
Misguided Transcriptional Elongation Causes Mixed Lineage Leukemia

EXPERIMENTAL CELL RESEARCH 227, 135–145 (1996)ARTICLE NO. 0258
Embryonic Lineage of Vascular Smooth Muscle Cells DeterminesResponses to Collagen Matrices and Integrin Receptor Expression
SHELDON L. THIESZEN,*,1 MICHAEL DALTON,†,1 PRESTON F. GADSON,†,2
ERIC PATTERSON,‡ AND THOMAS H. ROSENQUIST*,†
*Department of Pathology and Microbiology and †Department of Cell Biology and Anatomy, University of Nebraska Medical Center,Omaha, Nebraska 68198-6395; and ‡Creighton University, Department of Biomedical Sciences, Omaha, Nebraska 68990
presence of vascular smooth muscle cells (VSMC) de-Developmental studies have demonstrated that the rived from the neural crest [1–3]. Matrix pattern for-
vascular smooth muscle cells (VSMC) present within mation within the aortic arch arteries occurs in a sys-the elastic arteries are differentiated from two defini- tematic and predictable fashion to produce well-demar-tive origins, the neural crest and the mesoderm. Cells cated elastic laminae [1], highly ordered helices offrom these distinct progenitors differ in their ability collagen [4], and fibril-associated proteoglycan [5]. Mi-to determine long-range spatial order of the extracel- crosurgical removal of premigratory neural crest re-lular matrix, in proliferative responses, and in the ex- sults in the replacement of the normal VSM progenitorpression of critical proteins. The present study utilizes cells in the great vessels by surrogate cells from thecollagen gel contraction assays and the analysis of in- mesoderm, and the elastic extracellular matrix is con-tegrin receptor subunit expression to evaluate cell– genitally disordered [4–6].matrix interactions. In the presence of serum and
The absence of orderly matrix pattern formation intransforming growth factor-b1 (TGF) or TFG-b1 alone,neural crest-ablated embryos suggests a fundamentalVSMC isolated from the abdominal aorta (AA-VSMC)difference in the ability of VSMC derived from the neu-were found to contract collagen matrices to a signifi-ral crest vs those derived from mesoderm to establishcantly greater extent than VSMC from the thoracicspatial order and/or to interact with associated extra-aorta (TA-VSMC). However, in TA-VSMC, b1 integrincellular matrix proteins. In addition, regional differ-and gel contraction were stimulated only in the pres-ences in pattern formation between the aortic arch ar-ence of serum factors. Metabolic labeling and immuno-teries and more distal mesenchymal arteries in normalprecipitation of integrin subunits revealed that TGF-
b1 induced b1 and a5 integrin subunits in AA-VSMC development support these cell-specific differences. Tofour- and ninefold, respectively. AA-VSMC gel contrac- examine the mechanisms of matrix patterning by neu-tion stimulated by serum and TGF-b1 alone was inhib- ral crest and mesenchymal-derived VSMC, our labora-ited with anti-b1 integrin antibody by 70 and 100%, re- tory has undertaken studies evaluating these two typesspectively. However, the b1 integrin-specific antibody of VSMC isolated from the thoracic aorta (TA) and ab-inhibited serum-induced TA-VSMC gel contraction by dominal aorta (AA), respectively. Previously, TA-25%. The data suggest that vascular smooth muscle cell VSMC and AA-VSMC have been shown to differ in cellontogeny is an important determinant of cell function, proliferation and the activation of signal transductionphenotype, and response to growth factors such as cascades when treated with transforming growth factorTGF-b1 . q 1996 Academic Press, Inc.
b1 (TGF-b1) [7]. In TA-VSMC, activation of protein ki-nase C by translocation to the cellular membrane fol-lowed treatment with this growth factor; this cellularINTRODUCTIONresponse was absent in AA-VSMC. Similarly, the ex-pression of the c-jun, elastin, and b-actin genes hasDuring the development of the cardiac outflow tract,been shown to differ between these VSMC types [8].a high degree of order is established within the extra-More recently, our laboratory has reported the selectivecellular matrix, a process that is dependent upon theexpression of the proto-oncogene c-myb in TA-VSMCand examined its role in gene expression [9].
1 These authors contributed equally to the manuscript and should Although the mechanism of three-dimensional pat-be considered co-first authors.tern formation in the extracellular matrix is not yet2 To whom correspondence and reprint requests should be ad-
dressed. understood completely, integrin receptors are known
135 0014-4827/96 $18.00Copyright q 1996 by Academic Press, Inc.
All rights of reproduction in any form reserved.
AID ECR 3254 / 6i12$$$301 07-29-96 20:53:00 ecl AP: Exp Cell

136 THIESZEN ET AL.
between 4 and 6 h with a final concentration of 2–5 mCi/ml [3H]-to mediate interactions between cells and several sig-thymidine added at the times and for the periods indicated in thenificant components of their associated matrix. There isfigure legends. The cells were washed, and the incorporation of [3H]-
abundant in vitro [10–12] and in vivo [13–15] evidence thymidine into acid-insoluble material was determined as previouslythat integrin receptors function in the organization and described [7].remodeling of the extracellular matrix. The integrins Collagen gel contraction. Three-dimensional collagen gel matri-expressed by VSMC during development of the aorta ces were prepared as described previously [10, 12, 24]. A collagen
stock solution (2.5 mg/ml in 0.1% acetic acid) was prepared fromare thus potentially of great significance to the processlyophilized collagen type I isolated from bovine skin (Sigma). Embry-determining pattern formation of the elastic laminae.onic VSMC maintained in culture were detached with trypsin-EDTA,The treatment of a variety of cell types with TGF-b1 washed twice in Dulbecco’s phosphate-buffered saline (DPBS) with-
can readily alter the repertoire of integrin receptors out Ca2/ or Mg2/, and pelleted by centrifugation. Cell numbers weredetermined with quadruplicate cell counts using a hemocytometer.expressed [16–20]. These data, and the differential cel-Upon the final wash, cells were resuspended at the desired concen-lular activation by TGF-b1 of VSMC from different em-tration; in the majority of experiments, cell concentration was 5 1bryonic lineages, support integrin receptors as poten-106 cells/ml. A volume of 0.3 ml of the cell suspension was mixed
tial mediators of the differences in matrix pattern for- with 1.95 ml of gel-mix buffer containing 1.75 1 M199/0.1 N NaOH/mation. FBS (72:8:20). Finally, 0.75 ml of collagen stock solution was added
and 1.5 ml of the complete gel solution was immediately transferredThe purpose of this study was to investigate the in-into 35-mm bacteriologic petri dishes. Gel formation occurred withinteraction of TA-VSMC and AA-VSMC with collagenthe first hour of culture at 377C and gels were released from theand model the differences in matrix pattern formation, petri dish by gentle tapping. Gels were evaluated at 6-h intervals
utilizing three-dimensional collagen gels. Contraction for up to 48 h. Contraction was measured as the diameter of the gelof three-dimensional collagen matrices has been used ring in millimeters, using a magnifying loop.
