
Environmental heterogeneity, fungal parasitism and the demography of the grass Stipa leucotricha
8
Oecologia (1995) 103: 55-62 ORIGINAL PAPER Springer-Verlag 1995 Norma L. Fowler ? Keith Clay Environmental heterogeneity, fungal parasitism and the demography of the grass Stipa leucotricha Received: 16 June 1994 / Accepted: 31 January 1995 Abstract This study investigated the demographic con- sequences of fungal infection of a perennial grass, Stipa leucotricha. The rate of parasitism of this grass by the host-specific, systemic fungus Atkinsonella texensis varies over short distances. Infection was frequent (57% of plants) in "mottes" (clusters of woody plants) but rarer in adjacent open grasslands (9%). To test the hypothesis that the relative performance of infected and uninfected plants differed in the two habitats, infected and uninfected genotypes were collected from mottes in a central Texas population, propagated in the green- house and then transplanted into the same site in repli- cate plots within mottes, at the edges of mottes, and in open grassy areas. Demographic data were recorded for 30 months over three growing seasons. Plants were observed to lose and gain infection. Infection had no significant effect on plant survival, tiller number or dry mass although infected plants tended to be larger. Uninfected plants had a significantly higher probabil- ity of setting seed but there were no differences in seed production by reproductive plants. There were significant effects of planting environment on all of these measures. Motte edges were most favorable for S. leucotricha transplants while motte interiors were least favorable and open areas were intermediate. There was no evidence of habitat ? infection interactions; therefore the fungal infection had similar effects in different habitats. The high frequency of infection in motte habitats is best explained by more efficient con- tagious spread there. The favorability of motte edges for plant growth is substantially offset by higher infection rates at the edges of mottes. Key words Demography ? Environmental heterogeneity ? Fungal parasitism ? Reciprocal transplant ? Stipa leucotricha Introduction Populations of plants and other sessile organisms may experience very different environments over small spa- tial scales. Indeed, patchy environments, as continuous gradients or as mosaics, are probably the rule for plant populations (Fowler 1988c). This patchiness can have important effects on demographic parameters and the dynamics of populations (e.g., Quinn 1975; Grant and Antonovics 1978; Levin and Clay 1984), population genetic structure (Antonovics 1968; Stewart and Schoen 1987; Linhart 1988), and interactions of plant populations with other plant, animal, and microbial species (Crawley 1983; Burdon 1987; Fowler 1988c; Allen and Allen 1990; Louda et al. 1990). A common type of patchiness occurs in landscapes where vegetation dominated by woody plants meets herbaceous vegetation, as occurs at woodland margins and in savannas. One or a few trees can affect light intensity, soil properties, and the composition of the surrounding vegetation (Parker and M?ller 1982; Koop and Walker 1985; Archer et al. 1988; Belsky et al. 1989; Stuart-Hill and Tainton 1989). Interactions with her- bivores and pathogens may also vary over short dis- tances between adjacent wooded and open habitats. For example, Collinge and Louda (1988) found that rates of insect herbivory on Cardamine cordifolia were significantly higher in open sunny sites than in neigh- boring shaded sites (see also Huffaker and Kennett 1959; Lincoln and Mooney 1983). Similarly, Jarosz and Burdon (1988) found that wild barley, Hordeum leporinum, was more frequently infected by scald, Rhynchosporium secalis, under trees than in neighbor- ing open areas (see also Burdon et al. 1989). Such spa- tial variation in biotic interactions may restrict the distributions of many plant species even though they ?. L. Fowler (S) Department of Botany, University of Texas, Austin, TX 78713, USA K. Clay Department of Biology, Indiana University, Bloomington, IN 47405, USA This content downloaded from 129.116.71.180 on Mon, 9 Feb 2015 09:33:42 AM All use subject to JSTOR Terms and Conditions
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Environmental heterogeneity, fungal parasitism and the demography of the grass Stipa leucotricha

Oecologia (1995) 103: 55-62
ORIGINAL PAPER
Springer-Verlag 1995
Norma L. Fowler ? Keith Clay
Environmental heterogeneity, fungal parasitism and the demography
of the grass Stipa leucotricha
Received: 16 June 1994 / Accepted: 31 January 1995
Abstract This study investigated the demographic con-
sequences of fungal infection of a perennial grass, Stipa leucotricha. The rate of parasitism of this grass by the
host-specific, systemic fungus Atkinsonella texensis varies over short distances. Infection was frequent (57% of plants) in "mottes" (clusters of woody plants) but rarer in adjacent open grasslands (9%). To test the
hypothesis that the relative performance of infected and uninfected plants differed in the two habitats, infected and uninfected genotypes were collected from mottes in a central Texas population, propagated in the green- house and then transplanted into the same site in repli- cate plots within mottes, at the edges of mottes, and in
open grassy areas. Demographic data were recorded for 30 months over three growing seasons. Plants were observed to lose and gain infection. Infection had no
significant effect on plant survival, tiller number or dry mass although infected plants tended to be larger. Uninfected plants had a significantly higher probabil- ity of setting seed but there were no differences in seed production by reproductive plants. There were
significant effects of planting environment on all of these measures. Motte edges were most favorable for S. leucotricha transplants while motte interiors were least favorable and open areas were intermediate. There was no evidence of habitat ? infection interactions; therefore the fungal infection had similar effects in
different habitats. The high frequency of infection in
motte habitats is best explained by more efficient con-
tagious spread there. The favorability of motte edges for plant growth is substantially offset by higher infection rates at the edges of mottes.
