., 1998). Consequently, they share many biological activities on hematopoietic cells, embryonic stem cells, and hepato- cytes. Thus, chronic administration of cytokines to an- imals induces pleiotropic effects including weight loss (Henderson et al., 1994; Bordet et al., 1999; Haase et al., 1999) and synthesis of acute-phase proteins (Dittrich et al., 1994; Jin et al., 1996). Similar side effects have also been described after administration of recombinant CNTF protein in amyotrophic lateral sclerosis patients (Cedarbaum et al., 1995; Miller et al., 1996). Further- more, administration of CT-1, a protein initially charac- terized for its hypertrophic effect on cardiomyocytes, induces cardiac hypertrophy in adult mice (Jin et al., 1996; Bordet et al., 1999). These results suggested that neuroprotective effects of cytokines might be counter- balanced by their systemic toxicity. In order to increase the specificity of CT-1 for neurons we have decided to couple it with a neurotropic carrier. As carrier, we choose TTC, the C-terminal fragment of the tetanus toxin heavy chain. Tetanus toxin (TeNT), a potent neurotoxin produced by the anaerobic bacterium Clostridium tetani, causes spastic paralysis by blocking the neurotransmitter re- lease from spinal cord interneurons (for review see Schiavo et al., 2000). TeNT has a well-documented ca- pacity for neuronal binding and internalization (Critch- 1 To whom correspondence and reprint requests should be ad- dressed. Fax: 133 4 91 26 97 57. E-mail: [email protected]. doi:10.1006/mcne.2001.0979, available online at http://www.idealibrary.com on Molecular and Cellular Neuroscience 17, 842– 854 (2001) MCN 1044-7431/01 $35.00 Copyright © 2001 by Academic Press All rights of reproduction in any form reserved. 842
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Neuronal Targeting of Cardiotrophin-1 by Coupling with Tetanus Toxin C Fragment

Jd
stgma
d
doi:10.1006/mcne.2001.0979, available online at http://www.idealibrary.com on
Molecular and Cellular Neuroscience 17, 842–854 (2001)MCN
Neuronal Targeting of Cardiotrophin-1 byCoupling with Tetanus Toxin C FragmentT. Bordet,* L. Castelnau-Ptakhine,* F. Fauchereau,*G. Friocourt,* A. Kahn,* and G. Haase†,1
*INSERM U.129, Institut Cochin de Genetique Moleculaire, 24, Rue du Faubourg Stacques, 75014 Paris, France; and †INSERM U.382, Institut de Biologie du Developpemente Marseille, Campus de Luminy, Case 907, 13288 Marseille, France
awsmSHnt
mti1nbtcct
bsl
Cardiotrophin-1 (CT-1) is a potent neurotrophic factor formotoneurons but its clinical use in motor neuron diseasesis precluded by side effects on the heart and liver. Weexplored the possibility of targeting CT-1 to neurons bycoupling with the tetanus toxin fragment TTC. Geneticfusion proteins between CT-1 or GFP and TTC were pro-duced in Escherichia coli and assayed in vitro. In contrastto uncoupled CT-1 or GFP, TTC-coupled proteins boundwith high affinity to cerebral neurons and spinal cordmotoneurons and were rapidly internalized. Glia, hepato-cytes, or cardiomyocytes did not show detectable bindingor uptake of TTC-coupled proteins. Similar to CT-1, TTC-coupled CT-1 induced IL-6 secretion by KB cells, acti-vated Reg-2 gene expression, and promoted motoneuronurvival in a dose-dependent manner. In vivo studies willest whether TTC-coupled CT-1 might be targeted to de-enerating spinal cord or brain-stem motoneurons andigrate trans-synaptically to cortical motoneurons, which
re also affected in amyotrophic lateral sclerosis.
INTRODUCTION
The IL-6-related cytokines leukemia inhibitory factor(LIF), ciliary neurotrophic factor (CNTF), and cardiotro-phin-1 (CT-1) increase motoneuron survival in primaryculture and after peripheral nerve axotomy in neonates(Arakawa et al., 1990; Martinou et al., 1992; Pennica et al.,1996; Sendtner et al., 1990). Their therapeutic use inmotor neuron disease treatment has been proposed andthey have been tested in murine models of motor neu-ron disease. LIF and CNTF have protective effects inwobbler mice (Mitsumoto et al., 1994; Ikeda et al., 1995),
Sp
1 To whom correspondence and reprint requests should be ad-ressed. Fax: 133 4 91 26 97 57. E-mail: [email protected].
842
nd we and others have shown that CNTF and CT-1,hen delivered by cellular or viral gene transfer, have
trong neuroprotective effects in the pmn (progressiveotor neuronopathy) mouse (Sendtner et al., 1992;
agot et al., 1995; Bordet et al., 1999; Haase et al., 1999).owever, the action of cytokines is not restricted toeurons. Indeed, these proteins signal via the ubiqui-
ous LIFRb/gp130 receptor complex and the Jak/STATsignaling pathway (for review see Heinrich et al., 1998).Consequently, they share many biological activities onhematopoietic cells, embryonic stem cells, and hepato-cytes. Thus, chronic administration of cytokines to an-imals induces pleiotropic effects including weight loss(Henderson et al., 1994; Bordet et al., 1999; Haase et al.,1999) and synthesis of acute-phase proteins (Dittrich etal., 1994; Jin et al., 1996). Similar side effects have alsobeen described after administration of recombinantCNTF protein in amyotrophic lateral sclerosis patients(Cedarbaum et al., 1995; Miller et al., 1996). Further-
ore, administration of CT-1, a protein initially charac-erized for its hypertrophic effect on cardiomyocytes,nduces cardiac hypertrophy in adult mice (Jin et al.,996; Bordet et al., 1999). These results suggested thateuroprotective effects of cytokines might be counter-alanced by their systemic toxicity. In order to increasehe specificity of CT-1 for neurons we have decided toouple it with a neurotropic carrier. As carrier, wehoose TTC, the C-terminal fragment of the tetanusoxin heavy chain.
Tetanus toxin (TeNT), a potent neurotoxin producedy the anaerobic bacterium Clostridium tetani, causespastic paralysis by blocking the neurotransmitter re-ease from spinal cord interneurons (for review see
chiavo et al., 2000). TeNT has a well-documented ca-acity for neuronal binding and internalization (Critch-1044-7431/01 $35.00Copyright © 2001 by Academic Press
All rights of reproduction in any form reserved.

sized
843Neuronal Targeting of Cardiotrophin-1
ley et al., 1985; Rogers and Snyder, 1981). When admin-istered to animals, the toxin is taken up selectively bynerve endings at the neuromuscular junction and gainsaccess to the CNS via retrograde axonal transportwithin motoneurons and trans-synaptic migration tointerneurons (Price et al., 1975; Schwab et al., 1979). The150-kDa TeNT protein is composed of a heavy chainwhich mediates binding and retrograde transport and alight chain which is responsible for most of the toxicity.The TTC fragment, corresponding to the carboxy-termi-nal 451-amino-acid fragment of the heavy chain, retainsthe neuronal binding and uptake properties of the ho-lotoxin but is nontoxic (Bizzini et al., 1977; Evinger andErichsen, 1986; Fishman and Carrigan, 1987; Manning etal., 1990; Morris et al., 1980; Simpson, 1985). Thus it hasbeen proposed that TTC could be used to target and
FIG. 1. Western blot analysis of TTC-coupled proteins. (A) Schemaheavy (H) chain, the C-terminal fragment of tetanus toxin heavy chafusion proteins. (B–D) Western blot of TTC-coupled CT-1 (B, C) or Gprotein preparations with TTC in the C-terminal position (i.e., GGST-TTC/CT-1 and GST-TTC/GFP both contained a single smallerprotein was loaded; molecular weight standards are indicated.
deliver proteins to motoneurons (Bizzini et al., 1980).Chemical conjugation of large proteins with TTC has
indeed been shown to enhance their uptake by neuronsin tissue culture (Dobrenis et al., 1992) and also afterintramuscular or systemic injection into animals(Beaude et al., 1990; Fishman and Savitt, 1989; Fishmanet al., 1990). More recently, several proteins have beengenetically fused to TTC: superoxide dismutase-1(SOD-1) (Francis et al., 1995), b-galactosidase (Coen etal., 1997), and Diphtheria toxin (Francis et al., 2000). Thefusion proteins were internalized by neuronal cells andremained enzymatically active.
