
Efficacy of ketoenols on insecticide resistant field populations of two-spotted spider mite...
7
Efficacy of ketoenols on insecticide resistant field populations of two-spotted spider mite Tetranychus urticae and sweet potato whitefly Bemisia tabaci from Greece Aris Ilias a, b, 1 , Emmanouil Roditakis a,1 , Maria Grispou a , Ralf Nauen c , John Vontas b , Anastasia Tsagkarakou a, * a Hellenic Agricultural Organisation-Demeter, NAGREF, Plant Protection Institute of Heraklion, P.O. Box 2228, 71003 Heraklion, Greece b Faculty of Biotechnology and Applied Biology, Department of Biology, University of Crete, 71409 Heraklion, P.O. Box 2208, Greece c Bayer CropScience, Research Pest Control, D-40789 Monheim, Germany article info Article history: Received 9 May 2012 Received in revised form 20 July 2012 Accepted 29 July 2012 Keywords: Spiromesifen Spirodiclofen Acaricides Resistance management abstract Tetranychus urticae (two-spotted spider mite) and Bemisia tabaci (sweet potato whitefly), two of the most important agricultural pests worldwide, have developed resistance to almost all chemical classes of insecticides. Here we investigated the efficacy of the ketoenols spirodiclofen and spiromesifen against, respectively, T. urticae and B. tabaci populations from Greece with variable levels of resistance to other pesticides in order to evaluate their potential role in insecticide resistance management strategies for sustainable control of both pests. No resistance of T. urticae populations against spirodiclofen was observed. Nine out of ten T. urticae populations exhibited moderate levels of resistance to pirimiphos-methyl (up to 23-fold), pyridaben (up to 39-fold) and fenazaquin (up to 42-fold). Two of them exhibited high resistance ratios (RR) to bifenthrin (RR ¼ 81, 351) and one to fenbutatin oxide (RR ¼ 146). Moreover these nine pop- ulations were not cross resistant to spirodiclofen (RR < 5). A single population from greenhouse roses exhibited high resistance levels to most of the pesticides tested (RR ¼ 8413, 1494, 434, and 74 to bifenthrin, fenbutatin oxide, abamectin and pirimiphos-methyl, respectively) showed decreased susceptibility to spi- rodiclofen (RR ¼ 12). In B. tabaci the variation in response to spiromesifen treatments was very low among the eleven field populations. Most of them exhibited high levels of resistance to imidacloprid, and alpha- cypermethrin. The LC 50 of spiromesifen ranged from 4.5 to 14 mg/l and the corresponding LC 95 values were in general well below the recommended field doses. Ketoenol resistance risk potential was also investigated by applying constant selection pressure under laboratory conditions against populations from both species, but no increase in LC 50 s was detected. Ketoenols can be used for resistance management purposes for both pests in Greece as an effective chemical tool in rotation with other pesticides. Ó 2012 Elsevier Ltd. All rights reserved. 1. Introduction The two-spotted spider mite Tetranychus urticae Koch (Acari: Tetranychidae) and the sweet potato whitefly Bemisia tabaci (Gen- nadius) (Homoptera: Aleyrodidae) are two of the most important pests of many crops worldwide. Moreover B. tabaci has been shown to transmit more than 100 plant-pathogenic virus species (Jones, 2003). Both pests have a unique ability to develop resistance to major pesticides rapidly due to their short life-cycle, high fecundity, mode of reproduction and strong selection by repeated insecticide treatments to keep populations under economic damage thresholds (Knowles, 1997; Denholm et al., 1998; Van Leeuwen et al., 2010). In Greece, both T. urticae and B. tabaci infest a wide range of greenhouse, open field vegetable and ornamental crops. Neon- icotinoids, pyrethroids and organophosphates have been mainly used for B. tabaci control, while avermectins, METI’s (mitochondrial electron transport inhibitors) and pyrethroids are most commonly applied against T. urticae in Greece. In a recent survey, high resis- tance levels to the neonicotinoid imidacloprid and the pyrethroid a-cypermethrin were reported in field populations of B. tabaci from Greece (Roditakis et al., 2009). Resistance data for T. urticae pop- ulations from Greece have been limited to OPs and pyrethroids (Tsagkarakou et al., 1996, 2002, 2009a; Khajehali et al., 2010). In addition, growers often claim serious problems in controlling this pest by using other insecticides. * Corresponding author. Tel.: þ30 2810 302300; fax: þ30 2810 245858. E-mail address: [email protected] (A. Tsagkarakou). 1 Contributed equally to this work. Contents lists available at SciVerse ScienceDirect Crop Protection journal homepage: www.elsevier.com/locate/cropro 0261-2194/$ e see front matter Ó 2012 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.cropro.2012.07.024 Crop Protection 42 (2012) 305e311
Transcript of Efficacy of ketoenols on insecticide resistant field populations of two-spotted spider mite...

at SciVerse ScienceDirect
Crop Protection 42 (2012) 305e311
Contents lists available
Crop Protection
journal homepage: www.elsevier .com/locate/cropro
Efficacy of ketoenols on insecticide resistant field populations of two-spottedspider mite Tetranychus urticae and sweet potato whitefly Bemisia tabacifrom Greece
Aris Ilias a,b,1, Emmanouil Roditakis a,1, Maria Grispou a, Ralf Nauen c, John Vontas b,Anastasia Tsagkarakou a,*
aHellenic Agricultural Organisation-Demeter, NAGREF, Plant Protection Institute of Heraklion, P.O. Box 2228, 71003 Heraklion, Greeceb Faculty of Biotechnology and Applied Biology, Department of Biology, University of Crete, 71409 Heraklion, P.O. Box 2208, GreececBayer CropScience, Research Pest Control, D-40789 Monheim, Germany
a r t i c l e i n f o
Article history:Received 9 May 2012Received in revised form20 July 2012Accepted 29 July 2012
Keywords:SpiromesifenSpirodiclofenAcaricidesResistance management
* Corresponding author. Tel.: þ30 2810 302300; faE-mail address: [email protected] (A. Tsagkar
