
Protective effect of N-acetylcysteine against carmustine-induced myelotoxicity in rats
Upload
independentCategory
view
1download
0
FromdaScofin
SupptoPoPoSunaCCda
AuthPresBo
RepPa20
Thetom
0741Cophttp
Effects of N-acetylcysteine on skeletal musclestructure and function in a mouse model ofperipheral arterial insufficiencyBruno T. Roseguini, PhD,a Leonardo M. Silva, BS,a Tatiana G. Polotow, MS,b Marcelo P. Barros, PhD,b
Caden Souccar, PhD,c and Sang W. Han, PhD,a São Paulo, Brazil
Objective: Abnormalities in skeletal muscle structure and function are important contributors to exercise intolerance andfunctional decline in peripheral arterial disease. In this study, we tested the hypothesis that administration of N-ace-tylcysteine (NAC) would improve fatigue resistance and ameliorate the histopathological changes in skeletal muscle in amouse model of peripheral arterial disease. We also anticipated that NAC treatment would lower the levels of biomarkersof oxidative damage in the ischemic muscle.Methods: Male Balb/c mice were subjected to bilateral ligation of the femoral artery and, after 2 weeks of recovery,received daily intraperitoneal injections of either NAC (150 mg/kg) or saline for 15 days. At the end of the treatment, theextensor digitorium longus (EDL) and soleus muscles were excised for assessment of contractile function in vitro andhistological analysis. Free malondialdehyde and protein carbonyl levels were measured in the gastrocnemius muscle.Results: In the soleus muscle, force after 10 minutes of submaximal tetanic stimulation (60 Hz, 300 ms trains, 0.3trains/s) was higher (P < .05) in NAC-treated animals (45% 6 3% of the initial value; n [ 7) when compared withcontrols (30.3% 6 3%; n [ 8). No differences were found in fatigue development between groups in the EDL muscle(ligated NAC, 35.7% 6 1.9%; ligated saline, 37.5% 6 1.1%). In addition, there was a tendency for lower levels of con-nective tissue deposition in the soleus of animals treated with NAC (n [ 6) when compared with those that received onlysaline (n [ 9) (ligated NAC, 16% 6 2% vs ligated saline, 24% 6 2%; P [ .057). No differences were found in lipidperoxidation or protein carbonyl levels between ligated saline and ligated NAC groups.Conclusions: Taken together, these results indicate that treatment with NAC improves fatigue resistance in the soleus butnot the EDL muscle in a model of peripheral arterial insufficiency. (J Vasc Surg 2013;-:1-10.)
Clinical Relevance: Despite the increasing burden of peripheral arterial disease (PAD) and its detrimental consequenceson the quality of life of the patients, few pharmacological therapies have shown to evoke meaningful effects on functionalperformance in these individuals. N-acetylcysteine is approved for clinical use, has minimal side effects and mostimportant, has shown to consistently improve exercise performance in animals and humans. In this study, we showed, forthe first time, that treatment with this drug at a dose amenable for clinical application evoked marked effects on fatigueresistance in the soleus muscle in a mouse model of PAD. These encouraging findings set the stage for translational studiesto determine the acute and long-term impact of this drug on walking capacity in patients with PAD.
the Department of Biophysics, Escola Paulista de Medicina, Universi-de Federal de São Pauloa; the Institute of Physical Activity and Sportsiences (ICAFE), Universidade Cruzeiro do Sulb; and the DepartmentPharmacology, Section of Natural Products, Escola Paulista de Medic-a, Universidade Federal de São Paulo.c
orted by grants from the Sao Paulo Research Foundation e FAPESPDr Souccar (2010/51344-3) and Dr Han (2011/00859-6) and astdoctoral Fellowship to Dr Roseguini (Fapesp 2011/18197-0). Drlotow was supported by a M.Sc. fellowship by the Programa deporte à Pós-graduação de Instituições de Ensino Particulares, Coorde-ção de Aperfeiçoamento de Pessoal de Nível Superior (PROSUP/APES, Brazil). Dr Barros is an established investigator of the Nationalouncil of Technological and Scientific Development (Bolsa Produtivi-de em Pesquisa, Nivel 2, 307474/2012-7, CNPq, Brazil).or conflict of interest: none.ented in abstract form at the 2013 Experimental Biology Meeting inston, Mass, April 20-24, 2013.
rint requests: Sang W. Han, PhD, Department of Biophysics, Escolaulista de Medicina, Universidade Federal de São Paulo, Rua Mirassol7, São Paulo, SP, Brazil 04044-010 (e-mail: [email protected]).editors and reviewers of this article have no relevant financial relationshipsdisclose per the JVS policy that requires reviewers to decline review of anyanuscript for which they may have a conflict of interest.-5214/$36.00yright � 2013 by the Society for Vascular Surgery.://dx.doi.org/10.1016/j.jvs.2013.10.098
Peripheral arterial disease (PAD) is a common manifes-tation of atherosclerosis that affects a growing proportionof the elderly population.1,2 Patients with this conditionhave severe exercise intolerance, faster rates of functionaldecline, and poorer quality of life than their healthy coun-terparts.3-5 The genesis of exercise intolerance in PAD ismultifactorial,6 but a growing number of studies indicatethat alterations in skeletal muscle structure and functionplay a decisive role.7,8 It is well known that patients withPAD have lower extremity muscle weakness and reducedmuscle endurance.8 Histological and biochemical studieshave also documented a number of abnormalities in thecalf muscles from these individuals, including the presenceof degenerating and regenerating fibers, connective tissueproliferation, fat accumulation, mitochondrial dysfunction,and a marked increase in biomarkers of oxidative dam-age.9-11 Importantly, these adverse calf muscle characteris-tics are predictors of functional decline and all-causemortality in this population.12,13
Despite the growing acceptance that these myopathicchanges contribute to the pathophysiology of PAD,12
few therapeutic strategies to date have been shown to
1

JOURNAL OF VASCULAR SURGERY2 Roseguini et al --- 2013
effectively minimize or reverse these abnormalities. Forinstance, exercise training, widely accepted as the best sin-gle treatment option for this condition, has been shown toactually worsen ischemic damage in the calf muscle of pa-tients with PAD, as revealed by an increase in the numberof denervated fibers and the amount of connective tissue.14
The impact of pharmacological agents that have success-fully shown to improve walking tolerance in PAD, suchas the phosphodiesterase inhibitor cilostazol,15 on skeletalmuscle is largely unknown. A growing number of studiesindicate that antioxidant supplementation could be a usefulstrategy to prevent exercise-induced oxidative damage andinflammation in ischemic skeletal muscle,16-18 but thelong-term consequences of these agents on skeletal musclemorphology and fatigue resistance in PAD remain unex-plored. Therefore, there is an urgent need for developmentof novel strategies that ameliorate these skeletal muscleabnormalities and restore functional capacity in thesepatients.
