
Differential Shh, Bmp and Wnt gene expressions during craniofacial development in mice
Upload
independentCategory
view
0download
0
www.landesbioscience.com Epigenetics 343
Epigenetics 5:4, 343-351; May 16, 2010; © 2010 Landes Bioscience
REsEaRch papER REsEaRch papER
*Correspondence to: Carmen Jerónimo; Email: [email protected]: 12/10/09; Accepted: 03/10/10Previously published online: www.landesbioscience.com/journals/epigenetics/article/11749
Introduction
Cancer is a major public health concern in the Western world, with an increasing incidence rate that indicates it will continue to be a significant health burden in the future.1 For the year 2009 in the USA it is estimated that 321,020 new cases of prostate, bladder and renal carcinomas (the most common types of genito-urinary cancer) will be diagnosed (21.7% of all new cancer cases) and will account for 9.7% of all cancer-related deaths.1 Biologic characterization of those tumors is likely to have a strong impact on public health, allowing implementation of new early detection strategies and development of more efficient therapies.
The “classical,” also called canonical, Wingless-type (Wnt) signalling pathway is one of the most relevant pathways involved in normal cell growth and differentiation.2 Furthermore, this pathway plays a central role in tumorigenesis as its inappropriate
Epigenetic regulation of Wnt signaling pathway in urological cancer
Vera L. costa,1,3 Rui henrique,1,4,6 Franclim R. Ribeiro,2,3 João R. carvalho,1,3 Jorge Oliveira,5 Francisco Lobo,5 Manuel R. Teixeira2,3,6 and carmen Jerónimo1,3,6,*
1cancer Epigenetics Group; and 2cancer Genetics Group; Research center of the portuguese Oncology Institute at porto; porto, portugal; Departments of 3Genetics; 4pathology; and 5Urology; portuguese Oncology Institute at porto; Rua Dr. antónio Bernardino de almeida; porto, portugal; 6Department of pathology and Molecular
Immunology; (IcBas) Institute of Biomedical sciences abel salazar; University of porto; porto, portugal
Key words: promoter methylation, bladder cancer, renal cell tumors, prostate cancer, wnt-signaling pathway
Abbreviations: ACTB, beta actin; APC, adenomatous polyposis coli; BCa, primary transitional cell carcinoma of bladder; CDH1, E-cadherin (epithelial); CTNNB1, catenin (cadherin-associated protein), beta 1; HPRT1, hypoxanthine; NBM, normal bladder mucosa; NPT, normal prostate tissue; PCa, prostate carcinoma; QMSP, quantitative methylation-specific PCR; RCT, renal cell
tumors; SFRP1, secreted frizzled-related protein 1; WIF1, WNT inhibitory factor 1
activation is a common feature of many human cancers, lead-ing to deregulation of cell proliferation and differentiation.3 In normal conditions, CTNNB1 is controlled by the upstream regu-lators of the Wnt pathway.2 Those regulators (or antagonists), comprise extracellular inhibitors, membrane/cytosolic elements and nuclear components.4 Two major events implicated in aber-rant constitutive Wnt signaling pathway activation are genetic and epigenetic alterations.4 The ultimate consequence of these is the accumulation of β-catenin, which is translocated to the nucleus, causing inappropriate transcription of a large set of genes that will lead to neoplastic transformation through deregulation of cell proliferation and differentiation.3
Some of the Wnt pathway antagonists are epigenetically dereg-ulated in several cancer models4 and many studies attempted to characterize the role of epigenetic alterations of the Wnt path-way in several human cancers.4-11 That group of genes includes
constitutive activation of the Wnt signaling pathway is a common feature of solid tumors and contributes to uncontrolled cell-growth and impaired differentiation. We hypothesized that gene silencing mediated through aberrant promoter methylation of upstream Wnt antagonist genes might result in β-catenin accumulation, resulting in constitutive Wnt activation. Wnt antagonist genes (SFRP1, WIF1, APC and CDH1) and CTNNB1 promoter methylation was examined in genomic DNa extracted from 12 urological cancer cell lines and correlated with CTNNB1 mRNa expression. promoter methylation status was then assessed in 36 Bca, 30 pca, 31 RcT, and normal bladder mucosa (15), prostate (10) and renal (5) tissue samples. Finally, CTNNB1 mRNa relative expression levels were correlated with Wnt antagonist gene methylation status in RcT.
Methylation was found in at least one Wnt antagonist gene and the CTNNB1 promoter was unmethylated in all cancer cell lines tested. When gene methylation levels were compared between cancer cell lines with high and low CTNNB1 mRNa expression, a trend was found for increased CDH1 promoter methylation levels in the former. Bca and pca tumors demonstrated high frequency of promoter methylation at all tested genes. In RcT, CTNNB1 was unmethylated in all cases and the overall frequency of promoter methylation at the remainder genes was lower. Interestingly, median CTNNB1 mRNa expression levels were significantly higher in RcTs methylated in at least one Wnt antagonist gene promoter.
We concluded that epigenetic deregulation of Wnt pathway inhibitors may contribute to aberrant activation of Wnt signaling pathway in bladder, prostate and renal tumors.
