
Biological properties of garlic and garlic-derived organosulfur compounds
Upload
independentCategory
view
1download
0
This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Steroids 75 (2010) 272–281
Contents lists available at ScienceDirect
Steroids
journa l homepage: www.e lsev ier .com/ locate /s tero ids
Effect of dietary garlic and onion on biliary proteins and lipid peroxidation whichinfluence cholesterol nucleation in bile
Satyakumar Vidyashankar, Kari Sambaiah, Krishnapura Srinivasan ∗
Department of Biochemistry and Nutrition, Central Food Technological Research Institute, CSIR, Mysore 570 020, India
a r t i c l e i n f o
Article history:Received 2 August 2009Received in revised form27 December 2009Accepted 6 January 2010Available online 14 January 2010
Keywords:Cholesterol gallstonesCholesterol nucleation factorsBiliary proteinsDietary garlic and onionLipid peroxidationModel bile system
a b s t r a c t
Formation of cholesterol gallstones in gallbladder is controlled by procrystallizing and anticrystallizingfactors present in bile. Dietary garlic and onion have been recently observed to possess anti-lithogenicpotential in experimental mice. In this investigation, the role of biliary proteins from rats fed lithogenicdiet or garlic/onion-containing diet in the formation of cholesterol gallstones in model bile was studied.Cholesterol nucleation time of the bile from lithogenic diet group was prolonged when mixed withbile from garlic or onion groups. High molecular weight proteins of bile from garlic and onion groupsdelayed cholesterol crystal growth in model bile. Low molecular weight (LMW) proteins from the bileof lithogenic diet group promoted cholesterol crystal growth in model bile, while LMW protein fractionisolated from the bile of garlic and onion groups delayed the same. Biliary LMW protein fraction wassubjected to affinity chromatography using Con-A and the lectin-bound and unbound fractions werestudied for their influence on cholesterol nucleation time in model bile. Major portion of biliary LMWproteins in lithogenic diet group was bound to Con-A, and this protein fraction promoted cholesterolnucleation time and increased cholesterol crystal growth rate, whereas Con-A unbound fraction delayedthe onset of cholesterol crystallization. Biliary protein from garlic/onion group delayed the crystallizationand interfered with pronucleating activity of Con-A bound protein fraction. These data suggest that apartfrom the beneficial modulation of biliary cholesterol saturation index, these Allium spices also influencecholesterol nucleating and antinucleating protein factors that contribute to their anti-lithogenic potential.
© 2010 Elsevier Inc. All rights reserved.
1. Introduction
Cholesterol gallstones (CGS) occur primarily due to supersatura-tion of cholesterol in bile, leading to its precipitation (nucleation)in bile. Cholesterol precipitation is controlled by various factorssuch as mucin glycoprotein, calcium ions, bilirubin and low molec-ular weight proteins present in the bile [1,2]. Biliary proteins playa pivotal role in both promoting and inhibiting cholesterol precip-itation. Biliary cholesterol supersaturation is frequently reportedin patients with gallstones [3]. A rapid cholesterol nucleationtime and an increased cholesterol crystal growth rate are the twomost important indices that influence CGS formation and theyare affected by cholesterol supersaturation [4]. Apart from bil-iary lipid components, biliary proteins can influence cholesterolnucleation time and cholesterol crystal growth rate [5–8]. Proteinsthat enhance the nucleation of cholesterol are called pronucleatingand those which inhibit or prevent the nucleation of cholesterol,are called antinucleating factors. Both pronucleation and antinu-
∗ Corresponding author. Tel.: +91 821 2514876; fax: +91 821 2517233.E-mail address: [email protected] (K. Srinivasan).
cleation factors are found in human bile as well as in the bile ofexperimental animals such as mice, rats, gerbils and Prairie dogs.Many proteins that bind to the lectin Conconavalin-A such as �-acid glycoprotein [9,10], haptoglobin [11] and aminopeptidase-N9[12] have been shown to promote cholesterol nucleation and crys-tal growth. Protein binding to the Helix pomatia lectin [13] and arecently introduced family of cholesterol crystal associated pro-teins [14] appear to act as inhibitors of cholesterol crystallization.However, with the sole exception of apolipoproteins, no inhibitorprotein has so far been identified and fully characterized.
In contrast to the assumed direct interaction of glycoproteinsduring cholesterol crystal growth process, another biliary inhibitorprotein, apo A-I has been reported either to partially shift the dis-tribution of cholesterol in bile from vesicles towards the morestable mixed micellar form [15] or to stabilize cholesterol carri-ers such as phospholipids lamellae [16]. This increases the stabilityand appears to be based on its amphiphillic properties, allowingintegration of apo A-I in to lipid aggregates. These studies indicatethe complexity of the system in terms of the large number of fac-tors, which may affect cholesterol precipitation in bile. Apart fromthese factors, yet another factor such as lipid peroxides in bile mayplay a crucial role in the cholesterol crystallization [17]. Since an
0039-128X/$ – see front matter © 2010 Elsevier Inc. All rights reserved.doi:10.1016/j.steroids.2010.01.003

S. Vidyashankar et al. / Steroids 75 (2010) 272–281 273
increase in the fraction of peroxidized lipids in the aged populationand the increasing frequency of gallbladder stone formation in theelderly support this hypothesis.
We have recently shown that dietary Allium spices garlicand onion are effective in reducing the formation of CGS underlithogenic conditions [18] and also in the regression of the pre-formed CGS [19]. Hence, in order to understand the mechanismof cholesterol crystal nucleation and the probable effect of proteinspresent in biles of rats fed these spices, they were tested with super-saturated model biles for cholesterol nucleation. The role of lipidperoxidation on the onset of cholesterol nucleation in rat biles fedthese two test spices was also studied in normal and CGS-inducedconditions.
2. Experimental
2.1. Materials
Acrylamide, bovine serum albumin, bile acids (kit), bilesalts (kit), commassie brilliant blue, dipalmitoyl phosphatidylcholine, EDTA, HEPES, 3�-hydroxysteroid dehydrogenase, heparin,hydrazine hydrate, hydrogen peroxide, low molecular weight pro-tein standard kit for electrophoresis, manganese chloride, NADPH,N,N-methylene-bis-acrylamide, sodium dodecyl sulfate, sodiummetaperiodate, triglyceride purifier, 1,1,3,3-tetraethoxypropane,TEMED, thiobarbituric acid, tripalmitin, Tris–HCl, ethyl urethane,xanthine, xanthine oxidase and Sephadex G-100 were purchasedfrom M/s Sigma Chemical Co. (St. Louis, USA). Polyethylene tub-ing (PE-10) was purchased from M/s Thomas Scientific Co. (NewJersey, USA). Con-A sepharose was obtained from M/s BangaloreGenei (Bangalore, India). dl-Alanine, choline chloride, dini-trophenyl hydrazine, l-aspartate, �-cellulose, mercaptoethanol,l-methionine and sodium pyruvate were purchased from HiMe-dia Laboratories (Mumbai, India). Casein was purchased from M/sNimesh Corporation (Mumbai, India). All other chemicals usedwere of finest quality available and solvents were distilled beforeuse. Dehydrated onion (Allium cepa) and garlic (Allium sativum)powders used in the experiment were a generous gift from IndoNissin Foods Ltd. (Bangalore, India).
