
Effect of relative humidity and origin of isolates of Neozygites tanajoae (Zygomycetes:...
8
Biological Control 39 (2006) 489–496 www.elsevier.com/locate/ybcon 1049-9644/$ - see front matter © 2006 Elsevier Inc. All rights reserved. doi:10.1016/j.biocontrol.2006.08.003 EVect of relative humidity and origin of isolates of Neozygites tanajoae (Zygomycetes: Entomophthorales) on production of conidia from cassava green mite, Mononychellus tanajoa (Acari: Tetranychidae), cadavers Italo Delalibera Jr. a,b,¤ , Clarice G.B. Demétrio c , Bryan F.J. Manly c , Ann E. Hajek b a Department of Entomology, Plant Pathology and Agricultural Zoology, ESALQ-USP, CP9, Piracicaba-SP 13418-900, Brazil b Department of Entomology, Cornell University, Ithaca, NY 14853-2601, USA c Exact Sciences Department, ESALQ-USP, CP9, Piracicaba-SP 13418-900, Brazil Received 5 April 2006; accepted 7 August 2006 Available online 15 August 2006 Abstract Neozygites tanajoae is a very specialized fungus pathogenic to the cassava green mite (CGM), Mononychellus tanajoa, an important cassava pest introduced to Africa from the Neotropics. Conidial discharge from cadavers of CGM that died from infections with 14 iso- lates of N. tanajoae collected from diverse climates of Brazil was quantiWed to help select potential candidate strains for introduction to Africa. Studies aimed to identify isolates with lower requirements for relative humidity for sporulation and isolates that discharged more conidia during short periods of moisture. At 96 § 0.5% RH, production of conidia was variable and even isolates from the Brazilian semi- arid region, e.g., Petrolina and Itaberaba, produced few conidia. SigniWcant diVerences in the numbers of conidia produced by diverse Brazilian isolates were observed after 6, 9 and 12 h at 100% RH. At 100% RH, production of primary conidia increased considerably from an average of 57 § 4 conidia at 6 h to 509 § 37 conidia at 12 h. The isolate sporulating least (BIN21) discharged only 45.7% of the number of conidia produced by isolate BIN1, one of the isolates producing the most spores. Results from this study demonstrate that diVerences in production of conidia among isolates should be considered when selecting Neozygites isolates for new biological control introductions. © 2006 Elsevier Inc. All rights reserved. Keywords: Entomophthorales; Mononychellus tanajoa; Sporulation; Primary conidia; Capilliconidia; Biological control; Fungal pathogen 1. Introduction The pathogenic entomophthoralean fungus Neozygites tanajoae Delalibera, Humber and Hajek is an important natural enemy of the cassava green mite (CGM), Monony- chellus tanajoa (Bondar), in northeastern Brazil (Delalibera et al., 2000, 1992; Elliot et al., 2000). CGM became one of the most important pests of cassava (Manihot esculenta Crantz) in Africa after its introduction from the Neotrop- ics. Classical biological control programs have been initi- ated to introduce Brazilian isolates of N. tanajoae to Africa. To date, the two Brazilian isolates of N. tanajoae released in Benin diVered in levels of establishment and infection (Hountondji et al., 2002). Comparisons of several N. tana- joae isolates in the laboratory indicated variability in both virulence and mummiWcation, with some Brazilian isolates being highly virulent (Delalibera and Hajek, 2004). This observation suggests that additional releases of one (or more) highly virulent Brazilian isolate(s) to Africa could improve the CGM biological control program. The density of a pathogen population is one of the most important factors for triggering the initiation of an epizo- otic (Tanada and Kaya, 1993); high densities of the host and both high levels of infection and abundant spore * Corresponding author. Fax: +55 19 3429 4338. E-mail address: [email protected] (I. Delalibera).
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Effect of relative humidity and origin of isolates of Neozygites tanajoae (Zygomycetes:...

Biological Control 39 (2006) 489–496
www.elsevier.com/locate/ybcon
EVect of relative humidity and origin of isolates of Neozygites tanajoae (Zygomycetes: Entomophthorales) on production of
conidia from cassava green mite, Mononychellus tanajoa (Acari: Tetranychidae), cadavers
Italo Delalibera Jr. a,b,¤, Clarice G.B. Demétrio c, Bryan F.J. Manly c, Ann E. Hajek b
a Department of Entomology, Plant Pathology and Agricultural Zoology, ESALQ-USP, CP9, Piracicaba-SP 13418-900, Brazilb Department of Entomology, Cornell University, Ithaca, NY 14853-2601, USA
c Exact Sciences Department, ESALQ-USP, CP9, Piracicaba-SP 13418-900, Brazil
Received 5 April 2006; accepted 7 August 2006Available online 15 August 2006
Abstract
Neozygites tanajoae is a very specialized fungus pathogenic to the cassava green mite (CGM), Mononychellus tanajoa, an importantcassava pest introduced to Africa from the Neotropics. Conidial discharge from cadavers of CGM that died from infections with 14 iso-lates of N. tanajoae collected from diverse climates of Brazil was quantiWed to help select potential candidate strains for introduction toAfrica. Studies aimed to identify isolates with lower requirements for relative humidity for sporulation and isolates that discharged moreconidia during short periods of moisture. At 96§ 0.5% RH, production of conidia was variable and even isolates from the Brazilian semi-arid region, e.g., Petrolina and Itaberaba, produced few conidia. SigniWcant diVerences in the numbers of conidia produced by diverseBrazilian isolates were observed after 6, 9 and 12 h at 100% RH. At 100% RH, production of primary conidia increased considerably froman average of 57§ 4 conidia at 6 h to 509§ 37 conidia at 12 h. The isolate sporulating least (BIN21) discharged only 45.7% of the numberof conidia produced by isolate BIN1, one of the isolates producing the most spores. Results from this study demonstrate that diVerencesin production of conidia among isolates should be considered when selecting Neozygites isolates for new biological control introductions.© 2006 Elsevier Inc. All rights reserved.
Keywords: Entomophthorales; Mononychellus tanajoa; Sporulation; Primary conidia; Capilliconidia; Biological control; Fungal pathogen
1. Introduction
The pathogenic entomophthoralean fungus Neozygitestanajoae Delalibera, Humber and Hajek is an importantnatural enemy of the cassava green mite (CGM), Monony-chellus tanajoa (Bondar), in northeastern Brazil (Delaliberaet al., 2000, 1992; Elliot et al., 2000). CGM became one ofthe most important pests of cassava (Manihot esculentaCrantz) in Africa after its introduction from the Neotrop-ics. Classical biological control programs have been initi-
* Corresponding author. Fax: +55 19 3429 4338.E-mail address: [email protected] (I. Delalibera).