To analyze the effects of TGF-b1 on collagen gel contraction, cellsextensively to model the ability of fibroblasts to reorga-were prepared as above and mixed with a serum-free gel-mix buffer,nize connective tissues [for review, see 21]. Here we1.75 1 M199/0.1 N NaOH/DPBS with 7.5% bovine serum albuminreport that AA-VSMC contracts a collagen matrix to (BSA). TGF-b1 (Sigma) was added to the cell suspension and gel mix
a significantly greater extent than TA-VSMC in the prior to the addition of collagen. Control gels were not treated withpresence of serum or TGF-b1 . The expression of inte- the growth factor and contained only BSA. Collagen gel contraction
assays were also performed with each cell type in the presence of agrin subunits by both VSMC types has been evaluatedchick-specific b1 integrin monoclonal antibody (W1B10, Sigma) toto investigate a potential mechanism for differences inevaluate the role of this integrin subunit in gel contraction. For thesematrix pattern formation. The basal levels of integrin inhibition studies, cells were washed, counted, and resuspended at
receptor expression, and responses to TGF-b1 , differ the concentrations described. The cells were then mixed with the b1
integrin antibody and incubated on ice for 30 min. Cells were thenbetween AA-VSMC and TA-VSMC. These differencesmixed with the FBS or TGF-b1 gel buffers. Antibody concentrationsin receptor expression parallel data from collagen gelwere based on total volume of all reagents in a complete gel andcontraction assays, and together, these results suggestranged from 0.01 to 25 mg/ml. Inhibition of gel contraction was deter-
that VSMC ontogeny is a significant determinant of mined as a percentage of change in gel diameter (change in diametercell phenotype and function. of W1B10 gels/change in diameter of control gels). Statistical analysis
was performed using a t test on paired data and statistical signifi-cance was determined at P õ 0.01.MATERIALS AND METHODS
Metabolic cell labeling and immunoprecipitation. VSMC wereCell culture. Chick embryo VSMC were isolated for primary cell prepared for experiments by passaging cells at high densities (4–5
culture by enzymatic digestion of vessels isolated from fertilized Hy- 1 104 cells/cm2), resulting in uniformly confluent monolayers withinline chicken eggs incubated for 14–18 days. The thoracic aorta and 2–3 days. Confluent cultures were washed twice with DPBS andproximal portions of the brachiocephalic arteries were dissected from placed in serum-free medium without methionine and cysteine /embryos to obtain VSMC differentiated from neural crest cells [22]. 0.1% BSA for 24–30 h. Integrin synthesis was stimulated with TGF-VSMC of mesodermal origin were isolated from the abdominal seg- b1 (2 ng/ml). Metabolic labeling of proteins was carried out in time-ments of the aorta distal to the renal arteries. Connective tissue was course studies with 180–200 mCi/ml [35S]methionine (Trans Label-removed from the harvested vessels under a dissecting microscope ICN) for the final 6 h of growth factor treatment.and the arteries were digested with collagenase and elastase [23]. For the isolation of cellular proteins, cultures were rinsed withThe digested tissues were then washed and treated with 0.05% tryp- DPBS and scraped in RIPA buffer (20 mM Tris–HCl, 1 mM EDTA,sin-EDTA at 377C for 10 min. The trypsin digestion was stopped and 150 mM NaCl at pH 7.5) containing protease inhibitors (1 mMwith media containing 10% fetal bovine serum (FBS). Following cen- phenylmethylsulfonyl fluoride and 0.15 mM aprotinin). This cell sus-trifugation, the pellet was resuspended in Medium 199 (M199) sup- pension was centrifuged and the pellet resuspended in RIPA bufferplemented with penicillin/streptomycin (100 units/ml), L-glutamine / protease inhibitors and 1% Nonidet-P40. Following brief sonica-(2 mM ), Hepes buffer, and 10% FBS and seeded into flasks as tissue tion, cell suspensions were incubated on ice and then centrifuged atexplants. VSMC were utilized for experiments at the second or third 15,000g for 30 min. An aliquot of the supernate was utilized to deter-cell passage. For analysis of cell culture purity, cells were plated mine incorporation of radiolabel by scintillation counting. Samplesin four-well chamber slides (Lab Tech) and confirmed as VSMC by were normalized using the scintillation counts and precleared bymorphology and by their reaction with an antibody specific for adding 10 mg normal rabbit IgG and/or 50 ml of protein A–Sepharosesmooth muscle a-actin (Sigma). at 47C for 1.5 h and centrifuging at 15,000g for 5 min.
