
Sermons, by the Rev. John Ewing, D.D., later pastor of the First ...

This article was downloaded by: [UNAM Ciudad Universitaria]On: 11 August 2011, At: 11:48Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
International Journal of AcarologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/taca20
Presence of intradermal chigger mite Hannemaniahylae (Ewing, 1925) (Acari: Leeuwenhoekiidae) in thetroglophile frog Eleutherodactylus longipes (Anura:Brachycephalidae) at Los Riscos Cave, Querétaro,MexicoAdriana Espino del Castillo a , Ricardo Paredes-León b & Juan B. Morales-Malacara a ca Laboratorio de Acarología “Anita Hoffmann”, Facultad de Ciencias, Universidad NacionalAutónoma de México, Avenida Universidad 3000, Ciudad Universitaria, C. P., 04510, DistritoFederal, Méxicob Colección Nacional de Ácaros, Instituto de Biología, Universidad Nacional Autónoma deMéxico, Avenida Universidad 3000, Ciudad Universitaria, C. P., 04510, Distrito Federal,Méxicoc Unidad Multidisciplinaria de Docencia e Investigación, Facultad de Ciencias, UniversidadNacional Autónoma de México, Campus Juriquilla, Boulevard Juriquilla 3001, C. P., 76230,Querétaro, México
Available online: 11 Aug 2011
To cite this article: Adriana Espino del Castillo, Ricardo Paredes-León & Juan B. Morales-Malacara (2011): Presenceof intradermal chigger mite Hannemania hylae (Ewing, 1925) (Acari: Leeuwenhoekiidae) in the troglophile frogEleutherodactylus longipes (Anura: Brachycephalidae) at Los Riscos Cave, Querétaro, Mexico, International Journal ofAcarology, 37:5, 427-440
To link to this article: http://dx.doi.org/10.1080/01647954.2010.525522
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching and private study purposes. Any substantial or systematicreproduction, re-distribution, re-selling, loan, sub-licensing, systematic supply or distribution in any form toanyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses shouldbe independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims,proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly inconnection with or arising out of the use of this material.

International Journal of AcarologyVol. 37, No. 5, October 2011, 427–440
PRESENCE OF INTRADERMAL CHIGGER MITE HANNEMANIA HYLAE(EWING, 1925) (ACARI: LEEUWENHOEKIIDAE) IN THE TROGLOPHILE FROG
ELEUTHERODACTYLUS LONGIPES (ANURA: BRACHYCEPHALIDAE)AT LOS RISCOS CAVE, QUERÉTARO, MEXICO
Adriana Espino del Castillo1, Ricardo Paredes-León2 and Juan B. Morales-Malacara1,3
1. Laboratorio de Acarología “Anita Hoffmann”, Facultad de Ciencias, Universidad Nacional Autónoma deMéxico. Avenida Universidad 3000, Ciudad Universitaria, C. P. 04510; Distrito Federal, México
(e-mail: [email protected]); 2. Colección Nacional de Ácaros, Instituto de Biología, UniversidadNacional Autónoma de México. Avenida Universidad 3000, Ciudad Universitaria, C. P. 04510; Distrito Federal,
México (e-mail: [email protected]); 3. Unidad Multidisciplinaria de Docencia e Investigación, Facultadde Ciencias, Universidad Nacional Autónoma de México, Campus Juriquilla. Boulevard Juriquilla 3001,
C. P. 76230, Querétaro, México (e-mail: [email protected]).
(Received 11 December 2009; accepted 7 September 2010)
ABSTRACT – For the first time, we report the presence of Hannemania hylae (Ewing) parasitizingthe long-footed robber frog Eleutherodactylus longipes (Baird) in a cave environment in QueretaroState, Mexico. A morphological comparison was performed between H. hylae and its closest speciesHannemania bufonis Loomis and Welbourn and Hannemania monticola Welbourn and Loomis. Themain characters to separate these species are the size of ocular plate, the length of some scutal setae,and the shape and length of some tarsal setae of the legs I and II. We inspected 47 frogs, the prevalenceof infestation was 23.4%, mean abundance was 1.8 mites per host, mean intensity 7.9 mites per infestedhost, and the range intensity was 1–30 mites. Larvae of H. hylae were only collected during the dryseason. A brief discussion is provided about the period of larval stage on the host and seasonality iscompared with similar studies.Key words – Acari, Leeuwenhoekiidae, Hannemania, Anura, Eleutherodactylus, Mexico, cave.
INTRODUCTION
Chigger mites of the genus HannemaniaOudemans, are endoparasites in amphibians (frogs,toads, and salamanders) occurring in the Americasfrom the USA to Argentina. There are approx-imately 25 described species (Alzuet and Mauri,1985 (1987); Wohltmann et al., 2006), and sevenhave been recorded from Mexico: Hannemania anu-rae Welbourn and Loomis, Hannemania bufonisLoomis and Welbourn, Hannemania hylae (Ewing),Hannemania mexicana Hoffmann, Hannemania mon-ticola Welbourn and Loomis, Hannemania pelaeziHoffmann, and Hannemania saxicola Welbourn
and Loomis. However, because of morphologicalsimilarities Hoffmann (1990) considered thatH. bufonis and H. monticola could be synonymsof H. hylae.
The larva of H. hylae was described by Ewing(1925) as Trombicula hylae from the type host Hylaarenicolor Cope at Cottonwood Creek, San DiegoCounty, CA, USA [the population of this locality isnow recognized as a different taxon of Hyla areni-color and is named Pseudacris cadaverina (Cope), sensuDuellman (2001)]. The original description is veryshort, and lacks figures and many important taxo-nomical characters and measurements. Shortly there-after, Ewing (1926) described the nymph and an adult
ISSN 0164-7954 print/ISSN 1945-3892 online© 2011 Taylor & Francis; printed 11 August 2011DOI: 10.1080/01647954.2010.525522http://www.informaworld.com
Dow
nloa
ded
by [
UN
AM
Ciu
dad
Uni
vers
itari
a] a
t 11:
48 1
1 A
ugus
t 201
1