1 Contributed equally to this work.
0261-2194/$ e see front matter � 2012 Elsevier Ltd.http://dx.doi.org/10.1016/j.cropro.2012.07.024
a b s t r a c t
Tetranychus urticae (two-spotted spider mite) and Bemisia tabaci (sweet potato whitefly), two of the mostimportant agricultural pests worldwide, have developed resistance to almost all chemical classes ofinsecticides. Here we investigated the efficacy of the ketoenols spirodiclofen and spiromesifen against,respectively, T. urticae and B. tabaci populations from Greece with variable levels of resistance to otherpesticides in order to evaluate their potential role in insecticide resistance management strategies forsustainable control of both pests. No resistance of T. urticae populations against spirodiclofenwas observed.Nine out of ten T. urticae populations exhibited moderate levels of resistance to pirimiphos-methyl (up to23-fold), pyridaben (up to 39-fold) and fenazaquin (up to 42-fold). Two of them exhibited high resistanceratios (RR) to bifenthrin (RR ¼ 81, 351) and one to fenbutatin oxide (RR ¼ 146). Moreover these nine pop-ulations were not cross resistant to spirodiclofen (RR < 5). A single population from greenhouse rosesexhibited high resistance levels to most of the pesticides tested (RR¼ 8413, 1494, 434, and 74 to bifenthrin,fenbutatin oxide, abamectin and pirimiphos-methyl, respectively) showed decreased susceptibility to spi-rodiclofen (RR ¼ 12). In B. tabaci the variation in response to spiromesifen treatments was very low amongthe eleven field populations. Most of them exhibited high levels of resistance to imidacloprid, and alpha-cypermethrin. The LC50 of spiromesifen ranged from 4.5 to 14 mg/l and the corresponding LC95 values werein general well below the recommended field doses. Ketoenol resistance risk potential was also investigatedby applying constant selection pressure under laboratory conditions against populations from both species,but no increase in LC50s was detected. Ketoenols can be used for resistance management purposes for bothpests in Greece as an effective chemical tool in rotation with other pesticides.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
The two-spotted spider mite Tetranychus urticae Koch (Acari:Tetranychidae) and the sweet potato whitefly Bemisia tabaci (Gen-nadius) (Homoptera: Aleyrodidae) are two of the most importantpests of many crops worldwide. Moreover B. tabaci has been shownto transmit more than 100 plant-pathogenic virus species (Jones,2003). Both pests have a unique ability to develop resistance tomajor pesticides rapidly due to their short life-cycle, high fecundity,mode of reproduction and strong selection by repeated insecticide
x: þ30 2810 245858.akou).
All rights reserved.
treatments to keep populations under economic damage thresholds(Knowles, 1997; Denholm et al., 1998; Van Leeuwen et al., 2010).
In Greece, both T. urticae and B. tabaci infest a wide range ofgreenhouse, open field vegetable and ornamental crops. Neon-icotinoids, pyrethroids and organophosphates have been mainlyused for B. tabaci control, while avermectins, METI’s (mitochondrialelectron transport inhibitors) and pyrethroids are most commonlyapplied against T. urticae in Greece. In a recent survey, high resis-tance levels to the neonicotinoid imidacloprid and the pyrethroida-cypermethrinwere reported in field populations of B. tabaci fromGreece (Roditakis et al., 2009). Resistance data for T. urticae pop-ulations from Greece have been limited to OPs and pyrethroids(Tsagkarakou et al., 1996, 2002, 2009a; Khajehali et al., 2010). Inaddition, growers often claim serious problems in controlling thispest by using other insecticides.

Fig. 1. T. urticae (TU) and B. tabaci (GR) sampling sites.
Table 1Name, location, collection date, and host plant of Tetranychus urticae and Bemisiatabaci field populations.
Population Location Collection date Host planta
Tetranychus urticaeTU1 Tympaki June 2006 Melon e GTU2 Tympaki June 2006 Tomato e FTU3 Ierapetra June 2006 Cucumber, beans e GTU4 Vianos June 2006 Bean e FTU5 Ierapetra July 2006 Water melon, tomato e GTU6 Zakros Sept.2006 Eggplant e FTU7 Marathonas Febr. 2007 Rose e GTU9 Pyrgos March 2007 Cucumber e GTU11 Pyrgos March 2007 Carnation e GTU12 Aigio March 2007 Lemon e FBemisia tabaciGR157 Tympaki June 2006 Okra e FGR163 Vianos June 2006 Cucumber e FGR164 Vianos June 2006 Cucumber e GGR165 Antiskari July 2006 Melon e GGR187 Vianos May 2007 Cucumber e GGR188 Ierapetra May 2007 Pepper e GGR191 Vianos June 2007 Bean e GGR192 Ierapetra June 2007 Pepper e GGR194 Tympaki June 2007 Mellon e GGR258 Tympaki June 2008 Eggplant e GGR259 Kolimpari June 2008 Cucumber, zucchini e F
a The name of the host plants is followed by a G or F for greenhouse and field,respectively.
A. Ilias et al. / Crop Protection 42 (2012) 305e311306
Spiromesifen and spirodiclofen belong to the class of spirocyclictetronic acid derivatives (ketoenols), recently developed andregistered in the agrochemical market in several countries (Nauenet al., 2003). Spirodiclofen has been commercialised for the controlof several mite species (Rauch and Nauen, 2002;Wachendorff et al.,2002). Spiromesifen is an insecticide especially active againstwhiteflies with additional activity against tetranychidmites (Nauenet al., 2002). Ketoenols have been shown to interfere with pestdevelopment as well as female fecundity and fertility (Nauen,2005; Kontsedalov et al., 2008). They have a distinct target-sitecompared to other insecticide classes, i.e. they act on acetyl-CoA-carboxylase (ACCase), a key enzyme to lipid biosynthesis (Nauen,2005; Bretschneider et al., 2007). Spiromesifen and spirodiclofenwere introduced to the Greek market in late 2008 and 2009,respectively. Before the time of sampling they had never beenapplied to the field to the best of our knowledge.
Here we examined the baseline efficacy of spirodiclofen andspiromesifen against spider mite and whitefly field populationsfrom Greece respectively with no previous exposure to ketoenols,but a variable pattern of resistance against a number of registeredinsecticides used for their control. More specifically we examined:a) the levels of resistance of 10 T. urticae field populations fromGreece to abamectin (avermectins), bifenthrin (pyrethroid),pirimiphos-methyl (OP), fenbutatin oxide (organotin), pyridabenand fenazaquin (METI), b) the presence of cross resistance to spi-rodiclofen and spiromesifen in T. urticae and B. tabaci respectivelyand c) the risk of development of resistance by artificial selectionpressure under laboratory conditions.
2. Materials and methods
2.1. Spider mite and whitefly populations
Ten T. urticae populations (6 from Crete and 4 from mainlandGreece) were collected in 2006 and 2007 from greenhouse and fieldcrops, most of them with extensive spraying history and failurereports (Fig. 1). A detailed record for each population is given inTable 1. Two populations derived from greenhouses: populationTU7 collected on rose plants in Marathonas near Athens and TU11from carnation plants in Peloponnesus. Populations TU1, TU2,TU3, TU4, TU5 and TU9 were collected from extended vegetableproduction areas where abamectin and bifenthrin were mainlyused for the control of spider mites. Population TU6 was collectedon vegetables in a family garden in a location of East Crete isolatedfrom intensive cultures. Laboratory strain ALL was derived bymixing of 7 populations, 5 from Crete (TU1, TU2, TU3, TU4 and TU5)and 2 from Peloponnesus (TU9 and TU11) and was used for labo-ratory selections with spirodiclofen.