2.2. Animal diets
The animals were fed with AIN-76 semi-purified diet. The basalcontrol diet consisted of: sucrose, 60%; casein, 20%; cellulose, 5%;AIN-76 mineral mix, 3.5%; AIN-76 vitamin mix, 1%; dl-methionine,0.3%; choline chloride, 0.2% and refined peanut oil, 10%. Lithogenicdiet was prepared by supplementing 0.5% cholesterol and 0.25% bilesalts (1:1 mixture of sodium cholate and sodium deoxycholate) tothe AIN-76 basal diet. Various test diets were prepared by addinggarlic powder (0.6%) or onion powder (2%) to the AIN-76 basal diet.The diets were made isocaloric by varying the sucrose content. Heatprocessing of garlic and onion was done by adding the respectivefreeze-dried powder to the boiling water and boiled for 15 min withconstant stirring as practiced normally in the Indian culinary andused as heat-processed spices in these experiments. The diets weremade isocaloric by varying sucrose concentration. All these dietswere prepared by mixing the ingredients in a mechanical mixerand pellets were prepared using hand-operated pelletizer. Dietswere stored at 4 ◦C in air-tight containers.
2.3. Animals
Animal experiment was carried out taking appropriate mea-sures to minimize pain or discomfort in accordance with theguidelines laid down regarding the care and use of animals for
experimental procedures and with due clearance from the Insti-tutional Animal Ethics Committee. Male albino rats [OUT-Wistar,IND-cft (2c)] weighing about 140 g obtained from ExperimentalAnimal Production Facility of this institute were used. The animalswere placed in individual cages in an approved animal house facil-ity with 12 h light and dark cycles with temperature 25 ± 2 ◦C andfed fresh diets daily. The animals had free access to food and waterthroughout the study. The food intake and growth of the animalswere monitored at regular intervals. The rats were grouped into 6groups with 8 animals/group and fed with different experimentaldiets for 8 weeks. At the end of the experimental period, bile wascollected after cannulating the bile duct as described below.
2.4. Bile duct cannulation
Rats were anaesthetized with ethyl urethane (1.2 g/kg bodyweight) by intra-peritoneal injection. Laparatomy was performedand common bile duct was cannulated with intramedic PE-10polyethylene tubing (Thomas Scientific Co., USA). Bile was collectedfor 3 h in sterile tubes during which the body temperature of theanimals was maintained at 37 ◦C using incandescent lamps. Thevolume of the collected bile was measured and stored at −20 ◦Cuntil further analysis.
2.5. Biliary lipids and cholesterol saturation index (CSI)
Biliary lipids were extracted by the method of Bligh and Dyer[20]. The upper methanolic layer was used for bile acid and uronicacid analysis and lower chloroform layer was used for choles-terol and phospholipid estimation. Total bile acids in methanoliclayer were estimated by using 3∝-hydroxysteroid dehydrogenase[21]. Cholesterol was estimated by the method of Searcy andBergquist [22]. Phospholipids were quantitated by ferrous ammo-nium thiocyanate method using dipalmitoyl phosphatidyl cholineas standard [23]. Cholesterol saturation index (CSI) in the bile wascalculated by using the cholesterol, phospholipids and bile acids asinput data using Windows 98, Microsoft Excel statistical program.The coefficient for the fifth degree polynomial equation was usedin the program taken from Carey’s table [24].
2.6. Total protein and glycoprotein
Total proteins were measured according to Lowry et al. [25] withBSA as a reference standard. Glycoprotein was estimated accordingto Mantle and Allen [26] using mucin as a reference standard.
2.7. Biliary lipid peroxidation induced by hydrogen peroxide
Hepatic bile was incubated in two sets with hydrogen peroxide(0.1 mM), ferrous sulfate (0.2 mM) and ascorbic acid (0.25 mM) for30 min for the induction of lipid peroxidation. One set was usedfor total lipid extraction and used for measuring lipid peroxidation.The other set was used to study the nucleation of cholesterol atdifferent time intervals by incubating at 37 ◦C.
2.8. Evaluation of cholesterol nucleation time in bile samples
Rat hepatic bile from lithogenic diet group and various otherdietary groups was mixed in various ratios of 90:10, 75:25 and50:50, and incubated with 0.04% sodium azide in sterile tubeswith air-tight caps for 21 days at 37 ◦C. An aliquot of the bile waswithdrawn at different time intervals during this incubation andobserved under polarized microscope for precipitation and appear-ance of cholesterol monohydrate crystals. The earliest day on whichany crystal appeared was considered as the cholesterol nucleationtime of that particular bile sample. At the end of the incubation

274 S. Vidyashankar et al. / Steroids 75 (2010) 272–281
period, all the bile samples were extracted and lipid compositionwas determined. CSI was also calculated from the lipid compositionfor the individual as well as mixed bile samples.
2.9. Preparation of model bile
Model biles of predetermined CSI were prepared according tothe procedure of Kibe et al. [27]. Sodium taurocholate in methanolwas mixed with phosphatidylcholine and cholesterol in chloro-form. The mixture was dried at 45 ◦C under a mild stream ofnitrogen and subsequently for 24 h under reduced pressure. Thelipid film was reconstituted in 10 mM Tris, 150 mM sodium chlo-ride, and 3 mM sodium azide, and incubated for 16 h at 56 ◦C andsubsequently stored at 37 ◦C in dark under nitrogen.
2.10. Preparation of cholesterol monohydrate crystals (seedcrystals)
Cholesterol monohydrate crystals were prepared according toIgimi and Carey [28]. Five grams of cholesterol was dissolved in400 mL hot ethanol (95%, v/v) and the solution was slowly cooledto room temperature and kept at 4 ◦C for 3 days. Large choles-terol crystals were harvested by filtration (0.45 �m), washed with100 mL water and resuspended in 50 mL water containing 3 mMsodium azide. The suspension was sonicated for 60 s and cen-trifuged at 1000 × g for 5 min. The supernatant was collected andthe pellet was resuspended, re-sonicated and centrifuged. Theprocedure was repeated until about 200 mL of supernatant wasobtained. For the preparation of seed crystals, the pooled cloudysupernatants were stored at 4 ◦C for 4 days. During this period, asmall amount of sediment formed. The supernatant was decantedand filtered through 0.8 �m filter. The filtrate was passed througha 0.22 �m filter and the concentrate was then resuspended inwater containing 3 mM sodium azide thus producing a seed crystalsuspension with a crystal size ranging between 0.22 and 0.8 �m.Cholesterol concentration in the final seed crystal suspension wasdetermined [29].
2.11. Cholesterol crystal growth assay
Cholesterol crystal growth assay was carried out separately inseeded and unseeded sets to study the effect of different biliaryprotein fractions from rats of different diet groups on nucleationof cholesterol crystals and other parameters. The crystal growthof seeded set was initiated by the addition of 10 �g crystallinecholesterol per mL of model bile. Aliquots of solutions of pro-teins of interest were placed in screw-capped vials and filteredmodel bile was added to these vials. Both model bile and thevials with proteins were pre-equilibrated at 37 ◦C before the twowere mixed. An aliquot of model bile mixture was withdrawnat different time intervals and diluted with Tris buffered saline.The absorbance at 900 nm was measured [29]. The crystal growthparameters were calculated from the crystal growth curves [29];(1) maximum growth rate [crystal growth index (Ig)] = maximalslope of experimental curve/maximal slope of control, (2) final crys-tal concentration [crystal index (Ic)] = final crystal concentration ofexperimental/final crystal concentration of control and (3) onsettime of crystal detection [time index (It)] = onset time of experi-mental/onset time of control.