The present study was designed to evaluate the impactof the compound N-acetylcysteine (NAC) on the structuraland functional phenotype of different muscles in a mousemodel of peripheral arterial insufficiency. We chose NACbecause of the following reasons: first, NAC is a non-specific antioxidant,19 a vasodilator,20 and has been consis-tently associated with improvements in fatigue resistance ina variety of animal preparations as well as in clinical popu-lations21,22; second, in diseases associated with myopathicchanges, such as Duchenne muscular dystrophy, treatmentwith NAC has shown to abrogate adverse remodeling inboth cardiac and skeletal muscle as evidenced by reducedfibrosis and reduced number of fibers with centralnuclei.23,24 Therefore, we tested the hypothesis that2 weeks of daily administration of NAC would improve fa-tigue resistance, ameliorate the histopathological changes,and reduce oxidative damage in the skeletal muscle ofmice subjected to bilateral ligation of the femoral artery.
METHODS
Animals. Experiments were conducted on 14- to 15-week-old male Balb/c mice. This strain was chosenbecause these animals reportedly have fewer native collat-eral arteries,25 poorer collateral growth and blood flowrecovery in response to arterial occlusion,25,26 and, asshown recently, impaired myocyte regenerative capacitywhen compared with other strains.27 Animals were ob-tained from the Animal Facility at the Instituto Nacional deFarmacologia e Biologia Molecular, Escola Paulista deMedicina/Universidade Federal de São Paulo. All animalswere housed under a controlled 12-hour/12-hour light/dark cycle and at 22� 6 2� C with free access to food andwater. All procedures were performed in accordance withthe Guide for the Care and Use of Laboratory Animals ofthe U.S. National Institutes of Health (Bethesda, Md), andwere approved by the Research Ethics Committee of theFederal University of São Paulo, Brazil (Approval number:CEP 0212/12).
Femoral artery ligation. Surgical induction of hin-dlimb ischemia was performed under general anesthesiawith an intraperitoneal injection of ketamine (40 mg/kg)and xylazine (20 mg/kg). The femoral artery was exposed,separated from the vein and nerve, and occluded immedi-ately distal to the epigastric artery with nylon sutures(6-0). The animals were allowed 2 weeks of recoverybefore the experiments. As outlined in the Discussion(see Methodological considerations), the recovery timebefore the onset of treatment was given to allow for the nat-ural compensation in response to the ischemic insult tooccur. In this model, calf blood flow and oxygenation typi-cally attain steady-state levels within 2 weeks followingfemoral artery ligation.26 In order to minimize the effect ofinteranimal variability on outcome, ligated animals thatprogressed to severe ischemia and amputation were ex-cluded. A sham-operated control group was also included inthe study. Animals from this group underwent a similarsurgical procedure except for the ligation of the artery.
Protocol. After 2 weeks of recovery from the surgicalprocedure, the animals were randomized to treatment withNAC (150 mg/kg dissolved in 0.1 mL saline; Sigma-Aldrich, St. Louis, Mo)28 or saline (0.1 mL). The animalsreceived daily intraperitoneal injections for 15 consecutivedays. In the first subset of experiments (n ¼ 21), the ani-mals were euthanized at the end of the treatment for theassessment of skeletal muscle function in vitro. The lastdose of NAC/saline was given 1 hour prior to sacrifice. In asecond subset of experiments (n ¼ 20), the extensor dig-itorium longus (EDL) and soleus muscles were harvestedfor histological determination of fiber size, central nucle-ation, and collagen production. Lastly, in a third subset ofanimals (n ¼ 20), the gastrocnemius muscle of both legswas harvested under anesthesia for biochemical determi-nation of lipid peroxidation and protein carbonyl levels.The number of animals used for each experimentaloutcome was chosen based on prior reports that investi-gated the effects of NAC treatment in a mouse modelDuchenne muscular dystrophy.23,24,28
Contractile function and fatigue of isolated mus-cles. Isometric contractile properties of the slow-twitchsoleus and the fast-twitch EDL muscles were evaluatedin vitro as described previously.29 These two muscles werechosen because: (1) both allow for sutures to be tied in theproximal and distal tendons, and (2) have contrasting fibertype compositions, which allows for the investigation ofwhether this parameter influences treatment outcomes.Briefly, mice were anesthetized with an intraperitoneal in-jection of ketamine (40 mg/kg) and xylazine (20 mg/kg),and the entire hindlimb was removed and kept in Tyrodesolution with the following composition (in mM): 135NaCl; 15 NaHCO3; 5 KCl, 2 CaCl2, 1 MgCl2, 1NaH2PO4, and 11 glucose, pH 7.3 to 7.4 after bubblingwith 95% O2 þ 5% CO2. The soleus and EDL muscleswere then carefully isolated and tied at the proximal anddistal tendons with braided surgical silk. The muscleswere transferred to a temperature-controlled organ bathcontaining 5 mL of Tyrode solution at 25�C bubbled

Table I. Muscle weights and contractile properties of isolated extensor digitorum longus (EDL) and soleus muscles fromsham-operated controls, ligated animals treated intraperitoneally with N-acetylcysteine (NAC; 150 mg/kg), and ligatedanimals treated with saline
EDL Soleus
Sham-operated Ligated saline Ligated NAC Sham-operated Ligated saline Ligated NAC
Muscle weight, mg 15.8 6 0.4a 8.4 6 1 7.9 6 1 9.7 6 0.2b 5.5 6 1.1 7.4 6 0.9Muscle weight/body weight, mg/g 0.51 6 0.01a 0.29 6 0.03 0.27 6 0.03 0.31 6 0.006b 0.19 6 0.03 0.25 6 0.03Pt, mN 54.6 6 1.4a 31.2 6 1.8 31.8 6 2.7 39.1 6 1.1a 31.6 6 0.7 29.5 6 1.1TPT, ms 28.0 6 0.01a 25.5 6 0.5 25.4 6 0.6 33.8 6 0.5 32.7 6 0.4 32.5 6 0.7½ RT, ms 16.6 6 0.2 16.0 6 0.4 16.4 6 0.2 26.8 6 1.0 25.1 6 1.0 27.1 6 1.7P0, mN 429.4 6 14.7a 183.2 6 27.0 179.6 6 27.4 247.4 6 11.2a 110.2 6 21.3 154.3 6 20.6sP0, N/cm2 45.6 6 0.5a 36.3 6 1.5 37.3 6 1.4 40.2 6 1.6b 34.1 6 1.2 35.5 6 1.2
P0, Maximum tetanic force; Pt, maximum twitch force; ½ RT, one-half relaxation time; SP0, specific force; TPT, time to peak twitch.Values are as mean 6 standard error.aP < .05 vs ligated saline and ligated NAC.bP < .05 vs ligated saline.
