
Effects of mood stabilizers on brain reward processes in rats: Studies using the intracranial...
10
Effects of mood stabilizers on brain reward processes in rats: Studies using the intracranial self-stimulation paradigm Maria Mavrikaki a , George G. Nomikos b , George Panagis a, ⁎ a University of Crete, School of Social Sciences, Department of Psychology, Laboratory of Behavioral Neuroscience, 74100 Rethymno, Crete, Greece b Takeda Global Research & Development Center, Inc., Deerfield, Illinois 60015, USA Received 23 September 2008; received in revised form 5 November 2008; accepted 12 November 2008 KEYWORDS Brain stimulation reward; Mood stabilizers; d-Amphetamine; Reinforcement; Animal model Abstract Bipolar disorder is characterized by dysregulated motivation and increased hedonistic drive. d- Amphetamine induces manic symptoms in humans and exacerbates mania in bipolar disorder patients, effects that are counteracted by mood stabilizers. We utilized intracranial self-stimulation (ICSS) to examine how lithium (LiCl), valproate (VPA) or their combination that is commonly used in the clinic affect brain reward function in rats, and how these drugs affect d-amphetamine's reward-facilitating effects. Acute intraperitoneal (i.p.) administration of LiCl (100, 200 mg/kg), VPA (400 mg/kg) or combined administration of subthreshold doses of LiCl (50 mg/kg) and VPA (200 mg/kg) increased ICSS thresholds. LiCl (100 mg/kg) and combined administration of LiCl and VPA (50 and 200 mg/kg), but not VPA alone (200, 400 mg/kg), attenuated d-amphetamine's reward- facilitating effects. These results suggest that ICSS combined with d-amphetamine constitutes a useful model to explore the elation and increased hedonistic drive observed in bipolar patients and ultimately help to identify novel pharmacotherapies for bipolar disorder. © 2008 Elsevier B.V. and ECNP. All rights reserved. 1. Introduction Bipolar disorder is a common, severe, chronic and often life- threatening neuropsychiatric disorder. It is characterized by progressive and spontaneously alternating episodes of mania and depression, usually interspersed with periods of relatively normal mood (Belmaker, 2004; Goodwin and Jamison, 2007). The precise etiology and pathophysiology of bipolar disorder remain largely unknown. This might be related inter alia to the absence of an adequate animal model to study its pathophysiol- ogy. Undoubtedly, this fact has also largely hampered the development of novel therapeutics for bipolar disorder. Thus, lithium and anticonvulsants remain for many years the primary treatments for bipolar patients (Baldessarini, 2002; Fountoula- kis et al., 2007; Kahn et al., 2000; Kasper, 2003; Moller and Nasrallah, 2003), although adherence is often poor, since many patients fail to respond adequately or tolerate them. The cycling nature of bipolar disorder presents a unique difficulty in any attempts to model it in experimental ⁎ Corresponding author. Fax: +30 28310 77548. E-mail address: [email protected] (G. Panagis). 0924-977X/$ - see front matter © 2008 Elsevier B.V. and ECNP. All rights reserved. doi:10.1016/j.euroneuro.2008.11.002 www.elsevier.com/locate/euroneuro European Neuropsychopharmacology (2009) 19, 205–214
Transcript of Effects of mood stabilizers on brain reward processes in rats: Studies using the intracranial...

www.e l sev i e r. com/ loca te / eu roneu ro
European Neuropsychopharmacology (2009) 19, 205–214
Effects of mood stabilizers on brain reward processesin rats: Studies using the intracranialself-stimulation paradigmMaria Mavrikaki a, George G. Nomikos b, George Panagis a,⁎
a University of Crete, School of Social Sciences, Department of Psychology, Laboratory of Behavioral Neuroscience,74100 Rethymno, Crete, Greeceb Takeda Global Research & Development Center, Inc., Deerfield, Illinois 60015, USA
Received 23 September 2008; received in revised form 5 November 2008; accepted 12 November 2008
⁎ Corresponding author. Fax: +30 283E-mail address: [email protected]
0924-977X/$ - see front matter © 200doi:10.1016/j.euroneuro.2008.11.002
KEYWORDSBrain stimulation reward;Mood stabilizers;d-Amphetamine;Reinforcement;Animal model
Abstract
Bipolar disorder is characterized by dysregulated motivation and increased hedonistic drive. d-Amphetamine induces manic symptoms in humans and exacerbates mania in bipolar disorderpatients, effects that are counteracted bymood stabilizers. We utilized intracranial self-stimulation(ICSS) to examine how lithium (LiCl), valproate (VPA) or their combination that is commonly usedin the clinic affect brain reward function in rats, and how these drugs affect d-amphetamine's
reward-facilitating effects. Acute intraperitoneal (i.p.) administration of LiCl (100, 200 mg/kg),VPA (400 mg/kg) or combined administration of subthreshold doses of LiCl (50 mg/kg) and VPA(200 mg/kg) increased ICSS thresholds. LiCl (100 mg/kg) and combined administration of LiCl andVPA (50 and 200 mg/kg), but not VPA alone (200, 400 mg/kg), attenuated d-amphetamine's reward-facilitating effects. These results suggest that ICSS combined with d-amphetamine constitutes auseful model to explore the elation and increased hedonistic drive observed in bipolar patients andultimately help to identify novel pharmacotherapies for bipolar disorder.© 2008 Elsevier B.V. and ECNP. All rights reserved.10 77548.c.gr (G. Panagis).
8 Elsevier B.V. and ECNP. All ri
1. Introduction
Bipolar disorder is a common, severe, chronic and often life-threatening neuropsychiatric disorder. It is characterized byprogressive and spontaneously alternating episodes of maniaand depression, usually interspersed with periods of relativelynormal mood (Belmaker, 2004; Goodwin and Jamison, 2007).The precise etiology and pathophysiology of bipolar disorder
ght
remain largely unknown. This might be related inter alia to theabsence of an adequate animal model to study its pathophysiol-ogy. Undoubtedly, this fact has also largely hampered thedevelopment of novel therapeutics for bipolar disorder. Thus,lithium and anticonvulsants remain for many years the primarytreatments for bipolar patients (Baldessarini, 2002; Fountoula-kis et al., 2007; Kahn et al., 2000; Kasper, 2003; Moller andNasrallah, 2003), although adherence is often poor, since manypatients fail to respond adequately or tolerate them.
The cycling nature of bipolar disorder presents a uniquedifficulty in any attempts to model it in experimental
s reserved.

206 M. Mavrikaki et al.
animals. Thus, mostmodels tend to focus only on the one poleof the disorder. Although several valid models for thedepressive state are available, there are no ideal animalmodels for the manic pole of the disease. Nevertheless, avariety of certain pharmacologically- or physiologically-elicited behaviors in animals might represent certain facetsof the disease (Einat et al., 2003; Einat, 2007). Since a clinicalhallmark in the diagnosis of bipolar disorder is the presence ofmanic symptoms, an adequate animal model should resemblesome features of the manic state of the disease (i.e. themanic episode), such as euphoria, increased hedonistic driveand motivation, increased sexual drive, hyperactivity,insomnia, risk-taking behavior or aggressiveness.
Several behavioral tests for such constituents are available.An animalmodel thatmay be particularly useful in the study ofthe motivational state of an organism is the intracranial self-stimulation (ICSS) paradigm (Carlezon and Chartoff, 2007;Markou and Koob, 1992). This behavioral paradigm is based onthe discovery by Olds and Milner (1954) that rats willrepeatedly press a lever to stimulate specific components ofthe brain reward circuit. A number of studies have confirmedthe validity of this technique for the assessment of both therewarding and the anhedonic effects of drugs or othermanipulations (Kornetsky, 1985; Markou and Koob, 1991;Paterson et al., 2000; Roybal et al., 2007; Slattery et al.,2007; Wise and Munn, 1995; Wise, 1996). Thus, ICSS behaviormay represent a useful behavioral measurement of the brainreward function with which to study preclinically thischaracteristic of mania. Interestingly, Roybal et al. (2007) ina very elegant study have recently shown that Clock mutantmice that display an overall behavioral profile, whichresembles the symptomatology of human mania, demonstratean increase in the reward value of cocaine, sucrose and ICSS.Chronic administration of lithium returns these behavioralresponses to wild-type levels. Furthermore Clockmutant micedisplay a hyperfunctioning dopaminergic system, which is welldocumented that it regulates brain reward function andmood.
