
Enhanced Microscopic Definition of Campylobacter jejuni 81-176 Adherence to, Invasion of,...
11
INFECTION AND IMMUNITY, Nov. 2008, p. 5294–5304 Vol. 76, No. 11 0019-9567/08/$08.000 doi:10.1128/IAI.01408-07 Copyright © 2008, American Society for Micrology. All Rights Reserved. Enhanced Microscopic Definition of Campylobacter jejuni 81-176 Adherence to, Invasion of, Translocation across, and Exocytosis from Polarized Human Intestinal Caco-2 Cells Lan Hu, 1 Ben D. Tall, 2 Sherill K. Curtis, 2 and Dennis J. Kopecko 1 * Laboratory of Enteric and Sexually Transmitted Diseases, FDA Center for Biologics Evaluation and Research, 29 Lincoln Drive, NIH Campus, Bldg. 29/420, Bethesda, Maryland 20892, 1 and FDA Center for Food Safety and Applied Nutrition, Laurel, Maryland 20708 2 Received 18 October 2007/Returned for modification 21 November 2007/Accepted 26 August 2008 Campylobacter jejuni-mediated pathogenesis involves gut adherence and translocation across intestinal cells. The current study was undertaken to examine the C. jejuni interaction with and translocation across differ- entiated Caco-2 cells to better understand Campylobacter’s pathogenesis. The efficiency of C. jejuni 81-176 invasion of Caco-2 cells was two- to threefold less than the efficiency of invasion of INT407 cells. Adherence- invasion analyses indicated that C. jejuni 81-176 adhered to most INT407 cells but invaded only about two-thirds of the host cells over 2 h (two bacteria/cell). In contrast, only 11 to 17% of differentiated Caco-2 cells were observed to bind and internalize either C. jejuni strain 81-176 or NCTC 11168, and a small percentage of infected Caco-2 cells contained 5 to 20 internalized bacteria per cell after 2 h. Electron microscopy revealed that individual C. jejuni cells adhered to the tips of host cell microvilli via intimate flagellar contacts and by lateral bacterial binding to the sides of microvilli. Next, bacteria were observed to bind at the apical host membrane surface via presumed interactions at one pole of the bacterium and with host membrane protrusions located near intercellular junctions. The latter contacts apparently resulted in coordinated, localized plasma membrane invagination, causing simultaneous internalization of bacteria into an endosome. Passage of this Campylobacter endosome intracellularly from the apical surface to the basolateral surface occurred over time, and bacterial release apparently resulted from endosome-basolateral membrane fusion (i.e., exocytosis). Bacteria were found intercellularly below tight junctions at 60 min postinfection, but not at earlier times. This study revealed unique host cell adherence contacts, early endocytosis-specific structures, and a presumptive exocytosis component of the transcellular transcytosis route. Campylobacter jejuni is a leading bacterial cause of human diarrheal disease. The U.S. Centers for Disease Control and Prevention estimates that C. jejuni causes illness annually in 1% of the population in the United States. Campylobacter infection usually causes watery to bloody diarrhea with fever, abdominal cramps, and the presence of fecal leukocytes. The results of intestinal biopsies of patients, infected primates, and several other experimental model animals, as well as cell cul- ture assays, have demonstrated that C. jejuni is able to invade enterocytes and suggest that gut adherence, invasion, and mucosal translocation are important steps in pathogenesis (20, 29, 46). The intestinal mucosa forms a barrier that protects against invasion of the host by nonpathogenic bacteria residing in the intestinal lumen. Some enteropathogens, such as Salmonella, Shigella, and Yersinia, have specific tissue-invading capabilities and can physically breach the intestinal mucosal barrier (32). Pathogen-induced traversal of the human intestinal mucosa may involve crossing of absorptive enterocytes or passage through the specialized M cells (14–16). In addition to trans- cellular entry, some bacteria can disrupt the tight junctions between enterocytes and cross via paracellular mechanisms (47). Salmonella enterica serovar Typhimurium may also cross the intestinal mucosa by sequestering itself within luminal CD18-positive mononuclear cells that shuttle the microbe paracellularly across the villus epithelium into the systemic circulation (48). Shigella (45) and possibly Campylobacter (49) may also breach the intestinal barrier by transcytosis across M cells. C. jejuni has previously been reported to undergo trans- cellular translocation across the mucosa; there have also been reports of paracellular translocation of Campylobacter (7, 9–12, 25, 28, 35, 47, 49). Cultured mammalian cells are commonly used as a simple model for investigating the host epithelial cell-bacterium interaction that is more easily controlled than whole ani- mals. Caco-2 cells were derived from transformed human colonic carcinoma cells. During 7 to 10 days in culture, these cells form polarized monolayers that have intercellular tight junctions and defined apical and basolateral surfaces. The differentiated apical surface has dense brush border mi- crovilli containing normal intestinal brush border enzymes and antigens (13, 18, 43, 44). A key advantage of differen- tiated Caco-2 cells is that they provide a substrate mono- layer similar to that for which Campylobacter shows a natu- ral tropism in vivo. This cell line allows assessment of the ability of C. jejuni to penetrate and pass through a differ- entiated intestinal epithelial barrier, a process that appears to be important in causing overall colonic damage and the * Corresponding author. Mailing address: Laboratory of Enteric and Sexually Transmitted Diseases, FDA Center for Biologics Evaluation and Research, 29 Lincoln Drive, NIH Campus, Bldg. 29/420, Bethesda, MD 20892. Phone: (301) 496-1893. Fax: (301) 402-8701. E-mail: dennis [email protected]. Published ahead of print on 2 September 2008. 5294 at FDA BIOSCIENCES LIBRARY on October 30, 2008 iai.asm.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Enhanced Microscopic Definition of Campylobacter jejuni 81-176 Adherence to, Invasion of,...

INFECTION AND IMMUNITY, Nov. 2008, p. 5294–5304 Vol. 76, No. 110019-9567/08/$08.00�0 doi:10.1128/IAI.01408-07Copyright © 2008, American Society for Micrology. All Rights Reserved.
Enhanced Microscopic Definition of Campylobacter jejuni 81-176Adherence to, Invasion of, Translocation across, and Exocytosis
from Polarized Human Intestinal Caco-2 Cells�
Lan Hu,1 Ben D. Tall,2 Sherill K. Curtis,2 and Dennis J. Kopecko1*Laboratory of Enteric and Sexually Transmitted Diseases, FDA Center for Biologics Evaluation and Research, 29 Lincoln Drive,
NIH Campus, Bldg. 29/420, Bethesda, Maryland 20892,1 and FDA Center for Food Safety and Applied Nutrition,Laurel, Maryland 207082
Received 18 October 2007/Returned for modification 21 November 2007/Accepted 26 August 2008
Campylobacter jejuni-mediated pathogenesis involves gut adherence and translocation across intestinal cells.The current study was undertaken to examine the C. jejuni interaction with and translocation across differ-entiated Caco-2 cells to better understand Campylobacter’s pathogenesis. The efficiency of C. jejuni 81-176invasion of Caco-2 cells was two- to threefold less than the efficiency of invasion of INT407 cells. Adherence-invasion analyses indicated that C. jejuni 81-176 adhered to most INT407 cells but invaded only abouttwo-thirds of the host cells over 2 h (two bacteria/cell). In contrast, only 11 to 17% of differentiated Caco-2 cellswere observed to bind and internalize either C. jejuni strain 81-176 or NCTC 11168, and a small percentage ofinfected Caco-2 cells contained 5 to 20 internalized bacteria per cell after 2 h. Electron microscopy revealedthat individual C. jejuni cells adhered to the tips of host cell microvilli via intimate flagellar contacts and bylateral bacterial binding to the sides of microvilli. Next, bacteria were observed to bind at the apical hostmembrane surface via presumed interactions at one pole of the bacterium and with host membrane protrusionslocated near intercellular junctions. The latter contacts apparently resulted in coordinated, localized plasmamembrane invagination, causing simultaneous internalization of bacteria into an endosome. Passage of thisCampylobacter endosome intracellularly from the apical surface to the basolateral surface occurred over time,and bacterial release apparently resulted from endosome-basolateral membrane fusion (i.e., exocytosis).Bacteria were found intercellularly below tight junctions at 60 min postinfection, but not at earlier times. Thisstudy revealed unique host cell adherence contacts, early endocytosis-specific structures, and a presumptiveexocytosis component of the transcellular transcytosis route.
Campylobacter jejuni is a leading bacterial cause of humandiarrheal disease. The U.S. Centers for Disease Control andPrevention estimates that C. jejuni causes illness annually in1% of the population in the United States. Campylobacterinfection usually causes watery to bloody diarrhea with fever,abdominal cramps, and the presence of fecal leukocytes. Theresults of intestinal biopsies of patients, infected primates, andseveral other experimental model animals, as well as cell cul-ture assays, have demonstrated that C. jejuni is able to invadeenterocytes and suggest that gut adherence, invasion, andmucosal translocation are important steps in pathogenesis(20, 29, 46).