1049-9644/$ - see front matter © 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.biocontrol.2006.08.003
ated to introduce Brazilian isolates of N. tanajoae to Africa.To date, the two Brazilian isolates of N. tanajoae released inBenin diVered in levels of establishment and infection(Hountondji et al., 2002). Comparisons of several N. tana-joae isolates in the laboratory indicated variability in bothvirulence and mummiWcation, with some Brazilian isolatesbeing highly virulent (Delalibera and Hajek, 2004). Thisobservation suggests that additional releases of one (ormore) highly virulent Brazilian isolate(s) to Africa couldimprove the CGM biological control program.
The density of a pathogen population is one of the mostimportant factors for triggering the initiation of an epizo-otic (Tanada and Kaya, 1993); high densities of the hostand both high levels of infection and abundant spore

490 I. Delalibera Jr. et al. / Biological Control 39 (2006) 489–496
production on and/or in hosts are integral to high levels ofinfection by a fungal pathogen. Neozygites species producethree types of spores. After CGM die from N. tanajoaeinfections, primary conidia are actively ejected from swol-len, light brown cadavers, referred to as mummies. Neozyg-ites primary conidia germinate to form a secondary type ofconidia, the infective and more persistent capilliconidia.Capilliconidia of entomophthoralean fungi, including thoseof Neozygites fresenii (Nowakowski) Remaudière and Kel-ler and N. tanajoae are able to withstand lower relativehumidities than primary conidia (Oduor et al., 1996a,b;Uziel and Kenneth, 1991). In addition, for persistence, Neo-zygites species produce longer lived resting spores withincadavers.
High relative humidity is widely recognized as a criticalfactor for reproduction by fungi (Boucias and Pendland,1998; Hajek et al., 2001). Most entomophthoralean fungirequire high humidities to produce spores. However, atleast one species, Entomophthora muscae (Cohn) Fresenius,is less vulnerable to humidity and is able to sporulate fromcadavers of house Xies, Musca domestica L., at relativehumidities as low as 20% (Mullens and Rodriguez, 1985).Another entomophthoralean species was reported by New-man and Carner (1975) to sporulate across a broader rangeof relative humidities; Entomophthora gammae (DPandoragammae) (Weiser) sporulated from cadavers of Pseudoplu-sia includens (Walker) larvae at 80–100% RH. N. tanajoaehas been documented causing epizootics in CGM popula-tions in dry regions in Brazil, yet we assume that this fungusrequires some level of moisture to produce conidia thatsubsequently produce infective capilliconidia. Many areasin Africa where CGM is a problem are dry or have a longdry season. We hypothesized that fungal strains from dryareas of Brazil could be better adapted to the long dry sea-son in Africa when moisture availability might be marginal.An isolate that initiates sporulation rapidly and at a lowrelative humidity would be most suitable for use in thesemi-arid regions of Africa where cassava is a major crop.Cassava is grown in a wide range of climates throughoutthe tropical and subtropical areas of Africa. Even an isolatethat requires only a short period of high relative humidityat night to discharge conidia may meet its needs for mois-ture in order to sporulate in drier areas (e.g., Ben’Ze’ev andKenneth, 1981; see Carruthers and Soper, 1987).
To investigate the potential for further introductions ofN. tanajoae to Africa, we studied variability among N. tana-joae isolates in requirements for the amount of moisturenecessary for spore production. We evaluated productionof both primary conidia and capilliconidia by isolates ofN. tanajoae and the closely related Neozygites Xoridana(Weiser and Muma) Remaudière and Keller at diVerenthumidity levels. Dead, infected CGM (D mummies) werecollected in nine states of Brazil from areas with diVerentclimates, from the coastal rain forest to semi-arid regionswith <400 mm of rainfall per year. The mummies werestored at ¡10 °C, and the group of mummies from each col-lection site was assigned an isolate number. Fresh mummies
were produced in the laboratory by exposing healthy CGMto sporulating Weld-collected mummies, and only thesefresh mummies were used in the sporulation studiesreported here. Studies compared sporulation of diVerentmummies from diVerent locations when mummies wereexposed to two levels of high humidity for varying lengthsof time.
2. Materials and methods
2.1. Study organisms
The Wfteen isolates of the mite pathogenic fungusN. tanajoae and one isolate of N. Xoridana used in thisstudy (Table 1) have been characterized previously basedon 18 S rRNA gene sequences, host ranges, nutritionalrequirements for growth in vitro, tolerances to cold (4 °C)and abilities to withstand speciWc cryopreservation tech-niques (Delalibera and Hajek, 2004; Delalibera et al., 2003,2004a,b). Isolate BIN34 from Palmira, Colombia origi-nated from Tetranychus urticae Koch. All other isolateswere from CGM. BIN35 is a native isolate from Cotonou,Benin collected before any release of Brazilian isolates inBenin. All isolates are stored at the University of SãoPaulo, Piracicaba, São Paulo, Brazil. Except for the lastassay during which a Brazilian and an Beninese N. tanajoaeisolate were compared using CGM from Cotonou, Benin,all mites used originated from laboratory colonies fromCruz das Almas, Bahia, Brazil.
2.2. Sporulation studies
2.2.1. Inoculum preparationSporulation of N. tanajoae can be induced from hyphal
bodies produced in vitro but virulence of the resulting capilli-conidia is very low (Delalibera et al., 2003). Very few N. tana-joae strains have been isolated in culture media and thecollection of isolates is maintained mostly as mummiWedmites at ¡10°C. For these reasons all experiments were con-ducted using mummiWed mites. Before use in this study,cadavers were thawed and the fungus was then grown in vivoin the laboratory using the methods for pathogenicity testsdescribed by Delalibera and Hajek (2004). The mummiesresulting from laboratory infections were used in the sporula-tion studies. Mummies of each isolate were placed individu-ally on 1.5-cm diameter cassava leaf discs in Petri dishes at100% RH and 25§2 °C for sporulation over 16h. To infectmites to produce mummies for sporulation studies, 25 smallCGM females were transferred to each leaf disc. Petri disheswere kept at 70 §10% RH, 12:12 L:D at 25§2 °C. For theWrst 3 days, the plates were kept closed (100% RH) duringthe dark phase to induce further conidiogenesis in order tomaximize infection. For the purpose of sporulation assaysdescribed below only newly formed mummiWed cadavers(swollen brown fungus-killed cadavers) less than 12 h-oldand of similar size were used. Because infected mites thatdied and mummiWed were produced at 70§10% RH, no
I. Delalibera Jr. et al. / Biological Control 39 (2006) 489–496 491
sporulation occurred prior to experiments. Each replicateconsisted of mummiWed mites produced using mites from thesame plant and infected on the same day to ensure that fun-gal isolate would be the most important variable.