JOURNAL OF VASCULAR SURGERYVolume -, Number - Roseguini et al 3
continuously with carbogen. The distal tendon was tied toa holder fixed in the organ bath and the proximal tendonwas attached to a force transducer (model FT03; GrassInstruments, West Warwick, RI). The muscles were stim-ulated to contract isometrically using electrical fieldstimulation (S-88 Grass-stimulator; Grass Instruments)delivered via two platinum wire electrodes. The output ofthe force transducer was recorded and analyzed using aPowerLab system (ADInstruments, Colorado Springs,Colo). After being mounted, muscle optimal length (L0)was determined from micromanipulations of muscle lengthand a series of isometric twitch contractions. The temper-ature was then increased to 32�C, and the muscles wereallowed to equilibrate for 10 minutes in the presence of 10-mMD-tubocurarine in order to prevent muscle contractionevoked by electrical nerve stimulation. The resting muscletension was then readjusted, and the following parameterswere measured in response to supramaximal square-wavepulses (0.5 ms): peak twitch tension (Pt), time to peaktension (TPT), and one-half relaxation time (½ RT). Todefine force-frequency characteristics, we measured force inresponse to stimulus frequencies of 15, 30, 50, 80, 120,and 150 every 2 minutes. Pulse and train durations were0.5 and 300 ms for the EDL muscle and 0.5 and 800 msfor the soleus. Relative tension developed (P) was ob-tained, and the force-frequency curves were computedfrom the maximum tetanic tension (P0). The curves werefit with a four-parameter Hill Equation (SigmaPlot 11.0;Systat Software, San Jose, Calif) to obtain the F50, which isthe frequency at 50% of maximal force.
Two minutes after the force-frequency protocol, themuscles underwent a fatigue protocol consisting of seriesof tetanic contractions for a total time of 15 minutes(60 Hz, 0.3 ms pulse duration). As the EDL and soleusmuscles have marked differences in susceptibility to fatigue,distinct protocols were used for each muscle to provokea reduction in force to approximately 40% of theinitial value. Thus, in the EDL, the protocol consisted of12 contractions/min for 10 minutes followed by 18
contractions/min for an additional 5 minutes. In the so-leus, tetanic contractions were evoked at a rate of 18 con-tractions/min for 10 minutes and then at 24 contractions/min for the final 5 minutes. Before unmounting the musclefrom the myograph, muscle length was measured with cal-ipers. The muscles were then trimmed of their tendons andnonmuscle tissue and weighed on an analytic balance.Cross-sectional area (CSA) was determined as describedby Close.30 Absolute forces are expressed as N/cm2.Forces measured during the fatigue protocol were normal-ized for the force obtained in the first contraction of thetrial.
Biochemical assays. Free malondialdehyde (MDA)(Bioxytech MDA-586; Oxis International Inc, FosterCity, Calif), a well-known marker of lipid peroxidation, andprotein carbonyl (Cayman Chemical, Ann Arbor, Mich),a marker of protein oxidation, were measured in thegastrocnemius muscle using commercially available assaykits. This muscle was chosen because it is importantlyaffected by the ischemic insult in this animal model31 andprovided the required amount of starting material toconduct these assays, as detailed in the kit’s manual. Forthe MDA assay, the entire frozen gastrocnemius of one legwas homogenized in lysis buffer (CelLytic MT Cell LysisReagent; Sigma-Aldrich) containing a protease inhibitorcocktail (P8340; Sigma-Aldrich) and centrifuged at10.000 � g for 10 minutes at 4�C. The supernatant wasthen used to perform the assay according to manufacturer’sinstructions. For the protein carbonyl assay, the gastroc-nemius muscle was homogenized in 50 mM MES buffer,centrifuged at 10.000 � g for 10 minutes at 4�C andtreated with 1% streptomycin sulfate. The supernatant wasthen used to perform the assay according to manufacturer’sinstructions. Protein concentration was determined usingthe Bradford protein assay (Bio-rad; Bio-Rad, Hercules,Calif).32
Histological analysis. After removal, soleus and EDLmuscles were rinsed with saline solution and pinned to aSylgard-coated petri dish containing 10% neutral buffered

Fig 1. Relative force-frequency relationships (P/Po) in theextensor digitorum longus (EDL; top panel) and soleus (lowerpanel) muscles. Values are as mean 6 standard error. #P < .05sham-operated controls vs ligated N-acetylcysteine (NAC); *P <
.05 Sham-operated controls vs ligated saline and ligated NAC;þP < .05 ligated saline vs sham-operated controls and ligatedNAC; &P < .05 sham-operated controls vs ligated saline.
JOURNAL OF VASCULAR SURGERY4 Roseguini et al --- 2013
formalin (Sigma-Aldrich). After approximately 1 hour, themuscles were transferred to a small container with formalinand allowed to fix for 24 hours prior to histological pro-cessing. The muscles were then sectioned at the mid-bellyand embedded in paraffin. Serial transverse sections (4 mm)were cut from the muscle samples and routinely stainedwith hematoxylin and eosin for determination of fiber CSAand proportion of centrally nucleated fibers, and withPicrosirius Red for determination of collagen deposition.Images were taken using an Olympus BX40 microscopeand a Q-color 3 digital camera (Olympus America, CenterValley, Pa). Nonoverlapping images of the entire musclecross-section were taken and analyzed using the ImagePro-
Express software (Media Cybernetics, Silver Spring, Md).The outlines of on average 130 fibers per muscle weremanually traced for determination of mean CSA. Collagendeposition was expressed as a percentage of the total tissuearea occupied by Sirius Red-stained muscle tissue.
Statistical analysis. Statistical analysis was conductedusing a commercially available software (SigmaPlot 11).Differences in fatigue characteristics were tested usingtwo-way repeated-measures analysis of variance and thepost hoc Tukey test. The percentage of fibers with centralnucleation was compared between ligated groups usingStudent t-test. The other variables were compared betweenthe three experimental groups using a one-way analysis ofvariance. Statistical significance was accepted when P <.05. Data are means 6 standard error of the mean.