Mites were reared for 1e2 generations in the laboratory prior tobioassays. All mites were maintained on detached bean leaves thatwere deposited on a piece of moist cotton to prevent contaminationand escape. Rearing conditions were 25 � 2 �C, 70 � 5% RH anda 16:8 h light: dark photoperiod. Each colonywas initiated by> 300females and males coming from the field. Species identificationwas performed morphologically by examining the male aedeagousof w10 males from each population. All populations belonged toT. urticae and no mixture of more than one tetranychid species wasfound.
Two laboratory reference T. urticae strains were also used: SAMBinitially collected on Sambucus nigra (L.) in the Netherlands andobtained from the Centre de Biologie et de Gestion des Populations,Montpellier, France and GSS originally collected from an unknownhost from Germany and obtained from Bayer Crop Science. Bothhad been maintained in the laboratory without any treatmentsince 1965.
Eleven B. tabaci populations (Fig. 1) were collected in Creteduring a large survey of B. tabaci resistance to imidacloprid,a-cypermethrin and pirimiphos-methyl (Roditakis et al., 2009).Neonicotinoids, OPs and pyrethroids were the most commonlyused classes of insecticides for their control in the particulargreenhouses (Roditakis et al., 2009). Most of them exhibited highlevels of resistance (>100-fold) to imidacloprid and a-cypermeth-rin (Roditakis et al., 2009). All field populations from Crete wereQ-type B. tabaci (Tsagkarakou et al., 2009b), considered now as theMED species (Dinsdale et al., 2010).

A. Ilias et al. / Crop Protection 42 (2012) 305e311 307
A detailed record for each population is given in Table 1. Thefield populations were used in bioassays without prior rearing inthe laboratory. Population GR164was used for laboratory selectionswith spiromesifen. Two laboratory strains, GRMAL-RN and GRMAL-RP, resistant to a-cypermethrin and imidacloprid respectively, werealso used to evaluate cross resistance with spiromesifen. Thesewere maintained under selection pressure on cotton plants formore than two years prior the spiromesifen toxicity assays at 24 �C.
2.2. Chemicals
Commercial formulations of pirimiphos-methyl 500 g/l EC(Actellic, Syngenta, UK), bifenthrin 100 g/l EC (Talstar, FMC, Athens,Greece), pyridaben 200 g/l EC (Nexter, Du Pont, Greece), fenazaquin200 g/l SC (Pride, Elanko, Greece), fenbutatin oxide 550 g/l SC (TNT,Alpha, Greece), and abamectin 18 g/l EC (Vertimec, Syngenta,Greece) were used. Spirodiclofen and spiromesifen were ofanalytical grade and formulated by preparing stock solutions ina 3:1 (v/v) mixture of N,N-dimethylformamide and alkylar-ylpolyglycolether (emulsifier) and subsequently diluted withdeionized water. Spirodiclofen was tested on larvae of mites andspiromesifen on 2nd instar nymphs of whiteflies. All other pesti-cides were tested on adult females.
2.3. Toxicological bioassays
2.3.1. Bioassay on mite larvaeTen to fifteen adult females of T. urticaewere transferred into the
area of a sticky barrier (35 mm in diameter) on the upper leafsurface of a bean plant at the stage of the two cotyledons fullyexpanded. Oviposition was terminated by removing all femalesafter 24 h. When larvae hatched the bean leaves were placed onmoistened cotton wool in a plastic Petri dish. The individual Petridishes were then immediately sprayed with 2 ml aqueous solutionof spirodiclofen using a precision Potter Spray Tower at 1 barpressure in order to obtain a homogenous spray film (5.1 mg � 0.7aqueous acaricide deposit/cm2). Four replicates of five concentra-tions plus a control were tested. Controls were sprayedwith solventsolution alone. The treated discs were subsequently transferred to25 (�1) �C, 50e60% RH and 16:8 h light: dark photoperiod.Mortality was assessed after the emergence of adults on thesolvent-treated control leaves (approximately 6 days).
2.3.2. Bioassays on adult mitesBean leaves (cotyledons fully expanded) were placed on wet
cotton in plastic Petri dishes for conducting bioassays. Twenty-fiveadult females were transferred onto a leaf with a fine brush insidea 35 mm disc delimitated by insect glue (Dr. Trapp’s Gluekillers,Stac Ltd., Athens, Greece) to prevent mites from escaping. Indi-vidual Petri dishes were then sprayed with a precision Potter SprayTower (Burkard, Rickmansworth, Hertfordshire, UK) (Potter, 1952)with 1 ml aqueous solution of insecticide at 1 bar pressure in orderto obtain a homogenous spray film (2.4 mg � 0.3 aqueous acaricidedeposit/cm2). Four replicates of five concentrations plus a controlwere tested. Controls were sprayed with distilled water alone. Themites were then placed at 25 (�1) �C, 50e60% RH and 16:8 h light:dark photoperiod. Mortality was assessed after 48 h for bifenthrin,pirimiphos-methyl, abamectin, and fenbutatin oxide and after 24 hfor pyridaben and fenazaquin.
2.3.3. Bioassays on whitefly nymphsThe bioassay on whitefly larvae is described in Nauen and
Konanz (2005). In brief, 16 leaves from 4 to 5 medium size cottonplants were cut in rectangle shape to fit in ventilated boxes andwere hanged in horizontal position from a fixed point. The
ventilated box minimised whitefly escapes and provided optimumenvironment for egg laying. A rubber band was fixed on the poly-centric box with silicone allowing the necessary flexibility of thebox avoiding injuries of the leaf stem during handling.
Sixty mixed sex whiteflies coming from the field were placed ineach ventilated box and were allowed to oviposit for 24 h. Thewhiteflies were then removed and the eggs were monitored dailyuntil they developed to 2nd instar nymphs (approximately 8e10days at 24 �C). At least 2 leaves per concentration were dipped inthe insecticide dilution for 5 s. Five concentrations of spiromesifen(ranging from 0.01 to 100 mg/l) were used. Solvent solutionwas used for the control treatment. Mortality was estimated after10e12 days (approximately 20e22 days after egg laying in total).Nymphs that reached pupa or adult stage (emerged) wereconsidered alive. Dead nymphs were observed on the leaf surfacedehydrated or with inhibited/abnormal development.
2.4. Artificial selection
2.4.1. Selections with spirodiclofenArtificial selection for resistance to spirodiclofen was under-
taken for the T. urticae strain ALL for 14 generations. ALL resultedfrom amix of seven populations with different origins representingthus a broader genetic pool. Four hundred eighty adult femaleswere transferred onto 8 bean leaves (60 adult females/leaf) andhatched larvae were sprayed with a single concentration of spi-rodiclofen. Roughly 2000 larvae were treated in each selection. Theconcentration used at the beginning of the selection period was1 mg/l (lower than LC50s of the populations from which the strainderived from). Subsequent concentrations used caused w70%mortality in each generation.