The intestinal mucosa forms a barrier that protects againstinvasion of the host by nonpathogenic bacteria residing in theintestinal lumen. Some enteropathogens, such as Salmonella,Shigella, and Yersinia, have specific tissue-invading capabilitiesand can physically breach the intestinal mucosal barrier (32).Pathogen-induced traversal of the human intestinal mucosamay involve crossing of absorptive enterocytes or passagethrough the specialized M cells (14–16). In addition to trans-cellular entry, some bacteria can disrupt the tight junctions
between enterocytes and cross via paracellular mechanisms(47). Salmonella enterica serovar Typhimurium may also crossthe intestinal mucosa by sequestering itself within luminalCD18-positive mononuclear cells that shuttle the microbeparacellularly across the villus epithelium into the systemiccirculation (48). Shigella (45) and possibly Campylobacter (49)may also breach the intestinal barrier by transcytosis across Mcells. C. jejuni has previously been reported to undergo trans-cellular translocation across the mucosa; there have also beenreports of paracellular translocation of Campylobacter (7, 9–12,25, 28, 35, 47, 49).
Cultured mammalian cells are commonly used as a simplemodel for investigating the host epithelial cell-bacteriuminteraction that is more easily controlled than whole ani-mals. Caco-2 cells were derived from transformed humancolonic carcinoma cells. During 7 to 10 days in culture, thesecells form polarized monolayers that have intercellular tightjunctions and defined apical and basolateral surfaces. Thedifferentiated apical surface has dense brush border mi-crovilli containing normal intestinal brush border enzymesand antigens (13, 18, 43, 44). A key advantage of differen-tiated Caco-2 cells is that they provide a substrate mono-layer similar to that for which Campylobacter shows a natu-ral tropism in vivo. This cell line allows assessment of theability of C. jejuni to penetrate and pass through a differ-entiated intestinal epithelial barrier, a process that appearsto be important in causing overall colonic damage and the
* Corresponding author. Mailing address: Laboratory of Enteric andSexually Transmitted Diseases, FDA Center for Biologics Evaluationand Research, 29 Lincoln Drive, NIH Campus, Bldg. 29/420, Bethesda,MD 20892. Phone: (301) 496-1893. Fax: (301) 402-8701. E-mail: [email protected].
� Published ahead of print on 2 September 2008.
5294
at FD
A B
IOS
CIE
NC
ES
LIBR
AR
Y on O
ctober 30, 2008 iai.asm
.orgD
ownloaded from

occasional bacteremia associated with Campylobacter en-terocolitis (9, 10, 26, 28).
C. jejuni 81-176 is a well-characterized strain which exhibitsrelatively high levels of epithelial cell invasion in cultured hostcell assays and causes a bloody colitis, as demonstrated inhuman challenge studies (6; D. Trible, unpublished data). Cellculture invasion assays conducted with inhibitors of cytoskel-etal function initially demonstrated that this strain invadesepithelial cells apically via a novel mechanism that is stronglymicrotubule (MT) dependent (38). Subsequent immunofluo-rescence studies of this 81-176 invasion process confirmed theinvolvement of MTs, demonstrated that the MT-associatedmolecular motor dynein is required for this uptake process,and defined the kinetics of INT407 cell invasion by C. jejuni81-176 (19). Recent work has also shown that Ca2� releasefrom host intracellular stores is essential for C. jejuni 81-176internalization into host cells (21). In addition, host signaltransduction studies have suggested that strain 81-176 interactsat filipin III-sensitive membrane microdomains of the host cellsurface with G-protein-coupled receptors, a process which ac-tivates phosphatidylinositol 3-kinase and mitogen-activatedprotein kinases that appear to be intimately involved in eventscontrolling 81-176 internalization (22). Very recently, the rhoGTPases Rac1 and Cdc42 have been implicated in the 81-176invasion process (31). It is also noteworthy that other research-ers, using different invasion assay conditions, have observedmixed involvement of both MTs and microfilaments in 81-176invasion (5, 36). Watson and Galan (51) recently demonstratedthat following internalization into host cells C. jejuni 81-176survives within modified endosomes that avoid fusion withlysosomes.
Based on previous Campylobacter pathogenesis studies withcultured cells, animals, and humans (4–8, 10, 12, 17, 19–22,23–40, 46, 49–53), we hypothesize that during the first fewhours of infection C. jejuni adheres to and crosses differenti-ated intestinal epithelial cells by a transcellular process whichdoes not disrupt transepithelial electrical resistance (TER)(transcytosis is apical endocytosis followed by basolateral exo-cytosis). Thus, a kinetic examination of C. jejuni internalizationand microscopic analyses of the events involving adherence,invasion, and transcytosis over a 4-h period were conductedwith the goal of obtaining an enhanced understanding ofCampylobacter-host cell interactions. Importantly, this studyprovided new information about entry kinetics and revealednew specific bacterium-host cell interactions involved in epi-thelial cell translocation that have not been observed in previ-ous electron microscope (EM) studies of Campylobacter entrymechanisms (4, 10, 26, 28, 31).
MATERIALS AND METHODS
Bacterial strains, cell lines, media, and culture conditions. The experimentswere conducted using C. jejuni 81-176, a well-studied strain that was originallyobtained from a Campylobacter disease outbreak in Minnesota (30). A 2� cheYmutant of 81-176 designated RY213 (53), which is basically nonadherent andnoninvasive, was used as a nonadherent control. Also, genome-sequenced strainNCTC 11168 was employed for adherence comparisons. C. jejuni was grown inMueller-Hinton biphasic medium and on Mueller-Hinton agar (Difco) under aCampylobacter microaerophilic atmosphere containing 10% O2, 5% CO2, and85% N2. Immortalized human colonic cancer cells (Caco-2) and human embry-onic intestinal epithelial cells (INT407), obtained from the American Type Cul-ture Collection, were maintained in liquid nitrogen and cultured in minimal
essential media with 20% (for Caco-2 cells) or 10% (for INT407 cells) heat-inactivated fetal calf serum (FCS) (Gibco), 2 mM L-glutamine, and 0.1 mMnonessential amino acids, as recommended by the American Type Culture Col-lection. The relative state of cell differentiation and maintenance of tight junc-tions were monitored by determining monolayer TER with a dual-voltage ohm-meter (Millicel-ERS; Millipore) (3), and brush border organization was verifiedby EM.
Invasion assays. An invasion assay was performed essentially as describedpreviously (19). Epithelial cells at a concentration of 105 cells/ml were added toeach well of a 24-well plate and incubated for 1 to 15 days depending on thedegree of differentiation desired. The cell culture medium was replaced daily.Bacteria in minimal essential medium were added to 1 ml of culture medium(containing heat-inactivated FCS as described above) in each well. The multi-plicity of infection (MOI) was varied, as indicated below. Infected monolayers,which were not centrifuged, were typically incubated for 2 h at 37°C in a 5%CO2-95% air atmosphere to allow invasion to occur. For time course analyses,the invasion period was varied from 0 to 120 min. Following the invasion period,the monolayer was washed three times with Earle’s balanced salt solution andincubated for another 2 h in fresh tissue culture medium with 10% FCS plus 100�g/ml gentamicin to kill extracellular bacteria. After the gentamicin-killing pe-riod, the infected monolayers were washed as described above, and the host cellswere lysed using 0.1% Triton X-100 in phosphate-buffered saline (PBS) for 15min at room temperature on an orbital shaker. Following serial dilution in PBS,released intracellular bacteria were enumerated by colony counting on Mueller-Hinton agar incubated at 37°C under microaerophilic conditions. Each internal-ization assay was performed simultaneously in two separate wells and was re-peated on at least three separate occasions. The results were expressed asmeans � standard errors of the means. Control experiments confirmed that 100�g/ml gentamicin killed essentially all (�99.99%) extracellular bacteria within2 h; any remaining bacteria after gentamicin killing had no appreciable effect oncalculation of the invasion kinetics over the time period studied.
Fluorescence microscopy. To distinguish extracellular bacteria from internal-ized bacteria, an immunofluorescence procedure that rendered internalized bac-teria red and noninternalized bacteria green was used. For these assays, host cellswere grown on 13-mm glass coverslips placed in 24-well culture plates prior tobacterial infection and enumeration. At various times postinfection, each mono-layer was washed three times in PBS and exposed to rabbit anti-C. jejuni 81-176antibody (kindly provided by P. Guerry-Kopecko) for 1 h. After the monolayerwas washed with PBS, fluorescein isothiocyanate-conjugated anti-rabbit mono-clonal antibody (Sigma) was added for an additional 1 h, which was followed bywashing. Then the monolayer was permeabilized with 0.1% Triton X-100 for 5min, treated with primary anti-C. jejuni antibody for 1 h, washed, and incubatedfor 1 h with Texas Red-conjugated goat anti-rabbit monoclonal antibody(Sigma). After three washes in PBS, the preparations were viewed with a ZeissMC100 fluorescence microscope. One hundred individual host cells in each fieldwere examined to determine the presence and numbers of intracellular (red)bacteria. A general assessment of bacterial adherence was also conducted.
To assess the number of host cells containing associated C. jejuni bacteria,limited experiments were conducted after a 30-min infection period that wasfollowed by permeabilization with Triton X-100, treatment with anti-C. jejuniantibody, and then treatment with fluorescein isothiocyanate-conjugated anti-rabbit monoclonal antibody. Total cell-associated bacteria (adherent bacteriaplus internalized bacteria) were enumerated by scanning 100 host cells in a field.Three different fields were counted, and the data were averaged for each prep-aration.