RESULTS
Mice with bilateral ligation of the femoral arteryincluded in the study had no signs of ischemic injury otherthan mild darkening of the nails in some animals. Bodyweights were not different (P ¼ .226) between groupsbefore (sham-operated controls, 28.1 6 0.6 g; ligatedNAC, 28.3 6 0.4 g; ligated saline, 27.5 6 0.2 g) or aftertreatment (sham-operated controls, 30.8 6 0.6 g; ligatedNAC, 29.0 6 0.5 g; ligated saline, 29.2 6 0.3 g).
Contractile properties. EDL and soleus muscleweights and contractile properties are shown in Table I.Ligated animals had lower EDL muscle mass, Pt, absolute(P0) and specific (sP0) forces, and a slightly faster TPT (P <.05) compared with sham-operated controls. In the soleus,muscle mass and sP0 in the sham-operated controls werehigher compared with the ligated saline group (P < .05),whereas Pt and P0 differed from both the ligated saline andthe ligated NAC group (P < .05). The relative force-frequency relationships are depicted in Fig 1. In the EDLmuscle, ligated animals developed higher relative forces(P < .05) at the lower stimulation frequencies (30-80 Hz)when compared with sham-operated controls. A similarresponse was observed in the soleus muscle at frequencies15 to 30 Hz, but only in the ligated saline group (Fig 1).There were no statistical differences between groups in theF50 in the EDL or in the soleus (data not shown).
Fatigue characteristics. Fig 2 depicts fatigue charac-teristics in the EDL and soleus muscles. Overall, forcedecreased to a similar extent in all three groups during thefatigue trial in the EDL muscle. On the other hand, thedecline in force was markedly different in the entire stim-ulation period between ligated NAC and ligated salinegroups in the soleus muscle (Fig 2). For instance, forceafter 10 minutes of submaximal tetanic stimulation wassubstantially higher (P < .05) in NAC-treated animals(45.0% 6 3.0% of the initial value) when compared withligated saline (30.3% 6 3.0%). Force development was alsohigher (P < .05) in sham-operated controls whencompared with ligated saline during the first 5 minutes ofstimulation. There were no differences in the fatiguecharacteristics between ligated NAC and sham-operatedcontrols.

Fig 2. Fatigue characteristics in the extensor digitorum longus(EDL; top panel) and soleus (lower panel) muscles. Changes inforce are expressed relatively to the force developed in the firstcontraction of the protocol. The fatigue protocol consisted of 12tetanic contractions/min for 10 minutes and 18 contractions/minfor 5 minutes in the EDL muscle. In the soleus, the fatigue pro-tocol consisted of 18 contractions/min for 10 minutes and 24contractions/min for 5 min in the soleus muscle. Values are asmean 6 standard error. *P < .05 ligated N-acetylcysteine (NAC)vs ligated saline. #P < .05 sham-operated controls vs ligated saline.
JOURNAL OF VASCULAR SURGERYVolume -, Number - Roseguini et al 5
Muscle morphology. As shown in Table II, mean fi-ber CSA was higher in the EDL muscle of nonligatedcontrols when compared with ligated saline (P ¼ .014)and ligated NAC (P ¼ .051). Likewise, in the soleusmuscle, mean fiber CSA was on average 34% higher in non-ligated controls compared with ligated animals, althoughthis difference did not achieve statistical significance (P ¼.107). Assessment of the percentage of central nucleation(Fig 3) in the muscles revealed that NAC treatment tendedto reduce this parameter in both the EDL (ligated NAC,48% 6 14% vs ligated saline, 74% 6 3%; P ¼ .062) and
soleus muscles (ligated NAC, 58% 6 14% vs ligated saline,83% 6 8%; P ¼ .128).
Collagen deposition. Analysis of Picrosirius Red-stained sections revealed that collagen production was sub-stantially greater in ligated animals when compared withsham-operated controls (P < .05) in the EDL muscle,while there were no differences between the two ligatedgroups for this muscle (P ¼ .31; Fig 4). In the soleusmuscle, the area of collagen deposition was also lower insham-operated controls when compared with ligated saline(P < .01). Treatment with NAC tended to reduce the areaof fibrosis in treated animals when compared with thosethat received only saline (ligated NAC, 16%6 2% vs ligatedsaline, 24% 6 2%; P ¼ .057). Importantly, althoughcollagen infiltration tended to be lower in sham-operatedcontrols when compared with the ligated NAC group,this difference did not attain statistical significance (P ¼.072; Fig 4).
Biomarkers of oxidative damage. Levels of freemalondialdehyde (MDA) and protein carbonyl in thegastrocnemius muscle are shown in Fig 5. Free MDA wasnearly 44% lower in the sham-operated controls comparedwith the ligated groups (P < .05). Protein carbonyl contentfollowed a similar trend, but there were no statistical dif-ferences between groups for this parameter (P ¼ .09).Treatment with NAC did not cause a significant change inthe levels of MDA or protein carbonyls.
DISCUSSION
This study demonstrated that 2 weeks of treatmentwith NAC ameliorated the pathophysiology of the soleusbut not the EDL muscle in a mouse model of PAD. Thiscompound had marked effects on fatigue resistance in thesoleus muscle. However, NAC did not affect the levels ofmarkers of lipid peroxidation or protein oxidation. Overall,this is the first study to show that NAC, a safe and clinicallyapproved drug, has meaningful effects on skeletal muscle inperipheral arterial insufficiency.
Methodological considerations. A critical aspect inthe preclinical search for novel therapies for PAD is thatthe animal model should exhibit physiopathological fea-tures found in patients. Most important, it is imperativethat the therapy begins to be administered when theremodeling in response to the ischemic insult has beenresolved.33,34 Unfortunately, the vast majority of studiesin preclinical models of PAD start the treatment immedi-ately or shortly after induction of ischemia. As pointed outpreviously,33,34 this design is inappropriate as the therapyhas the potential to interact with the ongoing remodelingprocess triggered by surgically induced acute ischemia. Inthe present study, we have attempted to incorporate theseimportant concepts in the experimental design. First, wechose the Balb/c mouse strain because, as others haveshown previously,26,27 these mice develop histopatholog-ical features in response to ischemia that mimic the ab-normalities found in the skeletal muscle of patients withmoderate to severe PAD, such as central nucleation,inflammation, and fibrosis (Figs 3 and 4) as well as

Table II. Analysis of fiber cross-sectional area in the extensor digitorum longus (EDL) and soleus muscles from sham-operated controls, ligated animals treated intraperitoneally with N-acetylcysteine (NAC; 150 mg/kg), and ligated animalstreated with saline
EDL Soleus
Sham-operated Ligated saline Ligated NAC Sham-operated Ligated saline Ligated NAC
Mean cross-sectional area, mm2 975.3 6 93.1a 588.4 6 66.6 641.1 6 100 902.4 6 68.5 607.1 6 105 581.8 6 126
Values are as mean 6 standard error.aP < .05 vs ligated saline.