Key words Demography ? Environmental
heterogeneity ?
Fungal parasitism ?
Reciprocal transplant
? Stipa leucotricha
Introduction
Populations of plants and other sessile organisms may experience very different environments over small spa- tial scales. Indeed, patchy environments, as continuous
gradients or as mosaics, are probably the rule for plant populations (Fowler 1988c). This patchiness can have
important effects on demographic parameters and the
dynamics of populations (e.g., Quinn 1975; Grant and Antonovics 1978; Levin and Clay 1984), population genetic structure (Antonovics 1968; Stewart and Schoen 1987; Linhart 1988), and interactions of plant populations with other plant, animal, and microbial
species (Crawley 1983; Burdon 1987; Fowler 1988c; Allen and Allen 1990; Louda et al. 1990).
A common type of patchiness occurs in landscapes where vegetation dominated by woody plants meets herbaceous vegetation, as occurs at woodland margins and in savannas. One or a few trees can affect light intensity, soil properties, and the composition of the
surrounding vegetation (Parker and M?ller 1982; Koop and Walker 1985; Archer et al. 1988; Belsky et al. 1989; Stuart-Hill and Tainton 1989). Interactions with her- bivores and pathogens may also vary over short dis- tances between adjacent wooded and open habitats. For
example, Collinge and Louda (1988) found that rates of insect herbivory on Cardamine cordifolia were
significantly higher in open sunny sites than in neigh- boring shaded sites (see also Huffaker and Kennett
1959; Lincoln and Mooney 1983). Similarly, Jarosz and Burdon (1988) found that wild barley, Hordeum
leporinum, was more frequently infected by scald,
Rhynchosporium secalis, under trees than in neighbor- ing open areas (see also Burdon et al. 1989). Such spa- tial variation in biotic interactions may restrict the distributions of many plant species even though they
?. L. Fowler (S) Department of Botany, University of Texas, Austin, TX 78713, USA
K. Clay Department of Biology, Indiana University, Bloomington, IN 47405, USA
This content downloaded from 129.116.71.180 on Mon, 9 Feb 2015 09:33:42 AMAll use subject to JSTOR Terms and Conditions

56 OECOLOGIA 103(1995) ? Springer-Verlag
are physiologically capable of growing in a wider range of habitats.
In this study we investigated the effects of environ- mental heterogeneity and fungal infection on the
demography of the perennial grass Stipa leucotricha. This grass grows in several distinct habitats created by clusters of woody plants ("mottes") in central Texas
grasslands. These habitats differ in the frequency of infection of the host grass S. leucotricha by the fungus Atkinsonella texensis, as well as in herbaceous vegeta- tion, light, and so on. The primary objective of this
study was to test the hypothesis that the effect of fun-
gal infection is conditional on environment, i.e. that
infected and uninfected plants differ in their response to environmental heterogeneity. To do this we experi-
mentally determined (1) how the survival, growth, and
fecundity of S. leucotricha vary among habitats, (2) how
infection alters the same traits, and (3) whether there
are interactions between habitat type and fungal infec-
tion such that the impact of disease varies with envi-
ronment. Multi-species interactions that vary as a
function of environmental heterogeneity are potentially
important in many plant populations but have
rarely been investigated experimentally in natural
communities.
Materials and methods
Study organisms
Stipa leucotricha Trin. and Rupr. (Texas wintergrass, Texas spear- grass; henceforth referred to as Stipa) is a caespitose perennial grass found in grasslands, prairies, and brushy areas of Oklahoma and Texas south to Coahuila and Nuevo Leon (Correli and Johnston 1970). On the eastern Edwards Plateau in central Texas, where this
study was conducted, juniper (Juniperus ashei Buchh.) and other woody plants often grow in discrete clusters (mottes) scattered throughout a grassland matrix (Fowler 1988a, 1990). Stipa is abun- dant at the edges of and inside mottes, but also occurs in adjacent open grassland, where it is usually less abundant and shorter in stature (N. Fowler, personal observation). Active growing periods occur in the spring (when it flowers) and in the fall. Flowering culms
produce a terminal panicle of 5-25 chasmogamous spikelets, each
containing a single flower and seed, and a smaller number of cleis- togamous flowers and seeds at the lowest nodes, surrounded by the leaf sheaths, called axillary cleistogenes (Dyksterhuis 1945).
The grass is frequently infected by the fungal parasite A. tex- ensis (Diehl) Leuchtmann and Clay (Diehl 1950; Leuchtmann and
Clay 1989). This host-specific fungus forms systemic infections by growing epiphytically on meristems, leaf primordia and developing inflorescences (Leuchtmann and Clay 1989). Panicles of chasmog- amous flowers are aborted in infected plants although the axillary cleistogenes may still be produced. Instead of chasmogamous flowers a fungal fruiting body (stroma) is produced. The transfer of conidia between stromata on different plants results in the pro- duction of meiotic ascospores, which can contagiously infect new hosts (Leuchtmann and Clay 1989). The exact mechanism of con-
tagious spread is not known but in the related A. hypoxylon ascospores are apparently washed into leaf axils (K. Clay, personal observation). Stipa plants are sometimes observed with both aborted, stroma-bearing inflorescences and normal, seed-bearing inflorescences. Microscopic examinations have indicated that the
fungus is absent rather than asymptomatic in normal inflorescences on infected plants (K. Clay, personal observation).