428 Espino del Castillo et al. 2011
(without specifying sex) of this species, with an outlineof its life history, and an illustration of an engorgedlarva. To date, there have been at least three redescrip-tions or reviews of the larva of H. hylae: Hoffmann(1952), based on topotypes of the same sample as theholotype deposited at United States National MiteCollection (USNMC) but without measurements ofimportant taxonomic characters of the scutum; Gould(1956) redescribed H. hylae from the holotype andother specimens from California; and Loomis andWelbourn (1969) redescribed the species from topo-types. However, the available information regardingH. hylae has never been compiled to aptly describe thelarva, given the considerable variation among differentpopulations.
This chigger has been recorded on several hostsin the USA: Arizona and Utah ex Hyla areni-color, California ex Pseudacris cadaverina, and Texasex Hyla arenicolor and Eleutherodactylus guttilatus(Cope) (Loomis and Welbourn, 1969; Jung et al.,2001); Mexico: Baja California ex Pseudacris cadav-erina, Morelos ex Lithobates berlandieri (Baird) andEleutherodactylus sp., San Luis Potosí ex Lithobatessp. and, Sonora ex Hyla arenicolor, Scaphiopus couchiiBaird and Incilius mazatlanensis (Taylor) (Hoffmann,1969, 1990; Loomis and Welbourn, 1969; ; Hoffmannand López-Campos, 2000; Goldberg et al., 2002);and, Argentina: Misiones ex Hypsiboas curupi García,Faivovich and Haddad (Alzuet and Mauri, 1985(1987); García et al., 2007).
The long-footed robber frog Eleutherodactyluslongipes (Baird), of the family Brachycephalidae, isendemic to Mexico, being distributed along the SierraMadre Oriental in isolated localities of the states ofCoahuila, Nuevo León, Tamaulipas, Hidalgo, SanLuis Potosí, and Querétaro (Lynch, 1970; Lemos-Espinal and Smith, 2007; Frost, 2009; Santos-Barrera& Canseco-Márquez, 2009). It inhabits moderateelevations from 350 to 2000 m above sea level, andshows a strong tendency to occupy cave environments(Taylor, 1939; Lynch, 1970; Lemos-Espinal and Smith,2007; Santos-Barrera and Canseco-Márquez, 2009)and its partial endogenous circadian rhythmicityconfirms its troglophilic nature (Espino del Castilloet al., 2009).
The aims of this contribution are to report, forthe first time, the parasitism of the frog E. longipes bythe chigger mite H. hylae, to describe the infestationlevel, to compare it with similar studies, and, addition-ally, to compare this chigger with its closest speciesH. bufonis and H. monticola.
MATERIALS AND METHODS
Locality – Los Riscos Cave (21◦11′38′′N, 99◦30′50′′W) belongs to the El Abra limestone or karst
formation, and its geographical location is in the SierraGorda, State of Querétaro, Mexico (Alencaster et al.,1999). It is located 3 km north-east from Puente deDios up the river Jalpan, at 1122 m above sea level.
This cave represents a mixed underground system(horizontal and vertical) with eight zones, includingtunnels and galleries. The cave is mainly horizontal,with a depth of 35 m and a longitude of 550 m (Fig. 1)(Lazcano-Sahagún, 1986; Espino del Castillo et al.,2009). A detailed description of the cave can be foundin Espino del Castillo et al. (2009).
Field techniques – We carried out nine visitsto the cave, each consisted of 2 days of samplingof amphibians and their associated mites (November2005; April, June, October, and December 2006;March and October 2007; October 2008; April 2009).The search for frogs was carried out along each ofthe cave zones during day – twilight periods. In thezones where the frogs were encountered puddles orpools of water did not occur and the frogs weremore commonly observed resting on top of wallsor crevices and in some cases under rocks (Figs. 2and 3). Some frogs were examined for parasites insitu, others were collected and transported to thelaboratory to confirm identification, and to examinethem for parasites. Frog specimens were deposited atUniversidad Nacional Autónoma de México (UNAM)in the Museo de Zoología, Facultad de Ciencias(MZFC) and Colección Nacional de Anfibios yReptiles, Instituto de Biología, UNAM (CNAR), andthe mites are deposited at the Colección Nacionalde Ácaros, Instituto de Biología, UNAM (CNAC),and at Morales-Malacara (MM) Collection, housedat Laboratorio de Acarología, Facultad de Ciencias,UNAM.
Laboratory techniques – The frogs were exam-ined under a stereoscopic microscope (OlympusSZ series). Chigger mites were only found onlimbs and digits of their hosts, in cavities belowthe cuticle (Fig. 4). Chiggers were excised withneedles.
Some mites (31 specimens, CNAC006517;006527–006529; 006672–006698) were cleared withlactophenol and mounted on microscope slides insemi-permanent mounts (Hoyer’s medium). Theset of the measurements of the scutum setae anddistances between setae was made following Goffet al. (1982): AW (anterior width), PW (posteriorwidth), SB (distance between sensillary bases), ASB(distance from sensillary bases to extreme anteriormargin), PSB (distance from sensillary bases toextreme posterior margin), AP (distance from antero-lateral setal base to posterolateral setal base on oneside), AM (length of anteromedian seta), AL (lengthof anterolateral seta), PL (length of posterolateralseta) and S (length of sensilla). Measurements andobservations of diagnostic characters were carried out
Dow
nloa
ded
by [
UN
AM
Ciu
dad
Uni
vers
itari
a] a
t 11:
48 1
1 A
ugus
t 201
1

