
Effect of prey stages of the two-spotted mite Tetranychus urticae on functional response of the...
12
Acta Phytopathologica et Entomologica Hungarica 46 (2), pp. 277–288 (2011) DOI: 10.1556/APhyt.46.2011.2.12 0238–1249/$ 20.00 © 2011 Akadémiai Kiadó, Budapest Effect of Prey Stages of the Two-Spotted Mite Tetranychus urticae on Functional Response of the Coccinellid Predator Stethorus gilvifrons M. A. OSMAN 1 and M. H. BAYOUMY 2* 1 Faculty of Agriculture, Zoology Department, Mansoura University, Mansoura 35516, Egypt 2 Faculty of Agriculture, Entomology Department, Mansoura University, Mansoura 35516, Egypt (Received: 23 February 2011; accepted: 8 April 2011) The effect of prey stages of the two-spotted spider mite Tetranychus urticae on the functional response of the coccinellid predator Stethorus gilvifrons was studied at 25±1 °C, 65±10% rh, and 16:8 h (L:D photoperiod). The functional response of S. gilvifrons female, to increasing prey stage density from 10−100 preys per patch, was of the curvilinear shape depicting Roger’s type II response with the highest consumption rate of larval prey stage. Results of the maximum likelihood analyses confirmed this type. By plotting a type II functional response model and polynomial logistic regression model to the numbers and proportions of consumed stages of T. urticae, respectively, the resulting regression lines fitted the data well. However, a composite waving form that appeared to be a decelerating (type II) response at low to medium prey densities and an accelerating (type III) response at high prey densities. We assume that the small size of experimental arena or predator feeding behaviour at low and high prey densities may have induced the modified type II response. A sensitivity analysis of the functional response model shows the crucial effect of prey stages on predation rates by S. gilvifrons. Theoretically, based on the low- est handling time for S. gilvifrons on larval stage of T. urticae, the maximum numbers of mites that could consume by a single female of S. gilvifrons within 24 h period were 77.42 larvae. These results improve our understanding of the interaction among S. gilvifrons and T. urtica stages and provide insights into the control of T. urticae. Keywords: ladybeetles, Tetranychidae, Roger “predator-random” equation, handling time, attack rate. Predation is assumed to be one of the significant biotic mortality factors reducing in- sect pest populations, and using them in insect pest management programs has been re- ceiving increased attention because of the current need to reduce the exclusive use of insecticides for pest control (Sarmento et al., 2007). Stethorus gilvifrons Mulsant (Coleoptera: Coccinellidae) is a biological control agent of the two spotted spider mite, Tetranychus urticae Koch (Acari: Tetranychidae), which is a serious pest of different crops and ornamental plants over the world (e.g. Zhang, 2003; Alatawi et al., 2005). It feeds vo- raciously on different growth stages of spider mites (Tetranchidae) and the closely related Tenuipalpididae, which known as false spider mites or flat mites (Afshari, 1999). Prior to the release of natural enemy in a biological control program, it is essential to evaluate its efficiency under laboratory conditions. One informative method for assess- *Corresponding author; e-mail: [email protected] Aphyt_46(2011)2_3tord-imp:Aphyt_46(2011)2_2_tord 11/17/11 4:09 PM Page 277
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Effect of prey stages of the two-spotted mite Tetranychus urticae on functional response of the...

Acta Phytopathologica et Entomologica Hungarica 46 (2), pp. 277–288 (2011)DOI: 10.1556/APhyt.46.2011.2.12
0238–1249/$ 20.00 © 2011 Akadémiai Kiadó, Budapest
Effect of Prey Stages of the Two-Spotted MiteTetranychus urticae
on Functional Response of the Coccinellid PredatorStethorus gilvifrons
M. A. OSMAN1 and M. H. BAYOUMY2*
1Faculty of Agriculture, Zoology Department, Mansoura University, Mansoura 35516, Egypt2Faculty of Agriculture, Entomology Department, Mansoura University, Mansoura 35516, Egypt
(Received: 23 February 2011; accepted: 8 April 2011)
The effect of prey stages of the two-spotted spider mite Tetranychus urticae on the functional response ofthe coccinellid predator Stethorus gilvifrons was studied at 25±1 °C, 65±10% rh, and 16:8 h (L:D photoperiod).The functional response of S. gilvifrons female, to increasing prey stage density from 10−100 preys per patch,was of the curvilinear shape depicting Roger’s type II response with the highest consumption rate of larval preystage. Results of the maximum likelihood analyses confirmed this type. By plotting a type II functional responsemodel and polynomial logistic regression model to the numbers and proportions of consumed stages of T. urticae,respectively, the resulting regression lines fitted the data well. However, a composite waving form that appeared tobe a decelerating (type II) response at low to medium prey densities and an accelerating (type III) response at highprey densities. We assume that the small size of experimental arena or predator feeding behaviour at low and highprey densities may have induced the modified type II response. A sensitivity analysis of the functional responsemodel shows the crucial effect of prey stages on predation rates by S. gilvifrons. Theoretically, based on the low-est handling time for S. gilvifrons on larval stage of T. urticae, the maximum numbers of mites that could consumeby a single female of S. gilvifrons within 24 h period were 77.42 larvae. These results improve our understandingof the interaction among S. gilvifrons and T. urtica stages and provide insights into the control of T. urticae.
Keywords: ladybeetles, Tetranychidae, Roger “predator-random” equation, handling time, attack rate.
