
Toxicity, inheritance of fenpyroximate resistance, and detoxification-enzyme levels in a...
6
Toxicity, inheritance of fenpyroximate resistance, and detoxification-enzyme levels in a laboratory-selected fenpyroximate-resistant strain of Tetranychus urticae Koch (Acari: Tetranychidae) Recep Ay * , Fatma Ebru Kara Süleyman Demirel University, Faculty of Agriculture, Plant Protection Department, 32260 Çünür, Isparta, Turkey article info Article history: Received 19 May 2010 Received in revised form 6 October 2010 Accepted 20 November 2010 Keywords: Tetranychus urticae Fenpyroximate Detoxifying enzymes Inheritance Resistance abstract A strain (BEYO 2) of Tetranychus urticae was selected with fenpyroximate for 14 selections. The resulting strain (named FPY 14) became resistant to fenpyroximate. The present study examines the inheritance of fenpyroximate resistance, the toxicity of some insecticides and acaricides, detoxification enzymes [esterase, glutathione S-transferase (GST), and monooxygenase (P450)], and the synergistic ratios of certain synergists [piperonyl butoxide (PBO), S-benzyl-O,O-diisopropyl phosphorothioate (IBP), and tri- phenyl phosphate (TPP)] in the FPY 14 strain of T. urticae. A spray tower-Petri dish method was used in the selection and toxicity studies. The level of fenpyroximate resistance was 64.43-fold higher in the FPY 14 strain and 1.06-fold higher in the BEYO 2 strain compared to the GSS (German susceptible strain) strain. The FPY 14 strain was 7.80-, 6.90-, 6.43-, 4.78- and 2.78-fold more resistant to abamectin, chlorpyrifos, propargite, clofentezine and amitraz, respectively. Fenpyroximate resistance is inherited as an incompletely dominant trait with no sex linkage. None of the synergists showed a significant synergistic effect. In the FPY 14 strain, the activities of esterase, GST and P450 enzymes were 1.92-,1.06- and 3.96-fold higher, respectively, when compared to the susceptible GSS strain. The P450 and esterase enzymes might play a role in the mechanism of resistance to fenpyroximate. Ó 2010 Elsevier Ltd. All rights reserved. 1. Introduction Phytophagous mites are serious pests that can cause significant yield losses in many important agricultural crops, including fruits, cotton, vegetables and ornamentals (Van Leeuwen et al., 2007). Considering its worldwide distribution and its effects on a large number of host plants, Tetranychus urticae is one of the most significant pests in the world. One of the major obstacles to the control of T. urticae is its ability to rapidly develop resistance to many important acaricides, even after only a few applications (Stumpf et al., 2001). T. urticae develops resistances to many different classes of acaricides when they are heavily used, resulting in a huge demand for acaricides with novel modes of action (Suh et al., 2006). A number of detoxification enzymes have been shown to be involved in the development of resistance by insects and mites (Yu, 2008). These detoxification enzymes include cyto- chrome P450 monooxygenases, esterases and glutathione S- transferases (GSTs) (Tsagkarakou et al., 2002; Yang et al., 2002; Kim et al., 2004; Van Leeuwen et al., 2005; Kang et al., 2006; Van Leeuwen and Tirry, 2007). Fenpyroximate belongs to the mitochondrial electron transport inhibitor (METI) group of acaricides. METI acaricides (tebufenpyrad, fenpyroximate, pyradaben and fenazaquin) are potent xenobiotics that mainly act by inhibiting complex I (NADH: ubiquinone oxidoreductase) of the mitochondrial respiratory chain (Holingworth and Ahamadsahib, 1995). Due to the resistances developed by pests as a result of their frequent use, METIs have largely lost their capacity to control pests. However, they are still being used on many crops worldwide to control T. urticae (Van Leeuwen et al., 2009). The present study examines selection for fenpyroximate resis- tance in T. urticae, a species that causes widespread damage to greenhouse crops and orchards in Turkey. The study also analyses changes in the levels of detoxification enzymes in fenpyroximate- resistant strains, the effects of three different synergists [Piperonyl butoxide (PBO), triphenyl phosphate (TPP) and S-benzyl-O,O- diisopropyl phosphorothioate (IBP)] in combination with fenpyr- oximate, the effects of other pesticides on resistant strains and the mode of inheritance of fenpyroximate resistance. * Corresponding author. Tel.: þ902462114852; fax: þ902462114885. E-mail address: [email protected] (R. Ay). Contents lists available at ScienceDirect Crop Protection journal homepage: www.elsevier.com/locate/cropro 0261-2194/$ e see front matter Ó 2010 Elsevier Ltd. All rights reserved. doi:10.1016/j.cropro.2010.11.012 Crop Protection 30 (2011) 605e610
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Toxicity, inheritance of fenpyroximate resistance, and detoxification-enzyme levels in a...

lable at ScienceDirect
Crop Protection 30 (2011) 605e610
Contents lists avai
Crop Protection
journal homepage: www.elsevier .com/locate/cropro
Toxicity, inheritance of fenpyroximate resistance, and detoxification-enzymelevels in a laboratory-selected fenpyroximate-resistant strain of Tetranychusurticae Koch (Acari: Tetranychidae)
Recep Ay*, Fatma Ebru KaraSüleyman Demirel University, Faculty of Agriculture, Plant Protection Department, 32260 Çünür, Isparta, Turkey
a r t i c l e i n f o
Article history:Received 19 May 2010Received in revised form6 October 2010Accepted 20 November 2010
Keywords:Tetranychus urticaeFenpyroximateDetoxifying enzymesInheritanceResistance
* Corresponding author. Tel.: þ902462114852; fax:E-mail address: [email protected] (R. Ay).