2.4.2. Selections with spiromesifenTo examine the possibility of B. tabaci developing resistance to
spiromesifen we performed laboratory selections with the fieldpopulation GR164 which exhibited among the higher LC50s(LC50 ¼ 12 mg/l) and didn’t exhibit high levels of resistance toimidacloprid and a-cypermethrin. The procedure was the same asthe previously described toxicological bioassay method. Cottonleaves infested with 2nd instar nymphs were all immersed ina single concentration of spiromesifen. Selections were applied infour successive generations with spiromesifen concentrationsadjusted to wLC70 mg/l for each generation.
2.5. Data analysis
In dose response bioassays, mortality data were analysed byprobit analysis based on Finney (Finney, 1964) using the Probitsoftware 3.3 (Praxeme) (Raymond et al., 1993). This software teststhe linearity of dose-mortality response and provides the slope, thelethal concentrations (LC) and the 95% confidence limits (CL) of theLC for each mortality line.
Pair wise comparisons were used to investigate for correlationsbetween the responses to different insecticides. A two-tailed testwas applied to investigate the significance of Pearson productmoment correlation. Analysis was conducted with the SPSS 11statistical program (SPSS Inc., Chicago, IL).
3. Results and discussion
3.1. Resistance status of T. urticae populations from Greece againstseveral insecticides
The results of the toxicological assays with T. urticae populationsfrom different parts of Greece are shown in Table 2. Only one field
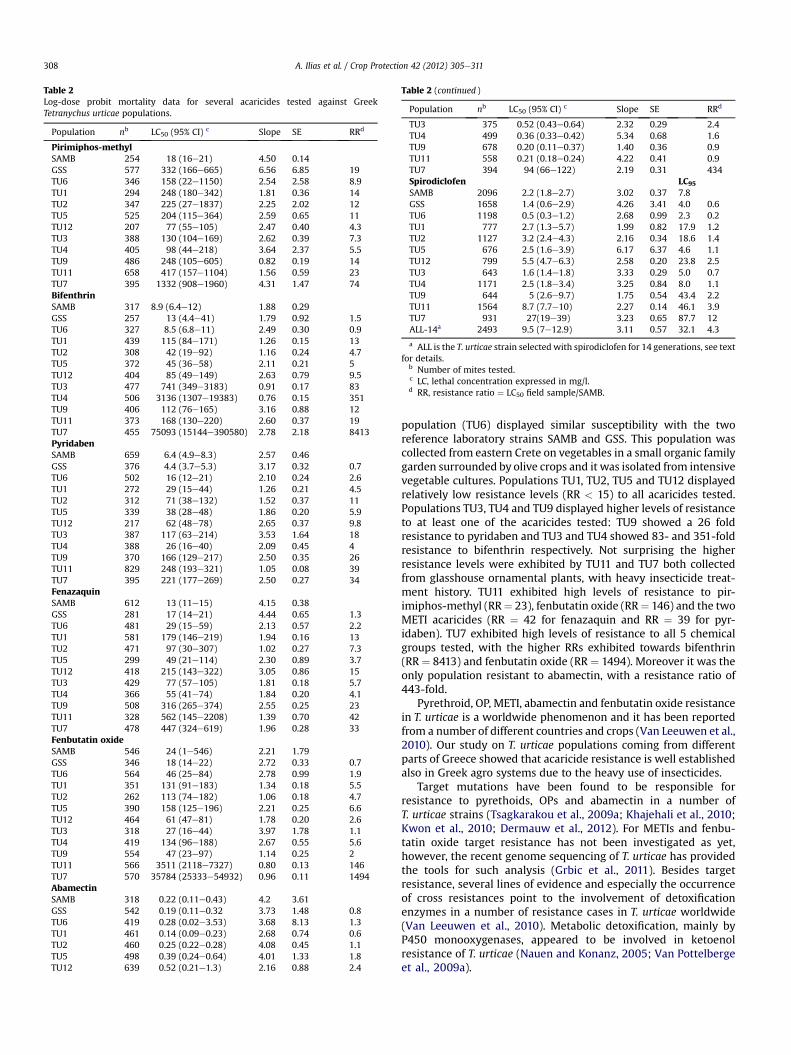
Table 2Log-dose probit mortality data for several acaricides tested against GreekTetranychus urticae populations.
Population nb LC50 (95% CI) c Slope SE RRd
Pirimiphos-methylSAMB 254 18 (16e21) 4.50 0.14GSS 577 332 (166e665) 6.56 6.85 19TU6 346 158 (22e1150) 2.54 2.58 8.9TU1 294 248 (180e342) 1.81 0.36 14TU2 347 225 (27e1837) 2.25 2.02 12TU5 525 204 (115e364) 2.59 0.65 11TU12 207 77 (55e105) 2.47 0.40 4.3TU3 388 130 (104e169) 2.62 0.39 7.3TU4 405 98 (44e218) 3.64 2.37 5.5TU9 486 248 (105e605) 0.82 0.19 14TU11 658 417 (157e1104) 1.56 0.59 23TU7 395 1332 (908e1960) 4.31 1.47 74BifenthrinSAMB 317 8.9 (6.4e12) 1.88 0.29GSS 257 13 (4.4e41) 1.79 0.92 1.5TU6 327 8.5 (6.8e11) 2.49 0.30 0.9TU1 439 115 (84e171) 1.26 0.15 13TU2 308 42 (19e92) 1.16 0.24 4.7TU5 372 45 (36e58) 2.11 0.21 5TU12 404 85 (49e149) 2.63 0.79 9.5TU3 477 741 (349e3183) 0.91 0.17 83TU4 506 3136 (1307e19383) 0.76 0.15 351TU9 406 112 (76e165) 3.16 0.88 12TU11 373 168 (130e220) 2.60 0.37 19TU7 455 75093 (15144e390580) 2.78 2.18 8413PyridabenSAMB 659 6.4 (4.9e8.3) 2.57 0.46GSS 376 4.4 (3.7e5.3) 3.17 0.32 0.7TU6 502 16 (12e21) 2.10 0.24 2.6TU1 272 29 (15e44) 1.26 0.21 4.5TU2 312 71 (38e132) 1.52 0.37 11TU5 339 38 (28e48) 1.86 0.20 5.9TU12 217 62 (48e78) 2.65 0.37 9.8TU3 387 117 (63e214) 3.53 1.64 18TU4 388 26 (16e40) 2.09 0.45 4TU9 370 166 (129e217) 2.50 0.35 26TU11 829 248 (193e321) 1.05 0.08 39TU7 395 221 (177e269) 2.50 0.27 34FenazaquinSAMB 612 13 (11e15) 4.15 0.38GSS 281 17 (14e21) 4.44 0.65 1.3TU6 481 29 (15e59) 2.13 0.57 2.2TU1 581 179 (146e219) 1.94 0.16 13TU2 471 97 (30e307) 1.02 0.27 7.3TU5 299 49 (21e114) 2.30 0.89 3.7TU12 418 215 (143e322) 3.05 0.86 15TU3 429 77 (57e105) 1.81 0.18 5.7TU4 366 55 (41e74) 1.84 0.20 4.1TU9 508 316 (265e374) 2.55 0.25 23TU11 328 562 (145e2208) 1.39 0.70 42TU7 478 447 (324e619) 1.96 0.28 33Fenbutatin oxideSAMB 546 24 (1e546) 2.21 1.79GSS 346 18 (14e22) 2.72 0.33 0.7TU6 564 46 (25e84) 2.78 0.99 1.9TU1 351 131 (91e183) 1.34 0.18 5.5TU2 262 113 (74e182) 1.06 0.18 4.7TU5 390 158 (125e196) 2.21 0.25 6.6TU12 464 61 (47e81) 1.78 0.20 2.6TU3 318 27 (16e44) 3.97 1.78 1.1TU4 419 134 (96e188) 2.67 0.55 5.6TU9 554 47 (23e97) 1.14 0.25 2TU11 566 3511 (2118e7327) 0.80 0.13 146TU7 570 35784 (25333e54932) 0.96 0.11 1494AbamectinSAMB 318 0.22 (0.11e0.43) 4.2 3.61GSS 542 0.19 (0.11e0.32 3.73 1.48 0.8TU6 419 0.28 (0.02e3.53) 3.68 8.13 1.3TU1 461 0.14 (0.09e0.23) 2.68 0.74 0.6TU2 460 0.25 (0.22e0.28) 4.08 0.45 1.1TU5 498 0.39 (0.24e0.64) 4.01 1.33 1.8TU12 639 0.52 (0.21e1.3) 2.16 0.88 2.4
