
Ecological strategies in California chaparral: interacting effects of soils, climate, and fire on...
10
Plant Ecology & Diversity iFirst, 2011, 1–10 Ecological strategies in California chaparral: interacting effects of soils, climate, and fire on specific leaf area Brian Anacker a *, Nishanta Rajakaruna b , David Ackerly c , Susan Harrison d , Jon Keeley e and Michael Vasey f a University of California, Davis, Davis, USA; b College of the Atlantic, 105 Eden Street, Bar Harbor, USA; c University of California, Berkeley, Berkeley, USA; d University of California, Davis, Davis, USA; e US Geological Survey, Three Rivers, USA; f University of California, Santa Cruz, Santa Cruz, USA (Received 29 June 2011; final version received 16 October 2011) Background: High values of specific leaf area (SLA) are generally associated with high maximal growth rates in resource-rich conditions, such as mesic climates and fertile soils. However, fire may complicate this relationship since its frequency varies with both climate and soil fertility, and fire frequency selects for regeneration strategies (resprouting versus seeding) that are not independent of resource-acquisition strategies. Shared ancestry is also expected to affect the distribution of resource-use and regeneration traits. Aims: We examined climate, soil, and fire as drivers of community-level variation in a key functional trait, SLA, in chaparral in California. Methods: We quantified the phylogenetic, functional, and environmental non-independence of key traits for 87 species in 115 plots. Results: Among species, SLA was higher in resprouters than seeders, although not after phylogeny correction. Among communities, mean SLA was lower in harsh interior climates, but in these climates it was higher on more fertile soils and on more recently burned sites; in mesic coastal climates, mean SLA was uniformly high despite variation in soil fertility and fire history. Conclusions: We conclude that because important correlations exist among both species traits and environmental filters, interpreting the functional and phylogenetic structure of communities may require an understanding of complex interactive effects. Keywords: chaparral; climate; fire; sandstone; serpentine; soil; specific leaf area Introduction Environmental conditions limit the range of viable eco- logical strategies in a community, creating assemblages in which functionally similar species tend to co-occur (Diamond 1975; Weiher and Keddy 2001; Webb et al. 2002; Cavender-Bares et al. 2009). For example, water or nutri- ent scarcity may select for species with traits conferring low mean and variance in resource acquisition and growth, such as small thick leaves and high below-ground invest- ment (Grubb 1977; Grime 1979; Diaz et al. 1998; Westoby et al. 2002; Westoby and Wright 2006). For continuous resource-acquisition traits, values are typically measured across single environmental gradients, or in two contrast- ing habitats, with the expectation that the gradient will filter community composition by limiting the range of trait val- ues found at each site (Knops and Reinhart 2000; Ackerly et al. 2002; Meinzer 2003; Shipley and Almeida-Cortez 2003; Burns 2004; Hoffmann et al. 2005). In turn, such trait filtering leads to the expectation that communities will be phylogenetically clustered along gradients, since close relatives often share trait values (Webb et al. 2002; Cavender-Bares et al. 2009; Willis et al. 2010). The expectation of close matching between envi- ronmental filters, functional traits, and evolutionary *Corresponding author. Email: [email protected] relatedness may not be met, however, when a single trait reflects multiple abiotic factors, such as plant height responding to both light availability and disturbance (McGill et al. 2006); when environmental gradients are not independent of one another, such as correlations among temperature, moisture availability, and fire (Westerling et al. 2006); or when traits are physiologically and/or evolution- arily linked to one another, such as seed mass and seed number (Stearns 1992; Price 1997; Westoby et al. 2002; Vesk and Westoby 2004). Thus, successful interpretation of the environmental filtering of ecological assemblages may depend on understanding the functional, environmental, and phylogenetic relationships among key traits. One of the most widely studied resource-use traits in plants is specific leaf area (SLA), also known as leaf area/dry mass (Reich et al. 1997; Cornelissen et al. 2003 and references therein). High SLA tends to be associated with high maximal rates of photosynthesis and growth, and thus with high rates of nutrient consumption and leaf turnover. Low SLA tends to reflect the opposing strategy of slow nutrient consumption, low growth rates and slow leaf turnover (Niinemets 2001; Westoby et al. 2002; Wright et al. 2004; Wright et al. 2005; Westoby and Wright 2006; Cornwell and Ackerly 2009; but see Lusk et al. 2010). ISSN 1755-0874 print/ISSN 1755-1668 online © 2011 Botanical Society of Scotland and Taylor & Francis http://dx.doi.org/10.1080/17550874.2011.633573 http://www.tandfonline.com Downloaded by [University of California Davis] at 08:21 18 November 2011
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Ecological strategies in California chaparral: interacting effects of soils, climate, and fire on...

Plant Ecology amp DiversityiFirst 2011 1ndash10
Ecological strategies in California chaparral interacting effects of soils climate and fire onspecific leaf area
Brian Anackera Nishanta Rajakarunab David Ackerlyc Susan Harrisond Jon Keeleye and Michael Vaseyf
aUniversity of California Davis Davis USA bCollege of the Atlantic 105 Eden Street Bar Harbor USA cUniversity of CaliforniaBerkeley Berkeley USA dUniversity of California Davis Davis USA eUS Geological Survey Three Rivers USA fUniversity ofCalifornia Santa Cruz Santa Cruz USA
(Received 29 June 2011 final version received 16 October 2011)
Background High values of specific leaf area (SLA) are generally associated with high maximal growth rates in resource-richconditions such as mesic climates and fertile soils However fire may complicate this relationship since its frequency varieswith both climate and soil fertility and fire frequency selects for regeneration strategies (resprouting versus seeding) that arenot independent of resource-acquisition strategies Shared ancestry is also expected to affect the distribution of resource-useand regeneration traitsAims We examined climate soil and fire as drivers of community-level variation in a key functional trait SLA in chaparralin CaliforniaMethods We quantified the phylogenetic functional and environmental non-independence of key traits for 87 species in115 plotsResults Among species SLA was higher in resprouters than seeders although not after phylogeny correction Amongcommunities mean SLA was lower in harsh interior climates but in these climates it was higher on more fertile soils and onmore recently burned sites in mesic coastal climates mean SLA was uniformly high despite variation in soil fertility and firehistoryConclusions We conclude that because important correlations exist among both species traits and environmental filtersinterpreting the functional and phylogenetic structure of communities may require an understanding of complex interactiveeffects
Keywords chaparral climate fire sandstone serpentine soil specific leaf area
Introduction
Environmental conditions limit the range of viable eco-logical strategies in a community creating assemblagesin which functionally similar species tend to co-occur(Diamond 1975 Weiher and Keddy 2001 Webb et al 2002Cavender-Bares et al 2009) For example water or nutri-ent scarcity may select for species with traits conferringlow mean and variance in resource acquisition and growthsuch as small thick leaves and high below-ground invest-ment (Grubb 1977 Grime 1979 Diaz et al 1998 Westobyet al 2002 Westoby and Wright 2006) For continuousresource-acquisition traits values are typically measuredacross single environmental gradients or in two contrast-ing habitats with the expectation that the gradient will filtercommunity composition by limiting the range of trait val-ues found at each site (Knops and Reinhart 2000 Ackerlyet al 2002 Meinzer 2003 Shipley and Almeida-Cortez2003 Burns 2004 Hoffmann et al 2005) In turn suchtrait filtering leads to the expectation that communitieswill be phylogenetically clustered along gradients sinceclose relatives often share trait values (Webb et al 2002Cavender-Bares et al 2009 Willis et al 2010)
The expectation of close matching between envi-ronmental filters functional traits and evolutionary
Corresponding author Email blanackerucdavisedu
relatedness may not be met however when a singletrait reflects multiple abiotic factors such as plant heightresponding to both light availability and disturbance(McGill et al 2006) when environmental gradients are notindependent of one another such as correlations amongtemperature moisture availability and fire (Westerling et al2006) or when traits are physiologically andor evolution-arily linked to one another such as seed mass and seednumber (Stearns 1992 Price 1997 Westoby et al 2002Vesk and Westoby 2004) Thus successful interpretation ofthe environmental filtering of ecological assemblages maydepend on understanding the functional environmentaland phylogenetic relationships among key traits
One of the most widely studied resource-use traitsin plants is specific leaf area (SLA) also known as leafareadry mass (Reich et al 1997 Cornelissen et al 2003and references therein) High SLA tends to be associatedwith high maximal rates of photosynthesis and growthand thus with high rates of nutrient consumption and leafturnover Low SLA tends to reflect the opposing strategyof slow nutrient consumption low growth rates and slowleaf turnover (Niinemets 2001 Westoby et al 2002 Wrightet al 2004 Wright et al 2005 Westoby and Wright 2006Cornwell and Ackerly 2009 but see Lusk et al 2010)
ISSN 1755-0874 printISSN 1755-1668 onlinecopy 2011 Botanical Society of Scotland and Taylor amp Francishttpdxdoiorg101080175508742011633573httpwwwtandfonlinecom
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
2 B Anacker et al
The value of SLA as an indicator of broad resource-usestrategies is demonstrated by its negative correlations withwater use efficiency nitrogen per unit leaf mass photo-synthetic capacity leaf toughness leaf longevity leaf drymatter content and stomatal conductance (Reich et al1997 Niinemets 2001 Wright et al 2002 Reich et al2003 Wright et al 2004 Hoffmann et al 2005 Wrightet al 2005 Paula and Pausas 2006 Saura Mas et al 2009)While individuals within species may vary in their SLA(Messier et al 2010) among-species variation in SLA isgenerally considerably greater (Cornelissen et al 2003)especially in heterogeneous environments (Cornwell andAckerly 2009)
Community assembly along gradients of climaticfavourability or soil fertility is expected to be stronglyorganised by resource-use strategy as measured by SLASpecies expressing higher SLA values should be found atthe more favourable ends of these gradients ie in moremesic (abundant rainfall less extreme temperatures) cli-mates and on more fertile soils (Lamont and Markey 1995Hopper et al 1997 Wisheu et al 2000 Ackerly et al2002 Hoffmann et al 2005 Cornwell and Ackerly 2009)However these straightforward expectations may be toosimplistic if SLA is linked functionally or phylogeneticallyto other non-resource traits or if there is a lack of inde-pendence among the environmental gradients selecting forfunctional traits
In this article we examined the proposition that theresource-use strategies expressed in chaparral communi-ties as reflected by the distribution of community-levelSLA values along gradients are shaped not only by cli-mate and soil fertility but also by fire Chaparral is theCalifornian term for evergreen shrubland vegetation associ-ated with Mediterranean climates within these climates ittends to be found on well-drained and nutrient-poor soilssuch as sandstone and serpentine Recurrent crown firesare a universal characteristic of chaparral and chaparralshrubs show alternative strategies for post-fire regenera-tion (Keeley and Keeley 2000) Obligate resprouters arespecies that survive fire and re-establish by resproutingfrom underground roots rhizomes or aerial buds obli-gate seeders are those that are killed by fire and recruitfrom soil seed banks and species using both strategies arecalled lsquofacultative seedersrsquo (Table 1) These regenerationsyndromes are not independent of resource-use strategiesResprouting which is favoured by high fire frequency andseverity because it permits quick regeneration comes atthe cost of reduced growth because it requires high alloca-tion to below-ground storage Seeding which is favouredby lower fire frequency and severity requires enhancedadaptation to summer drought because of the extremelyhigh mortality at the seedling stage (Keeley and Zedler1978 Keeley 1981 Keeley et al 2006 Pausas et al 2006Verduacute and Pausas 2007) Thus although SLA is not afire-adaptive trait per se fire regimes may affect it indi-rectly by the post-fire reproductive strategies for whichthey select (Clarke 2002a Clarke 2002b Clarke et al2005)
An additional link between fire and resource-use strate-gies lies in the environmental correlations among climatesoils and fire Fire regimes are affected by climate bothdirectly (ie fire is more frequent where the dry season islonger and more severe) and indirectly through the effectsof climate on plant biomass (ie fire may be more fre-quent or severe where productive climates lead to highfuel loads) Fire regimes may also be affected by soilsfor example plant communities on extremely infertile soilssuch as serpentine may have less frequent and severe firebecause of their lower biomass accumulation and discon-tinuous canopy cover leading to a higher prevalence oflsquoseederrsquo shrub species on these soils (Safford and Harrison2004)
In the context of Californian chaparral we askedwhether it is reasonable to expect a dependable matchingof a single trait such as SLA to individual environmentalfilters or whether interaction terms representing the cor-relations among traits and among filters are required for acorrect interpretation of environmental filtering We testedthe following hypotheses (H1) SLA is only affected byclimate (H2) SLA is only affected by soil fertility (H3)SLA is only affected by fire frequency and (H4) SLA isaffected by an interaction of several or all of these potentialenvironmental filters
Materials and methods
Study system
We examined 115 plots of chaparral community composi-tion (Figure 1) The plots are from two datasets referredto as lsquoMcLaughlinrsquo and lsquoCentral Coastrsquo The McLaughlinplots were sampled on two soils (sandstone and serpentine)and two fire histories (recently burned and unburned) theCentral Coast plots were sampled on one soil (sandstone)in two climatic zones (maritime and interior)
The McLaughlin plot data were collected atMcLaughlin University of California Reserve locatedin Napa Lake and Yolo counties California USAThe McLaughlin dataset includes 80 plots fromtwo soil types serpentine (n = 40) and sandstone(n = 40) (Safford and Harrison 2004) Half of the plots oneach soil type burned in an arson fire on 16 October 1999removing an average of 85 of the original shrub coverhalf of the plots did not burn We refer to these plots aslsquorecently burnedrsquo and lsquounburnedrsquo respectively
