
Effects of past growth trends and current water use strategies on Scots pine and pubescent oak...
16
1 23 European Journal of Forest Research ISSN 1612-4669 Eur J Forest Res DOI 10.1007/s10342-013-0768-0 Effects of past growth trends and current water use strategies on Scots pine and pubescent oak drought sensitivity T. Morán-López, R. Poyatos, P. Llorens & S. Sabaté
Transcript of Effects of past growth trends and current water use strategies on Scots pine and pubescent oak...

1 23
European Journal of Forest Research ISSN 1612-4669 Eur J Forest ResDOI 10.1007/s10342-013-0768-0
Effects of past growth trends and currentwater use strategies on Scots pine andpubescent oak drought sensitivity
T. Morán-López, R. Poyatos, P. Llorens& S. Sabaté

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag Berlin Heidelberg. This e-offprint is
for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

ORIGINAL PAPER
Effects of past growth trends and current water use strategieson Scots pine and pubescent oak drought sensitivity
T. Moran-Lopez • R. Poyatos • P. Llorens •
S. Sabate
Received: 19 June 2013 / Revised: 26 November 2013 / Accepted: 6 December 2013
� Springer-Verlag Berlin Heidelberg 2013
Abstract Drought-induced decline is affecting Pinus syl-
vestris populations in southern Europe, with very little
impact on the more drought-tolerant Quercus pubescens.
Although multiple studies have investigated interspecific
differences in water use and growth strategies, the link
between these two processes and how they vary within
drought-exposed populations remains poorly understood.
Here, we analysed tree ring and sap flow data from P. syl-
vestris and Q. pubescens stands in the Pyrenees in order to
(1) evaluate differences in climate–growth responses among
species, (2) disentangle the role of past growth trends and
water use strategies in individual trees drought sensitivity
and (3) assess whether such intraspecific patterns vary
between species. Both species have suffered recent climatic
constraints related to increased aridity. However, the effects
of past growth trends and current water use traits on drought
sensitivity varied among them. Initially, fast-growing
‘drought-sensitive’ pines displayed a higher gas exchange
potential but were more sensitive to evaporative demand and
soil moisture. They also showed lower water use efficiency
for growth (WUEBAI) and current growth decline. In con-
trast, initially, slow-growing ‘drought-tolerant’ pines
showed the opposite water use traits and currently maintain
the highest growth rates. In comparison, neither current
WUEBAI nor recent growth trends varied across Q. pubes-
cens climate–growth groups. Nonetheless, ‘drought-sensi-
tive’ oaks showed the lowest gas exchange potential and the
highest growth rates under milder conditions. Our results
show a strong effect of past growth trends and current water
use strategies on tree resilience to increased aridity, which is
more evident in P. sylvestris.
Keywords Climate–growth relationships � Drought-
induced decline � Pinus sylvestris � Quercus pubescens �Sap flow � Water use efficiency (WUE)
Introduction
Climate change projections predict a general increment in
mean temperatures as well as changes in the seasonal dis-
tribution of precipitation in the Mediterranean basin (IPCC
2007). In the case of the Iberian Peninsula, a general trend
of temperature increments between 3 and 4 �C on average
accompanied by reduced precipitations from 10 to 25 % is
expected by the end of the century (Christensen and
Christensen 2007). Particularly, throughout the Pyrenees,
symptoms of more intense summer droughts and lower
winter precipitations have already been observed, and
enhanced water shortage is expected towards the end of the
present century (Lopez-Moreno and Nogues-Bravo 2005).
Communicated by R. Matyssek.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s10342-013-0768-0) contains supplementarymaterial, which is available to authorized users.
T. Moran-Lopez (&) � S. Sabate
Department of Ecology, University of Barcelona,
08028 Barcelona, Spain
e-mail: [email protected]
T. Moran-Lopez
Department of Biogeography and Global Change, National
Natural Science Museum (MNCN-CSIC), Serrano 115dpo.,
28006 Madrid, Spain
R. Poyatos � S. Sabate
CREAF, Cerdanyola del Valles, 08193 Barcelona, Spain
P. Llorens
Institute of Environmental Assessment and Water Research
(IDAEA-CSIC), Barcelona, Spain
123
Eur J Forest Res
DOI 10.1007/s10342-013-0768-0
Author's personal copy

Mediterranean montane forests are of particular interest
since changes in their composition can be used as early
global change sensors (Weber et al. 2007). The coexistence
of Eurosiberian populations at their southern distribution
limits and more xeric-adapted species makes these eco-
systems particularly sensitive to land use and/or climate
changes (Gimmi et al. 2010). For instance, a significant
shift from boreal Scots pines (Pinus sylvestris L.) to sub-
Mediterranean pubescent oaks (Quercus pubescens Willd.)
in dry valleys of the Alps has already been observed
(Gimmi et al. 2010; Rigling et al. 2013). In the Iberian
Peninsula, enhanced climatic constraints on radial growth
(Andreu et al. 2007; Martinez-Vilalta et al. 2008), and
drought-driven defoliation and mortality (Galiano et al.
2010; Heres et al. 2012) are all evident signs of Scots
pine’s intrinsic vulnerability to water deficits. Increased
competition due to the abandonment of management
practices may also aggravate the effects of drought on
demographic rates (Martinez-Vilalta et al. 2012; Vila-
Cabrera et al. 2011). Poor Scots pine regeneration in
declining populations together with an increased recruit-
ment of Quercus species (Galiano et al. 2010) points at a
potential substitution of Scots pine by more drought-tol-
erant species in dry, unmanaged sites (Vila-Cabrera et al.
2012).
Scots pine and pubescent oak show contrasting physio-
logical and growth responses to drought, which may
underlie directional vegetation shifts with increased aridity.
Under atmospheric and edaphic drought, Scots pine dis-
plays a strict stomatal control, while pubescent oak main-
tains moderate transpiration rates (Poyatos et al. 2008;
Zweifel et al. 2007) thanks to its tolerance of lower xylem
water potentials without suffering substantial cavitation
and its ability to extract water from deeper soil layers
(Valentini et al. 1992; Zweifel et al. 2007). Moreover,
Scots pine’s late bud-burst and growth initiation makes it
especially vulnerable to changes in summer water avail-
ability, as opposed to pubescent oak, which takes advan-
tage of higher water availability in spring by earlier radial
growth and leaf expansion (Zweifel et al. 2006). Accord-
ingly, Scots pine radial growth shows comparatively
stronger correlations with climatic variables associated
with summer water availability (Weber et al. 2007; Eil-
mann et al. 2011). In addition to species-specific variation,
climatic controls on radial growth can vary due to local
factors (e.g. Weber et al. 2007; Vachiano et al. 2012) and
also within a population (e.g. Heres et al. 2012). Moreover,
tree-level characteristics such as size, previous growth rate
or age mediate the effects of extreme drought events on P.
sylvestris growth (Martinez-Vilalta et al. 2012; Zang et al.
2012). In comparison, intrapopulation variability in Q pu-
bescens climate–growth responses has been comparatively
less studied.
Despite the existing wealth of dendroecological and sap
flow studies, the link between climate–growth relationships
and physiological regulation of tree water use has remained
relatively unexplored. In this regard, high water use effi-
ciency (WUE) for growth, estimated as the annual girth
increment per volume of water transpired (Breda and
Granier 1996), would probably confer a competitive
advantage in water-limited forests. Indeed, intrinsic WUE
measured in tree rings has shown considerable intrapopu-
lation variability, associated with drought-induced decline
processes in P. sylvestris (Voltas et al. 2013; Heres et al.
2013) and deciduous oaks such as Quercus robur (Levanic
et al. 2011).
Here, we analyse mid-term water use strategies and
long-term growth–climate relationships in Q. pubescens
and P. sylvestris stands in the pre-Pyrenees (NE Iberian
Peninsula), focusing on differences between species and
intrapopulation variability. Using a combination of den-
droecological and sap flow techniques, we (1) assessed
radial growth patterns of both species and their possible
relationships with climatic trends and (2) analysed intra-
population variability in drought sensitivity and its asso-
ciation with water use strategies and recent growth trends.
We hypothesised that P. sylvestris growth would be more
sensitive to reduce water availability, especially during the
recent decades, compared to Q. pubescens (H1). We also
postulated that both species would display intrapopulation
variation in drought responses (H2a), whereby drought-
sensitive trees would display higher initial growth rates and
a more strict stomatal control (H2b). As a result of
enhanced drought constraints in P. sylvestris, (H3) intra-
population variability in water use efficiency for growth
(WUEBAI) would be more tightly linked to growth sensi-
tivity to drought in P. sylvestris than in Q. pubescens.
Materials and methods
Study area
The experimental plots are part of the Vallcebre research
area (42�120N, 1�490E), located in the Eastern Pyrenees (NE
Iberian Peninsula). Climate is sub-Mediterranean, with an
average air temperature of 9 �C (measured at 1,260 m.a.s.l.)
and a mean annual precipitation of 826 ± 206 mm and an
annual potential evapotranspiration of about 823 mm (La-
tron et al. 2008). During the central part of the growing
season (April–August), potential evapotranspiration
(515.86 ± 4.14 mm, 1950–2010 mean ± SE) exceeds pre-
cipitation (470 ± 17.3 mm). Mudstone and sandstone are
the principal underlying lithologies in this area.
Three plots, all located within ca. 1.5 km from the vil-
lage of Vallcebre, were chosen for this study: two Scots
Eur J Forest Res
123
Author's personal copy