2.12. Separation of vesicles and micelles from model bile andcholesterol nucleation of vesicles and micelles
Supersaturated model bile (CSI = 1.5) was incubated with pro-tein of interest at 37 ◦C for 48 h and chromatographed on Sephadex
G-100 column (75 cm × 1 cm). Protein and lipids in the peak frac-tions were determined using standard procedures [30]. The modelbiles were subjected to gel filtration chromatography to separatevesicles and micelles. The vesicles and micelles were incubatedwith Concanavalin-A bound biliary protein (200 �g) separatedfrom the rat bile of different diet groups to study the effect of dietaryAllium spices on the nucleation of vesicles and micelles.
2.13. Separation of biliary proteins by gel filtration and affinitychromatography
Concentrated rat bile was chromatographed on Sephadex G-100 column (75 cm × 1 cm). The elution buffer contained 10 mMTris, 150 mM NaCl [29]. For the separation of low molecularweight (LMW) glycoprotein by lectin affinity chromatography, thelyophilized LMW protein fraction obtained by gel filtration wassolubilized in the eluting buffer and chromatographed on affinitycolumn [29].
2.14. Electrophoresis
Mini slab gel electrophoresis was performed according toLammelli’s procedure to determine the purity and approximatemolecular weight of the isolated biliary proteins [31]. Runninggels were made of 10% acrylamide in 0.375 M Tris–HCl, 0.1% SDSat pH 8.8. Stacking gels were made of 4% acrylamide in 0.125 MTris–HCl, 0.1% SDS at pH 7.8. Protein samples were mixed withan equal volume of sample buffer (0.1 M Tris–HCl, pH 6.8, 2%SDS, 8% 2-mercaptoethanol, and 0.005% bromophenol blue) andthen boiled for 10 min before loading on the gel. Low molecularweight protein markers were co-electrophoresed. Electrophore-sis of the proteins was performed at a constant current of 40 mAfor 5 h. Staining was done with Coommassie blue and destainedin acetic acid:methanol:water (10:25:65, v/v) overnight. Approxi-mate molecular weight of Concanavalin-A bound protein fractionwas determined.
2.15. Statistical analysis
Statistical analysis was carried out using Windows 98 Microsoftexcel statistical software and Prism Graphpad statistical software.Results were analyzed and the significance level was calculatedusing Tukey–Kramer multiple comparison test and results are con-sidered significant at P < 0.05.
3. Results
3.1. Cholesterol nucleation time in hepatic bile from different dietgroups and their mixtures
Cholesterol nucleation time of hepatic bile of rats from variousdiet groups is presented in Table 1. Bile of lithogenic diet grouphaving a CSI of 2.16 nucleated on the 5th day. On the other hand,nucleation of cholesterol in the bile of rats fed with garlic (raw),garlic (heat-processed), onion (raw) and onion (heat-processed)occurred on 18th, 16th, 19th and 20th day, respectively. Their CSIwas also significantly lower compared to that of LG diet group.Cholesterol nucleation time of mixtures of bile from different dietgroups is presented in Table 2. The mixture containing the bile oflithogenic group and the bile of garlic or onion group in the ratioof 90:10 nucleated between 7th and 8th days of incubation. Nucle-ation time for other bile mixtures containing the bile of lithogenicgroup and the bile of Allium spice group in the ratio of 75:25 and50:50 was increased as the content of the bile from spice groupincreased in the mixture (Table 2). When the bile of lithogenic dietgroup was mixed with the bile of other diet groups in the ratio of
S. Vidyashankar et al. / Steroids 75 (2010) 272–281 275
Table 1Lipid composition, Cholesterol Saturation Index, cholesterol nucleation time in the bile from rats fed various diets.
Dietary group Lipids (mM) Total lipid (g/dL) CSI Cholesterol nucleationtime (days)
Bile acid Phospholipid Cholesterol
Basal control (C) 182.6 ± 12.1 20.3 ± 1.23 4.40 ± 0.33 10.7 ± 0.51 0.53 ± 0.09 NDLithogenic (LG) 140.1 ± 7.86a 25.6 ± 2.05a 32.4 ± 2.34a 10.1 ± 0.43 2.16 ± 0.23a 5C + garlic (raw) 173.3 ± 6.00b 20.4 ± 1.04b 15.9 ± 2.01b 10.7 ± 0.28 1.06 ± 0.07b 18C + garlic (heat processed) 152.5 ± 15.7b 17.7 ± 1.01b 20.5 ± 2.88b 9.65 ± 0.75 1.30 ± 0.08b 16C + onion (raw) 151.0 ± 12.4b 21.4 ± 1.84b 16.8 ± 1.76b 9.72 ± 0.55 1.15 ± 0.15b 19C + onion (heat processed) 175.8 ± 11.0b 19.3 ± 1.00b 12.5 ± 1.80b 10.6 ± 0.58 0.95 ± 0.13b 20
Each value is the mean ± SD of 6 samples. ND: crystals not detected even on 21 days.a Statistically significant when compared to basal control group at P < 0.01.b Statistically significant when compared to LG group at P < 0.01.
50:50, the cholesterol nucleation time of the same was significantlyprolonged to 15th (raw garlic), 14th (heat-processed garlic), 15th(raw onion), and 16th day (heat-processed onion) (Table 2).
3.2. Effect of dietary garlic and onion on lipid peroxidation in bileand cholesterol nucleation time
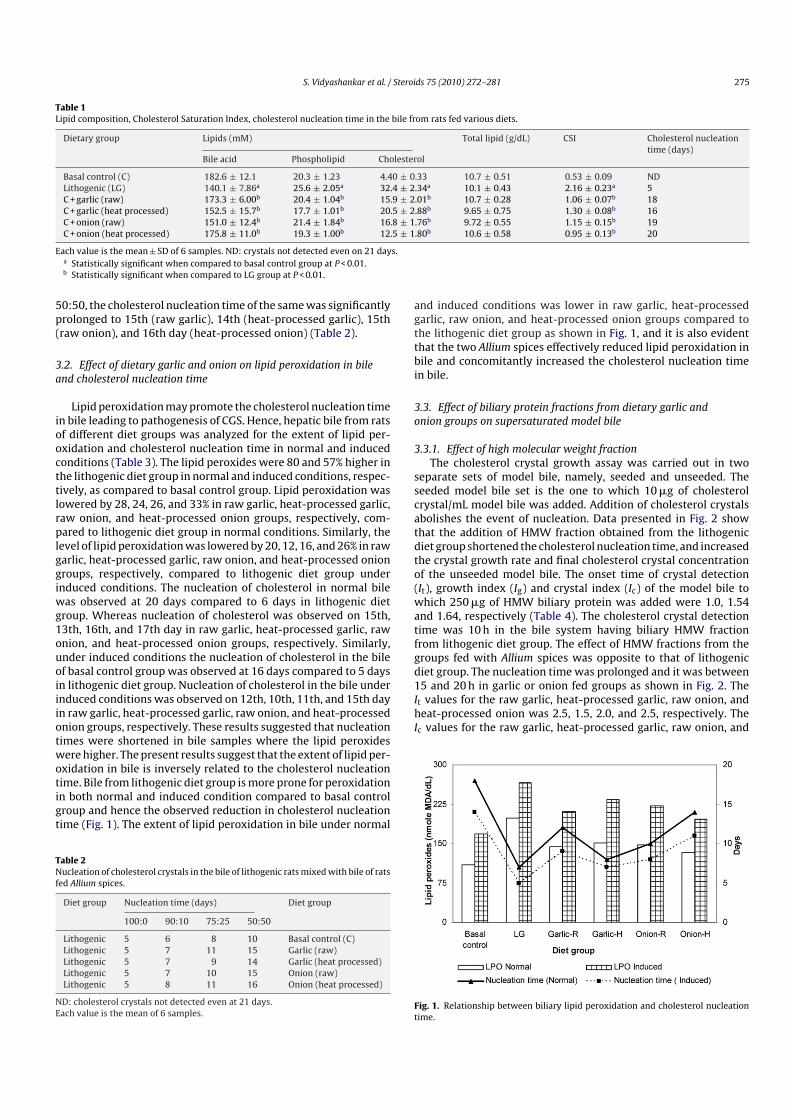