Predation is assumed to be one of the significant biotic mortality factors reducing in-sect pest populations, and using them in insect pest management programs has been re-ceiving increased attention because of the current need to reduce the exclusive use ofinsecticides for pest control (Sarmento et al., 2007). Stethorus gilvifrons Mulsant(Coleoptera: Coccinellidae) is a biological control agent of the two spotted spider mite,Tetranychus urticae Koch (Acari: Tetranychidae), which is a serious pest of different cropsand ornamental plants over the world (e.g. Zhang, 2003; Alatawi et al., 2005). It feeds vo-raciously on different growth stages of spider mites (Tetranchidae) and the closely relatedTenuipalpididae, which known as false spider mites or flat mites (Afshari, 1999).
Prior to the release of natural enemy in a biological control program, it is essentialto evaluate its efficiency under laboratory conditions. One informative method for assess-
*Corresponding author; e-mail: [email protected]
Aphyt_46(2011)2_3tord-imp:Aphyt_46(2011)2_2_tord 11/17/11 4:09 PM Page 277

ing the efficiency of natural enemies is the study of their foraging behavior including func-tional response (Fathipour et al., 2006; Bayoumy et al., 2009).. Functional response is thenumber of preys successfully consumed per predator as a function of prey density(Solomon, 1949). It is a commonly measured attribute of natural enemies. Practitioners ofinundative biological control may use functional response to estimate the appropriate num-bers of bio-control agents to be released in order to bring about an immediate reduction inpest numbers (Mills and Lacan, 2004).. Holling (1965) proposed three types of functionalresponses: type I, a linear rise to a plateau; type II, a curvilinear rise to a plateau; and typeIII, a sigmoid curve rising to a plateau, which then levels off under the influence of han-dling time or satiation (Hassell, 2000). Among the types of functional responses, types IIand III have received the most attention (Allahyari et al., 2004). Coccinellids are one of theimportant groups of predatory insects because of their biocontrol potential (e.g. Obrycki,1998; Atlihan et al., 2010). Although they generally show a type II functional response(e.g. Ofuya and Akingbohungbe, 1988; Uygun and Atlihan, 2000), the other types of func-tional responses are also reported for the group (e.g. Hassell et al., 1977; Hu et al., 1989).On the other hand, the response of a single species of coccinellid predator can differ be-tween various types of prey (Hu et al., 1989) and the response of different stages of a pred-ator can differ within a single type of prey (Koch et al., 2003). Here, we present a studyaddressing the influences of prey stage densities on the predation rate (i.e. functional re-sponse) of S. gilvifrons. This is one part of a comprehensive evaluation of the biologicalcontrol potential of S. gilvifrons and its interaction with T. urticae.
Many early studies of various species of Stethorini concluded that they are ‘‘highdensity predators” unable to regulate spider mite populations at low densities (e.g. Baileyand Caon, 1986; Tanigoshi and McMurtry, 1977). Congdon et al. (1993) challenged this‘‘high density predator” designation and asserted that high-density spider mite populationshave become common only since the advent of synthetic insecticides (Huffaker et al., 1970).Haji-Zadeh et al. (1993) and Afshari (1999) have conducted some bionomic studies on S.gilvifrons, however, no study in detail has been conducted on the influence of T. urticaestages on its functional responses. Only Sohrabi and Shishehbor (2007) reported a type IIIresponse of S. gilvifrons on Tetranychus turkestani Ugarov and Nikolski. Therefore, theobjective of this study is to quantify the effect of prey stage densities on consumption rateof S. gilvifrons for improving our understanding of prey-predator interaction and to knowsome information about the prey stage, which could be maximized mass rearing programfor this predator. Thus, this study might lead to develop a better strategy for biological con-trol of T. urticae.
Materials and Methods
1. Stock culture of Tetranychus urticae and Stethorus gilvifrons
Tetranychus urticae, used as prey for S. gilvifrons, was reared on the whole plants ofpotted beans, Phaseolus vulgaris L. at 26±3 °C, 65±10% rh, and 16:8 h (L:D photope-
Osman, Bayoumy: Coccinellid predator Stethorus gilvifrons
Acta Phytopathologica et Entomologica Hungarica 46, 2011
278
Aphyt_46(2011)2_3tord-imp:Aphyt_46(2011)2_2_tord 11/17/11 4:09 PM Page 278

riod). The initial population of S. gilvifrons was initially originated from caster plants (Rici-nus communis L.) in the pupal stage located at Mansoura district, Egypt. The collectedpupae were kept in Eppendorf tubes and maintained in the incubator at 25±1 °C and 16:8h (L:D photoperiod) until adult emergence. Then, the stock culture of S. gilvifrons wasstarted using all stages of T. urticae on caster leaves. One leaf disc placed upside-down onwater saturated cotton (2 cm thick) in a plastic box (20×10×5 cm). Water added daily tocover the base of the box to prevent the mites from escaping. A surplus of all stages of T.urticae was brushed daily onto the leaf arenas using a soft brush and funnel. The stock cul-ture was maintained in the incubator at 25±1, 65±10% rh, and 16:8 h (L:D photoperiod).The leaf discs were renewed once a week. The predatory mites used in the experiment werereared for two successive generations prior to this study (i.e. functional response).
2. General experimental conditions
Functional response of S. gilvifrons was conducted onto the modified Huffaker cell(Overmeer, 1985) provided with caster leaf disc as an experimental arena. The tested den-sities of T. urticae for S. gilvifrons were transferred from potted bean leaves to caster leafdisc using a soft brush. Prior to the functional response test, the predators collected fromthe stock culture in their pupal stage and places in Petri dishes. The emerged adults wereprovided with T. urticae as a source for food for 48 h, then separated individually in Ep-pendorf tubes and starved for another 24 h before used in the experiment to standardize theirhunger.