Local distribution and infection frequency
To quantify infection rates of Stipa by A. texensis in mottes and in open grassland areas, a series of 30 m transects was run in different sites in spring 1983. Three sites typical of Stipa habitat were selected. The Pedernales Falls State Park site, in Blanco Co., Texas, 80 km west of Austin, was the driest site and had the lowest proportion of area in mottes. The Gill Ranch site, in Travis Co., 15 km west of Austin, had a greater proportion of area in mottes. The Johnny Moore Road site, Travis Co. (near Austin), was a mosaic of open grassy areas, mottes, and scattered stands of honey mesquite (Prosopis glandulosa Torr.). The number of transects through each habitat in each site is given in Table I. Every 30 cm along each transect, the nearest Stipa plant was scored as nonflowering (and therefore of unknown infection status), as uninfected (possessing uninfected inflorescences), or as infected (possessing aborted inflorescences). If no plants occurred within 15 cm of the point, a blank was recorded. No plants were scored twice.
Reciprocal transplant experiment
To determine the separate and joint effects of fungal infection and habitat on Stipa performance a reciprocal transplant experiment was conducted. Flowering plants were dug up from Pedernales Falls State Park in spring 1984. The site was a typical mosaic of mottes and open grassy areas (see Fowler 1984, 1988b). At least 25 plants per infection class were collected. Plants were collected sufficiently far apart to insure that each was a different genotype. Due to the rarity and small size of infected plants in open areas, we were able to collect both infected and uninfected plants only from within mottes.
Plants were potted in a standard soil mix (2:1:1 sterile topsoil: perlite : peat) and maintained in a greenhouse. In spring 1985 plants were divided into ramets (clones) two to three tillers in size. Each ramet (henceforth referred to as a plant) was planted in the same soil mix in a plastic tube (Ray Leach "Conetainer", Canby, Ore, USA). Deaths of some plants resulted in an unbalanced number of ramets per genotype and of genotypes per origin. Therefore geno- type was not included in the statistical analyses.
Plants were transplanted into a site at Pedernales Falls State Park adjacent to the collection site in December 1985. Four tran- sects were located, each extending from a plot inside a large motte to a plot in an adjacent open grassy area. A third plot (the "edge" plot) was situated along each transect just inside the edge of the motte canopy. The distances between adjacent plots along a tran- sect varied from 3 to 10 m. In each motte and edge plot, the canopy was entirely composed of Juniperus ashei, except in one motte plot where Diospyros texana Scheele was also present. Holes of similar
depth and diameter to the plastic tubes were dug in a grid ( 15 cm
spacing between rows and columns) in each of the 12 plots. A total of 218 individual plants propagated from infected and uninfected motte plants were transplanted in December 1985. There were more
plants of infected than uninfected origin (138 versus 80) due to differences in original size and hence survival rates in the green- house. Plants were randomly assigned to plots and transects so that each plot and transect received approximately equal numbers of individuals from each origin. Each transplant was ringed with a red
plastic-coated wire and its grid location and initial tiller number were recorded.
Light level, air temperature, and relative humidity were mea- sured at each of the 12 experimental plots. Soil characteristics were not measured although they may have affected plant performance. While temperatures and humidities were very similar among plots, plots at the edge and inside mottes received only 25% and 20%
respectively of the average photosynthetic photon flux density (PPFD) received by plots in the open, as measured by a LiCor L?-109SA sensor at ground level when the sun was unobscured by clouds. Motte plots were shaded all day. Edge plots were shaded
This content downloaded from 129.116.71.180 on Mon, 9 Feb 2015 09:33:42 AMAll use subject to JSTOR Terms and Conditions

OECOLOGIA 103(1995) ? Springer-Verlag 57
at midday in June, when light readings were taken, but received direct sun in the morning (transect 3) or in the afternoon (transects 1,2, and 4).
Four censuses were made in spring 1986 (census 1), fall 1986 (census 2), spring 1987 (census 3), and spring 1988 (census 4). At each census the number of vegetative tillers per plant was recorded. At census 3, when plants first flowered, and at census 4 the num- ber of reproductive tillers and the number of terminal spikelets per tiller were also recorded, along with aborted inflorescences pro- duced by infected plants. After census 4, the above-ground portions of all surviving plants were harvested, dried and weighed, and the number of cleistogenes determined by dissection.
Statistical analyses
Runs tests (Siegel 1956) were made of infection frequencies for each 30 m transect in all three sites to determine whether infected plants were clumped. Contingency tables with ?2 tests were used to iden- tify significant differences in infection frequencies between mottes and open areas within each site.
The total fecundity of each plant during the experiment was obtained by adding the number of chasmogamous seeds produced in spring 1987 and spring 1988 and the number of cleistogamous seeds produced in spring 1988. This procedure slightly underestimates true fecundity because cleistogamous seeds were not counted in spring 1987, as that would have required destructive sampling. Total fecun-
dity, the number of tillers, and above-ground dry mass per plant in
spring 1988 were each log transformed to improve normality. These three variables were analyzed by analyses of variance
(ANOVA) that had origin (infected or uninfected), planting envi- ronment (motte interior, motte edge, open grassland), and transect (1-4) as independent variables. All possible interaction terms were included in these ANOVAs. Origin and planting environment were treated as fixed effects and transect as a random effect. The type III sums of squares (SAS 1985) were used because the data sets were unbalanced. Chi-square tests were used to evaluate the effects of origin, planting environment and transect on survival and the prob- ability of setting seed.