pine stands and one oak stand. The lower Scots pine stand
(Cal Sort plot) is located in an abandoned terraced slope, at
an elevation of 1,260 m.a.s.l. Soil depth is ca. 65 cm and
has a sandy-loam texture (Rubio 2005). Stand density is
2,165 stems ha-1, and leaf area index (LAI) is 2.4 m2 m-2
(Poyatos et al. 2008). Tree mean diameter at breast height
(dbh) is 15.0 ± 0.7 cm, and the estimated average age of
dominant and co-dominant trees is 64 ± 6 years (n = 20).
The upper Scots pine stand (Cal Parisa plot) is located in a
southeast-facing slope (10–30 % of hillslope gradient) at
1,400 m.a.s.l. Within this plot, the soil has a silty-loam
texture. Stand density is 2,400 stems ha-1, and LAI is ca.
2.0 m2 m-2 (Llorens et al. 1997). Mean dbh is 17.3 cm,
and age of dominant and co-dominant trees is 63 ± 5 years
(n = 15). The oak stand (Cal Barrol plot) is located at
1,100 m.a.s.l., on a substrate formed by a loamy matrix and
limestone blocks (Rubio 2005). Tree density is 828 trees
ha-1, and whole-stand LAI is 3.35 ± 0.5 m2 m-2 (Muzylo
et al. 2012). Tree mean dbh is 21.1 ± 1.4 cm. Based on
tree ring analyses of wood cores extracted at breast height
(n = 14), the age of Q. pubescens trees was estimated to be
at least ca. 80 years old. At the plot level, a surrogate for
intra-specific competition for belowground resources was
calculated as the number of neighbours within a buffer
distance of 2.5 and 5 m from individual trees. This index
was significantly correlated with distances to the nearest
neighbours in both pine and oak stands (Pearson test
-0.54, P \ 0.05 and -0.68, P \ 0.01, respectively). Hence,
distance to nearest neighbour was used to assess focal trees
competence status for water sources in further analyses.
Meteorological data
Meteorological monitoring of Vallcebre research catch-
ments started in 1989 (Llorens and Gallart 1992). In situ
measured rainfall (P, mm), air temperatures (T, �C) and
potential evapotranspiration (PET, mm) series (1989–2010)
were used in this study. Potential evapotranspiration, based
on the Penman–Monteith FAO method, was calculated as
described in Llorens et al. 2010.
In order to extend the climatic time series back in time
(period 1956–1988), Vallcebre climatic data were esti-
mated from nearby meteorological stations belonging to
the Spanish Meteorological Agency (AEMET) at Berga
(12 km from the study site) and La Molina (20 km from
the study site). We fitted generalised mixed models, and a
good agreement was found for both air temperatures
(R2 = 0.97) and precipitation (R2 = 0.84). During this
period (1956–1988), PET was estimated using the Thorn-
thwaite method and rescaled using a fitted relationship
obtained between Thornthwaite PET and Penman–Mon-
teith PET (R2 = 0.92). Two indexes of climatic drought
stress were calculated: (1) the ratio between annual
precipitation and potential evapotranspiration (P/PET;
UNEP 1992), which was used to analyse the drought
responses of tree growth and (2) a drought index (Di),
estimated as the difference between accumulated precipi-
tation and potential evapotranspiration from April to
August (Rigling et al. 2013). The value of Di was used to
compare the drought conditions in our study site with other
locations.
Tree ring chronology building and BAI calculation
In April 2010, 15 and 14 dominant or co-dominant trees
were sampled from the lower and the upper Scots pine
stands, respectively; 14 trees were sampled in the oak plot.
Mean dbh was 21.4 ± 2.0 cm for P. sylvestris and
23.9 ± 2.2 for Q. pubescens. Two cores were extracted to
the pith at breast height from each tree. All samples were
visually cross-dated and measured using Windendro soft-
ware (Regent Instrument Inc. 2002). The resulting series
underwent a cross-dating quality control with the statistical
programme COFECHA (Holmes, 1983). Subsequently,
series of the same stem were averaged, and annual basal
area increment estimated (BAI, mm2/year). A preliminary
analysis of growth trends in both Scots pine plots showed
that there were no significant differences between them
(P [ 0.05); therefore, all Scots pines were pooled together
for further analyses.
Sap flow, canopy stomatal conductance and water use
efficiency for growth
In the lower Scots pine plot and the oak plot, sap flow,
meteorological variables and soil moisture (0–30 cm) were
measured continuously and simultaneously from June 2003
to September 2005. A maximum of 14 P. sylvestris and 12
Q. pubescens were equipped with sap flow sensors. Tech-
nical details on the experimental setup of both plots can be
found in Poyatos et al. (2005). Sap flow density in the outer
xylem was measured using heat dissipation probes
according to Granier (1985). The procedure to upscale sap
flow in the outer xylem to the whole tree level was the
same for all trees and involved the correction for radial
variability in sap flow density and the estimation of sap-
wood depth using site-specific allometric relationships
(Poyatos et al. 2007; 2008). Tree leaf areas were also
obtained using local allometric relationships obtained from
forest inventory data (Gracia et al. 2000–2004; http://www.
creaf.uab.es/iefc/).
Time series of daily tree sap flow contained some
missing data (5.43 % on average). Since we were inter-
ested in total transpiration during the central part of the
growing season (June, July and August), days with missing
data were gap-filled using either fitted relationships
Eur J Forest Res
123
Author's personal copy