3. Functional response
To evaluate the functional response of S. gilvifrons, to change in the prey stage den-sity of the two spotted spider mite, individual predator females (72 h old) were exposed toten prey density levels (viz. 10, 20, 30, 40, 50, 60, 70, 80, 90, 100), of each tested stage (i.e.nymph, larva, and adult) for 24 h at 25±1 °C, 65±10% rh, and 16:8 h (L:D photoperiod).In case of adult mite females, newly females transferred directly from the stock culture tothe experimental arena to avoid oviposition of females in the experimental arena. Fifteenreplications of each prey density were set up simultaneously. After 24 h, the predators wereremoved from the experimental arenas and the consumed mites were recorded. In order toevaluate the ratio of natural mortality of prey, control treatments were performed with theabove-mentioned prey densities, but in the absence of predators. The mite stages consumedby N. includens were not replaced during the experimental. Only replicates with the pred-ator surviving after 24 h exposure time were considered.
4. Data analysis
Data from functional response experiments were used to establish the type and pa-rameters of functional response using the approach developed by Juliano (2001). This ap-proach includes two sequential analyses. Initially, the shape (type) of functional response
Osman, Bayoumy: Coccinellid predator Stethorus gilvifrons
Acta Phytopathologica et Entomologica Hungarica 46, 2011
279
Aphyt_46(2011)2_3tord-imp:Aphyt_46(2011)2_2_tord 11/17/11 4:09 PM Page 279

must be determined, typically by determining if the data fit a types II or III functional re-sponse. A polynomial logistic regression of the proportion of attacked preys (Na/N0) vs.the initial number of prey offered (N0) is established for distinguishing between both types:
(1)
where Na is the number of prey attacked, N0 is the initial prey density, and the parametersP0, P1, P2 and P3 are the constant, linear, quadratic and cubic parameters, respectively,related to the slope of the curve. Maximum likelihood estimates of parameters P0 to P3 wereobtained by applying logistic regression (CATMOD procedure in SAS software) (SAS,2000) to a dichotomous variable that equaled 0 for surviving preys and 1 for attacked preys.As in many cases a cubic or higher order expression are necessaries for good fits.
The curves of Na/N0 vs. N0 when the response is a type II, have the shape of a neg-ative exponential (proportion of attacked preys decaying exponentially) and when the re-sponse is a type III have a dome-shape, with a maximal of proportion attacked and then acontinuous decline. These curves may both be fit by quadratic (or higher order) polynomi-als expressions. In the type II curve, the linear (P1 in Eq. 1) and quadratic (P2 in Eq. 1) termwould be negative (initially decreasing) whereas in the type III curve the linear term wouldbe positive (initially increasing) and the quadratic term negative (Trexler et al., 1988; Ju-liano, 2001). Thus, one criterion for separating type I to type III functional responses is toanalyze the proportion of prey attacked and to test for significant positive or negative lin-ear coefficients from Eq. 1. Linear term not significantly different from 0 indicates a typeI functional response, a significant negative indicates a type II response, and a significantpositive indicates a type III (Juliano, 2001).
Once the functional response type was determined from logistic regression analysesand log likelihood-ratio tests, the second part of the analyses uses an iterative non-linearleast squares regression (NLIN procedure with Proc method of SAS) (SAS, 2000) to esti-mate the parameters of functional response (Juliano, 2001). Data were fit to “random-pred-ator” equation (Eq. 2) (Roger, 1972), a modification of Holling’s disk equation (Holling,1959), that is regarded as more appropriate because it considers prey density to be affectedby prey consumption (Hazzard and Ferro, 1991) as well as because the experiment wascarried out without prey replacement during the course of the experiment:
(2)
where T is the time of exposure (24 h), a is the rate of successful attack, and Th is thehandling time.
Values of Th, estimated by non-linear least squares regression, were used to calcu-late maximum attack rate as T/Th (Hassell, 2000), that represent the maximal number ofpreys that could be consumed by a predator of S. gilvifrons during 24 h.
N N a T N Ta h a= − −( ) { }0 1 exp
N
N
P P N P N P N
P P N Pa
0
0 1 0 2 02
3 03
0 1 0 21=
+ + + ++ + + +exp ( )
exp ( NN P N02