Lipid peroxidation may promote the cholesterol nucleation timein bile leading to pathogenesis of CGS. Hence, hepatic bile from ratsof different diet groups was analyzed for the extent of lipid per-oxidation and cholesterol nucleation time in normal and inducedconditions (Table 3). The lipid peroxides were 80 and 57% higher inthe lithogenic diet group in normal and induced conditions, respec-tively, as compared to basal control group. Lipid peroxidation waslowered by 28, 24, 26, and 33% in raw garlic, heat-processed garlic,raw onion, and heat-processed onion groups, respectively, com-pared to lithogenic diet group in normal conditions. Similarly, thelevel of lipid peroxidation was lowered by 20, 12, 16, and 26% in rawgarlic, heat-processed garlic, raw onion, and heat-processed oniongroups, respectively, compared to lithogenic diet group underinduced conditions. The nucleation of cholesterol in normal bilewas observed at 20 days compared to 6 days in lithogenic dietgroup. Whereas nucleation of cholesterol was observed on 15th,13th, 16th, and 17th day in raw garlic, heat-processed garlic, rawonion, and heat-processed onion groups, respectively. Similarly,under induced conditions the nucleation of cholesterol in the bileof basal control group was observed at 16 days compared to 5 daysin lithogenic diet group. Nucleation of cholesterol in the bile underinduced conditions was observed on 12th, 10th, 11th, and 15th dayin raw garlic, heat-processed garlic, raw onion, and heat-processedonion groups, respectively. These results suggested that nucleationtimes were shortened in bile samples where the lipid peroxideswere higher. The present results suggest that the extent of lipid per-oxidation in bile is inversely related to the cholesterol nucleationtime. Bile from lithogenic diet group is more prone for peroxidationin both normal and induced condition compared to basal controlgroup and hence the observed reduction in cholesterol nucleationtime (Fig. 1). The extent of lipid peroxidation in bile under normal
Table 2Nucleation of cholesterol crystals in the bile of lithogenic rats mixed with bile of ratsfed Allium spices.
Diet group Nucleation time (days) Diet group
100:0 90:10 75:25 50:50
Lithogenic 5 6 8 10 Basal control (C)Lithogenic 5 7 11 15 Garlic (raw)Lithogenic 5 7 9 14 Garlic (heat processed)Lithogenic 5 7 10 15 Onion (raw)Lithogenic 5 8 11 16 Onion (heat processed)
ND: cholesterol crystals not detected even at 21 days.Each value is the mean of 6 samples.
and induced conditions was lower in raw garlic, heat-processedgarlic, raw onion, and heat-processed onion groups compared tothe lithogenic diet group as shown in Fig. 1, and it is also evidentthat the two Allium spices effectively reduced lipid peroxidation inbile and concomitantly increased the cholesterol nucleation timein bile.
3.3. Effect of biliary protein fractions from dietary garlic andonion groups on supersaturated model bile
3.3.1. Effect of high molecular weight fractionThe cholesterol crystal growth assay was carried out in two
separate sets of model bile, namely, seeded and unseeded. Theseeded model bile set is the one to which 10 �g of cholesterolcrystal/mL model bile was added. Addition of cholesterol crystalsabolishes the event of nucleation. Data presented in Fig. 2 showthat the addition of HMW fraction obtained from the lithogenicdiet group shortened the cholesterol nucleation time, and increasedthe crystal growth rate and final cholesterol crystal concentrationof the unseeded model bile. The onset time of crystal detection(It), growth index (Ig) and crystal index (Ic) of the model bile towhich 250 �g of HMW biliary protein was added were 1.0, 1.54and 1.64, respectively (Table 4). The cholesterol crystal detectiontime was 10 h in the bile system having biliary HMW fractionfrom lithogenic diet group. The effect of HMW fractions from thegroups fed with Allium spices was opposite to that of lithogenicdiet group. The nucleation time was prolonged and it was between15 and 20 h in garlic or onion fed groups as shown in Fig. 2. TheIt values for the raw garlic, heat-processed garlic, raw onion, andheat-processed onion was 2.5, 1.5, 2.0, and 2.5, respectively. TheIc values for the raw garlic, heat-processed garlic, raw onion, and
Fig. 1. Relationship between biliary lipid peroxidation and cholesterol nucleationtime.
276 S. Vidyashankar et al. / Steroids 75 (2010) 272–281
Table 3Effect of dietary Allium spices on lipid peroxidation induced by hydrogen peroxide in rat bile and on cholesterol nucleation time.
Diet group Lipid peroxides as MDA (nmol/dL) Cholesterol nucleation time (days)
Uninduced Induced with hydrogen peroxide Uninduced Induced withhydrogen peroxide
Basal control 110.4 ± 3.89 168.9 ± 3.45 20 16Lithogenic 198.9 ± 5.77a 265.3 ± 7.89a 6 5Garlic (raw) 143.3 ± 8.34b 210.9 ± 8.78b 15 12Garlic (heat processed) 151.7 ± 9.44b 234.4 ± 6.44b 13 10Onion (raw) 147.3 ± 6.21b 221.7 ± 11.4b 16 11Onion (heat processed) 132.5 ± 3.29b 196.8 ± 7.47b 17 15
Values are mean ± SD of 6 samples/group.a Statistically significant when compared to basal control group at P < 0.01.b Statistically significant when compared to LG group at P < 0.01.
heat-processed onion were 1.33, 1.57, 1.47, and 1.15, respectively.The Ig values for the raw garlic, heat-processed garlic, raw onion,and heat-processed onion were 1.31, 1.5, 1.42, and 1.18, respec-tively (Table 4). In the seeded set of the model bile, addition of HMWprotein from lithogenic diet group increased the crystal growthrate and final crystal concentration. But dietary groups of garlic oronion brought down the crystal growth rate and final crystal con-centration (Fig. 2). Thus, feeding of Allium spices produced marked
Fig. 2. Effect of high molecular weight biliary proteins on crystal growth in (A)unseeded and (B) seeded model bile sets.
Table 4Influence of HMW protein fraction from bile of rats fed Allium spices on cholesterolcrystal growth parameters tested in model bile.
Diet group Protein (�g) Seeded Unseeded
Ig Ic Ig Ic It
Basal control 250 1.05 1.04 1.08 1.08 2.5Lithogenic 250 1.23 1.35 1.54 1.64 1.0Garlic (raw) 250 1.15 1.20 1.31 1.33 2.5Garlic (heat processed) 250 1.19 1.29 1.50 1.57 1.5Onion (raw) 250 1.17 1.25 1.42 1.47 2.0Onion (heat processed) 250 1.09 1.09 1.18 1.15 2.5
Each value is the mean of 4 samples.Ig—growth index; Ic—crystal index; It—onset time of crystal detection.Data calculated from Fig. 2.
reduction in It, Ig and Ic values in the model supersaturated bilesystem (Table 4).