Fig 3. A, Representative photomicrographs of cross-sections of the extensor digitorum longus (EDL) and soleusmuscles processed for hematoxylin and eosin staining. Scale bar, 50 mm. B, Proportion of centrally nucleated fibers inligated N-acetylcysteine (NAC; n ¼ 6) and ligated saline (n ¼ 9). Muscles from sham-operated controls had no centralnuclei, and therefore, this group is not shown in the graph. Values are as mean 6 standard error.
JOURNAL OF VASCULAR SURGERY6 Roseguini et al --- 2013
increased levels of markers of oxidative damage (Fig 5).9,10
Second and most important, we started the treatment withNAC 2 weeks after the surgical occlusion of the femoralartery. We chose this time point because, in this specificmouse model, calf blood flow, as measured by magneticresonance imaging, and tissue oxygen tension, measured byelectron paramagnetic resonance (EPR) oximetry, are sta-ble within 14 days.26 Furthermore, edema and tissueswelling in the calf muscle that occur characteristically inthe first few days after surgical occlusion have largely beenresolved after 14 days of recovery. Thus, since most of thenatural compensation in response to the ischemic insult isresolved at this time point, it is possible to speculate that
the potential for an interaction between the systemic effectsof the surgical occlusion and treatment with NAC is largelyminimized.
NAC and skeletal muscle pathophysiology. Histo-pathological and imaging studies in patients with PADhave consistently documented a number of adverse adapta-tions in the calf muscles such as atrophy, connective tissueproliferation, and higher muscle percent fat.10,12,35 Thesubstitution of healthy, contractile tissue by fat and con-nective tissue is a common feature of chronic muscle dis-orders such as Duchenne muscular dystrophy that leads toirreversible derangement of muscle tissue organization andnegatively impacts muscle function.36,37 In the present
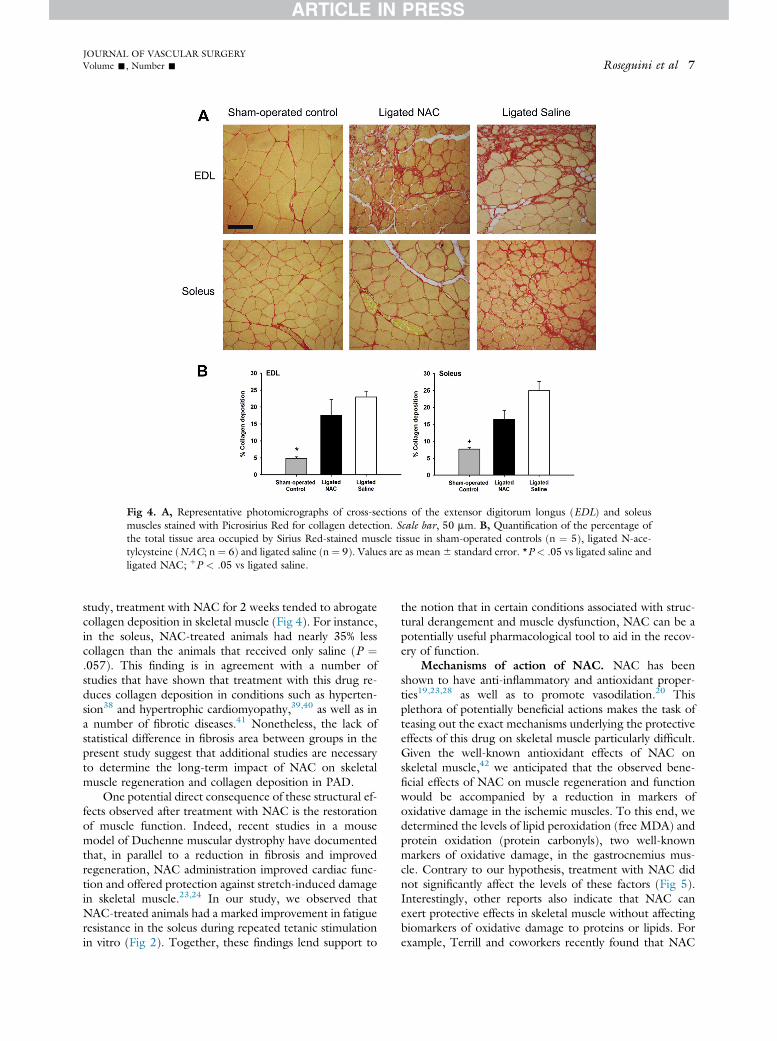
Fig 4. A, Representative photomicrographs of cross-sections of the extensor digitorum longus (EDL) and soleusmuscles stained with Picrosirius Red for collagen detection. Scale bar, 50 mm. B, Quantification of the percentage ofthe total tissue area occupied by Sirius Red-stained muscle tissue in sham-operated controls (n ¼ 5), ligated N-ace-tylcysteine (NAC; n ¼ 6) and ligated saline (n ¼ 9). Values are as mean6 standard error. *P < .05 vs ligated saline andligated NAC; þP < .05 vs ligated saline.
JOURNAL OF VASCULAR SURGERYVolume -, Number - Roseguini et al 7
study, treatment with NAC for 2 weeks tended to abrogatecollagen deposition in skeletal muscle (Fig 4). For instance,in the soleus, NAC-treated animals had nearly 35% lesscollagen than the animals that received only saline (P ¼.057). This finding is in agreement with a number ofstudies that have shown that treatment with this drug re-duces collagen deposition in conditions such as hyperten-sion38 and hypertrophic cardiomyopathy,39,40 as well as ina number of fibrotic diseases.41 Nonetheless, the lack ofstatistical difference in fibrosis area between groups in thepresent study suggest that additional studies are necessaryto determine the long-term impact of NAC on skeletalmuscle regeneration and collagen deposition in PAD.