We therefore hypothesized that fusion of the neuro-trophic factor cardiotrophin-1 to TTC might increase itsneuronal binding while leaving intact its genuine func-tion as a survival-promoting protein for neurons. As afirst step to test this hypothesis, we have constructedrecombinant cardiotrophin-1/TTC and also GFP (green
presentation of the tetanus neurotoxin (TeNT) with its light (L) andTC), and the TTC-coupled CT-1 and GFP proteins produced as GST) revealed with anti-CT-1 (B) and anti-TTC (C, D) antibodies. WhileT-1/TTC and GST-GFP/TTC) were all full size, preparations offragment of 78 kDa apparent molecular weight. Per lane, 50 ng of
tic rein (TFP (DST-C
fluorescent protein)/TTC fusion proteins and investi-gated their biological properties by using various bind-

e aloCT-1
844 Bordet et al.
ing, internalization, signaling, and neuronal survivalassays.
RESULTS
Expression of TTC-Coupled Proteins
Hybrid proteins of cardiotrophin-1 and the C-ter-minal fragment of tetanus toxin were produced asgenetic fusions. Because in these fusion proteins, theTTC domain might affect the correct folding or thebiological properties of CT-1 and vice versa, two
FIG. 2. In vitro cell surface binding of TTC-coupled proteins in primneuronal cells were incubated for 1 h at 0°C with 5 mg/ml TTC-coupC), or GFP (D, E) immunostaining when exposed to, respectively, TPunctate fluorescent signals were detected on the plasma membrancontacts. No labeling was observed after incubation with uncoupled
types of fusion proteins were designed with TTCplaced in either N- or C-terminal position of CT-1
(Fig. 1A). As a control, fusion proteins between GFPand TTC and also uncoupled CT-1, GFP, and TTCproteins were engineered. We previously showedthat fully biologically active CT-1 could be producedin Escherichia coli (Bordet et al., 1999). Recombinantproteins were therefore expressed in E. coli as solubleGST fusion proteins and purified on affinity glutathi-one columns. When analyzed by Western blot, puri-fied GST-CT-1/TTC and GST-TTC/CT-1 proteinswere stained by both anti-CT-1 and anti-TTC anti-bodies at the expected size (100 kDa; Figs. 1B–1D).Protein preparations of the C-terminal constructs
ultures of embryonic mouse brains. Three days after plating, primaryroteins and fixed. Neuronal cells displayed strong TTC (A), CT-1 (B,A), CT-1/TTC (B), TTC/CT-1 (C), GFP/TTC (D), or TTC/GFP (E).ng both cell bodies and neurites, with strong labeling at synaptic(F) or GFP (not shown). Scale bar, 15 mm.
ary cled pTC (
(GST-CT-1/TTC and GST-GFP/TTC) and also of theuncoupled constructs were free of degradation prod-
(tTdti ere ins
845Neuronal Targeting of Cardiotrophin-1
ucts. Protein preparations of GST-TTC/CT-1 andGST-TTC/GFP contained not only full-length formsbut also a single smaller fragment of 78 kDa (Figs. 1Cand 1D) resulting probably from a premature stopduring protein synthesis or from protein degrada-tion. Most of the following experiments were there-fore done with the CT-1/TTC and GFP/TTC proteinsharboring TTC in C-terminal position. The GST do-mains were proteolytically cleaved and the resulting
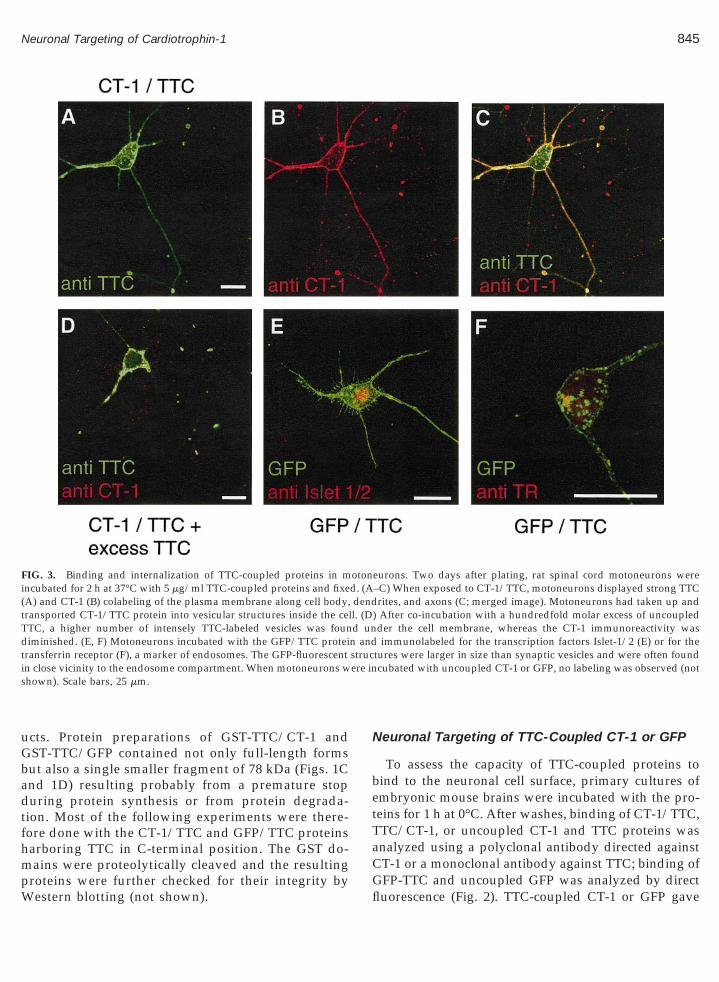
FIG. 3. Binding and internalization of TTC-coupled proteins in mincubated for 2 h at 37°C with 5 mg/ml TTC-coupled proteins and fixeA) and CT-1 (B) colabeling of the plasma membrane along cell body,ransported CT-1/TTC protein into vesicular structures inside the ceTC, a higher number of intensely TTC-labeled vesicles was founiminished. (E, F) Motoneurons incubated with the GFP/TTC protei
ransferrin receptor (F), a marker of endosomes. The GFP-fluorescentn close vicinity to the endosome compartment. When motoneurons whown). Scale bars, 25 mm.
proteins were further checked for their integrity byWestern blotting (not shown).