3.3.2. Effect of low molecular weight fractionData presented in Fig. 3 show that the addition of LMW fraction
obtained from the lithogenic diet group shortened the cholesterolnucleation time, increased the crystal growth rate and final choles-terol crystal concentration of the unseeded model bile. The Ig, Ic,and It of the model bile to which 250 �g of LMW biliary proteinwas added were 1.36, 1.58 and 1.0, respectively (Table 5). Thecholesterol crystal detection time was 2 h in the model bile sys-tem having biliary LMW fraction from lithogenic group. The effectof LMW fractions from the groups fed with Allium spices was oppo-site to that from lithogenic diet group. Cholesterol nucleation timewas prolonged in garlic and onion fed groups as shown in Fig. 3.The It values for the raw garlic, heat-processed garlic, raw onion,and heat-processed onion were 3.0, 2.0, 2.5, and 3.41, respectively,compared to 1.0 in lithogenic diet group. The Ic values for theraw garlic, heat-processed garlic, raw onion, and heat-processedonion were 1.36, 1.50, 1.44, and 1.22, respectively, compared to1.58 in lithogenic diet group. The Ig values for the raw garlic, heat-
Table 5Influence of LMW protein fraction from bile of rats fed different diets on crystalgrowth parameters tested in model bile.
Diet group Protein (�g) Seeded Unseeded
Ig Ic Ig Ic It
Basal control 250 1.16 1.15 1.13 1.14 2.0Lithogenic 250 1.19 1.66 1.36 1.58 1.0Garlic (raw) 250 1.17 1.44 1.27 1.36 3.0Garlic (heat processed) 250 1.16 1.62 1.33 1.50 2.0Onion (raw) 250 1.15 1.51 1.31 1.44 2.5Onion (heat processed) 250 1.21 1.33 1.20 1.22 3.41
Each value is the mean of 4 samples.Ig—growth index; Ic—crystal index; It—onset time of crystal detection.Data calculated from Fig. 3.
S. Vidyashankar et al. / Steroids 75 (2010) 272–281 277
Fig. 3. Effect of low molecular weight biliary proteins on crystal growth in (A)unseeded and (B) seeded model bile sets.
processed garlic, raw onion, and heat-processed onion were 1.27,1.33, 1.31, and 1.20, respectively, compared to 1.36 in lithogenicdiet group (Table 5). In the seeded set of model bile, additionof biliary LMW protein from lithogenic diet group increased thecholesterol crystal growth rate and final cholesterol crystal concen-tration. But dietary garlic and onion brought down the cholesterolcrystal growth rate and final crystal concentration (Fig. 3). Thus,feeding of Allium spices produced marked reduction in It, Ig and Icvalues in the model supersaturated bile system (Table 5).
3.4. Effect of biliary glycoprotein fractions separated by affinitylectin chromatography on cholesterol nucleation and crystalgrowth rates in model bile system
The biliary LMW protein fraction obtained from different dietarygroups on gel filtration was further purified on Con-A lectin affinitycolumn to separate sugar-specific glycoprotein (Fig. 4). The con-centration of the protein bound to Con-A column was higher inlithogenic group compared to those from Allium spice groups. Addi-tion of Con-A bound biliary protein fraction from lithogenic dietgroup to unseeded supersaturated model bile shortened the choles-
Fig. 4. (A and B) Affinity lectin column (Con-A) chromatography of low molecularweight protein fractions of bile obtained from different dietary groups.
terol nucleation time, increased the cholesterol crystal growth rateand final cholesterol crystal concentration (Fig. 5). When 100 �g ofCon-A bound biliary protein fraction from garlic and onion groupswas added to the model bile system, the cholesterol nucleationtime was 10–15 h, and the cholesterol crystal growth rate andfinal cholesterol crystal concentration were lesser than that of thelithogenic group. On the other hand, addition of Con-A bound pro-tein fraction from lithogenic group to the seeded model bile systemhad a higher cholesterol crystal growth rate and final crystal con-centration (Fig. 5). But the addition of Con-A bound fraction fromgarlic and onion groups reduced the cholesterol crystal growthrate and final crystal concentration. The data indicated that bil-iary proteins from lithogenic group accelerated the nucleation ofcholesterol crystal and crystal growth, whereas the proteins iso-lated from spice groups inhibited the cholesterol nucleation andcrystal growth rates.
3.5. Effect of Conconavalin-A bound biliary glycoprotein fractionon cholesterol nucleation time of vesicles and micelles
Cholesterol is highly insoluble in aqueous medium of the bileand it is transported in the form of cholesterol–phospholipidstructure called vesicles and cholesterol–phospholipid–bile saltstructure called micelles. Cholesterol crystal nucleation occursfrom vesicles. In this context, the effect of Con-A bound glycopro-tein fraction isolated from the bile of rats fed spices on cholesterolcrystal nucleation was studied in vesicles and micelles from model
278 S. Vidyashankar et al. / Steroids 75 (2010) 272–281
Fig. 5. Effect of Con-A bound biliary protein fraction on crystal growth in (A)unseeded and (B) seeded model bile set.
bile separated by gel filtration. Vesicles and micelles from the modelbile were incubated in sterile vials and observed for cholesterolcrystal nucleation under polarized microscope. Initially, no crys-tals were seen in either vesicles or micelles. Vesicles and micellesseparated from the control model bile nucleated on 12th and 18thday, respectively (Table 6). When 200 �g Con-A bound biliary pro-tein fraction isolated from lithogenic group was added to vesicles ormicelles, cholesterol nucleation occurred on 5th day and 10th day,respectively. Similarly, when protein fraction from spice groups
Table 6Effect of Con-A bound protein fraction from the bile of rats fed various diet on thenucleation time on vesicles and micelles separated from model bile.
Diet group Protein (�g) Nucleation time (days)
Vesicles Micelles
Model bile – 12 18Basal control 200 16 NDLithogenic 200 5 10Garlic (raw) 200 9 19Garlic (heat processed) 200 7 15Onion (raw) 200 8 16Onion (heat processed) 200 12 20
Each value is the mean of 4 samples.
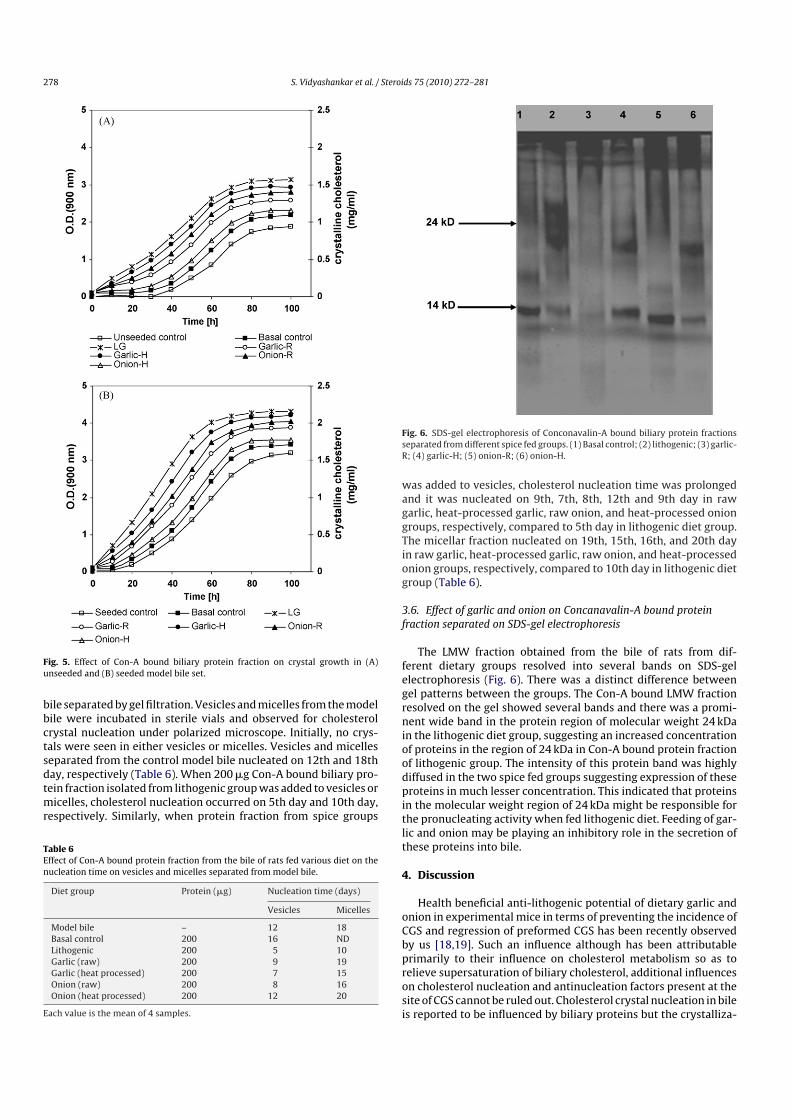
Fig. 6. SDS-gel electrophoresis of Conconavalin-A bound biliary protein fractionsseparated from different spice fed groups. (1) Basal control; (2) lithogenic; (3) garlic-R; (4) garlic-H; (5) onion-R; (6) onion-H.