The most frequently used pharmacological model for maniais the psychostimulant-induced hyperactivity. Psychostimu-lants, including d-amphetamine, induce manic symptoms inhumans and exacerbate mania in individuals with bipolardisorder (Anand et al., 2000; Angrist et al., 1987; Brauer anddeWit, 1996; Flemenbaum, 1974; Jacobs and Silverstone, 1986;Strakowski and Sax, 1998; Van Kammen and Murphy, 1975;Zacny et al., 1992). In general, these effects are counteractedby mood stabilizers, like lithium and valproate (Bell et al.,2005a,b; Silverstoneet al., 1998;Willsonet al., 2005).However,psychostimulants do not only inducemanic-like hyperactivity inexperimental animals. Psychostimulant administration affectsmore than one facet of mania-like behavior (i.e. increasesactivity, reduces sleep, produces risk-taking behavior, increaseshedonistic drive and induces euphoria). Moreover, most peopleabuse these drugs because of the elating effects they induce totheir mood. Interestingly, one of themain symptoms of mania isthe propensity towards drug abuse, and research has consis-tently shown that bipolar patients have an extremely high rateof co-occurring substance use disorder (Altamura, 2007; Cerulloand Strakowski, 2007; Maremmani et al., 2008; McIntyre et al.,2007). Accordingly, there is evidence for an abnormal brainreward systemactivation in bipolar patients (Abler et al., 2008).
Psychostimulants have been reported to facilitate ICSSbehavior in rats (Bossert and Franklin, 2003; Cassens and
Mills, 1973; Gallistel and Karras, 1984; Kornetsky andEsposito, 1979; Wise and Munn, 1993; Wise, 1996), indicatinga hyperfunction of brain reward system. This might be wellrelated to the increases in elation and hedonistic driveobserved in bipolar patients. On the other hand, the effectsof mood stabilizers on ICSS have not been studied in depth. Alimited number of studies have shown that lithium hasopposite effects on threshold for ICSS (Cassens and Mills,1973; Tomasiewicz et al., 2006), whereas according toTomasiewicz et al. (2006) valproic acid (VPA) did not affectICSS thresholds. Furthermore, although in the clinicalpractice a number of patients are treated with adjunctivetreatment, which combines both lithium and valproate, ithas not been investigated how this treatment may affectbrain reward function. Similarly, it has not been investigatedif mood stabilizers like lithium and VPA affect the reinforcingactions of amphetamine in the ICSS paradigm.
The present study was designed to compare the effects ofacute administration of lithium chloride (LiCl), VPA and theircombined administration on ICSS behavior in rats. Further-more, we studied the effects of LiCl, VPA and their combinedadministration on the reward-facilitating effects of d-amphetamine in ICSS.
We believe that using the ICSS paradigm we can explorethe hedonistic properties that amphetamine produces inexperimental animals. Thus, potentially this model can beused to explore the therapeutic actions of mood stabilizers,to explore the common denominator in their actions or evenin the development and testing of novel compounds withsimilar therapeutic effects.
2. Experimental procedures
2.1. Animals and surgery
Male Sprague–Dawley rats weighting 300–370 g at the time ofsurgery were used. Before surgery they were housed in groups ofthree and maintained on a 12 h light–12 h dark cycle with freeaccess to food and water. The animals were anaesthetized withintramuscular (i.m.) injection of ketamine hydrochloride (100 mg/kg) and xylazine (10 mg/kg). Atropine sulphate (0.6 mg/kg, i.m.)was injected to reduce bronchial secretion. The animals wereimplanted with a monopolar stimulation electrode aimed at themedial forebrain bundle (MFB) at the level of lateral hypothalamus(2.56 mm posterior to bregma, 1.8 mm lateral from midsaggitalsuture, and 8.6 below the outer flat skull), according to Paxinos andWatson (2007). The electrodes were constructed from 0.25 mmstainless steel wire insulated with Epoxylite except for the conicallyshaped tip. The anode was an uninsulated stainless steel wireconnected to an amphenol pin. Five miniature skull screws, theelectrode and the anode were secured to the skull with acrylicdental cement. Following implantation and for the entire durationof the experiments, the animals were housed individually.
Experiments were conducted in accordance with the 1996 Guidefor the care and Use of Laboratory Animals (NIH).
2.2. Apparatus and procedures for self-stimulation
After one week recovery, the rats were tested for self-stimulation inan operant chamber made of transparent Plexiglas (25 cm wide,25 cm deep and 30 cm high). Each chamber was equipped with astainless-steel poke device (lever) 4 cm wide and protruded 2 cmfrom the left side at a height of 4 cm from the bottom. Each bar-press triggered a constant current stimulator (Med Associates, St.Albans, VT) that delivered a 0.4-s train of rectangular cathodal

207Effects of mood stabilizers on brain reward processes in rats
pulses of constant duration (0.1 ms) and intensity (250 µA) andvariable frequency (15–100 Hz, i.e. 6–40 number of pulses/0.4 s).The pulse frequency, i.e. the number of pulses within a train, wasprogressively increased up to 40 per stimulation train until thesubject showed vigorous self-stimulation. During the acquisitionphase the animals were trained to self-stimulate for at least threeconsecutive days (1 h daily), using stimulation parameters thatmaintained near maximal bar-pressing rates. After self-stimulationhas been acquired and stabilized for a given pulse frequency, ratswere trained to self-stimulate using four alternating series ofascending and descending pulse frequencies. The pulse frequencywas varied by steps of approximately 0.1 log units. Each frequencywas tested within trials of 60 s in duration, followed by an extinctionperiod of 30 s. For each trial, there was an initial “priming” phaseduring which the animals received three trains of stimulation atthe frequency which was available for the specific trial. A rate-frequency determination session lasted about 45 min. One rate-frequency curve was established daily, for 10–14 days, depending onthe period when the self-stimulation indices (i.e. curve shift andthreshold measure) were stable. The stimulation parameters,intracranial self-stimulation sessions and data collection werecontrolled by a computer.
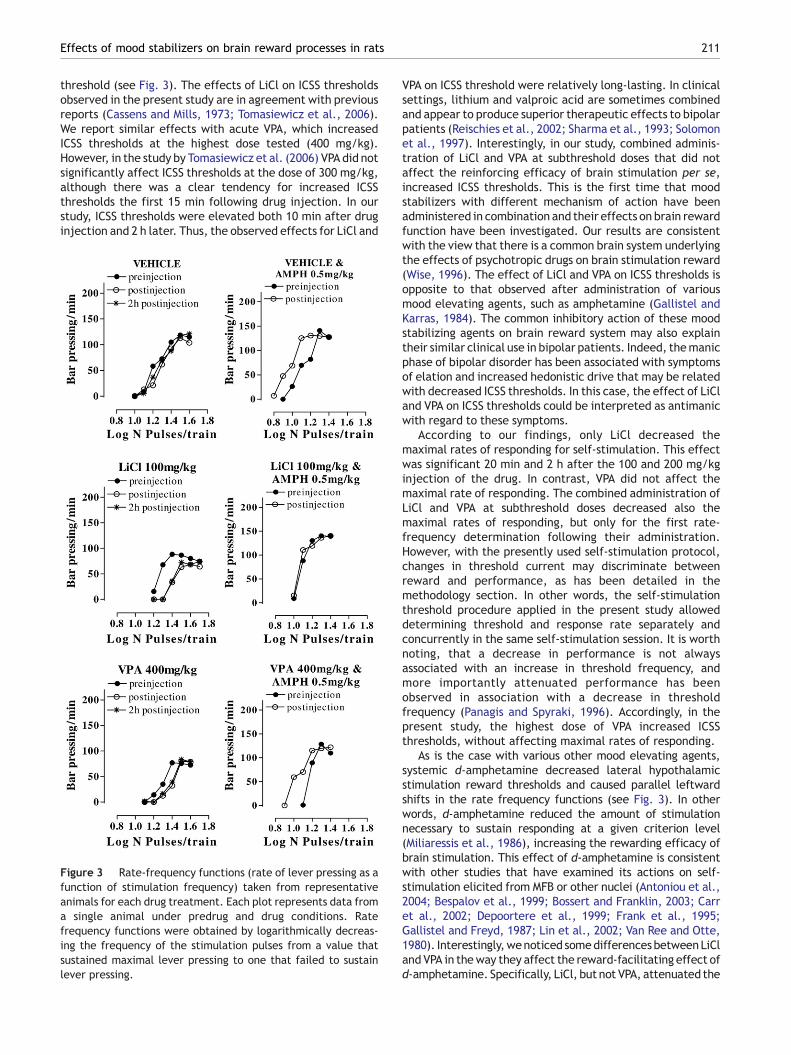
Unequivocally, intracranial self-stimulation behavior has theadvantage of not being affected by satiation or dysphoric effects,which are potentially modulated by various drug treatments. On theother hand, since both mood stabilizers and amphetamine seem toaffect motor activity/performance capacity in a dose-dependentmanner, the use of a reward selective measure, like the curve-shift,was requisite. In this method, plotting the responses of the animalsagainst the various pulse frequencies yields a sigmoidal rate-frequencycurve as shown in Fig. 3. Shifts in the lateral position of the curveprovide a selective measure of stimulation-produced reward, aselegantly demonstrated by Edmonds and Gallistel (1974), whilevertical shifts provide information on motor/performance capacity.Furthermore, this method offers quantitative scaling of drug-inducedchanges in reward (see, Campbell et al., 1985) that is useful whencomparing the effects of different drugs. In other words, the rate-frequency method appears to have reward selectivity that is requiredin psychopharmacological research (Liebman, 1983; Markou and Koob,1992; Miliaressis et al., 1986).