At McLaughlin soil fertility is correlated with fire fre-quency and intensity where chaparral on relatively fertilesandstone burns significantly more often than chaparral onvery infertile serpentine (time since fire 186 plusmn 31 yearson sandstone vs 737 plusmn 39 years for serpentine) andwith higher severity (Safford and Harrison 2004) Fire isless frequent and severe on serpentine because soil infer-tility limits biomass accumulation as shown by positiveseverityndashbiomass correlations within soil types Given theirproximity the plots at McLaughlin do not differ in climate
The Central Coast data include 35 sandstone plots inthe coastal ranges of California spanning climatic zones
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
Effects of climate soil and fire on specific leaf area 3
Table 1 Expected characteristics of chaparral species with regard to post-fire regeneration strategy
Post-fire regeneration strategy
Characteristic Resprouter Seeder References
Plant levelAllocation to
storageHigh Low Pate et al 1990 Bell et al 1996 Bell 2001 Langley et al 2002
Knox and Clarke 2005 but see Chew and Bonser 2009Growth rate Low High Pate et al 1990 Bell et al 1996 Bell 2001 Bond and Midgley
2003 Schwilk and Ackerly 2005 but see Chew and Bonser2009
Height Uncertain Uncertain Midgley 1996 Bond and Midgley 2003 Falster and Westoby2005 Menges 2007
Rooting depth Deep Shallow Bell et al 1996 Bell 2001Fecundity Low High Lamont 1985 Verduacute 2000 Bell 2001 Bond and Midgley 2001
Lamont and Wiens 2003
Ecological strategyFire Tolerate Avoid Odion and Davis 2000 Clarke 2002a Clarke 2002b Clarke and
Knox 2002 Russell-Smith et al 2003 Pausas et al 2004Clarke et al 2005 Franklin et al 2005 Ojeda et al 2005Keeley et al 2006
DroughtAridity Avoid Tolerate Pausas et al 2004 Clarke et al 2005 Lloret et al 2005 Ojedaet al 2005 Pratt et al 2008 Saura Mas et al 2009 but seeLamont and Markey 1995
Soil infertility Avoid Tolerate Lamont and Markey 1995 Wisheu et al 2000 Safford andHarrison 2004
Shading Tolerate Avoid Hawkes and Menges 1996 Menges 2007 Pratt et al 2008Lifespan Long Short Bond and Midgley 2003 Lamont and Wiens 2003 Chew and
Bonser 2009Competitive
abilityUncertain Uncertain Schmalzer 2003 Clarke et al 2005 Schwilk and Ackerly 2005
Chew and Bonser 2009 Clarke and Knox 2009
from maritime (n = 17) to interior (n = 18) (Figure 1) Thisclimate transition integrates decreased presence of a sum-mer marine fog layer increased temperatures decreasedrelative humidity and decreased summer water availabilityField data from chaparral plots during the summer 2009showed large differences between maritime and interiorclimate zones (respectively) in daily average daily leaf wet-ness (502 plusmn 19 vs 38 plusmn 07) vapour pressure deficit(0323 plusmn 0027 vs 2154 plusmn 0090 kPa) average daily max-imum temperature (200 plusmn 04 vs 318 plusmn 04 C) andaverage daily soil water content (123 plusmn 01 vs 23 plusmn 02)(means plusmn one standard error Vasey et al in review)
Fire occurs less frequently in the maritime than theinterior climatic zone due to a shorter fire season associ-ated with cooler less seasonal temperatures higher rela-tive humidity lower evapotranspiration and higher rainfall(Odion and Tyler 2002) However chaparral in the maritimeclimatic zone is found on infertile shallow and sandy soilsrelative to chaparral in the interior climatic zone becausethese edaphic factors favour the persistence of chaparralvegetation (Griffin 1978 Sawyer et al 2009)
Plant community composition and soil variables
At McLaughlin woody plant community composition andcover was measured in 2002 Visual cover was estimated asa measure of species abundances for species in each of five1 m2 quadrats spaced evenly along the centre-line of eachplot Cover values were then averaged for each species and
each plot For each plot 50 g of soil were collected fromthe centre of each quadrat at 5ndash10 cm depth mixed andthen analysed by A amp L Western Agricultural LaboratoriesModesto California USA for organic matter K Mg Caand Bray P
At the Central Coast plots the plant compositionand cover of woody plants for each of the 35 plotseach 50 20 m was surveyed between 2008 and 2009(M Vasey et al unpublished) Visual cover was estimatedin 10 quadrats each 10 10 m to provide a mean abun-dance per species per plot Soil was collected from eachquadrat at 5ndash10 cm depth mixed together and analysed byBrookside Laboratories New Knoxville Ohio USA fororganic matter K Mg Ca and Bray P
Climate and fire variables
To obtain climate data for each Central Coast plot weintersected plot locations with maps of 10 macrocli-mate variables from the climate mapping system for theUnited States PRISM (httpwwwprismoregonstateedu)Variables were based on 30-year averages and had a res-olution of 1 km2 Variables were relative humidity ()potential evapotranspiration (cm) annual temperature (C)minimum winter temperature (C) maximum summer tem-perature (C) temperature seasonality (standard deviation)annual precipitation winter precipitation summer precip-itation and precipitation seasonality (coefficient of varia-tion)
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
4 B Anacker et al
sandstoneserpentine
maritimeinterior
McLaughlin
San Francisco
Santa Cruz
Santa Barbara
0 64 128km
Figure 1 Locations of 115 chaparral study plots in westernCalifornia Eighty plots were surveyed at McLaughlin aUniversity of California Natural Reserve stratified by soil type(sandstone and serpentine) and burn history (recently burned andunburned) In addition 35 plots were surveyed on sandstone soilsin the Central Coast region these plots were stratified by climaticzone (maritime and interior)
To obtain data on fire regimes we intersected plotlocations with raster grids (250 m2) of lsquofuel rankrsquo andlsquofire rotationrsquo from the California Department of Forestryand Fire Protectionrsquos Fire and Resource AssessmentProgram (CDF 2003 Fire Regime and Condition ClassEdition 03_2 httpfrapcdfcagovdatafrapgisdataselectasptheme=5) Fuel rank is based on topography vegeta-tive fuel loads and climate and varied from 1 for lsquolittle orno fire hazardrsquo to 3 for lsquovery high fire hazardrdquo The fire rota-tion data are also given in classes and are based on annualacres burned vegetation type development status eleva-tion and the bioregion values range from 1 for lsquomoderatefrequencyrsquo to 3 for lsquohigh frequencyrsquo
Trait data ndash SLA and post-fire regeneration strategy
Mean SLA (mm2 mg1) was obtained for the 87 speciesfound in the plots (see Supplementary Table 1 inAppendix 1 available via the supplementary tab on the arti-clersquos online page at httpdxdoiorg101080175508742011633573) from several sources primarily from newfield measurements For 66 of the species SLA was mea-sured from new field collections where 10 leaves werecollected from each of 10 individuals Individuals were
sampled from at least three unique populations To accountfor phenotypic plasticity of SLA with respect to soil forspecies occurring on both serpentine and non-serpentine(34 of the 66 species) five individuals were sampled fromeach soil for the others all individuals were sampledfrom one soil type Mean SLA per species was determinedusing a standard protocol (Cornelissen et al 2003) Fornine species SLA data were available from The EcologicalFlora of California (httpucjepsberkeleyeduefc) so newfield measurements were not sought For an additional12 Arctostaphylos that were very geographically rare wemeasured SLA using our own dried herbarium specimensTo validate using SLA from dried leaves versus fresh leavesfor Arctostaphylos we measured SLA from both dry leavesand from fresh leaves for 20 additional more commonArctostaphylos taxa and found that the two measures werehighly correlated (r2 = 088 n = 20 b = 101 intercept= 009 P lt 0001) To examine intraspecific variation ofSLA by soil type soil type-specific SLA means were alsocalculated for the 34 species collected on two soils In allanalyses described below mean SLA was log-transformedto meet the assumptions of parametric analyses It is pos-sible that the SLA values we determined for these specieswould differ if measured in other environments than thoseselected because genotypes and environments interact todetermine phenotypes
For each of the 87 species we determined post-fireregeneration strategy using Hickman (1993) published lit-erature and expert opinion by the authors (primarily JK)We designated each species as an obligate resprouter obli-gate seeder or facultative resprouter (can resprout or recruitfrom seed following fire) Only 20 of our species can bothresprout and post-fire seed suggesting a negative trade-off between resprouting capacity and propagule persistence(Pausas 2001 Pausas and Verdu 2005)
Analyses
Correlated environmental filters We tested if field-measured edaphic variables remotely sensed macroclimatevariables andor remotely sensed fire attributes differedamong climatic zones (interior maritime) using one-wayANOVA Relationships of soil fertility fire frequency andintensity and recent burn history at McLaughlin describedabove are reported elsewhere (Safford and Harrison 2004)and thus new analyses were not conducted here
Correlated traits At the species level (n = 87) we testedwhether SLA differed by post-fire regeneration strategyusing one-way ANOVA and also repeated this analy-sis using a correction for phylogenetic non-independence(comparative analysis by independent contrasts (CAIC)details in Appendix 2 available online as before)
The percentage of obligate seeders in each plot wascalculated percentages were weighted by abundance todown-weight the relative influence of rare species on
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
Effects of climate soil and fire on specific leaf area 5
the description of functional structure The abundance-weighted percentage for each plot was calculated as fol-lows (sum abundance obligate seeders sum abundanceof all species) 100 We calculated community-level post-fire strategy with obligate seeders as the numerator becauseboth facultative resprouters and obligate resprouters areecologically similar in the sense that they can both resproutFor McLaughlin we tested if the percentage of obligateseeders differed by soil type burn history and their interac-tion (two-way ANOVA) Prior to analysis the percentage ofobligate seeders was arc-sine transformed For the CentralCoast dataset we tested if the percentage of seeders differedamong climatic zones using one-way ANOVA
Environmental correlates of SLA At the community levelwe tested if the mean abundance-weighted SLA observedin each plot at McLaughlin differed by soil type burn his-tory and their interaction (two-way ANOVA) For each plotthe mean abundance-weighted SLA was calculated by sum-ming the product of each speciesrsquo SLA and its proportionof total cover in that plot and then dividing by the numberof species in that plot We used the proportion of a speciescover in each plot as our metric of abundance rather thanabundance alone To examine for intraspecific variation inSLA by soil type we compared the SLA of sandstonecollections with the SLA of serpentine collections for the34 species collected from both soil types using a pairedt-test For the Central Coast we tested if abundance-weighted mean SLA differed among climatic zones usingone-way ANOVA
Environmental filtering of SLA To test for filtering of SLAthe range of SLA for each community was calculated(rangeobs) and compared with a null expectation of therange of SLA at the observed level of richness (rangeexp)(sensu Cornwell and Ackerly 2009) We randomly sam-pled the species pool by data set 9999 times for eachlevel of observed species richness among the plots andcalculated the mean range of SLA values for each level ofspecies richness Communities in which a filter is operatingwill have rangeexp gt rangeobs We tested if the distribu-tion of rangeexp rangeobs was significantly greater thanzero using a one-tailed t-test We then tested if rangeexp rangeobs in each plot differed by soil type burn history andclimatic zone using a two-way ANOVA All analyses wereperformed in R version 120 (R Development Core Team2010)
Results
Correlated environmental filters
Serpentine plots had a significantly lower CaMg ratiohigher organic matter content and lower P and K levelsthan the sandstone plots The less fertile maritime plots hada significantly lower CaMg ratio and lower P level than theinterior plots (Table 2) There were higher fuel levels andalso more frequent fires in interior plots than in maritime
plots (Table 2) For the macroclimate variables interiorplots were shown to be significantly more arid than mar-itime plots (lower relative humidity higher potential evap-otranspiration higher summer temperature lower wintertemperature and higher temperature seasonality) althoughprecipitation did not differ by zone (Supplementary Table 2in Appendix 1 available online)
Correlated traits
At the species level mean SLA was significantly higher inobligate resprouters than in either facultative resproutersor obligate seeders (one-way ANOVA F2 84 = 164P lt 0001) (Figure 2) This relationship was not signif-icant after correction for phylogenetic non-independence(P = 0327) (Appendix 2 available online) demonstratingshared variation among higher taxa
The abundance-weighted percentage of seeders wassignificantly lower on sandstone soils compared to serpen-tine soils (Figure 3a) but this soil contrast disappeared inrecently burned plots (two-way ANOVA soil F1 76 = 262P lt 0001 burn history F1 76 = 219 P lt 0001 soil burn history F176 = 294 P = 009 Figure 3a) For theCentral Coast plots there was a higher abundance-weightedpercentage of obligate seeders in the low-fire maritime cli-matic zone compared with the high-fire interior climaticzone (Table 2 Figure 3c) The Central Coast compared withMcLaughlin had a lesser proportion of obligate resprouters(040 vs 054) more obligate seeders (038 vs 027)and roughly the same proportion of facultative seeders(022 vs 019)
Environmental correlates of SLA
In the McLaughlin plots abundance-weighted mean SLAwas significantly higher on sandstone soils compared withserpentine soils (Figure 3b) regardless of burn history(two-way ANOVA soil F1 76 = 203 P lt 0001 burn his-tory F1 76 = 10 P = 031 soil burn history F1 76 = 03P = 059) Plant abundances were significantly lower onserpentine soils but this difference was only present amongthe unburned plots (Figure 3c two-way ANOVA soil F1 76
= 61 P = 0015 burn history F1 76 = 6905 P lt 0001soil burn history F1 76 = 49 P = 003) In the CentralCoast plots abundance-weighted mean SLA was not sig-nificantly different between climatic zones (Figure 3e) norwas abundance (Figure 3f)
Conspecifics had 18 higher mean SLA on sandstonethan serpentine soil (paired t-test t = 231 df = 33P = 0027) At the community level in contrast meanunweighted SLA was even greater with values in sand-stone plots 32 higher than in serpentine Thus variationin SLA among communities was due to both compositionalturnover and intraspecific variation (ie phenotypic plastic-ity) but species replacement played a larger role similarto previous results for Californian woody plants (Cornwelland Ackerly 2009)
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
6 B Anacker et al
Table 2 Environmental differences by soil type (McLaughlin plots sandstone serpentine) and climatic zone (Central Coast plotsinterior maritime)
McLaughlin Central Coast
Sandstone Serpentine F P Interior Maritime F P
EdaphicCaMg ratio 34 plusmn 22 05 plusmn 05 691 lt 0001 69 plusmn 16 45 plusmn 34 72 0011Bray P (ppm) 70 plusmn 26 34 plusmn 18 530 lt 0001 376 plusmn 236 144 plusmn 335 55 0025K (ppm) 2278 plusmn 628 1285 plusmn 713 440 lt 0001 2211 plusmn 767 1726 plusmn 886 30 0094Organic