between sap flow of neighbouring trees (R2 = 0.68 on
average) or values for days with similar environmental
conditions.
Midday canopy stomatal conductance (Gs,md) was cal-
culated with the simplified Penman–Monteith equation for
aerodynamically rough canopies (Whitehead and Jarvis
1981):
Gs;md ¼ckJL;md
qcpDmd
ð1Þ
where c is the psychrometric constant (kPa K-1), k is latent
heat of vaporisation of water (J kg-1), q is air density
(kg m-3), cp is specific heat of air at constant pressure
(J kg-1 K-1), and D (kPa) is vapour pressure deficit, and
JL,md is leaf-area-based sap flow averaged for the midday
hours (11:00 to 14:00) and converted to molar units
(mmol m-2 s-1).
Water use efficiency for growth (Breda and Granier
1996) was calculated here as the individual basal area
increment divided by the volume of water transpired from
early June to the end of August (WUEBAI). As current-year
growth could depend on physiological processes occurring
during the previous year, WUEBAI was calculated based on
both current and previous-year transpiration.
Data analysis
All data analyses, unless otherwise stated, were carried out
using the R statistical software (2011). Linear mixed
models were fitted using nlme package (Pinheiro et al.
2012), selecting the model with the lowest Akaike infor-
mation criterion (AIC). In order to evaluate climate–growth
relationships, Pearson correlations between BAI and aver-
age air temperatures and total rainfall were estimated on a
monthly basis and for all individual trees. Preliminary
examination of mean annual temperatures showed consid-
erably more years with positive anomalies over the last
two, and BAI curves revealed more erratic and acute peaks
(decades compared to previous years 1956–1989) (Fig. 1).
Hence, water availability effects on tree growth in both
time periods (1956/1960–1990 and 1991–2010) were
assessed using linear mixed models. Our response variable
was ln(BAI ? 1), and predictor variables were summer
P/PET, time period and the interaction between both. Tree
identity was included as a random variable, and year was
introduced with a first-order autoregressive structure in
order to account for temporal autocorrelation. Subse-
quently, summer P/PET effects on growth were analysed
separately for both periods.
In order to evaluate intrapopulation variability in
drought sensitivity, individual climate–growth correlation
coefficients were introduced into a principal components
analysis. In this analysis, only climatic variables which had
shown the highest signals on tree growth during the entire
period (1960–1990) were considered. The first component
of the PCA (PC1) explained 61 and 58 % of the variance
for P. sylvestris and Q. pubescens, respectively. PC1 was
thus interpreted as a measure of drought tolerance since it
was highly related to individual trees’ responses to summer
temperature and precipitation. In both species, trees which
loadings in the first component were clearly related to
Fig. 1 a Annual rainfall (mm) and b mean annual temperature (�C)
from 1956 to 2010. Continuous line indicates linear regression of
temporal trends. c Basal area increment (mm2/year, mean ± standard
error) of Q. pubescens (open circles) and P. sylvestris (black circles)
Eur J Forest Res
123
Author's personal copy

positive temperature effects on growth and lower summer
precipitation responses were considered ‘drought tolerant’,
those which displayed the opposite pattern were considered
‘drought sensitive’ and those with loadings close to zero
were considered as ‘intermediate’ (cf. ‘Results’ section).
Additionally, a k-means analysis was performed so as to
ensure climate–growth groups coherence. PCA and k-
means analysis were carried out with Primer 6 (Primer-e
Ltd, Lutton, UK) and Gingko software (De Caceres et al.
2007), respectively.
Linear models were used to disentangle the relative
effect of water use strategies and initial growth rates
(ln(BAI60–90 ? 1) from 1960 to 1990) on individual trees
drought tolerance. Our response variable was individual
trees’ projection on the first axis of the PCA in the direction
of positive temperature effects (PC1 for oaks and -PC1 for
pines), and the explanatory variables were individual pines
WUEBAI, ln(BAI60–90 ? 1) and their interaction. Besides,
changes in competitive status of trees according to drought
tolerance and time period were tested through linear mixed
models. Our response variable (BAIrel) was calculated as:
BAIrel ¼ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
BAIi�
BAI
q
ð2Þ
where BAIi is individual tree BAI and BAI is average stand
BAI. Predictor variables were drought-tolerance group
membership, time period and both variables interaction.
Tree identity was introduced as random effect.
In order to relate drought sensitivity of growth with tree-
specific stomatal control of gas exchange, we summarised
the variation in Gs,md with respect to vapour pressure
deficit and soil water content, using two well-known
mathematical formulations with strong physiological
foundations. The sensitivity of Gs,md to vapour pressure
deficit (D, kPa) was analysed using the following rela-
tionship (Oren et al. 1999):
Gs;md ¼ Gs;ref � m ln D ð3Þ
This equation was fitted using the quantile regression
technique as implemented in the R package quantreg
(Koenker Team 2008). We chose s = 0.95 (i.e. 95th
quantile), and we assumed that it was representative of the
relationship between Gs,md and ln D when other factors
were optimal (Cade and Noon 2003). The obtained
intercept and slope equalled Gs,ref (reference Gs at
D = 1 kPa) and m (sensitivity to vapour pressure deficit,
-dGs,md/d ln D), respectively (Oren et al. 1999). Only data
with average midday D [ 0.6 kPa were analysed, to avoid
potential errors in Gs estimation under low evaporative
demand (Ewers and Oren 2000).
Canopy conductance usually shows a threshold-like
response to soil water availability (e.g. Granier et al. 2000).
The response of Gs,md to soil water content was thus
analysed using a linear segmented regression, which
identifies the break point in soil water content below which
Gs,md starts to decline in a linear fashion (Muggeo 2008).
Preliminary analyses for Q. pubescens showed very little
sensitivity of Gs,md to soil moisture within 0–30 cm, in line
with previous results (Poyatos et al. 2008); hence, we only
carried out the analyses for Scots pine. The soil water
content threshold for transpiration decline was thus con-
sidered as an indicator of tree-level sensitivity to water
availability in the soil.
Results
Climate trends
Annual rainfall in the study site (Fig. 1a) significantly
decreased during the period 1956–2010 (R2 = 0.18,
P \ 0.05) with an average reduction of 5.75 mm/year. For
the same period, mean annual temperature (Fig. 1b) sig-
nificantly increased (R2 = 0.32, P \ 0.05), at an average
rate of 0.02 �C/year. Besides, the analysis of mean annual
temperatures revealed two different periods over the last
decades with respect to the frequency of positive temper-
ature anomalies. Between 1956 and 1990, 14.3 % of the
years showed positive temperature anomalies while it
amounted to 40 % over the 1991–2010 period. Such dif-
ferences between periods were not found for the frequency
of negative anomalies of annual rainfall (23.5 and 33.3 %
of years with negative anomalies, respectively). Regarding
summer drought, in the 1956–1989 period, the value of Di
(mean ± SE) was 4.11 mm ± 21.83, while during
1991–2010 Di dropped to -126 ± 31.06 mm and
remained below -300 mm for 14.2 % of the years.
Growth patterns and climate–growth relationships
Growth trends differed between species at the beginning of
the study period, but, from the 1970s onwards, P. sylvestris
and Q. pubescens showed similar growth trends. The time
series of BAI did not reveal an acute average loss of growth
potential for any species, but, over the last decades, an
enhanced jagged pattern in BAI was observed (Fig. 1c).
Pinus sylvestris BAI was correlated with both previous-
year and current-year climatic conditions (Fig. 2a, c).
During the 1960–1990 period, annual growth was mainly
positively correlated with winter temperatures (Fig. 2a). In
contrast, negative correlations between BAI and tempera-
ture were typically observed during the 1991–2010 period,
especially for current summer months. Moreover, the weak
relationships between BAI and rainfall observed during
1960–1990 turned to high, positive correlations during the
1991–2010 period (Fig. 2c). Accordingly, summer P/PET
Eur J Forest Res
123
Author's personal copy