Scanning and transmission EM. The ability of C. jejuni to pass throughconfluent, polarized Caco-2 cell monolayers grown on Transwell filters wasdetermined as a measure of bacterial translocation across the host epithelial cellbarrier. Polarized Caco-2 monolayers were prepared by seeding 105 cells on aTranswell clear polyester membrane (pore size, 3 �m; diameter, 6.5 mm; Milli-pore Corp.), followed by growth for �7 days. The integrity of tight junctions wasmonitored by measuring TER. Preliminary studies revealed that the Caco-2 cellsdifferentiated faster in the presence of 20% FCS than in the presence of lowerserum concentrations. Both 7- and 15-day-old Caco-2 monolayers displayed highTER (�1,000 �/cm2), developed dense microvilli, and showed identical C. jejuniinvasion kinetics (Fig. 1).
For scanning EM, filters carrying monolayers infected with C. jejuni 81-176 orNCTC 11168 were washed three times in PBS and then immersed for 2 h at 4°Cin primary fixative (pH 7.2) consisting of 4% paraformaldehyde, 1% glutaralde-hyde, and 0.1 M sodium cacodylate. The specimens were rinsed three times in 0.1M cacodylate buffer (pH 7.2), postfixed for 1.5 h in 1% osmium tetroxide in 0.1M cacodylate buffer (pH 7.2), rinsed three times in 0.1 M cacodylate buffer (pH7.2), dehydrated gradually using 30, 50, and 70% ethanol, and brought to room
VOL. 76, 2008 C. JEJUNI TRANSLOCATION ACROSS POLARIZED Caco-2 CELLS 5295
at FD
A B
IOS
CIE
NC
ES
LIBR
AR
Y on O
ctober 30, 2008 iai.asm
.orgD
ownloaded from
temperature in 95% ethanol. Each specimen was placed in a wire mesh basketand dropped into a small beaker containing absolute ethanol. After dehydrationusing three additional 10-min changes of absolute ethanol, each specimen wasimmersed in liquid carbon dioxide and dried using a Denton DCP-1 critical pointdryer. Each specimen was mounted on a stub, coated with gold-palladium usinga Denton vacuum evaporator, and examined using a Hitachi S4700 field emissionscanning electron microscope. The large number of washes should have elimi-nated casual bacterial binding, leaving intimate contacts for viewing.
For transmission EM, infected monolayers were grown as described above onTranswell filters. Monolayers infected with C. jejuni 81-176 for different timeperiods were washed three times with PBS and then fixed in 1% glutaraldehydein 0.1 M sodium cacodylate buffer for 24 h at 4°C. Fixed, infected monolayerswere embedded, sectioned, stained, and observed by transmission EM as previ-ously described (23).
RESULTS
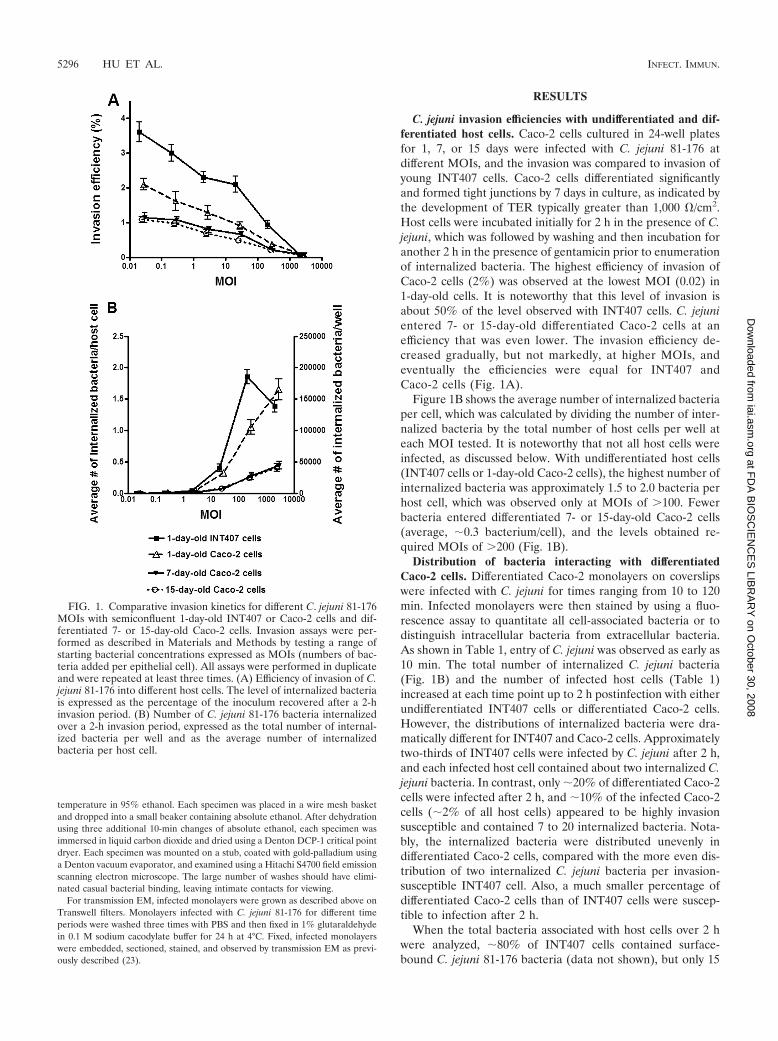
C. jejuni invasion efficiencies with undifferentiated and dif-ferentiated host cells. Caco-2 cells cultured in 24-well platesfor 1, 7, or 15 days were infected with C. jejuni 81-176 atdifferent MOIs, and the invasion was compared to invasion ofyoung INT407 cells. Caco-2 cells differentiated significantlyand formed tight junctions by 7 days in culture, as indicated bythe development of TER typically greater than 1,000 �/cm2.Host cells were incubated initially for 2 h in the presence of C.jejuni, which was followed by washing and then incubation foranother 2 h in the presence of gentamicin prior to enumerationof internalized bacteria. The highest efficiency of invasion ofCaco-2 cells (2%) was observed at the lowest MOI (0.02) in1-day-old cells. It is noteworthy that this level of invasion isabout 50% of the level observed with INT407 cells. C. jejunientered 7- or 15-day-old differentiated Caco-2 cells at anefficiency that was even lower. The invasion efficiency de-creased gradually, but not markedly, at higher MOIs, andeventually the efficiencies were equal for INT407 andCaco-2 cells (Fig. 1A).
Figure 1B shows the average number of internalized bacteriaper cell, which was calculated by dividing the number of inter-nalized bacteria by the total number of host cells per well ateach MOI tested. It is noteworthy that not all host cells wereinfected, as discussed below. With undifferentiated host cells(INT407 cells or 1-day-old Caco-2 cells), the highest number ofinternalized bacteria was approximately 1.5 to 2.0 bacteria perhost cell, which was observed only at MOIs of �100. Fewerbacteria entered differentiated 7- or 15-day-old Caco-2 cells(average, �0.3 bacterium/cell), and the levels obtained re-quired MOIs of �200 (Fig. 1B).
Distribution of bacteria interacting with differentiatedCaco-2 cells. Differentiated Caco-2 monolayers on coverslipswere infected with C. jejuni for times ranging from 10 to 120min. Infected monolayers were then stained by using a fluo-rescence assay to quantitate all cell-associated bacteria or todistinguish intracellular bacteria from extracellular bacteria.As shown in Table 1, entry of C. jejuni was observed as early as10 min. The total number of internalized C. jejuni bacteria(Fig. 1B) and the number of infected host cells (Table 1)increased at each time point up to 2 h postinfection with eitherundifferentiated INT407 cells or differentiated Caco-2 cells.However, the distributions of internalized bacteria were dra-matically different for INT407 and Caco-2 cells. Approximatelytwo-thirds of INT407 cells were infected by C. jejuni after 2 h,and each infected host cell contained about two internalized C.jejuni bacteria. In contrast, only �20% of differentiated Caco-2cells were infected after 2 h, and �10% of the infected Caco-2cells (�2% of all host cells) appeared to be highly invasionsusceptible and contained 7 to 20 internalized bacteria. Nota-bly, the internalized bacteria were distributed unevenly indifferentiated Caco-2 cells, compared with the more even dis-tribution of two internalized C. jejuni bacteria per invasion-susceptible INT407 cell. Also, a much smaller percentage ofdifferentiated Caco-2 cells than of INT407 cells were suscep-tible to infection after 2 h.
When the total bacteria associated with host cells over 2 hwere analyzed, �80% of INT407 cells contained surface-bound C. jejuni 81-176 bacteria (data not shown), but only 15
FIG. 1. Comparative invasion kinetics for different C. jejuni 81-176MOIs with semiconfluent 1-day-old INT407 or Caco-2 cells and dif-ferentiated 7- or 15-day-old Caco-2 cells. Invasion assays were per-formed as described in Materials and Methods by testing a range ofstarting bacterial concentrations expressed as MOIs (numbers of bac-teria added per epithelial cell). All assays were performed in duplicateand were repeated at least three times. (A) Efficiency of invasion of C.jejuni 81-176 into different host cells. The level of internalized bacteriais expressed as the percentage of the inoculum recovered after a 2-hinvasion period. (B) Number of C. jejuni 81-176 bacteria internalizedover a 2-h invasion period, expressed as the total number of internal-ized bacteria per well and as the average number of internalizedbacteria per host cell.