Antibodies to specific integrin subunits (10 mg) were then addedDNA synthesis. Growth factors were added directly to quiescentVSMCs in Medium 199 as described above. The cells were pulsed to the supernates and incubated overnight at 47C with continuous
AID ECR 3254 / 6i12$$$301 07-29-96 20:53:00 ecl AP: Exp Cell

137ONTOGENY AND VASCULAR SMOOTH MUSCLE CELL PHENOTYPE
agitation. Following a 2- to 4-h incubation with 20 ml of protein A– tions. VSMC seeded into collagen type I gels contractSepharose (Sigma), the immune complexes and beads were pelleted and reorganize the collagen matrix, yielding a denseby centrifugation for 5 min at 15,000g. The pelleted beads were
collagen ‘‘disc.’’ Initial studies revealed that the con-washed five times with RIPA wash buffer (20 mM Tris, 1 mM EDTA,traction process was dependent on cell numbers as0.15 M NaCl, 1% Tween, and protease inhibitors) and then boiled
for 5 min in 50 ml RIPA wash buffer containing 2 mM b-mecaptoetha- previously reported [24, 27]. Subsequent studiesnol. The Sepharose was pelleted by centrifugation and the supernate were performed with 7.5 1 105 cells/gel, which pro-mixed with loading buffer (42.5 mM Tris, 15% glycerol, 9% sodium duced a rapid and very consistent contraction of thedodecyl sulfate, and 0.05% bromophenol blue). Samples were ana-
collagen matrix. Time-course studies in the presencelyzed by polyacrylamide gel electrophoresis on 7–7.5% gels, and theof TGF-b1 or serum showed that contraction of thegels were dried and used for autoradiography or exposure to a Phos-
phorimager (Molecular Dynamics). Antibodies used for immunopre- gels was detectable after 6 h,that most of the gel con-cipitation of integrin subunits were the W1B10 antibody specific for traction occurred during the first 12 h, and that bythe b1 subunit and polyclonal antibodies to the a5 and a3 subunits 24 h the process was essentially complete (Fig. 1A).(Chemicon).
When comparing the contraction of collagen gels byAnalysis of b1 integrin mRNA. Confluent monolayers of VSMCthe two types of VSMC in the presence and absencewere obtained as described above and were blocked in serum-freeof TGF-b1 , it was apparent that AA-VSMC producedmedia (M199 / 0.1% BSA) for 24–30 h prior to treatment with TGF-
b1 . RNA was extracted from growth factor or serum plus TGF-b1- a greater reduction in gel diameter. Cell numberstreated cells by rinsing the cell layers twice and adding 1 ml of Tri were determined by averaging quadruplicate countsReagent (Molecular Research Center Inc., Cincinnati, OH) directly and experiments were repeated multiple times toto the monolayer. Culture dishes were scraped and the cell lysate
eliminate the possibility that this effect was the re-was extracted with chloroform and isopropanol precipitated. RNAwas used directly for reverse transcription–polymerase chain reac- sult of errors in cell numbers. The extent of the differ-tion (RT-PCR) or stored at 707C before use. Reverse transcription ences between AA- and TA-VSMC-mediated gel con-was performed on 100 ng of total cellular RNA in a 30-ml reaction traction varied in the assays utilizing serum. How-volume for 3 h at 377C. The reaction consisted of 800 U Superscript
ever, when TGF-b1 was added to the cultures, theII and 3 ml of 101 reaction buffer (Life Technologies), 10 mM dithi-contraction of collagen gels by AA-VSMC remainedolthreitol, 200 mM dATP, dCTP, dGTP, and dTTP, and 150 ng of
random hexamer primers. Upon completion of the reverse tran- statistically greater than that by TA-VSMC (Fig. 1).scriptase reaction, 3 U of RNase H was added to each reaction, incu- As shown in Fig. 2A, contraction assays were per-bated for 60 min at 377C, and then boiled for 10 min to terminate the formed in the presence of TGF-b1 , but serum wasRNase reaction. Aliquots of 10 ml were taken from these completed
omitted. Serum-free collagen gels supplemented withreactions for RT-PCR amplification. PCR was carried out in 100-ml7.5% BSA were prepared as described under Materi-reaction volumes containing the following components: 2.5 U Taq
DNA polymerase and 101 reaction buffer (Life Technologies), 200 als and Methods. TGF-b1 was added directly to themM dATP, dCTP, dGTP, and dTTP, 0.1 mM each oligonucleotide cell suspension and incorporated within the collagenprimer, and 2.5 mM magnesium chloride. The reaction cycles were gel. The influence of TGF-b1 on collagen gel contrac-957C for 1 min, 507C for 1 min, and 727C for 3 min. The final cycle
tion was analyzed through time-course and concen-concluded with a 7-min extension at 727C. Amplified products weretration experiments. The treatment of both VSMCanalyzed by electrophoresis on 2% agarose gels and stained with
ethidium bromide for visualization. Band intensity of negative im- types with TGF-b1 revealed that AA-VSMC wereages was scanned and quantified using Molecular Dynamics instru- stimulated to contract the collagen matrix to aments and software. greater extent. The results showed a dose-dependentThe quality of RNA was evaluated by the amplification of reverse
increase in gel contraction. At concentrations of 0.05,transcription products using primers designed to amplify b-actin.Primer sequences were 5*-AAGAGGTATCCTGACCCTGAA-3 * for 0.1, and 1 ng/ml, gel contraction mediated by AA-the upstream primer and 5*-ACCTGACCATCAGGGAGTTCA-3 * for VSMC was significantly greater than TA-VSMC gelthe downstream primer as previously reported [25]. This primer pair contraction. Mean diameters from four experimentsresults in the amplification of a 557-bp fragment which was used to
for AA-VSMC were 21 mm for 0.05 ng/ml and 14 mmnormalize the amount of RNA in all reactions. The expression of b1
at 0.1 and 1 ng/ml, respectively, vs 30.9 mm for TA-integrin mRNA was evaluated by a primer set which was designedagainst the chick integrin cDNA sequence [26]. The upstream and VSMC at all the concentrations examined.downstream primers were 5*-ATGGCCGAGACTAATTTAA-3 * and Inhibition of collagen gel reorganization by a b15*-TCTCAGTTTGGCTGGTGTTGT-3 *, respectively. Amplification integrin antibody (W1B10) was utilized to assess thewith this primer set produces a 623-bp fragment.
contribution of this matrix receptor to gel contrac-tion. The W1B10 antibody is directed to the extracel-
RESULTS lular domain of the b1 integrin and can disrupt cellu-lar adhesion to extracellular matrix proteins. VSMCReorganization of Collagen Matriceswere incubated with the antibody prior to incorpora-tion into collagen gels. Collagen gels were otherwiseTo investigate the differences we have reported in
matrix pattern formation by TA-VSMC and AA- prepared as in previous experiments. AA-VSMC cul-tures incubated with TGF-b1 induced gel contractionVSMC, we have utilized collagen gel contraction
assays as an in vitro model of cell–matrix interac- 20% less than serum-treated cells; however, AA-
AID ECR 3254 / 6i12$$$301 07-29-96 20:53:00 ecl AP: Exp Cell

138 THIESZEN ET AL.
FIG. 1. Analysis of VSMC collagen gel contraction in response to serum and TGF b1 . (A) AA-VSMC and TA-VSMC were incorporatedinto a three-dimensional collagen matrix in the presence of BSA, TGF-b1 , or 10% FBS at the times indicated along the abscissa. Gels wereprepared in the absence of serum by combining cells with gel buffer, collagen type I, and PBS containing 7.5% BSA. Gel diameter wasmeasured after 6, 12, and 24 h of incubation. Values reported within the graph represent the mean gel diameters obtained in eightconsecutive experiments. (B) Example of collagen gel contraction by the two types of VSMC. Control gels containing FBS or BSA alonewere run in parallel with gels treated with TGF-b1 during a 24-h incubation. Photographs were taken after 24 h of culture.