Because initial plant size can affect later performance, we checked for differences in plant size at the time of transplanting. Tiller num- ber per plant at transplanting was converted to a classified variable of 4 classes: tiller number = 1, 2, 3, or > 3. Tiller number is com- monly used as a non-destructive measure of plant size (e.g., Grant and Antonovics 1978; Fowler 1990; Clay 1990). Infected and unin- fected plants did not differ significantly in initial tiller number
(?]=2.??).
Results
Infection frequency
A significantly higher proportion of flowering plants were infected in mottes (which included motte edges) than in open grasslands at each site (57% versus 9% infection rate) (Table 1). The frequency of flowering plants exceeded 88% at all three sites and transects. Runs tests indicated a significant clumping of infected
plants in 4 of 8 transects in two of the sites (Table 1).
Effects of origin
There was no statistically significant effect of origin on
survival, biomass or tiller production, although plants
Table 1 Frequency of infection of the grass Stipa leucotricha by the fungus Atkinsonella texensis in mottes (clusters of woody plants) and in open grassland areas. There were 100 points per transect; not all had a S. leucotricha plant. Percentage infection calculated for flowering plants only (NS not significant)
Site, transect ? Percent Percent ?2 test Runs test no. and flowering infected (for each (for each habitat site) transect)
Pedernales 1 - motte 81 90% 63% ? < 0.0001
2-open 83 89% 5% ? < 0.0001 NS 3-open 96 96% 7% ? < 0.0001
Gill Ranch 4-motte 98 90% 53% ? < 0.0001 5-motte 75 96% 44% ? < 0.0001 NS 6-open 83 100% 18% ? < 0.005
Johnny Moore Road 7-motte 100 99% 66% ? < 0.0001 NS 8 - open 100 100% 5% NS
(Totals) all motte 354 94% 57% all open 362 96% 9%
Table 2 Results of analyses of variance of the effects of planting environment, transect, and origin on three measures of plant performance.
Effect df F
Aboveground dry mass/plant (log transformed) Planting environment 2 8.79* Transect 3 2.87** Origin 1 7.02 NS Environment x transect 6 3.22** Environment x origin 2 1.70 NS Transect x origin 3 0.17 NS Environment x transect x origin 6 0.95 NS Residual 114
No. tillers/plant, census 4 (log transformed) Planting environment 2 6.85* Transect 3 2.48 NS Origin 1 1.31 NS Environment x transect 6 2.56* Environment x origin 2 1.90 NS Transect x origin 3 0.33 NS Environment x transect x origin 6 0.93 NS Residual 115
Total seedset/plant (log transformed; reproducing plants only) Planting environment 2 17.99** Transect 3 1.96 NS Origin 1 9.72 ?=0.0525 Environment x transect 6 0.84 NS Environment x origin 2 2.35 NS Transect x origin 3 0.23 NS Environment x transect x origin 5 0.83 NS Residual 55
*P < 0.05; **P < 0.01; NS not significant
of uninfected origin weighed less and had fewer tillers
at the final census than plants of infected origin (Table 2, effect of "origin"; Fig. la, b). Origin did not
significantly affect survival to the final census
(?2 = 0.00001). There was a marginally significant trend
(P < 0.0525) for reproductive infected plants to pro- duce fewer seeds than reproductive uninfected plants (Table 2; Fig. Id).
This content downloaded from 129.116.71.180 on Mon, 9 Feb 2015 09:33:42 AMAll use subject to JSTOR Terms and Conditions
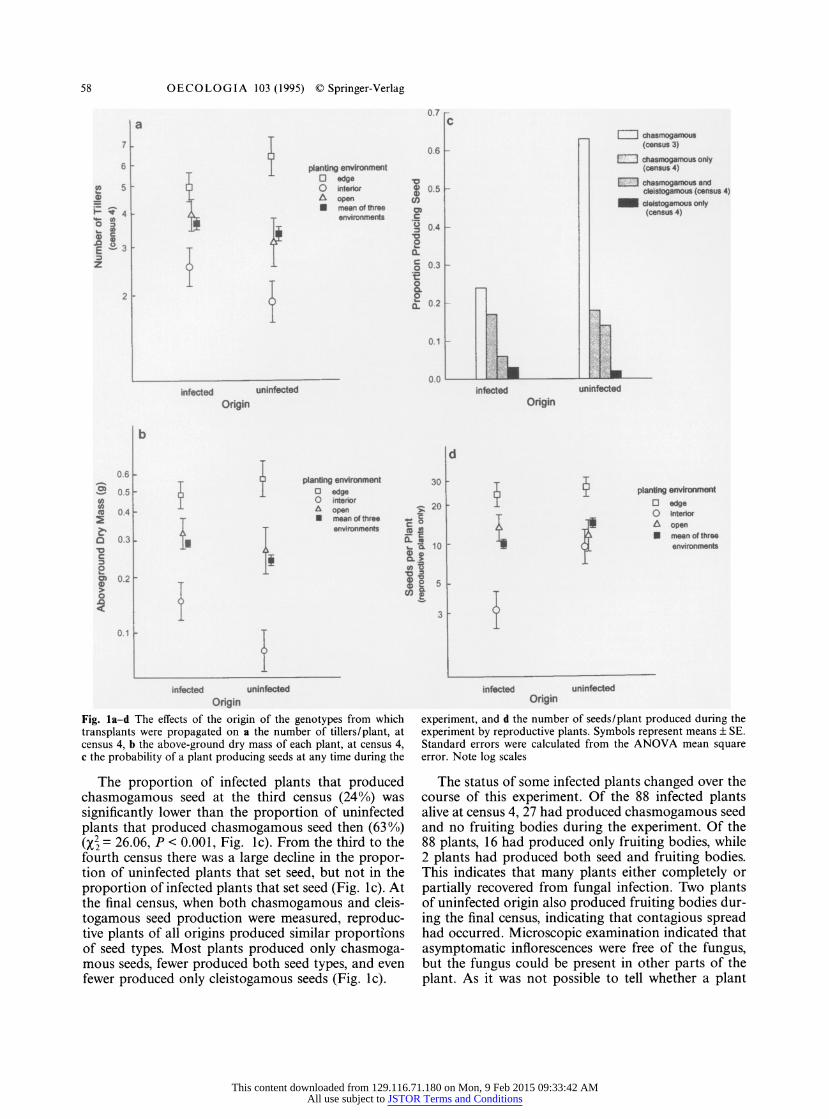
58 OECOLOGIA 103 (1995) ? Springer-Verlag
S2 f
? JO E ~ 3
planting environment D edge O interior ? open ? mean of three
environments
infected uninfected
Origin
0.7
0.6
1 0.5 CO
c ? 0.4 -a o ?