Drug testing began for each animal when the function relatingbar-pressing rate to pulse frequency (the rate-frequency function)was stable for at least three consecutive days. The criterion forstability was met when the frequency thresholds did not vary bymore than 0.1 log units.
2.3. Drugs and drug treatment
Lithium chloride (LiCl) (Sigma-Aldrich, St. Louis, MO, U.S.A.), valproicacid (VPA) (Sigma-Aldrich, St. Louis, MO, U.S.A.) and d-amphetaminesulfate salt (AMPH) (Sigma-Aldrich, St. Louis, MO, U.S.A.) weredissolved in 0.9% NaCl and injected intraperitoneally (i.p.) at a volumeof 1 mg/kg of body weight. The doses of d-amphetamine represent theweight of its salt form. The doses of the mood stabilizers tested arewithin the range of doses regularly used in a plethora of otherbehavioral and functional studies.
In the first study, three groups of animals were used to evaluatethe effects of the acute administration of LiCl, VPA, LiCl and VPA orvehicle on brain stimulation reward. Each drug or vehicle self-stimulation test consisted of a pre-drug and two post-drug rate-frequency function determinations (for 45 min each). The brainexposure of lithium ions is a relatively slow process, although lithiumbrain levels are directly related to the magnitude and the duration ofits plasma levels (Morrison et al., 1971). On the other hand VPA israpidly absorbed after i.p. administration and shows its maximaldistribution both in the brain and in serum 30 min after i.p.injection, whereas 1 h after administration both brain and serumlevels are markedly declined (Aly and Abdel-Latif, 1980). Thus, one
of the aims of the first study was to evaluate the effects of acuteLiCl, VPA or their combination at two different time points (45 minduration each), considering the pharmacokinetics of these drugs(see also below).
In the second study, four groups of animals were used to evaluatethe effects of the acute administration of LiCl, VPA, LiCl and VPA orvehicle on the reward facilitating effects of d-amphetamine. Theself-stimulation test consisted of a predrug and a postdrug rate-frequency function determination (for 45 min each). Rats werepretreated with mood stabilizers (LiCl, VPA or their combination)followed by d-amphetamine (AMPH) or vehicle.
The order of testing for various doses of each drug treatment wascounterbalanced according to a Latin square design. Furthermore, a3-day period was allowed between injections, as in our previousstudies we have observed that this period is considered sufficient forthe self-stimulation behavior to return to pretreatment levels andnot being affected by a prior administration.
2.4. Behavioral studies
2.4.1. Study 1. Effects of systematically administered LiCl, VPA orcombination of LiCl and VPA on brain stimulation rewardEight rats on each experiment received various doses of LiCl (0, 12.5,25, 50, 100, 200 mg/kg) or VPA (0, 100, 200, 400 mg/kg) or variouscombinations of LiCl (0, 25, 50 mg/kg) and VPA (0, 100, 200 mg/kg) ina randomized order. In the 3rd experiment (combined administrationof LiCl and VPA) each animal received two i.p. injections in aconsecutive manner. After a postinjection interval of 20, 10 and15 min, respectively, for each drug treatment (i.e., LiCl, VPA or theircombination), the rats were placed in the operant chamber and thefirst postdrug session began. The second postdrug session started120 min after drug administration.
2.4.2. Study 2. Effects of LiCl, VPA or LiCl and VPA combination onthe reward-facilitating effect of d-amphetamineEight rats on each experiment were pretreated with various doses ofLiCl (0, 50, 100 mg/kg) or VPA (0, 200, 400 mg/kg) or variouscombinations of LiCl (0, 25, 50 mg/kg) and VPA (0, 100, 200 mg/kg)followed by AMPH (0, 0.5, 1 mg/kg). In the 3rd experiment (combinedadministrationof LiCl andVPA followedbyAMPH) thedoses of 25mg/kgLiCl and 100 mg/kg VPA were selected as non-efficacious doses, whilethe doses of 50 mg/kg LiCl and 200 mg/kg VPA as well as 50 mg/kg LiCland 100 mg/kg VPA increased the ICSS threshold, as we have observedin the first study. The injection of LiCl, VPA or LiCl and VPA (twoseparate i.p. injections in a consecutive manner) was given immedi-ately following the baseline session. The rats were then given aninjection of AMPH20, 10 and 15min, respectively, after the LiCl, VPA orLiCl and VPA administration. After an additional 5-min postinjectioninterval, the ratswere placed in the operant chamber and the postdrugsession began.
2.5. Data analysis and statistics
The analysis was performed on two aspects of data obtained from therate-frequency curve: the intracranial self-stimulation (ICSS) thresholdand themaximum rate of responding or asymptote. The estimates wereprocured using the Gompertz sigmoid model (Coulombe and Miliaressis,1987):
f Xð Þ = ae�eb xi�xð Þ
In this equalization, α represents the asymptote videlicet themaximum rate of responding, whereasXi (X at inflection) represents thethreshold frequency. The ICSS threshold is the pulse number producing36.7% of the asymptotic rate. Parameter b represents an index of theslope whereas e is the base of natural logarithms.
The posttreatment threshold and asymptote values wereexpressed as percentage of pretreatment values. In the first study,

208 M. Mavrikaki et al.
the significance of the drug and time effect was statisticallyevaluated initially using two-way analyses of variance (ANOVA)with repeated measures followed, whenever appropriate, bycorrelated t-test using Bonferroni's adjustment for multiple compar-isons. In the second study, the significance of the drug effect wasstatistically evaluated using two-way ANOVA with repeated mea-sures (treatment 1×treatment 2). When the interaction wassignificant, we considered Bonferroni's inequality approach and theanalysis of simple effects was tested in a
p =The sumof ps for themain plus interaction effects
Number of simple effects
For our data, the six simple effects were tested at p=0.025. Thesignificance of simple effects was evaluated using repeated measuresANOVA followed, whenever appropriate, by correlated t-test usingBonferroni's adjustment for multiple comparisons.
2.6. Histology
Following the completion of the experiment, the animals were givena lethal dose of sodium pentothal. The location of the terminalstimulation site was then marked according to the protocoldescribed by Vlachou et al. (2006). The brains were then removedand stored in a solution of 10% formalin until sectioned and stainedfor verification of the electrode tips. Only the rats in which tracksfrom the electrode were verified to be located in the MFB at thelevel of lateral hypothalamus were included in this study.