VSMC gel contraction stimulated by TGF-b1 was in- gels, but maximum inhibition in TA-VSMC gels wasonly 25% (Figs. 2C and 2D). The mean diameters ofhibited 100% by the W1B10 antibody (Figs. 2B and
2C). In the presence of serum, the W1B10 antibody control and antibody-treated gels were statisticallydifferent (P õ 0.01) for both AA- and TA-VSMC. Thisinhibited gel contraction by up to 70% in AA-VSMC
AID ECR 3254 / 6i12$$$301 07-29-96 20:53:00 ecl AP: Exp Cell
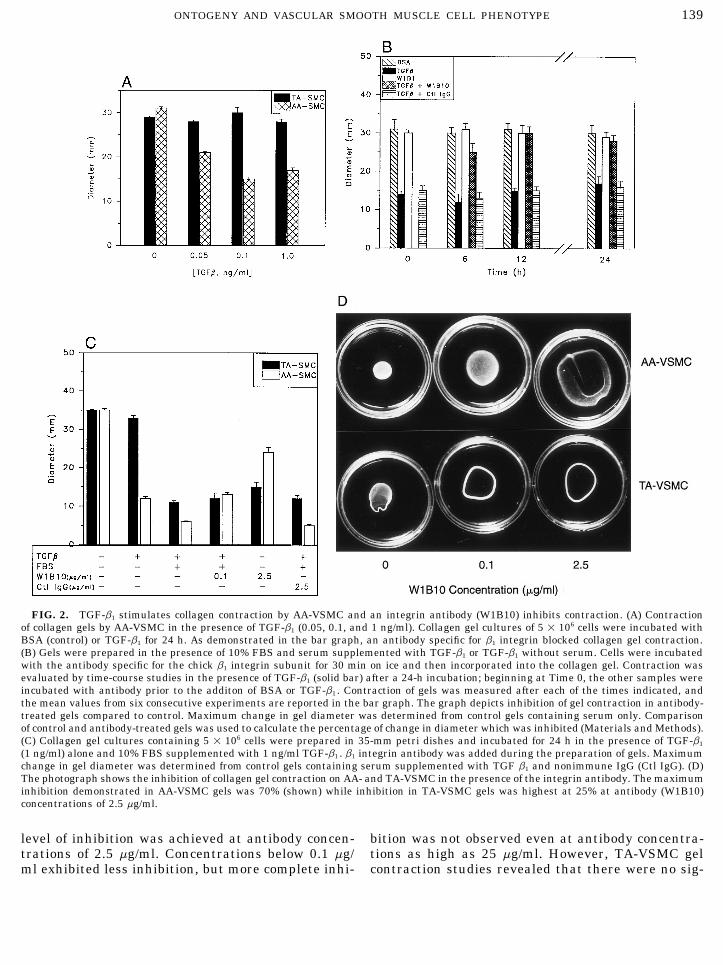
139ONTOGENY AND VASCULAR SMOOTH MUSCLE CELL PHENOTYPE
FIG. 2. TGF-b1 stimulates collagen contraction by AA-VSMC and an integrin antibody (W1B10) inhibits contraction. (A) Contractionof collagen gels by AA-VSMC in the presence of TGF-b1 (0.05, 0.1, and 1 ng/ml). Collagen gel cultures of 5 1 106 cells were incubated withBSA (control) or TGF-b1 for 24 h. As demonstrated in the bar graph, an antibody specific for b1 integrin blocked collagen gel contraction.(B) Gels were prepared in the presence of 10% FBS and serum supplemented with TGF-b1 or TGF-b1 without serum. Cells were incubatedwith the antibody specific for the chick b1 integrin subunit for 30 min on ice and then incorporated into the collagen gel. Contraction wasevaluated by time-course studies in the presence of TGF-b1 (solid bar) after a 24-h incubation; beginning at Time 0, the other samples wereincubated with antibody prior to the additon of BSA or TGF-b1 . Contraction of gels was measured after each of the times indicated, andthe mean values from six consecutive experiments are reported in the bar graph. The graph depicts inhibition of gel contraction in antibody-treated gels compared to control. Maximum change in gel diameter was determined from control gels containing serum only. Comparisonof control and antibody-treated gels was used to calculate the percentage of change in diameter which was inhibited (Materials and Methods).(C) Collagen gel cultures containing 5 1 106 cells were prepared in 35-mm petri dishes and incubated for 24 h in the presence of TGF-b1
(1 ng/ml) alone and 10% FBS supplemented with 1 ng/ml TGF-b1 . b1 integrin antibody was added during the preparation of gels. Maximumchange in gel diameter was determined from control gels containing serum supplemented with TGF b1 and nonimmune IgG (Ctl IgG). (D)The photograph shows the inhibition of collagen gel contraction on AA- and TA-VSMC in the presence of the integrin antibody. The maximuminhibition demonstrated in AA-VSMC gels was 70% (shown) while inhibition in TA-VSMC gels was highest at 25% at antibody (W1B10)concentrations of 2.5 mg/ml.