3 03+ )
Osman, Bayoumy: Coccinellid predator Stethorus gilvifrons
Acta Phytopathologica et Entomologica Hungarica 46, 2011
280
Aphyt_46(2011)2_3tord-imp:Aphyt_46(2011)2_2_tord 11/17/11 4:09 PM Page 280

Results
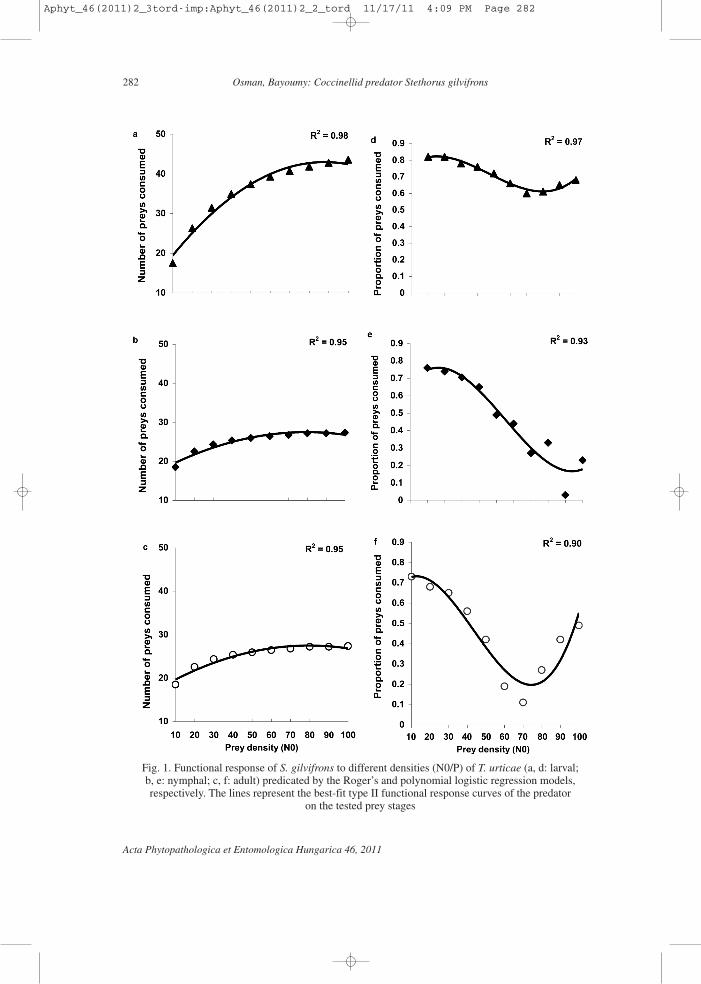
Prey consumption by S. gilvifrons increased from 10 to 60, 10 to 55.93 and 10 to35.73 individuals with increasing in density of larvae, nymphs and adults of T. urticae from10 to 60, 10 to 60 and 10 to 50, respectively. In contrast, the proportion of consumed preysby S. gilvifrons decreased from 1 to 0.43, 1 to 0.28 and 1 to 0.13 with increase in the den-sity of larvae, nymphs and adults of T. urticae from 10 to 100, respectively. The increasein prey consumption and the decrease in proportion of consumed preys by the predatorwith the increase in densities of prey stages were curvilinear in shape.
The accelerated decrease in the proportion of consumed stages of T. urticae in rela-tion to their density gave a good fit to the description of a type II functional response. Re-sults of the maximum likelihood analyses confirmed this observation (Table 1). By plottinga type II functional response and polynomial logistic regression models to the number andproportions of consumed stages of T. urticae, respectively, the resulting regression linesfitted the data well (Fig. 1). However, the curve for the proportion consumed was similarfor the predator on the three tested prey stages: a waving form that appeared to be a com-bination of decelerating (as in type II) at low-medium prey densities, followed by acceler-ating (as in type III) at high prey densities (Fig. 1d, e, f). However, the parameters ofpolynomial regression analysis that distinguishes type II (when it is negative) from type III(when it is positive) indicated neither a type II nor type III response (Table 1). The signif-
Osman, Bayoumy: Coccinellid predator Stethorus gilvifrons
Acta Phytopathologica et Entomologica Hungarica 46, 2011
281
Table 1
Maximum likelihood estimates from logistic regression of the proportion of different stages of T. urticaeconsumed by S. gilvifrons as a function of initial prey density
*A significant negative estimate for the parameter P1 indicates that the slope of the functional responsecurve is declining, thus a type II functional response
Prey stages Parameter Estimate SE Chi-Square (χ2) P-value
Intercept (P0) 131.2 17.23 58.01 ***
Larval*Linear (P1) −4.426 0.62 50.44 ***
Quadratic (P2) 0.0501 0.0075 45.24 **
Cubic (P3) −0.00019 0.000029 41.67 *
Intercept (P0) 31.2786 5.6602 30.54 ***
Nymphal*Linear (P1) −0.8991 0.2203 16.65 ***
Quadratic (P2) 0.00865 0.00282 9.43 **
Cubic (P3) −0.00003 0.000012 5.92 *
Intercept (P0) 21.6752 2.1967 97.36 ***
Adult*Linear (P1) −0.7456 0.0952 61.36 ***
Quadratic (P2) 0.00856 0.00134 40.86 ***
Cubic (P3) −0.00003 0.000006 32.15 ***
Aphyt_46(2011)2_3tord-imp:Aphyt_46(2011)2_2_tord 11/17/11 4:09 PM Page 281
Osman, Bayoumy: Coccinellid predator Stethorus gilvifrons
Acta Phytopathologica et Entomologica Hungarica 46, 2011
282
Fig. 1. Functional response of S. gilvifrons to different densities (N0/P) of T. urticae (a, d: larval; b, e: nymphal; c, f: adult) predicated by the Roger’s and polynomial logistic regression models,respectively. The lines represent the best-fit type II functional response curves of the predator
on the tested prey stages
Aphyt_46(2011)2_3tord-imp:Aphyt_46(2011)2_2_tord 11/17/11 4:09 PM Page 282

icantly negative parameters of P1 for S. gilvifrons in relation to different stages of T. urticae,suggested that the slopes of the functional response curves were declining, which is char-acteristics of a type II functional response.
The highest instantaneous rate of attack (a) and the lowest handling time (Th) for S.gilvifrons were related to the adult and larval stages of T. urticae, respectively (Table 2).Theoretically, based on the lowest handling time for S. gilvifrons on larval stage of T. ur-ticae, the maximum numbers of T. urticae that could be consumed by a single female of S.gilvifrons within 24 h period were 77.42 larvae (24 h/Th).
Discussion
Functional response is a widely used test for understanding the prey–predator orhost–parasitoid interaction and allows the determination of which predator/parasitoidspecies is the most effective at killing a particular prey/host. The exactness of functional re-sponse as a comparison tool in above-mentioned fields is highly related to models and dataanalysis; the use of inappropriate models and methods of analysis may result in an incor-rect estimation.