Vol. 37, No. 5 Internat. J. Acarol. 429
Fig. 1. Entry of Los Riscos Cave.
Dow
nloa
ded
by [
UN
AM
Ciu
dad
Uni
vers
itari
a] a
t 11:
48 1
1 A
ugus
t 201
1

430 Espino del Castillo et al. 2011
Fig. 2. The main frog habitat in a wall crevice inside Los Riscos Cave.
Dow
nloa
ded
by [
UN
AM
Ciu
dad
Uni
vers
itari
a] a
t 11:
48 1
1 A
ugus
t 201
1

Vol. 37, No. 5 Internat. J. Acarol. 431
Fig. 3. Eleutherodactylus longipes (Baird) in Los Riscos Cave.
Dow
nloa
ded
by [
UN
AM
Ciu
dad
Uni
vers
itari
a] a
t 11:
48 1
1 A
ugus
t 201
1
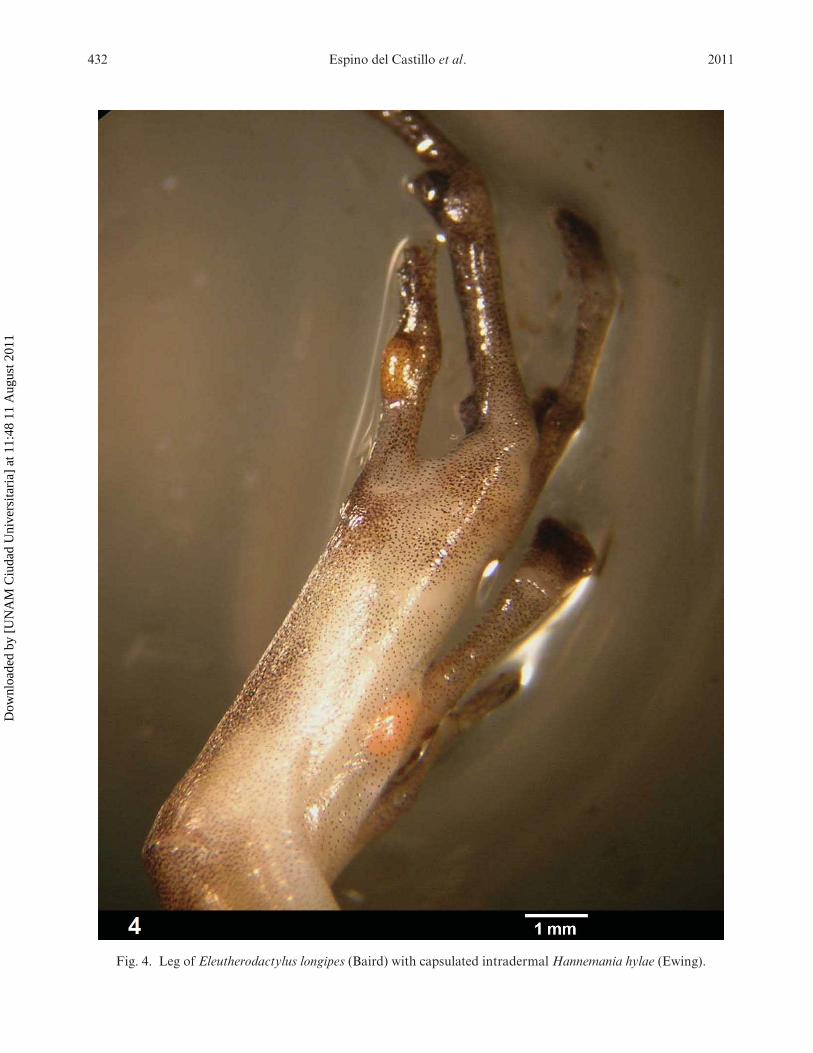
432 Espino del Castillo et al. 2011
Fig. 4. Leg of Eleutherodactylus longipes (Baird) with capsulated intradermal Hannemania hylae (Ewing).
Dow
nloa
ded
by [
UN
AM
Ciu
dad
Uni
vers
itari
a] a
t 11:
48 1
1 A
ugus
t 201
1