Neuronal Targeting of TTC-Coupled CT-1 or GFP
To assess the capacity of TTC-coupled proteins tobind to the neuronal cell surface, primary cultures ofembryonic mouse brains were incubated with the pro-teins for 1 h at 0°C. After washes, binding of CT-1/TTC,TTC/CT-1, or uncoupled CT-1 and TTC proteins wasanalyzed using a polyclonal antibody directed againstCT-1 or a monoclonal antibody against TTC; binding of
eurons. Two days after plating, rat spinal cord motoneurons were–C) When exposed to CT-1/TTC, motoneurons displayed strong TTCrites, and axons (C; merged image). Motoneurons had taken up and
) After co-incubation with a hundredfold molar excess of uncoupledder the cell membrane, whereas the CT-1 immunoreactivity wasimmunolabeled for the transcription factors Islet-1/2 (E) or for the
tures were larger in size than synaptic vesicles and were often foundcubated with uncoupled CT-1 or GFP, no labeling was observed (not
otond. (Adend
ll. (Dd un
n andstruc
GFP-TTC and uncoupled GFP was analyzed by directfluorescence (Fig. 2). TTC-coupled CT-1 or GFP gave

fdaNGcbabc
swThCRc
846 Bordet et al.
rise to labeling profiles very similar to that of uncou-pled TTC: punctate fluorescent signals were detected onthe plasma membrane along both cell bodies and pro-cesses (Figs. 2A–2E). No differences were observed withrespect to the N- or C-terminal position of the TTCfragment in the TTC-coupled proteins. Uncoupled CT-1(Fig. 2F) or GFP (not shown) did not give rise to asignal.
To study the uptake of the TTC-coupled proteins,cultures of embryonic rat spinal cord motoneuronswere incubated for 2 h at 37°C with CT-1/TTC orGFP/TTC and then analyzed by confocal microscopy(Fig. 3). When double immunolabeling was performedwith antibodies against CT-1 and TTC, all motoneuronsexposed to the CT-1/TTC protein exhibited a strongfluorescence signal not only on the plasma membranebut also in the cytoplasm. Within axons, dendrites, andperikarya, vesicular structures were strongly labeled.The CT-1 and TTC labeling profiles were almost iden-tical (Figs. 3A–3C), suggesting that the CT-1/TTC pro-tein had not been degraded following internalization.When motoneurons were incubated with 5 mg/ml CT-1/TTC and a hundredfold molar excess of uncoupledTTC, the intensity of the TTC fluorescence and thenumber of internalization vesicles were much in-creased, whereas the CT-1 fluorescence was reduced(Fig. 3D). Motoneurons incubated with GFP/TTC (Figs.3E and 3F) displayed a labeling profile very similar tothat of motoneurons exposed to CT-1/TTC, whereasthose exposed to uncoupled GFP or CT-1 showed nosignal (not shown). Together, these findings indicatethat the TTC-coupled proteins had been internalizedthrough interaction with the TTC receptor. Severaltypes of neurons, e.g., hippocampal neurons, take upTTC into synaptic vesicles (Matteoli et al., 1996; Verde-rio et al., 1999). In our spinal motoneuron cultures how-ever, the TTC-coupled proteins were enriched in struc-tures which were larger in size than synaptic vesicles(Figs. 3E and 3F) and not immunoreactive for the syn-aptic vesicle marker synaptotagmin or the a-adaptinsubunit of the adaptor complex AP2 (not shown). Afraction of the large GFP-positive internalization struc-tures could be detected adjacent to endosomes whichwere identified by transferrin receptor immunostaining(Fig. 3F).
By immunofluorescence studies we next examinedwhether binding and internalization of TTC-coupledproteins occurred specifically in neurons. We chose tostudy in particular hepatocytes and cardiomyocytesbecause these two cell types express the tripartite
CT-1 receptor, possess the intracellular CT-1 signal-ing machinery, and are responsible for CT-1-inducedFr
toxic side effects such as secretion of acute-phaseproteins from liver or cardiac hypertrophy. Primaryhepatocytes were obtained by in vivo collagenase per-fusion of mice and cultured for 2 days. As shown inFig. 4A, hepatocytes incubated for 2 h at 37°C with 5mg/ml GFP/TTC did not bind or take up the coupledprotein. Cardiomyocytes were differentiated in vitrorom embryoid bodies and identified as bona fide car-iomyocytes by spontaneous rhythmic contractionsnd positive desmin immunostaining (not shown).o uptake or internalization of CT-1/TTC (Fig. 4B) orFP/TTC (not shown) could be observed in these
ultures. In contrast, CT-1/TTC fusion proteinsound efficiently to neurons in mixed brain culturess shown by coimmunostaining of CT-1 and dou-lecortin, a microtubule-associated protein specifi-ally expressed in neurons (Fig. 4D) (Francis et al.,
1999). In these cultures, GFAP-positive glial cells didnot bind or internalize GFP/TTC or CT-1/TTC (Fig.4C and data not shown).
Signaling Activity and Survival Responses Inducedby TTC-Coupled CT1
The functionality of the CT-1 domain in the TTC-coupled proteins was first tested in the KB epidermoidcancer cell line. KB cells express approximately 5000binding sites for the tripartite CT-1 receptor, bind CT-1with very high affinity, and, following CT-1 receptoractivation, secrete interleukin-6 (IL-6) in a time- anddose-dependent manner (Robledo et al., 1997a,b). Toassess the ability of the TTC-coupled CT-1 proteins toactivate the CT-1 receptor, KB cells were cultured in thepresence of increasing concentrations of CT-1/TTC andTTC/CT-1 proteins. After 48 h, cell supernatants wereharvested and the amount of IL-6 was determined byELISA. As shown in Fig. 5, CT-1/TTC, and with slightlylower potency also TTC/CT-1, induced the secretion ofIL-6 in a dose-dependent manner similar to that ofuncoupled CT-1. In contrast, no IL-6 synthesis was de-tected after exposure of KB cells to uncoupled TTC.
In motoneurons, CT-1 receptor activation and signal-ing leads to induction of the Reg-2 (PAP-1) gene (Live-ey et al., 1997; Nishimune et al., 2000). Motoneuronsere therefore cultured for 24 h in the presence ofTC-coupled CT-1 proteins or control proteins and thenarvested for RT-PCR analysis. As shown in Fig. 6A,T-1/TTC, TTC/CT-1, and uncoupled CT-1 inducedeg-2 gene expression, whereas uncoupled TTC or glial-ell-derived neurotrophic factor (GDNF) had no effect.
inally, we analyzed the survival response of motoneu-ons cultured for 3 days in the presence of varying
stsh
1Ctmwtca
mtm(s
hvmwcssa
TnadCCau
847Neuronal Targeting of Cardiotrophin-1
concentrations of TTC-coupled or uncoupled proteins.In these experiments, GST fusion proteins were usedinstead of thrombin-cleaved proteins because evennanomolar concentrations of thrombin can induce apo-ptotic death of motoneurons in culture (Turgeon et al.,1998) thereby precluding survival analysis. Figure 6Bdemonstrates that GST-CT-1/TTC enhanced the sur-vival of motoneurons in a dose-dependent manner, inthe same concentration range as GST-CT-1 or a com-mercial CT-1 preparation. Preparations of GST-TTC orcommercial TTC had a minor but statistically not sig-nificant effect on motoneuron survival.