The response of S. gilvifrons to the tested stages was characterized by a reduction inthe proportion of prey consumed with increasing prey density, reaching a minimum con-sumption of ca. 0.43, 0.28 and 0.13 for larvae, nymphs and adults of T. urticae at the high-est prey density (100 T. urticae larvae, nymphs, adults). Decreasing in proportion of preyconsume with increasing prey density is common for arthropods predator as a result of sa-tiation level and handling time required for each prey item consumed (Holling, 1961).
For each density, predation by S. gilvifrons varied between the stages of T. urticae.At density of 10, 20, 30 and 40 T. urticae, all prey individuals were consumed, what sug-gests that prey density was below the level necessary for survival. At prey density of 50, theproportion of prey consumed was 0.82 and 0.98, and 0.73 on the larvae, nymphs, and adultsof T. urticae, respectively. Each adult female may consume 30–60 mites per day for S.punctum (LeConte) (Tanigoshi and McMurtry, 1977), S. punctillum Weise (Roy et al.,2003), S. japonicusKamiya (Mori et al., 2005), and S. tridensGordon (Fiaboe et al., 2007).According to Houck (1991), the feeding behavior of Stethorus varies with prey density and
Osman, Bayoumy: Coccinellid predator Stethorus gilvifrons
Acta Phytopathologica et Entomologica Hungarica 46, 2011
283
Table 2
Parameter estimates (95% confidence interval) of the attack constant [a (h–1)], handling time [Th (h)],and maximum number of killed preys (T/Th) for S. gilvifrons females preying different stages of T.
urticae derived from Holling disc equation
Prey stages
aAsymptotic 95% CI
Th
Asymptotic 95% CIT/Th
lower upper lower upper
Larval 0.08±0.008 0.0673 0.1003 0.31±0.02 0.2752 0.3528 77.42
Nymphal 0.11±0.02 0.0753 0.1493 0.46±0.03 0.4050 0.5182 52.17
Adult 0.214±0.09 0.0432 0.3843 0.83±0.05 0.7250 0.9264 28.92
Aphyt_46(2011)2_3tord-imp:Aphyt_46(2011)2_2_tord 11/17/11 4:09 PM Page 283

the results found agree with this statement. At higher prey densities, the predators spend lesstime searching the prey resulting in higher prey consumption than in lower densities butpredators require time to attack the prey. High prey density in this study could have inter-fered with the predation process in two different ways: (a) positively, by inducing abandonof prey not completely consumed or (b) negatively, due to the interference caused by theincreased numbers of prey running into the arena and around the predators. Some attrac-tive characteristics of Stethorini for mite biological control are their prey consumption,longevity and high reproductive capacity.
The type of functional response and its parameters for a natural enemy could be af-fected by some factors such as: host plant, temperature and type of prey or host ( Wang andFerro, 1998; De Clercq et al., 2000; Mohaghegh et al., 2001), and prey stages (Nwilene andNachman, 1996; Vantornhout, 2006). Although the type of functional response for S. gil -vifrons did not alter among the different stages of T. urticae in this study, the effects ofthese factors are reflected in the values of the attack rate and the handling time.
Although the logistic regression determined the shape of the functional responses ofS. gilvifrons to be a type II response and significant models could be fitted, the parameter es-timates of these functional responses were not always significantly different from zero. Thelack of significant parameters complicates ecological interpretation of the estimated func-tional responses. Therefore, ecologists normally face difficulties in determining functionalresponse when the curve lies between type II and type III. Hence, a polynomial regressionanalysis that can best determine the functional response is highly needed (Trexler et al.,1988). However, Juliano (pers. comm. with I. Vantornhout) reported that the best guide todistinguish type II from type III response is to observe the shape of the fitted curve. Desig-nation to either one of the Holling’s functional response types may be an oversimplification.The proportion of preys that were consumed by S. gilvifrons exhibited the same character-istic curve on the tested stages of prey: a composite waving form that appeared to be a de-celerating type II at low to medium prey densities and an accelerating type III at medium athigh prey densities. The explanation of this phenomenon may be difficult to explain, how-ever such a composite functional response may be induced by the small size of experimen-tal arena and/or at high prey densities, S. gilvifrons continued to feed even when satiatedand continued to attack mites, initiate feeding, but then subsequently abandon the prey withminimal feeding of bodily parts. Byeon et al. (2011) reported another waving form of func-tional response for the braconid, Aphidius colemani Viereck and the aphelinid Aphelinusasychis Walker parasitoids attacking the green peach aphid, Myzus persicae (Sulzer). Theyfound a composite waving form that appeared to be a decelerating type II at low host den-sity and an accelerating-and decelerating type III at medium to high host density.
A type II functional response is the most common type reported for most coccinel-lid predators (Isikber, 2005). For a type II response, there should be a decline in the pro-portion of consumed preys as the density increases, so that the linear term should benegative. The significant negative values for the linear parameter obtained in this studyconfirm the type II response for S. gilvifrons on all tested prey stages. The type II responseis similarly reported for Stethorus vagans (Blackburn) (Ullah, 2000), S. japonicus (Gotohet al., 2004), and S. tridens (Britto et al., 2009), all preying on T. urticae.