matter ()43 plusmn 09 52 plusmn 15 95 0003 50 plusmn 38 65 plusmn 18 22 0152
FireFuel rank1 na na na na 21 plusmn 05 16 plusmn 07 66 0015Fire rotation na na na na 26 plusmn 06 15 plusmn 11 153 0001
Notes Mean plusmn one standard deviation P values are reported from one-way ANOVA of each variable with soil type or climatic zone Degrees of freedomfor McLaughlin are 1 and 78 for the factor and the residuals respectively Degrees of freedom for Central Coast are 1 and 33 for the factor and theresiduals respectively1Higher fuel ranks correspond to higher potential fire hazard
10
15
20
25
30
35
40
All species (n = 87) found in plots
Post-fire regeneration strategy
log(
SLA
mm
2 m
g1)
Obligateresprouters
Facultativeresprouters
Obligateseeders
a
b
b
Figure 2 Relationship of SLA and post-fire regeneration strat-egy among the 87 species considered here Letters above box-plots indicate significant differences based on a post-hoc TukeyrsquosHSD test Open circles indicate observations considered to beoutliers
Environmental filtering of SLA
The range of SLA values for the McLaughlin plots wassignificantly lower than expected under the null modelindicating environmental filtering (54 of 80 had rangeexp rangeobs lt 0 P = 0004) (Figures 4a b) Filtering strength(rangeexp rangeobs) was significantly higher on sandstonethan serpentine (one-way ANOVA soil F1 78 = 157 P =00002 Table 2) Filtering strength was also related to burnhistory and soil type burn history (two-way ANOVAsoil F1 76 = 184 P lt 0001 burn history F1 76 = 116P = 0001 soil burn history F1 76 = 42 P = 0044) Theinteraction term reflected that filtering was significant forunburned plots on both soils and for recently burned plotson serpentine but not recently burned plots on sandstoneThe range of SLA values for the Central Coast plots was not
significantly lower than expected (17 of 35 had rangeexp rangeobs lt 0 P = 0378) indicating a lack of filtering byclimatic zone (Table 2 Figure 4c)
Discussion
Among Californian chaparral communities varying in cli-mate soil fertility and fire histories we found that SLAa critical indicator of resource-use strategy did not followthe simple patterns anticipated from considering individ-ual filters Within the McLaughlin data SLA was lower oninfertile serpentine soils as expected but fire also played arole in broadening the range of SLA present on more fer-tile sandstone soils (relaxing filtering) Within the CentralCoast data SLA did not follow any simple expectationsbeing higher neither in more equable maritime climatesnor in the interior where soils are more fertile and fires aremore frequent These results are most consistent with thehypothesis that SLA is affected by an interaction of climatesoil fertility and fire Key to understanding this interac-tion are the correlations we found at both the species andcommunity levels between SLA and post-fire regenerationstrategies and also the environmental correlations betweenclimate soils and fire frequency
Obligate post-fire seeders had significantly lower SLAthan obligate post-fire resprouters (Figure 2) This contra-dicts the notion that the reduced need for storage of theseeding strategy should lead to relatively higher growthrates and associated higher SLA in seeders (Table 1) Thecorrelation we observed may reflect a physiological trade-off in which obligate seeding imposes strong selection fordrought tolerance at the seedling stage (Ackerly 2004)Infertile soils may also select for drought-tolerant seed-ers due to imposing arid conditions (high rock contenthigh exposure to direct sun high rates of soil evapora-tion) (Ojeda et al 2005 Pratt et al 2008) While seedershave low allocation to roots and shallow rooting depth andthus a limited ability to avoid drought by reaching water inlower soil horizons they may tolerate soil aridity if they
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
Effects of climate soil and fire on specific leaf area 7
020406080
100120140
McLaughlin
Soil and Burn historyasin
(abu
ndan
cew
eigh
ted
o
blig
ate
seed
ers)
B U B USandstone Serpentine
a
aa
bA
0001020304050607
McLaughlin
Soil and burn historyA
bund
ance
wei
ghte
d lo
g(S
LA)
B U B USandstone Serpentine
a a
b b
B
0
50
100
150
McLaughlin
Soil and burn history
Abu
ndan
ce
B U B USandstone Serpentine
a
b
a
c
C
020406080
100120140
Central Coast
Climatic zoneasin
(abu
ndan
cew
eigh
ted
o
blig
ate
seed
ers)
Interior Maritime
Climatic zone
Interior Maritime
Climatic zone
Interior Maritime
a
bD
0001020304050607
Central Coast
Abu
ndan
cew
eigh
ted
log(
SLA
)
a
a
E
0
50
100
150
Central Coast
Abu
ndan
ce
aaF
Figure 3 Plant community attributes for serpentine and sandstone soils (andashc) and interior and maritime climatic zones (dndashf) Recentfire history is indicated for the McLaughlin data as follows B recently burned U unburned Letters above boxplots indicate significantdifferences based on a post-hoc Tukeyrsquos HSD test Open circles indicate observations considered to be outliers
have leaf-level strategies such as low SLA that providean advantage under low-resource conditions (Poorter et al2009 Saura Mas et al 2009) Post-fire resprouters in con-trast avoid drought by having deeper rooting systems (Bellet al 1996 Bell 2001)
The correlation between post-fire regeneration strategyand SLA was not significant in an analysis of phyloge-netically independent contrasts reflecting the influence ofspecies-rich lineages (ie Arctostaphylos and CeanothusAppendices 1 and 2 available online) that share particu-lar trait combinations (eg post-fire seeding and low SLA)through common inheritance This means that the obser-vation of low SLA and obligate seeding in a communityis in part due to common descent In other words anenvironmental filter may be directly selecting for obligateseeding and thereby indirectly selecting for the corre-lated low SLA value or vice versa Similarly a study of37 plants of the Iberian peninsula showed phylogeneticnon-independence among post-fire regeneration strategyand fruit type (Pausas and Verdu 2005) Such patterns ofphylogenetic non-independence in traits are well knownand form the basis of phylogenetic clustering in communitystructure (Webb et al 2002 Anacker 2011)
The observed patterns of post-fire regeneration strategyconfirmed that fire was more frequent on sandstone rela-tive to serpentine (obligate seeders were more common inserpentine chaparral) and that fire was more frequent in
the interior than on the coast (obligate seeders more com-mon in maritime chaparral) In the arid interior the infertileconditions of serpentine soil limit fuel accumulation cre-ating local fire refugia and favouring seeders Along thecoast cool and wet climates limit the length and severityof the fire season and the flammability of fuels leadingto low regional fire frequencies favouring seeders lowsoil fertility is likely important in determining the balancebetween woodland and chaparral (Griffin 1978 Sawyeret al 2009) rather than affecting fire regime Anotherfactor favouring obligate seeding in maritime chaparralis that longer time intervals between fires may increasethe severity of fires when they do occur lowering adultsurvivorship
Chaparral on serpentine soils at McLaughlin had sig-nificantly lower SLA than chaparral on sandstone evenimmediately following a fire (Figure 3) suggesting thatsoils play the dominant role in shaping SLA in this settingHowever our observation that low-SLA obligate seedersare much more prevalent on infrequently burning serpen-tine stands (Figure 3) suggests that fire does interact withsoil to determine community-level SLA over longer timeperiods In our Central Coast data SLA did not differbetween maritime and interior plots even though mesicmaritime climates should promote high SLA near the coastwhile both frequent fire and more fertile soils are predictedto lead to higher SLA in the interior Abundant evidence
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
8 B Anacker et al
5 10 15
00
05
10
15
20
25
30
McLaughlin sandstone
Richness
Ran
ge o
f lo
g(S
LA)
burnedunburned
expectedA
5 10 15
00
05
10
15
20
25
30
McLaughlin serpentine
Richness
Ran
ge o
f lo
g(S
LA)
burnedunburned
B
5 10 15
00
05
10
15
20
25
30
Central Coast sandstone
Richness
Ran
ge o
f lo
g(S
LA)
maritimeinterior
C
Figure 4 The range of SLA observed in chaparral plots versusnull expectation Points below the line have lower observed rangesthan expected by chance
from other studies shows that SLA tends to be higher inproductive habitats (reviewed in Cornelissen et al 2003Westoby and Wright 2006 Poorter et al 2009) such as ourmaritime plots where water availability is high Thus oneof our more striking findings is that the globally predomi-nant role of climate in shaping SLA can be nullified by themore localised effects of soil fertility and fire if these gra-dients happen to vary in the opposite direction with respectto their effects on SLA
Conclusions
We found that community structure in Californianchaparral is shaped by the environmental filtering of a keyresource-acquisition trait ndash SLA ndash by the simultaneous
effects of climate soil fertility and fire Interactiveeffects of these three filters arose from two factors thephylogenetic and functional non-independence of SLA andanother critical trait (post-fire regeneration strategy) andthe non-independence of the three environmental filtersthemselves Our results provide a clear warning that incor-rect inferences about functional and phylogenetic commu-nity structure may be reached when either traits or filtersare considered in isolation
AcknowledgementsWe thank Naveed Davoodian for assistance in compiling relevantliterature and early discussions on the role of fire and substrateon reproductive strategies in the California flora and Tom Parkerand Tanner Harris for comments on an earlier version of thismanuscript
Notes on contributorsBrian Anacker is a post doctoral researcher at University ofCalifornia Davis His research addresses the historical and eco-logical controls of diversity in Californian plant communities
Nishanta Rajakaruna is a professor of botany at College of theAtlantic Bar Harbor ME USA His research focuses on thefactors and mechanisms driving plant speciation and communityassembly on serpentine and other edaphically harsh environments
David Ackerly is an associate professor at University ofCalifornia Berkeley His research addresses the ecological andevolutionary processes that shape the functional diversity ofterrestrial plants with a particular focus on the California flora
Susan Harrison is a professor at University of California DavisThe major theme of her research is understanding the processesthat shape and maintain plant species diversity at the landscapescale where small-scale forces such as competition and facilita-tion interact with large-scale forces such as niche evolution anddispersal
Jon Keeley is a research ecologist with the US Geological Surveyand adjunct professor at University of California Los AngelesHis research focuses on the ecological impacts and history ofwildfires in Mediterranean-climate ecosystems
Michael Vasey is a graduate student at University of CaliforniaSanta Cruz His research is focused on the importance of themarine layer (coastal fog) to the ecology and diversity of chaparralvegetation and manzanitas
ReferencesAckerly DD 2004 Functional strategies of chaparral shrubs in
relation to seasonal water deficit and disturbance EcologicalMonographs 7425ndash44
Ackerly D Knight C Weiss S Barton K Starmer K 2002Leaf size specific leaf area and microhabitat distribution ofchaparral woody plants contrasting patterns in species leveland community level analyses Oecologia 130449ndash457
Anacker BL 2011 Phylogenetic patterns of serpentine plantendemism and diversity In Harrison SP Rajakaruna Neditors Serpentine as a model in evolution and ecologyBerkeley California USA University of California Pressp 49ndash70
Bell DT 2001 Ecological response syndromes in the floraof southwestern Western Australia fire resprouters versusreseeders The Botanical Review 67417ndash440
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
Effects of climate soil and fire on specific leaf area 9
Bell TL Pate JS Dixon KW 1996 Relationships between fireresponse morphology root anatomy and starch distributionin south-west Australian Epacridaceae Annals of Botany77357
Bond WJ Midgley JJ 2001 The persistence niche ecology ofsprouting in woody plants Trends in Ecology and Evolution1645ndash51
Bond WJ Midgley JJ 2003 The evolutionary ecology of sprout-ing in woody plants International Journal of Plant Sciences164103ndash114
Burns K 2004 Patterns in specific leaf area and the structureof a temperate heath community Diversity and Distributions10105ndash112
Cavender-Bares J Kozak KH Fine PVA Kembel SW 2009 Themerging of community ecology and phylogenetic biologyEcology Letters 12693ndash715
Chew SJ Bonser SP 2009 The evolution of growth rate resourceallocation and competitive ability in seeder and resprouter treeseedlings Evolutionary Ecology 23723ndash735
Clarke PJ 2002a Habitat insularity and fire response traitsevidence from a sclerophyll archipelago Oecologia132582ndash591
Clarke PJ 2002b Habitat islands in fire-prone vegetation do land-scape features influence community composition Journal ofBiogeography 29677ndash684
Clarke PJ Knox KJE 2002 Post-fire response of shrubs inthe tablelands of eastern Australia do existing modelsexplain habitat differences Australian Journal of Botany 5053ndash62
Clarke PJ Knox KJE 2009 Trade offs in resource alloca-tion that favour resprouting affect the competitive ability ofwoody seedlings in grassy communities Journal of Ecology971374ndash1382
Clarke PJ Knox KJE Wills KE Campbell M 2005 Landscapepatterns of woody plant response to crown fire distur-bance and productivity influence sprouting ability Ecology93544ndash555
Cornelissen JHC Lavorel S Garnier E Diaz S Buchmann NGurvich DE Reich PB Steege H Morgan HD HeijdenMGA 2003 A handbook of protocols for standardisedand easy measurement of plant functional traits worldwideAustralian Journal of Botany 51335ndash380
Cornwell WK Ackerly DD 2009 Community assembly andshifts in plant trait distributions across an environmen-tal gradient in coastal California Ecological Monographs79109ndash126
Diamond JM 1975 Assembly of species communities In CodyML Diamond JM editors Ecology and evolution of com-munities Cambridge MA USA Harvard University Pressp 342ndash444
Diaz S Cabido M Casanoves F 1998 Plant functional traits andenvironmental filters at a regional scale Journal of VegetationScience 9113ndash122
Falster DS Westoby M 2005 Tradeoffs between height growthrate stem persistence and maximum height among plantspecies in a post fire succession Oikos 11157ndash66
Franklin J Syphard AD He HS Mladenoff DJ 2005 Altered fireregimes affect landscape patterns of plant succession in thefoothills and mountains of southern California Ecosystems8885ndash898
Griffin JR 1978 Maritime chaparral and endemic shrubs of theMonterey Bay region California Madrono 2565ndash81
Grime JP 1979 Plant strategies and vegetation processesChichester UK John Wiley amp Sons Ltd
Grubb PJ 1977 The maintenance of species-richness in plantcommunities the importance of the regeneration nicheBiological Reviews 52107ndash145
Hawkes CV Menges ES 1996 The relationship between openspace and fire for species in a xeric Florida shrubland Bulletinof the Torrey Botanical Club 12381ndash92
Hickman JC editor 1993 The Jepson Manual Higher plantsof California Berkeley California USA University ofCalifornia Press