344 Epigenetics Volume 5 Issue 4
Results
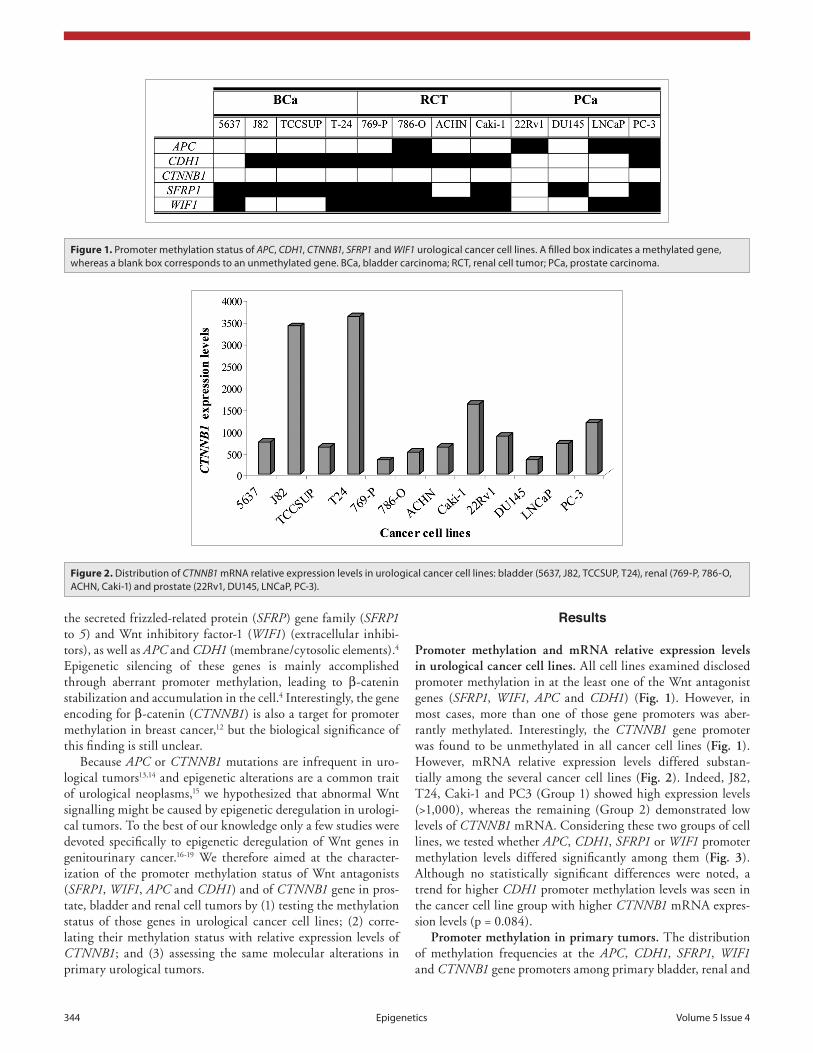
Promoter methylation and mRNA relative expression levels in urological cancer cell lines. All cell lines examined disclosed promoter methylation in at the least one of the Wnt antagonist genes (SFRP1, WIF1, APC and CDH1) (Fig. 1). However, in most cases, more than one of those gene promoters was aber-rantly methylated. Interestingly, the CTNNB1 gene promoter was found to be unmethylated in all cancer cell lines (Fig. 1). However, mRNA relative expression levels differed substan-tially among the several cancer cell lines (Fig. 2). Indeed, J82, T24, Caki-1 and PC3 (Group 1) showed high expression levels (>1,000), whereas the remaining (Group 2) demonstrated low levels of CTNNB1 mRNA. Considering these two groups of cell lines, we tested whether APC, CDH1, SFRP1 or WIF1 promoter methylation levels differed significantly among them (Fig. 3). Although no statistically significant differences were noted, a trend for higher CDH1 promoter methylation levels was seen in the cancer cell line group with higher CTNNB1 mRNA expres-sion levels (p = 0.084).
Promoter methylation in primary tumors. The distribution of methylation frequencies at the APC, CDH1, SFRP1, WIF1 and CTNNB1 gene promoters among primary bladder, renal and
the secreted frizzled-related protein (SFRP) gene family (SFRP1 to 5) and Wnt inhibitory factor-1 (WIF1) (extracellular inhibi-tors), as well as APC and CDH1 (membrane/cytosolic elements).4 Epigenetic silencing of these genes is mainly accomplished through aberrant promoter methylation, leading to β-catenin stabilization and accumulation in the cell.4 Interestingly, the gene encoding for β-catenin (CTNNB1) is also a target for promoter methylation in breast cancer,12 but the biological significance of this finding is still unclear.
Because APC or CTNNB1 mutations are infrequent in uro-logical tumors13,14 and epigenetic alterations are a common trait of urological neoplasms,15 we hypothesized that abnormal Wnt signalling might be caused by epigenetic deregulation in urologi-cal tumors. To the best of our knowledge only a few studies were devoted specifically to epigenetic deregulation of Wnt genes in genitourinary cancer.16-19 We therefore aimed at the character-ization of the promoter methylation status of Wnt antagonists (SFRP1, WIF1, APC and CDH1) and of CTNNB1 gene in pros-tate, bladder and renal cell tumors by (1) testing the methylation status of those genes in urological cancer cell lines; (2) corre-lating their methylation status with relative expression levels of CTNNB1; and (3) assessing the same molecular alterations in primary urological tumors.