0261-2194/$ e see front matter � 2010 Elsevier Ltd.doi:10.1016/j.cropro.2010.11.012
a b s t r a c t
A strain (BEYO 2) of Tetranychus urticae was selected with fenpyroximate for 14 selections. The resultingstrain (named FPY 14) became resistant to fenpyroximate. The present study examines the inheritance offenpyroximate resistance, the toxicity of some insecticides and acaricides, detoxification enzymes[esterase, glutathione S-transferase (GST), and monooxygenase (P450)], and the synergistic ratios ofcertain synergists [piperonyl butoxide (PBO), S-benzyl-O,O-diisopropyl phosphorothioate (IBP), and tri-phenyl phosphate (TPP)] in the FPY 14 strain of T. urticae. A spray tower-Petri dish method was used inthe selection and toxicity studies. The level of fenpyroximate resistance was 64.43-fold higher in the FPY14 strain and 1.06-fold higher in the BEYO 2 strain compared to the GSS (German susceptible strain)strain. The FPY 14 strain was 7.80-, 6.90-, 6.43-, 4.78- and 2.78-fold more resistant to abamectin,chlorpyrifos, propargite, clofentezine and amitraz, respectively. Fenpyroximate resistance is inherited asan incompletely dominant trait with no sex linkage. None of the synergists showed a significantsynergistic effect. In the FPY 14 strain, the activities of esterase, GST and P450 enzymes were 1.92-, 1.06-and 3.96-fold higher, respectively, when compared to the susceptible GSS strain. The P450 and esteraseenzymes might play a role in the mechanism of resistance to fenpyroximate.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
Phytophagous mites are serious pests that can cause significantyield losses in many important agricultural crops, including fruits,cotton, vegetables and ornamentals (Van Leeuwen et al., 2007).Considering its worldwide distribution and its effects on a largenumber of host plants, Tetranychus urticae is one of the mostsignificant pests in the world. One of the major obstacles to thecontrol of T. urticae is its ability to rapidly develop resistance tomany important acaricides, even after only a few applications(Stumpf et al., 2001). T. urticae develops resistances to manydifferent classes of acaricides when they are heavily used, resultingin a huge demand for acaricides with novel modes of action (Suhet al., 2006). A number of detoxification enzymes have beenshown to be involved in the development of resistance by insectsand mites (Yu, 2008). These detoxification enzymes include cyto-chrome P450 monooxygenases, esterases and glutathione S-transferases (GSTs) (Tsagkarakou et al., 2002; Yang et al., 2002; Kim
þ902462114885.
All rights reserved.
et al., 2004; Van Leeuwen et al., 2005; Kang et al., 2006; VanLeeuwen and Tirry, 2007).
Fenpyroximate belongs to the mitochondrial electron transportinhibitor (METI) group of acaricides. METI acaricides (tebufenpyrad,fenpyroximate, pyradaben and fenazaquin) are potent xenobioticsthat mainly act by inhibiting complex I (NADH: ubiquinoneoxidoreductase) of the mitochondrial respiratory chain(Holingworth and Ahamadsahib, 1995). Due to the resistancesdeveloped by pests as a result of their frequent use, METIs havelargely lost their capacity to control pests. However, they are stillbeing used on many crops worldwide to control T. urticae (VanLeeuwen et al., 2009).
The present study examines selection for fenpyroximate resis-tance in T. urticae, a species that causes widespread damage togreenhouse crops and orchards in Turkey. The study also analyseschanges in the levels of detoxification enzymes in fenpyroximate-resistant strains, the effects of three different synergists [Piperonylbutoxide (PBO), triphenyl phosphate (TPP) and S-benzyl-O,O-diisopropyl phosphorothioate (IBP)] in combination with fenpyr-oximate, the effects of other pesticides on resistant strains and themode of inheritance of fenpyroximate resistance.

R. Ay, F.E. Kara / Crop Protection 30 (2011) 605e610606
2. Materials and methods
2.1. Strains, selections and rearing
The original strain of T. urticae (termed BEYO 2) was establishedusing mites collected from a commercial bean (Phaseolus vulgarisL.) greenhouse in Antalya Province in June 2003. The BEYO 2 strainwas grown from June 2003 to October 2006 without any pesticideexposure. The strain developed resistance to fenpyroximate after 14rounds of selection with fenpyroximate from October 2006 toAugust 2007. The resulting strain was named “FPY 14”. A suscep-tible strain (GSS) was obtained from Rothamsted ExperimentalStation, Harpenden, England, in 2001 and then reared underlaboratory conditions. All T. urticae strains were continuouslygrown on pinto-bean plants, P. vulgaris L., under laboratory condi-tions [26 � 2 �C, 60% � 5% relative humidity (RH) with a 16-hphotoperiod].
2.2. Chemicals
The materials used for the main study consisted of fenpyrox-imate (Meteor� SC 50 g/l), which is a mitochondrial electrontransport inhibitor (METI) acaricide, and three synergists: piper-onyl butoxide (PBO), S-benzyl-O,O-diisopropyl phosphorothioate(IBP) and triphenyl phosphate (TPP). IBP and TPP are esteraseenzyme inhibitors (Kim et al., 2004; Kang et al., 2006; Wang andWu, 2007), while PBO is an esterase and P450 inhibitor (Stumpfand Nauen, 2002; Young et al., 2005; Kim et al., 2006; VanLeeuwen and Tirry, 2007). Chemicals containing the formamidineamitraz (Kortraz 20 EC 200 g/l), the mite-growth regulator clo-fentezine (Apollo SC 500 g/l), the sulphite ester propargite (KomiteEC 588 g/l), the pyrethroid bifenthrin (Talstar EC 100 g/l), theorganophosphate chlorpyrifos (Dursban 4 EC 480 g/l), or the aver-mectin abamectin (Agrimec 18 g/l) were used in subsequentresistance trials conducted on the resistant strain.