5296 HU ET AL. INFECT. IMMUN.
at FD
A B
IOS
CIE
NC
ES
LIBR
AR
Y on O
ctober 30, 2008 iai.asm
.orgD
ownloaded from
to 20% of differentiated Caco-2 cells had associated C. jejuni81-176 bacteria. Next, the host cell interactions of strain 81-176were compared to those of NCTC 11168 and the nonadherent2� cheY mutant of 81-176, RY213, over a 30-min infectionperiod (Table 2). Although strain 81-176 showed increasedadherence to undifferentiated INT407 cells, NCTC 11168 in-teracted equally well with differentiated and undifferentiatedhost cells. It is noteworthy that the adherence of mutantRY213 was markedly reduced, as previously reported (53).
Surface interactions of C. jejuni with Caco-2 cells early ininfection. Scanning EM was used to examine the early inter-action and binding of C. jejuni at the surface of differentiatedhost cells. Caco-2 cells that were grown on porous Transwellmembranes for 1 week formed confluent differentiated mono-layers, which was confirmed by the presence of dense apicalmicrovilli and tight intercellular junctions, as determined byhigh TER (see Materials and Methods).
These monolayers were infected, without centrifugation ofbacteria onto the monolayers, with C. jejuni at an MOI of 10 to100; the higher MOI made it easier to observe bound bacteriaby scanning EM, but the flagellar interactions appeared to beidentical at all MOIs. By 10 min postinfection, scanning EM
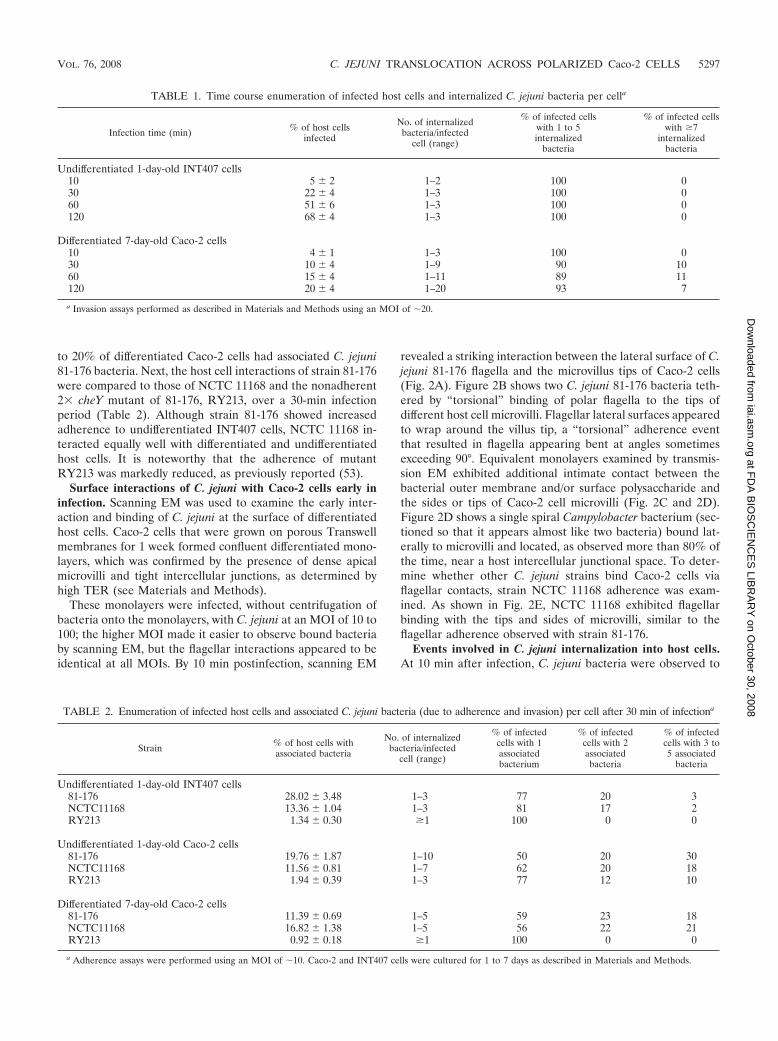
revealed a striking interaction between the lateral surface of C.jejuni 81-176 flagella and the microvillus tips of Caco-2 cells(Fig. 2A). Figure 2B shows two C. jejuni 81-176 bacteria teth-ered by “torsional” binding of polar flagella to the tips ofdifferent host cell microvilli. Flagellar lateral surfaces appearedto wrap around the villus tip, a “torsional” adherence eventthat resulted in flagella appearing bent at angles sometimesexceeding 90°. Equivalent monolayers examined by transmis-sion EM exhibited additional intimate contact between thebacterial outer membrane and/or surface polysaccharide andthe sides or tips of Caco-2 cell microvilli (Fig. 2C and 2D).Figure 2D shows a single spiral Campylobacter bacterium (sec-tioned so that it appears almost like two bacteria) bound lat-erally to microvilli and located, as observed more than 80% ofthe time, near a host intercellular junctional space. To deter-mine whether other C. jejuni strains bind Caco-2 cells viaflagellar contacts, strain NCTC 11168 adherence was exam-ined. As shown in Fig. 2E, NCTC 11168 exhibited flagellarbinding with the tips and sides of microvilli, similar to theflagellar adherence observed with strain 81-176.
Events involved in C. jejuni internalization into host cells.At 10 min after infection, C. jejuni bacteria were observed to
TABLE 1. Time course enumeration of infected host cells and internalized C. jejuni bacteria per cella
Infection time (min) % of host cellsinfected
No. of internalizedbacteria/infected
cell (range)
% of infected cellswith 1 to 5internalized
bacteria
% of infected cellswith �7
internalizedbacteria
Undifferentiated 1-day-old INT407 cells10 5 � 2 1–2 100 030 22 � 4 1–3 100 060 51 � 6 1–3 100 0120 68 � 4 1–3 100 0
Differentiated 7-day-old Caco-2 cells10 4 � 1 1–3 100 030 10 � 4 1–9 90 1060 15 � 4 1–11 89 11120 20 � 4 1–20 93 7
a Invasion assays performed as described in Materials and Methods using an MOI of �20.
TABLE 2. Enumeration of infected host cells and associated C. jejuni bacteria (due to adherence and invasion) per cell after 30 min of infectiona
Strain % of host cells withassociated bacteria
No. of internalizedbacteria/infected
cell (range)
% of infectedcells with 1associatedbacterium
% of infectedcells with 2associatedbacteria
% of infectedcells with 3 to5 associated
bacteria
Undifferentiated 1-day-old INT407 cells81-176 28.02 � 3.48 1–3 77 20 3NCTC11168 13.36 � 1.04 1–3 81 17 2RY213 1.34 � 0.30 �1 100 0 0
Undifferentiated 1-day-old Caco-2 cells81-176 19.76 � 1.87 1–10 50 20 30NCTC11168 11.56 � 0.81 1–7 62 20 18RY213 1.94 � 0.39 1–3 77 12 10
Differentiated 7-day-old Caco-2 cells81-176 11.39 � 0.69 1–5 59 23 18NCTC11168 16.82 � 1.38 1–5 56 22 21RY213 0.92 � 0.18 �1 100 0 0
a Adherence assays were performed using an MOI of �10. Caco-2 and INT407 cells were cultured for 1 to 7 days as described in Materials and Methods.