Figure 1. promoter methylation status of APC, CDH1, CTNNB1, SFRP1 and WIF1 urological cancer cell lines. a filled box indicates a methylated gene, whereas a blank box corresponds to an unmethylated gene. Bca, bladder carcinoma; RcT, renal cell tumor; pca, prostate carcinoma.
Figure 2. Distribution of CTNNB1 mRNa relative expression levels in urological cancer cell lines: bladder (5637, J82, TccsUp, T24), renal (769-p, 786-O, achN, caki-1) and prostate (22Rv1, DU145, LNcap, pc-3).

www.landesbioscience.com Epigenetics 345
tumors methylated in at the least one of those genes (Table 1). Interestingly, median CTNNB1 mRNA relative expression levels were significantly higher in the latter group (p = 0.01).
prostate tumors is depicted in Figure 4. Bladder and prostate cancers showed high frequency of promoter methylation at the various loci, includ-ing CTNNB1 in most cases (69.4% and 73.3%, respectively). In addition, more than one of the Wnt pathway antagonist genes was found to be methylated in the majority of cases. Remarkably, no CTNNB1 promoter methylation was detected in any of the renal tumors. Moreover, the overall frequency of promoter methylation at the remain-ing genes was lower than that of prostate and bladder carcinomas. Indeed, 41.9% of the renal tumors examined were unmethylated at all gene promoters tested.
The distribution of gene promoter methylation levels in tissue samples is displayed in Figure 5. CTNNB1 promoter methylation levels were sig-nificantly higher in bladder and prostate cancer compared to their normal tissues (p < 0.001 for both), but not in renal tumors. Although this was the only significant difference found for bladder tissues, prostate carcinomas also showed signifi-cantly higher levels of APC promoter methylation (p < 0.001). Interestingly, higher levels of SFRP1 promoter methylation were found in normal pros-tate tissue compared to prostate carcinoma (p < 0.001).
CTNNB1 expression and Wnt antagonist gene promoter methylation in renal tumors. Since renal cell tumors were all unmethylated at the CTNNB1 promoter (thus removing a possible interference of direct epigenetic regulation of this gene), we com-pared CTNNB1 mRNA levels among renal tumors unmethy-lated at the APC, CDH1, SFRP1 and WIF1 gene promoters and
Figure 4. Distribution of promoter methylation of APC, CDH1, CTNNB1, SFRP1 and WIF1 in primary bladder cancers (B, [a]), renal cell tumors (R, [B]) and prostate cancers (p, [c]). a filled box indicates a methylated gene, whereas a blank box corresponds to an unmethylated gene. Methylation was consid-ered when the ratio was equal to, or higher than, the median value across the respective normal samples.
Figure 3. Distribution of promoter methylation levels (target gene/ACTB x 1000) of APC, CDH1, CTNNB1, SFRP1 and WIF1 among bladder, kidney and prostate cancer cell lines. Group 1 includes J82, T-24, caki-1, pc-3, which displayed high (>1,000) CTNNB1 mRNa relative expression levels, whereas Group 2 refers to the remaining cell lines, which displayed low expression levels.

346 Epigenetics Volume 5 Issue 4
tumors, as well as cancer cell lines, allowing for a comparative analysis of epigenetic regulation of the canonical Wnt pathway in the three most common urological tumors.
The 12 urological cancer cell lines tested in this study were unmethylated at the CTNNB1 promoter, although relative mRNA expression levels differed markedly among them. These findings emphasize the pivotal regulatory role of the upstream Wnt pathway antagonist genes in this setting. Thus, constitutive Wnt pathway activation might be hypothetically accomplished by epigenetic silencing of those genes. Indeed, we found pro-moter methylation in at least one of the four genes tested in all cell lines, and in most cases (10 out of 12) at the least two of the antagonist genes were methylated. It was then expected that can-cer lines with higher CTNNB1 mRNA expression levels would display higher promoter methylation levels at Wnt antagonist genes. Median SFRP1, WIF1 and CDH1 promoter methylation levels were indeed higher in cell lines expressing higher levels of CTNNB1 mRNA, but no statistically significant differences were depicted, at the least partially, due to the small number of cancer cell lines in each group. Because we did not test all genes involved
Discussion
Abnormal Wnt signaling pathway, resulting in uncontrolled cell proliferation and impaired differentiation, is a common event in human cancer. Mutations in APC or CTNNB1 genes (which are mutually exclusive) are one of the most frequent causes of consti-tutive Wnt pathway activation in some tumors (e.g., colo-rectal cancer). However, in urological tumors APC or CTNNB1 muta-tions are infrequent.13,14 Because promoter methylation serves as an alternative mechanism for gene silencing,4 we hypothesized that abnormal Wnt signaling in the most common urological tumors might be caused by epigenetic deregulation. In fact, we have previously found that APC promoter methylation is com-mon in PCa.20
Overall, our results are in agreement with previously published reports on APC, CDH1, SFRP1 and WIF1 promoter methylation in bladder,17,21,22 kidney18,19 and prostate cancer,16 notwithstand-ing differences in methodology, sample size and population. However, to the best of our knowledge, our study is the first to assess simultaneously bladder, prostate and kidney primary
Figure 5A. Distribution of promoter methylation levels (target gene/ACTB x 1000) of APC, CDH1, CTNNB1, SFRP1 and WIF1 among bladder tissue samples.