2.3. Bioassays
These tests were conducted based on the method described byAy (2005). The acaricide used in the present study was a commer-cial formulation of fenpyroximate. A prepared suspension of fen-pyroximate was sprayed onto the internal surfaces, both lids andbases, of 60-mm-diameter plastic Petri dishes and allowed to dryfor 30 min. For each application, 1 ml of the fenpyroximatesuspension was sprayed onto each base and lid pair using a Potterspray tower (Burckard Auto-Load, Rickmansworth, Herts, UK) at100 kPa. Adult female mites (w30) were transferred into each dishusing a brush, and the dishes were closed and sealed with Parafilmto prevent the spider mites from escaping. During the first 24 hafter treatment, the mites in the dish were kept at 26 � 2 �C,60 � 5% RH with a 16-h photoperiod. Survival of individual miteswas determined by touching each mite with a fine brush. Mitesunable to walk at least a distance equivalent to their body lengthwere considered dead. Mortality tests were performed before eachexperiment to determine the concentration range that causedapproximately 10e95% mortality. Each experiment was conductedin triplicate using seven different concentrations (plus a distilledwater-only control). Pooled data were analysed using a probitanalysis (POLO PC) (LeOra software, 1994) and the LC50, LC60 andLC90 with 95% CL were estimated. LC50 values of the selected strainwere compared to those of the susceptible strain (GSS). A resistanceratio (RR) was calculated according to the following formula:RR ¼ LC50 value of the resistant strain/LC50 value of the GSS strain.
2.4. Selection for resistance
Females of the original strain (BEYO 2) were selected for fen-pyroximate resistance under laboratory conditions from October2006 to August 2007. Selection experiments were performedaccording to the method proposed by Yang et al. (2002), with somemodifications. At least 400 adult female mites of the BEYO 2 strainwere transferred into Petri dishes (40 mites/Petri dish) and treatedwith fenpyroximate concentrations equal to the LC60 for that cycle.After 24 h, surviving mites were transferred back to plants, and thepopulations were allowed to regenerate. The selection cycle wasrepeated two or three generations (approximately 15e20 days)later to ensure sufficient increase in the population size. A bioassayusing the same acaricide used for selection was conducted peri-odically on each mite strain when the number of surviving miteschanged in the Petri dishes used for selection. The new LC60 wasdetermined and applied to each new population, thereby exertingselection pressure.
2.5. Toxicity of other pesticides to the fenpyroximate-selected strain
Additional pesticides were evaluated using the fenpyroximate-resistant (FPY 14) and susceptible (GSS) strains of T. urticae. Thepesticides used in this experiment were amitraz, abamectin, clo-fentezine, propargite, bifenthrin and chlorpyrifos. All of the pesti-cides used were commercially available in Turkey. The bioassaymethod used for all pesticides was the same as that describedpreviously for the toxicity test. Themortality data for each pesticidefor the GSS and FPY 14 strains were subjected to probit analysis(POLO PC) (LeOra, 1994). Resistance ratios were calculated bydividing the LC50 of each resistant strain by the LC50 of thesusceptible strain.
2.6. Crossing experiment
By placing 15 teleiochrysalis-stage females of one strain and 30adult males of the other strain on the upper side of a primary beanleaf placed on wet wool in a Petri dish, the GSS and FPY 14 strainswere reciprocally crossed to produce hybrid F1 females so as toestimate the dominance of resistance. Directly after moulting,diploid females were fertilised by the haploid males and began tolay eggs one day later. Males and females were removed five dayslater. If mating was successful, the haploidediploid mating systemresulted in F1 females and males. The F1 females were then trans-ferred to clean bean plants, and a bioassay was conducted whenthey reached maturity. The bioassay method was the same as thatdescribed previously for the toxicity test. The experiment wasconducted in triplicate using seven serially diluted concentrations(plus a distilled water-only control) covering the 10e90% mortalityrange. The following formula was used to estimated the degree ofdominance (D) of the resistant trait in the F1 females from bothreciprocal crosses: D ¼ (2X2 � X1 � X3)/(X1 � X3), where X1 is thelogarithm of the LC50 of the resistant strain, X2 is the logarithm ofthe LC50 of the F1 females and X3 is the logarithm of the LC50 of thesusceptible strain (Stone, 1968). This formula gives a value of �1 ifresistance is fully recessive, a value of 0 if there is no dominance,and a value of þ1 if resistance is fully dominant.
2.7. Synergism test
The effects of fenpyroximate and synergists were tested usingthe methods developed by Kim et al. (2004). Mixed-functionoxidase (MFO), the esterase inhibitor piperonyl butoxide (PBO) andtwo other esterase inhibitors [S-benzyl-O,O-diisopropyl phosphor-othioate (IBP) and triphenyl phosphate (TPP)] were used to inhibit

Table 1Selection for resistance to fenpyroximate in a strain of T. urticae; estimation of LC50 (mg a.i./l distilled water) and resistance ratio.