One potential direct consequence of these structural ef-fects observed after treatment with NAC is the restorationof muscle function. Indeed, recent studies in a mousemodel of Duchenne muscular dystrophy have documentedthat, in parallel to a reduction in fibrosis and improvedregeneration, NAC administration improved cardiac func-tion and offered protection against stretch-induced damagein skeletal muscle.23,24 In our study, we observed thatNAC-treated animals had a marked improvement in fatigueresistance in the soleus during repeated tetanic stimulationin vitro (Fig 2). Together, these findings lend support to
the notion that in certain conditions associated with struc-tural derangement and muscle dysfunction, NAC can be apotentially useful pharmacological tool to aid in the recov-ery of function.
Mechanisms of action of NAC. NAC has beenshown to have anti-inflammatory and antioxidant proper-ties19,23,28 as well as to promote vasodilation.20 Thisplethora of potentially beneficial actions makes the task ofteasing out the exact mechanisms underlying the protectiveeffects of this drug on skeletal muscle particularly difficult.Given the well-known antioxidant effects of NAC onskeletal muscle,42 we anticipated that the observed bene-ficial effects of NAC on muscle regeneration and functionwould be accompanied by a reduction in markers ofoxidative damage in the ischemic muscles. To this end, wedetermined the levels of lipid peroxidation (free MDA) andprotein oxidation (protein carbonyls), two well-knownmarkers of oxidative damage, in the gastrocnemius mus-cle. Contrary to our hypothesis, treatment with NAC didnot significantly affect the levels of these factors (Fig 5).Interestingly, other reports also indicate that NAC canexert protective effects in skeletal muscle without affectingbiomarkers of oxidative damage to proteins or lipids. Forexample, Terrill and coworkers recently found that NAC

Fig 5. Free malondialdehyde (panel A) and protein carbonyl(panel B) levels in the gastrocnemius muscles of sham-operatedcontrols (n ¼ 6), ligated N-acetylcysteine (NAC; n ¼ 6) andligated saline (n ¼ 8). Values are as mean 6 standard error. *P <
.05 vs ligated saline and ligated NAC.
JOURNAL OF VASCULAR SURGERY8 Roseguini et al --- 2013
treatment in the mdx mice prevented exercise-inducedmyonecrosis but failed to prevent the changes in skeletalmuscle lipid and protein oxidation levels.43 Together, theseobservations suggest that an alternative mechanism plays amajor role in the positive adaptation triggered by thiscompound in skeletal muscle. One salutary effect of NACthat can be particularly meaningful for PAD is its actions onthe vasculature. Studies in vitro documented that NACpromotes vasodilation in part by activating voltage-gatedKþ channels.20 Recently, Nyberg and coworkers44 haveshown that intravenous infusion of NAC markedlyenhanced nitric oxide bioavailability in the systemic circu-lation and in the musculature of sedentary aging humans.Based on these reports, it is tempting to speculate thatrepeated administration of NAC may have enhanced bloodflow recovery in the ligated animals, thereby improvingmuscle regeneration. Future studies should determine thelong-term consequences of NAC on blood flow andvascular structure in this model of PAD.
Fiber type differences. A key observation of the pre-sent study was that the effects of NAC were mostly evidentin the slow-twitch soleus muscle, whereas minimal effects,especially in regard to fatigue resistance, were noted in the
fast-twitch EDL. These findings indicate that NAC canevoke substantially different responses depending onmuscle fiber type composition. The mechanistic basis un-derlying this apparent muscle-specific effect of NAC are notclear at this point. It is worth noting that postural, slow-twitch muscles such as the soleus have higher basaloxidative stress45,46 and higher resting blood flow thanpredominantly white muscles.47 Given that NAC hasantioxidant and vasoactive properties,20 it is conceivablethat its effects in resting conditions would be particularlymeaningful for the regenerative process in the soleusmuscle. Nonetheless, these findings highlight the impor-tance of future studies to evaluate the therapeutic effects ofthis drug in muscles composed of different fiber types.
Clinical applications. Despite the increasing burdenof PAD and its detrimental consequences on the qualityof life of the patients, few pharmacological therapies haveshown to evoke meaningful effects on functional perfor-mance in these individuals.15 Pentoxifylline and cil-ostazol, the two medications currently approved in theUnited States for symptomatic improvement in PAD, areknown to have rather modest effects on walking toler-ance.48 The development of novel pharmacological ap-proaches capable of increasing exercise tolerance in patientswith this condition is therefore highly desirable. NAC isapproved for clinical use, has minimal side effects, and it iswell-tolerated during long-term administration.49-51 Mostimportantly, NAC administration has shown to be consis-tently associated with improvements in exercise perfor-mance in animals and humans.21 For example, Koechlinand coworkers52 reported that short-term administration ofNAC increased time to exhaustion during knee extensionexercise and attenuated exercise-induced oxidative stress inpatients with chronic obstructive pulmonary disease. In thisstudy, we showed for the first time that treatment with thisdrug with a dose amenable for clinical application evokedmarked effects on fatigue resistance in the soleus muscle ina mouse model of PAD (Fig 2). Taken together, theseencouraging findings set the stage for translational studiesto determine the acute and long-term impact of this drugon walking capacity in patients with PAD.
Limitations. One important limitation of this study isthat we evaluated the effects of only one dose of NAC(150 mg/kg) given intraperitoneally. This dose was chosenbecause it has previously shown to significantly attenuateinflammation, lipid peroxidation, and myonecrosis in thediaphragm of mdx mice when given daily over 14 consec-utive days.28 Importantly, when normalizing to body sur-face area, the dose used in the present investigation wouldbe equivalent to w12 mg/kg in humans, which equates to851 mg of NAC for a 70 kg person.53 This is considered alow dose, as most clinical studies typically use doses rangingfrom 1200 to 1800 mg per day.49-52 Nonetheless, giventhe expected species differences in the pharmacokinetics ofthe drug, additional studies are warranted to determine theoptimal dose to maximize the benefits of NAC.
It is also important to highlight that, although the an-imal model employed in the present investigation has some

JOURNAL OF VASCULAR SURGERYVolume -, Number - Roseguini et al 9
of the myopathic changes commonly found in patients withPAD, the surgical induction of ischemia does not mimicthe complexity and chronicity of the atherosclerotic pathol-ogy and its associated impact on human skeletal muscle.Indeed, as opposed to the animals used in this study,elderly patients with PAD have multiple comorbiditiesand are typically treated with pharmacological agents thatare known to have an impact on skeletal muscle, such asstatins and angiotensin-converting enzyme inhibitors.15
Therefore, direct extrapolation of the findings from thisstudy to the clinical setting requires caution.
The authors thank Dr Leonardo Ferreira for his adviceand assistance in method development and are also gratefulto Antonio Souza for his assistance in the histologicalprocessing.