Table 2 (continued )
Population nb LC50 (95% CI) c Slope SE RRd
TU3 375 0.52 (0.43e0.64) 2.32 0.29 2.4TU4 499 0.36 (0.33e0.42) 5.34 0.68 1.6TU9 678 0.20 (0.11e0.37) 1.40 0.36 0.9TU11 558 0.21 (0.18e0.24) 4.22 0.41 0.9TU7 394 94 (66e122) 2.19 0.31 434Spirodiclofen LC95
SAMB 2096 2.2 (1.8e2.7) 3.02 0.37 7.8GSS 1658 1.4 (0.6e2.9) 4.26 3.41 4.0 0.6TU6 1198 0.5 (0.3e1.2) 2.68 0.99 2.3 0.2TU1 777 2.7 (1.3e5.7) 1.99 0.82 17.9 1.2TU2 1127 3.2 (2.4e4.3) 2.16 0.34 18.6 1.4TU5 676 2.5 (1.6e3.9) 6.17 6.37 4.6 1.1TU12 799 5.5 (4.7e6.3) 2.58 0.20 23.8 2.5TU3 643 1.6 (1.4e1.8) 3.33 0.29 5.0 0.7TU4 1171 2.5 (1.8e3.4) 3.25 0.84 8.0 1.1TU9 644 5 (2.6e9.7) 1.75 0.54 43.4 2.2TU11 1564 8.7 (7.7e10) 2.27 0.14 46.1 3.9TU7 931 27(19e39) 3.23 0.65 87.7 12ALL-14a 2493 9.5 (7e12.9) 3.11 0.57 32.1 4.3
a ALL is the T. urticae strain selected with spirodiclofen for 14 generations, see textfor details.
b Number of mites tested.c LC, lethal concentration expressed in mg/l.d RR, resistance ratio ¼ LC50 field sample/SAMB.
A. Ilias et al. / Crop Protection 42 (2012) 305e311308
population (TU6) displayed similar susceptibility with the tworeference laboratory strains SAMB and GSS. This population wascollected from eastern Crete on vegetables in a small organic familygarden surrounded by olive crops and it was isolated from intensivevegetable cultures. Populations TU1, TU2, TU5 and TU12 displayedrelatively low resistance levels (RR < 15) to all acaricides tested.Populations TU3, TU4 and TU9 displayed higher levels of resistanceto at least one of the acaricides tested: TU9 showed a 26 foldresistance to pyridaben and TU3 and TU4 showed 83- and 351-foldresistance to bifenthrin respectively. Not surprising the higherresistance levels were exhibited by TU11 and TU7 both collectedfrom glasshouse ornamental plants, with heavy insecticide treat-ment history. TU11 exhibited high levels of resistance to pir-imiphos-methyl (RR¼ 23), fenbutatin oxide (RR¼ 146) and the twoMETI acaricides (RR ¼ 42 for fenazaquin and RR ¼ 39 for pyr-idaben). TU7 exhibited high levels of resistance to all 5 chemicalgroups tested, with the higher RRs exhibited towards bifenthrin(RR ¼ 8413) and fenbutatin oxide (RR ¼ 1494). Moreover it was theonly population resistant to abamectin, with a resistance ratio of443-fold.
Pyrethroid, OP, METI, abamectin and fenbutatin oxide resistancein T. urticae is a worldwide phenomenon and it has been reportedfrom a number of different countries and crops (Van Leeuwen et al.,2010). Our study on T. urticae populations coming from differentparts of Greece showed that acaricide resistance is well establishedalso in Greek agro systems due to the heavy use of insecticides.
Target mutations have been found to be responsible forresistance to pyrethoids, OPs and abamectin in a number ofT. urticae strains (Tsagkarakou et al., 2009a; Khajehali et al., 2010;Kwon et al., 2010; Dermauw et al., 2012). For METIs and fenbu-tatin oxide target resistance has not been investigated as yet,however, the recent genome sequencing of T. urticae has providedthe tools for such analysis (Grbic et al., 2011). Besides targetresistance, several lines of evidence and especially the occurrenceof cross resistances point to the involvement of detoxificationenzymes in a number of resistance cases in T. urticae worldwide(Van Leeuwen et al., 2010). Metabolic detoxification, mainly byP450 monooxygenases, appeared to be involved in ketoenolresistance of T. urticae (Nauen and Konanz, 2005; Van Pottelbergeet al., 2009a).