Hoffmann WA Franco AC Moreira MZ Haridasan M 2005Specific leaf area explains differences in leaf traits betweencongeneric savanna and forest trees Functional Ecology19932ndash940
Hopper SD Brown AP Marchant NG 1997 Plants of WesternAustralian granite outcrops Journal of the Royal Society ofWestern Australia 80141ndash158
Keeley JE 1981 Reproductive cycles and fire regimes InProceedings of the Conference Fire Regimes and EcosystemProperties vol General Technical Report WO-26 USDAForest Service p 231ndash277
Keeley JE Fotheringham CJ Baer-Keeley M 2006 Demographicpatterns of postfire regeneration in Mediterranean-climate shrublands of California Ecological Monographs76235ndash255
Keeley JE Keeley SC 2000 Chaparral North American terres-trial vegetation 2204ndash253
Keeley JE Zedler PH 1978 Reproduction of chaparral shrubsafter fire a comparison of sprouting and seeding strategiesAmerican Midland Naturalist 99142ndash161
Knops JMH Reinhart K 2000 Specific leaf area along anitrogen fertilization gradient American Midland Naturalist265ndash272
Knox KJE Clarke PJ 2005 Nutrient availability induces contrast-ing allocation and starch formation in resprouting and obligateseeding shrubs Functional Ecology 19690ndash698
Lamont BB 1985 The comparative reproductive biology ofthree Leucospermum species (Proteaceae) in relation to fireresponses and breeding system Australian Journal of Botany33139ndash145
Lamont BB Markey A 1995 Biogeography of fire-killedand resprouting Banksia species in south-western AustraliaAustralian Journal of Botany 43283ndash303
Lamont BB Wiens D 2003 Are seed set and speciation ratesalways low among species that resprout after fire and whyEvolutionary Ecology 17277ndash292
Langley J Drake B Hungate B 2002 Extensive belowground car-bon storage supports roots and mycorrhizae in regeneratingscrub oaks Oecologia 131542ndash548
Lloret F Estevan H Vayreda J Terradas J 2005 Fire regenerativesyndromes of forest woody species across fire and climaticgradients Oecologia 146461ndash468
Lusk C Onoda Y Kooyman R Gutierrez Girun A 2010Reconciling species level vs plastic responses of evergreenleaf structure to light gradients shade leaves punch abovetheir weight New Phytologist 186429ndash438
McGill BJ Enquist BJ Weiher E Westoby M 2006 Rebuildingcommunity ecology from functional traits Trends in Ecologyamp Evolution 21178ndash185
Meinzer FC 2003 Functional convergence in plant responses tothe environment Oecologia 1341ndash11
Menges ES 2007 Integrating demography and fire managementan example from Florida scrub Australian Journal of Botany55261ndash272
Messier J McGill BJ Lechowicz MJ 2010 How do traits varyacross ecological scales A case for trait based ecologyEcology Letters 13838ndash848
Midgley JJ 1996 Why the worldrsquos vegetation is not totally dom-inated by resprouting plants because resprouters are shorterthan reseeders Ecography 1992ndash95
Niinemets U 2001 Global-scale climatic controls of leaf dry massper area density and thickness in trees and shrubs Ecology82453ndash469
Odion D Tyler C 2002 Are long fire-free periods needed tomaintain the endangered fire-recruiting shrub Arctostaphylosmorroensis (Ericaceae) Conservation Ecology 64 [online]URL httpwwwconsecolorgvol6iss2art4
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
10 B Anacker et al
Odion DC Davis FW 2000 Fire soil heating and the formationof vegetation patterns in chaparral Ecological Monographs70149ndash169
Ojeda F Brun FG Vergara JJ 2005 Fire rain and the selection ofseeder and resprouter life-histories in fire-recruiting woodyplants New Phytologist155ndash165
Pate JS Froend RH Bown BJ Hansen A Kuo J 1990 Seedlinggrowth and storage characteristics of seeder and resprouterspecies of Mediterranean-type ecosystems of SW AustraliaAnnals of Botany 65585
Paula S Pausas J 2006 Leaf traits and resprouting abil-ity in the Mediterranean basin Functional Ecology 20941ndash947
Pausas J Verdu M 2005 Plant persistence traits in fire-prone ecosystems of the Mediterranean basin a phylogeneticapproach Oikos 109196ndash202
Pausas JG 2001 Resprouting vs seeding ndash a Mediterraneanperspective Oikos 94193ndash194
Pausas JG Bradstock RA Keith DA Keeley JE 2004 Plantfunctional traits in relation to fire in crown-fire ecosystemsEcology 851085ndash1100
Pausas JG Keeley JE VerdUacute M 2006 Inferring differential evo-lutionary processes of plant persistence traits in NorthernHemisphere Mediterranean fire-prone ecosystems Ecology9431ndash39
Poorter H Niinemets Poorter L Wright IJ Villar R 2009Causes and consequences of variation in leaf mass perarea (LMA) a meta analysis New Phytologist 182565ndash588
Pratt RB Jacobsen AL Mohla R Ewers FW Davis SD 2008Linkage between water stress tolerance and life history typein seedlings of nine chaparral species (Rhamnaceae) Journalof Ecology 961252ndash1265
Price T 1997 Correlated evolution and independent contrastsTransactions of the Royal Society B Biological Sciences352519ndash529
Reich PB Walters MB Ellsworth DS 1997 From tropics to tun-dra global convergence in plant functioning Proceedings ofthe National Academy of Sciences of the United States ofAmerica 9413730ndash13734
Reich PB Wright IJ Cavender-Bares J Craine JM Oleksyn JWestoby M Walters MB 2003 The evolution of plant func-tional variation traits spectra and strategies InternationalJournal of Plant Sciences 164143ndash164
Russell-Smith J Whitehead P Cook G Hoare J 2003 Responseof Eucalyptus-dominated savanna to frequent fires lessonsfrom Munmarlary 1973ndash1996 Ecological Monographs73349ndash375
Safford HD Harrison SP 2004 Fire effects on plant diver-sity in serpentine vs sandstone chaparral Ecology 85539ndash548
Saura Mas S Shipley B Lloret F 2009 Relationship betweenpost fire regeneration and leaf economics spectrum inMediterranean woody species Functional Ecology 23103ndash110
Sawyer J Keeler-Wolf T Evans J 2009 A Manual ofCalifornia Vegetation Second Edition Sacramento CAUSA California Native Plant Society
Schmalzer PA 2003 Growth and recovery of oak-saw palmettoscrub through ten years after fire Natural Areas Journal235ndash13
Schwilk DW Ackerly DD 2005 Is there a cost to resproutingSeedling growth rate and drought tolerance in sprouting andnonsprouting Ceanothus (Rhamnaceae) 1 American Journalof Botany 92404ndash410
Shipley B Almeida-Cortez J 2003 Interspecific consistency andintraspecific variability of specific leaf area with respect toirradiance and nutrient availability Ecoscience 1074ndash79
Stearns S 1992 The evolution of life histories Oxford UKOxford University Press
Verduacute M 2000 Ecological and evolutionary differences betweenMediterranean seeders and resprouters Journal of VegetationScience 11265ndash268
Verduacute M Pausas JG 2007 Fire drives phylogenetic clusteringin Mediterranean Basin woody plant communities Ecology951316ndash1323
Vesk PA Westoby M 2004 Sprouting ability across diverse dis-turbances and vegetation types worldwide Journal of Ecology92310ndash320
Webb CO Ackerly DD McPeek MA Donoghue MJ 2002Phylogenies and community ecology Annual Review ofEcology and Systematics 33475ndash505
Weiher E Keddy PA 2001 Ecological assembly rules per-spectives advances retreats Cambridge UK CambridgeUniversity Press
Westerling AL Hidalgo HG Cayan DR Swetnam TW 2006Warming and earlier spring increase western US forest wild-fire activity Science 313940
Westoby M Falster DS Moles AT Vesk PA Wright IJ 2002Plant ecological strategies Some leading dimensions ofvariation between species Annual Review of Ecology andSystematics 33125ndash159
Westoby M Wright IJ 2006 Land-plant ecology on thebasis of functional traits Trends in Ecology amp Evolution21261ndash268
Willis CG Halina M Lehman C Reich PB Keen A McCarthyS Cavender-Bares J 2010 Phylogenetic community structurein Minnesota oak savanna is influenced by spatial extent andenvironmental variation Ecography 33565ndash577
Wisheu IC Rosenzweig ML Olsvig-Whittaker L Shmida A2000 What makes nutrient-poor Mediterranean heathlandsso rich in plant diversity Evolutionary Ecology Research2935ndash955
Wright IJ Reich PB Cornelissen JHC Falster DS GroomPK Hikosaka K Lee W Lusk CH Niinemets OleksynJ 2005 Modulation of leaf economic traits and trait rela-tionships by climate Global Ecology and Biogeography14411ndash422
Wright IJ Reich PB Westoby M Ackerly DD Baruch Z BongersF Cavender-Bares J Chapin T Cornelissen JHC DiemerM 2004 The worldwide leaf economics spectrum Nature428821ndash827
Wright IJ Westoby M Reich PB 2002 Convergence towardshigher leaf mass per area in dry and nutrient poor habitats hasdifferent consequences for leaf life span Journal of Ecology90534ndash543
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011

2 B Anacker et al
The value of SLA as an indicator of broad resource-usestrategies is demonstrated by its negative correlations withwater use efficiency nitrogen per unit leaf mass photo-synthetic capacity leaf toughness leaf longevity leaf drymatter content and stomatal conductance (Reich et al1997 Niinemets 2001 Wright et al 2002 Reich et al2003 Wright et al 2004 Hoffmann et al 2005 Wrightet al 2005 Paula and Pausas 2006 Saura Mas et al 2009)While individuals within species may vary in their SLA(Messier et al 2010) among-species variation in SLA isgenerally considerably greater (Cornelissen et al 2003)especially in heterogeneous environments (Cornwell andAckerly 2009)
Community assembly along gradients of climaticfavourability or soil fertility is expected to be stronglyorganised by resource-use strategy as measured by SLASpecies expressing higher SLA values should be found atthe more favourable ends of these gradients ie in moremesic (abundant rainfall less extreme temperatures) cli-mates and on more fertile soils (Lamont and Markey 1995Hopper et al 1997 Wisheu et al 2000 Ackerly et al2002 Hoffmann et al 2005 Cornwell and Ackerly 2009)However these straightforward expectations may be toosimplistic if SLA is linked functionally or phylogeneticallyto other non-resource traits or if there is a lack of inde-pendence among the environmental gradients selecting forfunctional traits
In this article we examined the proposition that theresource-use strategies expressed in chaparral communi-ties as reflected by the distribution of community-levelSLA values along gradients are shaped not only by cli-mate and soil fertility but also by fire Chaparral is theCalifornian term for evergreen shrubland vegetation associ-ated with Mediterranean climates within these climates ittends to be found on well-drained and nutrient-poor soilssuch as sandstone and serpentine Recurrent crown firesare a universal characteristic of chaparral and chaparralshrubs show alternative strategies for post-fire regenera-tion (Keeley and Keeley 2000) Obligate resprouters arespecies that survive fire and re-establish by resproutingfrom underground roots rhizomes or aerial buds obli-gate seeders are those that are killed by fire and recruitfrom soil seed banks and species using both strategies arecalled lsquofacultative seedersrsquo (Table 1) These regenerationsyndromes are not independent of resource-use strategiesResprouting which is favoured by high fire frequency andseverity because it permits quick regeneration comes atthe cost of reduced growth because it requires high alloca-tion to below-ground storage Seeding which is favouredby lower fire frequency and severity requires enhancedadaptation to summer drought because of the extremelyhigh mortality at the seedling stage (Keeley and Zedler1978 Keeley 1981 Keeley et al 2006 Pausas et al 2006Verduacute and Pausas 2007) Thus although SLA is not afire-adaptive trait per se fire regimes may affect it indi-rectly by the post-fire reproductive strategies for whichthey select (Clarke 2002a Clarke 2002b Clarke et al2005)
An additional link between fire and resource-use strate-gies lies in the environmental correlations among climatesoils and fire Fire regimes are affected by climate bothdirectly (ie fire is more frequent where the dry season islonger and more severe) and indirectly through the effectsof climate on plant biomass (ie fire may be more fre-quent or severe where productive climates lead to highfuel loads) Fire regimes may also be affected by soilsfor example plant communities on extremely infertile soilssuch as serpentine may have less frequent and severe firebecause of their lower biomass accumulation and discon-tinuous canopy cover leading to a higher prevalence oflsquoseederrsquo shrub species on these soils (Safford and Harrison2004)
In the context of Californian chaparral we askedwhether it is reasonable to expect a dependable matchingof a single trait such as SLA to individual environmentalfilters or whether interaction terms representing the cor-relations among traits and among filters are required for acorrect interpretation of environmental filtering We testedthe following hypotheses (H1) SLA is only affected byclimate (H2) SLA is only affected by soil fertility (H3)SLA is only affected by fire frequency and (H4) SLA isaffected by an interaction of several or all of these potentialenvironmental filters
Materials and methods
Study system
We examined 115 plots of chaparral community composi-tion (Figure 1) The plots are from two datasets referredto as lsquoMcLaughlinrsquo and lsquoCentral Coastrsquo The McLaughlinplots were sampled on two soils (sandstone and serpentine)and two fire histories (recently burned and unburned) theCentral Coast plots were sampled on one soil (sandstone)in two climatic zones (maritime and interior)
The McLaughlin plot data were collected atMcLaughlin University of California Reserve locatedin Napa Lake and Yolo counties California USAThe McLaughlin dataset includes 80 plots fromtwo soil types serpentine (n = 40) and sandstone(n = 40) (Safford and Harrison 2004) Half of the plots oneach soil type burned in an arson fire on 16 October 1999removing an average of 85 of the original shrub coverhalf of the plots did not burn We refer to these plots aslsquorecently burnedrsquo and lsquounburnedrsquo respectively
At McLaughlin soil fertility is correlated with fire fre-quency and intensity where chaparral on relatively fertilesandstone burns significantly more often than chaparral onvery infertile serpentine (time since fire 186 plusmn 31 yearson sandstone vs 737 plusmn 39 years for serpentine) andwith higher severity (Safford and Harrison 2004) Fire isless frequent and severe on serpentine because soil infer-tility limits biomass accumulation as shown by positiveseverityndashbiomass correlations within soil types Given theirproximity the plots at McLaughlin do not differ in climate
The Central Coast data include 35 sandstone plots inthe coastal ranges of California spanning climatic zones
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
Effects of climate soil and fire on specific leaf area 3
Table 1 Expected characteristics of chaparral species with regard to post-fire regeneration strategy
Post-fire regeneration strategy
Characteristic Resprouter Seeder References
Plant levelAllocation to
storageHigh Low Pate et al 1990 Bell et al 1996 Bell 2001 Langley et al 2002
Knox and Clarke 2005 but see Chew and Bonser 2009Growth rate Low High Pate et al 1990 Bell et al 1996 Bell 2001 Bond and Midgley
2003 Schwilk and Ackerly 2005 but see Chew and Bonser2009
Height Uncertain Uncertain Midgley 1996 Bond and Midgley 2003 Falster and Westoby2005 Menges 2007
Rooting depth Deep Shallow Bell et al 1996 Bell 2001Fecundity Low High Lamont 1985 Verduacute 2000 Bell 2001 Bond and Midgley 2001
Lamont and Wiens 2003
Ecological strategyFire Tolerate Avoid Odion and Davis 2000 Clarke 2002a Clarke 2002b Clarke and