VOL. 76, 2008 C. JEJUNI TRANSLOCATION ACROSS POLARIZED Caco-2 CELLS 5297
at FD
A B
IOS
CIE
NC
ES
LIBR
AR
Y on O
ctober 30, 2008 iai.asm
.orgD
ownloaded from
5298
at FD
A B
IOS
CIE
NC
ES
LIBR
AR
Y on O
ctober 30, 2008 iai.asm
.orgD
ownloaded from

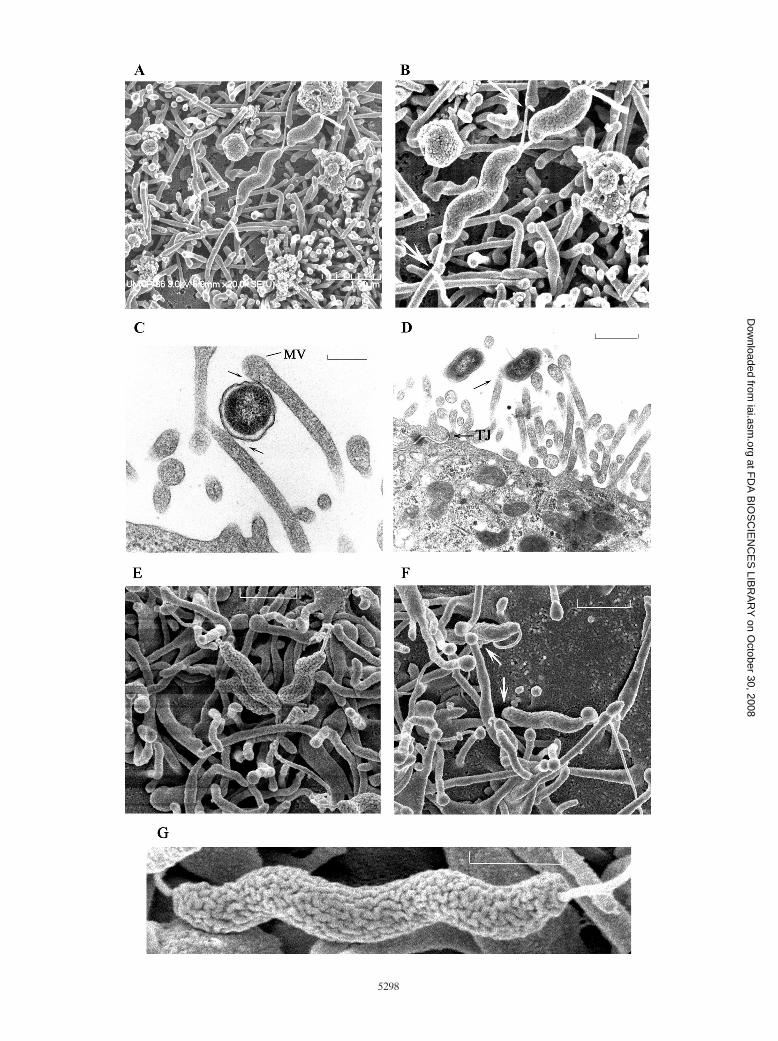
bind via flagella to Caco-2 cell microvilli (Fig. 2). These eventsoften occurred near a tight junction. Although the infectedmonolayers were washed thoroughly three times in PBS beforefixatives were added, we occasionally observed Campylobacterbound perpendicularly to the host cell surface. Because of theconsiderable washing during preparation, we believe that thisevent was not happenstance and reflected specific binding atone pole of the bacterium to the plasma membrane (Fig. 3A).We presumed that the polar flagella may have been bent out ofthe plane of sectioning and that only the proximal tips of theflagella could be seen (Fig. 3A). Figure 3B shows several bac-teria apparently in different stages of the internalization pro-cess. These bacteria are associated with an irregularly shapedmembrane protrusion, which may represent shortened, co-alesced microvilli at the Caco-2 cell surface. Several bacteriaappear to be bound to microvilli or vestigial microvilli. Onebacterium has apparently just been internalized and is in anendosome, as verified recently (51). At least two bacteria ap-pear to have been internalized previously and are in endo-somes which have moved to the base of a membrane protru-sion. The membrane protrusions appear to be activated sitesfor the uptake of multiple C. jejuni bacteria. Transmission EMexamination at 60 min revealed an occasional bacterium in thejunctional space (Fig. 3C), while another bacterium was adja-cent to this space but appeared to be transitioning from or tothe junction.
At 1 h postinfection, C. jejuni 81-176 bacteria were stillattached via flagellar contacts to microvilli, but the microvilliwere thicker and shorter and many of the microvilli had col-lapsed, leaving smooth areas visible on the host cell surface(Fig. 2F). Some bacteria appeared to be engulfed by invagina-tion of the plasma membrane (Fig. 2F). A highly enlargedtransmission EM micrograph of C. jejuni NCTC 11168 re-vealed the previously unobserved, convoluted, brainlike, spiralsurface of C. jejuni that may play a role in interaction with hostcells (Fig. 2G).
In contrast to the observations made with Caco-2 infectedcells, EM of INT407 cells during the initiation of Campy-lobacter invasion revealed a similar, but smaller, host cell mem-brane extension, which was about the size of Campylobacter.Thus, Campylobacter cells interacted at the INT407 cell surfaceand caused formation of a small membrane extension, a pro-trusion, or coalesced microvilli through which the Campy-lobacter cells were internalized by a membrane invaginationevent (Fig. 3D); also, there was an adjacent tight junction.
C. jejuni 81-176 translocation involves a discrete exocytosisevent. By 60 min postinfection, the ultrastructural evidence
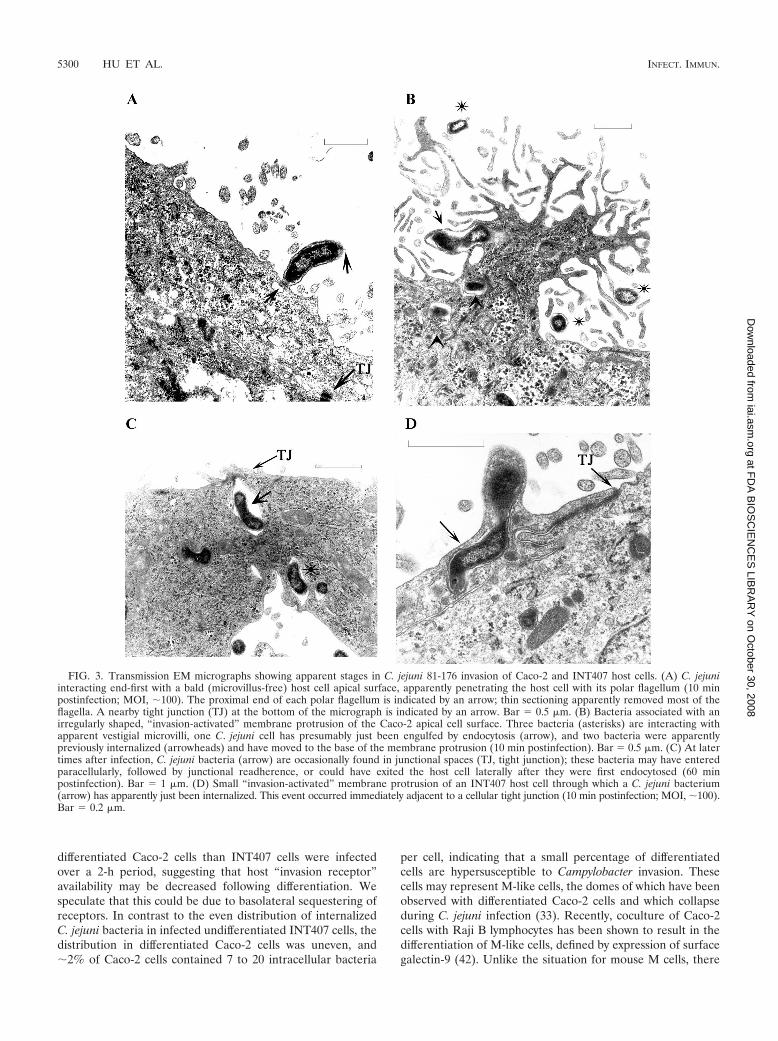
revealed movement of internalized Campylobacter cells withinendosomal vacuoles from the host cell apical domain to thebasolateral host cell domain. Figure 4A shows three bacteria inseparate endosomes, at least two of which are associated withthe perinuclear region. Figure 4B shows a Caco-2 cell contain-ing as many as 16 bacteria, and most of these bacteria wereclearly within endosomes near the basolateral surface at 1 hpostinfection. After moving within endosomes, first perinucle-arly and then to the basolateral host surface (Fig. 4C and D),bacteria are released by an apparent exocytosis event at thebasolateral host cell surface. Figure 4C shows two bacterianear the basolateral surface; one bacterium is outside the hostcell but above the transwell filter surface, and one bacterium isapparently in the process of being released from a cell byexocytosis. Figure 4D is an enlarged image of the exocytosisevent shown in Fig. 4C. The endosome containing C. jejuni haspresumably fused with the basolateral host cell membrane,creating a pore through which the bacterium is being releasedto the extracellular space, completing a transcellular transcy-tosis process. This exocytosis event is not a simple reversal ofthe endocytosis mechanism shown in Fig. 3B and presumablyoccurs by a separate, unique process. However, although weconsider it less likely based on our chronological studies, wecannot rule out the possibility that this event represented ba-solateral endocytosis of bacteria that entered paracellularly.
DISCUSSION
The current report describes an extension of our previousstudies aimed at characterizing C. jejuni 81-176 invasion. Thekinetics of C. jejuni 81-176 entry into differentiated Caco-2cells showed a pattern with increasing MOIs similar to thepattern observed for INT407 cells, but the overall level ofbacterial uptake was lower (Fig. 1). The invasion efficiency washighest at the lowest MOI (0.02), suggesting that single bacte-ria can trigger their own entry (29). Bacterial internalizationinto 7- or 15-day-old differentiated Caco-2 cells was reducedmost notably (Fig. 1B). Bacterium-host cell association assaysrevealed that �80% of INT407 cells contained adherent bac-teria but that only 15 to 20% of differentiated Caco-2 cellscontained bound bacteria. For example, entry into 1-day-oldINT407 cells occurred as early as 10 min and increased over2 h, after which 68% of the host cells were consistently infectedwith two internalized bacteria per host cell (Table 1). In strik-ing contrast, only 20% of polarized Caco-2 cells had internal-ized C. jejuni bacteria after 2 h, and the number of internalizedbacteria per host cell varied from 1 to 20. Thus, far fewer
FIG. 2. Scanning and transmission EM images of C. jejuni adherence to Caco-2 monolayers. (A) Scanning EM showing C. jejuni 81-176 boundvia flagella to tips of Caco-2 cell microvilli (10 min postinfection; MOI, �100). Bar � 1.5 �m. (B) Enlargement of panel A, showing the lateralsurfaces of flagella binding torsionally to tips of host cell microvilli (arrows). (C) Transmission EM showing cross-section of C. jejuni 81-176exhibiting intimate bacterial surface contact (arrows) with sides and tips of microvilli (MV) (10 min postinfection). Bar � 0.2 �m. (D) TransmissionEM of a longitudinal section of a single C. jejuni 81-176 cell adhering to a microvillus of a host cell (arrow) at 10 min postinfection. Thin sectioningeliminated the middle of the spiral bacterium. Note that the interaction occurs close to a tight junction (TJ). Bar � 0.5 �m. (E) Scanning EM ofC. jejuni NCTC 11168 showing flagella binding to tips of Caco-2 cell microvilli (60 min postinfection; MOI, 10). Bar � 1.0 �m. (F) Scanning EMshowing C. jejuni 81-176 flagella interacting with shortened microvilli and an adjacent host cell surface devoid of microvilli. The arrows indicatebacteria that are apparently being internalized by host membrane invagination (60 min postinfection; MOI, �10). Bar � 1.0 �m. (G) Enlargedscanning EM micrograph of C. jejuni NCTC 11168 showing the brainlike, convoluted surface of this spiral bacterium, which is bound via flagellarcontacts to host cell microvilli. Bar � 0.5 �m.