2.2.2. Experimental systemThe mummiWed mites were placed individually in the cen-
ter of a reticle area of 18mm by 18 mm on a glass microscopeslide inside of a 9 cm-Petri dish. The Petri dishes were placedon top of 7 cm-high perforated plastic tubes inside plasticchambers (9£12£11cm). The chambers were hermeticallysealed with paraWlm and kept at 23.5§1 °C in the dark. Rel-ative humidity levels in each chamber were adjusted using400 ml of sulfuric acid solutions (Stevens, 1916; Solomon,1951) and distilled water was used for 100% RH. Densities ofthe solutions were measured using a densimeter at the begin-ning and end of each replicate and the humidity was checkedusing a hygrometer sensor (Hanna Instruments HI8564)inside a replicate chamber. For each mummy, total primaryconidia and capilliconidia were counted in each grid (4 mm2)using phase contrast microscopy at 100£ magniWcation.
2.3. Experiments
2.3.1. Screen for minimum relative humidity requirementsPreliminary screening tests using fungal isolates BIN4
and BIN8 were carried out to determine the lowest relativehumidity at which N. tanajoae sporulates. Four densities ofsulfuric acid solution were tested; 1.035; 1.040; 1.045 and1.050 g/ml giving relative humidities of 96.4§0.1%;96.3§0.9%; 95.3§0.5% and 95.1§0.2%, respectively. SixmummiWed mites per isolate were placed in each chamberwith three chambers per relative humidity£ time combina-tion. Evaluations were made after 12 and 18 h. Based onthese results, sporulation of all isolates were compared
using the density of 1.042 g/ml sulfuric acid (96§0.5% RH),which was considered as a very low RH permitting minimalsporulation, and at 100% RH as the maximal RH.
2.3.2. N. tanajoae from Northeastern BrazilThe Wrst bioassay was conducted to compare sporulation
of 12 N. tanajoae isolates from Northeastern Brazil. OnemummiWed mite of each isolate was placed on a microscopeslide inside a plastic chamber as described above. For thisbioassay, three chambers were set up for each relative humid-ity level (96% versus 100% RH) for each isolate. One cham-ber at each relative humidity was randomly chosen to bechecked after 6 h. Another chamber at each RH was evalu-ated after 9h, and the remaining chambers were evaluatedafter 12h. This procedure was repeated 15 times with eachreplicate totaling 96 mummies per relative humidity£ timefor each isolate.
2.3.3. Neozygites from selected places in Brazil, Benin and Colombia
After the bioassay with only N. tanajoae from North-eastern Brazil, two Brazilian isolates (BIN1 and BIN8)were selected for subsequent comparison with the Colom-bian N. Xoridana isolate (BIN34) and a Beninese isolate ofN. tanajoae (BIN35). Two additional Brazilian isolateswere added for this assay, one from a warm and dry region,Petrolina (BIN31), and another from a colder and wetregion, Mandiocaba (BIN28). Sixteen and 24 replicateswere conducted at 100% RH and 96§ 0.5% RH, respec-tively, for 6, 9 and 12 h.
2.3.4. Brazilian and Beninese isolates of N. tanajoae challenging Beninese CGM
In this assay conducted in Benin, the isolate BIN8 fromBrazil, selected from the previous assays, was compared
Table 1Origins of isolates of Neozygites tanajoae and N. Xoridana
a BIN, Banco de Isolados de Neozygites, Isolates are stored at the University of São Paulo, Piracicaba, SP, Brazil.b BR, Brazil; CO, Colombia; BE, Benin. States: BA, Bahia; CE, Ceará; AL, Alagoas; PE, Pernambuco; PI, Piaui; PR, Paraná; VA, Valey.
IDa Location (City, State)b Countryb Latitude Longitude Assay
N. tanajoaeBIN1 Piritiba, BA BR 11°41�32�S 40°35�39�W 1, 2BIN4 Ipirá, BA BR 12°09�25� S 39°44�30�W 1BIN8 Cruz das Almas, BA BR 12°40�19�S 39°06�23�W 1, 2, 3BIN10 Tianguá, CE BR 04°06�33�S 40°53�15�W 1BIN11 Rio Real, BA BR 11°37�09�S 37°49�35�W 1BIN13 Porto Real do Colégio, AL BR 10°07�13�S 36°43�56�W 1BIN15 São M. dos Campos, AL BR 9°45�16�S 36°04�03�W 1BIN18 Caruaru, PE BR 8°12�29�S 35°58�06�W 1BIN20 Itapajé, CE BR 3°41�25�S 39°35�54�W 1BIN21 Floriano, PI BR 6°38�28�S 42°52�14�W 1BIN22 Araripina, PE BR 7°30�03�S 40°27�42�W 1BIN25 Itaberaba, BA BR 12°23�48�S 40°15�00�W 1BIN28 Mandiocaba, PR BR 23°05�S 52°27�W 2BIN31 Petrolina, PE BR 9°09�S 40°22�W 2BIN35 Cotonou BE 6°25�N 2°18�E 2, 3
N. XoridanaBIN34 Palmira, VA CO 3°32�N 76°16�W 2
492 I. Delalibera Jr. et al. / Biological Control 39 (2006) 489–496
again with isolate BIN35 from Benin, but this time the iso-lates were produced using a colony of CGM from Cotonou,Benin. As soon as the infected mites became mummiWedthey were placed individually on microscope slides above asponge bed soaked with water in 9-cm Petri dishes (100%RH) at 25.5§3.5 °C for 16 h. Over 5 days, conidial dis-charge by mummiWed mites from both isolates was evalu-ated with numbers of mummies used depending onavailability each day. Since almost all primary conidia hadgerminated to form capilliconidia after 16 h, only capilli-conidia were quantiWed. The total numbers of capilliconidiaproduced by 38 mummiWed mites for BIN8 (the isolatefrom Cruz das Almas, Brazil) and 48 mummiWed mites forBIN35 (the isolate from Cotonou, Benin) were counteddirectly by microscope.