Vol. 37, No. 5 Internat. J. Acarol. 433
with a Nikon Optiphot 2 (Japan) phase-contrast anddifferential-interference-contrast microscope.
The identification of the mites was made consult-ing specialized literature (Loomis and Welbourn, 1969;Brennan and Goff, 1977; Hoffmann, 1990; Krantz andWalter, 2009). It was not possible to obtain all the mea-surements from all the mounted specimens because ofinconsistencies in the mounting process, and becausethe cuticle of the mites was broken in some cases.Unmounted mites (CNAC006699–006702) were pre-served in vials with absolute ethanol. Some of thesewere prepared for imaging using a scanning electronicmicroscope (JSM-5310LV, JEOL). Also, we extend ourobservations by checking a few additional specimens,including those analysed by Hoffmann (1952), referredtoasH.hylaeanddepositedatCNAC(006703–006705).The infestation level was analysed according to theparameters proposed by Bush et al. (1997).
In addition to specimens of H. hylae, weexamined and measured seven paratype specimensof H. bufonis deposited at Ohio State AcarologyLaboratory (OSAL0081587–0081593) at Ohio StateUniversity, Columbus, OH, USA during a visit to thatlaboratory. These specimens were observed and mea-sured using a Zeiss Axioscope (Germany) microscope.All measurements are in micrometers. Type specimensof H. monticola were not available to review.
RESULTS
Taxonomical notes – Eleutherodactylus longipesfrom los Riscos Cave was parasitized only withH. hylae (Ewing, 1925) (Figs. 5 and 6). The specimensmatch the description and redescriptions of H. hylaein some important measurements of scutum, such asAP distance and AM, PL and S setae. However, thesemites are slightly larger in AW, PW, SB, PSB distancesand slightly shorter in ASB distance and AL setae(Table 1). Additional characters of the leg chaetotaxy(not shown) also match with data published by Loomisand Welbourn (1969).
According to Loomis and Welbourn (1969),Welbourn and Loomis (1970) and data obtained fromH. hylae from Los Riscos Cave, this species resem-bles H. bufonis and H. monticola; but differs fromH. monticola in having PL shorter 35–56 (63–79 inH. monticola) and differs from H. bufonis in havingparasubterminala I nude, tarsala I subequal to tarsalaII, PL short (less than 55), AP wide (17 or more), PWless than 64, PSB less than 25 and, with a large ocularplate (Fig. 7).
With the data obtained from specimens ofH. hylae from Los Riscos Cave we made a compar-ison of the main characters used to separate it from
its closest species, H. bufonis (based on paratypesdeposited at OSAL) and H. monticola (with data of theoriginal description) (Table 2).
Ecological notes and infestation level – Weencountered frogs on all sampling occasions exceptone (October 2007). Of 47 frogs examined for endopar-asitic chiggers, 11 were found to be parasitized (preva-lence 23.4%). From these 11 frogs were collected87 mites, all of the species H. hylae (mean abun-dance: 1.8 mites per host and mean intensity: 7.9 mitesper infested host). The range of intensity was fromone mite to 30 mites per frog. Larvae of H. hylaewere only encountered during the dry season [April2006 (19 mites), March 2007 (12 mites), and April2009 (56 mites)]. These results are different to thoseobtained in previous studies with the three speciesof Hannemania considered here (Table 3). All frogswith mites were found in a zone of complete dark-ness except for a single frog that was found at theentrance of the cave (see Espino del Castillo et al.,2009 for details of the distribution of the frogs insidethe cave). The mites were more abundant on digits andthighs than any other region of the body of E. longipes(Fig. 4).
DISCUSSION
The presence of H. hylae in the long-footed rob-ber frog E. longipes represents a new host record andextends the range of H. hylae in the State of Querétaro.This chigger shows a wide range of host utilization:it has been collected in association with frog speciesof more than one family. However, despite the appar-ent disjunct distribution and wide host range, H. hylaeexhibits minimal morphological variation throughoutits various hosts and localities (at least between thepopulations from Querétaro and California) (Table 1).The variation in length of ASB (Table 1) observed invarious studies is probably attributable to inconsisten-cies in the methodology for measuring this structure(for example, Loomis and Welbourn (1969) consideredthe nasus of scutum, unlike the other studies). Thesingle record of H. hylae from Argentina needs to becorroborated.
The specimens from San Luis Potosi, Mexicoreferred by Hoffmann (1952) as H. hylae correspondto H. monticola, based on the diagnostic character pro-posed by Welbourn and Loomis (1970) to separate thisspecies from H. hylae (length of the PLs setae 63–79).In addition, after revision of two of the five speci-mens reported by Hoffmann (1952), we observed thatthese specimens also coincide with other charactersof H. monticola in length of AM, AL, S, the size oftarsalae I and II, and parasubterminala.
Dow
nloa
ded
by [
UN
AM
Ciu
dad
Uni
vers
itari
a] a
t 11:
48 1
1 A
ugus
t 201
1

434 Espino del Castillo et al. 2011
Fig. 5. Hannemani hylae (Ewing), dorsal view (larva).
In relation to the morphological comparisonsbetween H. hylae and H. bufonis we can conclude thatthese species are very close and that the main differ-ences were found in the size of the ocular plate, somescutal setae, tarsalae I and II, and the form of para-subterminalae. The ocular plate is larger in H. hylae(37 × 19) than H. bufonis (24 × 11), tarsala I is shorterthan tarsala II in H. hylae (20 < 22), but tarsala I islonger than tarsala II in H. bufonis (21 > 19). Withregard to H. monticola, the main differences could beseen in the size of ocular plate (larger in H. hylae), thelength of AM, AL, PL, tarsalae I and II, subterminalaI, and pretarsala II (Table 2).
However, according to Hoffmann (1990) thesedifferences could be considered as intraspecific varia-tion, and in this case H. bufonis and H. monticola fallin synonym with H. hylae. This hypothesis needs to be
investigated with a detailed analysis of type specimensand multiple samples of the three species, and perhapsmolecular evidence.
With regard to the body sites preferred byH. hylae, mites were more abundant among digits andthighs that any region of the body of E. longipes.This has been reported previously by Ewing (1926)for H. hylae on Pseudacris cadaverina and by Junget al. (2001) for mites of Hannemania sp. fromLithobates berlandieri and Anaxyrus punctatus (Bairdand Girard), and H. hylae from Eleutherodacylus guti-latus and Hyla arenicolor. Also Malone and Paredes-León (2005) obtained H. monticola mites from thehind legs and hind feet of Eleutherodactylus marnockii.Furthermore, Hatano et al. (2007) report that theregions most commonly infested (by undeterminedHannemania) in Hylodes phyllodes Heyer and Cocroft
Dow
nloa
ded
by [
UN
AM
Ciu
dad
Uni
vers
itari
a] a
t 11:
48 1
1 A
ugus
t 201
1