VOL. 76, 2008 C. JEJUNI TRANSLOCATION ACROSS POLARIZED Caco-2 CELLS 5299
at FD
A B
IOS
CIE
NC
ES
LIBR
AR
Y on O
ctober 30, 2008 iai.asm
.orgD
ownloaded from
differentiated Caco-2 cells than INT407 cells were infectedover a 2-h period, suggesting that host “invasion receptor”availability may be decreased following differentiation. Wespeculate that this could be due to basolateral sequestering ofreceptors. In contrast to the even distribution of internalizedC. jejuni bacteria in infected undifferentiated INT407 cells, thedistribution in differentiated Caco-2 cells was uneven, and�2% of Caco-2 cells contained 7 to 20 intracellular bacteria
per cell, indicating that a small percentage of differentiatedcells are hypersusceptible to Campylobacter invasion. Thesecells may represent M-like cells, the domes of which have beenobserved with differentiated Caco-2 cells and which collapseduring C. jejuni infection (33). Recently, coculture of Caco-2cells with Raji B lymphocytes has been shown to result in thedifferentiation of M-like cells, defined by expression of surfacegalectin-9 (42). Unlike the situation for mouse M cells, there
FIG. 3. Transmission EM micrographs showing apparent stages in C. jejuni 81-176 invasion of Caco-2 and INT407 host cells. (A) C. jejuniinteracting end-first with a bald (microvillus-free) host cell apical surface, apparently penetrating the host cell with its polar flagellum (10 minpostinfection; MOI, �100). The proximal end of each polar flagellum is indicated by an arrow; thin sectioning apparently removed most of theflagella. A nearby tight junction (TJ) at the bottom of the micrograph is indicated by an arrow. Bar � 0.5 �m. (B) Bacteria associated with anirregularly shaped, “invasion-activated” membrane protrusion of the Caco-2 apical cell surface. Three bacteria (asterisks) are interacting withapparent vestigial microvilli, one C. jejuni cell has presumably just been engulfed by endocytosis (arrow), and two bacteria were apparentlypreviously internalized (arrowheads) and have moved to the base of the membrane protrusion (10 min postinfection). Bar � 0.5 �m. (C) At latertimes after infection, C. jejuni bacteria (arrow) are occasionally found in junctional spaces (TJ, tight junction); these bacteria may have enteredparacellularly, followed by junctional readherence, or could have exited the host cell laterally after they were first endocytosed (60 minpostinfection). Bar � 1 �m. (D) Small “invasion-activated” membrane protrusion of an INT407 host cell through which a C. jejuni bacterium(arrow) has apparently just been internalized. This event occurred immediately adjacent to a cellular tight junction (10 min postinfection; MOI, �100).Bar � 0.2 �m.
5300 HU ET AL. INFECT. IMMUN.
at FD
A B
IOS
CIE
NC
ES
LIBR
AR
Y on O
ctober 30, 2008 iai.asm
.orgD
ownloaded from

are no commercially available antibodies to identify humanM cells, which prevented confirmation of this interestingpossibility.
Both scanning EM and transmission EM were used to ex-amine the early Campylobacter–Caco-2 cell interactions in dif-ferentiated monolayers grown on Transwell filters. ScanningEM revealed unique torsional contacts between the sides offlagella and microvillus tips, in contrast to previous reports offlagellar tip adherence (26, 28), possibly due to the higher-resolution images obtained in the current study. Both C. jejuni81-176 and NCTC 11168 were typically tethered at the hostsurface by each polar flagellum binding laterally to differentmicrovillus tips (Fig. 2 A, B, E, and G), an interaction whichsuggests that there is specific binding between the sides of
flagella and components of the rounded tips of microvilli.Transmission EM revealed additional contacts between thebacterial cell surface and the sides of microvilli (Fig. 2C andD). The latter contacts may involve capsular polysaccharide,Peb1, JlpA, or other outer membrane proteins which havebeen reported to be involved in adherence (2, 24, 39–41). C.jejuni has a brainlike convoluted surface (Fig. 2G), which mayplay a role in host cell-bacterium interactions. We speculatethat ligands located at the apical surface (instead of deep in thefolds) of the convolutions may be more active in host cellinteractions. The bacterium-host cell interactions describedabove were readily observed early and throughout the invasionperiod. We presume that they represent different types ofadherence, some of which may be reversible.
FIG. 4. Intracellular transport within endosomes and C. jejuni 81-176 exocytosis from Caco-2 cells. (A) Caco-2 cell containing three intracel-lular bacteria. One bacterium (arrow) close to apical surface appears to have just been internalized. The remaining two bacteria (asterisks) are inseparate endosomes and have migrated to the perinuclear region (N, nucleus) (30 min postinfection; MOI, 100). Bar � 0.5 �m. (B) Multiplebacteria in endosomes at the basolateral host cell surface at 1 h postinfection. The membrane filter is below the cell. Bar � 1 �m. (C) At 45 to90 min postinfection, C. jejuni endosomes are transported to the basolateral surface (membrane filter) and are released by endosome-plasmamembrane fusion, resulting in bacterial exocytosis. Bar � 1 �m. (D) Enlargement of panel C, more clearly showing the exocytosis event at thebasolateral surface of the host cell. The arrow indicates a pore created by membrane fusion of the endosome with the host basolateral membrane.The edge of the transwell filter is at bottom right of the micrograph.
VOL. 76, 2008 C. JEJUNI TRANSLOCATION ACROSS POLARIZED Caco-2 CELLS 5301
at FD
A B
IOS
CIE
NC
ES
LIBR
AR
Y on O
ctober 30, 2008 iai.asm
.orgD
ownloaded from

Flagella have long been implicated as Campylobacter ad-hesins (34, 37). Flagellum-dependent motility and the flagel-lum itself appear to be essential for C. jejuni invasion of epi-thelial cells, but the mechanism(s) remains unknown (1, 12, 50,52, 53). It seems unlikely that the Campylobacter flagellar ad-herence is casual because the infected monolayers werewashed six times with Earle’s balanced salt solution prior tofixation for scanning EM, a process that involves an additional10 washes and buffer changes. Although most EM studies wereperformed with an MOI of 100, using an MOI of 10 resulted insimilar bacterium-host cell interactions, but the frequencieswere much reduced. We speculate that C. jejuni flagellum-hostcell interactions represent a primary adherence mechanismthat augments the contact of “invasion-specific” bacterial li-gands with host membrane receptors in lipid rafts, which resultin signal transduction events that lead to bacterial internaliza-tion. Since �10% of the invasion ability can be restored tononflagellated C. jejuni cells by centrifugation onto a mono-layer (50), we suggest that “invasion-specific” C. jejuni ligandsmay be located at the bacterial poles, which are normallyadjacent to the polar flagella in wild-type C. jejuni. StrainNCTC 11168 is known to be less invasive than 81-176, possiblydue to reduced numbers of “invasion-specific” ligands or ex-pression of surface factors that interfere with these ligands.
C. jejuni cells were observed to interact typically with mi-crovilli at the apical Caco-2 cell surface adjacent to intercellu-lar junctions, as noted previously (28). Very infrequently,Campylobacter bacteria were observed bound perpendicular tothe host cell, possibly with one polar flagellum interacting withthe host cell (Fig. 3A); a similar EM observation of perpen-dicular bacterial interaction was recently reported (31). This isa relatively difficult event to observe, possibly because theorganism is internalized rapidly, and it is technically difficult tofind a thin section containing such a physically space-limitedevent. Figure 3A shows that the local host membrane surfacelacks the typical dense microvilli shown in Fig. 2D, and thismay reflect a differentiated M-like cell surface. We suggest thatintimate bacterium-host cell contact, as shown in Fig. 3A, B, orD, may be “invasion specific” and that binding of bacterialligands with specific host plasma membrane receptors mayhave triggered a localized collapse of the terminal F-actin web,leading to shortening and coalescence of microvilli. Previously,we and other workers have reported that pretreatment ofmonolayers with cytochalasin D actually enhances Campy-lobacter invasion (19, 38, 51), possibly by eliminating theterminal actin web. Thus, initial actin depolymerization andsubsequent polymerization are probably involved in Campy-lobacter entry. In fact, a recent report showed that Rac1 andCdc42 are involved and that there may be membrane rufflingprior to C. jejuni invasion (31). We have no evidence of effectorsecretion into host cells via the bacterium-host cell interactionshown in Fig. 3A, but we cannot exclude this interesting pos-sibility. Konkel and coworkers (27) suggested that effectorsecretion is important in Campylobacter invasion.