DISCUSSION
We previously showed that CT-1 gene transfer inmice suffering from progressive motor neuronopathypromotes a long-term efficient slowdown of nerve de-generation but has only a modest effect in prolonginglife span (Bordet et al., 1999). This discrepancy wasurprising since results obtained with the administra-ion of other neurotrophic factors in pmn mice have
FIG. 5. Production of IL-6 by KB cells after incubation with TTC-coupled CT-1. Following CT-1 receptor activation, KB epidermoidcancer cells secrete interleukin-6 (IL-6). The KB cells were incubatedwith increasing concentrations of TTC-coupled proteins or controlproteins and IL-6 synthesis was measured 48 h later in supernatantsby ELISA. Supernatants from KB cells exposed to CT-1/TTC or touncoupled CT-1 induced a similar dose-dependent increase in IL-6synthesis; supernatants from KB cells exposed to TTC/CT-1 in theconcentration range 0.1 to 10 ng/ml tended to contain less IL-6. NoIL-6 immunoreactivity was found in TTC- or mock-incubated KB cellsupernatants. Mean results of quadruplicate wells are shown.
hown a correlation between nerve protection and en-anced life span (Sagot et al., 1995; Haase et al., 1997,
br
999). Thus we speculated that the beneficial effects ofT-1 on axonal protection were counterbalanced by its
oxic side effects. A more targeted delivery of CT-1 tootoneurons might therefore be advantageous. Heree report the construction and biological characteriza-
ion of a fusion protein, CT-1/TTC, between mouseardiotrophin-1 and the C-terminal fragment of the tet-nus toxin heavy chain.We first analyzed the functionality of the TTC do-ain of the fusion proteins by investigating their ability
o bind specifically to neuronal surfaces. The TTC do-ain mediates the binding of the tetanus holotoxin
Morris et al., 1980) with its neuronal receptor, whicheems to be made up of gangliosides GD1b or GT1b (for
review see Halpern and Neale, 1995) and an as yetill-characterized protein of 15 kDa molecular weight(Herreros et al., 2000). In our immunofluorescence ex-periments, we found that the binding of TTC-coupledCT-1 proteins was specific to neurons and was verysimilar to that observed with uncoupled TTC. Puncta ofimmunoreactivity were detected all along perikaryaand neuronal processes with strong staining at synapticcontacts. We found that glial cells, hepatocytes, andcardiomyocytes did not bind or internalize these pro-teins, whereas motoneurons and cerebral neurons wereable to take up TTC-coupled proteins at a high rate.These observations indicate that the TTC-coupled pro-teins have conserved the neurotropism of TTC. Usingdouble immunolabeling and confocal analysis, TTC-coupled CT-1 or GFP was detected in vesicular struc-tures located in axons, dendrites, and perikarya. Inhippocampal neurons, these structures were identifiedas synaptic vesicles (Matteoli et al., 1996). In our study,
owever, these structures were larger than synapticesicles, were not immunoreactive for synaptic vesiclearkers, and could eventually be found in close contactith endosomes. These observations suggest that TTC-
oupled proteins do not enter spinal motoneurons byynaptic vesicle endocytosis or that they rapidly exitynaptic vesicles and are en route toward structuresdjacent to the endosome.In the past, several proteins have been coupled to
TC and retained their activity when bound and inter-alized into neurons. All these proteins were enzymesnd not signaling proteins. We therefore studied inetail the functional properties of the TTC-coupledT-1 molecules. In particular, we demonstrated thatT-1 coupled C-terminally to TTC (CT-1/TTC) wasble to induce IL-6 secretion in KB cells as efficiently asncoupled CT-1. The TTC/CT-1 preparation tended to
e slightly less efficient in the lower concentrationange, which might be due to the presence of a trun-
wf ltured rnali
848 Bordet et al.
cated form. By using cultures of highly purified spinalcord motoneurons, we were able to show that TTC-coupled CT-1 proteins induced the up-regulation of theCT-1-responsive gene Reg-2. Finally, TTC-coupled CT-1was as efficient as CT-1 to promote motoneuron cellsurvival. Thus, interaction of the TTC domain with itsreceptor on motoneurons does not preclude CT-1 recep-tor activation and survival signaling. It can be specu-lated that the same CT1/TTC molecule can interactwith both CT1 and TTC receptors at a given moment.
FIG. 4. Absence of detectable binding or uptake of TTC-coupled protestudied by incubating various cell cultures for 2 h at 37°C with 5 mg/ml
ere not labeled after incubation with GFP/TTC (A) or CT-1/TTC (B). Hor desmin (not shown) and b-tubulin (B). (C, D) In mixed cerebral cuoublecortin immunolabeling (D). Neurons but not glia bound and inte
The similar ranges of motoneuron survival observedwith uncoupled CT-1 and TTC-coupled CT-1 suggest
that this is the case. For a definitive answer, detailedquantitative experiments with the TTC-coupled CT-1proteins will be required in order to determine their onand off rates on motoneurons.
This study suggests that cytokines might be targeted toneurons by coupling with TTC. Future in vivo applicationswill be based on the construction of gene therapy vectorsaimed toward continuous production of CT-1/TTC pro-teins in different organs. When produced in skeletal mus-cle, the high affinity of TTC for motoneurons and its rapid
nonneuronal cells. The neuronal tropism of TTC-coupled proteins wascoupled proteins. (A, B) Primary hepatocytes (A) or cardiomyocytes (B)ocytes were counterstained with DAPI; cardiomyocytes stained positives, glia were identified by GFAP immunolabeling (C) and neurons byzed TTC-coupled CT-1 or GFP. Scale bars, 15 mm.
ins inTTC-epat
uptake should allow a specific targeting of the TTC-cou-pled cytokine to the corresponding motor pool. Recently,

si
tes n
849Neuronal Targeting of Cardiotrophin-1
intramuscular injection of a LacZ/TTC plasmid in Xeno-pus tadpoles was shown to drive the in vivo synthesis ofthe fusion protein and its uptake by connected motoneu-rons (Coen et al., 1999). Alternatively, it might be advan-tageous to release TTC-coupled cytokines into the blood-stream because neurons with projections outside theblood–brain barrier can rapidly take up systemically ad-ministered TTC and transport it to their cell bodies in theCNS (Fishman and Carrigan, 1988). Finally, the property
FIG. 6. Increased Reg-2 gene expression and survival in motoneuropinal cord motoneuron cultures 24 h after addition of the proteinsnduced the expression of the CT-1-responsive gene Reg-2, whereas u
Control reactions were performed using primers specific for b-actin; oa Reg-2 cDNA plasmid as template; 2, RT-PCR control without RT.2 days in the presence of the indicated concentration (in ng/ml) ofsimilar to that of uncoupled GST-CT-1 or recombinant CT-1. The effecat the optimal concentration of GST-CT-1 was set as 100%; NB indicaof counts in quadruplicate wells.
of TTC to migrate trans-synaptically might be exploited todeliver TTC-coupled cytokines to upper (cortical) mo-
toneurons, which are also affected in amyotrophic lateralsclerosis (ALS) (Fig. 7). Indeed, after intramuscular injec-tion of TTC-coupled proteins, Coen et al. (1997) andFigueiredo et al. (1997) demonstrated retrograde axonaltransport of TTC-coupled proteins in lower motoneuronsand trans-synaptic transfer to connected higher order neu-rons.