level of inhibition was achieved at antibody concen- bition was not observed even at antibody concentra-tions as high as 25 mg/ml. However, TA-VSMC geltrations of 2.5 mg/ml. Concentrations below 0.1 mg/
ml exhibited less inhibition, but more complete inhi- contraction studies revealed that there were no sig-
AID ECR 3254 / 6i12$$$301 07-29-96 20:53:00 ecl AP: Exp Cell

140 THIESZEN ET AL.
TABLE 1
Effect of TGFb1 on Proliferation of CulturedAA- and TA-VSMC
[3H]Thymidine incorporation (dpm/well)
AA TA AA TA
Diluent 4072 { 316 4143 { 410 4118 { 431 4067 { 385TGF-b
(1 ng/ml) 4182 { 394 4092 { 366 5245 { 474 4843 { 37012 h 24 h
Note. AA- and TA-VSMC were cultured and growth arrested for48 h before addition on 2 ng/ml TGF-b1 in the presence of type Icollagen. Incubation was continued for 12 and 24 h after the additionof 2 mCi/ml/[3H]thymidine. Data given are the mean { SEM of tripli-cate experiments from five experiments.
nificant differences between cells treated with serumalone and cultures incubated with serum supple-
FIG. 4. The expression of the b1 integrin subunit was evaluatedmented with TGF-b1 (data not shown). As demon-by immunoprecipitation. AA-VSMC and TA-VSMC cultures werestrated in Table 1 and Fig. 3, AA- and TA-VSMC grow grown to confluency and blocked for 24 h in serum-free media (0.1%
at the same rate in the presence of type I collagen. BSA) without methionine and cysteine. Cells were metabolically la-beled with [35S]methionine during the last 6 h of TGF-b1 treatment.These data suggest that the difference in collagen gelThe b1 subunit was then immunoprecipitated from detergent solublecontraction between AA- and TA-cells was indepen-cell fractions with the W1B10 antibody. Precipitated proteins weredent of cell growth.analyzed by polyacrylamide gel electrophoresis and gels were driedand evaluated with a Molecular Dynamics Phosphorimager. A repre-Immunoprecipitation of Integrin Subunitssentative image of the proteins immunoprecipitated from treated
The differences in collagen gel reorganization, and cultures is shown (A). As previously demonstrated by other investiga-tors, the b1 antibody precipitates proteins of 110 kDa (b1 subunit)the extent of W1B10 inhibition, suggested that the AA-and a subunits which migrate as a broad band of 140–160 kDa. (B)The relative intensity of the bands is graphed vs time. Basal levelsof the b1 subunit are twofold higher in AA-VSMC (Time Å 0), andwith TGF-b1 treatment, the levels in AA-VSMC (solid line) werefourfold higher than in TA-VSMC (dashed line) at 6 h. Expressionof the a subunits in the 140- to 160-kDa band followed a similar butslightly delayed time-course of expression.
VSMC and TA-VSMC might be expressing a differentcomplement of integrin receptors. Cell surface inte-grins expressed by VSMC might thus offer an explana-tion for the differences in matrix pattern formation dur-ing development of the aorta. Metabolic labeling andimmunoprecipitation of the b1 integrin protein ex-pressed by both VSMC types were undertaken to testthis hypothesis. Cells were blocked for a minimum of24 h using serum-free medium containing 0.1% BSAand then treated with TGF-b1 in a time-course study.Proteins were labeled with [35S]methionine during thefinal 6 h of growth factor treatment. Control cultureswere not treated with TGF-b1 and remained in M199/ 0.1% BSA. Immunoprecipitation of detergent-solubleFIG. 3. Cell growth of AA- and TA-VSMC in the presence of TGF-
b1 and collagen type I. Cells were cultured on type I collagen-coated cellular proteins with the W1B10 antibody produced adishes in growth media containing 7.5% BSA in the presence of 2 band of 100–110 kDa upon SDS–PAGE (Fig. 4). Inng/ml TGF-b1 for the times indicated. Cell numbers were counted addition, a broad band of 140–160 kDa, correspondingwith a hemacytometer, and each column represents the mean and
to a subunits, was coprecipitated as previously re-standard deviation of quadruplicate determinations from six experi-ments. ported by others [28–31].
AID ECR 3254 / 6i12$$$301 07-29-96 20:53:00 ecl AP: Exp Cell

141ONTOGENY AND VASCULAR SMOOTH MUSCLE CELL PHENOTYPE
Immunoprecipitation of cell lysates demonstrated approximately 6 h (Fig. 4). In AA-VSMC growth factortreatment resulted in a twofold increase in a expres-that even in the absence of serum or TGF-b1 , cultures
of AA-VSMC expressed substantially higher levels of sion at 12 h. Expression then declined to control levelsat 24 h of treatment. TA-VSMC responded to treatmentthe b1 subunit than TA-VSMC. Figure 3 is representa-
tive of the differences between these cell types. The with a slight but steady increase in a subunit expres-sion throughout the 24-h period, reaching levels equalamount of labeled protein immunoprecipitated from
AA-VSMC cultures was approximately three times that to the AA-VSMC. Previous studies have identified anddesignated the a subunits migrating within this bandisolated from TA-VSMC cultures. The timing and in-
duction of b1 expression following TGF-b1 treatment as a3 , a5a , and a5b [28], however, additional a subunitshave also been described [30]. In an attempt to furtherwas dramatically different between VSMC types. AA-
VSMC responded to growth factor treatment with a characterize these subunits, immunoprecipitation oftwo a integrin subunits was undertaken utilizing com-very rapid increase in b1 subunit expression (Fig. 4).
Expression was highest within the first 6 h of treat- mercially available antibodies reactive to avian pro-teins. Cells were blocked, treated, and labeled withment and declined with continued treatment, dropping
slightly below control levels at 24 h of treatment. At its [35S]methionine as before for evaluation in a time-course study. Detergent-soluble cell extracts were incu-peak, b1 expression in AA-VSMC was four- to fivefold
higher than the expression in TA-VSMC. The response bated with a3- and a5-specific polyclonal antiserum andimmunoprecipitated. Expression of the a5 subunit wasto TGF-b1 in TA-VSMC was more delayed, peaking at
12 h (1.5 times control levels) and rapidly returning to concordant with the pattern observed with the 140-to 160-kDa a band coprecipitated by the b1 integrinbaseline levels at 18 h.
The broad band of a subunits, which was coprecipi- antibody (Fig. 5A). The response to growth factor treat-ment was again different among AA-VSMC and TA-tated with the b1 subunit, also demonstrated that these
subunits were differentially regulated in TA-VSMC VSMC cultures. In AA-VSMC, the expression of a5 in-creased ninefold at 12 h and returned to near controland AA-VSMC. The expression of a subunits in re-
sponse to TGF-b1 followed a similar induction in time- levels during 24 h of treatment (Fig. 5B). This rapidand transient increase in a5 subunit expression wascourse studies as the b1 subunit, but was delayed byconfirmed in multiple experiments. In contrast, TA-VSMC responded to TGF-b1 treatment with a smallincrease (1.3 times control level) at 12 h and then stead-ily declined in a5 expression to below control levels.