was added to vesicles, cholesterol nucleation time was prolongedand it was nucleated on 9th, 7th, 8th, 12th and 9th day in rawgarlic, heat-processed garlic, raw onion, and heat-processed oniongroups, respectively, compared to 5th day in lithogenic diet group.The micellar fraction nucleated on 19th, 15th, 16th, and 20th dayin raw garlic, heat-processed garlic, raw onion, and heat-processedonion groups, respectively, compared to 10th day in lithogenic dietgroup (Table 6).
3.6. Effect of garlic and onion on Concanavalin-A bound proteinfraction separated on SDS-gel electrophoresis
The LMW fraction obtained from the bile of rats from dif-ferent dietary groups resolved into several bands on SDS-gelelectrophoresis (Fig. 6). There was a distinct difference betweengel patterns between the groups. The Con-A bound LMW fractionresolved on the gel showed several bands and there was a promi-nent wide band in the protein region of molecular weight 24 kDain the lithogenic diet group, suggesting an increased concentrationof proteins in the region of 24 kDa in Con-A bound protein fractionof lithogenic group. The intensity of this protein band was highlydiffused in the two spice fed groups suggesting expression of theseproteins in much lesser concentration. This indicated that proteinsin the molecular weight region of 24 kDa might be responsible forthe pronucleating activity when fed lithogenic diet. Feeding of gar-lic and onion may be playing an inhibitory role in the secretion ofthese proteins into bile.
4. Discussion
Health beneficial anti-lithogenic potential of dietary garlic andonion in experimental mice in terms of preventing the incidence ofCGS and regression of preformed CGS has been recently observedby us [18,19]. Such an influence although has been attributableprimarily to their influence on cholesterol metabolism so as torelieve supersaturation of biliary cholesterol, additional influenceson cholesterol nucleation and antinucleation factors present at thesite of CGS cannot be ruled out. Cholesterol crystal nucleation in bileis reported to be influenced by biliary proteins but the crystalliza-

S. Vidyashankar et al. / Steroids 75 (2010) 272–281 279
tion sequence due to proteins is not completely clear. Pronucleatingproteins such as mucin [32] and other glycoproteins [32,33] arethought to provide a nucleating matrix for cholesterol crystal for-mation. IgM and IgA can serve as pronucleating agents [6]. Theantinucleation proteins such as apolipoproteins (Apo-AI, Apo-AII)are also found in bile.
Bile samples from gallbladders of gallstone patients containcholesterol crystals [34,35], which cannot be completely removedwith the result that reliable crystal growth assays cannot be per-formed. Further, procedures used for the removal of cholesterolcrystal could also remove other critical components such as vesi-cles or proteins such as mucin and some LMW glycoproteins, whichare also important in the nucleation process [36]. Unlike native bile,model bile system is a simpler and clear system containing threecomponents, viz., cholesterol, phospholipids and bile acids. Theirassociation and dissociation during cholesterol crystal nucleationprocess can be very well studied in model bile system, and the influ-ence of any exogenous factor on the kinetics of cholesterol crystalnucleation can also be evaluated.
In the present study, hepatic biliary proteins from garlic andonion fed rats were examined for their influence on cholesterolcrystal nucleation. Rat hepatic bile which is essentially free ofany cholesterol crystals; and being very dilute and hence do notnucleate, was concentrated to simulate gallbladder bile. Duringconcentration, there was no precipitation of any biliary compo-nents. Concentration of hepatic bile by the gallbladder leads tolipid concentration. Upon concentrating the bile samples, thereis an increase in vesicular cholesterol:phospholipid ratio and anincreased propensity to nucleate cholesterol crystals [37]. Higherconcentrations of biliary components may lead to vesicular aggre-gation, which precedes cholesterol nucleation [38]. In the presentstudy, high cholesterol diet caused a near saturated condition inbile, which was made supersaturated by dehydration. Earlier stud-ies have shown a high cholesterol secretion in the bile of rats fed ahigh cholesterol diet and feeding garlic reduced the biliary choles-terol and increased bile secretion in rats [39].
Addition of bile from basal control group to that of lithogenicgroup had only little effect on the nucleation time although the CSIof the bile mixture (50:50) was >1. On the other hand, additionof bile from garlic or onion fed groups even at the lowest pro-portion markedly prolonged the nucleation time of the bile fromlithogenic diet group. These differences in the nucleation time canbe attributed to the presence of factors in the bile apart from choles-terol supersaturation. These factors may be proteins as suggestedby earlier reports [32,33]. It is suggested that lithogenic diet fed ratsmay secrete proteins in to bile which act as promoters of cholesterolcrystal nucleation, whereas rats fed Allium spices secrete biliaryproteins which act as antinucleators. An increase in the nucle-ation time in the bile of lithogenic diet group upon addition ofsmall quantity of bile from dietary garlic and onion groups sug-gests that the antinucleating factor present in these spice groupsprobably neutralizes the pronucleating factor of the lithogenic bileand hence prolongs the cholesterol nucleation time in the bile oflithogenic diet group. The bile of rats fed lithogenic diet probablycontains large quantity of pronucleating activity, but this pronucle-ating activity decreased upon feeding Allium spices. Thus, feedinggarlic and onion decreased the pronucleating activity which, sup-pressed the pronucleating activity.
Both high molecular weight and low molecular weight proteinfractions from the bile of lithogenic group shortened the cholesterolnucleation time in model bile indicating the presence of pronu-cleating factors. In contrast, both high molecular weight and lowmolecular weight protein fractions from the bile of dietary gar-lic and onion groups showed the presence of antinuleating factorswhich, exerted a dose-dependent effect in model bile system. Ear-lier, it has been reported that LMW protein fraction of hepatic bile
of rats from lithogenic diet group obtained by sepharose-4B gel fil-tration chromatography shortened the cholesterol nucleation timeand increased the crystal growth rate and final crystal concentra-tion in supersaturated model bile [40]. But with the LMW proteinfractions from the hepatic bile of rats on lithogenic diet group sup-plemented with curcumin and capsaicin, the nucleation times wereprolonged and crystal growth rates and final crystal concentrationswere decreased [40].