Not only neurotrophic factors but also other thera-peutic candidates for motor neuron diseases such as
ltured in the presence of TTC-coupled CT-1. (A) RT-PCR analysis ofonstrates that CT-1/TTC, TTC/CT-1, and uncoupled CT-1 proteinspled TTC or the unrelated neurotrophic factor GDNF had no effect.lanes, NB, neurobasal medium without trophic factors; 1, PCR withrvival analysis of motoneurons (MNs) which had been cultured for
roteins. GST-CT-1/TTC promoted the survival of MNs to an extentGST-TTC or recombinant TTC were not significant. Survival of MNs
eurobasal medium without trophic factors; error bars represent SEM
ns cudemncouther
(B) Suthe pts of
antiapoptotic proteins and chaperones might be cou-pled to TTC. The antiapoptotic protein Bcl-2, for exam-

am2trkt(
dtTfv
ptp
Gp
tPTf
T
prafm
850 Bordet et al.
ple, can reduce motoneuron degeneration in transgenicmutant SOD1 mice (Kostic et al., 1997; Azzouz et al.,2000), and SMN (survival motor neuron) transgenes canttenuate disease severity in mouse models of spinaluscular atrophy (Hsieh-Li et al., 2000; Monani et al.,
000). TTC coupling might also be used to modify theropism of gene therapy vectors. A retargeted adenovi-us vector, engineered by coupling TTC via an anti-nob antibody to the virus surface, was recently used toransduce motoneurons after intramuscular injectionSchneider et al., 2000).
In conclusion, many strategies to treat motor neuroniseases will require the targeting of the therapeutic agent
o the nervous system. The remarkable properties of theTC fragment might be useful for targeting neurotrophic
actors, other therapeutically relevant proteins, or evenirus vectors to degenerating motoneurons.
EXPERIMENTAL METHODS
TTC Fusion Constructs
Full-length TTC DNA (GenBank AF154828, nucleotides1 to 1356) was generated by PCR from the genomic DNAof the C. tetani strain (a gift from M. Popoff, PasteurInstitute). Two different PCR products were synthesizedowing to the C- or N-terminal position of TTC. For C-terminal fusion proteins, a BglII site in 59 and a SalI site in39 were introduced by PCR (59 GAAGATCTTATTCTA-AAAATCTGGATTGTTGGGT 39; 59 GCGTCGACTCTA-GATTTAATCATTTGTCCATCCTTTCATCTGTA 39). ForN-terminal fusion proteins, an initiation codon was intro-duced with a NcoI site and the TTC stop codon wasconverted in a BspHI site to be cloned in phase with thetransgene cDNA (59 T G C C A T G G A A A A T C T GG A T T G T T G G G T T G A T A 39; 59 TGTCATGAAAT-CATTTGTCCATCCTTCATCT 39). A pGEX-TTC con-struct was obtained by cloning the TTC NcoI/SalI frag-ment into pGEX-4T3 (Amersham Pharmacia).
The pGEX-CT-1 plasmid was obtained by insertingthe murine CT-1 cDNA in the pGEX vector digested byEcoRI and SalI (Bordet et al., 1999). To construct pGEX-CT-1/TTC, a BglII site was first introduced by PCR inpGEX-CT-1 in place of the stop codon of CT-1 cDNAand the TTC BglII/SalI fragment was then inserted. The
GEX-TTC/CT-1 plasmid was obtained by insertinghe TTC NcoI/BspHI sequence into NcoI-linearizedGEX-CT-1.The pGEX-GFP sequence was obtained by cloning the
FP cDNA from the pEGFP-C1 vector (Clontech) intoGEX-4T-3. pGEX-GFP/TTC was obtained by insertion of
he TTC BglII/SalI fragment into pGEX-GFP modified byCR to replace the GFP stop codon by a BglII site. pGEX-TC/GFP was obtained by inserting the TTC NcoI/BspHI
ragment into NcoI-linerarized pGEX-GFP.
TC Fusion Protein Production and Purification
In order to express GST fusion proteins, the pGEXlasmids were transformed into E. coli. Overnight bacte-ial cultures were diluted 1:100 in LB medium containingmpicillin and grown for 1 h at 37°C. Protein expressionrom the Ptac promoter was induced by the addition of 1
M isopropyl-b-d-thiogalactoside and 1 mM MgCl2. Af-ter 5 h incubation at 32°C, the bacteria were pelleted bycentrifugation for 20 min at 3000 rpm and washed withPBS. Cells were lysed for 1 h at 4°C in PBS containing 1%(v/v) Triton X-100, 1% (v/v) NP-40, 1 mg/ml lysosyme,and anti-proteases (Complete; Boehringer) and furtherdisrupted by sonication. The bacterial lysates were ultra-centrifuged at 30,000g for 10 min and the supernatantsincubated overnight at 4°C with affinity matrix gluta-thione–Sepharose B. Beads were collected on column andextensively washed with PBS and 1 M NaCl. Column-bound proteins were eluted by addition of 5 mM reducedglutathione in 50 mM Tris–HCl, pH 8, 0.3 M NaCl, fol-lowed by extensive dialysis against PBS and 5 to 10%(v/v) glycerol at 4°C. Some bound proteins were incu-bated with thrombin (10 units/ml; Sigma) in order to becleaved from the GST sequence and thereby eluted fromthe affinity column. Purified proteins were quantitated bya spectroscopic Coomassie assay (Pierce) and analyzed bySDS/PAGE. Western blots were reacted with primaryantibodies (goat anti-CT-1 (R&D Systems), dilution 1:500;monoclonal anti-TTC (Boehringer), dilution 1:200), sec-ondary antibodies (HRP-conjugated anti-goat and anti-mouse (DAKO), dilution 1:2,000), and the ECL-detectionkit (Amersham).
Cell Culture Methods
Primary neuronal cells were prepared from fetal mousebrain at stage E15. Brains were dissociated by treatmentwith trypsin (0.25% for 15 min at room temperature),followed by trituration with a fire-polished Pasteur pi-pette. Dissociated cells were plated on poly-l-lysine-treated glass coverslips at a density of 25,000 cells/cm2
and cultured in Dulbecco’s modified Eagle’s medium(DMEM; Life Technologies) supplemented with 2.5%(v/v) fetal calf serum and 2.5% (v/v) horse serum (LifeTechnologies). After 24 h, the culture medium was re-
placed by DMEM supplemented with 2% (v/v) B-27 (LifeTechnologies) and 2.5% (v/v) horse serum. Cultures en-
n
tBa
actawdbt12
851Neuronal Targeting of Cardiotrophin-1
riched in glial cells were obtained by adding 10% (v/v)fetal calf serum (FCS) to the culture medium. Hepatocyteswere isolated from C57Bl/6 adult mice by in vivo collage-
ase perfusion (Bergot et al., 1992). Cardiomyocytes wereobtained by first aggregating mouse ES cells into embry-oid bodies using the “hanging drop” method essentiallyas described by Maltsev et al. (1994). Embryoid bodieswere then cultured in four-well dishes on gelatin-treatedglass coverslips until spontaneous rhythmic contractionscould be monitored in foci. Motoneurons (MNs) werepurified from embryonic (E14) Sprague–Dawley rats, us-ing the 192 antibody that recognizes the low-affinity neu-rotrophin receptor p75NTR and following the proceduredescribed by Arce et al. (1999). The MNs were plated onpoly-l-ornithine- and laminin-coated coverslips and cul-tured in “complete” neurobasal medium (Neurobasal;Life Technologies) supplemented with 0.5 mM l-glu-tamine, 25 mM l-glutamate, 25 mM b-mercaptoethanol,2% (v/v) horse serum, and 2% B27 to which 1 ng/mlBNDF was added.