only showed positive, significant effects on P. sylvestris
growth over the last twenty years. Besides, a change in
growth sensitivity to summer P/PET over the last decades
was detected (Table 1).
Likewise, Q. pubescens climate–growth relationships
embraced both previous-year and current-year conditions
(Fig. 2). During the 1960–1990 period, positive correla-
tions between BAI and monthly temperature were gener-
ally observed; particularly, warmer springs were
accompanied by enhanced growth. In comparison, during
the 1991–2010 period, more negative correlations were
found (Fig. 2b). In contrast, an increased frequency of
positive correlations between BAI and precipitation was
observed during the 1991–2010 period, especially during
winter and summer months (Fig. 2d). When analysing the
complete Q. pubescens BAI time series, enhanced growth
sensitivity to summer P/PET was observed as well as a
positive and highly significant effect of P/PET on BAI
when the 1991–2010 period was analysed separately
(Table 1).
Intrapopulation variability in climate–growth
relationships, WUEBAI and recent BAI trends
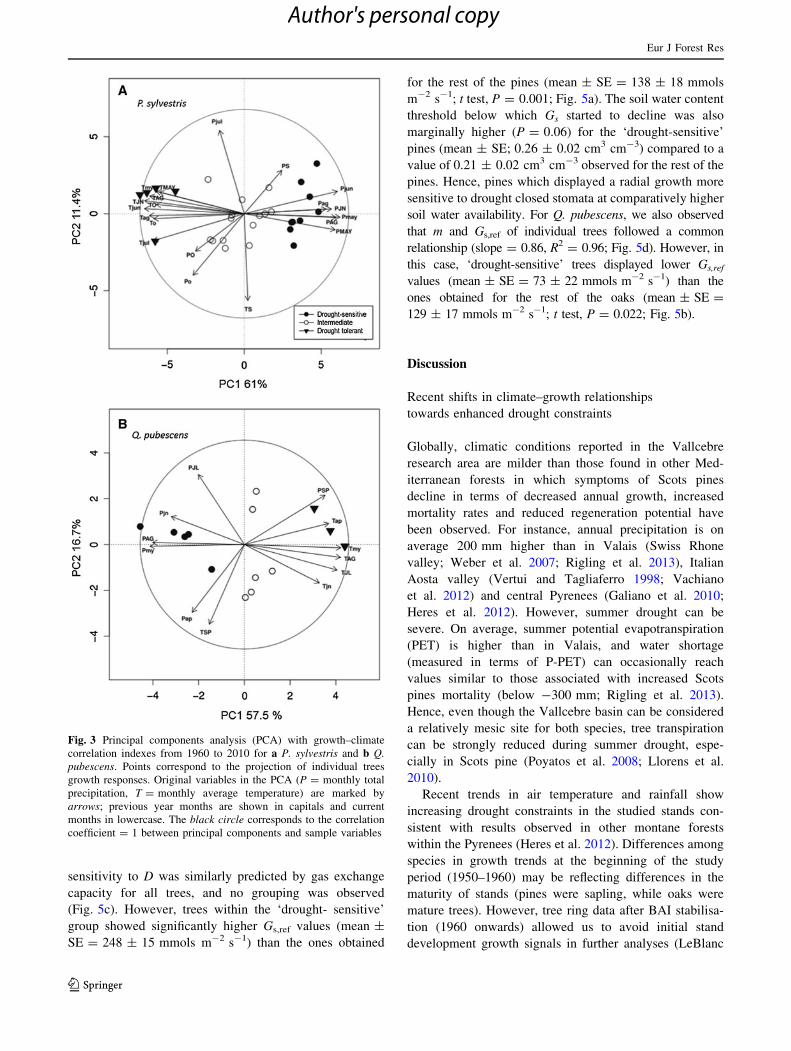
We observed intrapopulation differences in climate-
growth relationships of both species, associated with a
gradient of drought sensitivity (Fig. 3; cf. ‘Meterials and
methods’ section). Trees were grouped as ‘drought sensi-
tive’ (9 pines and 5 oaks), ‘drought tolerant’ (5 pines and 3
oaks) and ‘intermediate’ (14 pines and 6 oaks). PCA
groups were also coherent with k-means analysis (92.86 %
of coherence, TESS = 5.69, pseudo-F = 27.10). These
groups also showed different temporal growth trends
(Fig. 4a, b) but did not significantly differ in their distances
Fig. 2 Pearson correlation coefficient between basal area increment
(mm2/year) and a, b mean monthly temperatures (�C) or c, d monthly
rainfall (mm). Left panels correspond to P. sylvestris and right ones to
Q. pubescens. Black columns correspond to 1960–1990 period and
grey ones to 1991–2010. More than 50 % of trees with significant
correlations (P \ 0.05) are indicated by solid triangles and more than
30 % by solid dots. Previous year months are shown in capitals and
current months in lowercase
Eur J Forest Res
123
Author's personal copy

to the nearest neighbour (P = 0.58 and P = 0.87, in oaks
and pines, respectively).
‘Drought-sensitive’ P. sylvestris, which displayed higher
initial growth rates but current growth decline (Fig. 4a),
also showed significantly lower WUEBAI (Fig. 4c;
mean ± SE = 0.23 ± 0.01 mm2/l H2O, P \ 0.01). The
opposite pattern (no symptoms of growth decline as well as
high WUEBAI) was found in drought-tolerant trees (Fig. 4a,
c; mean ± SE = 1.01 ± 0.12 mm2/l H2O). In accordance,
WUEBAI and initial growth rates had positive and negative
effects, respectively, on P. sylvestris growth sensitivity to
drought (see Table 3), albeit only marginally for initial
growth. In addition, there was a significant shift of ten-
dency in the competitive status of trees, as measured by
their BAIrel, which varied across climate–growth groups
and the time period considered (Table 2; Fig. 4a).
In contrast, Q. pubescens growth trends and WUEBAI
did not show large differences among groups (Fig. 4, right
panels). Besides, neither water use strategies nor initial
growth rates had significant effects on drought tolerance
(Table 3). Nonetheless, some among-groups differences
were observed in growth trends, but only during the 1970s
and 1980s, when drought-sensitive trees showed a stronger
growth response (Table 2; Fig. 4b) to particularly mild
climatic conditions during that period (Fig. 1a). These
differences vanished during the 1990s, and recent growth
now shows a slight decline for all groups (Fig. 4b).
Differences in biomass allocation and size
between climate–growth groups
P. sylvestris growth was significantly related to basal area
(P\ 0.01), and all groups showed similar size–growth patterns
(Fig. S1). Similarly, accumulated transpiration was positively
related to tree leaf area, with analogous responses between
groups. Some but not all ‘drought-sensitive’ pines tended to
show remarkably higher leaf areas (Fig. 4e) although no dif-
ferences in leaf area were found between ‘drought-sensitive’
and ‘drought-tolerant’ pines (P = 0.10). However, we did
observe a significantly lower leaf to sapwood area ratio (AL:AS)
in ‘drought-sensitive’ pines (0.059 ± 0.002 m2 cm-2) com-
pared to ‘drought-tolerant’ pines (0.071 ± 0.003 m2 cm-2,
P \0.01). For Q. pubescens, BAI was highly related to tree
basal area (Fig. S1) and tree transpiration to leaf areas (Fig. 4f),
without any obvious difference among climatic growth sensi-
tivity groups. Nonetheless, ‘drought-sensitive’ oaks showed
the highest basal areas, and ‘drought-tolerant’ oaks showed the
smallest leaf areas (Figs. 4f and S1).
Tree-specific stomatal control related to growth
sensitivity to climate
With regard to P. sylvestris stomatal control, parameter
m of the stomatal response to D followed a common
relationship with respect to Gs,ref, meaning that stomatal
Table 1 Results of the linear mixed models of P. sylvestris and Q. pubescens BAI response to summer water availability, applied to different
time periods
Species Time period of
the analysis
Predictor variables Estimate SE t value P AIC
P. sylvestris 1960–2010 Intercept 6.11 0.08 73.78 <0.001 1467.86
P/PET 0.05 0.04 1.22 0.22
(1991–2010) 0.02 0.08 0.19 0.85
P/PET 3 1991–2010 0.32 0.06 5.68 <0.001
1960–1990 Intercept 6.16 0.09 65.30 <0.001 848.28
P/PET 0.05 0.04 1.21 0.23
1991–2010 Intercept 6.14 0.10 63.67 <0.001 631.31
P/PET 0.37 0.04 8.50 <0.001
Q. pubescens 1960–2010 Intercept 5.80 0.14 40.29 <0.001 429.61
P/PET 0.05 0.04 1.23 0.22
(1991–2010) -0.02 0.10 -0.17 0.87
P/PET 3 1991–2010 0.22 0.06 3.46 <0.001
1960–1990 Intercept 5.88 0.14 41.39 <0.001 319.25
P/PET 0.05 0.05 1.10 0.27
1991–2010 Intercept 5.78 0.17 33.54 <0.001 103.79
P/PET 0.28 0.04 6.50 <0.001
For the analysis covering the period 1960–2010, the response variable was ln(BAI ? 1) and predictor variables were P/PET (mean summer rainfall/
potential evapotranspiration) and time period (1960–1990 or 1991–2010). The period 1960–1990 was used as the reference level. Additional
analyses were also performed where ln(BAI ? 1) was modelled as a function of P/PET for the two separate periods (1960–1990 vs. 1991–2010).
Significant effects (P \ 0.05) are shown in bold. AIC is Akaike information criterion, and SE represents the standard error of the estimate
Eur J Forest Res
123
Author's personal copy
sensitivity to D was similarly predicted by gas exchange
capacity for all trees, and no grouping was observed
(Fig. 5c). However, trees within the ‘drought- sensitive’
group showed significantly higher Gs,ref values (mean ±
SE = 248 ± 15 mmols m-2 s-1) than the ones obtained
for the rest of the pines (mean ± SE = 138 ± 18 mmols
m-2 s-1; t test, P = 0.001; Fig. 5a). The soil water content
threshold below which Gs started to decline was also
marginally higher (P = 0.06) for the ‘drought-sensitive’
pines (mean ± SE; 0.26 ± 0.02 cm3 cm-3) compared to a
value of 0.21 ± 0.02 cm3 cm-3 observed for the rest of the
pines. Hence, pines which displayed a radial growth more
sensitive to drought closed stomata at comparatively higher
soil water availability. For Q. pubescens, we also observed
that m and Gs,ref of individual trees followed a common
relationship (slope = 0.86, R2 = 0.96; Fig. 5d). However, in
this case, ‘drought-sensitive’ trees displayed lower Gs,ref
values (mean ± SE = 73 ± 22 mmols m-2 s-1) than the
ones obtained for the rest of the oaks (mean ± SE =
129 ± 17 mmols m-2 s-1; t test, P = 0.022; Fig. 5b).
Discussion
Recent shifts in climate–growth relationships
towards enhanced drought constraints
Globally, climatic conditions reported in the Vallcebre
research area are milder than those found in other Med-
iterranean forests in which symptoms of Scots pines
decline in terms of decreased annual growth, increased
mortality rates and reduced regeneration potential have
been observed. For instance, annual precipitation is on
average 200 mm higher than in Valais (Swiss Rhone
valley; Weber et al. 2007; Rigling et al. 2013), Italian
Aosta valley (Vertui and Tagliaferro 1998; Vachiano
et al. 2012) and central Pyrenees (Galiano et al. 2010;
Heres et al. 2012). However, summer drought can be
severe. On average, summer potential evapotranspiration
(PET) is higher than in Valais, and water shortage
(measured in terms of P-PET) can occasionally reach
values similar to those associated with increased Scots
pines mortality (below -300 mm; Rigling et al. 2013).
Hence, even though the Vallcebre basin can be considered
a relatively mesic site for both species, tree transpiration
can be strongly reduced during summer drought, espe-
cially in Scots pine (Poyatos et al. 2008; Llorens et al.
2010).
Recent trends in air temperature and rainfall show
increasing drought constraints in the studied stands con-
sistent with results observed in other montane forests
within the Pyrenees (Heres et al. 2012). Differences among
species in growth trends at the beginning of the study
period (1950–1960) may be reflecting differences in the
maturity of stands (pines were sapling, while oaks were
mature trees). However, tree ring data after BAI stabilisa-
tion (1960 onwards) allowed us to avoid initial stand
development growth signals in further analyses (LeBlanc
Fig. 3 Principal components analysis (PCA) with growth–climate
correlation indexes from 1960 to 2010 for a P. sylvestris and b Q.
pubescens. Points correspond to the projection of individual trees
growth responses. Original variables in the PCA (P = monthly total
precipitation, T = monthly average temperature) are marked by
arrows; previous year months are shown in capitals and current
months in lowercase. The black circle corresponds to the correlation
coefficient = 1 between principal components and sample variables
Eur J Forest Res
123
Author's personal copy