Vol. 37, No. 5 Internat. J. Acarol. 435
Fig. 6. Hannemania hylae (Ewing), lateral view (larva).
were the lateral body and the hind limbs, particularlythe thighs, dorsal leg, and lateral and dorsal body. Theconcentration of mites in these particular body partsmay be the result of their propensity for coming in con-tact with the surfaces that unengorged larvae inhabit(Ewing, 1926).
The infestation levels from the published reportsof the three Hannemania species considered in thisstudy, compared with our own data (Table 3), havevarious differences that can be attributed to severalfactors, such as host characteristics (size, habits, habi-tat preferences, home range, population size), the studylocality (geographical location, weather, vegetation,and physiographic features), collection effort, sam-ple size, the seasonal availability of host species, andcertainly the habits of the chigger mite. For exam-ple, at Los Riscos Cave we have a small sample of
E. longipes (n = 47) with a low level of infestation byH. hylae (23.4% with 1–30 mites), but Jung et al. (2001)reported a high infestation level by the same chiggermite (81% with 1–105 mites) in a smaller sample ofHyla arenicolor (n = 25). They proposed two hypothe-ses to explain the high infestation levels (comparingtwo different populations of another host and itsmites), the first relating to the higher densities of frogsin one site that could elicit higher transmission anddensities of parasite chiggers in the zone and the sec-ond relating to the better survival of post-larval instarsin certain habitats (Jung et al., 2001). We consideredthese hypotheses as they relate to E. longipes at LosRiscos Cave and Hyla arenicolor in Texas, USA; forthe first hypotheses the breeding aggregation habit ofHyla arenicolor (Canseco-Marquez, personal commu-nication) should favor a greater abundance of H. hylae,
Dow
nloa
ded
by [
UN
AM
Ciu
dad
Uni
vers
itari
a] a
t 11:
48 1
1 A
ugus
t 201
1

436 Espino del Castillo et al. 2011
Tab
le1.
Scut
alm
easu
rem
ents
ofla
rvae
ofH
anne
man
iahy
lae
from
diff
eren
tlo
calit
ies.
Loc
alit
yan
dho
stN
AW
PW
SBA
SBP
SBA
PA
MA
LP
LS
Ref
eren
ce
San
Ber
nard
ino
Co.
,C
A,U
SA1
4560
2234
2415
3334
5176
Gou
ld(1
956)
exP
seud
acri
sca
dave
rina
San
Die
goC
o.,C
A,
USA
148
6625
3422
1730
3450
75G
ould
(195
6)
exP
seud
acri
sca
dave
rina
San
Die
goC
o.,C
A,
USA
(Hol
otyp
e)1
4560
2331
2418
26–
––
Gou
ld(1
956)
exP
seud
acri
sca
dave
rina
San
Die
goC
o.,C
A,
USA
(Top
otyp
es)
2147
62.2
25.2
52.6
2419
.526
.631
44.1
70L
oom
isan
dW
elbo
urn
(196
9)ex
Pse
udac
ris
cada
veri
na
(44–
49)
(59–
65)
(23–
29)
(50–
55)
(23–
25)
(17–
21)
(24–
30)
(28–
34)
(39–
49)
(68–
78)
San
Die
goC
o.,C
A,
USA
(Top
otyp
e)1
47.1
60.1
23.6
29.8
22.3
1827
.330
.441
.565
CN
AC
0067
03T
his
stud
yex
Pse
udac
ris
cada
veri
naQ
ueré
taro
,Mex
ico
3150
.669
.927
.230
.829
2026
.228
.345
.179
Thi
sst
udy
exE
leut
hero
dact
ylus
long
ipes
(46–
59)
(64–
77)
(23–
32)
(28–
37)
(25–
31)
(17–
24)
(20–
32)
(21–
37)
(35–
56)
(62–
87)
Dow
nloa
ded
by [
UN
AM
Ciu
dad
Uni
vers
itari
a] a
t 11:
48 1
1 A
ugus
t 201
1