3. Results
3.1. Study 1
3.1.1. Experiment 1. Effects of systematically administeredLiCl on brain stimulation rewardThe changes in ICSS threshold and asymptotic rate of respondingafter systemic injection of LiCl are presented in Fig. 1. Two-wayANOVA with repeated measures demonstrated a significant drugeffect [F(5,35)=20.320, pb0.001], but neither significant timepostinjection effect [F(1,7)=0.881, pN0.05] nor interaction be-tween drug and time [F(5,35)=0.973, pN0.05], on the ICSSthreshold. Furthermore, paired sample t-tests using Bonferroni'sadjustment formultiple comparisons revealed that LiCl significantlyincreased self-stimulation thresholds at 100 mg/kg and 200 mg/kg(pb0.05). Two-way ANOVA with repeated measures indicated asignificant drug effect [F(5,35)=24.452, pb0.001], but neithersignificant time postinjection effect [F(1,7)=1.113, pN0.05] norinteraction between drug and time [F(5,35)=0.349, pN0.05], onthe asymptotic rate of responding. Paired sample t-tests showedthat this effect on themaximal response ratewas significant for thedose of 100 mg/kg and 200 mg/kg (pb0.05).
3.1.2. Experiment 2. Effects of systematically administeredVPA on brain stimulation rewardThe changes in ICSS threshold and asymptotic rate of respondingafter systemic injection of VPA are presented in Fig. 1. Two-wayANOVA with repeated measures demonstrated a significant drugeffect [F(3,21)=14.046, pb0.001], but no significant time post-injection effect [F(1,7)=3.004, pN0.05] or interaction betweendrug and time [F(5,21)=0.809, pN0.05], on the ICSS threshold.Furthermore, paired sample t-tests revealed that VPA significantlyincreased self-stimulation thresholds only at the highest dosetested, 400 mg/kg (pb0.05). Two-way ANOVA with repeatedmeasures indicated no significant drug effect [F(3,21)=0.369,
pN0.05] or time postinjection effect [F(1,7)=0.019, pN0.05] orinteraction between drug and time [F(3,21)=0.015, pN0.05] on theasymptotic rate of responding.
3.1.3. Experiment 3. Effects of systematically administeredcombination of LiCl and VPA on brain stimulation rewardThe changes in ICSS threshold and asymptotic rate of respondingafter systemic, combined injection of LiCl and VPA are presented inFig. 1. Two-way ANOVA with repeated measures demonstrated asignificant drug effect [F(4,28)=7.273, pb0.001], but no significanttime postinjection effect [F(1,7)=11.912, pN0.05] or interactionbetween drug and time [F(4,28)=1.698, pN0.05], on the ICSSthreshold. Paired sample t-tests revealed that only the combinationof 50mg/kgLiCl and200mg/kgVPA (pb0.05) significantly increasedself-stimulation thresholds. Two-way ANOVA with repeated mea-sures indicated no significant dose effect [F(4,28)=11.912, pN0.05]on the asymptotic rate of responding or time effect [F(1,28)=2.720,pN0.05] or interaction between drug and time [F(4,28)=2.706,pN0.05].
3.2. Study 2
3.2.1. Experiment 1. Effects of LiCl on the reward-facilitating effect of d-amphetamineFig. 2 presents the changes in self-stimulation threshold andasymptotic rate of responding after systemic injection of LiCl orits vehicle and d-amphetamine or its vehicle. Two-way ANOVAwith repeated measures showed a statistical significant inter-action of LiCl and AMPH [F(4,28)=3.255, pb0.05] on the ICSSthreshold. Repeated measures on dose 0 of LiCl showed a sta-tistical significant effect of AMPH [F(2,14)=93.228, pb0.001].Furthermore, paired samples t-test using Bonferroni's adjust-ment for multiple comparisons showed that d-amphetamine(0.5, 1 mg/kg, i.p.) produced a statistical significant decrease inself-stimulation threshold (pb0.001). Repeated measures onthe simple effect of 0.5 mg/kg of AMPH demonstrated astatistical significant effect of LiCl [F(2,14)=20.098, pb0.001].Paired-samples t-test showed that acute administration ofLiCl (100 mg/kg, i.p.) blocked the reward-facilitating effect ofd-amphetamine (0.5 mg/kg, i.p.) on ICSS (pb0.05). On thecontrary, LiCl (50 mg/kg, i.p.) did not block this effect (pN0.05)Repeated measures on the simple effect of 1 mg/kg AMPH showedthat LiCl had no effect [F(2,14)=4.420, pN0.025] on the reward-facilitating effect of the highest dose of d-amphetamine (1mg/kg,i.p.). Two-way ANOVA with repeated measures showed a statis-tical significant interaction of LiCl and AMPH [F(4,27) = 3.860,pb0.05] on the asymptotic rate of responding. In contrast,repeated measures on the dose 0 of LiCl showed that AMPH hadno effect on asymptote [F(2,14)=2.591, pN0.025].
3.2.2. Experiment2.EffectsofVPAon the reward-facilitatingeffect of d-amphetamineFig. 2 presents the changes in self-stimulation threshold andasymptotic rate of responding after systemic injection of VPA or itsvehicle and d-amphetamine or its vehicle. Repeated measuresANOVA showeda statistical significant interactionof VPAandAMPH[F(4,28)=7.276, pb0.001] on the ICSS threshold. Repeatedmeasures on dose 0 of VPA demonstrated a statistical significanteffect of AMPH [F(2,14)=91.039, pb0.001]. Furthermore, pairedsamples t-test using Bonferroni's adjustment for multiple compar-isons showed that d-amphetamine (0.5, 1 mg/kg, i.p.) produced a
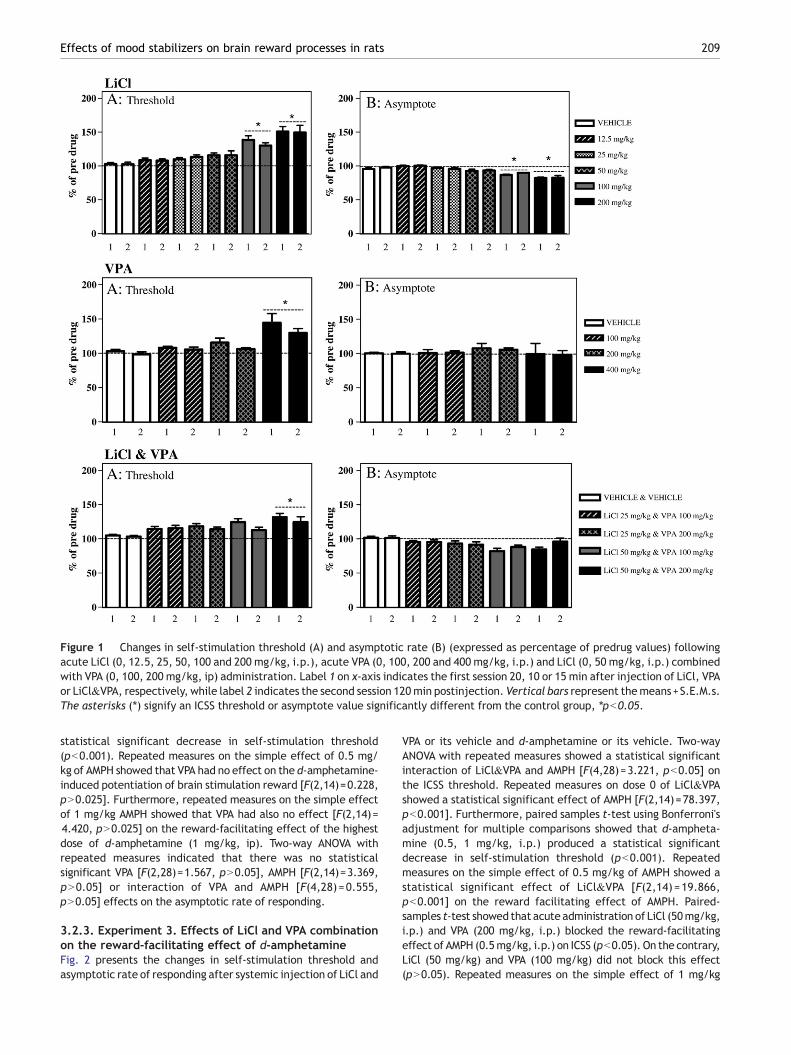
Figure 1 Changes in self-stimulation threshold (A) and asymptotic rate (B) (expressed as percentage of predrug values) followingacute LiCl (0, 12.5, 25, 50, 100 and 200 mg/kg, i.p.), acute VPA (0, 100, 200 and 400 mg/kg, i.p.) and LiCl (0, 50 mg/kg, i.p.) combinedwith VPA (0, 100, 200 mg/kg, ip) administration. Label 1 on x-axis indicates the first session 20, 10 or 15 min after injection of LiCl, VPAor LiCl&VPA, respectively, while label 2 indicates the second session 120min postinjection. Vertical bars represent themeans+S.E.M.s.The asterisks (⁎) signify an ICSS threshold or asymptote value significantly different from the control group, ⁎pb0.05.