1990). Since the 1970s, P. sylvestris and Q. pubescens have
displayed, on average, similar growth patterns. In accor-
dance with climatic trends (Fig. 1a, b), climate–growth
relationships showed a change in tendency towards
enhanced summer drought constraints in recent years for
both species (Fig. 2; Table 1).
Fig. 4 Basal area increment (BAI; mm2/year) in P. sylvestris a and
Q. pubescens b trees in the three different climate–growth groups.
Error bars depict SEs of the mean. Relationship between accumu-
lated transpiration during the months of June to August (l H2O/tree)
and annual growth (BAI) for P. sylvestris (c) and Q. pubescens (d).
The slope of the linear regression (continuous line) between both
variables is water use efficiency expressed in terms of BAI (WUEBAI,
annual accumulated per annual growth). Relationship between leaf
area (m2) and June–August transpiration for P. sylvestris (e) and Q.
pubescens (f). In c–f, each point is an individual tree over 1 year and
represents the accumulated transpiration from June to August
(2003–2005). Note that the scale of the y axis in c and d is different
Eur J Forest Res
123
Author's personal copy

Pinus sylvestris growth was mainly related to winter
temperatures over the 1960–1990 period while linked to
milder summer droughts over the last two decades (Fig. 2a,
c; Table 1). This change in tendency is in accordance with
previous studies showing enhanced climatic constraints on
P. sylvestris growth over the last decades among different
Iberian populations (Martinez-Vilalta et al. 2008) and
consistent with our hypothesis H1. We also observed a shift
in climate–growth relationships for Q. pubescens, from
winter–spring temperatures constraints to summer water
availability limitations (Fig. 2b, d; Table 1). Such shifts
towards increased growth sensitivity to water availability
were quite unexpected here, given the comparatively lower
drought constraints on growth displayed by Q. pubescens
elsewhere (Weber et al. 2007).
Additionally, recent changes in forest structure may have
enhanced competition for belowground resources, espe-
cially water, in these Mediterranean mountain forests. The
studied P. sylvestris stands are located in old agricultural
terraces abandoned in the early 1950s and thus subjected to
forest densification processes (Poyatos et al. 2003). Stand
density in these areas (cf. ‘Materials and methods’ section)
is much higher than the average density of Scots pine stands
in Catalonia (629.18 trees ha-1; IEFC, Gracia et al. 2000–
2004; http://www.creaf.uab.es/iefc/). High stand basal areas
and densities reduce growth potential in P. sylvestris and
may aggravate climatic constraints (Vila-Cabrera et al.
2011; see Martinez-Vilalta et al. 2012 and references
therein). In fact, stand density has been pointed out as a
predisposing factor for drought-induced crown damage and
mortality in unmanaged Scots pines stands or plantations
(Galiano et al. 2010; Sanchez-Salguero et al. 2012). Similar
processes after the abandonment of management may have
occurred in the Q. pubescens stand, as it presently shows a
dense understory and a relatively high stand LAI (Muzylo
et al. 2012). Hence, the enhanced vulnerability to summer
drought observed for both species in this study is probably
the result of the combined effect of increased climatic
aridity and enhanced competition for water. Thinning
would reduce competition intensity and enhance growth,
but increasing drought conditions would probably continue
to constrain the maximum sustainable basal area, as
recently reported for a xeric P. sylvestris stand (Giuggiola
et al. 2013).
Table 2 Results of the linear mixed models relating tree relative BAI (BAIrel), as a measure of competitive status of individual trees, to climate–
growth group (drought tolerant, drought sensitive or intermediate) and time period (1960–1990 and 1991–2010), for P. sylvestris and Q.
pubescens
Species Predictor variables df F value P AIC
P. sylvestris Intercept 1373 734.15 <0.001 112.08
Group 26 0.65 0.531
Period 1373 16.57 <0.001
Group 3 period 1373 73.68 <0.001
Q. pubescens Intercept 663 205.08 <0.001 -137.54
Group 12 0.61 0.560
Period 663 10.87 <0.001
Group 3 period 663 30.62 <0.001
Significant effects (P \ 0.05) are shown in bold. AIC is Akaike information criterion, and SE represents the standard error of the estimate
Table 3 Results of the linear model of climate–growth characteristics as a function of water use efficiency for growth (WUEBAI) and ln-
transformed initial growth rates (ln(BAI60–90 ? 1))
Species Predictor variables Estimate SE t value P AIC
P. sylvestris Intercept 12.36 8.048 1.536 0.15 71.21
WUEBAI 6.423 1.742 3.688 <0.005
ln(BAI60–90 1 1) -2.562 1.206 -2.125 0.05
Q. pubescens (Intercept) -0.110 0.743 -0.269 0.793 64.50
WUEBAI 0.007 0.004 1.481 0.167
The response variable is the individual tree loading on the first principal component (PC1), resulting from the PCA of climate–growth
relationships (Fig. 3). We used values of -PC1 for P. sylvestris and PC1 for Q. pubescens, so that higher, positive values represent increasing
growth sensitivity to drought. Significant effects (P \ 0.05) are shown in bold. AIC is Akaike information criterion and SE represents the
standard error of the estimate
Eur J Forest Res
123
Author's personal copy