Binding and Internalization Assays ofTTC-Coupled Proteins
Primary cultures were used for experiments after 3days in vitro. Binding and internalization of TTC fusionproteins were assessed using a modified protocol (Fran-cis et al., 1995). In binding experiments cells were incu-bated for 1 h at 0°C with 5 mg/ml recombinant TTC-coupled proteins or control proteins; in internalizationexperiments cells were incubated for 2 h at 37°C withproteins diluted in 20 mM Tris–HCl, pH 8, 1 mM CaCl2,1 mM MgCl2, 0.25% BSA in PBS. Following removal ofhe proteins, cells were extensively washed with PBS.rain cultures were then fixed with methanol for 7 mint 220°C; hepatocytes, cardiomyocytes, or MNs were
fixed with 3.7% (w/v) formaldehyde in PBS for 30 minat room temperature. GFP activity was detected onfixed cells by direct fluorescence.
For immunohistochemical staining, brain cultureswere incubated in 2% (w/v) BSA and 0.02% (v/v) Tri-ton-X100 in Tris-buffered saline for 1 h at room temper-ture. Other cell types were incubated for 10 min in PBSontaining 50 mM lysine and 0.1% Triton X-100 andhen blocked in 2% (w/v) BSA, 2% (v/v) donkey serum,nd 2% (v/v) goat serum in PBS. Primary antibodiesere used for 1 h at room temperature at the followingilutions: anti-cardiotrophin-1 (R&D), 1:200; anti-dou-lecortin pep (Francis et al., 1999), 1:300; anti-synapto-agmin I (Chemicon), 1:200; anti-GFAP (Chemicon),
:200; anti-TTC (Fitzgerald), 1:200, anti-Islet1/2 mAbsD6 and 4D5 (Hybridoma Bank), 1:100, anti-b-tubulinAb-1 (Oncogene), 1:400; anti-desmin mAb D-8281(Sigma), 1:20; anti-transferrin receptor mAb B3/25(Odorizzi and Trowbridge, 1997), 1:500. After severalwashes, cells were incubated for 1 h with secondaryantibodies conjugated with fluorescein–isothiocyanate,Texas red, Cy3, or Cy5 (Jackson, West Grove, PA). Aftermounting, cells were analyzed with a Bio-Rad MRC-1024 or a Zeiss LSM 400 confocal laser microscope.Images were obtained in frame with filters set at 515–525 or 515–565 nm (FITC, GFP), 590 nm (Texas red,Cy3), or 590–810 nm (Cy5).
CT-1 Signaling and Neuronal Survival Assays
Cytokine bioassays were performed on the humanKB epidermoid cell line (ECACC No. 94050408) accord-ing to previously described protocol (Robledo et al.,1997a). Briefly, human KB epidermoid cells were platedin quadruplicate in 24-well plates at a concentration of2 3 105 cells/well and cultured for 48 h in DMEMcontaining 10% (v/v) FCS. After two rinses in DMEM,cells were incubated in 500 ml of culture medium con-taining serial dilutions of tested proteins ranging from0.01 to 100 ng/ml. After 48 h, supernatants were har-vested and their IL-6 content was determined by thehuman IL-6 ELISA Cytoscreen system (BIOSOURCEInternational, Camarillo, CA).
Reg-2 gene activation in motoneurons in response toTTC-coupled CT-1 was analyzed by reverse transcrip-tion PCR. MNs were seeded at a density of 15,000 cellson poly-l-ornithine- and laminin-coated 35-mm plasticdishes (Nunc) and cultured for 24 h in the presence of 5mg/ml CT-1/TTC, TTC/CT-1, or uncoupled CT-1 orTTC or 100 pg/ml GDNF. Cells were harvested, RNAsextracted with Trizol (Gibco), and cDNAs synthesizedwith Superscript II reverse transcriptase (Life Technol-ogies). PCR primers specific for Reg-2 were located inReg-2 exon 3 (59 CCGAATTCAGGAGAAGACTCTC-CGAAGA) and exon 6 (59 GGGTCGACTTAACCTG-TAAATTTGCAGA). PCRs were also performed onReg-2 plasmid DNA or using primers specific for b-ac-tin (Haase et al., 1999).
For survival analysis, motoneurons were seeded infour-well coated plastic dishes (f 15.5 mm; Nunc) at adensity of 1,000 cells per dish and cultured in completeneurobasal medium containing different concentrations ofGST-CT-1/TTC, GST-CT-1, GST-TTC, TTC (purchasedfrom Boehringer), or murine CT-1 (purchased from R&D).After 2 days in culture, motoneuron survival was quanti-
fied in quadruplicate wells by counting the number ofphase-bright neurons with long axonal processes.
ationper
852 Bordet et al.
ACKNOWLEDGMENTS
We thank Dr. M. Popoff (Institut Pasteur, Paris, France) for provid-ing genomic DNA from Clostridium tetani, members of the INSERMU.129 metabolism group for hepatocyte cultures, Dr. L. Miquerol(Marseille) for cardiomyocyte cultures, Drs. F. Francis (Paris) and I.Trowbridge (La Jolla) for antibodies against doublecortin and thetransferrin receptor. Some confocal images were obtained with thehelp of Mrs. I. Bouchaert (ICGM, Paris). We are indebted to Dr. C.
FIG. 7. Therapeutic strategy to prevent in vivo motoneuron degenreleased by skeletal muscle or peripheral organs, are expected to bindthis study, interaction of TTC-coupled CT-1 with the CT-1 receptorpathways and interaction with the TTC receptor leads to internalizmigration of TTC-coupled cytokines might allow their delivery to up
Henderson (Marseille) for critical comments on the manuscript andDrs. A. Le Bivic (Marseille) and T. Galli (Paris) for helpful discussions.
T.B. is a fellow of the Association Francaise contre les Myopathies(AFM). This work was supported by grants from INSERM, AFM, andFederation pour la Recherche Medicale.
REFERENCES
on by using TTC-coupled cytokines. TTC-coupled cytokines, whenh high affinity to nerve endings of lower motoneurons. As shown ininal cord motoneurons activates intracellular CT-1-specific survivalof the coupled protein. Subsequent retrograde and trans-synaptic
motoneurons, which are also affected in ALS.
eratiwit
of sp
Arakawa, Y., Sendtner, M., and Thoenen, H. (1990). Survival effect ofciliary neurotrophic factor (CNTF) on chick embryonic motoneu-

B
B
B
C
C
C
C
D
D
E
F
F
F
F
F
F
F
F
H
H
H
H
H
H
H
I
J
K
853Neuronal Targeting of Cardiotrophin-1
rons in culture: Comparison with other neurotrophic factors andcytokines. J. Neurosci. 10: 3507–3515.
Arce, V., Garces, A., de Bovis, B., Filippi, P., Henderson, C. E., Pett-mann, B., and deLapeyriere, O. (1999). Cardiotrophin-1 requiresLIFRb to promote survival of mouse motoneurons purified by anovel technique. J. Neurosci. Res. 55: 119–126.
Azzouz, M., Hottinger, A. F., Paterna, J.-C., Zurn, A. D., Aebischer, P.,and Bueler, H. (2000). Increased motoneuron survival and im-proved neuromuscular function in transgenic ALS mice after in-traspinal injection of an adeno-associated virus encoding Bcl-2.Hum. Mol. Genet. 9: 803–811.
Beaude, P., Delacour, A., Bizzini, B., Domuado, D., and Remy, M.-H.(1990). Retrograde axonal transport of an exogenous enzyme co-valently linked to B-IIb fragment of tetanus toxin. Biochem. J. 271:87–91.