? 0.3 r 8. ? ? 0.2
0.1
0.0
I l chasmogamous (census 3)
\ i chasmogamous only {census 4)
i ? chasmogamous and cleistogamous (census 4)
W??? cleistogamous only (census 4)
infected uninfected
Origin
0.6 ?
?2 0.5 r
5
Q ?D c
? o .O <
0.4
0.3 L
0.2 h
0.1 h
planting environment ? edge O interior ? open ? mean of three
environments
30
20
10
si co ??
planting environment D edge O interior ? open ? mean of three
environments
infected uninfected
Origin infected
Origin uninfected
Fig. la-d The effects of the origin of the genotypes from which transplants were propagated on a the number of tillers/plant, at census 4, b the above-ground dry mass of each plant, at census 4, c the probability of a plant producing seeds at any time during the
experiment, and d the number of seeds/plant produced during the experiment by reproductive plants. Symbols represent means ? SE. Standard errors were calculated from the ANOVA mean square error. Note log scales
The proportion of infected plants that produced chasmogamous seed at the third census (24%) was
significantly lower than the proportion of uninfected
plants that produced chasmogamous seed then (63%)
(?2 = 26.06, ? < 0.001, Fig. le). From the third to the
fourth census there was a large decline in the propor- tion of uninfected plants that set seed, but not in the
proportion of infected plants that set seed (Fig. Ic). At the final census, when both chasmogamous and cleis-
togamous seed production were measured, reproduc- tive plants of all origins produced similar proportions of seed types. Most plants produced only chasmoga- mous seeds, fewer produced both seed types, and even
fewer produced only cleistogamous seeds (Fig. Ic).
The status of some infected plants changed over the
course of this experiment. Of the 88 infected plants alive at census 4, 27 had produced chasmogamous seed
and no fruiting bodies during the experiment. Of the
88 plants, 16 had produced only fruiting bodies, while
2 plants had produced both seed and fruiting bodies.
This indicates that many plants either completely or
partially recovered from fungal infection. Two plants of uninfected origin also produced fruiting bodies dur-
ing the final census, indicating that contagious spread had occurred. Microscopic examination indicated that
asymptomatic inflorescences were free of the fungus, but the fungus could be present in other parts of the
plant. As it was not possible to tell whether a plant
This content downloaded from 129.116.71.180 on Mon, 9 Feb 2015 09:33:42 AMAll use subject to JSTOR Terms and Conditions

OECOLOGIA 103(1995) ? Springer-Verlag 59
10
8
? 7
F 6
H^5 o S
fi 3 ?
1.00 0.80 h
^ 0.60 h >S5 (A g 0.40
Q "D C
2 F > O
0.10 h
0.05
motte-interior motte-edge open
Planting Environment
motte-interior motte-edge open
Planting Environment
O ? a
transect 1 transect 2 transect 3 transect 4 mean
O ? D V
transect 1 transect 2 transect 3 transect 4 mean
0.6
0.5
? F <? CO 0.4 ?) C ? 3 "? S 0.3 ?_ c o ?
| ??2
0.1
0.0
40
30
-20
iS ?
Q. ? 10
L
I I chasmogamous (census 3)
I J chasmogamous only (census 4)
G?:1 chasmogamous and cleistogamous (census 4)
W??? cleistogamous only (census 4)
I
interior edge open
Planting Environment
O transect 1 ? transect 2 ? transect 3 V transect 4 ? mean
motte-interior motte-edge open Planting Environment
Fig. 2a-d The effects of the environment in which a transplant was grown on a the number of tillers/plant, at census 4, b the above- ground dry mass of each plant, at census 4, c the probability of a plant producing seeds at any time during the experiment, and
d the number of seeds/plant produced during the experiment by reproductive plants. Symbols represent means ? SE. Standard errors were calculated from the ANOVA mean square error. Note log scales
had completely lost the infection, the ANOVAs treated
all infected origin plants as a single class regardless of
their symptoms. Infected plants grown in edge and open plots were
further separated into one group that had produced
only seeds and a second group that had produced only
fruiting bodies. The two plants that produced both seed
and fruiting bodies were not included, nor were four
motte-grown plants that set seed, as no motte-grown
plants produced fungal fruiting bodies. On average, the
16 symptomatic plants were larger and produced more
biomass than the 23 asymptomatic plants (9.4 versus
5.7 tillers, 1.06g versus 0.66g; ? < 0.05 for both,
i-tests, log transformed values).