Vol. 37, No. 5 Internat. J. Acarol. 437
Fig. 7. Hannemania hylae (Ewing), dorsal scutum and ocular plates.
unlike the isolated environment and solitary habits ofE. longipes at Los Riscos Cave. Additionally, Lemos-Espinal and Smith (2007) suggest that some popu-lations of Hyla arenicolor (a desert-adapted species)could have close contact with water after exposure toextreme sunlight, and we considered that this behav-ior could be extended to mud or humid soil, whichwould be more suited to contact with the chiggers(which prefer the soil habitats) compared with themain sites occupied by E. longipes, which are rockcrevices and wall caves. The second hypothesis is alsolikely, given that the soil at Los Riscos Cave is notan adequate habitat for the growth of nymphs andadults; these stages were never observed inside the cave.Therefore, we considered that both hypotheses couldbe applicable to our own data. The low density offrogs at Los Riscos cave produces low transmission
and densities of chiggers, supporting the hypothesis ofhigher transmission of parasites when the host popula-tion is high, and for the second hypothesis also, the lowdensity of chiggers is the result of the soil of the cavenot being a suitable habitat for post-larval stages ofH. hylae.
Other studies reporting the infestation levels byHannemania larvae were not considered in our com-parisons (Duszynski and Jones, 1973; Hatano et al.,2007), because, in those studies, mites were not identi-fied to species level. Infestations in which two speciesof Hannemania coexist on the same host animalhave been reported, for example H. monticola andH. saxicola (Welbourn and Loomis, 1970). This phe-nomenon creates uncertainty when interpreting datafrom studies in which chiggers were not identified tospecies.
Dow
nloa
ded
by [
UN
AM
Ciu
dad
Uni
vers
itari
a] a
t 11:
48 1
1 A
ugus
t 201
1

438 Espino del Castillo et al. 2011
Tab
le2.
Com
pari
son
ofth
em
ean
mor
phol
ogic
aldi
ffer
ence
sam
ong
Han
nem
ania
bufo
nis,
Han
nem
ania
hyla
e,an
dH
anne
man
iam
onti
cola
.
Scut
umL
egI
Leg
II
Ocu
lar
Pal
pal
Mic
roP
aras
ub-
Sub-
Mic
ro-
Spec
ies
plat
eta
rsal
aA
PA
MA
LP
LT
arsa
lata
rsal
ate
rmin
ala
term
inal
aT
arsa
lage
nual
aP
reta
rsal
a
H.m
onti
cola
30×
1312
2135
4176
265
Nud
e26
266
10(d
ata
ofW
elbo
urn
and
Loo
mis
,197
0)
(19–
25)
(31–
45)
(39–
47)
(63–
79)
H.b
ufon
is24
×11
9.2
15.3
3236
.165
.221
4.7
Bra
nche
d22
.18
195.
1112
.3O
SAL
para
type
s(7
–11)
(14–
17)
(28–
40)
(31–
39)
(62–
68)
(4–5
)(2
1–24
)(4
–6)
(12–
14)
H.h
ylae
Los
37×
1912
2026
.228
4520
3.4
Nud
e18
.522
4.1
14.3
Ris
cos
Cav
e(9
–14)
(17–
24)
(20–
32)
(21–
36)
(35–
56)
(2–5
)(1
2–22
)(2
–5)
(9–2
0)
Tab
le3.
Infe
stat
ion
leve
lsby
Han
nem
ania
bufo
nis,
Han
nem
ania
hyla
e,an
dH
anne
man
iam
onti
cola
repo
rted
indi
ffer
ent
stud
ies.
Par
asit
eP
reva
lenc
eM
ean
Mea
nH
osts
Loc
alit
ysp
ecie
sn
(%)
inte
nsit
yab
unda
nce
Ran
geSo
urce
Pse
udac
ris
cada
veri
naC
A,U
SAH
.hyl
ae25
698
.9–
21.4
8–
37W
elbo
urn
and
Loo
mis
(197
5)A
naxy
rus
punc
tatu
sC
A,U
SAH
.buf
onis
131
46.8
–8
2–
9.6
Wel
bour
nan
dL
oom
is(1
975)
Ele
uthe
roda
ctyl
usgu
ttila
tus
TX
,USA
H.h
ylae
366
.6a
16.6
a25
a12
–38
Jung
etal
.(2
001)
Hyl
aar
enic
olor
TX
,USA
H.h
ylae
2581
––
3–
105
Jung
etal
.(2
001)
Inci
lius
maz
atla
nens
isSo
nora
,Mex
ico
H.b
ufon
is20
202b
–1
–4
Gol
dber
get
al.
(200
2)In
ciliu
sm
azat
lane
nsis
Sono
ra,M
exic
oH
.hyl
ae20
705b
–1
–12
Gol
dber
get
al.
(200
2)L
itho
bate
sta
rahu
mar
aeSo
nora
,Mex
ico
H.m
onti
cola
4233
.35b
–4
–8
Gol
dber
get
al.
(200
2)E
leut
hero
dact
ylus
mar
nock
iiT
X,U
SAH
.mon
tico
la94
8127
.6a
22.4
1–
152
Mal
one
and
Par
edes
-Leó
n(2
005)
Ele
uthe
roda
ctyl
uslo
ngip
esQ
ueré
taro
,Mex
ico
H.h
ylae
4723
.47.
91.
81
–30
Thi
sst
udy
Not
es:a
Dat
aca
lcul
ated
wit
hin
form
atio
nfr
omre
spec
tive
arti
cles
;bD
ata
prov
ided
as“m
ean”
wit
hout
spec
ifyi
ngw
heth
erm
ean
inte
nsit
yor
mea
nab
unda
nce.
Dow
nloa
ded
by [
UN
AM
Ciu
dad
Uni
vers
itari
a] a
t 11:
48 1
1 A
ugus
t 201
1