Intrapopulation variability in climate–growth
relationships
For both species, we observed intrapopulation clustering
of climate–growth relationships, supporting our hypothe-
sis H2a. Our results showed that ‘drought-sensitive’ P.
sylvestris, which displayed the highest initial growth rates,
have suffered a loss of growth potential over the last
decades (Fig. 4a). In contrast, ‘drought-tolerant’ pines,
which showed a more conservative initial growth,
increased their growth rates after the 1980s and currently
maintain the highest BAI (Fig. 4a). These results point to
a significant shift in the competitive status of individual
trees associated with drought sensitivity (Table 2). The
opposite pattern has been found in a recent study on the
effects of extreme drought events on P. sylvestris growth.
Fast-growing trees resulted more affected by drought in
terms of immediate and delayed growth but continued
having the highest basal area increments after drought.
Hence, intrapopulation competitive relationships were not
affected (Martinez-Vilalta et al. 2012). However, that
study focused on the effects of a single extreme drought
event in the 1980s while here we consider overall growth
trends over the last two decades. The present growth
decline of initially fast-growing trees (Fig. 4a) may be
reflecting a response to gradually increasing dryness
(Fig. 1a, b) rather than the effects of a single and acute
drought episode.
With regard to Q. pubescens, the PCA also splits up
oaks in three different groups with different drought tol-
erances (Fig. 3b). Although all climate–growth archetypes
are currently showing similar growth patterns, ‘drought-
sensitive’ oaks were favoured under enhanced water
availability during the 1970s–1980s, while under recent
drier conditions, all climate–growth groups showed incip-
ient symptoms of growth decline (Fig. 4b). Unlike for P.
sylvestris, climatic constraints are not as important drivers
of shifts in intraspecific competitive relationships of Q.
pubescens at the studied site since all groups currently
show similar growth trends.
Fig. 5 a Reference canopy stomatal conductance (Gs,ref) of drought-
sensitive pines (n = 3) with respect to the rest of the pines (drought
tolerant and intermediate, n = 10), b Gs,ref of drought-sensitive
pubescent oaks (n = 5) and the rest of the oaks (n = 7). Relationship
between Gs,ref and sensitivity to vapour pressure deficit (m) of P.
sylvestris (c) and Q. pubescens (d). **P \ 0.01, *P \ 0.05
Eur J Forest Res
123
Author's personal copy

Intrapopulation variability in climate–growth
relationships associated to water use regulation
Interestingly, for both species, this intrapopulation vari-
ability in climate–growth relationships seemed to be linked
to different water use strategies. ‘Drought-sensitive’ pines
(Fig. 3a) showed higher gas exchange potential (i.e. higher
Gs,ref) and lower WUEBAI compared to ‘drought-tolerant’
pines, which displayed lower Gs,ref and higher WUEBAI
(Figs. 4c,5a). No differences were found in the relationship
between m and Gs,ref among pines (Fig. 5c) suggesting that
‘drought-sensitive’ P. sylvestris did not differ in their rel-
ative stomatal regulation with respect to D (Poyatos et al.
2007). However, in absolute terms, they were indeed more
sensitive to D due to their higher values of m and Gs,ref.
‘Drought-sensitive’ trees also closed stomata at higher soil
water content values. As we only measured average plot
soil water content (i.e. not associated to individual trees),
this behaviour could result from either within-site spatial
variability in soil water availability for trees or differences
in rooting patterns between ‘drought-sensitive’ and
‘drought-tolerant’ pines. Enhanced physiological sensitiv-
ity to drought was related to reduced biomass allocation to
transpiring vs. conducting tissue, as ‘drought-sensitive’
pines showed consistently lower AL:AS compared to
‘drought-tolerant’ pines. These results are consistent with
patterns across climatic gradients (Poyatos et al. 2007) and
within-population differences between declining and non-
declining P. sylvestris (Poyatos et al. 2013).
The opposite pattern was observed for Q. pubescens,
whereby ‘drought-sensitive’ individuals showed reduced
Gs,ref and m values (Fig. 5b, d). In addition, sap flow is
relatively insensitive to superficial soil moisture in Q. pu-
bescens (Poyatos et al. 2008), so we could not explore
differences in soil water content thresholds for stomatal
closure. Both results agree with the ability of this species to
tap deep water sources (Valentini et al. 1992) and suggest
that ‘drought-sensitive’ trees may display comparatively
less developed root systems, thus explaining structural
constraints on gas exchange potential (i.e. reduced Gs,ref)
(Sperry et al. 1998). Overall, these findings show that
stomatal regulation of transpiration and climatic controls
on radial growth is tightly linked (hypothesis H2b), but in
different ways for P. sylvestris and Q. pubescens.
Other factors such as size (Zang et al. 2012) or crown
class membership (Martin-Benito et al. 2008) could also
underlie the observed intrapopulation variability in cli-
mate–growth relationships (but see Merian and Lebour-
geois (2011) for a counterexample of little size-related
variation in climate–growth relationships in P. sylvestris
and Quercus petraea). Our sampling strategy would have
minimised these size and social position effects as all trees
were codominant, showed similar heights and a small range
of dbh values. Indeed, our results show that climate–growth
groups were not associated with competition for below-
ground resources.
Implications for drought-induced decline processes
Even though no apparent symptoms of Scots pine drought-
induced decline have been observed in the study area, the
possibility that ‘drought-sensitive’ trees may have entered
a decline process cannot be discarded, given the gradual
decrease observed in BAI for ‘drought-sensitive’ and
‘intermediate’ trees (Fig. 4a). A long, gradual growth
decline has been shown to precede tree death in drought-
exposed Scots pine populations and has also been linked to
an enhanced climatic sensitivity of radial growth (Bigler
et al. 2006; Heres et al. 2012), as we have reported here.
Our results on intrapopulation variability in drought sen-
sitivity are consistent with coupling between climate,
growth and WUE (Voltas et al. 2013) and/or a limited
increase in WUE with increasing atmospheric CO2 con-
centration in P. sylvestris (Heres et al. 2013) as symptoms
of incipient drought-induced decline. Given the high stand
density in the study area, coupled with the current trend of
increasing aridity (Fig. 1), the studied population may
become vulnerable to drought-induced dieback in the fol-
lowing decades, as it has already been observed in other
unmanaged Scots pine populations in the Pyrenees (Gali-
ano et al. 2011). Nevertheless, ‘drought-tolerant’ individ-
uals increased their growth rates after 2005, a dry year
(Poyatos et al. 2008) which triggered forest decline epi-
sodes elsewhere in the Pyrenees (Heres et al. 2012). This
recovery occurred in response to a recent transient increase
in annual rainfall (Fig. 1a) and demonstrates that resilient
individuals can recover after drought.
In comparison, the evidence in scientific literature for
drought-induced decline in Q. pubescens is scarce (but see
Carnicer et al. 2011, Supp. Information, pp. 12–13). Despite
that in our study site Q. pubescens leaf water potentials
(Poyatos et al. 2008) are far from those values known to
induce leaf desiccation and senescence (Damesin et al.
1998), its moderate stomatal control in response to high
evaporative demand (Zweifel et al. 2007; Poyatos et al.
2008) can cause shoot dieback under extremely dry spells
(Zweifel et al. 2006; Nardini et al. 2013). According to a
recent monitoring programme of forest decline recently
started in Catalonia (NE Spain), 4 % of the total area
occupied by Q. pubescens was affected by crown dieback in
2011, including populations in the Pyrenees (Banque et al.
2011; P. Llorens, personal observation). These results are
consistent with the increasing drought constraints on growth
observed for Q. pubescens since the 1990s (Fig. 4b).
Other deciduous oaks in Europe have indeed shown
vulnerability to drought-induced decline (e.g. Breda et al.
Eur J Forest Res
123
Author's personal copy