Biliary proteins were purified on affinity lectin column to exam-ine whether sugar specificity of glycoproteins present in the bile ofspice fed groups would have any role in cholesterol nucleation. Ahigher concentration of Con-A bound activity was isolated fromthe bile of lithogenic diet group. This glucose or mannose specificactivity was shown to be a potent promoter of cholesterol nucle-ation. The SDS-PAGE profile indicated that this activity is associatedwith 24 kDa protein. On the other hand, a 130 kDa protein has beenshown to have cholesterol nucleation promoting activity in humans[32]. Our present observation is in agreement with the earlier reportthat a higher proportion of LMW proteins from the hepatic bile oflithogenic group was bound to Con-A, and this Con-A bound frac-tion showed a pronucleating effect [40]. It is possible that the Con-Abinding protein or its sugar moiety may interact with biliary lipidsresulting in the formation of lipoprotein or glycolipid complexes[41]. Such interaction may cause destabilization of vesicles andmicelles, the two major cholesterol carriers in the bile. A higherproportion of LMW proteins from the hepatic bile of rats fed cur-cumin and capsaicin are found to, respectively, bind to wheat germagglutinin and H. pomatia lectin, and these WGA-bound or H. poma-tia lectin-bound protein fractions showed a potent antinucleatingactivity in model bile [40].
The present study is the first to report such a novel antinucleat-ing activity present in the hepatic bile of rats fed garlic and onion.This study provides evidence that the anti-lithogenicity of Alliumspices is not only controlled by reducing hepatic and biliary choles-terol but perhaps also by causing the secretion of antinucleatingproteins in the bile and by suppressing the pronucleating activity.There is no evidence that biliary proteins are affected by dietarycomponents except indirectly by the fact that some bile acids stim-ulate secretion of mucin [42] and a high cholesterol diet feedingleads to hypersecretion of mucin in Prairie dogs [43].
In all the dietary groups, both cholesterol crystal promoting andinhibiting activities can be seen except for the differences in theirconcentration. The concentration of cholesterol nucleation promot-ing activity is higher in the lithogenic diet group than its inhibitingactivity. This study has evidenced that dietary components havea profound effect on cholesterol nucleation inhibiting or promot-ing activity. One possibility is that garlic and onion feeding mayreduce the promoting activity produced in response to the dietarycholesterol and may increase the synthesis or secretion of inhibit-ing activity. The present study has clearly demonstrated that theliver or bile duct of rats secretes factors influencing the nucleationof cholesterol crystal. Feeding cholesterol in the diet increases thepronucleating factors. This release of pronucleating factors may bethe result of the pathogenic effect of high cholesterol load on theliver. Feeding garlic or onion increases the antinucleation factor,which is the result of an antagonistic effect of these Allium spices.In this context, it is evident that feeding garlic and onion preventsCGS not only by reducing cholesterol levels in bile and increasingbile acids, but also by increasing the antinucleation factors.
Lipid peroxides may also play a vital role in CGS pathogenesis.Bile is a complex mixture of phospholipids, bile acids and choles-terol. But, little is known about the redox status of bile contributingto the pathogenesis of CGS. Conceivably, biliary phospholipid mayundergo peroxidation in conditions associated with oxidant stress.Peroxidation products may be factors responsible for the choles-terol nucleation and thus leading to the formation of CGS. Despite

280 S. Vidyashankar et al. / Steroids 75 (2010) 272–281
increasing evidence of an important role of oxidant stress in varioushepatobiliary diseases such as cholestasis [44] and cholangiocarci-noma [45], no attempt has been made to assess oxidant stress inbile. Earlier, Eder et al. [46] have reported that generation of lipidperoxidation products in model bile significantly reduced choles-terol crystal formation time indicating that lipid peroxidation mayplay a role in CGS pathogenesis. In vivo study also reports increasedlevels of MDA in the bile samples of patients with cholesterol gall-stones as compared to stone free patients [17]. Our results are inagreement with the earlier reports, where the bile of lithogenic dietgroup showed higher levels of MDA in normal and peroxidation-induced conditions. But Allium spices could effectively reduce thelevels of MDA formation in normal and peroxidation-induced con-ditions leading to the delayed onset of nucleation of cholesterolin hepatic bile. These observations indicated that dietary garlicand onion have a profound influence of promoting the antinu-cleating factors and inhibition of peroxidation in the bile. Thesefactors help in the stabilization of cholesterol carriers and keepingthe same in solubilized form, which is thermodynamically feasiblefrom inhibiting cholesterol from nucleating.
Thus, proteins of different sugar specificities coexist in bile;some of them have cholesterol nucleation promoting activity,while others have cholesterol nucleation inhibiting activity. Thedominance of one over the other determines the net cholesterolnucleation behaviour. Our study has demonstrated that factor(s)influencing the nucleation of cholesterol crystal is secreted by theliver or bile duct of rats. The present report is the first one showingthe presence of antinucleating proteins in hepatic bile of rats fedgarlic or onion. It is inferred that dietary garlic or onion leads to achange in the profile of hepatobiliary proteins in such a way thatantinucleating activity was manifested unlike in the lithogenic dietgroup where pronucleating activity was manifest. Hence the sug-gestion that the anti-lithogenicity of these dietary Allium spices isdue not merely to their ability to lower cholesterol saturation index[18,19], but also to their beneficial influence on the biliary proteinprofile.
Acknowledgements
The first author (SV) is grateful to Indian Council of Medi-cal Research, New Delhi, India for the award of senior researchfellowship. This work was supported by grant-in-aid from theDepartment of Science and Technology, Government of India, NewDelhi.
References
[1] Gallinger S, Harvey PR, Petrunka CN, Taylor RD, Strasberg SM. Effectof binding of ionised calcium on the in vitro nucleation of choles-terol and calcium bilirubinate in human gall bladder bile. Gut 1986;27:1382–6.
[2] Harvey PR, Rupar CA, Gallinger S, Petrunka CN, Strasberg SM. Quantitative andqualitative comparison of gall bladder mucus glycoprotein from patients withand without gall stones. Gut 1986;27:374–81.
[3] Holzbach RT, Marsh M, Olszewski M, Holan K. Cholesterol solubility in bile.Evidence that supersaturated bile is frequent in healthy man. J Clin Invest1973;52:1467–79.
[4] Holan K, Holzbach RT, Hermann RE, Cooperman AM, Claffey WJ. Nucleationtime: a key factor in the pathogenesis of cholesterol gallstone disease. Gas-troenterology 1979;77:611–7.
[5] Groen AK, Stout JP, Draper JA, Hoek FJ, Grijm R, Tygat GN. Choles-terol nucleation-influencing activity in T-tube bile. Hepatology 1988;8:347–52.
[6] Harvey PR, Upadhya AG, Strasberg SM. Immunoglobulins as nucleating pro-teins in the gallbladder bile of patients with cholesterol gallstones. J Biol Chem1991;266:13996–4003.
[7] Harvey PR, Strasberg SM. Will the real cholesterol nucleating and antinucleat-ing proteins please stand up? Gastroenterology 1993;104:646–50.
[8] Holzbach RT, Kibe A, Thiel E, Howell JH, Marsh M, Hermann RL. Biliary proteins,unique inhibitors of cholesterol crystal nucleation in human gallbladder bile. JClin Invest 1984;73:35–45.
[9] Abei M, Schwarzendrube J, Nuutinen H, Broughan TA, Kawczak P, WilliamsC, et al. Cholesterol crystallization—promoters in human bile: comparativepotencies of immunoglobulins, �-1 acid glycoprotein, phospholipase C andaminopeptidase N1. J Lipid Res 1993;34:1141–8.