www.landesbioscience.com Epigenetics 347
in Wnt signaling (e.g., those involved in non-canonical Wnt activation), genetic or epigenetic alterations occurring at other gene loci might also be implicated in CTNNB1 deregulation.
Interestingly, the findings in urological cancer cell lines did not translate faith-fully to primary bladder and prostate carcinomas. Indeed, CTNNB1 promoter methylation was detected in most of those primary cancers and methylation levels were statistically significantly higher in tumors compared to normal tissues pro-cured from non-cancerous bladders and prostates. This apparently paradoxical finding has been recently documented also in breast cancer12 and its biological signifi-cance remains unclear. Theoretically, can-cer cells methylated at the CTNNB1 gene might be less effective in constitutionally activating the Wnt signaling pathway, although the concomitant underexpression of the Wnt antagonist genes might over-come this effect. Remarkably, in our study, both bladder and prostate carcinomas pre-sented higher frequency of methylation at the APC and CDH1 promoters than renal tumors, which were all unmethylated at the CTNNB1 promoter. The association of APC and CDH1 promoter methylation has also been demonstrated in lobular breast cancer, although no accumulation of β-catenin was found in this tumor type.23 Because we did not determine CTNNB1 mRNA expression in primary prostate and bladder carcinomas, it is not possible to ascertain the net effect of promoter methy-lation at CTNNB1 and/or Wnt antagonist genes in those primary cancers.
Contrarily to most bladder and pros-tate carcinomas, the methylation status of CTNNB1 of renal cancer cell lines was con-sistent with that of primary tumors. This finding granted us the opportunity to test in vivo the effect of epigenetic regulation of upstream Wnt antagonist on CTNNB1 expression. As postulated, renal tumors with methylation in at least one of the Wnt pathway gene promoters did show a signifi-cantly increased CTNNB1 mRNA expres-sion. Thus, these findings provide indirect
Figure 5B and C. Distribution of promoter methylation levels (target gene/ACTB x 1000) of APC, CDH1, CTNNB1, SFRP1 and WIF1 among renal (B) and prostate (c) tissue samples.

348 Epigenetics Volume 5 Issue 4
the alteration of more than one inhibitor gene in the pathway to effectively lead to increased β-catenin, and consequent con-stitutive activation of the signaling pathway. Additional studies are needed to validate this hypothesis.
Because the CTNNB1 gene is located at chromosome 3p21, an area which is frequently deleted (approximately 50% of cases) in clear cell renal cell carcinoma, the most common type of RCT, our findings concerning CTNNB1 mRNA expres-sion and its correlation with promoter methylation might be affected by this result. Although we did not test our cases for 3p21 deletion, we believe that this feature is unlikely to have influenced our results. Indeed, the group of 17 RCTs designated as “Methylated” in Table 1, includes 13 clear cell renal cell car-cinomas (76% of cases), whereas these represent only 33% of cases (4) from the “Unmethylated” group. If CTNNB1 monoal-lelic deletion was indeed relevant for its downregulation, then an increase in CTNNB1 mRNA levels in the “Methylated” group would not be observed, as it was the case in this study.
Although methylation-induced silencing has been con-firmed as the most likely mechanism responsible for loss of SFRP1 expression in several cancer types,24 we found higher SFRP1 promoter methylation levels in normal prostate tis-sue compared to prostate carcinoma. Two previous studies on SFRP1 promoter methylation in PCa provided quite dissimi-lar results. Whereas Florl and co-workers found that less than 5% of PCa were methylated at the SFRP1 promoter,25 Lodygin et al. reported that more than 80% of PCa from their series were methylated,26 although the same set of primers was used in both studies. Our results are well in accordance with the former study (2 out of 30 cases showed SFRP1 promoter methylation in our series) and both the primers and probe were designed for the same region. Thus, methodological or populational differ-ences should account for the discrepant frequency reported by Lodygin and co-workers.26 These authors did not found SFRP1 methylation in non-cancerous prostate tissues, but they only tested benign prostatic hyperplasias (from the central area of the prostate) and not morphologically normal tissues from the peripheral zone (were most PCa arise), as we did. Remarkably, SFRP1 and other SFRP family members have been shown to have not only tumor suppressor activity but also tumor-pro-moting activity.24 Indeed, SFRP1 has shown antiapoptotic and increased cell proliferation activities.16 Moreover, overexpression of SFRP1 by prostatic tumor stroma may account for the previ-ously reported capacity of prostatic tumor stroma to provide a pro-proliferative paracrine signal to adjacent epithelial cells.16 Thus, it seems plausible that SFRP1 silencing through promoter methylation in normal prostate tissue helps to inhibit excessive cell proliferation. On the contrary, in prostate carcinomas an increase in SFRP1 expression might foster cell proliferation and evasion of apoptosis, eventually through non-canonical activa-tion of the Wnt signaling.27
Recently, suppression of Wnt signaling has been accom-plished with a novel small molecule (PKF118-310), causing inhibition of prostate cancer cell proliferation, holding prom-ise for prostate cancer prevention or treatment.28 Moreover, another compound—docosahexaenoic acid—has anti-tumoral
evidence that in renal cell tumors aberrant promoter methyla-tion of Wnt pathway antagonist might serve as an alternative to genetic mutations for accomplishing constitutive activation of the pathway. However, and contrarily to genetic mutations, more that one antagonist gene was found to be epigenetically altered in several primary renal tumors tested. A possible explanation for this finding may lie on the epigenetic heterogeneity of tumor cells, i.e., the progressive accumulation of methylated CpG dinu-cleotides gradually reduces gene expression levels, thus requiring
Table 1. CTNNB1 mRNa relative expression levels in renal cell tumors, either unmethylated at all Wnt antagonist genes (Unmethylated) or methylated in at least one of those genes (Methylated)
Methylation status CTNNB1 expression p value*
Unmethylated (n = 12) 153.15 0.010
Methylated (n = 17) 1029.43*Mann-Whitney test. Expression data was available in 29 of 31 cases.