The a3 subunit precipitated as a narrow band of ap-proximately 150 kDa and exhibited kinetics of expres-sion nearly opposite that of the a5 subunit (Fig. 6). AA-VSMC responded to growth factor stimulation with asteady 1.4-fold increase in a3 expression throughoutthe 24 h. Expression of a3 in TA-VSMC very rapidlyincreased to greater than eightfold above control levelsat 6 h, sixfold through 18 h, and remained at fourfoldat 24 h in the presence of TGF-b1 .
RNA Analysis
Total cellular RNA was harvested from confluentVSMC cultures following experimental treatments andreverse transcribed prior to PCR amplification with b-actin or b1 integrin primers. Experiments for AA-
FIG. 5. Immunoprecipitation of a5 integrin subunits in metaboli- VSMC were performed in serum-free medium. Ascally labeled AA-VSMC and TA-VSMC cultures. Detergent-soluble shown in Fig. 6, the induction of b1 integrin mRNA bycell fractions were immunoprecipitated with an a5-specific polyclonal
TGF-b1 in AA-VSMC was first observed at 0.05 ng/mlantibody (A). The expression of the a5 subunit was similar to thatand reached approximately sevenfold at 1 ng/ml. Inof the b1 subunit and the coprecipitated 140- to 160-kDa a complex,
with a peak in expression at 12 h of TGF-b1 treatment (B). In AA- TA-VSMCs, TGF-b1 induced b1 integrin mRNA at 1 ng/VSMC the expression of a5 was increased ninefold at 12 h and re- ml; however, concentrations of TGF-b1 as high as 5 ng/turned to near control levels during 24 h of treatment. In contrast, ml increased b1 integrin mRNA only 1.3-fold (data notTA-VSMC responded to TGF-b1 treatment with a slight increase (1.3
shown). The more than fivefold higher expression of b1times control level) at 12 h and a steady decline in a5 expression tobelow control levels. integrin mRNA in AA- vs TA-VSMC is supported by the
AID ECR 3254 / 6i12$$$301 07-29-96 20:53:00 ecl AP: Exp Cell

142 THIESZEN ET AL.
gree of collagen gel contraction of VSMC. In addition,we measured the expression of b1 integrin in TA-VSMCincubated with 10% FBS in the absence and presenceof TGF-b1 to determine whether there was a correlationbetween integrin expression and gel contraction. Asdemonstrated by the results in Fig. 7C, there was asixfold increase in b1 integrin transcripts in TA-VSMCincubated for 6, 12, and 24 h (lanes 1–4) with FBS asdetermined by RT-PCR. There was a slight increase inb1 integrin expression in the presence of TGF-b1 (lane6) compared to serum-treated cells without TGF-b1 .
DISCUSSION
Here we have shown that AA-VSMC and TA-VSMCdiffer in responses to three-dimensional collagen matri-ces and in the expression of integrin receptor subunitsin vitro. Two distinct embryological origins of VSMChave been recognized by laboratories studying neuralFIG. 6. Immunoprecipitation of the a3 subunit in AA-VSMC and
TA-VSMC. Cultures were treated and metabolically labeled as pre- crest cells for many years [32]. The significance ofviously described. (A and B) Immunoprecipitation of detergent-solu- VSMC ontogeny on cellular phenotype, however, hasble cell fractions with an a3-specific polyclonal antibody revealed that remained unknown. In our studies, the remarkable dif-AA-VSMC responded to growth factor stimulation with a 1.4-fold
ferences in matrix pattern formation within the greatincrease in a3 expression throughout the 24 h. Expression in TA-arteries following neural crest cell ablation in chickVSMC very rapidly increased to greater than eightfold that of control
levels at 6 h, sixfold through 18 h, and remained at a fourfold induc- embryos [4, 6], have suggested that neural crest- andtion at 24 h of TGF-b1 treatment. mesodermal-derived VSMC function differently in vivo.
Now, we offer an explanation for the differences ob-served in matrix pattern formation by analysis of inte-grin subunit expression.protein analyses. The complete analysis of b1 integrin
expression is presented in Figs. 7A and 7B. The PCR The reorganization of a collagen gel by fibroblastsresults in a matrix which grossly and histologically re-products were analyzed both by serial dilution and by
sequential cycles methods. The quantity of RNA ana- sembles the extracellular matrix of the dermis [24, 33]and has also been utilized as a model for tissue morpho-lyzed by sequential cycles was 100 ng, and the same
quantity was used for both TGF-b1-treated and control genesis during development [34]. The contraction ofcollagen gels by fibroblasts is mediated through thecultures. It is clear that both methods demonstrate a
comparable increase in b1 integrin mRNA from TGF- a2b1 integrin receptor [10, 11] and molecular studiesof integrin receptor function have shown that the cyto-b1-treated AA-VSMC. Highly expressed b-actin was
used to confirm the comparability of RNA amounts in plasmic domains of the a2 and a5 subunits are responsi-ble for the intracellular events required for the physicalRT-PCR. These data are in agreement with the immu-
noprecipitation studies and support the concept that contraction of collagen gels [12]. The most recognizedintegrin receptors for collagen are b1 subunits com-an increase in b1 integrin is directly related to the de-
FIG. 7. Analysis of b1 integrin mRNA in AA-VSMC and TA-VSMC by RT-PCR. (A) RNA from AA-VSMC cells with (solid circles) andwithout TGF b1 (open circles) treatment was analyzed by PCR. Results from analysis of quadruplicate serial dilutions and sequential cyclesare shown at the top and bottom, respectively. All serial dilutions were carried to 28 cycles. Sequential cycles were initiated with 100 ngof RNA in untreated and treated cells. (Bottom) AA-VSMC cultures were incubated without (solid circles) and with (open circles) 2 ng/mlTGF-b1 . (B) RT-PCR analysis of RNA from AA-VSMC using oligonucleotide primers complementary to the b1 integrin coding region. Cellswere treated without (lane 7) and with various concentrations of TGF-b1 : 0.01 ng/ml (lane 6); 0.02 ng/ml (lane 5); 0.05 ng/ml (lane 4); 0.1ng/ml (lane 3); 1 ng/ml (lane 2); 5 ng/ml (lane 1). (C) TA-VSMC were incubated in the absence (lane 1) and presence of serum for 6 h (lane2), 12 h (lane 3) and 24 h (lane 4). In addition, lanes 5 and 7 are samples from cells incubated in the absence of serum and TGF-b1 for 24h, respectively, and PCR products in lanes 6 and 8 are from cells treated with serum supplemented with 2 ng/ml TGF b1 and serum alonefor 24 h, respectively. b-actin PCR products are shown in lanes 1 through 7 to normalize for the amount of RNA used for each sample. Thesizes of molecular standards (M) are shown in B. For quantitation, an image (Molecular Dynamics) of a negative from a photograph ofethidium bromide-stained gels was obtained and stored by a personal computer. The area under the curve for each band was calculatedand plotted on the graphs shown.