Effects of environment and transect
Regardless of their origin, plants grown at the edge of
mottes had the greatest biomass and the most tillers
and, if they reproduced, set the most seed during the
experiment (Table 2, effect of "planting environment";
Fig. 2a, b, d). Survival through the experiment also
differed significantly among the three planting
This content downloaded from 129.116.71.180 on Mon, 9 Feb 2015 09:33:42 AMAll use subject to JSTOR Terms and Conditions

60 OECOLOGIA 103(1995) ? Springer-Verlag
100
??
I
II 60 S E
40
20 h
edge
open
interior
10 15 20 25 30
Months Since Planting
Fig. 3 Mortality (log scale) of transplants grown in different plant- ing environments during the experiment, pooled across all origins
environments (?^=8.55, ? < 0.05; Fig. 3). Plants
grown in the mottes generally had the lowest values for
these parameters and plants grown in the open were
intermediate. There was a significant environ- ment x transect interaction effect on biomass and tiller
number, indicating that the relative favorability of plots varied among transects (Table 2). The open plot in tran- sect 4 was especially favorable; it was shaded in late
afternoon, unlike other open plots, and may have pro- vided an environment more similar to that of the edge
plots. Plants in edge plots were more likely to produce
seeds in census 3 (?22= 4.52, ? < 0.10) and in census 4
(?22= 19.10, ? < 0.001) than plants in open or motte
plots (Fig. 2c). Plants within mottes set almost no seed. Of all the plants in edge plots, 23% produced cleis-
togamous seeds whereas less than 5% of plants in motte
and open plots produced cleistogamous seeds (?\- 12.69, ? < 0.01). Thus the relative production of cleis-
togamous seeds differed substantially between edge and
open plots.
Discussion
Stipa occupies a range of microenvironments created
by a mosaic of mottes in a grassland matrix. One of
our original objectives was to determine the quality of
different microenvironments for Stipa growth. The
edges of mottes were found to be more favorable for
Stipa than the interiors of mottes or the open grass- land. Plants in edge plots produced more tillers, bio-
mass, and seed, and they had higher rates of survival
than did plants in the open, despite competition from
woody plants. However, the frequency of infection by Atkinsonella texensis was significantly higher within motte habitats; data from three widely separated sites revealed that infection was on average 6 times more common in mottes than in open grasslands.
A second objective was to determine the effect of the fungal infection on Stipa demography. The most dramatic effect of infection is the production of fruit-
ing bodies in place of the terminal chasmogamous flowers. As a result, infected plants are largely steril-
ized, although some tillers may escape the infection and set seed. In this study, a substantial proportion of
initially infected plants ceased producing disease symp- toms (24% during census 3 and 23% during census 4) and two plants produced both seeds and fruiting bod- ies simultaneously. Microscopic examinations of such "mixed" plants indicated that asymptomatic portions had lost the infection. Greenhouse propagation and/or
transplant shock could have contributed to the loss of infection. In addition to losses of infection, two ini-
tially uninfected transplants became infected by A. tex- ensis over the course of the experiment. The level of
infection in Stipa populations is evidently dynamic due to contagious spread and loss of infection. Losses and
gains of infection among plants and plant parts have been documented in a variety of systemic fungi (Alexander and Antonovics 1988; Wennstrom 1994).
Despite its effects on fecundity, infection did not
impair the vegetative vigor of host plants. Initially infected plants produced more tillers and biomass than
initially uninfected plants (Fig. 1), although this difference was not significant. The measured differences are probably underestimates, given the loss of infection
by initially infected plants during the experiment. Supporting evidence is provided by the significantly
greater size of initially infected plants that produced fruiting bodies (indicating infection) compared to
plants of the same origin that produced seed (indicat-
ing loss of infection). Previous research has demon- strated that infection of the grass Danthonia spicata by the congeneric fungus A. hypoxy Ion enhances plant sur-
vival, tiller production, and interspecific competitive
ability relative to uninfected plants, although it reduces seed set (Clay 1984; Kelley and Clay 1987). Inocula- tions of seedlings with A. hypoxylon also significantly stimulated the rate of tillering in the absence of
flowering (Leuchtmann and Clay 1988). Similar effects have been reported in grasses and sedges infected by closely related fungi (Bradshaw 1959; Harberd 1961;
Clay 1990). The third objective of this study was to determine
whether there were significant origin ? environment
interactions, that is, whether the fitness consequences of fungal infection are conditional (sensu Bronstein
1994) upon the environment in which plants live.