Bergot, M. O., Diaz-Guerra, M. J., Puzenat, N., Raymondjean, M., andKahn, A. (1992). Cis-regulation of the L-type pyruvate kinase genepromoter by glucose, insulin and cyclic AMP. Nucleic Acids Res. 20:1871–1877.
izzini, B., Stoeckel, K., and Schwab, M. (1977). An antigenic polypep-tide fragment isolated from tetanus toxin: Chemical characteriza-tion, binding to gangliosides and retrograde axonal transport invarious neuron systems. J. Neurochem. 28: 529–542.
izzini, B., Grob, P., Glicksman, M. A., and Akert, K. (1980). Use of theB-IIb tetanus toxin derived fragment as a specific neuropharmaco-logical transport agent. Brain Res. 193: 221–227.
ordet, T., Schmalbruch, H., Pettmann, B., Hagege, A., Castelnau-Ptakhine, L., Kahn, A., and Haase, G. (1999). Adenoviral cardiotro-phin-1 gene transfer protects pmn mice from progressive motorneuronopathy. J. Clin. Invest. 104: 1077–1085.
edarbaum, J. M., Chapman, C., Charatan, M., Stambler, N., An-drews, L., Zhan, C., Radka, S., Morrisey, D., Lakings, D., Brooks,B. R., Sanjak, M., Pestronk, A., Florence, J., Mitsumoto, H., Szirony,K., Bittle, L., Ringel, S., Neville, H., and Brinkmann, J. (1995). Thepharmacokinetics of subcutaneously administered recombinant hu-man ciliary neurotrophic factor (rHCNTF) in patients with amyo-trophic lateral sclerosis: Relation to parameters of the acute-phaseresponse, by The ALS CNTF Treatment Study (ACTS) Phase I–IIStudy Group. Clin. Neuropharmacol. 18: 500–514.
oen, L., Osta, R., Maury, M., and Brulet, P. (1997). Construction ofhybrid proteins that migrate retrogradely and transynaptically intothe central nervous system. Proc. Natl. Acad. Sci. USA 94: 9400–9405.
oen, L., Kissa, K., le Mevel, S., Brulet, P., and Demeneix, B. A. (1999).A somatic gene transfer approach using recombinant fusion pro-teins to map muscle–motoneuron projections in Xenopus spinalcord. Int. J. Dev. Biol. 43: 823–830.
ritchley, D. R., Nelson, P. G., Habig, W. H., and Fishman, P. H.(1985). Fate of tetanus toxin bound to the surface of primary neu-rons in culture: Evidence for rapid internalization. J. Cell. Biol. 100:1499–1507.ittrich, F., Thoenen, H., and Sendtner, M. (1994). Ciliary neurotro-phic factor: Pharmacokinetics and acute-phase response in rat. Ann.Neurol. 35: 151–163.obrenis, K., Joseph, A., and Rattazzi, M. C. (1992). Neuronal lyso-somal enzyme replacement using fragment C of tetanus toxin. Proc.Natl. Acad. Sci. USA 89: 2297–2301.
vinger, C., and Erichsen, J. T. (1986). Transsynaptic retrograde trans-port of fragment C of tetanus toxin demonstrated by immunohis-tochemical localization. Brain Res. 380: 383–388.
igueiredo, D. M., Hallewell, R. A., Chen, L. L., Fairweather, N. F.,
Dougan, G., Savitt, J. M., Parks, D. A., and Fishman, P. S. (1997).Delivery of recombinant tetanus–superoxide dismutase proteins tocentral nervous system neurons by retrograde axonal transport.Exp. Neurol. 145: 546–554.
ishman, P. S., and Carrigan, D. R. (1987). Retrograde transneuronaltransfer of the C-fragment of tetanus toxin. Brain Res. 406: 275–279.
ishman, P. S., and Carrigan, D. R. (1988). Motoneuron uptake fromthe circulation of the binding fragment of tetanus toxin. Arch.Neurol. 45: 558–561.
ishman, P. S., and Savitt, J. M. (1989). Transsynaptic transfer ofretrogradely transported tetanus protein–peroxidase conjugates.Exp. Neurol. 106: 197–203.
ishman, P. S., Savitt, J. M., and Farrand, D. A. (1990). Enhanced CNSuptake of systemically administered proteins through conjugationwith tetanus C-fragment. J. Neurol. Sci. 98: 311–325.
rancis, F., Koulakoff, A., Boucher, D., Chafey, P., Schaar, B., Vinet,M. C., Friocourt, G., McDonnell, N., Reiner, O., Kahn, A., McCon-nell, S. K., Berwald-Netter, Y., Denoulet, P., and Chelly, J. (1999).Doublecortin is a developmentally regulated, microtubule-associ-ated protein expressed in migrating and differentiating neurons.Neuron 23: 247–256.
rancis, J. W., Hosler, B. A., Brown, R. H., and Fishman, P. S. (1995).CuZn superoxide dismutase (SOD-1): Tetanus toxin fragment Chybrid protein for targeted delivery of SOD-1 to neuronal cells.J. Biol. Chem. 270: 15434–15442.
rancis, J. W., Brown, R. H., Figueiredo, D., Remington, M. P.,Castillo, O., Schwarzschild, M. A., Fishman, P. S., Murphy, J. R.,and vanderSpek, J. C. (2000). Enhancement of diphteria toxin po-tency by replacement of the receptor binding domain with tetanustoxin C-fragment: A potential vector for delivering heterologousproteins to neurons. J. Neurochem. 74: 2528–2536.aase, G., Kennel, P., Pettmann, B., Vigne, E., Akli, S., Revah, F.,Schmalbruch, H., and Kahn, A. (1997). Gene therapy of murinemotor neuron disease using adenoviral vectors for neurotrophicfactors. Nat. Med. 3: 429–436.aase, G., Pettmann, B., Bordet, T., Villa, P., Vigne, E., Schmalbruch,H., and Kahn, A. (1999). Therapeutic benefit of ciliary neurotrophicfactor in progressive motor neuronopathy depends on the route ofdelivery. Ann. Neurol. 45: 296–304.alpern, J. L., and Neale, E. A. (1995). Neurospecific binding, inter-nalization, and retrograde axonal transport. Curr. Top. Microbiol.Immunol. 195: 221–241.einrich, P. C., Behrmann, I., Muller-Newen, G., Scharper, F., andGraeve, L. (1998). Interleukin-6-type cytokine signalling throughthe gp130/Jak/STAT pathway. Biochem. J. 334: 297–314.enderson, J. T., Seniuk, N. A., Richardson, P. M., Gauldie, J., andRoder, J. C. (1994). Systemic administration of ciliary neurotrophicfactor induces cachexia in rodents. J. Clin. Invest. 93: 2632–2638.erreros, J., Lalli, G., Montecucco, C., and Schiavo, G. (2000). Tetanustoxin fragment C binds to a protein present in neuronal cell linesand motoneurons. J. Neurochem. 74: 1941–1950.sieh-Li, H. M., Chang, J. G., Jong, Y. J., Wu, M. H., Wang, N. M.,Tsai, C. H., and Li, H. (2000). A mouse model for spinal muscularatrophy. Nat. Genet. 24: 66–70.
keda, K., Iwasaki, Y., Tagaya, N., Shiojima, T., and Kinoshita, M.(1995). Neuroprotective effect of cholinergic differentiation factor/leukemia inhibitory factor on wobbler murine motor neuron dis-ease. Muscle Nerve 18: 1344–1347.
in, H., Yang, R., Keller, G. A., Ryan, A., Ko, A., Finkle, D., Swanson,T. A., Li, W., Pennica, D., Wood, W. I., and Paoni, N. F. (1996). Invivo effects of cardiotrophin-1. Cytokine 8: 920–926.
ostic, V., Jackson-Lewis, V., de Bilbao, F., Dubois-Dauphin, M., and
Przedborski, S. (1997). Bcl-2: Prolonging life in a transgenic mousemodel of familial amyotrophic lateral sclerosis. Science 277: 559–562.