AID ECR 3254 / 6i12$$$301 07-29-96 20:53:00 ecl AP: Exp Cell

143ONTOGENY AND VASCULAR SMOOTH MUSCLE CELL PHENOTYPE
AID ECR 3254 / 6i12$$3254 07-29-96 20:53:00 ecl AP: Exp Cell

144 THIESZEN ET AL.
bined with a1 , a2 , or a3 subunits [35]. In addition, a5 sive specificities and may have important implicationsfor the regulation of cell function in vivo. TGF-b1 medi-subunits isolated from chick embryo fibroblasts bind
monomeric and fibrillar collagen with specific affinities ated changes in the expression of individual integrinsubunits are likely to alter the balance of functional[30]. Under the experimental conditions reported here,
AA-VSMC were significantly more aggressive in con- integrin complexes expressed on the surface of the cell,modifying adhesive interactions that determine cell mi-tracting a collagen matrix. Inhibition studies using a
b1-specific antibody showed that AA-VSMC gel contrac- gration, cell phenotype, and orientation in tissues dur-ing development and morphogenesis. Apparently,tion mediated by TGF-b1 is blocked completely by this
treatment. However, in the presence of serum, gel con- TGF-b1 acts at the transcriptional as well as the post-translational level to stimulate terminal maturation oftraction of both cell types could be only partially inhib-
ited by the antibody. Previous studies of fibroblast col- a and b subunits and transfer of subunits to the plasmamembrane. In addition to its effects on the levels oflagen gel contraction have also observed only a partial
inhibition by b1 integrin antibodies [10, 11]. This may integrin a and b subunits, TGF-b1 elevates expressionof a1(I) procollagen in TA-VSMCs with slightly differ-be the result of additional integrin receptors participat-
ing in the process of gel contraction or inefficient ent kinetics [9]. However, antisense c-myb oligonucleo-tides inhibit the expression of both integrins and a1(I)blockage of integrin receptor interactions with colla-
gen. The differences observed between inhibition of AA- procollagen expression in TA-VSMC (unpublished ob-servations) [9]. AA-VSMC do not express c-myb [9].VSMC and TA-VSMC gels support the former possibil-
ity, because b1 induction is the same for both cell types This parallel effect of c-myb on the expression of inte-grins and procollagen suggests the presence of commonin the presence of serum and AA-VSMC gel contraction
is inhibited to a greater extent by the b1 integrin anti- regulatory elements which may control the expressionof these proteins. In AA-VSMC, TGF-b1 increases thebody. Obviously, these two cell types may express dif-
ferent integrin receptor repertoires for interacting with synthesis of a and b integrins with similar kinetics,indicating that the same factor(s) controls the expres-the extracellular matrix in response to TGF-b1 alone,
as is the case for AA-VSMC, or in the presence of serum sion of both polypeptides, but TGF-b1 has little or noeffect on procollagen expression in AA-VSMC [9].factors. These results indicate either a fundamental
difference in the TGF-b1 receptor signal cascade or dif- Throughout this report, we have referred to the cellsunder study by their site of anatomical origin (i.e., AA-ferences in the factors that control integrin gene tran-
scription. Furthermore, our data support a previous and TA-VSMC). These terms were used for clarificationto the reader not intimately familiar with embryologi-report by Villaschi et al. [36], who demonstrated that
intimal smooth muscle cells contract collagen gels to cal studies of VSMC origin. However, the terms meso-dermal-derived and neural crest-derived VSMC area greater extent than medial smooth muscle cells. In
combination with the present report, the current infor- more accurate in the description of these cell types.For example, neural crest cells differentiate into VSMCmation suggests that intimal and medial smooth mus-
cle cells may differ in embryonic origin. which populate the media of the thoracic aorta, proxi-mal pulmonary arteries, and brachiocephalic and ca-In this report, AA-VSMC were shown to express sub-
stantially larger quantities of b1 integrin and a sub- rotid arteries [2, 3, 32]. Therefore, we suggest thatthese latter terms are preferable for future referenceunits of 140–160 kDa which coprecipitated with the b1
subunit. These cells of mesodermal origin exemplified to VSMC and their embryological origin.VSMC of neural crest and mesodermal origin havethis phenotype under control conditions and in the
presence of TGF-b1 , which stimulated the synthesis of previously been shown to differ in proliferation andsignal transduction pathways [7] and expression of ma-both the b1 and the a subunits. Immunoprecipitation
of a5 and a3 subunits also revealed differences in the trix and cytoskeletal proteins [8]. Now, in an attemptto clarify differences in in vivo matrix pattern forma-subunit expression. The presence of the a5 subunit par-
alleled the expression of b1 ; however, the a3 subunit tion, we have shown differences in integrin expression.Additional studies have demonstrated differences inwas expressed at higher levels in TA-VSMC in response
to TGF-b1 . The higher levels of b1 and a5 integrin sub- proliferation, c-myb expression, and cytokine expres-sion by cells of these distinct origins [9]. In combina-unit expression by AA-VSMC correlated well with the
increased collagen gel contraction observed by this cell tion, these data support the embryological origin ofVSMC as a significant determinant of cell phenotype.type. It is difficult to determine if additional subunits
are involved, given the limited number of commercially It is important that in the study of vascular processeswe recognize these differences. A large volume of re-available antibodies which are reactive with avian pro-
teins. The coexpression of multiple a subunits with a search in smooth muscle cell biology is driven by at-tempts to understand the process of atherosclerosis. Acommon b1 subunit by AA- and TA-VSMC can result
in a repertoire of integrin complexes with distinct adhe- limitation of such studies is the lack of uniformity in
AID ECR 3254 / 6i12$$$301 07-29-96 20:53:00 ecl AP: Exp Cell

145ONTOGENY AND VASCULAR SMOOTH MUSCLE CELL PHENOTYPE
15. Larjava, H., Salo, T., Haapasalmi, K., Kramer, R. H., and Heino,VSMC samples. Interpretation of in vitro studies, andJ. (1993) J. Clin. Invest. 92, 1425–1435.extrapolation to an in vivo disease process, may be com-
16. Heino, J., Ignotz, R. A., Hemler, M. E., Crouse, C., and Mas-plicated by a failure to recognize the importance of on-sague, J. (1989) J. Biol. Chem. 264, 380–388.