2.3.5. Data analysisThe experiment followed a completely randomized
design. All analyses used data for primary conidia and cap-illiconidia combined, except for the assay with a Brazilianand a Beninese N. tanajoae on Beninese CGM because pri-mary conidia were not counted. The eVect of isolate andtime on the number of capilliconidia and primary conidiareleased per mummiWed mite by each isolate at each relativehumidity was tested using a Poisson regression model(McCullagh and Nelder, 1989) which is a common methodfor the analysis of count data. With the standard Poissonregression model the variance of an observation Y is equalto the expected value �. However, it often happens that thecounts display more variation than this implies, a phenom-enon known as overdispersion (Hinde and Demétrio,1998a,b). This occurred for the data considered here, so thiswas compensated by the estimation of a heterogeneity fac-tor. The calculations were carried out in GenStat (LawesAgricultural Trust, 2005), which estimates the heterogene-ity factor as a standard option. The Poisson regressionmodel that was used assumes that the expected count isgiven by �DR exp(�0 + Timei + Isolatej), where R is thenumber of replicates of the experimental conditions, Timeirepresents an eVect for the observation time i (1, 2 or 3), andIsolatej represents an eVect by isolate (1–12 for the assaywith only N. tanajoae from Northeastern Brazil, and 1 to 6for the assay with Neozygites from selected places in Brazil,Benin and Colombia).
3. Results
3.1. Screen for minimum relative humidity requirements
Preliminary screening tests demonstrated that sporula-tion did not occur when the density of 1.05 g/ml of sulfuricacid solution was used (95.1§0.2% RH) and no signiWcantdiVerences were observed in production of conidia whenthe other 3 solutions were used (96.4§ 0.1%; 96.3§0.9%;95.3§ 0.5%), or between 12 and 18 h of sporulation for thefour relative humidity levels tested (data not presented).These results were used to decide that the minimum RH we
would use for comparisons of the sporulation of all isolateswould be 96§ 0.5% RH.
3.2. N. tanajoae from Northeastern Brazil
At the minimum RH tested (96§ 0.5% RH), the numberof primary conidia was highly variable among replicates ofthe same isolate within the same time and among diVerenttime intervals. For example, the average number of primaryconidia per mummiWed mite per isolate ranged from 1§1to 117§80 (mean§SE) after 9 h and from 0§0 to 47§ 24after 12 h. Formation of capilliconidia was not observed atthis minimum RH required for sporulation. At this RH, nosporulation was observed from 86.3% of the mummiWedcadavers. On the other hand, 2.4% of the mummies dis-charged more than 500 conidia each. Production of conidiawas not apparently diVerent among isolates at any timeinterval and because of the variability in the data, statisticalanalysis was not carried out for data from 96§0.5% RH.
At 100% RH, production of primary conidia was high;and after 12 h, 98.4% of the mummiWed mites had sporu-lated. No capilliconidia were observed at 6 h and produc-tion of capilliconidia was relatively low at 9 and 12 h.
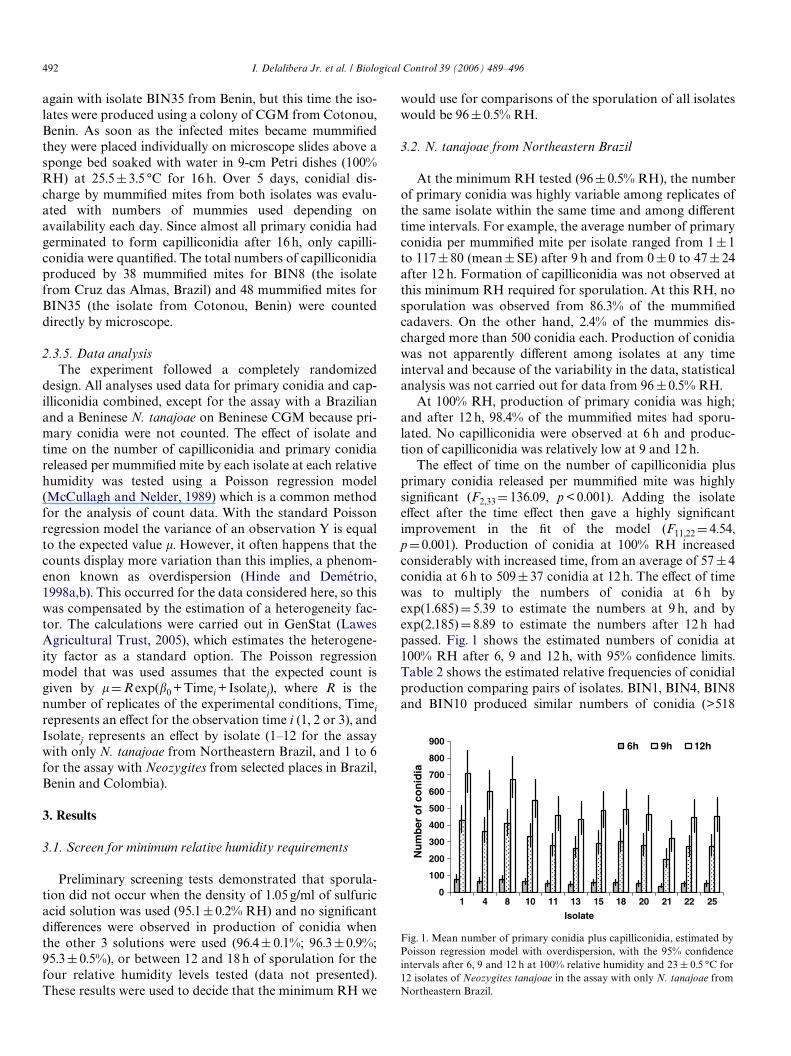
The eVect of time on the number of capilliconidia plusprimary conidia released per mummiWed mite was highlysigniWcant (F2,33D 136.09, p < 0.001). Adding the isolateeVect after the time eVect then gave a highly signiWcantimprovement in the Wt of the model (F11,22D 4.54,pD0.001). Production of conidia at 100% RH increasedconsiderably with increased time, from an average of 57§4conidia at 6 h to 509§37 conidia at 12 h. The eVect of timewas to multiply the numbers of conidia at 6 h byexp(1.685)D 5.39 to estimate the numbers at 9 h, and byexp(2.185)D 8.89 to estimate the numbers after 12 h hadpassed. Fig. 1 shows the estimated numbers of conidia at100% RH after 6, 9 and 12 h, with 95% conWdence limits.Table 2 shows the estimated relative frequencies of conidialproduction comparing pairs of isolates. BIN1, BIN4, BIN8and BIN10 produced similar numbers of conidia (>518
Fig. 1. Mean number of primary conidia plus capilliconidia, estimated byPoisson regression model with overdispersion, with the 95% conWdenceintervals after 6, 9 and 12 h at 100% relative humidity and 23 § 0.5 °C for12 isolates of Neozygites tanajoae in the assay with only N. tanajoae fromNortheastern Brazil.