209Effects of mood stabilizers on brain reward processes in rats
statistical significant decrease in self-stimulation threshold(pb0.001). Repeated measures on the simple effect of 0.5 mg/kg of AMPH showed that VPA had no effect on the d-amphetamine-induced potentiation of brain stimulation reward [F(2,14)=0.228,pN0.025]. Furthermore, repeated measures on the simple effectof 1 mg/kg AMPH showed that VPA had also no effect [F(2,14)=4.420, pN0.025] on the reward-facilitating effect of the highestdose of d-amphetamine (1 mg/kg, ip). Two-way ANOVA withrepeated measures indicated that there was no statisticalsignificant VPA [F(2,28)=1.567, pN0.05], AMPH [F(2,14)=3.369,pN0.05] or interaction of VPA and AMPH [F(4,28)=0.555,pN0.05] effects on the asymptotic rate of responding.
3.2.3. Experiment 3. Effects of LiCl and VPA combinationon the reward-facilitating effect of d-amphetamineFig. 2 presents the changes in self-stimulation threshold andasymptotic rate of responding after systemic injection of LiCl and
VPA or its vehicle and d-amphetamine or its vehicle. Two-wayANOVA with repeated measures showed a statistical significantinteraction of LiCl&VPA and AMPH [F(4,28)=3.221, pb0.05] onthe ICSS threshold. Repeated measures on dose 0 of LiCl&VPAshowed a statistical significant effect of AMPH [F(2,14)=78.397,pb0.001]. Furthermore, paired samples t-test using Bonferroni'sadjustment for multiple comparisons showed that d-ampheta-mine (0.5, 1 mg/kg, i.p.) produced a statistical significantdecrease in self-stimulation threshold (pb0.001). Repeatedmeasures on the simple effect of 0.5 mg/kg of AMPH showed astatistical significant effect of LiCl&VPA [F(2,14)=19.866,pb0.001] on the reward facilitating effect of AMPH. Paired-samples t-test showed that acute administration of LiCl (50mg/kg,i.p.) and VPA (200 mg/kg, i.p.) blocked the reward-facilitatingeffect of AMPH (0.5mg/kg, i.p.) on ICSS (pb0.05). On the contrary,LiCl (50 mg/kg) and VPA (100 mg/kg) did not block this effect(pN0.05). Repeated measures on the simple effect of 1 mg/kg
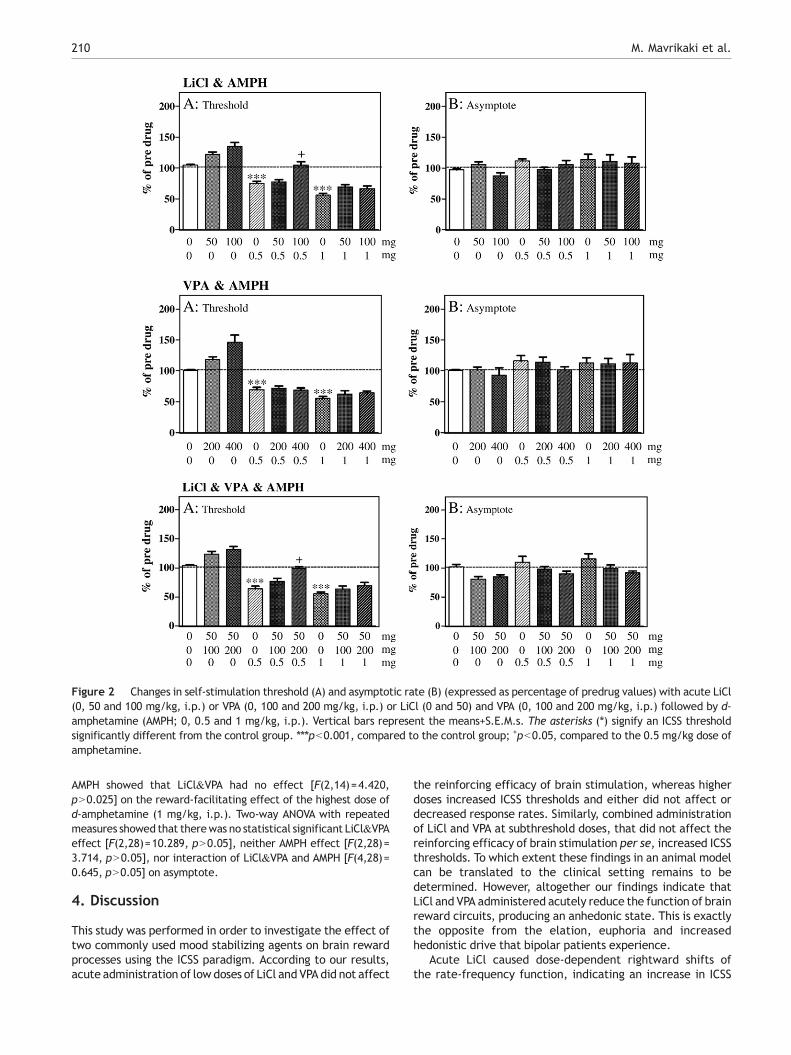
Figure 2 Changes in self-stimulation threshold (A) and asymptotic rate (B) (expressed as percentage of predrug values) with acute LiCl(0, 50 and 100 mg/kg, i.p.) or VPA (0, 100 and 200 mg/kg, i.p.) or LiCl (0 and 50) and VPA (0, 100 and 200 mg/kg, i.p.) followed by d-amphetamine (AMPH; 0, 0.5 and 1 mg/kg, i.p.). Vertical bars represent the means+S.E.M.s. The asterisks (⁎) signify an ICSS thresholdsignificantly different from the control group. ⁎⁎⁎pb0.001, compared to the control group; +pb0.05, compared to the 0.5 mg/kg dose ofamphetamine.
210 M. Mavrikaki et al.
AMPH showed that LiCl&VPA had no effect [F(2,14)=4.420,pN0.025] on the reward-facilitating effect of the highest dose ofd-amphetamine (1 mg/kg, i.p.). Two-way ANOVA with repeatedmeasures showed that therewas no statistical significant LiCl&VPAeffect [F(2,28)=10.289, pN0.05], neither AMPH effect [F(2,28)=3.714, pN0.05], nor interaction of LiCl&VPA and AMPH [F(4,28)=0.645, pN0.05] on asymptote.
4. Discussion
This study was performed in order to investigate the effect oftwo commonly used mood stabilizing agents on brain rewardprocesses using the ICSS paradigm. According to our results,acute administration of low doses of LiCl and VPA did not affect
the reinforcing efficacy of brain stimulation, whereas higherdoses increased ICSS thresholds and either did not affect ordecreased response rates. Similarly, combined administrationof LiCl and VPA at subthreshold doses, that did not affect thereinforcing efficacy of brain stimulation per se, increased ICSSthresholds. To which extent these findings in an animal modelcan be translated to the clinical setting remains to bedetermined. However, altogether our findings indicate thatLiCl and VPA administered acutely reduce the function of brainreward circuits, producing an anhedonic state. This is exactlythe opposite from the elation, euphoria and increasedhedonistic drive that bipolar patients experience.
Acute LiCl caused dose-dependent rightward shifts ofthe rate-frequency function, indicating an increase in ICSS
211Effects of mood stabilizers on brain reward processes in rats
threshold (see Fig. 3). The effects of LiCl on ICSS thresholdsobserved in the present study are in agreement with previousreports (Cassens and Mills, 1973; Tomasiewicz et al., 2006).We report similar effects with acute VPA, which increasedICSS thresholds at the highest dose tested (400 mg/kg).However, in the study by Tomasiewicz et al. (2006) VPA did notsignificantly affect ICSS thresholds at the dose of 300 mg/kg,although there was a clear tendency for increased ICSSthresholds the first 15 min following drug injection. In ourstudy, ICSS thresholds were elevated both 10 min after druginjection and 2 h later. Thus, the observed effects for LiCl and
Figure 3 Rate-frequency functions (rate of lever pressing as afunction of stimulation frequency) taken from representativeanimals for each drug treatment. Each plot represents data froma single animal under predrug and drug conditions. Ratefrequency functions were obtained by logarithmically decreas-ing the frequency of the stimulation pulses from a value thatsustained maximal lever pressing to one that failed to sustainlever pressing.