Strains Na Slope � SE LC50 (mg a.i./l)(95% CIb)
LC60 (mg a.i./l)(95% CIb)
LC90 (mg a.i./l)(95% CIb)
RR LC50c RR LC90
c
GSS 720 1.35 � 0.10 3.312.71e3.99
5.104.23e6.21
29.5021.60e44.19
e e
BEYO 2 715 1.38 � 0.11 3.512.88e4.23
5.434.46e6.51
29.9122.11e44.11
1.06 1.01
Sel 1 715 1.42 � 0.11 3.653.01e4.38
5.504.59e6.64
29.1527.49e42.31
1.10 0.99
Sel 2 714 1.29 � 0.10 4.283.15e5.64
6.745.10e9.03
42.4827.49e79.63
1.29 1.44
Sel 3 724 1.14 � 0.87 22.7816.30e32.61
37.9926.93e58.51
302.74162.82e796.89
6.88 10.26
Sel 4 639 1.14 � 0.10 23.6518.72e29.82
39.5032.45e49.09
317.03205.74e571.16
7.15 10.75
Sel 5 722 1.17 � 0.10 25.9020.28e32.69
42.7233.84e54.95
325.65216.91e564.28
7.82 11.04
Sel 6 723 1.24 � 0.11 49.7239.96e62.25
79.6763.58e103.20
539.76379.83e847.67
15.02 18.30
Sel 7 725 1.29 � 0.11 70.1756.56e87.57
110.2788.32e142.17
690.52491.44e1068.54
21.20 23.41
Sel 8 716 1.27 � 0.10 70.9557.50e87.79
112.3990.74e143.03
726.93495.96e1208.32
21.44 24.64
Sel 9 721 1.56 � 0.15 103.8782.52e128.29
151.07122.24e188.07
690.76501.63e1066.59
31.38 23.42
Sel 10 722 1.26 � 0.12 156.19122.35e200.27
247.76193.50e329.38
1611.581040.57e2964.52
47.19 54.63
Sel 11 721 1.30 � 0.12 172.02135.19e219.03
269.57211.95e354.08
1668.761099.30e2974.04
51.97 56.57
Sel 12 719 1.30 � 0.12 177.15140.92e222.69
277.05220.45e358.06
1701.141145.05e2912.92
53.52 57.67
Sel 13 720 1.41 � 0.12 195.55141.11e268.14
295.84216.89e421.28
1587.69974.90e3356.67
59.08 53.82
FPY 14 720 1.45 � 0.12 213.26142.84e316.76
319.10218.45e504.16
1637.86916.86e4441.48
64.43 55.52
a Total number of mites used.b Confidence interval.c Resistance ratio ¼ LC50 or LC90 value of resistance strain/LC50 or LC90 value of the GSS strain.
R. Ay, F.E. Kara / Crop Protection 30 (2011) 605e610 607
detoxification enzymes (Stumpf and Nauen, 2002; Young et al.,2005; Kim et al., 2004, 2006; Kang et al., 2006; Van Leeuwen andTirry, 2007; Wang and Wu, 2007). Synergists were dissolved inacetone and distilled water (1:1), and 1 ml of the suspension wassprayed on the base and lid of Petri dishes 30 min prior to acaricideapplication as in the toxicity test. Only acetone þ distilled waterwas applied to the control. Synergist solutions were prepared at thefollowing concentrations (mg l�1): PBO (500), IBP (400) and TPP(125). Synergistic ratios (SR) were calculated using the followingformula: SR ¼ LC50 of synergist-free fenpyroximate/LC50 of fen-pyroximate with synergist.
2.8. Enzymatic assay
Esterase assays using a-naphthyl acetate were performedaccording to the method of Stumpf and Nauen (2002). Bulkhomogenates of 20 adult females [prepared in 100 ml ice-cold 0.1 Msodium phosphate buffer, pH 7.5, containing 0.1% (w v�1) Triton X-100] were centrifuged at 10,000g. The supernatant was diluted 10-fold and used as the enzyme source. Aliquots (25 ml aliquots; 0.5mite equivalent) were added to the wells of a 96-well microplatecontaining 25 ml of 0.2 M sodium phosphate buffer, pH 6.0. Wellscontaining buffer only served as controls for the non-enzymaticreaction. The assay was started by adding 200 ml of substratesolution to each well, reaching a final volume of 250 ml. Thesubstrate solution consisted of 30 mg of fast Blue RR salt dissolvedin 50 ml of sodium phosphate buffer, pH 6.0, and 500 ml of 100 mM1-naphthyl acetate in acetone. Esterase activity was measuredcontinuously for 10 min at 450 nm and 25 �C in a Versamax kineticmicroplate reader (Molecular Devices) and was analysed usingSoftmax PRO software to fit kinetics plots by linear regression.
Glutathione S-transferase (GST) activity was assayed accordingto the method of Stumpf and Nauen (2002). GST activity wasdetermined using 1-chloro-2,4-dinitrobenzene and reducedglutathione (GSH) as substrates. Fifty adult females were homo-genised in 500 ml TriseHCL (0.05 M, pH 7.5) and centrifuged at10,000g for 5 min. The supernatant was collected for use in theenzymatic reactions. The total reaction volume in each microplatewell was 300 ml, consisting of 100 ml each of supernatant, CDNBcontaining 0.1% (v v�1) ethanol, and GSH in TriseHCL (0.05 M, pH7.5), giving final concentrations of 0.4 mM CDNB and 4 mM GSH.The change in absorbance was measured continuously for 5 min at340 nm and 25 �C using a Versamax kinetic microplate reader(Molecular Devices). The non-enzymatic reaction of CDNB withGSH in the absences of homogenate served as a control.
The O-demethoxylation of p-nitroanisole (PNOD) by cyto-chrome P450 monooxygenase was measured using the proceduresdeveloped by Rose et al. (1995). For the PNOD assay, 50 adultfemales were homogenised on ice in 200 ml of homogenisationbuffer (0.05 M TriseHCl þ 1.15% KCl þ 1 mM EDTA, pH 7.7). Thehomogenate was centrifuged for 20 min at 20,000g at 4 �C. To eachwell of a 96-well microplate, 100 ml of 2 mM p-nitroanisole solutionand 90 ml of supernatant were added. Themicroplatewas incubatedfor 5 min at 30 �C, and the reaction was initiated by adding of 10 mlof 9.6 mM NADPH. The non-enzymatic reaction of PNOD withNADPH in the absence of homogenate served as a control. Theabsorbance was read using a Versamax kinetic microplate reader(Molecular Devices) at 405 nm and 30 �C for 15 min.