0
100
200
300
400
500
600
700
800
900
1 4 8 10 11 13 15 18 20 21 22 25
Isolate
Nu
mb
er o
f co
nid
ia
6h 9h 12h
I. Delalibera Jr. et al. / Biological Control 39 (2006) 489–496 493
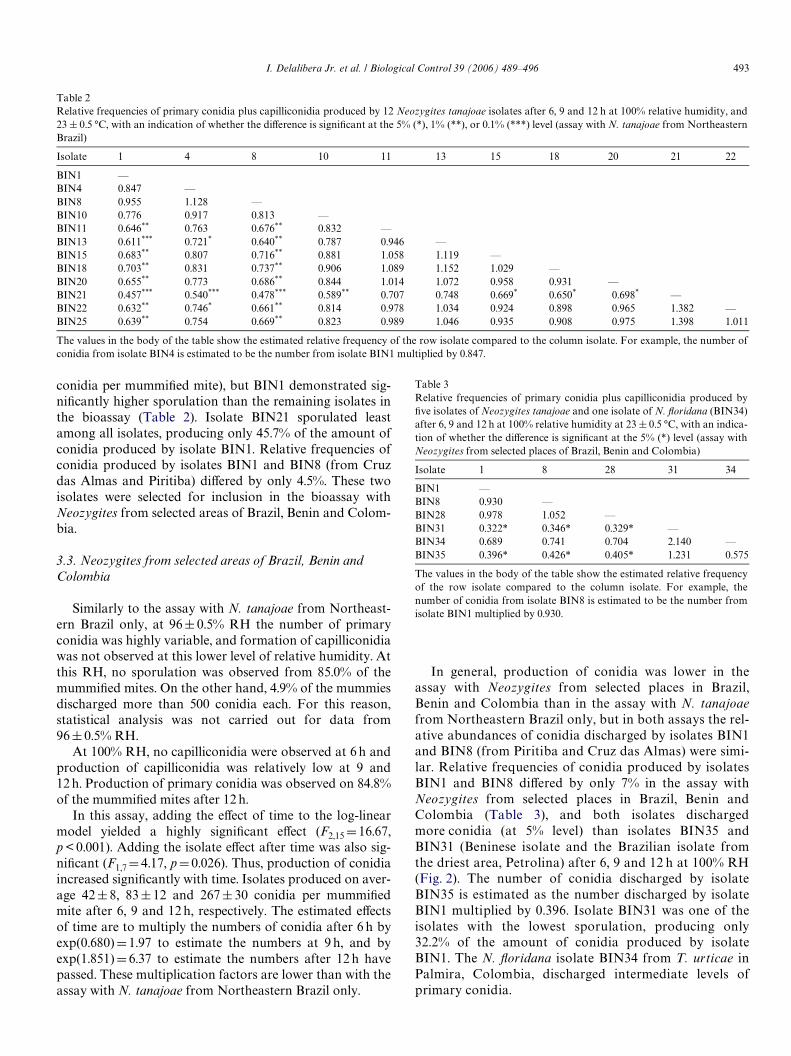
conidia per mummiWed mite), but BIN1 demonstrated sig-niWcantly higher sporulation than the remaining isolates inthe bioassay (Table 2). Isolate BIN21 sporulated leastamong all isolates, producing only 45.7% of the amount ofconidia produced by isolate BIN1. Relative frequencies ofconidia produced by isolates BIN1 and BIN8 (from Cruzdas Almas and Piritiba) diVered by only 4.5%. These twoisolates were selected for inclusion in the bioassay withNeozygites from selected areas of Brazil, Benin and Colom-bia.
3.3. Neozygites from selected areas of Brazil, Benin and Colombia
Similarly to the assay with N. tanajoae from Northeast-ern Brazil only, at 96§ 0.5% RH the number of primaryconidia was highly variable, and formation of capilliconidiawas not observed at this lower level of relative humidity. Atthis RH, no sporulation was observed from 85.0% of themummiWed mites. On the other hand, 4.9% of the mummiesdischarged more than 500 conidia each. For this reason,statistical analysis was not carried out for data from96§ 0.5% RH.
At 100% RH, no capilliconidia were observed at 6 h andproduction of capilliconidia was relatively low at 9 and12 h. Production of primary conidia was observed on 84.8%of the mummiWed mites after 12 h.
In this assay, adding the eVect of time to the log-linearmodel yielded a highly signiWcant eVect (F2,15D16.67,p < 0.001). Adding the isolate eVect after time was also sig-niWcant (F1,7D 4.17, pD0.026). Thus, production of conidiaincreased signiWcantly with time. Isolates produced on aver-age 42§ 8, 83§12 and 267§ 30 conidia per mummiWedmite after 6, 9 and 12 h, respectively. The estimated eVectsof time are to multiply the numbers of conidia after 6 h byexp(0.680)D1.97 to estimate the numbers at 9 h, and byexp(1.851)D6.37 to estimate the numbers after 12 h havepassed. These multiplication factors are lower than with theassay with N. tanajoae from Northeastern Brazil only.