VPA on ICSS threshold were relatively long-lasting. In clinicalsettings, lithium and valproic acid are sometimes combinedand appear to produce superior therapeutic effects to bipolarpatients (Reischies et al., 2002; Sharma et al., 1993; Solomonet al., 1997). Interestingly, in our study, combined adminis-tration of LiCl and VPA at subthreshold doses that did notaffect the reinforcing efficacy of brain stimulation per se,increased ICSS thresholds. This is the first time that moodstabilizers with different mechanism of action have beenadministered in combination and their effects on brain rewardfunction have been investigated. Our results are consistentwith the view that there is a common brain system underlyingthe effects of psychotropic drugs on brain stimulation reward(Wise, 1996). The effect of LiCl and VPA on ICSS thresholds isopposite to that observed after administration of variousmood elevating agents, such as amphetamine (Gallistel andKarras, 1984). The common inhibitory action of these moodstabilizing agents on brain reward system may also explaintheir similar clinical use in bipolar patients. Indeed, themanicphase of bipolar disorder has been associated with symptomsof elation and increased hedonistic drive that may be relatedwith decreased ICSS thresholds. In this case, the effect of LiCland VPA on ICSS thresholds could be interpreted as antimanicwith regard to these symptoms.
According to our findings, only LiCl decreased themaximal rates of responding for self-stimulation. This effectwas significant 20 min and 2 h after the 100 and 200 mg/kginjection of the drug. In contrast, VPA did not affect themaximal rate of responding. The combined administration ofLiCl and VPA at subthreshold doses decreased also themaximal rates of responding, but only for the first rate-frequency determination following their administration.However, with the presently used self-stimulation protocol,changes in threshold current may discriminate betweenreward and performance, as has been detailed in themethodology section. In other words, the self-stimulationthreshold procedure applied in the present study alloweddetermining threshold and response rate separately andconcurrently in the same self-stimulation session. It is worthnoting, that a decrease in performance is not alwaysassociated with an increase in threshold frequency, andmore importantly attenuated performance has beenobserved in association with a decrease in thresholdfrequency (Panagis and Spyraki, 1996). Accordingly, in thepresent study, the highest dose of VPA increased ICSSthresholds, without affecting maximal rates of responding.
As is the case with various other mood elevating agents,systemic d-amphetamine decreased lateral hypothalamicstimulation reward thresholds and caused parallel leftwardshifts in the rate frequency functions (see Fig. 3). In otherwords, d-amphetamine reduced the amount of stimulationnecessary to sustain responding at a given criterion level(Miliaressis et al., 1986), increasing the rewarding efficacy ofbrain stimulation. This effect of d-amphetamine is consistentwith other studies that have examined its actions on self-stimulation elicited from MFB or other nuclei (Antoniou et al.,2004; Bespalov et al., 1999; Bossert and Franklin, 2003; Carret al., 2002; Depoortere et al., 1999; Frank et al., 1995;Gallistel and Freyd, 1987; Lin et al., 2002; Van Ree and Otte,1980). Interestingly,wenoticed somedifferences betweenLiCland VPA in theway they affect the reward-facilitating effect ofd-amphetamine. Specifically, LiCl, but not VPA, attenuated the

212 M. Mavrikaki et al.
reward-facilitating effects of d-amphetamine in the ICSS.However, combined administration of LiCl and VPA atsubthreshold doses also attenuated the reward-facilitatingeffects of d-amphetamine. Apparently, VPA potentiated theeffect of lithium on this behavioral paradigm.
d-Amphetamine increases the release of dopamine fromneuronal terminals, resulting in increased concentration ofdopamine in the synapse. It is conceivable, therefore, that themechanism by which LiCl attenuated the reward-facilitatingeffects of d-amphetamine may be related to its effect ondopaminergic neurotransmission. Certainly, evidence is nowaccumulating which implies that LiCl, but not VPA, decreasesdopaminergic neurotransmission in the nucleus accumbens(Ichikawa et al., 2005b).We speculate that this neurochemicalalteration may counteract the reward facilitating effect ofamphetamine. Interestingly, VPA which in our study did notattenuate the reward-facilitating effects of d-amphetamine inthe ICSS also did not decrease the dopamine release in thenucleus accumbens, but rather preferentially increaseddopamine release in the prefrontal cortex (Ichikawa andMeltzer, 1999; Ichikawa et al., 2005a). Acute lithium admin-istration can thus decrease the release of dopamine in a brainarea related to the euphoric effects of d-amphetamine.Furthermore, since facilitation of DA release in the NACinduced by d-amphetamine is thought toworsen the symptomsof bipolar patients, it is tempting to speculate that lithiumproduced its effects by normalizing dopaminergic neurotrans-mission in this brain region. Additionally, there is also someevidence that lithium can inhibit other amphetamine-inducedbehaviors in experimental animals (Berggren et al., 1978;Borison et al., 1978; Cox et al., 1971; Gould et al., 2001).
The present results may be relevant, in part, to theclinical evidence that lithium is a more efficacious antimanicagent than valporic acid and other anticonvulsant drugs(Burgess et al., 2001; Frost and Messiha, 1983; Geddes et al.,2004; Swann et al., 1997). Thus, manic patients withdepressive features (i.e. patients with mixed, dysphoric ordepressive mania) respond better to valproic acid, than dopatients with pure manic episodes. On the other hand, acutelithium has been reported to be superior in patients withclassic (euphoric) mania. Furthermore, lithium has beensuggested to be more effective than valproic acid in reducingmanic relapses, whereas valproic acid is more effective thanlithium in delaying time to depressive relapse (Bowden,1995; Burgess et al., 2001; Geddes et al., 2004; Gyulai et al.,2003; Swann et al., 1997). All of these observationscorroborate our findings and are consistent with otherexperimental data in humans indicating that lithium treat-ment attenuates amphetamine-induced euphoric mood inman (Angrist and Gershon, 1979).
In summary, according to the findings of the present studylow doses of acute LiCl and VPA did not affect the reinforcingefficacy of brain stimulation, whereas higher doses increasedICSS thresholds. Similarly, combined administration of sub-threshold doses of LiCl and VPA increased ICSS thresholds.However, only LiCl, but not VPA, attenuated the reward-facilitating effects of d-amphetamine in the ICSS. Interest-ingly, combined administration of LiCl and VPA at subthres-hold doses also attenuated the reward-facilitating effects ofd-amphetamine. We suggest that the ICSS model combinedwith amphetamine administration will be useful to explore inrats the elation and increased hedonistic drive observed in
bipolar patients and ultimately help to identify novel drugtreatments for bipolar disorder.
Role of the funding source
This work was performed in the Department of Psychology andsupported by the Research Committee (KA 2761) and the Depart-ment of Psychology of the University of Crete. The University ofCrete had no further role in study design; in the collection, analysisand interpretation of data; in the writing of the report; and in thedecision to submit the paper for publication.
Contributors
MM performed the experiments and analyzed the data; GGN and GPdesigned the study, managed the literature searches and wrote thepaper. All authors contributed to and have approved the finalmanuscript.
Conflict of interest
All authors declare that they have no conflict of interest.
Acknowledgments
This study was supported by a grant from the Research Committee(KA 2761) and the Department of Psychology of the University ofCrete.
References
Abler, B., Greenhouse, I., Ongur, D., Walter, H., Heckers, S., 2008.Abnormal reward system activation in mania. Neuropsychophar-macology 33, 2217–2227.
Altamura, A.C., 2007. Bipolar spectrum and drug addiction. J. Affect.Disord. 99, 285.
Aly, M.I., Abdel-Latif, A.A., 1980. Studies on distribution andmetabolism of valproate in rat brain, liver, and kidney. Neurochem.Res. 5, 1231–1242.
Anand, A., Verhoeff, P., Seneca, N., Zoghbi, S.S., Seibyl, J.P., Charney,D.S., Innis, R.B., 2000. Brain SPECT imaging of amphetamine-induced dopamine release in euthymic bipolar disorder patients.Am. J. Psychiatry 157, 1108–1114.