An early membrane dynamic event observed during Campy-lobacter entry is the formation of an activated membrane pro-trusion, which may represent a coalescence of local microvilli.We speculate that invasion-specific bacterium-host cell contactmay promote host signaling events that activate the mem-brane in a localized region, leading to bacterial internaliza-
tion through this “activated” site (Fig. 3B). “Activation” mayencompass host signaling events, as well as cytoskeletal andmembrane alterations. In differentiated Caco-2 cells, this “ac-tivated” membrane protrusion is approximately the length ofone bacterium wide and 2 to 3 lengths high and can apparentlyundergo multiple bacterial internalization events. In contrast,the comparable invasion-specific, “activated” membrane ex-tension observed in undifferentiated INT407 cells during C.jejuni invasion is smaller and apparently engulfs a single or-ganism (Fig. 3D). Regardless of the host membrane protrusionsize, bacterial internalization apparently results from plasmamembrane invagination that begins at the bacterium-host cellcontact site (19, 26, 29, 31). As observed previously (28, 38),the bacteria are internalized into an endosome, which is trans-ported over time from the apical host surface to the basolateralhost surface (Fig. 4). Our previous confocal microscopic anal-yses suggested that C. jejuni 81-176 cells are transported inendosomes via the molecular motor dynein along MTs (19).Watson and Galan (51) recently reported that Campylobactercells reside and transit within special endosomes that avoidfusion with lysosomes. At later infection times, an occasionalbacterium is observed in the intercellular space below apicaljunctional adherence (Fig. 3C). This event has not been ob-served commonly or earlier in infection, leading us to suggestthat this bacterium just exited the host cell laterally; however,the possibility of paracellular entry from the apical surfacefollowed by junctional resealing cannot be ruled out (8). Basedon the current study, we suggest that C. jejuni bacteria inendosomes are transported basolaterally through the cell andcan undergo specific endosome-basolateral membrane fusion(Fig. 4D), an exocytosis process which releases C. jejuni sub-epithelially. This process does not visually appear to be a sim-ple reversal of the apical endocytic mechanism. Although weconsider a basolateral endocytosis event unlikely, this possibil-ity cannot be ruled out by the results of EM. Previous reportsindicated that the ability of C. jejuni to invade Caco-2 cells isnot linked to translocation ability (7, 17), also suggesting thatendocytosis and exocytosis are unique mechanistic events.Based on these observations, we speculate that C. jejuni bac-teria proficient in transcytosis across the epithelium may beable to alter the Campylobacter-endosome membrane so thatthey traffic through the cell and can fuse with the host plasmamembrane, resulting in basolateral exocytosis. Since mono-layer TER is decreased at later infection times (33) and alter-ations in tight junction proteins have been observed at 24 to48 h postinfection (8, 33), it seems reasonable to suggest thatintercellular junction damage may occur at later times, follow-ing C. jejuni transcytosis across the intestinal epithelium.
In summary, this study revealed new specific C. jejuni-hostcell interactions and structures involved in adherence to,invasion of, and translocation across differentiated Caco-2cells that were not observed in previous studies. C. jejuniinteractions with differentiated Caco-2 cells likely are morerepresentative of events that actually occur in the intestine;nevertheless, INT407 cells have provided much useful infor-mation on C. jejuni adherence and invasion mechanisms. Incontrast to the relatively consistent level of 2 internalizedbacteria per undifferentiated INT407 cell, certain differen-tiated Caco-2 cells appeared to be hypersusceptible to inva-sion and contained as many as 20 internalized C. jejuni
5302 HU ET AL. INFECT. IMMUN.
at FD
A B
IOS
CIE
NC
ES
LIBR
AR
Y on O
ctober 30, 2008 iai.asm
.orgD
ownloaded from

bacteria. Whether these hypersusceptible Caco-2 cells arehuman M-like cells could not be verified due to a lack ofavailable reagents. Campylobacter transcytosis appears toinvolve discrete endocytic and exocytic events not previouslyobserved by transmission EM. Together, these findings pro-vide an improved concept for C. jejuni 81-176 adherence,endocytosis, and exocytosis mechanisms and indicate poten-tially important processes and structures that should becharacterized further.
ACKNOWLEDGMENTS
We thank Timothy K. Maugel for technical help with the scanningEM and Michael Schmitt and Alain Debrabant for critical reviews ofthe manuscript.
REFERENCES
1. Alm, R. A., P. Guerry, and T. J. Trust. 1993. The Campylobacter sigma 54flaB flagellin promoter is subject to environmental regulation. J. Bacteriol.175:4448–4455.
2. Bacon, D. J., C. M. Szymanski, D. H. Burr, R. P. Silver, R. A. Alm, and P.Guerry. 2001. A phase-variable capsule is involved in virulence of Campy-lobacter jejuni 81-176. Mol. Microbiol. 40:769–777.
3. Beatty, W. L., S. Meresse, P. Gounon, J. Davoust, J. Mounier, P. J. San-sonetti, and J. P. Gorvel. 1999. Trafficking of Shigella lipopolysaccharide inpolarized intestinal epithelial cells. J. Cell Biol. 145:689–698.
4. Biswas, D., K. Itoh, and C. Sasakawa. 2000. Uptake pathways of clinical andhealthy animal isolates of Campylobacter jejuni into INT-407 cells. FEMSImmunol. Med. Microbiol. 29:203–211.
5. Biswas, D., K. Itoh, and C. Sasakawa. 2003. Role of microfilaments andmicrotubules in the invasion of INT407 cells by Campylobacter jejuni. Mi-crobiol. Immunol. 47:469–473.
6. Black, R. E., M. M. Levine, M. L. Clements, T. P. Hughes, and M. J. Blaser.1988. Experimental Campylobacter jejuni infection in humans. J. Infect. Dis.157:472–479.
7. Bras, A. M., and J. M. Ketley. 1999. Transcellular translocation of Campy-lobacter jejuni across human polarised epithelial monolayers. FEMS Micro-biol. Lett. 179:209–215.
8. Chen, M. L., Z. Ge, J. G. Fox, and D. B. Schauer. 2006. Disruption of tightjunctions and induction of proinflammatory cytokine responses in colonicepithelial cells by Campylobacter jejuni. Infect. Immun. 74:6581–6597.
9. Cruz, N., Q. Lu, X. Alvarez, and E. A. Deitch. 1994. Bacterial translocationis bacterial species dependent: results using the human Caco-2 intestinal cellline. J. Trauma 36:612–616.
10. Everest, H., H. Goossens, J. P., Butzler, D. Lloyd, S. Knutton, J. M. Ketley,and P. H. Williams. 1992. Differentiated Caco-2 cells as a model for entericinvasion by Campylobacter jejuni and C. coli. J. Med. Microbiol. 37:319–325.
11. Gaynor, E., C. S. Cawthraw, G. Manning., J. K. MacKichan, S. Falkow, andD. G. Newell. 2004. The genome-sequenced variant of Campylobacter jejuniNCTC 11168 and the original clonal clinical isolate differ markedly in colo-nization, gene expression, and virulence-associated phenotypes. J. Bacteriol.186:503–517.
12. Grant, C. C., M. E. Konkel, W. Cieplak, Jr., and L. S. Tompkins. 1993. Roleof flagella in adherence, internalization, and translocation of Campylobacterjejuni in nonpolarized and polarized epithelial cell cultures. Infect. Immun.61:1764–1771.
13. Grasset, E., J. Bernabeu, and M. Pinto. 1985. Epithelial properties of humancolonic carcinoma cell line Caco-2: effect of secretagogues. Am. J. Physiol.248:C410–C418.
14. Grutzkau, A., C. Hanski, H. Hahn, and E. O. Riecken. 1990. Involvement ofM cells in the bacterial invasion of Peyer’s patches: a common mechanismshared by Yersinia enterocolitica and other enteroinvasive bacteria. Gut 31:1011–1015.
15. Finlay, B. B., B. Gumbiner, and S. Falkow. 1988. Penetration of Salmonellathrough a polarized Madin-Darby canine kidney epithelial cell monolayer.J. Cell Biol. 107:221–230.
16. Finlay, B. B., and S. Falkow. 1990. Salmonella interactions with polarizedhuman intestinal Caco-2 epithelial cells. J. Infect. Dis. 162:1096–1106.
17. Harvey, P., T. Battle, and S. Leach. 1999. Different invasion phenotypes ofCampylobacter isolates in Caco-2 cell monolayers. J. Med. Microbiol. 48:461–46924.
18. Hidalgo, I. J., T. J. Raub, and R. T. Borchardt. 1989. Characterization of thehuman colon carcinoma cell line (Caco-2) as a model system for intestinalepithelial permeability. Gastroenterology 96:736–749.
19. Hu, L., and D. J. Kopecko. 1999. Campylobacter jejuni 81-176 associates withmicrotubules and dynein during invasion of human intestinal cells. Infect.Immun. 67:4171–4182.
20. Hu, L., and D. J. Kopecko. 2000. Interactions of Campylobacter with eukary-
otic cells: gut luminal colonization and mucosal invasion mechanisms, p.191–215. In I. Nachamkin and M. J. Blaser (ed.), Campylobacter, 2nd ed.ASM Press, Washington, DC.