A. Ilias et al. / Crop Protection 42 (2012) 305e311 309
3.2. Susceptibility of T. urticae to spirodiclofen
Slight decrease of susceptibility to spirodiclofen were recordedonly in four populations from continental Greece (RR ¼ 2e12),compared to the reference strain SAMB. The LC50s to spirodiclofenwere significantly correlated to LC50s to other 6 pesticides whenall the 12 T. urticae (ten Greek and two laboratory strains) wereconsidered (N ¼ 12, P < 0.05) but only three of these correlationsremained significant (between spirodiclofen and either METIs orwith fenbutatin oxide) when TU7 was removed from comparisons.The population TU7 exhibited the highest resistance ratio (12-fold)to spirodiclofen and very high RRs to all other pesticides. In additionTU7 had the heaviest spraying history (greenhouse roses) suggest-ing that an enhanced metabolism selected by the application ofother pesticide(s) possibly led to a reduced spirodiclofen suscepti-bility. Even so, the LC95 of TU7 (¼ 88 mg/l) is still below the rec-ommended field doses (96e144 mg/l) which explains the goodefficacy of this acaricide in field trials soon after our laboratoryexperiments (personal communication with the farmer). None ofthe populations of T. urticae from Crete showed cross-resistance tospirodiclofen. The lack of cross-resistance to spirodiclofen inT. urticae strains with very similar resistance patterns to otheracaricides, has previously been documented (Van Leeuwen et al.,2005). This further validated the results obtained in this study andconfirms that ketoenols are resistant breaking pesticides for spidermite populations in Europe.
Spirodiclofen was introduced in mite control programmes inGreece three years ago and so far no control failures were reported.Similarly, Nauen et al. (2002) reported full susceptibility of multi-resistant strains from different parts of the world to spirodiclofen.To examine the sustainability of spirodiclofen efficacy in the field,we established laboratory selections in the mixed strain ALL. Four-teen successive selections of ALL with spirodiclofen failed toincrease the resistance ratio: the LC50 of ALL_SP14 (the ALL strainafter 14 selections with spirodiclofen) was 9.5 mg/l (range 7e13 mg/l) and the LC50 of TU11, that displayed the higher LC50among the seven populations from which ALL derived from, was8.7 (8e10) mg/l. In line with our study, other artificial selectiontrials with spirodiclofen also revealed a low risk of resistancedevelopment (Rauch and Nauen, 2002). However, a laboratorystrain from Belgium exhibited high resistance levels to spirodiclofen(RR ¼ 274) after 1 year of selection (Van Pottelberge et al., 2009a),although the eggs were shown to be still quite susceptible. Besideslarva mortality, spirodiclofen has a pronounced effect on fecundityand fertility of female mites (Nauen, 2005; Kontsedalov et al., 2008).Van Pottelberge et al. (2009b) showed that in contrast to the
Table 3Log-dose probit mortality data for spiromesifen against B. tabaci populations collected in
Population Spiromesifen
na LC50 (95% CI) b Slope
GR165 2602 4.5 (4e5.1) 1.80GR187 2146 4.6 (4.1e5.1) 2.24GR191 2731 5.1 (3.1e8.6) 2.94GR157 1024 5.4 (2.5e11) 1.16GR188 1116 6.1 (5.4e6.7) 2.99GR194 3305 6.4 (5e8.1) 1.85GR259 2009 7.9 (7.2e8.6) 2.79GR163 3064 11.6 (9e15) 1.90GR164 1264 12 (7.1e19) 2.13GR258 1842 12 (8.8e16) 2.66GR192 2748 14 (12e15) 3.18GRMAL-RP 1252 1.1 (0.4e2.9) 0.98GRMAL-RN 1024 6.7 (5.1e8.8) 1.94
a Number of whiteflies tested.b LC, lethal concentration expressed in mg/l.c RR, resistance ratio, from Roditakis et al.(2006, 2009, 2010).
susceptible parental strain, the fecundity and fertility of resistantmites was not affected by treatment. In our study selections did notaffect female fecundity. The average number of larvae/per female inthe beginning of selections (ALL to ALLSP2, average number oflarvae/female ¼ 4.77 � 2.44) did not differ significantly from theaverage number of larvae/female at the end of selections (ALL_SP11to ALL_SP13, 4.89 � 1.80; ManneWhitney test P ¼ 0. 34).
High resistance levels to spirodiclofen have been also developedafter selections with this compound in Panonychus ulmi (RR> 7000,Kramer and Nauen, 2011) and Panonychus citri (RR ¼ 103, Yu et al.,2011) two other important plant feeding spider mite species. Thus,the potential of ketoenol resistance development appears to vary indifferent geographical areas and in different species.
3.3. Resistance of B. tabaci to spiromesifen
The toxicity of spiromesifen against B. tabaci populations withhigh levels of resistance against neonicotinoids and pyrethroids asdetermined in previous studies (Roditakis et al., 2009) was inves-tigated here. Due to the lack of susceptible strain it was not possibleto estimate the resistance levels, so the LC50s were used to comparethe tested populations. The eleven field collected B. tabaci pop-ulations exhibited low variation in the susceptibility to spi-romesifen (LC50 ¼ 4.5e14 mg/l) resulting in a 3-fold differencebetween the least and the most sensitive populations (Table 3).
The LC95s ranged from 19 to 85 mg/l and only in one population(GR157) it was over 100 mg/l. Given that the recommendedapplication rates for spiromesifen in Greece range from 96 to144 mg/l, which is above the LC95s determined in this study, it isexpected that spiromesifen would be efficient to control resistantB. tabaci populations. This is in line with the absence of field failurereports since the introduction of spiromesifen in the Greek market(4 years ago).
Absence of cross resistance was detected for spiromesifen inrelation to neonicotinoids and pyrethroids. By performing pair wisecomparisons of response (LC50) between spiromesifen and imida-cloprid and between spiromesifen and a-cypermethrin the Pearsonproductemoment correlation coefficient (R) was 0.046 and 0.214(n ¼ 11, P > 0.05) for the comparison of spiromesifen to imidaclo-prid and to a-cypermethrin respectively. LC50 data for imidaclopridand a-cypermethrin are published elsewhere (Roditakis et al.,2009). Furthermore the toxicity levels observed for the two resis-tant strains maintained to the laboratory under selection pressureGRMAL-RP resistant to a-cypermethrin and GRMAL-RN resistant toimidacloprid were comparable to that exhibited by the fieldcollected populations. The LC50s to spiromesifen were 1.1 and 6.7
Greece.
Imidacloprid a-cypermethrin
SE LC95 RRc RRc
0.10 37.16 73 2560.11 24.95 94 1010.87 18.62 120 1790.23 139.90 88 4260.23 21.41 353 1050.19 49.23 247 1420.13 30.74 323 7410.24 85.08 46 4780.47 69.44 7 420.35 49.19 369 11900.14 44.79 323 1790.21 50.31 >6210.18 47.61 958

A. Ilias et al. / Crop Protection 42 (2012) 305e311310
for GRMAL-RP and GRMAL-RN respectively thus further supportingthe absence of cross resistance.