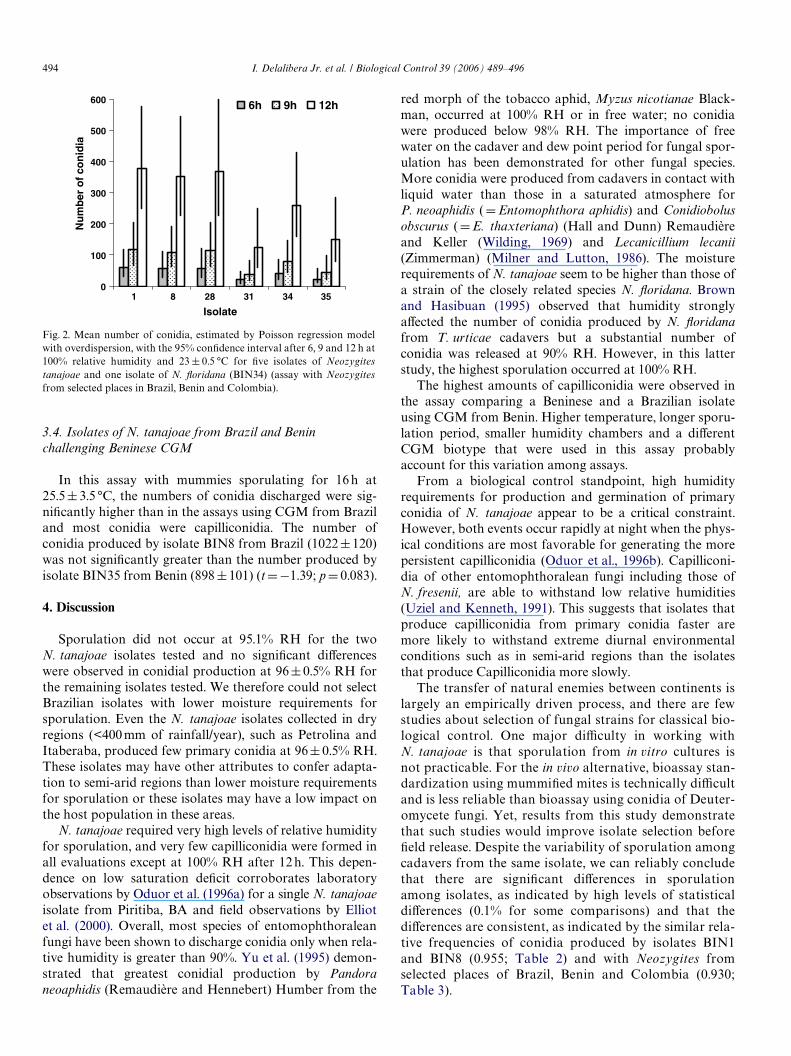
In general, production of conidia was lower in theassay with Neozygites from selected places in Brazil,Benin and Colombia than in the assay with N. tanajoaefrom Northeastern Brazil only, but in both assays the rel-ative abundances of conidia discharged by isolates BIN1and BIN8 (from Piritiba and Cruz das Almas) were simi-lar. Relative frequencies of conidia produced by isolatesBIN1 and BIN8 diVered by only 7% in the assay withNeozygites from selected places in Brazil, Benin andColombia (Table 3), and both isolates dischargedmore conidia (at 5% level) than isolates BIN35 andBIN31 (Beninese isolate and the Brazilian isolate fromthe driest area, Petrolina) after 6, 9 and 12 h at 100% RH(Fig. 2). The number of conidia discharged by isolateBIN35 is estimated as the number discharged by isolateBIN1 multiplied by 0.396. Isolate BIN31 was one of theisolates with the lowest sporulation, producing only32.2% of the amount of conidia produced by isolateBIN1. The N. Xoridana isolate BIN34 from T. urticae inPalmira, Colombia, discharged intermediate levels ofprimary conidia.
Table 3Relative frequencies of primary conidia plus capilliconidia produced byWve isolates of Neozygites tanajoae and one isolate of N. Xoridana (BIN34)after 6, 9 and 12 h at 100% relative humidity at 23 § 0.5 °C, with an indica-tion of whether the diVerence is signiWcant at the 5% (*) level (assay withNeozygites from selected places of Brazil, Benin and Colombia)
The values in the body of the table show the estimated relative frequencyof the row isolate compared to the column isolate. For example, thenumber of conidia from isolate BIN8 is estimated to be the number fromisolate BIN1 multiplied by 0.930.
Isolate 1 8 28 31 34
BIN1 —BIN8 0.930 —BIN28 0.978 1.052 —BIN31 0.322* 0.346* 0.329* —BIN34 0.689 0.741 0.704 2.140 —BIN35 0.396* 0.426* 0.405* 1.231 0.575
Table 2Relative frequencies of primary conidia plus capilliconidia produced by 12 Neozygites tanajoae isolates after 6, 9 and 12 h at 100% relative humidity, and23 § 0.5 °C, with an indication of whether the diVerence is signiWcant at the 5% (*), 1% (**), or 0.1% (***) level (assay with N. tanajoae from NortheasternBrazil)
The values in the body of the table show the estimated relative frequency of the row isolate compared to the column isolate. For example, the number ofconidia from isolate BIN4 is estimated to be the number from isolate BIN1 multiplied by 0.847.
Isolate 1 4 8 10 11 13 15 18 20 21 22
BIN1 —BIN4 0.847 —BIN8 0.955 1.128 —BIN10 0.776 0.917 0.813 —BIN11 0.646** 0.763 0.676** 0.832 —BIN13 0.611*** 0.721* 0.640** 0.787 0.946 —BIN15 0.683** 0.807 0.716** 0.881 1.058 1.119 —BIN18 0.703** 0.831 0.737** 0.906 1.089 1.152 1.029 —BIN20 0.655** 0.773 0.686** 0.844 1.014 1.072 0.958 0.931 —BIN21 0.457*** 0.540*** 0.478*** 0.589** 0.707 0.748 0.669* 0.650* 0.698* —BIN22 0.632** 0.746* 0.661** 0.814 0.978 1.034 0.924 0.898 0.965 1.382 —BIN25 0.639** 0.754 0.669** 0.823 0.989 1.046 0.935 0.908 0.975 1.398 1.011
494 I. Delalibera Jr. et al. / Biological Control 39 (2006) 489–496
3.4. Isolates of N. tanajoae from Brazil and Benin challenging Beninese CGM
In this assay with mummies sporulating for 16 h at25.5§ 3.5 °C, the numbers of conidia discharged were sig-niWcantly higher than in the assays using CGM from Braziland most conidia were capilliconidia. The number ofconidia produced by isolate BIN8 from Brazil (1022§120)was not signiWcantly greater than the number produced byisolate BIN35 from Benin (898§101) (tD¡1.39; pD 0.083).
4. Discussion
Sporulation did not occur at 95.1% RH for the twoN. tanajoae isolates tested and no signiWcant diVerenceswere observed in conidial production at 96§0.5% RH forthe remaining isolates tested. We therefore could not selectBrazilian isolates with lower moisture requirements forsporulation. Even the N. tanajoae isolates collected in dryregions (<400 mm of rainfall/year), such as Petrolina andItaberaba, produced few primary conidia at 96§ 0.5% RH.These isolates may have other attributes to confer adapta-tion to semi-arid regions than lower moisture requirementsfor sporulation or these isolates may have a low impact onthe host population in these areas.
N. tanajoae required very high levels of relative humidityfor sporulation, and very few capilliconidia were formed inall evaluations except at 100% RH after 12 h. This depen-dence on low saturation deWcit corroborates laboratoryobservations by Oduor et al. (1996a) for a single N. tanajoaeisolate from Piritiba, BA and Weld observations by Elliotet al. (2000). Overall, most species of entomophthoraleanfungi have been shown to discharge conidia only when rela-tive humidity is greater than 90%. Yu et al. (1995) demon-strated that greatest conidial production by Pandoraneoaphidis (Remaudière and Hennebert) Humber from the
Fig. 2. Mean number of conidia, estimated by Poisson regression modelwith overdispersion, with the 95% conWdence interval after 6, 9 and 12 h at100% relative humidity and 23 § 0.5 °C for Wve isolates of Neozygitestanajoae and one isolate of N. Xoridana (BIN34) (assay with Neozygitesfrom selected places in Brazil, Benin and Colombia).