The activity of all enzymes was analysed using Softmax PROsoftware and defined as mOD min�1 mg�1 proteins. The data wereanalysed via the General Linear Model (GLM) procedure of SAS(1999). Strains were used as the independent variable in the
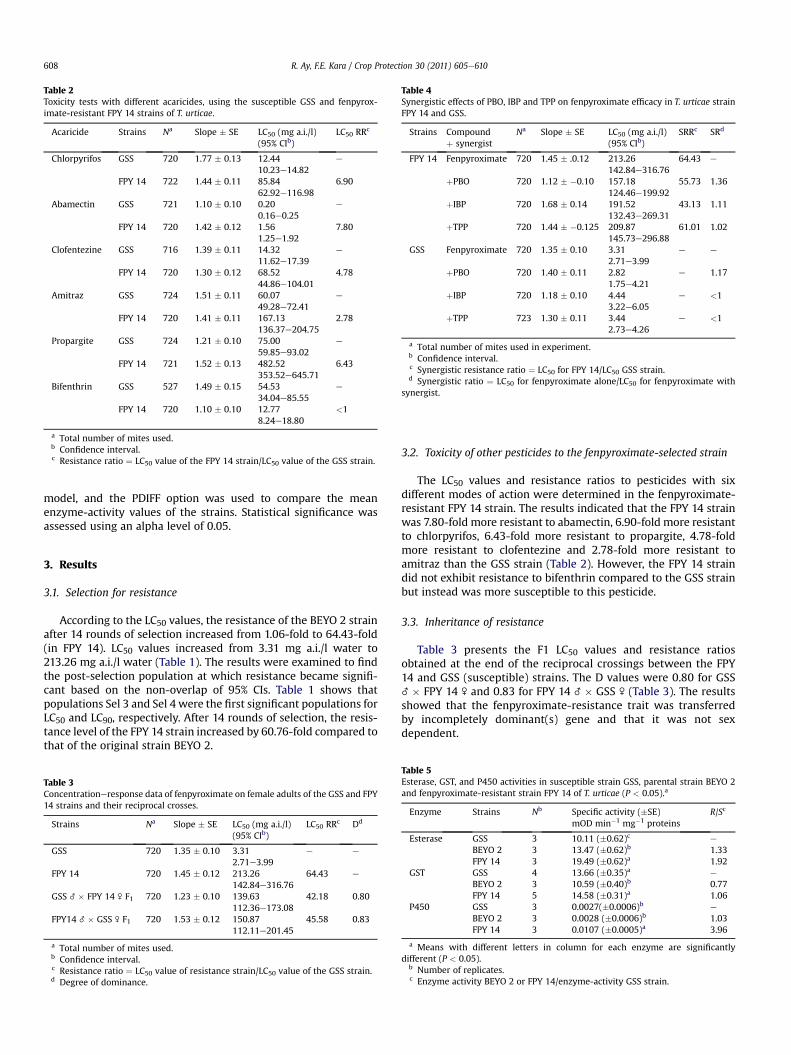
Table 2Toxicity tests with different acaricides, using the susceptible GSS and fenpyrox-imate-resistant FPY 14 strains of T. urticae.
Acaricide Strains Na Slope � SE LC50 (mg a.i./l)(95% CIb)
LC50 RRc
Chlorpyrifos GSS 720 1.77 � 0.13 12.4410.23e14.82
e
FPY 14 722 1.44 � 0.11 85.8462.92e116.98
6.90
Abamectin GSS 721 1.10 � 0.10 0.200.16e0.25
e
FPY 14 720 1.42 � 0.12 1.561.25e1.92
7.80
Clofentezine GSS 716 1.39 � 0.11 14.3211.62e17.39
e
FPY 14 720 1.30 � 0.12 68.5244.86e104.01
4.78
Amitraz GSS 724 1.51 � 0.11 60.0749.28e72.41
e
FPY 14 720 1.41 � 0.11 167.13136.37e204.75
2.78
Propargite GSS 724 1.21 � 0.10 75.0059.85e93.02
e
FPY 14 721 1.52 � 0.13 482.52353.52e645.71
6.43
Bifenthrin GSS 527 1.49 � 0.15 54.5334.04e85.55
e
FPY 14 720 1.10 � 0.10 12.778.24e18.80
<1
a Total number of mites used.b Confidence interval.c Resistance ratio ¼ LC50 value of the FPY 14 strain/LC50 value of the GSS strain.
Table 4Synergistic effects of PBO, IBP and TPP on fenpyroximate efficacy in T. urticae strainFPY 14 and GSS.
Strains Compoundþ synergist
Na Slope � SE LC50 (mg a.i./l)(95% CIb)
SRRc SRd
FPY 14 Fenpyroximate 720 1.45 � .0.12 213.26142.84e316.76
64.43 e
þPBO 720 1.12 � �0.10 157.18124.46e199.92
55.73 1.36
þIBP 720 1.68 � 0.14 191.52132.43e269.31
43.13 1.11
þTPP 720 1.44 � �0.125 209.87145.73e296.88
61.01 1.02
GSS Fenpyroximate 720 1.35 � 0.10 3.312.71e3.99
e e
þPBO 720 1.40 � 0.11 2.821.75e4.21
e 1.17
þIBP 720 1.18 � 0.10 4.443.22e6.05
e <1
þTPP 723 1.30 � 0.11 3.442.73e4.26
e <1
a Total number of mites used in experiment.b Confidence interval.c Synergistic resistance ratio ¼ LC50 for FPY 14/LC50 GSS strain.d Synergistic ratio ¼ LC50 for fenpyroximate alone/LC50 for fenpyroximate with
synergist.
R. Ay, F.E. Kara / Crop Protection 30 (2011) 605e610608
model, and the PDIFF option was used to compare the meanenzyme-activity values of the strains. Statistical significance wasassessed using an alpha level of 0.05.
3. Results
3.1. Selection for resistance
According to the LC50 values, the resistance of the BEYO 2 strainafter 14 rounds of selection increased from 1.06-fold to 64.43-fold(in FPY 14). LC50 values increased from 3.31 mg a.i./l water to213.26 mg a.i./l water (Table 1). The results were examined to findthe post-selection population at which resistance became signifi-cant based on the non-overlap of 95% CIs. Table 1 shows thatpopulations Sel 3 and Sel 4 were the first significant populations forLC50 and LC90, respectively. After 14 rounds of selection, the resis-tance level of the FPY 14 strain increased by 60.76-fold compared tothat of the original strain BEYO 2.
Table 3Concentrationeresponse data of fenpyroximate on female adults of the GSS and FPY14 strains and their reciprocal crosses.