Vol. 37, No. 5 Internat. J. Acarol. 439
We observed larvae of H. hylae (March–April2006–2009) on E. longipes, during months thatcoincide with the driest period at Los Riscos Cave(December–May) (CETENAL, 1986; CNA, 2004).Apparently, the larval stage of H. hylae is adaptedto the dry conditions found at Los Riscos Cave and,as a population, possibly possesses a shorter para-sitic period because of the relatively constant condi-tions (temperature and humidity) around Los RiscosCave. However, this short annual larval period con-trasts with other studies of H. hylae and closely relatedspecies. Welbourn and Loomis (1975) reported par-asitism by H. hylae during March–November 1969–1970 in Joshua Tree National Monument, CA, USA,and conclude that H. hylae has an extended period onthe host during the summer. Jung et al. (2001) encoun-tered H. hylae on frogs at Big Bend National Park, TX,USA during May–October 1998–1999. These peri-ods include the hottest months (May–June) and therainy season (mid-July through late September). Someparts of Big Bend National Park receive no precipi-tation (National Park Services, 2009). We think thatthe extended larval stage in the latter localities couldbe related to extreme conditions (dry and hot) in thesummer period. The mites extend their parasitic phaseuntil the rainy seasons or when the conditions oftemperature and humidity are better for the survivalsuccess of the free-living post-larval stages. This sup-position needs to be investigated with a detailed studyin different populations of H. hylae.
ACKNOWLEDGEMENTS
We are grateful to biologists Sara Soriano,Stephanie Ortega, Itzel Sigala, Miguel Hernández,Marisol Vega, Yunuén Avila, and Laura del Castillofor field assistance during the biospeleological expe-ditions, and to the people inhabiting the area of theLos Riscos Cave for their services. We would like tothank M. en C. Griselda Montiel and Dr. FaustoMéndez for extend permissions to collect mites andamphibians, respectively. Thanks to M. en C. GriseldaMontiel, Dr. Tila M. Pérez (CNAC), and Dr. HansKlompen (OSAL) for providing facilities to checksome of the mites deposited in their respective col-lections; to Dr. Silvia Espinosa Matías (Facultad deCiencias, UNAM) for producing the scanning electronmicrographs and to Eduardo Díaz del Castillo Lomelífor the support in photo edition; and to PhD candidateNathan D. Burkett-Cadena (Auburn University) fordetailed revision of the English language. This studywas partially supported by CONACyT in the form ofa graduate scholarship (42361) and PAEP, Posgrado enCiencias Biológicas, UNAM to R. P-L.
REFERENCES
Alencaster, G., R. Torres-Hernández, M. Tristán-González, R. Barbosa-Gudiño, R. López-Doncel,J. M. Pons and L. Omaña. 1999. El Abra forma-tion in the western part of the Valles-San LuisPotosí Plataform (Mexico). Erlanger geol. Abh.Sonderband. 3: 7–8.
Alzuet, A. D. B. and R. A. Mauri. 1985 (1987).Ácaros Trombiculidae del género HannemaniaOudemans, endoparásitos de anfibios. Rev. Soc.Entomol. Argentina 44(2): 111–116.
Brennan, J. M. and M. L. Goff. 1977. Keys to thegenera of chiggers of the Western Hemisphere(Acarina: Trombiculidae). J. Parasitol. 63(3):554–566.
Bush, A., K. Lafferty, J. Lotz and A. Shostak. 1997.Parasitology meets ecology on its own terms:Margolis et al. revisited. J. Parasitol. 83: 575–583.
CETENAL. 1986. Precipitación y probabilidad de lalluvia en la República Mexicana y su evaluación.Querétaro e Hidalgo. Serie climas. Instituto deGeografía. UNAM.
CNA. 2004. Registro Mensual de Precipitación Pluvialen mm. Inédito, In Anuario Estadístico deQuerétaro de Arteaga [cited 2009 May 15]. Avail-able from http://www.queretaro.gob.mx/sedesu/deseco/esteco/perfeco/anuest/PDF/Capitulos/C1.pdf
Duellman, W. E. 2001. The Hylid Frogs of MiddleAmerica. Natural History Museum of theUniversity of Kansas, Lawrence, KS, USA.1180 pp.
Duszynski, D. and K. Jones. 1973. The occurrenceof intradermal mites, Hannemania spp. (Acarina:Trombiculidae) in anurans in New Mexico with ahistological description of the tissue capsule. Int.J. Parasitol. 3(4): 531–538.
Espino del Castillo, A., G. Castaño-Meneses, M. J.Dávila-Montes, M. Miranda-Anaya, J. B.Morales-Malacara and R. Paredes-León.2009. Seasonal distribution and Circadianactivity in the troglophile long-footed rob-ber frog, Eleutherodactylus longipes (Anura:Brachycephalidae) at Los Riscos Cave, Querétaro,Mexico: field and laboratory studies. J. CaveKarst Stud. 71(1): 24–31.
Ewing, H. E. 1925. Two new chiggers (Trombicula lar-vae). Proc. Entomol. Soc. Wash. 27(7): 145–146.
Ewing, H. E. 1926. The life history and biology of thetree-toad chigger, Trombicula hylae Ewing. Ann.Entomol. Soc. Am. 19(3): 261–267.
Frost, D. R. 2009. Amphibian Species of theWorld: an Online Reference [Internet].Version 5.3. 12 February, 2009. American
Dow
nloa
ded
by [
UN
AM
Ciu
dad
Uni
vers
itari
a] a
t 11:
48 1
1 A
ugus
t 201
1