Osman, Bayoumy: Coccinellid predator Stethorus gilvifrons
Acta Phytopathologica et Entomologica Hungarica 46, 2011
284
Aphyt_46(2011)2_3tord-imp:Aphyt_46(2011)2_2_tord 11/17/11 4:09 PM Page 284

The handling time is the time predator spend in identifying, chasing, killing, eating,water ingestion, and other related time-consuming activities (Holling, 1965). It is a goodindicator of consumption rate and effectiveness of a predator because it reflects the cumu-lative effect of time taken during capturing, killing, subduing and digesting the prey (Veer-avel and Baskaran, 1997). Handling time of S. gilvifrons estimated in this study on T. urticaelarvae (0.31 h) is smaller than that of 0.46 and 0.83 h on T. urticae nymphs and adults.However, the highest attack rate was calculated for S. gilvifrons on T. urticae adults. Thismay be due to S. gilvifrons at high prey densities continued to feed even when satiated andcontinued to attack mites, initiate feeding, but then subsequently abandon the prey withminimal feeding of bodily parts. However, it takes a long time in dealing with each adultindividual of prey. This may be due to the bigger size and faster movement of adult preythan other immature stages or to the long time between the two successive meals. The han-dling time and the maximal number of consumed preys (T/Th) for S. gilvifrons on T. urticaenymphs in our study is similar to that reported for the same predator feeding on T. turkestaninymphs (Sohrabi and Shishehbor, 2007). The ratio between exposure time and handlingtime (T/Th) is an indicator of maximum theoretical predation, which may give an indica-tion of the number of predators that have to be introduced in the crop based on killing ef-ficiency to improve pest control. In this case, the maximum number of consumed preys(per day) by S. gilvifrons is 77.42 larvae of T. urticae. Based on Th values, the estimated the-oretical construct is higher than that measured empirically by the asymptotes of fitted re-lationships for all tested stages. This is because the theoretical number is calculated basedon the actual time of predator feeding activity, without consideration of non-feeding activ-ities (i.e. searching, water sap, rest), rather than the total time of experiment (T). The timeas prey and predator exposure (T) can be considered to be a function of: (1) time in non-feeding activities; and (2) time in feeding related activities (Holling, 1961). Therefore, thetheoretical number of prey consumed by a predator seems to be more valuable to predicatethe maximum daily consumption rate.
According to Berry et al. (1988) some problems are associated with estimating func-tional responses in the laboratory. Predators are not allow to leave a patch in search ofhigher densities where prey location is more efficient, and therefore predators consumemore prey at low density than they would probably do in nature. This may result in type IIfunctional responses when the underlying response is really type III (van Lenteren andBakker, 1976). However, such studies are useful in providing the first step to compare theefficiency of a natural enemy against different prey species or stages (Vantornhout, 2006)and to provide a valid means of comparing host-finding abilities of candidate natural ene-mies (Munyaneza and Obrycki, 1997).
Based on the inversely density-dependent predation for S. gilvifrons on different preystages, it seems to be more effective in low prey densities and this result has to be consid-ered in future inundative biological control programs. Hence, releasing the predator earlyin the beginning of growing season on low prey populations might be contributed towardmaximizing the efficiency of pest control. The results show that S. gilvifrons females in-crease predation rate as function of T. urticae larva densities. This fact, together with lowerhandling time on prey larvae indicates that S. gilvifrons could be consumed high numbers
Osman, Bayoumy: Coccinellid predator Stethorus gilvifrons
Acta Phytopathologica et Entomologica Hungarica 46, 2011
285
Aphyt_46(2011)2_3tord-imp:Aphyt_46(2011)2_2_tord 11/17/11 4:09 PM Page 285

of larvae per day than other stages. Therefore, the best time to release the predator in thefield is when most of population in the larval stage. Therefore, the results obtained in thisstudy can be useful for the further study of the development of management tactics forcontrol of T. urtica. The relevance of the laboratory experiments on prey consumption is,however, limited because densities used in the laboratory may be unrealistically high. In ad-dition, the time used by S. gilvifrons in searching for food may reduce its prey consump-tion capacity under natural conditions. Thus, the contribution of S. gilvifrons to control T.urticae in fields requires further studies on the influence of the predator age, large search-ing arenas, spatial complexity, number of predators in the search unit, and other environ-mental factors.
AcknowledgementsWe would like to thank Professor S. A. Juliano (Department of Biological Science, Illinois State Univer-
sity, Illinois, USA) for his valuable suggestions and statistical advices. Thanks Professor A. A. Osman (Economicentomology, Faculty of Agriculture, Zoology Department, Menofia University, Egypt) and F. Kozár (Plant Pro-tection Institute, Zoology Department, Hungarian Academy of Sciences, Budapest) for their helpful insights andcritical reviews of the manuscript.
LiteraturesAfshari, G. A. (1999): A survey on the ladybirds belong to genus Stethorus and study on the biology, prey
consumption and population dynamics of Stethorus gilvifrons in sugarcane farms in Khuzestan, Iran. M.Sc. Thesis, Ahvaz, Iran, 179 p.
Alatawi, F. J., Opit, G. P., Margolies, D. C. and Nechols, J. R. (2005): Within-plant distribution of two-spottedspider mites (Acari: Tetranychidae) on impatiens: Development of a presence–absence sampling. PlantJournal of Economic Entomology 98, 1040–1047.
Allahyari, H., Azmayeshfard, P.� and� Nozari, J.� (2004): Effects of host on functional response of offspring intwo populations of Trissolcus grandis on the sunn pest. – Journal of Applied Entomology 128, 39–43.
Atlihan, R., Bora, M. K., Alper, Y. and Hayrettin, O. (2010): Functional response of the coccinellid predatorAdalia fasciatopunctata revelierei to walnut aphid (Callaphis juglandis). Phytoparasitica 38, 23–29.
Bailey, P. and Caon, G. (1986): Predation on two spotted mite, Tetranychus urticaeKoch (Acarina: Tetranychidae)by Haplothrips victoriensis Bagnall (Thysanoptera: Phlaeothripidae) and Stethorus nigripes Kapur(Coleoptera: Coccinellidae) on seed Lucerne crops in South Australia. Australian Journal of Zoology 34,515–525.