AUTHOR CONTRIBUTIONS
Conception and design: BR, SH, CSAnalysis and interpretation: BR, LS, TP, MB, CS, SHData collection: BR, LS, TPWriting the article: BRCritical revision of the article: MB, SH, CSFinal approval of the article: BR, LS, TP, MB, CS, SHStatistical analysis: BRObtained funding: CS, SHOverall responsibility: BR
REFERENCES
1. Allison MA, Ho E, Denenberg JO, Langer RD, Newman AB,Fabsitz RR, et al. Ethnic-specific prevalence of peripheral arterial dis-ease in the United States. Am J Prev Med 2007;32:328-33.
2. McDermott MM. The magnitude of the problem of peripheral arterialdisease: epidemiology and clinical significance. Cleve Clin J Med2006;73(Suppl 4):S2-7.
3. McDermott MM, Liu K, Greenland P, Guralnik JM, Criqui MH,Chan C, et al. Functional decline in peripheral arterial disease: associ-ations with the ankle brachial index and leg symptoms. JAMA2004;292:453-61.
4. Regensteiner JG, Hiatt WR, Coll JR, Criqui MH, Treat-Jacobson D,McDermott MM, et al. The impact of peripheral arterial disease onhealth-related quality of life in the Peripheral Arterial Disease Aware-ness, Risk, and Treatment: New Resources for Survival (PARTNERS)Program. Vasc Med 2008;13:15-24.
5. Sieminski DJ, Gardner AW. The relationship between free-living dailyphysical activity and the severity of peripheral arterial occlusive disease.Vasc Med 1997;2:286-91.
6. Anderson JD, Epstein FH, Meyer CH, Hagspiel KD, Wang H, Berr SS,et al. Multifactorial determinants of functional capacity in peripheralarterial disease: uncoupling of calf muscle perfusion and metabolism.J Am Coll Cardiol 2009;54:628-35.
7. McDermott MM, Hoff F, Ferrucci L, Pearce WH, Guralnik JM,Tian L, et al. Lower extremity ischemia, calf skeletal muscle charac-teristics, and functional impairment in peripheral arterial disease. J AmGeriatr Soc 2007;55:400-6.
8. Regensteiner JG, Wolfel EE, Brass EP, Carry MR, Ringel SP,Hargarten ME, et al. Chronic changes in skeletal muscle histology andfunction in peripheral arterial disease. Circulation 1993;87:413-21.
9. Hedberg B, Angquist KA, Sjostrom M. Peripheral arterial insuffi-ciency and the fine structure of the gastrocnemius muscle. IntAngiol 1988;7:50-9.
10. Pipinos II, Swanson SA, Zhu Z, Nella AA, Weiss DJ, Gutti TL, et al.Chronically ischemic mouse skeletal muscle exhibits myopathy in
association with mitochondrial dysfunction and oxidative damage. AmJ Physiol Regul Integr Comp Physiol 2008;295:R290-6.
11. Pipinos II, Judge AR, Zhu Z, Selsby JT, Swanson SA, Johanning JM,et al. Mitochondrial defects and oxidative damage in patients withperipheral arterial disease. Free Radic Biol Med 2006;41:262-9.
12. McDermott MM, Ferrucci L, Guralnik J, Tian L, Liu K, Hoff F, et al.Pathophysiological changes in calf muscle predict mobility loss at 2-yearfollow-up in men and women with peripheral arterial disease. Circu-lation 2009;120:1048-55.
13. McDermott MM, Liu K, Tian L, Guralnik JM, Criqui MH, Liao Y,et al. Calf muscle characteristics, strength measures, and mortality inperipheral arterial disease: a longitudinal study. J Am Coll Cardiol2012;59:1159-67.
14. Hiatt WR, Regensteiner JG, Wolfel EE, Carry MR, Brass EP. Effect ofexercise training on skeletal muscle histology and metabolism in pe-ripheral arterial disease. J Appl Physiol 1996;81:780-8.
15. Berger JS, Hiatt WR. Medical therapy in peripheral artery disease.Circulation 2012;126:491-500.
16. Judge AR, Dodd SL. Oxidative damage to skeletal muscle following anacute bout of contractile claudication. Atherosclerosis 2003;171:219-24.
17. Judge AR, Dodd SL. Xanthine oxidase and activated neutrophils causeoxidative damage to skeletal muscle after contractile claudication. Am JPhysiol Heart Circ Physiol 2004;286:H252-6.
18. Pipinos II, Judge AR, Selsby JT, Zhu Z, Swanson SA, Nella AA, et al.The myopathy of peripheral arterial occlusive disease: part 2. Oxidativestress, neuropathy, and shift in muscle fiber type. Vasc EndovascularSurg 2008;42:101-12.
19. Zafarullah M, Li WQ, Sylvester J, Ahmad M. Molecular mechanisms ofN-acetylcysteine actions. Cell Mol Life Sci 2003;60:6-20.
20. Han WQ, Zhu DL, Wu LY, Chen QZ, Guo SJ, Gao PJ. N-ace-tylcysteine-induced vasodilation involves voltage-gated potassiumchannels in rat aorta. Life Sci 2009;84:732-7.
21. Ferreira LF, Reid MB. Muscle-derived ROS and thiol regulation inmuscle fatigue. J Appl Physiol 2008;104:853-60.
22. Hernandez A, Cheng A, Westerblad H. Antioxidants and skeletalmuscle performance: “common knowledge” vs. experimental evidence.Front Physiol 2012;3:46.
23. Whitehead NP, Pham C, Gervasio OL, Allen DG. N-Acetylcysteineameliorates skeletal muscle pathophysiology in mdx mice. J Physiol2008;586:2003-14.
24. Williams IA, Allen DG. The role of reactive oxygen species in the heartsof dystrophin-deficient mdx mice. Am J Physiol Heart Circ Physiol2007;293:H1969-77.
25. Chalothorn D, Faber JE. Strain-dependent variation in collateral cir-culatory function in mouse hindlimb. Physiol Genomics 2010;42:469-79.
26. Helisch A, Wagner S, Khan N, Drinane M, Wolfram S, Heil M, et al.Impact of mouse strain differences in innate hindlimb collateralvasculature. Arterioscler Thromb Vasc Biol 2006;26:520-6.
27. McClung JM, McCord TJ, Keum S, Johnson S, Annex BH,Marchuk DA, et al. Skeletal muscle-specific genetic determinantscontribute to the differential strain-dependent effects of hindlimbischemia in mice. Am J Pathol 2012;180:2156-69.