M
M
M
M
N
O
P
P
R
R
R
S
S
S
S
S
S
S
T
V
854 Bordet et al.
Livesey, F. J., O’Brien, J. A., Li, M., Smith, A. G., Murphy, L. J., andHunt, S. P. (1997). A Schwann cell mitogen accompanying regen-eration of motor neurons. Nature 390: 614–618.
Maltsev, V. A., Wobus, A. M., Rohwedel, J., Bader, M., and Hescheler,J. (1994). Cardiomyocytes differentiated in vitro from embryonicstem cells developmentally express cardiac-specific genes and ioniccurrents. Circ. Res. 75: 233–244.
Manning, K. A., Erichsen, J. T., and Evinger, C. (1990). Retrogradetransneuronal transport properties of fragment C of tetanus toxin.Neuroscience 34: 251–263.
Martinou, J. C., Martinou, I., and Kato, A. C. (1992). Cholinergicdifferentiation factor (CDF/LIF) promotes survival of isolated ratembryonic motoneurons in vitro. Neuron 8: 737–744.
Matteoli, M., Verderio, C., Rossetto, O., Iezzi, N., Coco, S., Schiavo, G.,and Montecucco, C. (1996). Synaptic vesicle endocytosis mediatesthe entry of tetanus neurotoxin into hippocampal neurons. Proc.Natl. Acad. Sci. USA 93: 13310–13315.iller, R. G., Bryan, W. W., Dietz, M. A., Munsat, T. L., Petajan, J. H.,Smith, S. A., and Goodpasture, J. C. (1996). Toxicity and tolerabilityof recombinant human ciliary neurotrophic factor in patients withamyotrophic lateral sclerosis. Neurology 47: 1329–1331.itsumoto, H., Ikeda, K., Klinkosz, B., Cedarbaum, J. M., Wong, V.,and Lindsay, R. M. (1994). The effects of ciliary neurotrophic factoron motor neuron dysfunction in wobbler mouse motor neurondisease. Ann. Neurol. 36: 142–148.onani, U. R., Sendtner, M., Coovert, D. D., Parsons, D. W., Andre-assi, C., Le, T. T., Jablonka, S., Schrank, B., Rossol, W., Prior, T. W.,Morris, G. E., and Burghes, A. H. M. (2000). The human centromericsurvival motor neuron gene (SMN2) rescues embryonic lethality inSmn2/2 mice and results in a mouse with spinal muscular atro-phy. Hum. Mol. Genet. 9: 333–339.orris, N. P., Consiglio, E., and Kohn, L. D. (1980). Interaction offragment B and C of tetanus toxin with neural and thyroid mem-branes and with gangliosides. J. Biol. Chem. 255: 6071–6076.ishimune, H., Vasseur, S., Wiese, S., Birling, M. C., Holtmann, B.,Sendtner, M., Iovanna, J. L., and Henderson, C. E. (2000). Reg-2 is amotoneuron neurotrophic factor and a signalling intermediate inthe CNTF survival pathway. Nat. Cell Biol. 2: 906–914.dorizzi, G., and Trowbridge, I. S. (1997). Structural requirements forbasolateral sorting of the human transferrin receptor in the biosyn-thetic and endocytic pathways of Madin–Darby canine kidney cells.J. Cell Biol. 137: 1255–1264.
ennica, D., Arce, V., Swanson, T. A., Vejsada, R., Pollock, R. A.,Armanini, M., Dudley, K., Phillips, H. S., Rosenthal, A., Kato, A. C.,and Henderson, C. E. (1996). Cardiotrophin-1, a cytokine present in
embryonic muscle, supports long-term survival of spinal motoneu-rons. Neuron 17: 63–74.rice, D. L., Griffin, J., Young, A., Peck, K., and Stocks, A. (1975).Tetanus toxin: Direct evidence for retrograde intraaxonal transport.Science 188: 945–947.
obledo, O., Chevalier, S., Froger, J., Barthelaix-Pouplard, A., Pen-nica, D., and Gascan, H. (1997a). Regulation of interleukin 6 expres-sion by cardiotrophin 1. Cytokine 9: 666–671.
obledo, O., Fourcin, M., Chevalier, S., Guillet, C., Auguste, P., Pou-plard-Barthelaix, A., Pennica, D., and Gascan, H. (1997b). Signalingof the cardiotrophin-1 receptor, evidence for a third receptor com-ponent. J. Biol. Chem. 272: 4855–4863.
ogers, T. B., and Snyder, S. H. (1981). High affinity binding oftetanus toxin to mammalian brain membranes. J. Biol. Chem. 256:2402–2407.
agot, Y., Tan, S. A., Baetge, E., Schmalbruch, H., Kato, A. C., andAebischer, P. (1995). Polymer encapsulated cell lines geneticallyengineered to release ciliary neurotrophic factor can slow downprogressive motor neuronopathy in the mouse. Eur. J. Neurosci. 7:1313–1322.
chiavo, G., Matteoli, M., and Montecucco, C. (2000). Neurotoxinsaffecting neuroexocytosis. Physiol. Rev. 80: 717–766.
chneider, H., Groves, M., Muhle, C., Reynolds, P. N., Knight, A.,Groves, M. T., Scaravilli, F., Curiel, D. T., Fairweather, N. F., andCoutelle, C. (2000). Re-targeting of adenoviral vectors to neuronsusing the Hc fragment of tetanus toxin. Gene Ther. 7: 1584–1592.
chwab, M. E., Suda, K., and Thoenen, H. (1979). Selective retrogradetranssynaptic transfer of a protein, tetanus toxin, subsequent to itsretrograde axonal transport. J. Cell. Biol. 82: 789–810.
endtner, M., Kreutzberg, G. W., and Thoenen, H. (1990). Ciliaryneurotrophic factor prevents the degeneration of motor neuronsafter axotomy. Nature 345: 440–441.
endtner, M., Schmalbruch, H., Stockli, K. A., Carroll, P., Kreutzberg,G. W., and Thoenen, H. (1992). Ciliary neurotrophic factor preventsdegeneration of motor neurons in mouse mutant progressive motorneuronopathy. Nature 358: 502–504.
impson, L. L. (1985). Pharmacological experiments on the bindingand internalization of the 50,000 dalton carboxyterminus of tetanustoxin at the cholinergic neuromuscular junction. J. Pharmacol. Exp.Ther. 234: 100–105.
urgeon, V. L., Lloyd, E. D., Wang, S., Festoff, B. W., and Houenou,L. J. (1998). Thrombin perturbs neurite outgrowth and inducesapoptotic cell death in enriched chick spinal motoneuron culturesthrough caspase activation. J. Neurosci. 18: 6882–6891.
erderio, C., Coco, S., Bacci, A., Rossetto, O., De Camilli, P., Mon-tecucco, C., and Matteoli, M. (1999). Tetanus toxin blocks the exo-
cytosis of synaptic vesicles clustered at synapses but not of synapticvesicles in isolated axons. J. Neurosci. 19: 6723–6732.Received December 15, 2000Revised February 19, 2001
Accepted February 26, 2001




