0
100
200
300
400
500
600
1 8 28 31 34 35
Isolate
Nu
mb
er o
f co
nid
ia
6h 9h 12h
red morph of the tobacco aphid, Myzus nicotianae Black-man, occurred at 100% RH or in free water; no conidiawere produced below 98% RH. The importance of freewater on the cadaver and dew point period for fungal spor-ulation has been demonstrated for other fungal species.More conidia were produced from cadavers in contact withliquid water than those in a saturated atmosphere forP. neoaphidis (DEntomophthora aphidis) and Conidiobolusobscurus (DE. thaxteriana) (Hall and Dunn) Remaudièreand Keller (Wilding, 1969) and Lecanicillium lecanii(Zimmerman) (Milner and Lutton, 1986). The moisturerequirements of N. tanajoae seem to be higher than those ofa strain of the closely related species N. Xoridana. Brownand Hasibuan (1995) observed that humidity stronglyaVected the number of conidia produced by N. Xoridanafrom T. urticae cadavers but a substantial number ofconidia was released at 90% RH. However, in this latterstudy, the highest sporulation occurred at 100% RH.The highest amounts of capilliconidia were observed inthe assay comparing a Beninese and a Brazilian isolateusing CGM from Benin. Higher temperature, longer sporu-lation period, smaller humidity chambers and a diVerentCGM biotype that were used in this assay probablyaccount for this variation among assays.
From a biological control standpoint, high humidityrequirements for production and germination of primaryconidia of N. tanajoae appear to be a critical constraint.However, both events occur rapidly at night when the phys-ical conditions are most favorable for generating the morepersistent capilliconidia (Oduor et al., 1996b). Capilliconi-dia of other entomophthoralean fungi including those ofN. fresenii, are able to withstand low relative humidities(Uziel and Kenneth, 1991). This suggests that isolates thatproduce capilliconidia from primary conidia faster aremore likely to withstand extreme diurnal environmentalconditions such as in semi-arid regions than the isolatesthat produce Capilliconidia more slowly.
The transfer of natural enemies between continents islargely an empirically driven process, and there are fewstudies about selection of fungal strains for classical bio-logical control. One major diYculty in working withN. tanajoae is that sporulation from in vitro cultures isnot practicable. For the in vivo alternative, bioassay stan-dardization using mummiWed mites is technically diYcultand is less reliable than bioassay using conidia of Deuter-omycete fungi. Yet, results from this study demonstratethat such studies would improve isolate selection beforeWeld release. Despite the variability of sporulation amongcadavers from the same isolate, we can reliably concludethat there are signiWcant diVerences in sporulationamong isolates, as indicated by high levels of statisticaldiVerences (0.1% for some comparisons) and that thediVerences are consistent, as indicated by the similar rela-tive frequencies of conidia produced by isolates BIN1and BIN8 (0.955; Table 2) and with Neozygites fromselected places of Brazil, Benin and Colombia (0.930;Table 3).

I. Delalibera Jr. et al. / Biological Control 39 (2006) 489–496 495
Decisions on which isolates to choose for release arecomplex, and bioassays comparing important attributes ofthe pathogen can Wrst eliminate the less appropriate isolates(e.g., those that do not infect or sporulate well). An exampleof an isolate not adapted to CGM is BIN21. This isolatewas collected from CGM but caused very low mummiWca-tion (Delalibera and Hajek, 2004) and sporulated verypoorly in this study. The N. Xoridana isolate (BIN34 fromColombia) was initially considered a candidate for biologi-cal control of CGM. Later, this isolate proved to be lessspeciWc and less pathogenic to CGM than N. tanajoae iso-lates from Brazil (Delalibera and Hajek, 2004). We demon-strated in this study that N. Xoridana (BIN34) alsoproduced fewer conidia than some of the Brazilian isolatesof N. tanajoae.
N. tanajoae was introduced to one district in Benin in1999 (Hountondji et al., 2002) using isolates Altal.brz andColal.brz, from Alto Alegre, BA and Cruz das Almas, BA,respectively. Unfortunately, these isolates were not avail-able for inclusion in the present study.
Releases of N. tanajoae are planned for the future forother countries in Africa. DiVerences in virulence againstCGM and production of conidia among isolates should beimportant factors when selecting isolates for further intro-ductions into Africa to reduce crop losses caused by CGM.In this study, we have demonstrated that BIN1, BIN4,BIN8, and BIN10, all from Northeastern Brazil, producedthe most spores. Among these four isolates, during bioas-says with CGM, BIN1 displayed the lowest LT50 andcaused the greatest percent mummiWcation in the shortestperiod of time (Delalibera and Hajek, 2004). BIN1 also dis-charged more conidia and caused greater percent mummiW-cation in a shorter period of time than the endemicBeninese isolate BIN35 (Delalibera and Hajek, 2004). Bio-assays also demonstrated that BIN11, BIN12 and BIN32from Northeastern Brazil are highly virulent to CGM (Del-alibera and Hajek, 2004) but, unfortunately, these isolatescould not be included in the present study. Along withBIN1, these latter isolates may have the best potential forfuture classical biological control releases in Africa.
Acknowledgments
This study was partially supported by CNPq (TheNational Council for ScientiWc and Technological Develop-ment), Brazil, and the United Nations Development Pro-gram (GLO/91/013) as part of a collaborative researchproject by the International Institute of Tropical Agricul-ture (IITA), Brazilian Agriculture Research Corporation(EMBRAPA) and International Center for Tropical Agri-culture (CIAT). We thank Gilberto J. de Moraes, RichardA. Humber, for scientiWc input, Adriano Medeiros andCorifeu F. Buzetti for their help with the Brazilian CGMassays; and Rachid Hanna, Andy Cherry, Fabien Houton-dji, Honoré Dossounon, Jacques Kuinsou, Roland Bocco,Hugues Baimey and Adande Marcellin for assistance in theBeninese CGM assay. We also thank IITA for giving space
and support for conducting the third bioassay in theirCotonou laboratories. Support for the trip to Cotonou wasprovided by the Richard BradWeld Research Award,Cornell University.
References
Ben’Ze’ev, I., Kenneth, R.G., 1981. Zoophthora radicans and Zoophthorapetchi, two species of the “Sphaerosperma group” attacking leaf-hop-pers and frog-hoppers. Entomophaga 26, 131–142.