28. de Senzi Moraes, Pinto R, Ferretti R, Moraes LH, Neto HS,Marques MJ, et al. N-Acetylcysteine treatment reduces TNF-alphalevels and myonecrosis in diaphragm muscle of mdx mice. Clin Nutr2013;32:472-5.
29. Souccar C, Goncalo Mdo C, Buck Hde S, Lima-Landman MT,Lapa AJ. Mild dystrophic damage in the androgen-sensitive levator animuscle of the mdx mouse. Neuromuscul Disord 2005;15:48-56.
30. Close RI. Dynamic properties of mammalian skeletal muscles. PhysiolRev 1972;52:129-97.
31. Kinnaird T, Stabile E, Burnett MS, ShouM, Lee CW, Barr S, et al. Localdelivery of marrow-derived stromal cells augments collateral perfusionthrough paracrine mechanisms. Circulation 2004;109:1543-9.
32. Bradford MM. A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dyebinding. Anal Biochem 1976;72:248-54.
33. Hoefer IE, van Royen N, Jost MM. Experimental models of arterio-genesis: differences and implications. Lab Anim (NY) 2006;35:36-44.

JOURNAL OF VASCULAR SURGERY10 Roseguini et al --- 2013
34. Ziegler MA, Distasi MR, Bills RG, Miller SJ, Alloosh M, Murphy MP,et al. Marvels, mysteries, and misconceptions of vascular compensationto peripheral artery occlusion. Microcirculation 2010;17:3-20.
35. McDermott MM, Greenland P, Liu K, Guralnik JM, Celic L,Criqui MH, et al. The ankle brachial index is associated with legfunction and physical activity: the Walking and Leg Circulation Study.Ann Int Med 2002;136:873-83.
36. Bernasconi P, Torchiana E, Confalonieri P, Brugnoni R, Barresi R,Mora M, et al. Expression of transforming growth factor-beta 1 indystrophic patient muscles correlates with fibrosis. Pathogenetic role ofa fibrogenic cytokine. J Clin Invest 1995;96:1137-44.
37. Zhou L, Lu H. Targeting fibrosis in Duchenne muscular dystrophy.J Neuropathol Exp Neurol 2010;69:771-6.
38. Bourraindeloup M, Adamy C, Candiani G, Cailleret M, Bourin MC,Badoual T, et al. N-acetylcysteine treatment normalizes serum tumornecrosis factor-alpha level and hinders the progression of cardiac injuryin hypertensive rats. Circulation 2004;110:2003-9.
39. Lombardi R, Rodriguez G, Chen SN, Ripplinger CM, Li W, Chen J,et al. Resolution of established cardiac hypertrophy and fibrosis andprevention of systolic dysfunction in a transgenic rabbit model of hu-man cardiomyopathy through thiol-sensitive mechanisms. Circulation2009;119:1398-407.
40. Marian AJ, Senthil V, Chen SN, Lombardi R. Antifibrotic effects ofantioxidant N-acetylcysteine in a mouse model of human hypertrophiccardiomyopathy mutation. J Am Coll Cardiol 2006;47:827-34.
41. Liu RM, Gaston Pravia KA. Oxidative stress and glutathione in TGF-beta-mediated fibrogenesis. Free Radic Biol Med 2010;48:1-15.
42. Medved I, Brown MJ, Bjorksten AR, Murphy KT, Petersen AC,Sostaric S, et al. N-acetylcysteine enhances muscle cysteine and gluta-thione availability and attenuates fatigue during prolonged exercise inendurance-trained individuals. J Appl Physiol 2004;97:1477-85.
43. Terrill JR, Radley-Crabb HG, Grounds MD, Arthur PG. N-Ace-tylcysteine treatment of dystrophic mdx mice results in protein thiolmodifications and inhibition of exercise induced myofibre necrosis.Neuromuscul Disord 2012;22:427-34.
44. Nyberg M, Blackwell JR, Damsgaard R, Jones AM, Hellsten Y,Mortensen SP. Lifelong physical activity prevents an age-related
reduction in arterial and skeletal muscle nitric oxide bioavailability inhumans. J Physiol 2012;590(Pt 21):5361-70.
45. Ji LL, Fu R, Mitchell EW. Glutathione and antioxidant enzymes inskeletal muscle: effects of fiber type and exercise intensity. J ApplPhysiol 1992;73:1854-9.
46. Powers SK, Jackson MJ. Exercise-induced oxidative stress: cellularmechanisms and impact on muscle force production. Physiol Rev2008;88:1243-76.
47. Armstrong RB, Laughlin MH. Metabolic indicators of fibre recruit-ment in mammalian muscles during locomotion. J Exp Biol 1985;115:201-13.
48. Stevens JW, Simpson E, Harnan S, Squires H, Meng Y, Thomas S,et al. Systematic review of the efficacy of cilostazol, naftidrofuryl oxalateand pentoxifylline for the treatment of intermittent claudication. Br JSurg 2012;99:1630-8.
49. De Mattia G, Bravi MC, Laurenti O, Cassone-Faldetta M, Proietti A,De Luca O, et al. Reduction of oxidative stress by oral N-acetyl-L-cysteine treatment decreases plasma soluble vascular cell adhesionmolecule-1 concentrations in non-obese, non-dyslipidaemic, normo-tensive, patients with non-insulin-dependent diabetes. Diabetologia1998;41:1392-6.
50. Jeremias A, Soodini G, Gelfand E, Xu Y, Stanton RC, Horton ES, et al.Effects of N-acetyl-cysteine on endothelial function and inflammationin patients with type 2 diabetes mellitus. Heart Int 2009;4:e7.
51. Martina V, Masha A, Gigliardi VR, Brocato L, Manzato E, Berchio A,et al. Long-term N-acetylcysteine and L-arginine administration re-duces endothelial activation and systolic blood pressure in hypertensivepatients with type 2 diabetes. Diabetes Care 2008;31:940-4.
52. Koechlin C, Couillard A, Simar D, Cristol JP, Bellet H, Hayot M, et al.Does oxidative stress alter quadriceps endurance in chronic obstructivepulmonary disease? Am J Respir Crit Care Med 2004;169:1022-7.
53. Reagan-Shaw S, Nihal M, Ahmad N. Dose translation from animal tohuman studies revisited. FASEB J 2008;22:659-61.
Submitted Aug 8, 2013; accepted Oct 24, 2013.
Copyright © 2022 FDOKUMEN




