Knox 2002 Russell-Smith et al 2003 Pausas et al 2004Clarke et al 2005 Franklin et al 2005 Ojeda et al 2005Keeley et al 2006
DroughtAridity Avoid Tolerate Pausas et al 2004 Clarke et al 2005 Lloret et al 2005 Ojedaet al 2005 Pratt et al 2008 Saura Mas et al 2009 but seeLamont and Markey 1995
Soil infertility Avoid Tolerate Lamont and Markey 1995 Wisheu et al 2000 Safford andHarrison 2004
Shading Tolerate Avoid Hawkes and Menges 1996 Menges 2007 Pratt et al 2008Lifespan Long Short Bond and Midgley 2003 Lamont and Wiens 2003 Chew and
Bonser 2009Competitive
abilityUncertain Uncertain Schmalzer 2003 Clarke et al 2005 Schwilk and Ackerly 2005
Chew and Bonser 2009 Clarke and Knox 2009
from maritime (n = 17) to interior (n = 18) (Figure 1) Thisclimate transition integrates decreased presence of a sum-mer marine fog layer increased temperatures decreasedrelative humidity and decreased summer water availabilityField data from chaparral plots during the summer 2009showed large differences between maritime and interiorclimate zones (respectively) in daily average daily leaf wet-ness (502 plusmn 19 vs 38 plusmn 07) vapour pressure deficit(0323 plusmn 0027 vs 2154 plusmn 0090 kPa) average daily max-imum temperature (200 plusmn 04 vs 318 plusmn 04 C) andaverage daily soil water content (123 plusmn 01 vs 23 plusmn 02)(means plusmn one standard error Vasey et al in review)
Fire occurs less frequently in the maritime than theinterior climatic zone due to a shorter fire season associ-ated with cooler less seasonal temperatures higher rela-tive humidity lower evapotranspiration and higher rainfall(Odion and Tyler 2002) However chaparral in the maritimeclimatic zone is found on infertile shallow and sandy soilsrelative to chaparral in the interior climatic zone becausethese edaphic factors favour the persistence of chaparralvegetation (Griffin 1978 Sawyer et al 2009)
Plant community composition and soil variables
At McLaughlin woody plant community composition andcover was measured in 2002 Visual cover was estimated asa measure of species abundances for species in each of five1 m2 quadrats spaced evenly along the centre-line of eachplot Cover values were then averaged for each species and
each plot For each plot 50 g of soil were collected fromthe centre of each quadrat at 5ndash10 cm depth mixed andthen analysed by A amp L Western Agricultural LaboratoriesModesto California USA for organic matter K Mg Caand Bray P
At the Central Coast plots the plant compositionand cover of woody plants for each of the 35 plotseach 50 20 m was surveyed between 2008 and 2009(M Vasey et al unpublished) Visual cover was estimatedin 10 quadrats each 10 10 m to provide a mean abun-dance per species per plot Soil was collected from eachquadrat at 5ndash10 cm depth mixed together and analysed byBrookside Laboratories New Knoxville Ohio USA fororganic matter K Mg Ca and Bray P
Climate and fire variables
To obtain climate data for each Central Coast plot weintersected plot locations with maps of 10 macrocli-mate variables from the climate mapping system for theUnited States PRISM (httpwwwprismoregonstateedu)Variables were based on 30-year averages and had a res-olution of 1 km2 Variables were relative humidity ()potential evapotranspiration (cm) annual temperature (C)minimum winter temperature (C) maximum summer tem-perature (C) temperature seasonality (standard deviation)annual precipitation winter precipitation summer precip-itation and precipitation seasonality (coefficient of varia-tion)
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
4 B Anacker et al
sandstoneserpentine
maritimeinterior
McLaughlin
San Francisco
Santa Cruz
Santa Barbara
0 64 128km
Figure 1 Locations of 115 chaparral study plots in westernCalifornia Eighty plots were surveyed at McLaughlin aUniversity of California Natural Reserve stratified by soil type(sandstone and serpentine) and burn history (recently burned andunburned) In addition 35 plots were surveyed on sandstone soilsin the Central Coast region these plots were stratified by climaticzone (maritime and interior)
To obtain data on fire regimes we intersected plotlocations with raster grids (250 m2) of lsquofuel rankrsquo andlsquofire rotationrsquo from the California Department of Forestryand Fire Protectionrsquos Fire and Resource AssessmentProgram (CDF 2003 Fire Regime and Condition ClassEdition 03_2 httpfrapcdfcagovdatafrapgisdataselectasptheme=5) Fuel rank is based on topography vegeta-tive fuel loads and climate and varied from 1 for lsquolittle orno fire hazardrsquo to 3 for lsquovery high fire hazardrdquo The fire rota-tion data are also given in classes and are based on annualacres burned vegetation type development status eleva-tion and the bioregion values range from 1 for lsquomoderatefrequencyrsquo to 3 for lsquohigh frequencyrsquo
Trait data ndash SLA and post-fire regeneration strategy
Mean SLA (mm2 mg1) was obtained for the 87 speciesfound in the plots (see Supplementary Table 1 inAppendix 1 available via the supplementary tab on the arti-clersquos online page at httpdxdoiorg101080175508742011633573) from several sources primarily from newfield measurements For 66 of the species SLA was mea-sured from new field collections where 10 leaves werecollected from each of 10 individuals Individuals were
sampled from at least three unique populations To accountfor phenotypic plasticity of SLA with respect to soil forspecies occurring on both serpentine and non-serpentine(34 of the 66 species) five individuals were sampled fromeach soil for the others all individuals were sampledfrom one soil type Mean SLA per species was determinedusing a standard protocol (Cornelissen et al 2003) Fornine species SLA data were available from The EcologicalFlora of California (httpucjepsberkeleyeduefc) so newfield measurements were not sought For an additional12 Arctostaphylos that were very geographically rare wemeasured SLA using our own dried herbarium specimensTo validate using SLA from dried leaves versus fresh leavesfor Arctostaphylos we measured SLA from both dry leavesand from fresh leaves for 20 additional more commonArctostaphylos taxa and found that the two measures werehighly correlated (r2 = 088 n = 20 b = 101 intercept= 009 P lt 0001) To examine intraspecific variation ofSLA by soil type soil type-specific SLA means were alsocalculated for the 34 species collected on two soils In allanalyses described below mean SLA was log-transformedto meet the assumptions of parametric analyses It is pos-sible that the SLA values we determined for these specieswould differ if measured in other environments than thoseselected because genotypes and environments interact todetermine phenotypes
For each of the 87 species we determined post-fireregeneration strategy using Hickman (1993) published lit-erature and expert opinion by the authors (primarily JK)We designated each species as an obligate resprouter obli-gate seeder or facultative resprouter (can resprout or recruitfrom seed following fire) Only 20 of our species can bothresprout and post-fire seed suggesting a negative trade-off between resprouting capacity and propagule persistence(Pausas 2001 Pausas and Verdu 2005)
Analyses
Correlated environmental filters We tested if field-measured edaphic variables remotely sensed macroclimatevariables andor remotely sensed fire attributes differedamong climatic zones (interior maritime) using one-wayANOVA Relationships of soil fertility fire frequency andintensity and recent burn history at McLaughlin describedabove are reported elsewhere (Safford and Harrison 2004)and thus new analyses were not conducted here
Correlated traits At the species level (n = 87) we testedwhether SLA differed by post-fire regeneration strategyusing one-way ANOVA and also repeated this analy-sis using a correction for phylogenetic non-independence(comparative analysis by independent contrasts (CAIC)details in Appendix 2 available online as before)
The percentage of obligate seeders in each plot wascalculated percentages were weighted by abundance todown-weight the relative influence of rare species on
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
Effects of climate soil and fire on specific leaf area 5
the description of functional structure The abundance-weighted percentage for each plot was calculated as fol-lows (sum abundance obligate seeders sum abundanceof all species) 100 We calculated community-level post-fire strategy with obligate seeders as the numerator becauseboth facultative resprouters and obligate resprouters areecologically similar in the sense that they can both resproutFor McLaughlin we tested if the percentage of obligateseeders differed by soil type burn history and their interac-tion (two-way ANOVA) Prior to analysis the percentage ofobligate seeders was arc-sine transformed For the CentralCoast dataset we tested if the percentage of seeders differedamong climatic zones using one-way ANOVA
Environmental correlates of SLA At the community levelwe tested if the mean abundance-weighted SLA observedin each plot at McLaughlin differed by soil type burn his-tory and their interaction (two-way ANOVA) For each plotthe mean abundance-weighted SLA was calculated by sum-ming the product of each speciesrsquo SLA and its proportionof total cover in that plot and then dividing by the numberof species in that plot We used the proportion of a speciescover in each plot as our metric of abundance rather thanabundance alone To examine for intraspecific variation inSLA by soil type we compared the SLA of sandstonecollections with the SLA of serpentine collections for the34 species collected from both soil types using a pairedt-test For the Central Coast we tested if abundance-weighted mean SLA differed among climatic zones usingone-way ANOVA
Environmental filtering of SLA To test for filtering of SLAthe range of SLA for each community was calculated(rangeobs) and compared with a null expectation of therange of SLA at the observed level of richness (rangeexp)(sensu Cornwell and Ackerly 2009) We randomly sam-pled the species pool by data set 9999 times for eachlevel of observed species richness among the plots andcalculated the mean range of SLA values for each level ofspecies richness Communities in which a filter is operatingwill have rangeexp gt rangeobs We tested if the distribu-tion of rangeexp rangeobs was significantly greater thanzero using a one-tailed t-test We then tested if rangeexp rangeobs in each plot differed by soil type burn history andclimatic zone using a two-way ANOVA All analyses wereperformed in R version 120 (R Development Core Team2010)
Results
Correlated environmental filters
Serpentine plots had a significantly lower CaMg ratiohigher organic matter content and lower P and K levelsthan the sandstone plots The less fertile maritime plots hada significantly lower CaMg ratio and lower P level than theinterior plots (Table 2) There were higher fuel levels andalso more frequent fires in interior plots than in maritime
plots (Table 2) For the macroclimate variables interiorplots were shown to be significantly more arid than mar-itime plots (lower relative humidity higher potential evap-otranspiration higher summer temperature lower wintertemperature and higher temperature seasonality) althoughprecipitation did not differ by zone (Supplementary Table 2in Appendix 1 available online)
Correlated traits
At the species level mean SLA was significantly higher inobligate resprouters than in either facultative resproutersor obligate seeders (one-way ANOVA F2 84 = 164P lt 0001) (Figure 2) This relationship was not signif-icant after correction for phylogenetic non-independence(P = 0327) (Appendix 2 available online) demonstratingshared variation among higher taxa
The abundance-weighted percentage of seeders wassignificantly lower on sandstone soils compared to serpen-tine soils (Figure 3a) but this soil contrast disappeared inrecently burned plots (two-way ANOVA soil F1 76 = 262P lt 0001 burn history F1 76 = 219 P lt 0001 soil burn history F176 = 294 P = 009 Figure 3a) For theCentral Coast plots there was a higher abundance-weightedpercentage of obligate seeders in the low-fire maritime cli-matic zone compared with the high-fire interior climaticzone (Table 2 Figure 3c) The Central Coast compared withMcLaughlin had a lesser proportion of obligate resprouters(040 vs 054) more obligate seeders (038 vs 027)and roughly the same proportion of facultative seeders(022 vs 019)
Environmental correlates of SLA
In the McLaughlin plots abundance-weighted mean SLAwas significantly higher on sandstone soils compared withserpentine soils (Figure 3b) regardless of burn history(two-way ANOVA soil F1 76 = 203 P lt 0001 burn his-tory F1 76 = 10 P = 031 soil burn history F1 76 = 03P = 059) Plant abundances were significantly lower onserpentine soils but this difference was only present amongthe unburned plots (Figure 3c two-way ANOVA soil F1 76
= 61 P = 0015 burn history F1 76 = 6905 P lt 0001soil burn history F1 76 = 49 P = 003) In the CentralCoast plots abundance-weighted mean SLA was not sig-nificantly different between climatic zones (Figure 3e) norwas abundance (Figure 3f)
Conspecifics had 18 higher mean SLA on sandstonethan serpentine soil (paired t-test t = 231 df = 33P = 0027) At the community level in contrast meanunweighted SLA was even greater with values in sand-stone plots 32 higher than in serpentine Thus variationin SLA among communities was due to both compositionalturnover and intraspecific variation (ie phenotypic plastic-ity) but species replacement played a larger role similarto previous results for Californian woody plants (Cornwelland Ackerly 2009)
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
6 B Anacker et al
Table 2 Environmental differences by soil type (McLaughlin plots sandstone serpentine) and climatic zone (Central Coast plotsinterior maritime)
McLaughlin Central Coast
Sandstone Serpentine F P Interior Maritime F P
EdaphicCaMg ratio 34 plusmn 22 05 plusmn 05 691 lt 0001 69 plusmn 16 45 plusmn 34 72 0011Bray P (ppm) 70 plusmn 26 34 plusmn 18 530 lt 0001 376 plusmn 236 144 plusmn 335 55 0025K (ppm) 2278 plusmn 628 1285 plusmn 713 440 lt 0001 2211 plusmn 767 1726 plusmn 886 30 0094Organic
matter ()43 plusmn 09 52 plusmn 15 95 0003 50 plusmn 38 65 plusmn 18 22 0152
FireFuel rank1 na na na na 21 plusmn 05 16 plusmn 07 66 0015Fire rotation na na na na 26 plusmn 06 15 plusmn 11 153 0001
Notes Mean plusmn one standard deviation P values are reported from one-way ANOVA of each variable with soil type or climatic zone Degrees of freedomfor McLaughlin are 1 and 78 for the factor and the residuals respectively Degrees of freedom for Central Coast are 1 and 33 for the factor and theresiduals respectively1Higher fuel ranks correspond to higher potential fire hazard
10
15
20
25
30
35
40
All species (n = 87) found in plots
Post-fire regeneration strategy
log(
SLA
mm
2 m
g1)
Obligateresprouters
Facultativeresprouters
Obligateseeders
a
b
b
Figure 2 Relationship of SLA and post-fire regeneration strat-egy among the 87 species considered here Letters above box-plots indicate significant differences based on a post-hoc TukeyrsquosHSD test Open circles indicate observations considered to beoutliers
Environmental filtering of SLA