[10] Abei M, Nuutinen H, Kawczak P, Schwarzendrube J, Pillay SP, Holzbach RT. Iden-tification of human biliary �-1 acid glycoprotein as a cholesterol crystallizationpromoter. Gastroenterology 1994;106:231–8.
[11] Yamashita G, Corradini SG, Secknus R, Takabayashi A, Williams C, Hays L, et al.Biliary haptoglobin, a potent promoter of cholesterol crystallization at physio-logical concentrations. J Lipid Res 1995;36:1325–33.
[12] Offiner GD, Gong D, Afdhal NT. Identification of a 130-kilodalton human bil-iary Conconavalin A binding protein as aminopeptidase N. Gastroenterology1994;106:755–62.
[13] Ohya T, Schwarzendrube J, Busch N. Isolation of a human biliary glycopro-tein inhibitor of cholesterol crystallization. Gastroenterology 1993;104:527–38.
[14] Busch N, Lammert F, Matern S. Cholesterol crystal morphology is changed bya new subgroup of lectin-bound biliary proteins. In: Cholestatic liver diseasesnew strategies for prevention and treatment of hepatobiliary and cholestaticliver diseases. London: Kluwer Academic Publishers; 1994. p. 130.
[15] Ahmed HA, Petroni ML, Abu-Hamdiyyah M, Jazrawi RP, Northfield TC.Hydrophobic/hydrophilic balance of proteins: a major determinant of choles-terol crystal formation in model bile. J Lipid Res 1994;35:211–9.
[16] Tao S, Tazuma S, Kajiyama G. Apolipoprotein A-I stabilizes phospholipid lamel-lae and thus prolongs nucleation time in model bile systems: an ultrastructuralstudy. Biochem Biophys Acta 1993;1166:25–30.
[17] Eder MI, Jungst D, Meyer G, Paumgartner C, Von Ritter C. Lipid peroxidationproducts are increased in lithogenic human gallbladder bile. Gastroenterology1995;114:548–52.
[18] Vidyashankar S, Sambaiah K, Srinivasan K. Dietary garlic and onion reduce theincidence of atherogenic diet induced cholesterol gallstone in experimentalmice. Br J Nutr 2009;101:1621–9.
[19] Vidyashankar S, Sambaiah K, Srinivasan K. Regression of pre-establishedcholesterol gallstones by dietary garlic and onion in experimental mice.Metabolism, 59; doi:10.1016/j.metabol.2009.12.032; 2010; in press.
[20] Bligh EG, Dyer WJ. A rapid method of total lipid extraction and purification. CanJ Biochem Physiol 1959;37:911–7.
[21] Turley SD, Dietschy JM. Reevaluation of 3�-hydroxysteroid dehydrogenaseassay for total bile acids. J Lipid Res 1970;19:924–8.
[22] Searcy RL, Bergquist LM. A new colour reaction for the quantification of serumcholesterol. Clin Chim Acta 1960;5:192–9.
[23] Stewart CJM. Colorimetric estimation of phospholipids with ammonium fer-rothiocyanate. Anal Biochem 1980;104:10–4.
[24] Carey MC. Critical tables for calculating the cholesterol saturation of native bile.J Lipid Res 1978;19:945–55.
[25] Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with theFolin phenol reagent. J Biol Chem 1951;193:265–75.
[26] Mantle M, Allen A. A colorimetric assay for glycoproteins based on the periodicacid/Schiff stain. Biochem Soc Trans 1978;6:607–9.
[27] Kibe A, Dudley MA, Halpern Z, Lynn MP, Breuer AC, Holzbach RT. Factorsaffecting cholesterol monohydrate crystal nucleation time in model systemsof supersaturated bile. J Lipid Res 1985;26:1102–11.
[28] Igimi H, Carey MC. Cholesterol gallstone dissolution in bile: dissolution kineticsof crystalline (anhydrate and monohydrate) cholesterol with chenodeoxy-cholate, ursodeoxycholate, and their glycine and taurine conjugates. J LipidRes 1981;22:254–70.
[29] Busch N, Matiuck N, Sahlin S, Pillay SP, Holzbach RT. Inhibition and promotionof cholesterol crystallization by protein factors from normal human gallbladderbile. J Lipid Res 1991;32:695–702.
[30] Pattison NR, Willis KE, Frampton CM. Comparative analysis of cholesterol trans-port in bile from patients with and without cholesterol gallstones. J Lipid Res1991;32:205–14.
[31] Lammelli UK. Cleavage of structural proteins during the assembly of the headof bacteriophage T4. Nature 1970;227:680–5.
[32] Lee TJ, Smith BF. Bovine gallbladder mucin promotes cholesterol crystal nucle-ation from cholesterol-transporting vesicles in supersaturated model bile. JLipid Res 1989;30:491–8.
[33] Burnstein MJ, Ilson RG, Petrunka CN, Taylor RD, Strasberg SM. Evidence fora potent nucleating factor in the gallbladder bile of patients with cholesterolgallstones. Gastroenterology 1983;85:801–7.
[34] Sedaghat A, Grundy SM. Cholesterol crystals and the formation of cholesterolgallstones. N Engl J Med 1980;302:1274–7.
[35] Yamazki K, Powers SP, LaRusso NF. Biliary proteins: assessment of quantitativetechniques and comparison in gallstone and nongallstone subjects. J Lipid Res1988;29:1055–63.
[36] Marianne AC, DeBruijn NC, Goldhoorn BG, Guido NJ, Groen AK. The validity ofthe cholesterol nucleation assay. Biochim Biophys Acta 1992;1138:41–5.
[37] Harvey PR, Upadhya AG, Toth JL, Strasberg SM. Lectin binding characteristics ofa cholesterol nucleation promoting protein. Clin Chim Acta 1989;185:185–9.
[38] Somjen GJ, Gilat T. A non-micellar mode of cholesterol transport in human bile.FEBS Lett 1983;156:265–8.
[39] Platel K, Srinivasan K. Stimulatory influence of select spices on bile secretionin rats. Nutr Res 2000;20:1493–503.
[40] Hussain MS, Chandrasekhara N. Biliary proteins from hepatic bile of rats fedcurcumin or capsaicin inhibit cholesterol crystal nucleation in supersaturatedmodel bile. Indian J Biochem Biophys 1994;31:407–12.

S. Vidyashankar et al. / Steroids 75 (2010) 272–281 281
[41] Afdhal NH, Smith BF. Cholesterol crystal nucleation: a decade long searchfor the missing link in gallstone pathogenesis. Hepatology 1990;11:699–770.
[42] O’leary DP, Murray FE, Turner BS, LaMont JT. Bile salts stimulate glycopro-tein release by guinea pig gallbladder in vitro. Hepatology 1991;13:957–61.
[43] Lee SP, LaMont JT, Carey MC. Role of gallbladder mucus hypersecretion in theevolution of cholesterol gallstones. J Clin Invest 1981;67:1712–23.
[44] Ohshio G, Miyachi Y, Kudo H, Niwa Y, Manabe T, Tobe T. Effects of sera frompatients with obstructive jaundice on the generation of oxygen intermediatesby normal polymorphonuclear leukocytes. Liver 1988;8:366–71.
[45] Toyokuni S, Okaoto K, Yodoi J, Hiai H. Persistent oxidative stress in cancer. FEBSLett 1995;358:1–3.
[46] Eder MI, Miquel JF, Jungst D, Paumgartner G, Von Ritter C. Reactive oxygenmetabolites promote cholesterol crystal formation in model bile: role of lipidperoxidation. Free Radic Biol Med 1996;20:743–9.
Copyright © 2022 FDOKUMEN




