Bayoumy, M. H. M., Abd El-Kareim, A. I., Abdel-Salam, A. H., Abdel-Baky, N. F. and Schopf. A. (2009):Functional responses of Encarsia perniciosi and Encarsia citrina to Quadraspidiotus perniciosus inresponse to temperature. Journal of Agricultural Sciences 34, 9673–9687.
Berry, J. S., Holtzer, T. O., Innis, G. S. and Logan, J. A. (1988): Simple order of prey preference technique formodelling the predator functional response. Experimental Applied of Acarology 5, 207–224.
Britto, E. P. J., Gondim Jr., M. G. C., Torres, J. B., Fiaboe, K. K. M., Moraes, G. J. and Knapp, M. (2009):Predation and reproductive output of the ladybird beetle Stethorus tridens preying on tomato red spider miteTetranychus evansi. BioControl 54, 363–398 (doi: 10.1007/s10526-008-9178-5).
Byeon, Y. W., Tudam, M., Kim, J. H. and Choi, M. Y. (2011): Functional responses of aphid parasitoids, Aphi diuscolemani (Hymenoptera: Braconidae) and Aphelinus asychis (Hymenoptera: Aphelinidae). BiocontrolSciences and Technology 21, 57–70.
Osman, Bayoumy: Coccinellid predator Stethorus gilvifrons
Acta Phytopathologica et Entomologica Hungarica 46, 2011
286
Aphyt_46(2011)2_3tord-imp:Aphyt_46(2011)2_2_tord 11/17/11 4:09 PM Page 286

Congdon, J. D., Dunham, A. E. and van Loben Sels, R. C. (1993): Delayed sexual maturity and demographics ofblanding’s turtles (Emydoidea blandingii): implications for conservation and management of long-livedorganisms. Conservation Biology 7, 826–833.
De Clercq, P., Mohaghegh, J. and Tirry, L. (2000): Effect of host plant on the functional response of the predatorPodisus nigrispinus (Heteroptera: Pentatomidae). Biological Control 18, 65–70.
Fathipour, Y., Hosseini, A., Talebi, A. and Moharramipour, S. (2006): Functional response and mutual interferenceof Diaeretiella rapae (Hymenoptera: Aphidiidae) on Brevicoryne brassicae (Homoptera: Aphididae).Entomological Fennica 17, 90–97.
Fiaboe, K. K. M., Gondim, M. G. C., de Moraes, G. J., Ogol, C. K. P. O. and Knapp, M. (2007): Bionomics ofthe acarophagous ladybird beetle Stethorus tridens fed Tetranychus evansi. Journal of Applied Entomology131, 355–361.
Gotoh, T., Nozawa, M. and Yamaguchi, K. (2004): Prey consumption and functional response of threeacarophagous species to eggs of the two-spotted spider mite in the laboratory. Applied Entomology andZoology 39, 97–105.
Haji-Zadeh, J., Kamali, G. K. and Assadi, H. B. (1993): Investigations on the functional response and populationsfluctuations of Stethorus gilvifrons on red spider mite, Panonychus ulmi (Koch) in Karaj vicinity [Iran].Applied Entomology and Phytopathology 61, 32–34.
Hassel, M. P., Lawton, J. H. and Beddington, J. R. (1977): Sigmoid functional response by invertebrate predatorsand parasitoids. Journal of Animal Ecology 46, 249–262.
Hassell, M. P. (2000): The Spatial and Temporal Dynamics of Host Parasitoid Interactions. Oxford Series inEcology and Evolution. Oxford University Press, London, UK, 198 p.
Hazzard, R. V. and Ferro, D. N. (1991): Feeding responses of adults Coleomegilla maculata (Coleoptera:Coccinellidae) to eggs of Colorado potato beetle (Coleoptera: Chrysomelidae) and green peach aphids(Homoptera: Aphididae). Environmental Entomology 20, 644–651.
Holling, C. S. (1959): Some characteristics of simple types of predation and parasitism. Canadian Entomology91, 385–398.
Holling, C. S. (1961): Principles of insect predation. Annual Review of Entomology 6, 163–183.Holling, C. S. (1965): Functional response of predators to prey density and its role in mimicry and population
regulation. Memoirs of Entomological Society of Canada 45, 3–60.Houck, M. A. (1991): Time and resource partitioning in Stethorus punctum (Coleoptera: Coccinellidae).
Environmental Entomology 20, 494–497.Hu, Y. S., Wang, Z. M., Ning, C. L., Pi, Z. Q. and Gao, G. Q. (1989): The functional response of Harmonia (Leis)
axyridis to their prey of Cinara sp. Natural Enemies of Insects 11, 164–168.Huffaker, C. B., van de Vrie, M. and McMurtry, J. A. (1970): Ecology of tetranychid mites and their natural
enemies: a review. II. Tetranychid populations and their possible control by predators: an evaluation.Hilgardia 40, 391–458.
Isikber, A. A. (2005): Functional response of two coccinellid predators, Scymnus levaillanti and Cycloneda san-uinea, to the cotton aphid, Aphis gossypii. Turkish Journal of Agriculture Forest 29, 347–355.
Juliano, S. A. (2001): Non-linear curve fitting: Predation and functional response curves. In: S. M. Scheiner andJ. Gurevitch (eds): Design and Analysis of Ecological Experiments. 2nd edition. Chapman and Hall, NewYork, pp. 178–196.
Koch, R. L., Hutchison, W. D., Venette, R. C. and Heimpel, G. E. (2003): Susceptibility of immature monarchbutterfly, Danaus plexippus (Lepidoptera: Nymphalidae: Danainae), to predation by Harmonia axyridis(Coleoptera: Coccinellidae). Biological Control 28, 265–270.