The range of SLA values for the McLaughlin plots wassignificantly lower than expected under the null modelindicating environmental filtering (54 of 80 had rangeexp rangeobs lt 0 P = 0004) (Figures 4a b) Filtering strength(rangeexp rangeobs) was significantly higher on sandstonethan serpentine (one-way ANOVA soil F1 78 = 157 P =00002 Table 2) Filtering strength was also related to burnhistory and soil type burn history (two-way ANOVAsoil F1 76 = 184 P lt 0001 burn history F1 76 = 116P = 0001 soil burn history F1 76 = 42 P = 0044) Theinteraction term reflected that filtering was significant forunburned plots on both soils and for recently burned plotson serpentine but not recently burned plots on sandstoneThe range of SLA values for the Central Coast plots was not
significantly lower than expected (17 of 35 had rangeexp rangeobs lt 0 P = 0378) indicating a lack of filtering byclimatic zone (Table 2 Figure 4c)
Discussion
Among Californian chaparral communities varying in cli-mate soil fertility and fire histories we found that SLAa critical indicator of resource-use strategy did not followthe simple patterns anticipated from considering individ-ual filters Within the McLaughlin data SLA was lower oninfertile serpentine soils as expected but fire also played arole in broadening the range of SLA present on more fer-tile sandstone soils (relaxing filtering) Within the CentralCoast data SLA did not follow any simple expectationsbeing higher neither in more equable maritime climatesnor in the interior where soils are more fertile and fires aremore frequent These results are most consistent with thehypothesis that SLA is affected by an interaction of climatesoil fertility and fire Key to understanding this interac-tion are the correlations we found at both the species andcommunity levels between SLA and post-fire regenerationstrategies and also the environmental correlations betweenclimate soils and fire frequency
Obligate post-fire seeders had significantly lower SLAthan obligate post-fire resprouters (Figure 2) This contra-dicts the notion that the reduced need for storage of theseeding strategy should lead to relatively higher growthrates and associated higher SLA in seeders (Table 1) Thecorrelation we observed may reflect a physiological trade-off in which obligate seeding imposes strong selection fordrought tolerance at the seedling stage (Ackerly 2004)Infertile soils may also select for drought-tolerant seed-ers due to imposing arid conditions (high rock contenthigh exposure to direct sun high rates of soil evapora-tion) (Ojeda et al 2005 Pratt et al 2008) While seedershave low allocation to roots and shallow rooting depth andthus a limited ability to avoid drought by reaching water inlower soil horizons they may tolerate soil aridity if they
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
Effects of climate soil and fire on specific leaf area 7
020406080
100120140
McLaughlin
Soil and Burn historyasin
(abu
ndan
cew
eigh
ted
o
blig
ate
seed
ers)
B U B USandstone Serpentine
a
aa
bA
0001020304050607
McLaughlin
Soil and burn historyA
bund
ance
wei
ghte
d lo
g(S
LA)
B U B USandstone Serpentine
a a
b b
B
0
50
100
150
McLaughlin
Soil and burn history
Abu
ndan
ce
B U B USandstone Serpentine
a
b
a
c
C
020406080
100120140
Central Coast
Climatic zoneasin
(abu
ndan
cew
eigh
ted
o
blig
ate
seed
ers)
Interior Maritime
Climatic zone
Interior Maritime
Climatic zone
Interior Maritime
a
bD
0001020304050607
Central Coast
Abu
ndan
cew
eigh
ted
log(
SLA
)
a
a
E
0
50
100
150
Central Coast
Abu
ndan
ce
aaF
Figure 3 Plant community attributes for serpentine and sandstone soils (andashc) and interior and maritime climatic zones (dndashf) Recentfire history is indicated for the McLaughlin data as follows B recently burned U unburned Letters above boxplots indicate significantdifferences based on a post-hoc Tukeyrsquos HSD test Open circles indicate observations considered to be outliers
have leaf-level strategies such as low SLA that providean advantage under low-resource conditions (Poorter et al2009 Saura Mas et al 2009) Post-fire resprouters in con-trast avoid drought by having deeper rooting systems (Bellet al 1996 Bell 2001)
The correlation between post-fire regeneration strategyand SLA was not significant in an analysis of phyloge-netically independent contrasts reflecting the influence ofspecies-rich lineages (ie Arctostaphylos and CeanothusAppendices 1 and 2 available online) that share particu-lar trait combinations (eg post-fire seeding and low SLA)through common inheritance This means that the obser-vation of low SLA and obligate seeding in a communityis in part due to common descent In other words anenvironmental filter may be directly selecting for obligateseeding and thereby indirectly selecting for the corre-lated low SLA value or vice versa Similarly a study of37 plants of the Iberian peninsula showed phylogeneticnon-independence among post-fire regeneration strategyand fruit type (Pausas and Verdu 2005) Such patterns ofphylogenetic non-independence in traits are well knownand form the basis of phylogenetic clustering in communitystructure (Webb et al 2002 Anacker 2011)
The observed patterns of post-fire regeneration strategyconfirmed that fire was more frequent on sandstone rela-tive to serpentine (obligate seeders were more common inserpentine chaparral) and that fire was more frequent in
the interior than on the coast (obligate seeders more com-mon in maritime chaparral) In the arid interior the infertileconditions of serpentine soil limit fuel accumulation cre-ating local fire refugia and favouring seeders Along thecoast cool and wet climates limit the length and severityof the fire season and the flammability of fuels leadingto low regional fire frequencies favouring seeders lowsoil fertility is likely important in determining the balancebetween woodland and chaparral (Griffin 1978 Sawyeret al 2009) rather than affecting fire regime Anotherfactor favouring obligate seeding in maritime chaparralis that longer time intervals between fires may increasethe severity of fires when they do occur lowering adultsurvivorship
Chaparral on serpentine soils at McLaughlin had sig-nificantly lower SLA than chaparral on sandstone evenimmediately following a fire (Figure 3) suggesting thatsoils play the dominant role in shaping SLA in this settingHowever our observation that low-SLA obligate seedersare much more prevalent on infrequently burning serpen-tine stands (Figure 3) suggests that fire does interact withsoil to determine community-level SLA over longer timeperiods In our Central Coast data SLA did not differbetween maritime and interior plots even though mesicmaritime climates should promote high SLA near the coastwhile both frequent fire and more fertile soils are predictedto lead to higher SLA in the interior Abundant evidence
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
8 B Anacker et al
5 10 15
00
05
10
15
20
25
30
McLaughlin sandstone
Richness
Ran
ge o
f lo
g(S
LA)
burnedunburned
expectedA
5 10 15
00
05
10
15
20
25
30
McLaughlin serpentine
Richness
Ran
ge o
f lo
g(S
LA)
burnedunburned
B
5 10 15
00
05
10
15
20
25
30
Central Coast sandstone
Richness
Ran
ge o
f lo
g(S
LA)
maritimeinterior
C
Figure 4 The range of SLA observed in chaparral plots versusnull expectation Points below the line have lower observed rangesthan expected by chance
from other studies shows that SLA tends to be higher inproductive habitats (reviewed in Cornelissen et al 2003Westoby and Wright 2006 Poorter et al 2009) such as ourmaritime plots where water availability is high Thus oneof our more striking findings is that the globally predomi-nant role of climate in shaping SLA can be nullified by themore localised effects of soil fertility and fire if these gra-dients happen to vary in the opposite direction with respectto their effects on SLA
Conclusions
We found that community structure in Californianchaparral is shaped by the environmental filtering of a keyresource-acquisition trait ndash SLA ndash by the simultaneous
effects of climate soil fertility and fire Interactiveeffects of these three filters arose from two factors thephylogenetic and functional non-independence of SLA andanother critical trait (post-fire regeneration strategy) andthe non-independence of the three environmental filtersthemselves Our results provide a clear warning that incor-rect inferences about functional and phylogenetic commu-nity structure may be reached when either traits or filtersare considered in isolation
AcknowledgementsWe thank Naveed Davoodian for assistance in compiling relevantliterature and early discussions on the role of fire and substrateon reproductive strategies in the California flora and Tom Parkerand Tanner Harris for comments on an earlier version of thismanuscript
Notes on contributorsBrian Anacker is a post doctoral researcher at University ofCalifornia Davis His research addresses the historical and eco-logical controls of diversity in Californian plant communities
Nishanta Rajakaruna is a professor of botany at College of theAtlantic Bar Harbor ME USA His research focuses on thefactors and mechanisms driving plant speciation and communityassembly on serpentine and other edaphically harsh environments
David Ackerly is an associate professor at University ofCalifornia Berkeley His research addresses the ecological andevolutionary processes that shape the functional diversity ofterrestrial plants with a particular focus on the California flora
Susan Harrison is a professor at University of California DavisThe major theme of her research is understanding the processesthat shape and maintain plant species diversity at the landscapescale where small-scale forces such as competition and facilita-tion interact with large-scale forces such as niche evolution anddispersal
Jon Keeley is a research ecologist with the US Geological Surveyand adjunct professor at University of California Los AngelesHis research focuses on the ecological impacts and history ofwildfires in Mediterranean-climate ecosystems
Michael Vasey is a graduate student at University of CaliforniaSanta Cruz His research is focused on the importance of themarine layer (coastal fog) to the ecology and diversity of chaparralvegetation and manzanitas
ReferencesAckerly DD 2004 Functional strategies of chaparral shrubs in
relation to seasonal water deficit and disturbance EcologicalMonographs 7425ndash44
Ackerly D Knight C Weiss S Barton K Starmer K 2002Leaf size specific leaf area and microhabitat distribution ofchaparral woody plants contrasting patterns in species leveland community level analyses Oecologia 130449ndash457
Anacker BL 2011 Phylogenetic patterns of serpentine plantendemism and diversity In Harrison SP Rajakaruna Neditors Serpentine as a model in evolution and ecologyBerkeley California USA University of California Pressp 49ndash70
Bell DT 2001 Ecological response syndromes in the floraof southwestern Western Australia fire resprouters versusreseeders The Botanical Review 67417ndash440
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
Effects of climate soil and fire on specific leaf area 9
Bell TL Pate JS Dixon KW 1996 Relationships between fireresponse morphology root anatomy and starch distributionin south-west Australian Epacridaceae Annals of Botany77357
Bond WJ Midgley JJ 2001 The persistence niche ecology ofsprouting in woody plants Trends in Ecology and Evolution1645ndash51
Bond WJ Midgley JJ 2003 The evolutionary ecology of sprout-ing in woody plants International Journal of Plant Sciences164103ndash114
Burns K 2004 Patterns in specific leaf area and the structureof a temperate heath community Diversity and Distributions10105ndash112
Cavender-Bares J Kozak KH Fine PVA Kembel SW 2009 Themerging of community ecology and phylogenetic biologyEcology Letters 12693ndash715
Chew SJ Bonser SP 2009 The evolution of growth rate resourceallocation and competitive ability in seeder and resprouter treeseedlings Evolutionary Ecology 23723ndash735
Clarke PJ 2002a Habitat insularity and fire response traitsevidence from a sclerophyll archipelago Oecologia132582ndash591
Clarke PJ 2002b Habitat islands in fire-prone vegetation do land-scape features influence community composition Journal ofBiogeography 29677ndash684
Clarke PJ Knox KJE 2002 Post-fire response of shrubs inthe tablelands of eastern Australia do existing modelsexplain habitat differences Australian Journal of Botany 5053ndash62
Clarke PJ Knox KJE 2009 Trade offs in resource alloca-tion that favour resprouting affect the competitive ability ofwoody seedlings in grassy communities Journal of Ecology971374ndash1382
Clarke PJ Knox KJE Wills KE Campbell M 2005 Landscapepatterns of woody plant response to crown fire distur-bance and productivity influence sprouting ability Ecology93544ndash555
Cornelissen JHC Lavorel S Garnier E Diaz S Buchmann NGurvich DE Reich PB Steege H Morgan HD HeijdenMGA 2003 A handbook of protocols for standardisedand easy measurement of plant functional traits worldwideAustralian Journal of Botany 51335ndash380
Cornwell WK Ackerly DD 2009 Community assembly andshifts in plant trait distributions across an environmen-tal gradient in coastal California Ecological Monographs79109ndash126
Diamond JM 1975 Assembly of species communities In CodyML Diamond JM editors Ecology and evolution of com-munities Cambridge MA USA Harvard University Pressp 342ndash444
Diaz S Cabido M Casanoves F 1998 Plant functional traits andenvironmental filters at a regional scale Journal of VegetationScience 9113ndash122
Falster DS Westoby M 2005 Tradeoffs between height growthrate stem persistence and maximum height among plantspecies in a post fire succession Oikos 11157ndash66
Franklin J Syphard AD He HS Mladenoff DJ 2005 Altered fireregimes affect landscape patterns of plant succession in thefoothills and mountains of southern California Ecosystems8885ndash898
Griffin JR 1978 Maritime chaparral and endemic shrubs of theMonterey Bay region California Madrono 2565ndash81
Grime JP 1979 Plant strategies and vegetation processesChichester UK John Wiley amp Sons Ltd
Grubb PJ 1977 The maintenance of species-richness in plantcommunities the importance of the regeneration nicheBiological Reviews 52107ndash145
Hawkes CV Menges ES 1996 The relationship between openspace and fire for species in a xeric Florida shrubland Bulletinof the Torrey Botanical Club 12381ndash92
Hickman JC editor 1993 The Jepson Manual Higher plantsof California Berkeley California USA University ofCalifornia Press
Hoffmann WA Franco AC Moreira MZ Haridasan M 2005Specific leaf area explains differences in leaf traits betweencongeneric savanna and forest trees Functional Ecology19932ndash940
Hopper SD Brown AP Marchant NG 1997 Plants of WesternAustralian granite outcrops Journal of the Royal Society ofWestern Australia 80141ndash158
Keeley JE 1981 Reproductive cycles and fire regimes InProceedings of the Conference Fire Regimes and EcosystemProperties vol General Technical Report WO-26 USDAForest Service p 231ndash277
Keeley JE Fotheringham CJ Baer-Keeley M 2006 Demographicpatterns of postfire regeneration in Mediterranean-climate shrublands of California Ecological Monographs76235ndash255
Keeley JE Keeley SC 2000 Chaparral North American terres-trial vegetation 2204ndash253
Keeley JE Zedler PH 1978 Reproduction of chaparral shrubsafter fire a comparison of sprouting and seeding strategiesAmerican Midland Naturalist 99142ndash161
Knops JMH Reinhart K 2000 Specific leaf area along anitrogen fertilization gradient American Midland Naturalist265ndash272
Knox KJE Clarke PJ 2005 Nutrient availability induces contrast-ing allocation and starch formation in resprouting and obligateseeding shrubs Functional Ecology 19690ndash698
Lamont BB 1985 The comparative reproductive biology ofthree Leucospermum species (Proteaceae) in relation to fireresponses and breeding system Australian Journal of Botany33139ndash145