However, no origin ? environment interactions were
detected; infected and uninfected plants performed
This content downloaded from 129.116.71.180 on Mon, 9 Feb 2015 09:33:42 AMAll use subject to JSTOR Terms and Conditions

OECOLOGIA 103(1995) ? Springer-Verlag 61
similarly in all environments. Given that there were no survival differences between infected and uninfected
plants, that there were no origin ? environment inter-
actions, and that infections were not disproportionately lost in the open plots, the most likely explanation for the significantly greater levels of infection in mottes is that contagious spread is more efficient in the motte environment.
The fitness of Stipa varied among the motte, edge and open habitats and between infection classes. Both habitat and fungal infection must be considered in any estimates of plant fitness in this mosaic environment. Uninfected plants were more fit, by all measures of
plant performance, at the edges of mottes than they were in the open, and more fit in the open than in the interiors of mottes. However, infection rates were
significantly lower in open areas, so that the propor- tion of plants sterilized by the fungus in open areas was lower. In the interior of mottes plant performance was poor and infection rates were high.
We used the data gathered in this study to estimate the average relative fitness of Stipa in mottes, motte
edges and open grassland. We assumed that
1. The probability of infection is 0.09 in open areas and 0.57 in the interiors and edges of mottes (Table 1). 2. The proportion of uninfected plants that set seed is 0.41 within mottes, 0.78 at motte edges and 0.42 in the
open (proportion of initially infected transplants set-
ting seed at census 3 and/or 4 in each habitat). 3. The mean number of seeds set by reproductive, unin- fected plants is 9.90 within mottes, 27.41 at motte edges and 11.11 in the open (Fig. 2d).
Because plants that remained infected throughout the course of the experiment set no cleistogamous seed, the seedset of infected plants was assumed to be 0.
Therefore, the expected fecundity of a plant is (1 ? the
probability of infection) ? (the probability of an unin- fected plant reproducing) ? (mean seedset of an unin- fected reproductive plant) :
1. At motte edges: 0.43 ? 0.78 ? 27.41 =9.19 seeds/plant 2. In the open: 0.91 ? 0.42 ? 11.11 = 4.25 seeds/plant 3. Within mottes: 0.43 ? 0.41 ? 9.90 = 1.75 seeds/plant
Thus, had our transplants been infected at a typical rate in each habitat, a transplant growing in an open grassy area would have a fitness of 0.46 relative to plants at the motte edge. The twofold higher rate of infection in motte edges would have substantially offset the 4.6-fold higher fecundity of uninfected plants in that habitat (mean seedset per uninfected transplant was 21.38 at motte edges, 4.67 in the open and 4.06 within
mottes). Plants within the mottes had a relative fitness of only 0.19, due to both a high rate of infection and to lower seedset if uninfected.
Over longer time spans, the difference in infection rates between motte edges and open areas may not offset the difference in habitat favorability as completely
as it does in the calculations above. Lower mortality rates in edge habitats (Fig. 3), coupled with faster
growth rates (Fig. 2a, b), would tend to produce greater size, and hence fecundity, differentials than those measured in this 3 year experiment. Nevertheless, differences in infection rates in this mosaic environment can substantially modify large fitness differentials created by other environmental factors.
These conclusions would be incorrect if infection affected a plant's responses to the environment; for example, if infection affected the relative favorabi-
lity of motte and open habitats for infected plants. However, all interaction effects involving infection were
non-significant. The hypothesis that the rarity of infected plants in open habitats results from their reduced fitness there is not supported by our data.
Acknowledgements We thank D. Riskind, W. Mengers, and the staff of Pedernales Falls State Park for permission to conduct this pro- ject there, and for facilitating our work in the Park; J. Russell Holman for assistance with all stages of the field work and subse- quent harvest and measurements; Emily Hsi for assistance collect- ing transect data; and the comments of anonymous reviewers on the manuscript. Support was provided by NSF grants BSR-8118968 and BSR-8600068 to N.L.F. and BSR-8400163 and BSR-8614972 to K.C.
References
Alexander HM, Antonovics J (1988) Disease spread and popula- tion dynamics of anther-smut infection of Sil?ne alba caused by the fungus Ustilago violacea. J Ecol 76: 91-104
Allen EB, Allen MF (1990) The mediation of competition by myc- orrhizae in successional and patchy environments. In: Grace JB, Tilman D (eds) Perspectives on plant competition. Academic Press, San Diego, pp 367-389
Antonovics J (1968) Evolution in closely adjacent plant popula- tions. VI. Manifold effects of gene flow. Heredity 23: 507-524
Archer S, Scifres C, Bassham CR, Maggio R (1988) Autogenic suc- cession in a subtropical savanna: conversion of grassland to thorn woodland. Ecol Monogr 58: 111-127
Belsky AJ, Amundson RG, Duxbury JM, Riha SJ, Ali AR, Mwonga SM (1989) The effects of trees on their physical, chemical, and biological environments in a semi-arid savanna in Kenya. J Appi Ecol 26: 1005-1024