Strains Na Slope � SE LC50 (mg a.i./l)(95% CIb)
LC50 RRc Dd
GSS 720 1.35 � 0.10 3.312.71e3.99
e e
FPY 14 720 1.45 � 0.12 213.26142.84e316.76
64.43 e
GSS _ � FPY 14 \ F1 720 1.23 � 0.10 139.63112.36e173.08
42.18 0.80
FPY14 _ � GSS \ F1 720 1.53 � 0.12 150.87112.11e201.45
45.58 0.83
a Total number of mites used.b Confidence interval.c Resistance ratio ¼ LC50 value of resistance strain/LC50 value of the GSS strain.d Degree of dominance.
3.2. Toxicity of other pesticides to the fenpyroximate-selected strain
The LC50 values and resistance ratios to pesticides with sixdifferent modes of action were determined in the fenpyroximate-resistant FPY 14 strain. The results indicated that the FPY 14 strainwas 7.80-fold more resistant to abamectin, 6.90-fold more resistantto chlorpyrifos, 6.43-fold more resistant to propargite, 4.78-foldmore resistant to clofentezine and 2.78-fold more resistant toamitraz than the GSS strain (Table 2). However, the FPY 14 straindid not exhibit resistance to bifenthrin compared to the GSS strainbut instead was more susceptible to this pesticide.
3.3. Inheritance of resistance
Table 3 presents the F1 LC50 values and resistance ratiosobtained at the end of the reciprocal crossings between the FPY14 and GSS (susceptible) strains. The D values were 0.80 for GSS_ � FPY 14 \ and 0.83 for FPY 14 _ � GSS \ (Table 3). The resultsshowed that the fenpyroximate-resistance trait was transferredby incompletely dominant(s) gene and that it was not sexdependent.
Table 5Esterase, GST, and P450 activities in susceptible strain GSS, parental strain BEYO 2and fenpyroximate-resistant strain FPY 14 of T. urticae (P < 0.05).a
Enzyme Strains Nb Specific activity (�SE)mOD min�1 mg�1 proteins
R/Sc
Esterase GSS 3 10.11 (�0.62)c e
BEYO 2 3 13.47 (�0.62)b 1.33FPY 14 3 19.49 (�0.62)a 1.92
GST GSS 4 13.66 (�0.35)a e
BEYO 2 3 10.59 (�0.40)b 0.77FPY 14 5 14.58 (�0.31)a 1.06
P450 GSS 3 0.0027(�0.0006)b e
BEYO 2 3 0.0028 (�0.0006)b 1.03FPY 14 3 0.0107 (�0.0005)a 3.96
a Means with different letters in column for each enzyme are significantlydifferent (P < 0.05).
b Number of replicates.c Enzyme activity BEYO 2 or FPY 14/enzyme-activity GSS strain.

R. Ay, F.E. Kara / Crop Protection 30 (2011) 605e610 609
3.4. Synergistic effects
The fenpyroximate-resistant FPY 14 and susceptible GSS strainswere exposed to the synergists PBO, IBP and TPP in combinationwith fenpyroximate. The synergistic effects of PBO, IBP and TTP onthe FPY 14 strain were 1.36-, 1.11- and 1.02-fold, respectively. In theGSS strain, on the other hand, only PBO had a synergistic effect(1.17-fold). When considered on the basis of LC50 confidenceintervals, however, none of the synergists showed a significantsynergistic effect (Table 4). The synergistic resistance ratios of theFPY 14 strain with PBO, IBP and TTP were calculated to be 55.73-,43.13- and 61.01-fold, respectively. According to the synergisticresistance ratios, the strongest synergistic effect was shown by IBP,followed by PBO. TTP had the weakest synergistic effect.
3.5. Biochemical analyses
All three enzymes tested in this study showed statisticallysignificant different activity levels in the fenpyroximate-resistantFPY 14 strain compared to the original BEYO 2 strain (P < 0.005)(Table 5). While esterase activity was significantly higher in BEYO 2when compared to the GSS strain (P< 0.05), GST activity was lower(P > 0.05). The activity of P450 was equal in BEYO 2 and thesusceptible strain (P ¼ 0.05) (Table 5). Compared to those observedin the GSS strain, the activities of esterase, GST and P450 in FPY 14increased by 1.92-fold, 1.06-fold and 3.96-fold, respectively.Compared to those observed in BEYO 2, the activities of esterase,GST and P450 in FPY 14 increased by 1.44-fold, 1.38-fold and 3.82-fold, respectively.
4. Discussion
The resistance of T. urticae to fenpyroximate in BEYO 2-derivedstrains was analysed via bioassays and biochemical methods. Fen-pyroximate resistance increased with increasing selection pressure.Other studies have indicated that fenpyroximate resistancedevelops in T. urticae strains according to the frequency andnumber of applications (Stumpf and Nauen, 2001; Devine et al.,2001; Sato et al., 2004; Kim et al., 2004; Van Pottelberge et al.,2009). Resistance-management programs concentrate on whethera strain that is resistant to a particular pesticide is also resistant toother pesticides. The fenpyroximate-resistant FPY 14 strain showsresistance to the avermectin abamectin, the organophosphatechlorpyrifos, the sulphite ester propargite, the mite-growth regu-lator clofentezine and the formamidine amitraz at low levels. Ay(2006) has reported that the BEYO 2 strain is 1.82-fold moreresistant to abamectin and 1.15-fold more resistant to amitrazcompared to the GSS strain in an earlier study in a bean greenhousein 2003. However, propargite susceptibility was detected in thesame study. In the current study, fenpyroximate selection increasedabamectin and propargite resistance. Kim et al. (2004) have per-formed fenpyroximate selection on 20 generations of T. urticae anddetected cross-resistance to 18 acaricides in the FR-20 strain. Theyalso detected a high level of cross-resistance to propargite (63.8-fold) and moderate resistance to abamectin (24.1-fold). Sato et al.(2004) have performed five rounds of fenpyroximate selection ona T. urticae strain and reported positive cross-resistance to pyr-idaben and dimethoate. However, they detected no cross-resis-tance between fenpyroximate and propargite, abamectin,milbectin, fenpropathrin or cyhexatin. Stumpf and Nauen (2001)have reported cross-resistance between the METI acaricides pyr-idaben, fenpyroximate and tebufenpyrad. Interestingly, no cross-resistance to the pyrethroid bifenthrin was found in the FPY 14strain in the current study. Ahmad et al. (2007) have reporteda negative correlation betweenmethomyl and bifenthrin resistance
in Spodoptera litura (F.); this correlation can be exploited in resis-tance management.