Mills, N. J. and Lacan, I. (2004): Ratio-dependence in the functional response of insect parasitoids: evidencefrom Trichogramma minutum foraging for eggs in small host patches. Ecological Entomology 29, 208–216.
Mohaghegh, J., De Clercq, P. and Tirry, L. (2001): Functional response of the predators Podisus maculiventris (say)and Podisus nigrispinus (Dallas) (Heteroptera: Pentatomidae) to the beet armyworm, Spodoptera exigua(Hubner) (Lepidoptera: Noctuidae): effect of temperature. Journal of Applied Entomology 125, 131–134.
Osman, Bayoumy: Coccinellid predator Stethorus gilvifrons
Acta Phytopathologica et Entomologica Hungarica 46, 2011
287
Aphyt_46(2011)2_3tord-imp:Aphyt_46(2011)2_2_tord 11/17/11 4:09 PM Page 287

Mori, K., Nozawa, M., Arai, K. and Gotoh, T. (2005): Life-history traits of the acaraphagous lady beetle, Stetho-rus japonicus at three constant temperatures. BioControl 50, 35–51.
Munyaneza, J. and Obrycki, J. J. (1997): Functional response of Coleomegilla maculata (Coleoptera:Coccinellidae) to Colorado potato beetle eggs (Coleoptera: Chrysomelidae). Biological Control 8, 215–224.
Nwilene, F. E. and Nachman, G. (1996): Functional responses of Iphiseius degenerans and Neoseiulus teke (Acari:Phytoseiidae) to changes in the density of the cassava green mite, Mononychellus tanajoa (Acari:Tetranychidae). Experimental Applied Acarology 20, 259–271.
Obrycki, J. J. (1998): Predaceous coccinellidae in biological control. Annual Review of Entomology 43, 295–321.Ofuya, T. I. and Akingbohungbe, A. E. (1988): Functional and numerical responses of Cheilomenes lunata
(Fabricius) (Coleoptera: Coccinellidae) feeding on the cowpea aphid, Aphis craccivora Koch (Homoptera:Aphididae). Insect Science Application, 9, 543–546.
Overmeer, W. P. J. (1985): Rearing and handling. In: W. Helle and M. W. Sabelis (eds): Spider Mites, TheirBiology, Natural Enemies and Control. Elsevier, Amsterdam, Vol. 1b, pp. 162–170.
Roger, D. (1972): Random search and insect population models. Journal of Animal Ecology 41, 369–383.Roy, M., Brodeur, J. and Cloutier, C. (2003): Effect of temperature on intrinsic rates of natural increase (rm) of a
coccinellid and its spider mite prey. BioControl 48, 57–72Sarmento, R. A., Pallini, A., Venzon, M., Souza, F. F., Molina-Rugama, A. J. and Oliveira, C. L. (2007): Functional
response of the predator Eriopis connexa (Coleoptera: Coccinellidae) to different prey types. BrazilianArchives of Biology and Technology 50, 121–126.
SAS Institute (2000): SAS/STAT user’s guide. Version 9.1 edition. SAS Institute Cary, NC.Sohrabi, F. and Shishehbor, P. (2007): Functional and numerical responses of Stethorus gilvifronsMuslant feeding
on strawberry spider mite, Tetranychus turkestani Ugarov and Nikolski. Pakistan Journal of BiologicalScience 10, 4563–4566.
Solomon, M. (1949): The natural control of animal populations. Journal of Animal Ecology 41, 369–383.Tanigoshi, L. K. and McMurtry, J. A. (1977): The dynamics of predation of Stethorus picipes (Coloptera:
Coccinellidae) and Typhlodromus floridanus on the prey Oligonychus punicae (Acarina: Phytoseiidae,Tetranychidae). 1. Comparative life history and life table studies. Hilgardia 45, 237–261.
Trexler, J. C., Charles, E. M. and Travis, J. (1988): How can the functional response best be determined? Oecologia76, 206–214.
Ullah, I. (2000): Aspects of the biology of the ladybird beetle Stethorus vegans (Blackburn) (Coleoptera:Coccinellidae). Doctoral thesis, Centre For Horticulture and Plant Sciences, University of Western Sydney,Hawkesbury, Richmond, NSW Australia, xvii, 183 p.
Uygun, N. and Atlihan, R. (2000): The effect of temperature on development and fecundity of Scymnus levaillanti.BioControl 45, 453–462.
van Lenteren, J. C. and Bakker, K. (1976): Functional responses in invertebrates. Netherland Journal of Zoology26, 567–572.
Vantornhout, I. (2006): Biology and ecology of the predatory mite Iphiseius degenerans (Berlese) (Acari:Phytoseiidae). Ph.D. thesis, Ghent University, Ghent, Belgium, 207 p.
Veeravel, R. and Baskaran, P. (1997): Functional and numerical responses of Coccinella transversalis andCheilomenes sexmaculata Fabr. feeding on the melon aphid, Aphis gossypii Glov. Insect Science and ItsApplication 17, 335–339.
Wang, B. and Ferro, D. N. (1998): Functional responses of Trichogramma ostriniae (Hym., Trichogrammatidae)to Ostrinia nubilalis (Lep., Pyralidae) under laboratory and field conditions. Environmental Entomology27, 752–758.
Zhang, Z. Q. (2003): Mite of Greenhouses. Identification, Biology, and Control. CABI Publishing, 244 p.
Osman, Bayoumy: Coccinellid predator Stethorus gilvifrons
Acta Phytopathologica et Entomologica Hungarica 46, 2011
288
Aphyt_46(2011)2_3tord-imp:Aphyt_46(2011)2_2_tord 11/17/11 4:09 PM Page 288




