At the time of experimentation spiromesifen has not beenregistered for pest control application in Greece, suggesting that ithad never been used against B. tabaci in the particular collectionarea. This is in agreement with the homogenous response of thefield collected populations to spiromesifen. Prabhaker et al. (2008)also performed baseline toxicity studies with spiromesifen onB. tabaci populations (type B) collected prior to the use of spi-romesifen in the field. In this study higher natural variability (29-fold) was reported.
Good performance of spiromesifen against immature stages hasbeen reported for greenhouse strains and field B- and Q-typeB. tabaci populations worldwide (Nauen and Konanz, 2005;Kontsedalov et al., 2008; Prabhaker et al., 2008; Fernandez et al.,2009). In Israel two years after the introduction of spiromesifenin the control programmes of B. tabaci no resistance was observed(Kontsedalov et al., 2008). In addition, the treatment of laboratorypopulations of B. tabaci (B- and Q-type) resistant to major insecti-cides, including neonicotinoids, pyriproxyfen and diafenthiuron,with spiromesifen showed no cross-resistance to these insecticides(Kontsedalov et al., 2008).
The selections of GR164 with spiromesifen did not result in anyincrease of the resistance levels after four successive selections. Incontrast a slight reduction in the susceptibility was observed. Thesurvival (13%) recorded at the last selection cycle with a concen-tration of 30 mg/l of spiromesifen for the selected population waslower than that of the parental population (27%) at the sameconcentration (c2 ¼ 11.34, df ¼ 1). Spiromesifen did not developresistance after three successive treatments in an imidaclopridresistant B. tabaci Q-type from Spain (Guthrie et al., 2003).
Taking together these results suggest that when used accordingto the manufacturer’s recommendations and in alteration withother chemical classes, ketoenols may be a valuable option for theimplementation of insecticide resistance management strategies tosustainably control B. tabaci in Greece.
In conclusion, ketoenols proved quite effective against T. urticaeand B. tabaci populations from Greece, which are resistant to otherpesticides. Moreover artificial selection with both chemicals did notincrease the resistance. The use of ketoenols in alternation withinsecticides with different mode of action classes as well as contin-uous monitoring of the efficacy situation should improve thesustainable chemical control of these two major pests in Greece.Genomic information (Wang et al., 2010; Grbic et al., 2011) for bothspecies will facilitate investigations of resistance mechanisms at themolecular level in the future aswell as the developmentof resistancediagnostics to be used in routine resistance monitoring initiatives.
Acknowledgements
The authors would like to thank A. Birouraki, E. Detsi, M. Hal-kiadaki, D. Katsirdaki, and M. Tsahelidou for their assistance in theexperiments and M. Navajas and A. Migeon (CBGP, Montpellier,France) for providing the SAMB strain. Part of this research wassupported by ‘HRAKLEITOS II’ grand to Aris Ilias (Operational Pro-gramme ‘Education and Life Long Learning’ co-funded by theEuropean Social Found and National Resources), by a grant from theHellenic Secretariat General for Research and Technology (THALIS)and by grants from Bayer Crop Science.
References
Bretschneider, T., Fischer, R., Nauen, R., 2007. Inhibitors of lipid synthesis (acetyl-CoA-carboxylase inhibitors). In: Krämer, W., Schirmer, U. (Eds.), Modern CropProtection Compounds. Wiley, Weinheim, pp. 909e925.
Denholm, I., Cahill, M., Dennehy, T.J., Horowitz, A.R., 1998. Challenges withmanaging insecticide resistance in agricultural pests, exemplified by thewhitefly Bemisia tabaci. Phil. Trans. R. Soc. (Lond. B) 353, 1757e1767.
Dermauw, W., Ilias, A., Riga, M., Tsagkarakou, A., Grbi�c, M., Tirry, L., Van Leeuwen, T.,Vontas, J., 2012. The cys-loop ligand-gated ion channel gene family of Tetra-nychus urticae: implications for acaricide toxicology and a novel mutationassociated with abamectin resistance. Insect Biochem. Mol. Biol. 42, 455e465.
Dinsdale, A., Cook, L., Riginos, C., Buckley, Y., De Barro, P.J., 2010. Refined globalanalysis of Bemisia tabaci (Gennadius) (Hemiptera: Sternorrhyncha: Aleyr-odoidea) mitochondrial CO1 to identify species level genetic boundaries. Ann.Entomol. Soc. Am. 103, 196e208.
Fernandez, E., Gravalos, C., Haro, P.J., Cifuentes, D., Bielza, P., 2009. Insecticideresistance status of Bemisia tabaci Q-biotype in south-eastern Spain. PestManag. Sci. 65, 885e891.
Finney, D.J., 1964. Probit Analysis. Cambridge University Press, Cambridge, UK.Grbic, M., Van Leeuwen, T., Clark, R.M., Rombauts, S., Rouze, P., Grbic, V.,
Osborne, E.J., Dermauw, W., Thi Ngoc, P.C., Ortego, F., Hernandez-Crespo, P.,Diaz, I., Martinez, M., Navajas, M., Sucena, E., Magalhaes, S., Nagy, L., Pace, R.M.,Djuranovic, S., Smagghe, G., Iga, M., Christiaens, O., Veenstra, J.A., Ewer, J.,Villalobos, R.M., Hutter, J.L., Hudson, S.D., Velez, M., Yi, S.V., Zeng, J.,PiresdaSilva, A., Roch, F., Cazaux, M., Navarro, M., Zhurov, V., Acevedo, G.,Bjelica, A., Fawcett, J.A., Bonnet, E., Martens, C., Baele, G., Wissler, L., Sanchez-Rodriguez, A., Tirry, L., Blais, C., Demeestere, K., Henz, S.R., Gregory, T.R.,Mathieu, J., Verdon, L., Farinelli, L., Schmutz, J., Lindquist, E., Feyereisen, R., Vande Peer, Y., 2011. The genome of Tetranychus urticae reveals herbivorous pestadaptations. Nature 479, 487e492.
Guthrie, F., Devine, G., Denholm, I., Nauen, R., 2003. Biological Evaluation of Spi-romesifen Against Bemisia tabaci and an Assessment of Resistance Risks. In:Proceedings of the BCPC Conference, vol. 2, pp. 795e800.
Jones, P.R., 2003. Plant viruses transmitted by whiteflies. Eur. J. Plant Pathol. 109,195e219.
Khajehali, J., Van Leeuwen, T., Grispou, M., Morou, E., Alout, H., Weill, M., Tirry, L.,Vontas, J., Tsagkarakou, A., 2010. Acetylcholinesterase point mutations inEuropean strains of Tetranychus urticae (Acari: Tetranychidae) resistant toorganophosphates. Pest Manag. Sci. 66, 220e228.
Knowles, C.O., 1997. Mechanisms of resistance to acaricides. In: Sjut, V. (Ed.),Molecular Mechanisms of Resistance to Agrochemicals. Springer, New York,pp. 57e77.