440 Espino del Castillo et al. 2011
Museum of Natural History, New York, USA[cited 2009 Mar 27]. Available from: http://research.amnh.org/herpetology/amphibia/
García, P. C. A., J. Faivovich and C. F. B. Haddad.2007. Redescription of Hypsiboas semigutta-tus, with the description of a new speciesof the Hypsiboas pulchellus group. Copeia 4:933–951.
Goff, M. L., R. B. Loomis, W. C. Welbourn and W. J.Wrenn. 1982. A glossary of chigger terminology(Acari: Trombiculidae). J. Med. Entomol. 19(3):221–238.
Goldberg, S. R., W. J. Wrenn and C. R. Bursey.2002. Bufo mazatlanensis (Sinaloa toad), Ranatarahumarae (Tarahumara frog). Ectoparasites.Herpetol. Rev. 33(4): 301–302.
Gould, D. J. 1956. The larval trombiculid mites ofCalifornia (Acarina: Trombiculidae). Univ. Calif.Publ. Entomol. 11(1): 1–116.
Hatano, F. H., D. Gettinger, M. Van Sluys andC. F. D. Rocha. 2007. Parasitism of Hylodes phyl-lodes (Anura: Cycloramphidae) by Hannemaniasp. (Acari: Trombiculidae) in an area of Atlanticforest, Ilha Grande, South east. Brazil Parasite14: 107–112.
Hoffmann, A. 1952. Contribuciones al conocimientode los trombicúlidos mexicanos (5a parte).Ciencia 12(3–4): 87–94.
Hoffmann, A. 1969. Ácaros parásitos de batracios yreptiles en México. Rev. Latinoam. Microbiol.Parasitol. 11: 209–216.
Hoffmann, A. 1990. Los trombiculidos de México(Acarida: Trombiculidae). Publicaciones Espec-iales del Instituto de Biología 2, UNAM, México.275 pp.
Hoffmann, A. and G. López-Campos. 2000.Biodiversidad de los ácaros en México.CONABIO, UNAM, México. 230 pp.
Jung, R. E., J. S. Claeson, E. Wallace and W. C.Welbourn, Jr. 2001. Eleutherodactylus guttila-tus (spotted chirping frog), Bufo punctatus (red-spotted toad), Hyla arenicolor (canyon tree frog),and Rana Berlandieri (Rio Grande leopard frog).Mite infestation. Herpetol. Rev. 32(1): 33–34.
Krantz, G. W. and D. E. Walter. 2009. A Manual ofAcarology. 3rd ed. Texas Tech University Press,Lubbock, TX, USA. 807 pp.
Lazcano-Sahagun, C. 1986. Las Cavernas de la SierraGorda. Tomo II. Universidad Autónoma deQuerétaro, Secretaría de Desarrollo Urbano y.
Ecología y Sociedad Mexicana de ExploracionesSubterráneas, México. 207 pp.
Lemos-Espinal, J. A. and H. M. Smith. 2007.Anfibios y Reptiles del Estado de Coahuila,México. Universidad Nacional Autónoma deMéxico (UNAM) y Comisión Nacional parael Conocimiento y Uso de la Biodiversidad(CONABIO), México. 550 pp.
Loomis, R. and W. C. Welbourn, Jr. 1969. A newspecies of Hannemania (Acarina, Trombiculidae)from Bufo punctatus of Western North America,with comments on Hannemania hylae (Ewing).Bull. South. Calif. Acad. Sci. 68(3): 160–168.
Lynch, J. 1970. A taxonomic revision of theLeptodactylid frog genus EleutherodactylusCope. Univ. Kans. Mus. Nat. Hist. 20(1): 1–45.
Malone, J. H. and R. Paredes-León. 2005.Characteristics of chigger mite (Hannemaniasp.) parasitism on Eleutherodactylus marnockii(Amphibia: Leptodactylidae). Texas J. Sci. 57(4):345–358.
National Park Services. 2009. Big Bend NationalPark, Texas [Internet]. National Park Services,Washington DC, USA; [cited 2009 Oct 27].Available from http://www.nps.gov/bibe/index.htm.
Santos-Barrera, G. and L. Canseco-Márquez.2009. Eleutherodactylus longipes. In: IUCN2010. IUCN Red List of ThreatenedSpecies. Version 2010.2 [cited 2010 Sep 8].Available from: http://www.iucnredlist.org/apps/redlist/details/56727/.
Taylor, E. 1939. New Species of Mexican Anura. Univ.Kans. Sci. Bull. 26(11): 385–401.
Welbourn, Jr., W. C. and R. Loomis. 1970. Three newspecies of Hannemania (Acarina, Trombiculidae)from amphibians of Western Mexico. Bull.South. Calif. Acad. Sci. 69(2): 65–73.
Welbourn, Jr., W. C. and R. Loomis. 1975.Hannemania (Acarina: Trombiculidae) andtheir anuran hosts at Fortynine Palms Oasis,Joshua Tree National Monument, California.Bull. South. Calif. Acad. Sci. 74(1): 15–19.
Wohltmann, A., J. Köhler and P. Martin. 2006.Endoparasitic mite infection of anuran amphib-ians from Bolivian montane rain forests, withdescriptions of two new Hannemania species(Acari: Parasitengona: Trombiculidae: Leeuwenhoekiinae). Org. Divers. Evol. 6: 141–150.
Dow
nloa
ded
by [
UN
AM
Ciu
dad
Uni
vers
itari
a] a
t 11:
48 1
1 A
ugus
t 201
1
Copyright © 2022 FDOKUMEN


![Poly[di(carboxylatophenoxy)phosphazene] is a potent adjuvant for intradermal immunization](https://static.fdokumen.com/doc/165x107/6335c6c4a1ced1126c0af097/polydicarboxylatophenoxyphosphazene-is-a-potent-adjuvant-for-intradermal-immunization.jpg)

