Angrist, B., Gershon, S., 1979. Variable attenuation of amphetamineeffects by lithium. Am. J. Psychiatry 136, 806–810.
Angrist, B., Corwin, J., Bartlik, B., Cooper, T., 1987. Early pharmaco-kinetics and clinical effects of oral D-amphetamine in normalsubjects. Biol. Psychiatry 22, 1357–1368.
Antoniou, K., Papathanasiou, G., Panagis, G., Nomikos, G.G.,Hyphantis, T., Papadopoulou-Daifoti, Z., 2004. Individual responsesto novelty predict qualitative differences in d-amphetamine-induced open field but not reward-related behaviors in rats.Neuroscience 123, 613–623.
Baldessarini, R.J., 2002. Treatment research in bipolar disorder:issues and recommendations. CNS. Drugs. 16, 721–729.
Bell, E.C., Willson, M.C., Wilman, A.H., Dave, S., Asghar, S.J.,Silverstone, P.H., 2005a. Lithium and valproate attenuatedextroamphetamine-induced changes in brain activation. Hum.Psychopharmacol. 20, 87–96.
Bell, E.C., Willson, M.C., Wilman, A.H., Dave, S., Silverstone, P.H.,2005b. Differential effects of chronic lithium and valproate onbrain activation in healthy volunteers. Hum. Psychopharmacol.20, 415–424.
Belmaker, R.H., 2004. Bipolar disorder. N. Engl. J. Med. 351, 476–486.

213Effects of mood stabilizers on brain reward processes in rats
Berggren, U., Tallstedt, L., Ahlenius, S., Engel, J., 1978. The effectof lithium on amphetamine-induced locomotor stimulation.Psychopharmacology (Berl) 59, 41–45.
Bespalov, A., Lebedev, A., Panchenko, G., Zvartau, E., 1999. Effectsof abused drugs on thresholds and breaking points of intracranialself-stimulation in rats. Eur. Neuropsychopharmacol. 9, 377–383.
Borison, R.L., Sabelli, H.C., Maple, P.J., Havdala, H.S., Diamond, B.I.,1978. Lithium prevention of amphetamine-induced ‘manic’ excite-ment and of reserpine-induced ‘depression’ in mice: possible role of2-phenylethylamine. Psychopharmacology (Berl) 59, 259–262.
Bossert, J.M., Franklin, K.B., 2003. Reinforcing versus anticonvul-sant drugs: effects on intracranial self-stimulation rate-fre-quency M50 indices. Behav. Brain Res. 144, 243–247.
Bowden, C.L., 1995. Predictors of response to divalproex andlithium. J. Clin. Psychiatry 56 (Suppl 3), 25–30.
Brauer, L.H., deWit, H., 1996. Subjective responses to d-amphetaminealone and after pimozide pretreatment in normal, healthyvolunteers. Biol. Psychiatry 39, 26–32.
Burgess, S., Geddes, J., Hawton, K., Townsend, E., Jamison, K.,Goodwin, G., 2001. Lithium for maintenance treatment of mooddisorders. Cochrane Database Syst. Rev. CD003013.
Campbell, K.A., Evans, G., Gallistel, C.R., 1985. Amicrocomputer-basedmethod for physiologically interpretable measurement of therewarding efficacy of brain stimulation. Physiol. Behav. 35, 395–403.
Carlezon Jr., W.A., Chartoff, E.H., 2007. Intracranial self-stimula-tion (ICSS) in rodents to study the neurobiology of motivation.Nat. Protoc. 2, 2987–2995.
Carr, K.D., Yamamoto, N., Omura, M., Cabeza de Vaca, S., Krahne, L.,2002. Effects of theD(3) dopamine receptor antagonist, U99194A, onbrain stimulation and d-amphetamine reward,motor activity, and c-fos expression in ad libitum fed and food-restricted rats. Psycho-pharmacology (Berl) 163, 76–84.
Cassens, G.P., Mills, A.W., 1973. Lithium and amphetamine: oppositeeffects on threshold of intracranial reinforcement. Psychopharma-cologia 30, 283–290.
Cerullo, M.A., Strakowski, S.M., 2007. The prevalence and signifi-cance of substance use disorders in bipolar type I and II disorder.Subst. Abuse Treat. Prev. Policy 2, 29.
Coulombe, D., Miliaressis, E., 1987. Fitting intracranial self-stimula-tion data with growth models. Behav. Neurosci. 101, 209–214.
Cox, C., Harrison-Read, P.E., Steinberg, H., Tomkiewicz, M., 1971.Lithium attenuates drug-induced hyperactivity in rats. Nature232, 336–338.
Depoortere, R., Perrault, G., Sanger, D.J., 1999. Intracranial self-stimulation under a progressive-ratio schedule in rats: effects ofstrength of stimulation, d-amphetamine, 7-OH-DPAT and halo-peridol. Psychopharmacology (Berl) 142, 221–229.
Edmonds, D.E., Gallistel, C.R., 1974. Parametric analysis of brainstimulation reward in the rat: III. effect of performance variableson the reward summation function. J. Comp. Physiol. Psychol.87, 876–883.
Einat, H., Manji, H.K., Belmaker, R.H., 2003. New approaches tomodeling bipolar disorder. Psychopharmacol. Bull. 37, 47–63.
Einat, H., 2007. Establishment of a battery of simple models forfacets of bipolar disorder: a practical approach to achieveincreased validity, better screening and possible insights intoendophenotypes of disease. Behav. Genet. 37, 244–255.
Flemenbaum, A., 1974. Does lithium block the effects ofamphetamine? A report of three cases. Am. J. Psychiatry131, 820–821.
Fountoulakis, K.N., Vieta, E., Siamouli, M., Valenti, M., Magiria, S.,Oral, T., Fresno, D., Giannakopoulos, P., Kaprinis, G.S., 2007.Treatment of bipolar disorder: a complex treatment for a multi-faceted disorder. Ann. Gen. Psychiatry 6, 27.
Frank, R.A., Tsibulsky, V., Grocki, S., Dashevsky, B., Kehne, J.H.,Schmidt, C.J., Sorensen, S.M., 1995. Mixed D2/5-HT2A antagon-ism of amphetamine-induced facilitation of brain stimulationreward. Pharmacol. Biochem. Behav. 52, 799–804.
Frost, R.E., Messiha, F.S., 1983. Clinical uses of lithium salts. BrainRes. Bull. 11, 219–231.
Gallistel, C.R., Karras, D., 1984. Pimozide and amphetamine haveopposing effects on the reward summation function. Pharmacol.Biochem. Behav. 20, 73–77.
Gallistel, C.R., Freyd, G., 1987. Quantitative determination of theeffects of catecholaminergic agonists and antagonists on therewarding efficacy of brain stimulation. Pharmacol. Biochem.Behav. 26, 731–741.
Geddes, J.R., Burgess, S., Hawton, K., Jamison, K., Goodwin, G.M.,2004. Long-term lithium therapy for bipolar disorder: systematicreview and meta-analysis of randomized controlled trials. Am. J.Psychiatry. 161, 217–222.
Goodwin, F.K., Jamison, K.R., 2007. Manic-depressive Illness:Bipolar Disorders and Recurrent Depression, Vol. Oxford Uni-versity Press, Oxford.
Gould, T.J., Keith, R.A., Bhat, R.V., 2001. Differential sensitivity tolithium's reversal of amphetamine-induced open-field activity intwo inbred strains of mice. Behav. Brain Res. 118, 95–105.
Gyulai, L., Bowden, C.L., McElroy, S.L., Calabrese, J.R., Petty, F.,Swann, A.C., Chou, J.C.,Wassef, A., Risch, C.S., Hirschfeld, R.M.,Nemeroff, C.B., Keck Jr., P.E., Evans, D.L., Wozniak, P.J., 2003.Maintenance efficacy of divalproex in the prevention of bipolardepression. Neuropsychopharmacology 28, 1374–1382.
Ichikawa, J., Meltzer, H.Y., 1999. Valproate and carbamazepineincrease prefrontal dopamine release by 5-HT1A receptoractivation. Eur. J. Pharmacol. 380, R1–3.