Boucias, D.G., Pendland, J.C., 1998. Principles of Insect Pathology. Klu-wer Academic Publishers, Boston.
Brown, G.C., Hasibuan, R., 1995. Conidial discharge and transmissioneYciency of Neozygites Xoridana, an entomopathogenic fungus infect-ing two-spotted spider mites under laboratory conditions. J. Invertebr.Pathol. 65, 10–16.
Carruthers, R.I., Soper, R.S., 1987. Fungal diseases. In: Fuxa, J.R., Tanada,Y. (Eds.), Epizootiology of Insect Diseases. Wiley, New York, pp. 357–416.
Delalibera Jr., I., Hajek, A.E., 2004. Pathogenicity and speciWcity of Neo-zygites tanajoae and Neozygites Xoridana (Zygomycetes: Entomoph-thorales) isolates pathogenic to the cassava green mite. Biol. Control30, 608–616.
Delalibera Jr., I., Gomez, D.R.S., de Moraes, G.J., de Alencar, J.A., Araujo,W.F., 1992. Infection of Mononychellus tanajoa (Acari: Tetranychidae)by the fungus Neozygites sp. (Zygomycetes: Entomophthorales) innortheastern Brazil. Fla. Entomol. 75, 145–147.
Delalibera Jr., I., de Moraes, G.J., Lapointe, S.L., da Silva, C.A.D., Tamai,M.A., 2000. Temporal variability and progression of Neozygites sp.(Zygomycetes: Entomophthorales) in populations of Mononychellustanajoa (Bondar) (Acari: Tetranychidae). An. Soc. Entomol. Brasil 29,523–535.
Delalibera Jr., I., Hajek, A.E., Humber, R.A., 2003. Use of cell culturemedia for cultivation of the mite pathogenic fungi Neozygites tanajoaeand N. Xoridana. J. Invertebr. Pathol. 84, 119–127.
Delalibera Jr., I., Hajek, A.E., Humber, R.A., 2004a. Neozygites tanajoaesp. nov., a pathogen of the cassava green mite. Mycologia 96 (5),1002–1009.
Delalibera Jr., I., Humber, R.A., Hajek, A.E., 2004b. Preservation ofin vitro cultures of the mite pathogenic fungus Neozygites tanajoae.Can. J. Microbiol. 50, 579–586.
Elliot, S.L., de Moraes, G.J., Delalibera Jr., I., da Silva, C.A.D., Tamai,M.A., Mumford, J.D., 2000. Potential of the mite-pathogenic fungusNeozygites Xoridana (Entomophthorales: Neozygitaceae) for control ofthe cassava green mite Mononychellus tanajoa (Acari: Tetranychidae).Bull. Entomol. Res. 90, 191–200.
Hajek, A.E., Wraight, S.P., Vandenberg, J.D., 2001. Control of arthropodsusing pathogenic fungi. In: Pointing, S.B., Hyde, K.D. (Eds.), Bio-exploitation of Filamentous Fungi. Fungal Diversity Press, HongKong, pp. 309–347.
Hinde, J.P., Demétrio, C.G.B., 1998a. Overdispersion: models and estima-tion. Comput. Stat. Data Anal. 27, 151–170.
Hinde, J.P., Demétrio, C.G.B., 1998b. Overdispersion: models and estima-tion, São Paulo: Associação Brasileira de Estatística, 73p.
Hountondji, F.C.C., Lomer, C.J., Hanna, R., Cherry, A.J., Dara, S.K.,2002. Field evaluation of Brazilian isolates of Neozygites Xoridana(Entomophthorales: Neozygitaceae) for the microbial control ofcassava green mite in Benin, West Africa. Biocontrol Sci. Technol.12, 361–370.
Lawes Agricultural Trust, 2005. GenStat, eighth ed. Rothamsted Agricul-tural Station, UK.
McCullagh, P., Nelder, J.A., 1989. Generalized Linear Models, second ed.Chapman & Hall, London. 511p.
Milner, R.J., Lutton, G.G., 1986. Dependence of Verticillium lecanii(Fungi: Hyphomycetes) on high humidities for infection and sporula-tion using Myzus persicae (Homoptera Aphididae) as host. Environ.Entomol. 15, 380–382.

496 I. Delalibera Jr. et al. / Biological Control 39 (2006) 489–496
Mullens, B.A., Rodriguez, J.L., 1985. Dynamics of Entomophthora muscae(Entomophthorales: Entomophthoraceae) conidial discharge fromMusca domestica (Diptera: Muscidae) cadavers. Environ. Entomol. 14,317–322.
Newman, G.G., Carner, G.R., 1975. Environmental factors aVecting conid-ial sporulation and germination of Entomophthora gammae. Environ.Entomol. 4, 615–618.
Oduor, G.I., de Moraes, G.J., Van der Geest, L.P.S., Yaninek, J.S., 1996a.Production and germination of primary conidia of Neozygites Xoridana(Zygomycetes: Entomophthorales) under constant temperatures,humidities, and photoperiods. J. Invertebr. Pathol. 68, 213–222.
Oduor, G.I., Yaninek, J.S., Van der Geest, L.P.S., de Moraes, G.J., 1996b.Germination and viability of capilliconidia of Neozygites Xoridana(Zygomycetes: Entomophthorales) under constant temperature,humidity, and light conditions. J. Invertebr. Pathol. 67, 267–278.
Solomon, M.E., 1951. Control of humidity with potassium hydroxide, sul-phuric acid and other solutions. Bull. Entomol. Res. 42, 543–553.
Stevens, N.E., 1916. A method of studying the humidity relations of fungiin culture. Phytopathology 6, 428–432.
Tanada, Y., Kaya, H.K., 1993. Insect Pathology. Academic Press, San Diego.Uziel, A., Kenneth, R.G., 1991. Survival of primary conidia and capillicon-
idia at diVerent humidities in Erynia (subgen. Zoophthora) spp. and inNeozygites fresenii (Zygomycotina: Entomophthorales), with specialemphasis on Erynia radicans. J. Invertebr. Pathol. 58, 118–126.
Wilding, N., 1969. EVect of humidity on the sporulation of Entomophthoraaphidis and E. thaxteriana. Trans. Brit. Mycol. Soc. 53, 126–130.
Yu, Z., Nordin, G.L., Brown, G.C., Jackson, D.M., 1995. Studies onPandora neoaphidis (Entomophthorales: Entomophthoraceae) infec-tious to the red morph of tobacco aphid (Homoptera: Aphididae).Environ. Entomol. 24, 962–966.




