Table 2. clinical and histopathological parameters of patients with urological tumors
Clinicopathological features
BCa RCT PCa
Patients, n 36 31 30
Gender, n (%)
Male 28 (77.8) 17 (54.8) n.a
Female 8 (22.2) 14 (45.2) n.a
Age, yr, median (range) 69.5 (50–89) 63 (36–86) 65 (40–70)
PSA, ng/mL, median (range)
n.a n.a8.03
(3.11–28.35)
Histologic subtype, n (%)
clear cell Rcc n.a 19 (61.3) n.a
papillary Rcc n.a 5 (16.1) n.a
chromophobe Rcc n.a 4 (12.9) n.a
Oncocytoma n.a 3 (9.7) n.a
Pathologic stage, n (%)
pTa-pTis 4 (11.1) n.a n.a
pT1 22 (61.1) 16 (51.6) n.a
pT2 3 (8.3) 6 (19.4) 17 (56.7)
pT3 5 (13.9) 8 (25.8) 13 (43.3)
pT4 2 (5.6) 1 (3.2) n.a
Grade, n (%)
1 3 (8.3) 1 (3.6) n.a
2 16 (44.4) 11 (39.3) n.a
3 16 (44.4) 13 (46.4) n.a
4 n.a 3 (10.7) n.a
Gleason score, n (%)
5 n.a n.a 1 (3.3)
6 n.a n.a 14 (46.7)
7 n.a n.a 15 (50.0)
n.a, not applicable; Bca, bladder carcinoma; RcT, renal cell tumor; pca, prostate carcinoma.

www.landesbioscience.com Epigenetics 349
Bisulfite treatment and QMSP. Sodium bisulfite conver-sion was performed as previously described.35 Briefly, 2 µg of genomic DNA were denatured in 0.3 M NaOH for 20 min at 50°C. The denatured DNA was diluted in 450 µl of a freshly prepared bisulfite solution, and the mixture was incubated at 70°C for 3 h in the dark. After incubation, the resulting bisulfite-converted DNA was desalted and purified through a column by using Wizard DNA purification resin (Wizard DNA Clean-Up System; Promega, Madison, WI, USA). The eluted DNA was denatured in 0.3 M NaOH, precipitated with 100% ethanol, resuspended and stored at -80°C.
The chemically modified DNA was amplified by fluorescence-based real-time methylation-specific PCR, using TaqMan tech-nology (PerkinElmer Corp., Foster City, CA, USA) as described previously.36 We performed QMSP in 96-well plates using an Applied Biosystems 7000 Sequence Detection System (Foster City, CA, USA).
APC, CDH1 and CTNNB1 genes were amplified using prim-ers and probe sequences previously reported.34 Primers and probe sequences used for the SFRP1 (GenBank: NM_003012) and WIF1 (GenBank: NM_007191) genes were the following, respectively: sense, 5'-TTC GGT CGT AGC AGT TTC G-3'; antisense, 5'-AAC GAC GCT ACG AAC TAA ATA-3'; and probe, 6FAM-5'-AAA ACG CGA AAC GAC GAA CGC-3'-TAMRA and sense, 5'-ACG CGT TTA GTC GTT TAA AC-3'; antisense, 5'-TAC CGA AAA AAC TCC TCG TA-3'; and probe, 6FAM-5'-TAC GCA ACC TAA CGC CGT CAA A-3'-TAMRA. To normalize for DNA input in each sample, β-actin (ACTB) was used as an internal reference gene, once its primers and probe were designed to amplify a region CpG nucleotides free.37
Fluorescence based real-time PCR assays were carried out in a reaction volume of 20 µL, consisting of 16.6 mM ammonium sulfate; 67 mM trizma preset; 6.7 mM magnesium chloride; 10 mM mercaptoethanol; 0.1% DMSO; 200 µM each of dATP, dCTP, dGTP and dTTP; 600 nM of each primer; 0.4 µL of Rox dye; 200 nM of probe; 1 unit of Platinum Taq polymerase (Invitrogen, Carlsbad, CA), and 2 µl of bisulfite-modified DNA as a template. To ensure the specificity of the analysis, each 96-well PCR plate had multiple water blanks, as a control for contamination (negative control), and completely methylated DNA at all CpGs as a positive control. All amplifications were performed at 95°C for 2 minutes, followed by 50 cycles of 95°C for 15 seconds, and 60°C for 1 minute. For the SFRP1 gene pro-moter QMSP assay, the reaction was carried out under the same conditions, except for the annealing temperature (62°C).