Kontsedalov, S., Gottlieb, Y., Ishaaya, I., Nauen, R., Horowitz, R., Ghanim, M., 2008.Toxicity of spiromesifen to the developmental stages of Bemisia tabaci biotypeB. Pest Manag. Sci. 65, 5e13.
Kramer, T., Nauen, R., 2011. Monitoring of spirodiclofen susceptibility in field pop-ulations of European red mites, Panonychus ulmi (Koch) (Acari: Tetranychidae),and the cross-resistance pattern of a laboratory-selected strain. Pest Manag. Sci.67, 1285e1293.
Kwon, D.H., Yoon, K.S., Clark, J.M., Lee, S.H., 2010. A point mutation in a glutamate-gated chloride channel confers abamectin resistance in the two-spotted spidermite, Tetranychus urticae Koch. Insect Mol. Biol. 19, 583e591.
Nauen, R., Bretschneider, T., Brück, E., Elbert, A., Reckmann, U., Wachendorff, U.,Tiemann, R., 2002. BSN 2060: A Novel Compound for Whitefly and Spider MiteControl. In: The BCPC Conference: Pests and Diseases. Proceedings of the Inter-national Conference, Brighton, UK, 18e21 November 2002, vol. 1, pp. 39e44.
Nauen, R., Bretschneider, T., Elbert, A., Fischer, R., Tiemann, R., 2003. Spirodiclofenand spiromesifen. Pestic. Outlook 12, 243e245.
Nauen, R., 2005. Spirodiclofen: mode of action and resistance risk assessment intetranychid pest mites. J. Pestic. Sci. 30, 272e274.
Nauen, R., Konanz, S., 2005. Spiromesifen as a new chemical option for resistancemanagement in whiteflies and spider mites. Pflanzenschutz Nachr. Bayer 58,485e502.
Prabhaker, N., Castle, S.J., Buckelew, L., Toscano, N.C., 2008. Baseline susceptibility ofBemisia tabaci B biotype (Hemiptera: Aleyrodidae) populations from Californiaand Arizona to spiromesifen. J. Econ. Entomol. 101, 174e181.
Rauch, N., Nauen, R., 2002. Spirodiclofen resistance risk assessment in Tetranychusurticae (Acari: Tetranychidae): a biochemical approach. Pest Biochem. Physiol.74, 91e101.
Raymond, M., Prato, G., Ratsira, D., 1993. Probit Analysis of Mortality Assays Dis-playing Quantal Response. Version 3.3 License 193019.
Roditakis, E., Tsagkarakou, A., Vontas, J., 2006. Identification of mutations in thepara sodium channel of Bemisia tabaci from Crete, associated with resistance topyrethroids. Pest Biochem. Physiol. 85, 161e166.
Roditakis, E., Grispou, M., Morou, E., Kristoffersen, J.B., Roditakis, N., Nauen, R.,Vontas, J., Tsagkarakou, A., 2009. Current status of insecticide resistance in Qbiotype Bemisia tabaci populations from Crete. Pest Manag. Sci. 65, 313e322.
Roditakis, E., Morou, E., Tsagkarakou, A., Riga, M., Nauen, R., Paine, M.J.I., Morin, S.,Vontas, J., 2010. Assessment of the Bemisia tabaci CYP6CM1vQ transcript andprotein levels in laboratory and field caught Imidacloprid resistant insects, andcross-metabolism potential of the recombinant enzyme. Insect Sci. 18, 25e29.
Tsagkarakou, A., Navajas, M., Lagnel, J., Gutierrez, J., Pasteur, N., 1996. Geneticvariability in Tetranychus urticae (Acari: Tetranychidae) from Greece: insecticideresistance and isozymes. J. Econ. Entomol. 89, 1354e1358.
Tsagkarakou, A., Pasteur, N., Cuany, A., Chevillon, C., Navajas, M., 2002. Mechanismsof resistance to organophosphates in Tetranychus urticae (Acari: Tetranychidae)from Greece. Insect Biochem. Mol. Biol. 32, 417e424.
Tsagkarakou, A., Van Leeuwen, T., Khajehali, J., Ilias, A., Grispou, M.,Williamson, M.S., Tirry, L., Vontas, J., 2009a. Identification of pyrethroid

A. Ilias et al. / Crop Protection 42 (2012) 305e311 311
resistance mutations in the para sodium channel of the two spotted spider miteTetranychus urticae (Acari: Tetranychidae). Insect Mol. Biol. 18, 583e593.
Tsagkarakou, A., Nikou, D., Roditakis, E., Sharvit, M., Morin, S., Vontas, J., 2009b.Molecular diagnostics for detecting pyrethroid and organophosphate resistancemutations in the whitefly Bemisia tabaci (Hemiptera: Aleyrodidae). Pest Bio-chem. Physiol. 94, 49e54.
Van Leeuwen, T., Van Pottelberge, S., Tirry, L., 2005. Comparative acaricidesusceptibility and detoxifying enzyme activities in field-collected resistant andsusceptible strains of Tetranychus urticae. Pest Manag. Sci. 61, 499e507.
Van Leeuwen, T., Vontas, J., Tsagkarakou, A., Dermauw, W., Tirry, L., 2010. Acaricideresistance mechanisms in the two-spotted spider mite Tetranychus urticae andother important Acari: a review. Insect Biochem. Mol. Biol. 40, 563e572.
Van Pottelberge, S., Van Leeuwen, T., Khajehali, J., Tirry, L., 2009a. Geneticand biochemical analysis of a laboratory-selected spirodiclofen-resistant
strain of Tetranychus urticae Koch (Acari: Tetranychidae). Pest Manag. Sci. 65,358e366.
Van Pottelberge, S., Khajehali, J., Van Leeuwen, T., Tirry, L., 2009b. Effects of spi-rodiclofen in a susceptible and resistant of Tetranychus urticae Koch (Acari:Tetranychidae). Exp. Appl. Acarol. 47, 301e309.
Wachendorff, U., Nauen, R., Schnorbach, H.J., Rauch, N., Elbert, A., 2002. The bio-logical profile of spirodiclofen (Envidor�) e a new selective tetronic acidacaricide. Pflanzensch Nachr. Bayer 55, 149e176.
Wang, X.W., Luan, J.B., Li, J.M., Bao, Y.Y., Zhang, C.X., Liu, S.S., 2010. De novo char-acterization of a whitefly transcriptome and analysis of its gene expressionduring development. BMC Genomics 11, 400.
Yu, D.Y., Wang, C.F., Yu, Y., Huang, Y.Q., Yao, J.A., Hu, J.F., 2011. Laboratory selectionfor spirodiclofen resistance and cross-resistance in Panonychus citri. African J.Biotech. 10, 3424e3429.


