togeny in determination of cell phenotype. 17. Janat, M. F., Argraves, W. S., and Liau, G. (1992) J. Cell. Phys-iol. 151, 588–595.
18. Cervella, P., Silengo, L., Pastore, C., and Altruda, F. (1993) J.REFERENCESBiol. Chem. 268, 5148–5155.
19. Kagami, S., Border, W. A., Ruoslaht, E., and Noble, N. A. (1993)1. Rosenquist, T. H., Fray-Gavalas, C., Waldo, K., and Beall, Lab. Invest. 69, 68–76.
A. C. (1990) Am. J. Anat. 189, 339–356.20. Gailit, J., Welch, M. P., and Clark, R. A. F. (1994) J. Invest.
2. Beall, A. C., and Rosenquist, T. H. (1990) Anat. Rec. 226, 360– Dermatol. 103, 221–227.366. 21. Grinnell, F. (1994) J. Cell. Biol. 124, 401–404.
3. Rosenquist, T. H., and Beall, A. C. (1990) Ann. N. Y. Acad. Sci. 22. Topouzis, S., Catravas, J. D., Ryan, J. W., and Rosenquist,588, 106–130. T. H. (1992) Circulation 71, 923–931.
4. Rosenquist, T. H., and Modis, L. (1991) Anat. Rec. 229, 116– 23. Foster, J. A., Rich, C. B., Horrigan, S., Miller, M., Dadd, C. A.,124. Baule, V., and Keene, D. R. (1988) Lab. Invest. 58, 667–673.
24. Bell, E., Ivarsson, B., and Merrill, C. (1979) Proc. Natl. Acad.5. Thieszen, S. L., and Rosenquist, T. H. (1995) Matrix Biol. 14,Sci. USA 76, 1274–1278.573–582.
25. Rugarli, E. I., Lutz, B., Kuratani, S. C., Wawersik, S., Borsani,6. Rosenquist, T. H., Beall, A. C., Modis, L., and Fishman, R.G., Ballabio, A., and Eichele, G. (1993) Nature Genet. 4, 19–26.(1990) Anat. Rec. 226, 347–359.
26. Tamkun, J. W., DeSimone, D. W., Fonda, D., Patel, R. S., Buck,7. Wrenn, R. W., Raeuber, C. L., Herman, L. E., Walton, W. J.,C., Horwitz, A. F., and Hynes, R. O. (1986) Cell 46, 271–282.and Rosenquist, T. H. (1993) In Vitro Cell. Dev. Biol. 29A, 73–
27. Guidry, C., and Grinnell, F. (1985) J. Cell Sci. 79, 67–81.78.28. Hynes, R. O., Marcantonio, E. E., Stepp, M. A., Urry, L. A., and8. Gadson, P. F., Rossignol, C., McCoy, J., and Rosenquist, T. H.
Yee, G. H. (1989) J. Cell. Biol. 109, 409–420.(1993) In Vitro Cell. Dev. Biol. 29A, 773–781.29. Hayashi, Y., Haimovich, B., Reszka, A., Boettiger, D., and Hor-
9. Gadson, P. F., Dalton, M. L., Patterson, E., Hutchinson, L., and witz, A. (1990) J. Cell. Biol. 110, 175–184.Rosenquist, T. H. (1995) Submitted for publication.
30. Potts, A. J., and Little, C. D. (1992) J. Cell. Physiol. 152, 558–10. Klein, C. E., Dressel, D., Steinmayer, T., Mauch, C., Eckes, B., 567.
and Krieg, T. (1991) J. Cell. Biol. 115, 1427–1436. 31. Enomoto, M. I., Boettiger, D., and Menko, A. S. (1993) Develop-11. Schiro, J. A., Chan, B. M. C., Roswit, W. T., Kassner, P. D., ment 155, 180–197.
Pentland, A. P., Hemler, M. E., Eisen, A. Z., and Kupper, T. S. 32. LeLievre, C. S., and LeDouarin, N. M. (1975) J. Embryol. Exp.(1991) Cell 67, 403–410. Morphol. 34, 125–154.
12. Chan, B. M. C., Kassner P. D., Schiro, J. A., Byers, H. R., Kup- 33. Grinnell, F., and Lamke, C. R. (1984) J. Cell Sci. 66, 51–63.per, T. S., and Hemler, M. E. (1992) Cell 68, 1051–1060. 34. Stopak, D., and Harris, A. K. (1982) Dev. Biol. 90, 383–398.
13. Albelda, S. M., and Buck, C. A. (1990) FASEB J. 4, 2868–2880. 35. Hynes, R. O. (1992) Cell 69, 11–25.36. Villaschi, S., Nicosia, R. F.,and Smith, M. R. (1994) In Vitro14. Cavani, A., Zambruno, G., Marconi, A., Manca, V., Marchetti,
M., and Giannetti, A. (1993) J. Invest. Dermatol. 101, 600–604. Cell. Dev. Biol. 30A, 589–595.
Received September 29, 1995Revised version received April 24, 1996
AID ECR 3254 / 6i12$$$301 07-29-96 20:53:00 ecl AP: Exp Cell
Copyright © 2022 FDOKUMEN




