Ichikawa, J., Chung, Y.C., Dai, J., Meltzer, H.Y., 2005a. Valproic acidpotentiates both typical and atypical antipsychotic-inducedprefrontal cortical dopamine release. Brain. Res. 1052, 56–62.
Ichikawa, J., Dai, J., Meltzer, H.Y., 2005b. Lithium differs fromanticonvulsant mood stabilizers in prefrontal cortical andaccumbal dopamine release: role of 5-HT(1A) receptor agonism.Brain Res. 1049, 182–190.
Jacobs, D., Silverstone, T., 1986. Dextroamphetamine-induced arousalin human subjects as amodel formania. Psychol.Med. 16, 323–329.
Kahn, D.A., Sachs, G.S., Printz, D.J., Carpenter, D., Docherty, J.P.,Ross, R., 2000. Medication treatment of bipolar disorder 2000:a summary of the expert consensus guidelines. J. Psychiatr. Pract.6, 197–211.
Kasper, S., 2003. Issues in the treatment of bipolar disorder. Eur.Neuropsychopharmacol. 13 (Suppl 2), S37–42.
Kornetsky, C., 1985. Brain-stimulation reward: a model for theneuronal bases for drug-induced euphoria. NIDA. Res. Monogr. 62,30–50.
Kornetsky, C., Esposito, R.U., 1979. Euphorigenic drugs: effects onthe reward pathways of the brain. Fed. Proc. 38, 2473–2476.
Liebman, J.M., 1983. Discriminating between reward and perfor-mance: a critical review of intracranial self-stimulation metho-dology. Neurosci. Biobehav. Rev. 7, 45–72.
Lin, D., Bruijnzeel, A.W., Schmidt, P., Markou, A., 2002. Exposure tochronic mild stress alters thresholds for lateral hypothalamicstimulation reward and subsequent responsiveness to ampheta-mine. Neuroscience 114, 925–933.
Maremmani, I., Pacini, M., Perugi, G., Deltito, J., Akiskal, H., 2008.Cocaine abuse and the bipolar spectrum in 1090 heroin addicts:clinical observations and a proposed pathophysiologic model.J. Affect. Disord. 106, 55–61.
Markou, A., Koob,G.F., 1991. Postcocaine anhedonia. An animalmodelof cocaine withdrawal. Neuropsychopharmacology 4, 17–26.
Markou, A., Koob, G.F., 1992. Construct validity of a self-stimulationthreshold paradigm: effects of reward and performance manip-ulations. Physiol. Behav. 51, 111–119.
McIntyre, R.S., McElroy, S.L., Konarski, J.Z., Soczynska, J.K., Bottas,A., Castel, S., Wilkins, K., Kennedy, S.H., 2007. Substance usedisorders and overweight/obesity in bipolar I disorder: prelimin-ary evidence for competing addictions. J. Clin. Psychiatry 68,1352–1357.

214 M. Mavrikaki et al.
Miliaressis, E., Rompre, P.P., Laviolette, P., Philippe, L., Coulombe, D.,1986. The curve-shift paradigm in self-stimulation. Physiol. Behav.37, 85–91.
Moller, H.J., Nasrallah,H.A., 2003. Treatment of bipolar disorder. J. Clin.Psychiatry 64 (Suppl 6), 9–17 discussion 28.
Morrison Jr., J.M., Pritchard, H.D., Braude, M.C., D'Aguanno, W.,1971. Plasma and brain lithium levels after lithium carbonate andlithium chloride administration by different routes in rats. Proc.Soc. Exp. Biol. Med. 137, 889–892.
Olds, J., Milner, P., 1954. Positive reinforcement produced byelectrical stimulation of septal area and other regions of ratbrain. J. Comp. Physiol. Psychol. 47, 419–427.
Panagis, G., Spyraki, C., 1996. Neuropharmacological evidence forthe role of dopamine in ventral pallidum self-stimulation.Psychopharmacology (Berl) 123, 280–288.
Paterson, N.E., Myers, C., Markou, A., 2000. Effects of repeatedwithdrawal from continuous amphetamine administration onbrain reward function in rats. Psychopharmacology (Berl) 152,440–446.
Paxinos, G., Watson, C., 2007. The rat brain in stereotaxiccoordinates, sixth ed. Elsevier, Amsterdam.
Reischies, F.M., Hartikainen, J., Berghofer, A., 2002. Initial lithiumand valproate combination therapy in acute mania. Neuropsy-chobiology 46 (Suppl 1), 22–27.
Roybal, K., Theobold, D., Graham, A., DiNieri, J.A., Russo, S.J.,Krishnan, V., Chakravarty, S., Peevey, J., Oehrlein, N., Birnbaum,S., Vitaterna, M.H., Orsulak, P., Takahashi, J.S., Nestler, E.J.,Carlezon Jr., W.A., McClung, C.A., 2007. Mania-like behaviorinduced by disruption of CLOCK. Proc. Natl. Acad. Sci. U. S. A.104, 6406–6411.
Sharma, V., Persad, E., Mazmanian, D., Karunaratne, K., 1993.Treatment of rapid cycling bipolar disorder with combinationtherapy of valproate and lithium. Can. J. Psychiatry 38, 137–139.
Silverstone, P.H., Pukhovsky, A., Rotzinger, S., 1998. Lithium doesnot attenuate the effects of D-amphetamine in healthy volun-teers. Psychiatry Res. 79, 219–226.
Slattery, D.A., Markou, A., Cryan, J.F., 2007. Evaluation of rewardprocesses in an animal model of depression. Psychopharmacology(Berl) 190, 555–568.
Solomon, D.A., Ryan, C.E., Keitner, G.I., Miller, I.W., Shea, M.T.,Kazim, A., Keller,M.B., 1997. Apilot studyof lithiumcarbonateplusdivalproex sodium for the continuationandmaintenance treatmentof patients with bipolar I disorder. J. Clin. Psychiatry 58, 95–99.
Strakowski, S.M., Sax, K.W., 1998. Progressive behavioral responseto repeated d-amphetamine challenge: further evidence forsensitization in humans. Biol. Psychiatry 44, 1171–1177.
Swann, A.C., Bowden, C.L., Morris, D., Calabrese, J.R., Petty, F.,Small, J., Dilsaver, S.C., Davis, J.M., 1997. Depression duringmania. Treatment response to lithium or divalproex. Arch. Gen.Psychiatry 54, 37–42.
Tomasiewicz, H.C., Mague, S.D., Cohen, B.M., Carlezon Jr., W.A.,2006. Behavioral effects of short-term administration of lithiumand valproic acid in rats. Brain Res. 1093, 83–94.
Van Kammen, D.P., Murphy, D.L., 1975. Attenuation of the euphoriantand activating effects of d- and l-amphetamine by lithium car-bonate treatment. Psychopharmacologia 44, 215–224.
Van Ree, J.M., Otte, A.P., 1980. Effects of (Des-Tyr1)-gamma-endorphin and alpha-endorphin as compared to haloperidol andamphetamine on nucleus accumbens self-stimulation. Neuro-pharmacology 19, 429–434.
Vlachou, S., Nomikos, G.G., Panagis, G., 2006. Effects of endo-cannabinoid neurotransmission modulators on brain stimulationreward. Psychopharmacology (Berl) 188, 293–305.
Willson, M.C., Bell, E.C., Dave, S., Asghar, S.J., McGrath, B.M.,Silverstone, P.H., 2005. Valproate attenuates dextroampheta-mine-induced subjective changes more than lithium.Eur. Neuropsychopharmacol. 15, 633–639.
Wise, R.A., 1996. Addictive drugs and brain stimulation reward.Annu. Rev. Neurosci. 19, 319–340.
Wise, R.A., Munn, E., 1993. Effects of repeated amphetamineinjections on lateral hypothalamic brain stimulation reward andsubsequent locomotion. Behav. Brain Res. 55, 195–201.
Wise, R.A., Munn, E., 1995. Withdrawal from chronic amphetamineelevates baseline intracranial self-stimulation thresholds. Psy-chopharmacology (Berl) 117, 130–136.
Zacny, J.P., Bodker, B.K., de Wit, H., 1992. Effects of setting on thesubjective and behavioral effects of d-amphetamine in humans.Addict. Behav. 17, 27–33.




