According to the data acquired from F1 generations obtainedthrough reciprocal crossing, fenpyroximate resistance in T. urticae isincompletelydominant andautosomal. VanPottelberge et al. (2009)have foundD values of 1.12 for GSS \�MR-VP _ and 0.93 for MR-VP\�GSS_ in F1 generations obtained bycrossing the fenpyroximate-resistant MR-VP strain with the susceptible GSS strain, indicatingthat resistance is dominant and is not sex dependent. Sato et al.(2004) have found D values of 0.511 for S \ � R _ and 0.580 for R\ � S _, indicating that resistance is incompletely dominant and isnot sex dependent. The results of the present study are consistentwith these earlier studies and others reported in the literature(Devine et al., 2001; Stumpf and Nauen, 2001).
The enzymes P450, GST and esterase and combinations thereofplay important roles in the metabolic detoxification of many acari-cides in mites (Kim et al., 2006). Van Pottelberge et al. (2009) havereported that P450 activity in response to treatment with the METIacaricides tebufenpyrad, fenpyroximate and pyridaben increased by23.5-fold in the MR-VP strain of T. urticae compared to the suscep-tible GSS strain (detected with 7-EFC O-demethylation) and thatthere were no significant differences between different substrates(4-nitrophenol, 1- and 2-naphthol) and resistant and susceptiblestrains in terms of esterase activity. Furthermore, GST activitydetected with CDNB and MCB substrates was significantly high inthe resistant strain; however, this difference was low with CDNB(1.2-fold). Kim et al. (2004) have found that esterase (2.5-fold with1-naphthyl acetate) and P450 (2.5-fold with PNOD) levels weresignificantly higher in the fenpyroximate-selected FR-20 straincompared to the susceptible strain; however, there was no signifi-cant difference in GST (1.2-fold with CDNB). Stumpf and Nauen(2001) have reported a significant correlation (0.88) between fen-pyroximate resistance and P450 activity in T. urticae.
Synergists are important factors in determining the effectivenessof enzymes in resistance mechanisms. IBP and TTP, which areesterase inhibitors, andPBO,which is an esterase andP450 inhibitor,exhibited no synergistic effects on the LC50 confidence intervals inthe present study. Van Pottelberge et al. (2009) have indicated thatfenpyroximate and PBO exhibit a low-level synergistic effect (1.9-fold) in theMETI-acaricide-resistant MR-VP strain. Kim et al. (2004)have found that fenpyroximate has a strong synergistic effect withPBO in the fenpyroximate-selected FR-20 strain (SR¼371) andweaksynergistic effects with IBP and TPP (SR¼ 3.6 and 4.4, respectively).Stumpf and Nauen (2001) have indicated that fenpyroximate andPBO have only a 3.3-fold synergistic effect in the METI-acaricides-resistant AKITA strain and that PBO has no significant synergisticeffect on theMETI-acaricides-resistant UK strain. Van Leeuwenet al.(2009) have reported that this lack of synergism in fenpyroximateresistance, together with the clearly monogenic inheritance of theresistance trait, suggests an altered target site. In the present study,the synergistic effect ratios of fenpyroximate and synergists werelowin theFPY14 strain;however, IBP (anesterase inhibitor) andPBO(an esterase andP450 inhibitor) decreased the synergistic resistanceratios from 64.43-fold to 43.13-fold and 55.73-fold, respectively, onthe basis of the LC50 values.
The results of the present study and previous studies collectivelysuggest that the enzymes P450 and esterase play a role in themechanisms of fenpyroximate resistance. However, the weakeffects of synergists on fenpyroximate resistance suggest thatadditional mechanisms are involved.
Acknowledgements
This study was supported by the Scientific and TechnologicalResearch Council of Turkey (TUBITAK, TOVAG-105O179).

R. Ay, F.E. Kara / Crop Protection 30 (2011) 605e610610
References
Ahmad, M., Arif, M.I., Ahmad, M., 2007. Occurrence of insecticide resistance in fieldpopulations of Spodoptera litura (Lepidoptera: Noctuidae) in Pakistan. Crop Prot.26, 809e817.
Ay, R., 2005. Determination of susceptibility and resistance of some greenhousepopulations strains of Tetranychus urticae Koch to chlorpyrifos (dursban 4) bythe petri dish-potter tower method. J. Pest Sci. 78, 139e143.
Ay, R., 2006. Antalya ili örtüaltı sebze üretim alanlarında zararlı olan Tetranychusurticae Koch popülasyonlarının bazı akarisitlere karsı tepkileri. A.Ü. Zir. Fak. Tar.Bilim. Derg. 12, 301e306.
Devine, G.J., Barberü, M., Denholm, I., 2001. Incidence and inheritance of resistanceto METI-acaricides in European strains of the two-spotted spider mite (Tetra-nychus urticae) (Acari: Tetranychidae). Pest Manag. Sci. 57, 443e448.
Holingworth, R.M., Ahammadsahib, K.I., 1995. Inhibitors of respiratory complex 1:mechanisms, pesticidal actions and toxicology. Rev. Pestic. Toxicol. 3, 277e302.
Kang, C.Y., Wu, G., Miyata, T., 2006. Synergism of enzyme inhibitors and mecha-nisms of insecticide resistance in Bemisia tabaci. (Gennadius.) (Hom., Aleyr-odidae). J. Appl. Entomol. 130, 377e385.