2006). For one of these species, Quercus robur, BAI
trends, wood anatomical traits and gas exchange patterns
revealed that fast-growing trees showed lower WUE and
wider vessels, which resulted in increased predisposition to
drought-related mortality (Levanic et al. 2011). Likewise,
the combined monitoring of water use and growth strate-
gies in potentially vulnerable populations of Q. pubescens
would shed some light on the physiology behind climatic
controls on growth under severe drought, and how these
mechanisms vary within a population.
Conclusions
Our results reveal increasing growth constraints over the last
decades for P. sylvestris and for Q. pubescens. These patterns
may be the result of a concomitant increase in aridity and the
lack of forest management. However, drought responses of
growth and physiological traits were not uniform within
species. Drought sensitivity was closely related to tree-spe-
cific past growth trends and water use regulation in P. syl-
vestris. In contrast, within-population differences in drought
sensitivity and water use traits did not result in different
growth trends in recent decades among Q. pubescens indi-
viduals. Here, we also show that the initially slowest-grow-
ing P. sylvestris eventually became more resilient and
recovered faster after recent drought-induced declines in
growth, leading to a shift in intrapopulation competitive
relationships. Although increasing drought constraints may
be contributing to the substitution of P. sylvestris by Q. pu-
bescens in dry sites (Galiano et al. 2010; Rigling et al. 2013),
we observed an incipient growth decline in Q. pubescens,
which may be punctuated by the occurrence of extreme
drought episodes. Overall, this study sheds some light on the
key physiological and growth regulation mechanisms, which
may be involved in the onset of gradual tree decline. How-
ever, future studies including larger spatial scales and
incorporating aridity and stand density gradients may be
needed in order to make species-level generalisations.
Acknowledgments This study was supported by MONTES
(CSD2008-0040) funded by Spanish Government. TM-L was bene-
ficiary of a ‘Fundacion la Caixa’ grant, and RP was supported by a
Juan de la Cierva postdoctoral fellowship. We acknowledge help of
Garcıa-Estrıngana P. and Latron J. in field data acquisition, Riera J.L.
in data analysis Gutierrez E. in dendroecological study, Delgado J. for
providing us long-term climatic databases and Valladares F. for
carefully reading this manuscript.
References
Andreu L, Gutierrez E, Macias M, Ribas M, Bosch O, Camarero JJ
(2007) Climate increases regional tree-growth variability in
Iberian pine forests. Glob Change Biol 13:804–815
Banque M, Vayreda J, Martınez-Vilalta J (2011) Decaıment dels
boscos de Catalunya (DeBosCat). Informe Annual. Barcelona,
Spain
Bigler C, Braker OU, Bugmann H, Dobbertin M, Rigling A (2006)
Drought as an inciting mortality factor in Scots pine stands of the
Valais, Switzerland. Ecosystems 9:330–343
Breda N, Granier A (1996) Intra- and interannual variations of
transpiration, leaf area index and radial growth of a sessile oak
stand (Quercus petraea). Ann Sci For 53:521–536
Breda N, Huc R, Granier A, Dreyer E (2006) Temperate forest trees
and stands under severe drought: a review of ecophysiological
responses, adaptation processes and long-term consequences.
Ann For Sci 63:625–644
Cade BS, Noon BR (2003) A gentle introduction to quantile
regression for ecologists. Front Ecol Environ 1:412–420
Carnicer J, Coll M, Ninyerola M, Pons X, Sanchez G, Penuelas J
(2011) Widespread crown condition decline, food web disruption
and amplified tree mortality with increased climate change-type
drought. PNAS 108:1474–1478
Christensen JH, Christensen OB (2007) A summary of the PRU-
DENCE model projections of changes in European climate by
the end of this century. Clim Change 81:7–30
Damesin C, Rambal S, Joffre R (1998) Co-occurrence of trees with
different leaf habit: a functional approach on Mediterranean
oaks. Acta Oecol 19:195–204
De Caceres M, Oliva F, Font X, Vives S (2007) Gingko, a program
for non-standard multivariate fuzzy analysis. Fuzze Set Syst
2:41–56
Eilmann B, Zweifel R, Buchmann N, Pannatier EG, Rigling A (2011)
Drought alters timing quantity and quality of wood formation in
Scots pine. J Exp Bot 62:2763–2771
Ewers BE, Oren R (2000) Analyses of assumptions and errors in the
calculation of stomatal conductance from sap flux measure-
ments. Tree Physiol 20:579–589
Galiano L, Martınez-Vilalta J, Lloret F (2010) Drought-induced
multifactor decline of scots pine in the Pyrenees and potential
vegetation change by the expansion of co-occurring oak species.
Ecosystems 13:978–991
Galiano L, Martinez-Vilalta J, Lloret F (2011) Carbon reserves and
canopy defoliation determine the recovery of Scots pine 4 yr
after a drought episode. New Phytol 190:750–759
Gimmi U, Wohlgemuth T, Rigling A, Hoffmann CW, Burgi M (2010)
Land-use and climate change effects in forest compositional
trajectories in a dry Central-Alpine valley. Ann For Sci
67:701–710
Giuggiola A, Bugmann H, Zingg A, Dobbertin M, Rigling A (2013)
Reduction of stand density increases drought resistance in xeric
Scots pine forests. Forest Ecol Manag 310:827–835
Gracia C, Burriel JA, Ibanez JJ, Mata T, Vayreda J (2000–2004)
Inventari Ecologic i Forestal de Catalunya, 10 volumes. CREAF,
Bellaterra, Spain
Granier A (1985) Une nouvelle methode pur la mesure du flux de seve
brute dans le tronc des arbres. Ann For Sci 42:193–200
Granier A, Loustau D, Breda N (2000) A generic model of forest
canopy conductance dependent on climate, soil water availability
and leaf area index. Ann For Sci 57:755–765
Heres AM, Martinez-Vilalta J, Lopez BC (2012) Growth patterns in
relation to drought-induced mortality at two Scots pine (Pinus
sylvestris L.) sites in NE Iberian Peninsula. Trees Struct Funct
26:621–630
Heres AM, Voltas J, Lopez BC, Martınez-Vilalta J (2013) Drought-
induced mortality selectively affects Scots pine trees that show
limited intrinsic water-use efficiency responsiveness to raising
atmospheric CO2. Func Plant Biol. doi:10.1071/FP13067
Holmes RL (1983) Computer-assisted quality control in tree-ring
dating and measurement. Tree Ring Bull 43:68–78
Eur J Forest Res
123
Author's personal copy