To determine the relative levels of fully methylated alleles in each sample, we used serial dilutions of the positive control DNA to construct the calibration curve. The values obtained, after QMSP analysis (mean quantity) for each target gene, were divided by the respective values of the internal reference gene. The ratio generated, which constitutes an index of the percentage of input copies of DNA that are fully methylated at the primer- and probe-binding sites, was then multiplied by 1000 for easier tabulation [methylation level = (target gene/ACTB) x 1000]. The methylation status of tumor samples was categorized for each gene (unmethylated vs. methylated) based on the distribution of
effects through induction of β-catenin degradation.29 Thus, pharmacological inhibition of Wnt signalling pathway might be a relevant anti-cancer therapeutic approach. In this context, our findings suggest that the methylation status of CTNNB1 and Wnt antagonist genes might indicate which urological tumors are more likely to benefit from this therapeutic strategy, and may thus serve as surrogate biomarkers for response prediction in these neoplasms.
In conclusion, our study suggests that epigenetic deregula-tion of Wnt pathway inhibitors may contribute to aberrant acti-vation of Wnt signaling in bladder, prostate and renal tumors, eventually contributing to increased cell proliferation and uncontrolled differentiation, thus playing an important role in urological carcinogenesis.
Materials and Methods
Cancer cell lines. Human cancer cell lines representative of each tumor type [bladder cancer: 5637, J82, T24, TCCSUP; renal cell cancer: 769-P, 786-O, ACHN, Caki-1; and prostate cancer: 22Rv1, DU145, LNCaP, PC-3] were obtained from the American Type Culture Collection (Manassas, VA). These cell lines were cultured according to manufacturer’s specifications, with 10% fetal bovine serum and antibiotics (100 units/mL penicillin and 100 µg/mL streptomycin), and maintained in a humidified atmosphere of 5% CO
2, 95% air at 37°C.
Patients and sample collection. Fresh-frozen samples, stored at -80°C, of primary transitional cell carcinoma of bladder (BCa, 36 cases), prostate carcinoma (PCa, 30 cases), and renal cell tumors (RCT, 31 cases) were collected for the purposes of this study. In addition, morphologically normal tissue samples were included, comprising 15 samples of bladder mucosa (NBM, obtained from patients submitted to radical prostatectomy for prostate cancer), 10 samples of normal prostate tissue (NPT, collected from cystoprostatectomy specimens of bladder cancer patients not harboring a simultaneous prostate carcinoma) and five samples of normal renal tissue, obtained from autopsies of patients without renal tumors. All samples were cut in cryostat for morphological control purposes. Then, the areas of interest were further sectioned for DNA extraction.
Routine pathological evaluation was performed in all cases in formalin-fixed and paraffin-embedded tissue fragments, to determine the tumor type,30 grade31,32 and pathological stage.33 Relevant clinical data were obtained from patient’s medical records (see Table 2). All patients were diagnosed and treated at the Portuguese Oncology Institute—Porto, Portugal, and their enrollment followed informed consent procedures. These stud-ies were approved by the institutional review board (Comissão de Ética) of the Portuguese Oncology Institute—Porto.
Nucleic acids extraction. Genomic DNA was extracted from cell lines and tissue samples using a standard technique, com-prising overnight digestion with proteinase K (20 mg/mL) in the presence of 10% SDS at 55°C, followed by phenol-chloroform extraction and precipitation with 100% ethanol.34 Total RNA was isolated using FastRNA Kit Green (QBiogene, Carlsbad, CA, USA) as per the manufacturer’s protocol.

350 Epigenetics Volume 5 Issue 4
normalized against the values of the internal reference gene to obtain a ratio that was then multiplied by 1000 for easier tabula-tion [(CTNNB1/HPRT1) x 1000].
Statistical analysis. The median and interquartile range of methylation levels were determined for each target gene. Differences in the distribution of methylation levels of the five genes among cancer cell line or tissue samples were assessed using the Mann-Whitney U test. Results were considered statistically significant at the two-sided 5% significance level. Statistical anal-yses were done using a computer-assisted program (SPSS version 15.0).
Acknowledgements
V.L.C. and F.R.R. are supported by scholar grants from Fundação para a Ciência e a Tecnologia (SFRH/BD/23374/2005, SFRH/BPD/26492/2006, respectively).