Bradshaw AD (1959) Population differentiation in Agrostis tenuis Sibth. II. The incidence and significance of infection by Epichloe typhina. New Phytol 58: 310-315
Bronstein JL (1994) Conditional outcomes in mutualistic interac- tions. Trends Ecol Evol 9: 214-217
Burdon JJ (1987) Diseases and plant population biology. Cambridge University Press, Cambridge, UK
Burdon J, Jarosz AM, Kirby GC (1989) Patterns and patchiness in plant-pathogen interactions - causes and consequences. Annu Rev Ecol Syst 20: 119-136
Clay ? (1984) The effect of the fungus Atkinsonella hypoxylon (Clavicipitaceae) on the reproductive system and demography of the grass Danthonia spicata. New Phytol 98: 165-175
Clay ? (1990) Comparative demography of three graminoids infected by systemic, clavicipitaceous fungi. Ecology 71: 558-570
Collinge SK, Louda SM (1988) Herbivory by leaf-miners in response to experimental shading of a native crucifer. Oecologia 75: 559-566
Correli DS, Johnston MC (1970) Manual of vascular plants of Texas. Texas Research Foundation, Austin
This content downloaded from 129.116.71.180 on Mon, 9 Feb 2015 09:33:42 AMAll use subject to JSTOR Terms and Conditions

62 OECOLOGIA 103(1995) ? Springer-Verlag
Crawley MJ (1983) Herbivory: the dynamics of animal-plant inter- actions. University of California Press, Berkeley, Calif
Diehl WW (1950) Balansia and the Balansiae in America. Agrie Monogr 4 USDA., Washington, DC
Dyksterhuis EJ (1945) Axillary cleistogenes in Stipa and their role in nature. Ecology 26: 195-199
Fowler NL (1984) Patchiness in patterns of growth and survival of two grasses. Oecologia 62: 424-428
Fowler NL (1988a) Grasslands, nurse trees, and coexistence: three views of community structure. In: Amos BB, Gelbach FR (eds) Edwards Plateau vegetation: plant ecological studies in central Texas. Baylor University Press, Waco, Texas, pp 91-100
Fowler NL (1988b) What is a safe site?: neighbor, litter, germina- tion date and patch effects. Ecology 69: 947-961
Fowler NL (1988c) The effects of environmental heterogeneity in space and time on the regulation of populations and communities. In: Davy AJ, Hutchings MJ, Watkinson AR (eds) Plant population ecology. Blackwell, Oxford, pp 249-269
Fowler NL (1990) The effects of competition and environmental heterogeneity on three coexisting grasses. J Ecol 78: 389-402
Grant MC, Antonovics J (1978) Biology of ecologically marginal populations of Anthoxanthum odorat urn. I. Phenetics and dynamics. Evolution 32: 822-838
Harberd DJ (1961) Note on choke disease of Festuca rubra. Scott Plant Breed Stn Rep 1961: 47-51
Huffaker CB, Kennett CE (1959) A ten-year study of vegetational changes associated with the biological control of Klamath weed. J Range Manage 12: 69-82
Jarosz AM, Burdon JJ (1988) The effect of small scale change on disease incidence and severity in a plant-pathogen interaction. Oecologia 75: 78-81
Kelley SE, Clay ? (1987) Interspecific competitive interactions and the maintenance of genotypic variation within the populations of two perennial grasses. Evolution 41: 92-103
Koop WT, Walker BH (1985) Interactions of woody and herba- ceous vegetation in a southern African savanna. J Ecol 73: 235-253
Leuchtmann A, Clay ? (1988) Experimental infection of host grasses and sedges with Atkinsonella hypoxylon and Balansia cyperi (Balansiae, Clavicipitaceae). Mycologia 80: 291-297
Leuchtmann A, Clay ? (1989) Morphological, cultural and mat- ing studies on Atkinsonella, including A. texensis. Mycologia 81: 692-701
Levin DA, Clay ? (1984) Dynamics of synthetic Phlox drummondii populations at the species margin. Am J Bot 71: 1040-1050
Lincoln DE, Mooney HA (1983) Herbivory on Diplacus aurantia- cus shrubs in sun and shade. Oecologia 64: 173-176
Linhart YB (1988) Intrapopulation differentiation in annual plants. III. The contrasting effects of intra- and interspecific competi- tion. Evolution 42: 1047-1064
Louda SM, Keeler KH, Holt RD (1990) Herbivore influences on plant performance and competitive interactions. In: Grace JB, Tilman D (eds) Perspectives on plant competition, Academic Press, New York, pp 413-444
Parker VT, Muller CH (1982) Vegetational and environmental changes beneath isolated live oak trees (Quercus agrifolio) in a California annual grassland. Am Midi Nat 107: 69-81
Quinn JA ( 1975) Variability among Danthonia sericea (Gramineae) populations in responses to substrate moisture levels. Am J Bot 62: 884-891
SAS Institute Ine (1985) Sas User's Guide: Statistics, Version 5 Edition. Cary, NC, USA
Siegel S (1956) Nonparametric statistics for the behavioral sciences. McGraw-Hill, New York
Stewart SC, Schoen DJ (1987) Pattern of phenotypic viability and fecundity selection in a natural population of Impatiens pallida. Evolution 41: 1290-1301
Stuart-Hill GC, Tainton ? M (1989) The competitive interaction between Acacia karroo and the herbaceous layer and how this is influenced by defoliation. J Appi Ecol 26: 285-298
Wennstrom A (1994) Systemic diseases on hosts with different growth forms. Oikos 69: 535-538
This content downloaded from 129.116.71.180 on Mon, 9 Feb 2015 09:33:42 AMAll use subject to JSTOR Terms and Conditions














![Тайны советской демографии [The Secrets of Soviet Demography]](https://static.fdokumen.com/doc/165x107/633ca7f270f9415ce7029283/tayni-sovetskoy-demografii-the-secrets-of-soviet-demography.jpg)