IPCC, Climate Change (2007) The physical science basis: working
group I contribution to the fourth assessment report of the IPCC.
Cambridge University Press, Cambridge
Koenker R, the R Development Core Team (2008) Quantreg: quantile
regression and related methods. R package version 4.81
Latron J, Soler M, Llorens P, Gallart F (2008) Spatial and temporal
variability of the hydrological response in a small Mediterranean
research catchment (Vallcebre, Eastern Pyrenees). Hydrol Pro-
cess 22:775–787
LeBlanc DC (1990) Relationships between breast-height and whole-
stem growth indices for red spruce on Whiteface Mountain, New
York. Can J For Res 20:1399–1407
Levanic T, Cater M, McDowell NG (2011) Associations between
growth, wood anatomy, carbon isotope discrimination and
mortality in a Quercus robur forest. Tree Physiol 31:298–308
Llorens P, Gallart F (1992) Small basin response in a Mediterranean
mountainous abandoned farming area- research design and
preliminary-results. Catena 19:309–320
Llorens P, Poch R, Latron J, Gallart F (1997) Rainfall interception by
a Pinus sylvestris forest patch overgrown in a Mediterranean
mountainous abandoned area.1. Monitoring design and results
down to the event scale. J Hydrol 199:331–345
Llorens P, Poyatos R, Latron J, Delgado J, Oliveras I, Gallart F (2010)
A multi-year study of rainfall and soil water controls on Scots
pine transpiration under Mediterranean mountain conditions.
Hydrol Process 24:3053–3064
Lopez-Moreno JI, Nogues-Bravo D (2005) A generalized additive
model for the spatial distribution of snowpack in the Spanish
Pyrenees. Hydrol Process 19:3167–3176
Martin-Benito D, Cherubini P, del Rio M, Canellas I (2008) Growth
response to climate and drought in Pinus nigra Arn. trees of
different crown classes. Trees Struct Funct 22:363–373
Martinez-Vilalta J, Lopez BC, Adell N, Badiella L, Ninyerola M
(2008) Twentieth century increase of Scots pine radial growth in
NE Spain shows strong climate interactions. Glob Change Biol
14:2868–2881
Martinez-Vilalta J, Lopez BC, Loepfe L, Lloret F (2012) Stand- and
tree-level determinants of the drought response of Scots pine
radial growth. Oecologia 168:877–888
Merian P, Lebourgeois F (2011) Size-mediated climate-growth
relationships in temperate forests: a multi-species analysis. For
Ecol Manag 261:1382–1391
Muggeo V (2008) Segmented: an R package to fit regression models
with broken-line relationships. Rnews 8:20–25
Mu _zyło A, Llorens P, Domingo F (2012) Rainfall partitioning in a
deciduous forest plot in leafed and leafless periods. Ecohydrol-
ogy 5:759–767
Nardini A, Battistuzzo M, Savi T (2013) Shoot desiccation and
hydraulic failure in temperate woody angiosperms during an
extreme summer drought. New Phytol 200:322–329
Oren R, Sperry JS, Katul GG, Pataki DE, Ewers BE, Phillips N,
Schafer KVR (1999) Survey and synthesis of intra- and
interspecific variation in stomatal sensitivity to vapour pressure
deficit. Plant Cell Environ 22:1515–1526
Pinheiro J, Bates D, DebRoy S, Sarkar D, the R Development Core
Team (2012) nlme: linear and nonlinear mixed effects models. R
package version 3.1-106
Poyatos R, Latron J, Llorens P (2003) Land use and land cover change
after agricultural abandonment—the case of a Mediterranean
Mountain Area (Catalan Pre-Pyrenees). Mt Res Dev 23:362–368
Poyatos R, Llorens P, Gallart F (2005) Transpiration of montane
Pinus sylvestris L. and Quercus pubescens Willd. forest stands
measured with sap flow sensors in NE Spain. Hydrol Earth Syst
Sci 9:493–505
Poyatos R, Martinez-Vilalta J, Cermak J, Ceulemans R, Granier A,
Irvine J, Kostner B, Lagergren F, Meiresonne L, Nadezhdina N,
Zimmermann R, Llorens P, Mencuccini M (2007) Plasticity in
hydraulic architecture of Scots pine across Eurasia. Oecologia
153:245–259
Poyatos R, Llorens P, Pinol J, Rubio C (2008) Response of Scots pine
(Pinus sylvestris L.) and pubescent oak (Quercus pubescens
Willd.) to soil and atmospheric water deficits under Mediterra-
nean mountain climate. Ann For Sci 65:306–319
Poyatos R, Aguade D, Galiano L, Mencuccini M, Martınez-Vilalta J
(2013) Drought-induced defoliation and long periods of near-
zero gas exchange play a key role in accentuating metabolic
decline of Scots pine. New Phytol 200:388–401
R Core Team (2011) R: A language and environment for statistical
computing. R Foundation for Statistical Computing, Vienna,
Austria. http://www.R-project.org/
Rigling A, Bigler C, Eilmann B, Feldmeyer-Christe E, Gimmi U,
Ginzler C, Graf U, Mayer P, Vacchiano G, Weber P, Wohlge-
muth T, Zweifel R, Dobbertin M (2013) Driving factors of a
vegetation shift from Scots pine to pubescent oak in dry Alpine
forests. Glob Change Biol 19:229–240
Rubio EC (2005) Hidrodinamica de los suelos de un area de montana
media mediterranea sometida a cambios de uso y cubierta, PhD
thesis, Universitat Autonoma de Barcelona, Barcelona, Spain
Sanchez-Salguero R, Navarro-Cerrillo RM, Swetman TW, Zavala
MA (2012) Is drought the main decline factor at the rear edge of
Europe? The case of southern Iberian pine plantations. Forest
Ecol Manag 271:158–169
Sperry JS, Adler FR, Campbell GS, Comstock JP (1998) Limitation
of plant water use by rhizosphere and xylem conductance: results
from a model. Plant Cell Environ 21:347–359
UNEP (1992) World atlas of desertification. Edward Arnold, London,
UK
Vachiano G, Garbarino M, Mondino EB, Motta R (2012) Evidences
of drought stress as predisposing factor to Scots pine decline in
Valle d’Aosta (Italy). Eur J For Res 132:989–1000
Valentini R, Mugnozza GES, Ehleringer JR (1992) Hydrogen and
carbon isotope ratios of selected species of a mediterranean
macchia ecosystem. Funct Ecol 6:627–631
Vertui F, Tagliaferro F (1998) Scots pine (Pinus sylvestris L.) die-
back by unknown causes in the Aosta Valley, Italy. Chemo-
sphere 36:1061–1065
Vila-Cabrera A, Martinez-Vilalta J, Vayreda J, Retana J (2011) Structural
and climatic determinants of demographic rates of Scots pine forests
across the Iberian Peninsula. Ecol Appl 21:1162–1172
Vila-Cabrera A, Rodrigo A, Martinez-Vilalta J, Retana J (2012) Lack
of regeneration and climatic vulnerability to fire of Scots pine
may induce vegetation shifts at the southern edge of its
distribution. J Biogeogr 39:488–496
Voltas J, Camarero JJ, Carulla D, Aguilera M, Ortiz A, Ferrio JP (2013) A
retrospective, dual-isotope approach reveals individual predisposi-
tions to winter-drought induced tree dieback in the southernmost
distribution limit of Scots pine. Plant Cell Environ 36:1435–1448
Weber P, Bugmann H, Rigling A (2007) Radial growth responses to
drought of Pinus sylvestris and Quercus pubescens in an inner-
Alpine dry valley. J Veg Sci 18:777–792
Whitehead D, Jarvis PG (1981) Coniferous forests and plantations. In:
Kozlowski TT (ed) Water deficits and plant growth. Academic
Press, New York, pp 49–152
Zang C, Pretzsch H, Rothe A (2012) Size-dependent responses to
summer drought in Scots pine, Norway spruce and common oak.
Trees Struct Funct 26:557–569
Zweifel R, Zimmermann L, Zeugin F, Newbery DM (2006) Intra-
annual radial growth and water relations of trees: implications
towards a growth mechanism. J Exp Bot 57:1445–1459
Zweifel R, Steppe K, Sterck FJ (2007) Stomatal regulation by
microclimate and tree water relations: interpreting ecophysiological
field data with a hydraulic plant model. J Exp Bot 56:2113–2131
Eur J Forest Res
123
Author's personal copy




