This study was funded by grants from Liga Portuguesa Contra o Cancro—Núcleo Regional do Norte, and the Calouste Gulbenkian Foundation (Project # 96474).
methylation levels across the respective normal tissue samples, i.e., levels equal to, or higher than, those median values were considered methylated. These criteria were based on our previous observations on promoter methylation of several genes in prostate and renal tumors and respective normal tissues.38-40
cDNA synthesis and quantitative reverse-transcription-PCR. Total RNA was reverse-transcribed using RevertAidTM H Minus first strand cDNA synthesis kit (Fermentas Inc., Glen Burnie, MD), with random hexamers as reaction primer. CTNNB1 (Hs00991809_g1) and the endogenous control HPRT1 (Hs99999909_m1) assays from Applied Biosystems were ampli-fied, and the real time quantitative gene expression was measured by the 7000 Sequence Detection System (Foster City, CA). PCR was performed according to manufacturer’s specifications in sep-arate wells for each primer/probe set and each sample was run in triplicate. Each plate included multiple nontemplate controls and serial dilutions of a positive control for constructing the standard curve on each plate. To determine the relative expression levels of CTNNB1 in each sample, the values of the target gene were
References1. Jemal A, Siegel R, Ward E, Hao Y, Xu J, Thun MJ.
Cancer statistics, 2009. CA Cancer J Clin 2009; 59:225-49.
2. Lustig B, Behrens J. The Wnt signaling pathway and its role in tumor development. J Cancer Res Clin Oncol 2003; 129:199-221.
3. Gavert N, Ben-Ze’ev A. β-catenin signaling in biologi-cal control and cancer. J Cell Biochem 2007; 102:820-8.
4. Aguilera O, Muñoz A, Esteller M, Fraga MF. Epigenetic alterations of the Wnt/β-catenin pathway in human disease. Endocr Metab Immune Disord Drug Targets 2007; 7:13-21.
5. Roth W, Wild-Bode C, Platten M, Grimmel C, Melkonyan HS, Dichgans J, et al. Secreted frizzled-related proteins inhibit motility and promote growth of human malignant glioma cells. Oncogene 2000; 19:4210-20.
6. Takada T, Yagi Y, Maekita T, Imura M, Nakagawa S, Tsao S-W, et al. Methylation-associated silencing of the Wnt antagonist SFRP1 gene in human ovarian cancers. Cancer Sci 2004; 95:741-4.
7. Suzuki H, Watkins DN, Jair K-W, Schuebel KE, Markowitz SD, Chen WD, et al. Epigenetic inactiva-tion of SFRP genes allows constitutive WNT signaling in colorectal cancer. Nat Genet 2004; 36:417-22.
8. Mazieres J, He B, You L, Xu Z, Lee AY, Mikami I, et al. Wnt inhibitory factor-1 is silenced by promoter methylation in human lung cancer. Cancer Res 2004; 64:4717-20.
9. Zou H, Molina JR, Harrington JJ, Osborn NK, Klatt KK, Romero Y, et al. Aberrant methylation of secreted frizzled-related protein genes in esophageal adenocar-cinoma and Barrett’s esophagus. Int J Cancer 2005; 116:584-91.
10. Ai L, Tao Q, Zhong S, Fields CR, Kim W-J, Lee MW, et al. Inactivation of Wnt inhibitory factor-1 (WIF1) expression by epigenetic silencing is a common event in breast cancer. Carcinogenesis 2006; 27:1341-8.
11. Takagi H, Sasaki S, Suzuki H, Toyota M, Maruyama R, Nojima M, et al. Frequent epigenetic inactiva-tion of SFRP genes in hepatocellular carcinoma. J Gastroenterol 2008; 43:378-89.
12. Hoque MO, Prencipe M, Poeta ML, Barbano R, Valori VM, Copetti M, et al. Changes in CpG islands promoter methylation patterns during ductal breast carcinoma progression. Cancer Epidemiol Biomarkers Prev 2009; 18:2694-700.
13. Verras M, Sun Z. Roles and regulation of Wnt signal-ing and β-catenin in prostate cancer. Cancer Lett 2006; 237:22-32.
14. Bilim V, Kawasaki T, Katagiri A, Wakatsuki S, Takahashi K, Tomita Y. Altered expression of β-catenin in renal cell cancer and transitional cell cancer with the absence of β-catenin gene mutations. Clin Cancer Res 2000; 6:460-6.
15. Henrique RM, Costa VL, Jeronimo C. Methylation-based biomarkers for early detection of urological cancer. Crit Rev Oncog 2007; 13:265-82.
16. Joesting MS, Perrin S, Elenbaas B, Fawell SE, Rubin JS, Franco OE, et al. Identification of SFRP1 as a candidate mediator of stromal-to-epithelial signaling in prostate cancer. Cancer Res 2005; 65:10423-30.
17. Urakami S, Shiina H, Enokida H, Kawakami T, Kawamoto K, Hirata H, et al. Combination analysis of methylated Wnt-antagonist family genes as a novel epigenetic biomarker panel for bladder cancer detec-tion. Clin Cancer Res 2006; 12:2109-16.
18. Urakami S, Shiina H, Enokida H, Hirata H, Kawamoto K, Kawakami T, et al. Wnt antagonist family genes as biomarkers for diagnosis, staging and prognosis of renal cell carcinoma using tumor and serum DNA. Clin Cancer Res 2006; 12:6989-97.
19. Gumz ML, Zou H, Kreinest PA, Childs AC, Belmonte LS, LeGrand SN, et al. Secreted frizzled-related protein 1 loss contributes to tumor phenotype of clear cell renal cell carcinoma. Clin Cancer Res 2007; 13:4740-9.