Lamont BB Markey A 1995 Biogeography of fire-killedand resprouting Banksia species in south-western AustraliaAustralian Journal of Botany 43283ndash303
Lamont BB Wiens D 2003 Are seed set and speciation ratesalways low among species that resprout after fire and whyEvolutionary Ecology 17277ndash292
Langley J Drake B Hungate B 2002 Extensive belowground car-bon storage supports roots and mycorrhizae in regeneratingscrub oaks Oecologia 131542ndash548
Lloret F Estevan H Vayreda J Terradas J 2005 Fire regenerativesyndromes of forest woody species across fire and climaticgradients Oecologia 146461ndash468
Lusk C Onoda Y Kooyman R Gutierrez Girun A 2010Reconciling species level vs plastic responses of evergreenleaf structure to light gradients shade leaves punch abovetheir weight New Phytologist 186429ndash438
McGill BJ Enquist BJ Weiher E Westoby M 2006 Rebuildingcommunity ecology from functional traits Trends in Ecologyamp Evolution 21178ndash185
Meinzer FC 2003 Functional convergence in plant responses tothe environment Oecologia 1341ndash11
Menges ES 2007 Integrating demography and fire managementan example from Florida scrub Australian Journal of Botany55261ndash272
Messier J McGill BJ Lechowicz MJ 2010 How do traits varyacross ecological scales A case for trait based ecologyEcology Letters 13838ndash848
Midgley JJ 1996 Why the worldrsquos vegetation is not totally dom-inated by resprouting plants because resprouters are shorterthan reseeders Ecography 1992ndash95
Niinemets U 2001 Global-scale climatic controls of leaf dry massper area density and thickness in trees and shrubs Ecology82453ndash469
Odion D Tyler C 2002 Are long fire-free periods needed tomaintain the endangered fire-recruiting shrub Arctostaphylosmorroensis (Ericaceae) Conservation Ecology 64 [online]URL httpwwwconsecolorgvol6iss2art4
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
10 B Anacker et al
Odion DC Davis FW 2000 Fire soil heating and the formationof vegetation patterns in chaparral Ecological Monographs70149ndash169
Ojeda F Brun FG Vergara JJ 2005 Fire rain and the selection ofseeder and resprouter life-histories in fire-recruiting woodyplants New Phytologist155ndash165
Pate JS Froend RH Bown BJ Hansen A Kuo J 1990 Seedlinggrowth and storage characteristics of seeder and resprouterspecies of Mediterranean-type ecosystems of SW AustraliaAnnals of Botany 65585
Paula S Pausas J 2006 Leaf traits and resprouting abil-ity in the Mediterranean basin Functional Ecology 20941ndash947
Pausas J Verdu M 2005 Plant persistence traits in fire-prone ecosystems of the Mediterranean basin a phylogeneticapproach Oikos 109196ndash202
Pausas JG 2001 Resprouting vs seeding ndash a Mediterraneanperspective Oikos 94193ndash194
Pausas JG Bradstock RA Keith DA Keeley JE 2004 Plantfunctional traits in relation to fire in crown-fire ecosystemsEcology 851085ndash1100
Pausas JG Keeley JE VerdUacute M 2006 Inferring differential evo-lutionary processes of plant persistence traits in NorthernHemisphere Mediterranean fire-prone ecosystems Ecology9431ndash39
Poorter H Niinemets Poorter L Wright IJ Villar R 2009Causes and consequences of variation in leaf mass perarea (LMA) a meta analysis New Phytologist 182565ndash588
Pratt RB Jacobsen AL Mohla R Ewers FW Davis SD 2008Linkage between water stress tolerance and life history typein seedlings of nine chaparral species (Rhamnaceae) Journalof Ecology 961252ndash1265
Price T 1997 Correlated evolution and independent contrastsTransactions of the Royal Society B Biological Sciences352519ndash529
Reich PB Walters MB Ellsworth DS 1997 From tropics to tun-dra global convergence in plant functioning Proceedings ofthe National Academy of Sciences of the United States ofAmerica 9413730ndash13734
Reich PB Wright IJ Cavender-Bares J Craine JM Oleksyn JWestoby M Walters MB 2003 The evolution of plant func-tional variation traits spectra and strategies InternationalJournal of Plant Sciences 164143ndash164
Russell-Smith J Whitehead P Cook G Hoare J 2003 Responseof Eucalyptus-dominated savanna to frequent fires lessonsfrom Munmarlary 1973ndash1996 Ecological Monographs73349ndash375
Safford HD Harrison SP 2004 Fire effects on plant diver-sity in serpentine vs sandstone chaparral Ecology 85539ndash548
Saura Mas S Shipley B Lloret F 2009 Relationship betweenpost fire regeneration and leaf economics spectrum inMediterranean woody species Functional Ecology 23103ndash110
Sawyer J Keeler-Wolf T Evans J 2009 A Manual ofCalifornia Vegetation Second Edition Sacramento CAUSA California Native Plant Society
Schmalzer PA 2003 Growth and recovery of oak-saw palmettoscrub through ten years after fire Natural Areas Journal235ndash13
Schwilk DW Ackerly DD 2005 Is there a cost to resproutingSeedling growth rate and drought tolerance in sprouting andnonsprouting Ceanothus (Rhamnaceae) 1 American Journalof Botany 92404ndash410
Shipley B Almeida-Cortez J 2003 Interspecific consistency andintraspecific variability of specific leaf area with respect toirradiance and nutrient availability Ecoscience 1074ndash79
Stearns S 1992 The evolution of life histories Oxford UKOxford University Press
Verduacute M 2000 Ecological and evolutionary differences betweenMediterranean seeders and resprouters Journal of VegetationScience 11265ndash268
Verduacute M Pausas JG 2007 Fire drives phylogenetic clusteringin Mediterranean Basin woody plant communities Ecology951316ndash1323
Vesk PA Westoby M 2004 Sprouting ability across diverse dis-turbances and vegetation types worldwide Journal of Ecology92310ndash320
Webb CO Ackerly DD McPeek MA Donoghue MJ 2002Phylogenies and community ecology Annual Review ofEcology and Systematics 33475ndash505
Weiher E Keddy PA 2001 Ecological assembly rules per-spectives advances retreats Cambridge UK CambridgeUniversity Press
Westerling AL Hidalgo HG Cayan DR Swetnam TW 2006Warming and earlier spring increase western US forest wild-fire activity Science 313940
Westoby M Falster DS Moles AT Vesk PA Wright IJ 2002Plant ecological strategies Some leading dimensions ofvariation between species Annual Review of Ecology andSystematics 33125ndash159
Westoby M Wright IJ 2006 Land-plant ecology on thebasis of functional traits Trends in Ecology amp Evolution21261ndash268
Willis CG Halina M Lehman C Reich PB Keen A McCarthyS Cavender-Bares J 2010 Phylogenetic community structurein Minnesota oak savanna is influenced by spatial extent andenvironmental variation Ecography 33565ndash577
Wisheu IC Rosenzweig ML Olsvig-Whittaker L Shmida A2000 What makes nutrient-poor Mediterranean heathlandsso rich in plant diversity Evolutionary Ecology Research2935ndash955
Wright IJ Reich PB Cornelissen JHC Falster DS GroomPK Hikosaka K Lee W Lusk CH Niinemets OleksynJ 2005 Modulation of leaf economic traits and trait rela-tionships by climate Global Ecology and Biogeography14411ndash422
Wright IJ Reich PB Westoby M Ackerly DD Baruch Z BongersF Cavender-Bares J Chapin T Cornelissen JHC DiemerM 2004 The worldwide leaf economics spectrum Nature428821ndash827
Wright IJ Westoby M Reich PB 2002 Convergence towardshigher leaf mass per area in dry and nutrient poor habitats hasdifferent consequences for leaf life span Journal of Ecology90534ndash543
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011

Effects of climate soil and fire on specific leaf area 3
Table 1 Expected characteristics of chaparral species with regard to post-fire regeneration strategy
Post-fire regeneration strategy
Characteristic Resprouter Seeder References
Plant levelAllocation to
storageHigh Low Pate et al 1990 Bell et al 1996 Bell 2001 Langley et al 2002
Knox and Clarke 2005 but see Chew and Bonser 2009Growth rate Low High Pate et al 1990 Bell et al 1996 Bell 2001 Bond and Midgley
2003 Schwilk and Ackerly 2005 but see Chew and Bonser2009
Height Uncertain Uncertain Midgley 1996 Bond and Midgley 2003 Falster and Westoby2005 Menges 2007
Rooting depth Deep Shallow Bell et al 1996 Bell 2001Fecundity Low High Lamont 1985 Verduacute 2000 Bell 2001 Bond and Midgley 2001
Lamont and Wiens 2003
Ecological strategyFire Tolerate Avoid Odion and Davis 2000 Clarke 2002a Clarke 2002b Clarke and
Knox 2002 Russell-Smith et al 2003 Pausas et al 2004Clarke et al 2005 Franklin et al 2005 Ojeda et al 2005Keeley et al 2006
DroughtAridity Avoid Tolerate Pausas et al 2004 Clarke et al 2005 Lloret et al 2005 Ojedaet al 2005 Pratt et al 2008 Saura Mas et al 2009 but seeLamont and Markey 1995
Soil infertility Avoid Tolerate Lamont and Markey 1995 Wisheu et al 2000 Safford andHarrison 2004
Shading Tolerate Avoid Hawkes and Menges 1996 Menges 2007 Pratt et al 2008Lifespan Long Short Bond and Midgley 2003 Lamont and Wiens 2003 Chew and
Bonser 2009Competitive
abilityUncertain Uncertain Schmalzer 2003 Clarke et al 2005 Schwilk and Ackerly 2005
Chew and Bonser 2009 Clarke and Knox 2009
from maritime (n = 17) to interior (n = 18) (Figure 1) Thisclimate transition integrates decreased presence of a sum-mer marine fog layer increased temperatures decreasedrelative humidity and decreased summer water availabilityField data from chaparral plots during the summer 2009showed large differences between maritime and interiorclimate zones (respectively) in daily average daily leaf wet-ness (502 plusmn 19 vs 38 plusmn 07) vapour pressure deficit(0323 plusmn 0027 vs 2154 plusmn 0090 kPa) average daily max-imum temperature (200 plusmn 04 vs 318 plusmn 04 C) andaverage daily soil water content (123 plusmn 01 vs 23 plusmn 02)(means plusmn one standard error Vasey et al in review)
Fire occurs less frequently in the maritime than theinterior climatic zone due to a shorter fire season associ-ated with cooler less seasonal temperatures higher rela-tive humidity lower evapotranspiration and higher rainfall(Odion and Tyler 2002) However chaparral in the maritimeclimatic zone is found on infertile shallow and sandy soilsrelative to chaparral in the interior climatic zone becausethese edaphic factors favour the persistence of chaparralvegetation (Griffin 1978 Sawyer et al 2009)
Plant community composition and soil variables
At McLaughlin woody plant community composition andcover was measured in 2002 Visual cover was estimated asa measure of species abundances for species in each of five1 m2 quadrats spaced evenly along the centre-line of eachplot Cover values were then averaged for each species and
each plot For each plot 50 g of soil were collected fromthe centre of each quadrat at 5ndash10 cm depth mixed andthen analysed by A amp L Western Agricultural LaboratoriesModesto California USA for organic matter K Mg Caand Bray P
At the Central Coast plots the plant compositionand cover of woody plants for each of the 35 plotseach 50 20 m was surveyed between 2008 and 2009(M Vasey et al unpublished) Visual cover was estimatedin 10 quadrats each 10 10 m to provide a mean abun-dance per species per plot Soil was collected from eachquadrat at 5ndash10 cm depth mixed together and analysed byBrookside Laboratories New Knoxville Ohio USA fororganic matter K Mg Ca and Bray P
Climate and fire variables
To obtain climate data for each Central Coast plot weintersected plot locations with maps of 10 macrocli-mate variables from the climate mapping system for theUnited States PRISM (httpwwwprismoregonstateedu)Variables were based on 30-year averages and had a res-olution of 1 km2 Variables were relative humidity ()potential evapotranspiration (cm) annual temperature (C)minimum winter temperature (C) maximum summer tem-perature (C) temperature seasonality (standard deviation)annual precipitation winter precipitation summer precip-itation and precipitation seasonality (coefficient of varia-tion)
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
4 B Anacker et al
sandstoneserpentine
maritimeinterior
McLaughlin
San Francisco
Santa Cruz
Santa Barbara
0 64 128km
Figure 1 Locations of 115 chaparral study plots in westernCalifornia Eighty plots were surveyed at McLaughlin aUniversity of California Natural Reserve stratified by soil type(sandstone and serpentine) and burn history (recently burned andunburned) In addition 35 plots were surveyed on sandstone soilsin the Central Coast region these plots were stratified by climaticzone (maritime and interior)
To obtain data on fire regimes we intersected plotlocations with raster grids (250 m2) of lsquofuel rankrsquo andlsquofire rotationrsquo from the California Department of Forestryand Fire Protectionrsquos Fire and Resource AssessmentProgram (CDF 2003 Fire Regime and Condition ClassEdition 03_2 httpfrapcdfcagovdatafrapgisdataselectasptheme=5) Fuel rank is based on topography vegeta-tive fuel loads and climate and varied from 1 for lsquolittle orno fire hazardrsquo to 3 for lsquovery high fire hazardrdquo The fire rota-tion data are also given in classes and are based on annualacres burned vegetation type development status eleva-tion and the bioregion values range from 1 for lsquomoderatefrequencyrsquo to 3 for lsquohigh frequencyrsquo
Trait data ndash SLA and post-fire regeneration strategy
Mean SLA (mm2 mg1) was obtained for the 87 speciesfound in the plots (see Supplementary Table 1 inAppendix 1 available via the supplementary tab on the arti-clersquos online page at httpdxdoiorg101080175508742011633573) from several sources primarily from newfield measurements For 66 of the species SLA was mea-sured from new field collections where 10 leaves werecollected from each of 10 individuals Individuals were
sampled from at least three unique populations To accountfor phenotypic plasticity of SLA with respect to soil forspecies occurring on both serpentine and non-serpentine(34 of the 66 species) five individuals were sampled fromeach soil for the others all individuals were sampledfrom one soil type Mean SLA per species was determinedusing a standard protocol (Cornelissen et al 2003) Fornine species SLA data were available from The EcologicalFlora of California (httpucjepsberkeleyeduefc) so newfield measurements were not sought For an additional12 Arctostaphylos that were very geographically rare wemeasured SLA using our own dried herbarium specimensTo validate using SLA from dried leaves versus fresh leavesfor Arctostaphylos we measured SLA from both dry leavesand from fresh leaves for 20 additional more commonArctostaphylos taxa and found that the two measures werehighly correlated (r2 = 088 n = 20 b = 101 intercept= 009 P lt 0001) To examine intraspecific variation ofSLA by soil type soil type-specific SLA means were alsocalculated for the 34 species collected on two soils In allanalyses described below mean SLA was log-transformedto meet the assumptions of parametric analyses It is pos-sible that the SLA values we determined for these specieswould differ if measured in other environments than thoseselected because genotypes and environments interact todetermine phenotypes
For each of the 87 species we determined post-fireregeneration strategy using Hickman (1993) published lit-erature and expert opinion by the authors (primarily JK)We designated each species as an obligate resprouter obli-gate seeder or facultative resprouter (can resprout or recruitfrom seed following fire) Only 20 of our species can bothresprout and post-fire seed suggesting a negative trade-off between resprouting capacity and propagule persistence(Pausas 2001 Pausas and Verdu 2005)
Analyses
Correlated environmental filters We tested if field-measured edaphic variables remotely sensed macroclimatevariables andor remotely sensed fire attributes differedamong climatic zones (interior maritime) using one-wayANOVA Relationships of soil fertility fire frequency andintensity and recent burn history at McLaughlin describedabove are reported elsewhere (Safford and Harrison 2004)and thus new analyses were not conducted here