21. Hu, L., R. B. Raybourne, and D. J. Kopecko. 2005. Ca2� release from hostintracellular stores and related signal transduction during Campylobacterjejuni 81-176 internalization into human intestinal cells. Microbiology 151:3097–3105.
22. Hu, L., J. P McDaniel, and D. J. Kopecko. 2006. Signal transduction eventsinvolved in human epithelial cell invasion by Campylobacter jejuni 81-176.Microb. Pathog. 40:91–100.
23. Huang, X. Z., B. Tall, W. R. Schwan, and D. J. Kopecko. 1998. Physicallimitations on Salmonella typhi entry into cultured human intestinal epithe-lial cells. Infect. Immun. 66:2928–2937.
24. Jin, S., A. Joe, J. Lynett, E. K. Hani, P. Sherman, and V. L. Chan. 2001. JlpA,a novel surface-exposed lipoprotein specific to Campylobacter jejuni, medi-ates adherence to host epithelial cells. Mol. Microbiol. 39:1225–1236.
25. Konkel, M. E., and L. A. Joens. 1989. Adhesion to and invasion of HEp-2cells by Campylobacter spp. Infect. Immun. 57:2984–2990.
26. Konkel, M. E., M. D. Corwin, L. A. Joens, and W. Cieplak. 1992. Factors thatinfluence the interaction of Campylobacter jejuni with cultured mammaliancells. J. Med. Microbiol. 37:30–37.
27. Konkel, M. E., J. D. Klena, V. Rivera-Amill, M. R. Monteville, D. Biswas, B.Raphael, and J. Mickelson. 2004. Secretion of virulence proteins fromCampylobacter jejuni is dependent on a functional flagellar export apparatus.J. Bacteriol. 186:3296–3303.
28. Konkel, M. E., D. J. Mead, S. F. Hayes, and W. Cieplak, Jr. 1992. Translo-cation of Campylobacter jejuni across human polarized epithelial cell mono-layer cultures. J. Infect. Dis. 166:308–315.
29. Kopecko, D. J., L. Hu, and K. J. Zaal. 2001. Campylobacter jejuni-microtu-bule-dependent invasion. Trends Microbiol. 9:389–396.
30. Korlath, J. A., M. T. Osterholm, L. A. Judy, J. C. Forfang, and R. A.Robinson. 1985. A point-source outbreak of campylobacteriosis associatedwith consumption of raw milk. J. Infect. Dis. 152:592–596.
31. Krause-Gruszczynska, M., M. Rohde, R. Hartig, H. Genth, G. Schmidt, T.Keo, W. Konig, W. G. Miller, M. E. Konkel, and S. Backert. 2007. Role of thesmall Rho GTPases Rac1 and Cdc42 in host cell invasion of Campylobacterjejuni. Cell. Microbiol. 9:2431–2444.
32. Lencer, W. I. 2001. Microbial invasion across the intestinal epithelial barrier.Pediatr. Res. 49:4–5.
33. MacCallum, A., S. P. Hardy, and P. H. Everest. 2005. Campylobacter jejuniinhibits the absorptive transport functions of Caco-2 cells and disrupts cel-lular tight junctions. Microbiology 151:2451–2458.
34. McSweegan, E., and R. I. Walker. 1986. Identification and characterizationof two Campylobacter jejuni adhesins for cellular and mucous substrates.Infect. Immun. 53:141–148.
35. Monteville, M. R., and M. E. Konkel. 2002. Fibronectin-facilitated invasionof T84 eukaryotic cells by Campylobacter jejuni occurs preferentially at thebasolateral cell surface. Infect. Immun. 70:6665–6671.
36. Monteville, M. R., J. E. Yoon, and M. E. Konkel. 2003. Maximal adherenceand invasion of INT 407 cells by Campylobacter jejuni requires the CadFouter-membrane protein and microfilament reorganization. Microbiology149:153–165.
37. Newell, D. G., and A. Pearson. 1984. The invasion of epithelial cell lines andthe intestinal epithelium of infant mice by Campylobacter jejuni/coli. J. Di-arrhoeal Dis. Res. 2:19–26.
38. Oelschlaeger, T. A., P. Guerry, and D. J. Kopeck. 1993. Unusual microtu-bule-dependent endocytosis mechanisms triggered by Campylobacter jejuniand Citrobacter freundii. Proc. Natl. Acad. Sci. USA 90:6884–6888.
39. Pei, Z., and M. J. Blaser. 1993. PEB1, the major cell-binding factor ofCampylobacter jejuni, is a homolog of the binding component in gram-negative nutrient transport systems. J. Biol. Chem. 268:18717–18725.
40. Pei, Z., C. Burucoa, B. Grignon, S. Baqar, X. Z. Huang, D. J. Kopecko, A. L.Bourgeois, J. L. Fauchere, and M. J. Blaser. 1998. Mutation in the peb1Alocus of Campylobacter jejuni reduces interactions with epithelial cells andintestinal colonization of mice. Infect. Immun. 66:938–943.
41. Pei, Z. H., R. T. Ellison III, and M. J. Blaser. 1991. Identification, purifica-tion, and characterization of major antigenic proteins of Campylobacter je-juni. J. Biol. Chem. 266:16363–16369.
42. Pielage, J. F., C. Cichon, L. Greune, M. Hirashima, T. Kucharzik, and M. A.Schmidt. 2007. Reversible differentiation of Caco-2 cells reveals galectin-9 asa surface marker molecule for human follicle-associated epithelia and Mcell-like cells. Int. J. Biochem. Cell Biol. 39:1886–1901.
43. Pinto, M., S. Robine-Leon, M. D. Appay, M. Kedinger, N. Triadou, E.Dussaulx, B. Lacroix, P. Simon-Assmann, K. Haffen, J. Fogh, and A.Zweibaum. 1983. Enterocyte-like differentiation and polarization of the hu-man colon carcinoma cell line Caco-2 in culture. Biol. Cell 47:323–330.
44. Rousset, M., G. Trugnan, J. L. Brun, and A. Zweibaum. 1986. Inhibition ofthe post-translational processing of microvillar hydrolases is associated witha specific decreased expression of sucrase-isomaltase and an increased turn-over of glucose in Caco-2 cells treated with monensin. FEBS Lett. 208:34–38.
45. Sansonetti, P. J., and A. Phalipon. 1999. M cells as ports of entry for
VOL. 76, 2008 C. JEJUNI TRANSLOCATION ACROSS POLARIZED Caco-2 CELLS 5303
at FD
A B
IOS
CIE
NC
ES
LIBR
AR
Y on O
ctober 30, 2008 iai.asm
.orgD
ownloaded from

enteroinvasive pathogens: mechanisms of interaction, consequences for thedisease process. Semin. Immunol. 11:193–203.
46. Skirrow, M. B., and M. J. Blaser. 2002. Campylobacter jejuni, p. 719–739. InM. J. Blaser, P. D. Smith, J. I. Ravdin, H. B. Greenberg, and R. L. Guerrant(ed.), Infections of the gastroitestinal tract, 2nd ed. Lippincott Williams andWilkins, Philadelphia, PA.
47. van Alphen, L. B., N. M. C. Bleumink-Pluym, K. D. Rochat, B. W. M. vanBalkom, M. M. S. M. Wosten, and J. P. M. van Putten. 2008. Active migra-tion into the subcellular space precedes Campylobacter jejuni invasion ofepithelial cells. Cell. Microbiol. 10:53–66.
48. Vazquez-Torres, A., J. Jones-Carson, A. J. Baumler, S. Falkow, R. Valdivia,W. Brown, M. Le, R. Berggren, W. T. Parks, and F. C. Fang. 1999. Extrain-testinal dissemination of Salmonella by CD18-expressing phagocytes. Nature401:804–808.
49. Walker, R. I., E. A. Schmauder-Chock, J. L. Parker, and D. Burr. 1988.
Selective association and transport of Campylobacter jejuni through M cellsof rabbit Peyer’s patches. Can. J. Microbiol. 34:1142–1147.
50. Wassenaar, T. M., N. M. Bleumink-Pluym, and B. A. van der Zeijst. 1991.Inactivation of Campylobacter jejuni flagellin genes by homologous recom-bination demonstrates that flaA but not flaB is required for invasion. EMBOJ. 10:2055–2061.
51. Watson, R. O., and J. E. Galan. 2008. Campylobacter jejuni survives withinepithelial cells by avoiding delivery to lysosomes. PLoS Pathog. 4:e14.
52. Yanagawa, Y., M. Takahashi, and T. Itoh. 1994. The role of flagella ofCampylobacter jejuni in colonization in the intestinal tract in mice and thecultured-cell infectivity. Nippon Saikingaku Zasshi 49:395–403.
53. Yao, R., D. H. Burr, P. Doig, T. J. Trust, H. Niu, and P. Guerry. 1994.Isolation of motile and non-motile insertional mutants of Campylobacterjejuni: the role of motility in adherence and invasion of eukaryotic cells. Mol.Microbiol. 14:883–893.
Editor: B. A. McCormick
5304 HU ET AL. INFECT. IMMUN.
at FD
A B
IOS
CIE
NC
ES
LIBR
AR
Y on O
ctober 30, 2008 iai.asm
.orgD
ownloaded from



