20. Henrique R, Ribeiro FR, Fonseca D, Hoque MO, Carvalho AL, Costa VL, et al. High promoter methyla-tion levels of APC predict poor prognosis in sextant biopsies from prostate cancer patients. Clin Cancer Res 2007; 13:6122-9.
21. Urakami S, Shiina H, Enokida H, Kawakami T, Tokizane T, Ogishima T, et al. Epigenetic inactivation of Wnt inhibitory factor-1 plays na important role in bladder cancer through aberrant canonical Wnt/β-catenin signaling pathway. Clin Cancer Res 2006; 12:383-91.
22. Neuhausen A, Florl AR, Grimm M-O, Schulz WA. DNA methylation alterations in urothelial carcinoma. Cancer Biol Ther 2006; 5:993-1001.
23. Sarrió D, Moreno-Bueno G, Hardisson D, Sánchez-Estévez C, Guo M, Herman JG, et al. Epigenetic and genetic alterations of APC and CDH1 genes in lobular breast cancer: relationships with abnormal E-cadherin and catenin expresion and microsatellite instability. Int J Cancer 2003; 106:208-15.
24. Rubin JS, Barshishat-Kupper M, Feroze-Merzoug F, Xi ZF. Secreted WNT antagonists as tumor suppressors: pro and con. Front Biosci 2006; 11:2093-105.
25. Florl AR, Steinhoff C, Muller M, Seifert H-H, Hader C, Engers R, et al. Coordinate hypermethylation at specific genes in prostate carcinoma precedes LINE-I hypomethylation. Br J Cancer 2004; 91:985-94.
26. Lodygin D, Epanchintsev A, Menssen A, Diebold J, Hermeking H. Functional epigenomics identifies genes frequently silenced in prostate cancer. Cancer Res 2005; 65:4218-27.
27. Joesting MS, Cheever TR, Volzing KG, Yamaguchi TP, Wolf V, Naf D. Secreted frizzled related protein 1 is a paracrine modulator of epithelial branching morpho-genesis, proliferation, and secretory gene expression in the prostate. Dev Biol 2008; 317:161-73.
28. Lu W, Tinsley HN, Keeton A, Qu Z, Piazza GA, Li Y. Suppression of Wnt/β-catenin signaling inhibits pros-tate cancer cell proliferation. European J Pharmacology 2009; 602:8-14.
29. Calviello G, Resci F, Serini S, Piccioni E, Toesca A, Boninsegna A, et al. Docosahexaenoic acid induces pro-teasome-dependent degradation of β-catenin, downreg-ulation of survivin, and apoptosis in human colorectal cancer cells not expressing COX-2. Carcinogenesis 2007; 28:1202-9.
30. Eble JN, Sauter G, Epstein JI, Sesterhenn IA. Part 1: Tumors of the kidney. In World health organization classification of tumors. Pathology and genetics of tumors of the urinary system and male genital organs. Lyon (France): IARC Press 2004; 9-43.
31. Gleason DF, Mellinger GT. Prediction of prognosis for prostatic adenocarcinoma by combined histological grading and clinical staging. J Urol 1974; 111:58-64.
32. Fuhrman SA, Lasky LC, Limas C. Prognostic sig-nificance of morphologic parameters in renal cell carcinoma. Am J Surg Pathol 1982; 6:655-63.
33. Sobin LH, Wittekind Ch, editors. TNM classification of malignant tumors, 6th ed. Geneva: UICC 2002.
34. Pearson H, Stirling D. DNA extraction from tissue. Methods Mol Biol 2003; 226:33-4.
35. Clark SJ, Harrison J, Paul CL, Frommer M. High sen-sitivity mapping of methylated cytosines. Nucleic Acids Res 1994; 22:2990-7.
36. Eads CA, Danenberg KD, Kawakami K, Saltz LB, Blake C, Shibata D, et al. MethyLight: a high-through-put assay to measure DNA methylation. Nucleic Acids Res 2000; 28:32.

www.landesbioscience.com Epigenetics 351
37. Eads CA, Lord RV, Wickramasinghe K, Long TI, Kurumboor SK, Bernstein L, et al. Epigenetic patterns in the progression of esophageal adenocarcinoma. Cancer Res 2001; 61:3410-8.
38. Henrique R, Jerónimo C, Teixeira MR, Hoque MO, Carvalho AL, Pais I, et al. Epigenetic heterogeneity of high-grade prostatic intraepithelial neoplasia: clues for clonal progression in prostate carcinogenesis. Mol Cancer Res 2006; 4:1-8.
39. Henrique R, Costa VL, Cerveira N, Carvalho AL, Hoque MO, Ribeiro FR, et al. Hypermethylation of cyclin D2 is associated with loss of mRNA expression and tumor development in prostate cancer. J Mol Med 2006; 84:911-8.
40. Costa VL, Henrique R, Ribeiro FR, Pinto M, Oliveira J, Lobo F, et al. Quantitative promoter methylation analysis of multiple cancer-related genes in renal cell tumors. BMC Cancer 2007; 7:133.
Copyright © 2022 FDOKUMEN




