Kim, Y.J., Lee, S.H., Lee, S.W., Ahn, Y.J., 2004. Fenpyroximate resistance in Tetranychusurticae (Acari: Tetranychidae) cross-resistance and biochemical resistancemechanisms. Pest Manag. Sci. 60, 1001e1006.
Kim, Y.J., Park, H.M., Cho, J.R., Ahn, Y.J., 2006. Multiple resistance and biochemicalmechanisms of pyridaben resistance in Tetranychus urticae (Acari: Tetranychi-dae). J. Econ. Entomol. 99, 954e958.
LeOra, 1994. Software, POLO e PC: A User’s Guide to Probit or Logit Analysis LeOraSoftware, Berkeley, CA. p. 28.
Rose, R., Barbhaiya, L., Roe, R., Rock, G., Hodgson, E., 1995. Cytochrome P-450-associated insecticide resistance and the development of biochemical diag-nostic assays in Heliothis virescens. Pestic. Biochem. Physiol. 51, 178e191.
SAS, 1999. Statistical Analysis Systems User’s Guide, eighth ed.. SAS Institute INC,Raleigh, North Carolina, USA.
Sato, M.E., Miyata, T., da Silva, M., Raga, A., De Souza Filho, M.F., 2004. Selections forfenpyroximate resistance and susceptibility, and inheritance, cross-resistanceand stability of fenpyroximate resistance in Tetranychus urticae Koch (Acari:Tetranychidae). Appl. Entomol. Zool. 39, 293e302.
Stone, B.F., 1968. A formula for determining degree of dominance in cases ofmonofactorial inheritance of resistance to chemicals. Bull. World Health Organ38, 325e326.
Stumpf, N., Nauen, R., 2001. Cross-Resistance, inheritance, and biochemistry ofmitochondrial electron transport inhibitor-acaricide resistance in Tetranychusurticae (Acari: Tetranychidae). J. Econ. Entomol. 94, 1577e1583.
Stumpf, N., Nauen, R., 2002. Biochemical markers linked to abamectin resistancein Tetranychus urticae (Acari: Tetranychidae). Pestic. Biochem. Physiol. 72,111e121.
Stumpf, N., Zebitz, C.P.W., Kraus, W., Moores, G.D., Nauen, R., 2001. Resistance toorganophosphates and biochemical genotyping of acetylcholinesterases inTetranychus urticae (Acari: Tetranychidae). Pestic. Biochem. Physiol. 69,131e142.
Suh, E., Koh, S.-Y., Lee, J.-H., Shin, K.-I., Cho, K., 2006. Evaluation of resistance patternto fenpyroximate and pyridaben in Tetranychus urticae collected from green-houses and apple orchards using lethal concentration-slope relationship. Exp.Appl. Acarol 38, 151e165.
Tsagkarakou, A., Pasteur, N., Cuany, A., Najavas, M., 2002. Mechanisms of resistanceto organophosphates in Tetranychus urticae (Acari: Tetranychidae) from Greece.Insect Biochem. Molec. 32, 417e424.
Van Leeuwen, T., Tirry, L., 2007. Esterase-mediated bifenthrin resistance in a mul-tiresistant strain of the two-spotted spider mite, Tetranychus urticae. PestManag. Sci. 63, 150e156.
Van Leeuwen, T., Pottelberge, S.V., Tirry, L., 2005. Comparative acaricidesusceptibility and detoxifying enzyme activities in field e collected resis-tant and susceptible strains of Tetranychus urticae. Pest Manag. Sci. 61,499e507.
Van Leeuwen, T., Van Pottelberge, S., Nauen, R., Tirry, L., 2007. Organophosphateinsecticides and acaricides antagonise bifenazate toxicity through esteraseinhibition in Tetranychus urticae. Pest Manag. Sci. 63, 1172e1177.
Van Leeuwen, T., Vontas, J., Tsagkarakou, A., Tirry, L., 2009. Mechanisms of acaricideresistance in the two-spotted spider mite Tetranychus urticae. In: Ishaaya, I.,Horowitz, A.R. (Eds.), Biorational Control of Arthropod Pests. Springer,Netherlands, pp. 347e393.
Van Pottelberge, S., Van Leeuwen, T., Nauen, R., Tirry, L., 2009. Resistance mech-anisms to mitochondrial electron transport inhibitors in a field-collectedstrain of Tetranychus urticae Koch (Acari: Teranychidae). Bull. Entomol. Res. 99,23e31.
Wang, L., Wu, Y., 2007. Cross-resistance and biochemical mechanisms of abamectinresistance in the B-type Bemisia tabaci. J. Appl. Entomol. 131, 98e103.
Yang, X., Buschman, L.L., Zhu, Y.K., Margolies, D.C., 2002. Susceptibility and detox-ifying enzyme activity in two spider mite species (Acari: Tetranychidae) afterselection with three insecticides. J. Econ. Entomol. 95, 399e406.
Young, S.J., Gunning, R.V., Moores, G.D., 2005. The effect of piperonyl butoxide onpyrethroid resistance associated esterases in Helicoverpa armigera (Hübner)(Lepidoptera: Noctuidae). Pest Manag. Sci. 61, 397e401.
Yu, S.J., 2008. Insecticide resistance. In: Yu, S.J. (Ed.), The Toxicology andBiochemistry of Insecticide. CRC Press, New York, pp. 2001e2230.
















![IJCBS RESEARCH PAPER VOL. 1 [ISSUE 3] JUNE, 2014 ISSN:-2349–2724 Light Emitting Diodes (LEDs) Reduce Vertimec, Resistance in Tetranychus urticae (Koch](https://static.fdokumen.com/doc/165x107/631ba468a906b217b9069031/ijcbs-research-paper-vol-1-issue-3-june-2014-issn-23492724-light-emitting.jpg)