Correlated traits At the species level (n = 87) we testedwhether SLA differed by post-fire regeneration strategyusing one-way ANOVA and also repeated this analy-sis using a correction for phylogenetic non-independence(comparative analysis by independent contrasts (CAIC)details in Appendix 2 available online as before)
The percentage of obligate seeders in each plot wascalculated percentages were weighted by abundance todown-weight the relative influence of rare species on
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
Effects of climate soil and fire on specific leaf area 5
the description of functional structure The abundance-weighted percentage for each plot was calculated as fol-lows (sum abundance obligate seeders sum abundanceof all species) 100 We calculated community-level post-fire strategy with obligate seeders as the numerator becauseboth facultative resprouters and obligate resprouters areecologically similar in the sense that they can both resproutFor McLaughlin we tested if the percentage of obligateseeders differed by soil type burn history and their interac-tion (two-way ANOVA) Prior to analysis the percentage ofobligate seeders was arc-sine transformed For the CentralCoast dataset we tested if the percentage of seeders differedamong climatic zones using one-way ANOVA
Environmental correlates of SLA At the community levelwe tested if the mean abundance-weighted SLA observedin each plot at McLaughlin differed by soil type burn his-tory and their interaction (two-way ANOVA) For each plotthe mean abundance-weighted SLA was calculated by sum-ming the product of each speciesrsquo SLA and its proportionof total cover in that plot and then dividing by the numberof species in that plot We used the proportion of a speciescover in each plot as our metric of abundance rather thanabundance alone To examine for intraspecific variation inSLA by soil type we compared the SLA of sandstonecollections with the SLA of serpentine collections for the34 species collected from both soil types using a pairedt-test For the Central Coast we tested if abundance-weighted mean SLA differed among climatic zones usingone-way ANOVA
Environmental filtering of SLA To test for filtering of SLAthe range of SLA for each community was calculated(rangeobs) and compared with a null expectation of therange of SLA at the observed level of richness (rangeexp)(sensu Cornwell and Ackerly 2009) We randomly sam-pled the species pool by data set 9999 times for eachlevel of observed species richness among the plots andcalculated the mean range of SLA values for each level ofspecies richness Communities in which a filter is operatingwill have rangeexp gt rangeobs We tested if the distribu-tion of rangeexp rangeobs was significantly greater thanzero using a one-tailed t-test We then tested if rangeexp rangeobs in each plot differed by soil type burn history andclimatic zone using a two-way ANOVA All analyses wereperformed in R version 120 (R Development Core Team2010)
Results
Correlated environmental filters
Serpentine plots had a significantly lower CaMg ratiohigher organic matter content and lower P and K levelsthan the sandstone plots The less fertile maritime plots hada significantly lower CaMg ratio and lower P level than theinterior plots (Table 2) There were higher fuel levels andalso more frequent fires in interior plots than in maritime
plots (Table 2) For the macroclimate variables interiorplots were shown to be significantly more arid than mar-itime plots (lower relative humidity higher potential evap-otranspiration higher summer temperature lower wintertemperature and higher temperature seasonality) althoughprecipitation did not differ by zone (Supplementary Table 2in Appendix 1 available online)
Correlated traits
At the species level mean SLA was significantly higher inobligate resprouters than in either facultative resproutersor obligate seeders (one-way ANOVA F2 84 = 164P lt 0001) (Figure 2) This relationship was not signif-icant after correction for phylogenetic non-independence(P = 0327) (Appendix 2 available online) demonstratingshared variation among higher taxa
The abundance-weighted percentage of seeders wassignificantly lower on sandstone soils compared to serpen-tine soils (Figure 3a) but this soil contrast disappeared inrecently burned plots (two-way ANOVA soil F1 76 = 262P lt 0001 burn history F1 76 = 219 P lt 0001 soil burn history F176 = 294 P = 009 Figure 3a) For theCentral Coast plots there was a higher abundance-weightedpercentage of obligate seeders in the low-fire maritime cli-matic zone compared with the high-fire interior climaticzone (Table 2 Figure 3c) The Central Coast compared withMcLaughlin had a lesser proportion of obligate resprouters(040 vs 054) more obligate seeders (038 vs 027)and roughly the same proportion of facultative seeders(022 vs 019)
Environmental correlates of SLA
In the McLaughlin plots abundance-weighted mean SLAwas significantly higher on sandstone soils compared withserpentine soils (Figure 3b) regardless of burn history(two-way ANOVA soil F1 76 = 203 P lt 0001 burn his-tory F1 76 = 10 P = 031 soil burn history F1 76 = 03P = 059) Plant abundances were significantly lower onserpentine soils but this difference was only present amongthe unburned plots (Figure 3c two-way ANOVA soil F1 76
= 61 P = 0015 burn history F1 76 = 6905 P lt 0001soil burn history F1 76 = 49 P = 003) In the CentralCoast plots abundance-weighted mean SLA was not sig-nificantly different between climatic zones (Figure 3e) norwas abundance (Figure 3f)
Conspecifics had 18 higher mean SLA on sandstonethan serpentine soil (paired t-test t = 231 df = 33P = 0027) At the community level in contrast meanunweighted SLA was even greater with values in sand-stone plots 32 higher than in serpentine Thus variationin SLA among communities was due to both compositionalturnover and intraspecific variation (ie phenotypic plastic-ity) but species replacement played a larger role similarto previous results for Californian woody plants (Cornwelland Ackerly 2009)
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
6 B Anacker et al
Table 2 Environmental differences by soil type (McLaughlin plots sandstone serpentine) and climatic zone (Central Coast plotsinterior maritime)
McLaughlin Central Coast
Sandstone Serpentine F P Interior Maritime F P
EdaphicCaMg ratio 34 plusmn 22 05 plusmn 05 691 lt 0001 69 plusmn 16 45 plusmn 34 72 0011Bray P (ppm) 70 plusmn 26 34 plusmn 18 530 lt 0001 376 plusmn 236 144 plusmn 335 55 0025K (ppm) 2278 plusmn 628 1285 plusmn 713 440 lt 0001 2211 plusmn 767 1726 plusmn 886 30 0094Organic
matter ()43 plusmn 09 52 plusmn 15 95 0003 50 plusmn 38 65 plusmn 18 22 0152
FireFuel rank1 na na na na 21 plusmn 05 16 plusmn 07 66 0015Fire rotation na na na na 26 plusmn 06 15 plusmn 11 153 0001
Notes Mean plusmn one standard deviation P values are reported from one-way ANOVA of each variable with soil type or climatic zone Degrees of freedomfor McLaughlin are 1 and 78 for the factor and the residuals respectively Degrees of freedom for Central Coast are 1 and 33 for the factor and theresiduals respectively1Higher fuel ranks correspond to higher potential fire hazard
10
15
20
25
30
35
40
All species (n = 87) found in plots
Post-fire regeneration strategy
log(
SLA
mm
2 m
g1)
Obligateresprouters
Facultativeresprouters
Obligateseeders
a
b
b
Figure 2 Relationship of SLA and post-fire regeneration strat-egy among the 87 species considered here Letters above box-plots indicate significant differences based on a post-hoc TukeyrsquosHSD test Open circles indicate observations considered to beoutliers
Environmental filtering of SLA
The range of SLA values for the McLaughlin plots wassignificantly lower than expected under the null modelindicating environmental filtering (54 of 80 had rangeexp rangeobs lt 0 P = 0004) (Figures 4a b) Filtering strength(rangeexp rangeobs) was significantly higher on sandstonethan serpentine (one-way ANOVA soil F1 78 = 157 P =00002 Table 2) Filtering strength was also related to burnhistory and soil type burn history (two-way ANOVAsoil F1 76 = 184 P lt 0001 burn history F1 76 = 116P = 0001 soil burn history F1 76 = 42 P = 0044) Theinteraction term reflected that filtering was significant forunburned plots on both soils and for recently burned plotson serpentine but not recently burned plots on sandstoneThe range of SLA values for the Central Coast plots was not
significantly lower than expected (17 of 35 had rangeexp rangeobs lt 0 P = 0378) indicating a lack of filtering byclimatic zone (Table 2 Figure 4c)
Discussion
Among Californian chaparral communities varying in cli-mate soil fertility and fire histories we found that SLAa critical indicator of resource-use strategy did not followthe simple patterns anticipated from considering individ-ual filters Within the McLaughlin data SLA was lower oninfertile serpentine soils as expected but fire also played arole in broadening the range of SLA present on more fer-tile sandstone soils (relaxing filtering) Within the CentralCoast data SLA did not follow any simple expectationsbeing higher neither in more equable maritime climatesnor in the interior where soils are more fertile and fires aremore frequent These results are most consistent with thehypothesis that SLA is affected by an interaction of climatesoil fertility and fire Key to understanding this interac-tion are the correlations we found at both the species andcommunity levels between SLA and post-fire regenerationstrategies and also the environmental correlations betweenclimate soils and fire frequency
Obligate post-fire seeders had significantly lower SLAthan obligate post-fire resprouters (Figure 2) This contra-dicts the notion that the reduced need for storage of theseeding strategy should lead to relatively higher growthrates and associated higher SLA in seeders (Table 1) Thecorrelation we observed may reflect a physiological trade-off in which obligate seeding imposes strong selection fordrought tolerance at the seedling stage (Ackerly 2004)Infertile soils may also select for drought-tolerant seed-ers due to imposing arid conditions (high rock contenthigh exposure to direct sun high rates of soil evapora-tion) (Ojeda et al 2005 Pratt et al 2008) While seedershave low allocation to roots and shallow rooting depth andthus a limited ability to avoid drought by reaching water inlower soil horizons they may tolerate soil aridity if they
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
Effects of climate soil and fire on specific leaf area 7
020406080
100120140
McLaughlin
Soil and Burn historyasin
(abu
ndan
cew
eigh
ted
o
blig
ate
seed
ers)
B U B USandstone Serpentine
a
aa
bA
0001020304050607
McLaughlin
Soil and burn historyA
bund
ance
wei
ghte
d lo
g(S
LA)
B U B USandstone Serpentine
a a
b b
B
0
50
100
150
McLaughlin
Soil and burn history
Abu
ndan
ce
B U B USandstone Serpentine
a
b
a
c
C
020406080
100120140
Central Coast
Climatic zoneasin
(abu
ndan
cew
eigh
ted
o
blig
ate
seed
ers)
Interior Maritime
Climatic zone
Interior Maritime
Climatic zone
Interior Maritime
a
bD
0001020304050607
Central Coast
Abu
ndan
cew
eigh
ted
log(
SLA
)
a
a
E
0
50
100
150
Central Coast
Abu
ndan
ce
aaF
Figure 3 Plant community attributes for serpentine and sandstone soils (andashc) and interior and maritime climatic zones (dndashf) Recentfire history is indicated for the McLaughlin data as follows B recently burned U unburned Letters above boxplots indicate significantdifferences based on a post-hoc Tukeyrsquos HSD test Open circles indicate observations considered to be outliers
have leaf-level strategies such as low SLA that providean advantage under low-resource conditions (Poorter et al2009 Saura Mas et al 2009) Post-fire resprouters in con-trast avoid drought by having deeper rooting systems (Bellet al 1996 Bell 2001)
The correlation between post-fire regeneration strategyand SLA was not significant in an analysis of phyloge-netically independent contrasts reflecting the influence ofspecies-rich lineages (ie Arctostaphylos and CeanothusAppendices 1 and 2 available online) that share particu-lar trait combinations (eg post-fire seeding and low SLA)through common inheritance This means that the obser-vation of low SLA and obligate seeding in a communityis in part due to common descent In other words anenvironmental filter may be directly selecting for obligateseeding and thereby indirectly selecting for the corre-lated low SLA value or vice versa Similarly a study of37 plants of the Iberian peninsula showed phylogeneticnon-independence among post-fire regeneration strategyand fruit type (Pausas and Verdu 2005) Such patterns ofphylogenetic non-independence in traits are well knownand form the basis of phylogenetic clustering in communitystructure (Webb et al 2002 Anacker 2011)
The observed patterns of post-fire regeneration strategyconfirmed that fire was more frequent on sandstone rela-tive to serpentine (obligate seeders were more common inserpentine chaparral) and that fire was more frequent in
the interior than on the coast (obligate seeders more com-mon in maritime chaparral) In the arid interior the infertileconditions of serpentine soil limit fuel accumulation cre-ating local fire refugia and favouring seeders Along thecoast cool and wet climates limit the length and severityof the fire season and the flammability of fuels leadingto low regional fire frequencies favouring seeders lowsoil fertility is likely important in determining the balancebetween woodland and chaparral (Griffin 1978 Sawyeret al 2009) rather than affecting fire regime Anotherfactor favouring obligate seeding in maritime chaparralis that longer time intervals between fires may increasethe severity of fires when they do occur lowering adultsurvivorship
Chaparral on serpentine soils at McLaughlin had sig-nificantly lower SLA than chaparral on sandstone evenimmediately following a fire (Figure 3) suggesting thatsoils play the dominant role in shaping SLA in this settingHowever our observation that low-SLA obligate seedersare much more prevalent on infrequently burning serpen-tine stands (Figure 3) suggests that fire does interact withsoil to determine community-level SLA over longer timeperiods In our Central Coast data SLA did not differbetween maritime and interior plots even though mesicmaritime climates should promote high SLA near the coastwhile both frequent fire and more fertile soils are predictedto lead to higher SLA in the interior Abundant evidence
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
8 B Anacker et al
5 10 15
00
05
10
15
20
25
30
McLaughlin sandstone
Richness
Ran
ge o
f lo
g(S
LA)
burnedunburned
expectedA
5 10 15
00
05
10
15
20
25
30
McLaughlin serpentine
Richness
Ran
ge o
f lo
g(S
LA)
burnedunburned
B
5 10 15
00
05
10
15
20
25
30
Central Coast sandstone
Richness
Ran
ge o
f lo
g(S
LA)
maritimeinterior
C
Figure 4 The range of SLA observed in chaparral plots versusnull expectation Points below the line have lower observed rangesthan expected by chance
from other studies shows that SLA tends to be higher inproductive habitats (reviewed in Cornelissen et al 2003Westoby and Wright 2006 Poorter et al 2009) such as ourmaritime plots where water availability is high Thus oneof our more striking findings is that the globally predomi-nant role of climate in shaping SLA can be nullified by themore localised effects of soil fertility and fire if these gra-dients happen to vary in the opposite direction with respectto their effects on SLA
Conclusions
We found that community structure in Californianchaparral is shaped by the environmental filtering of a keyresource-acquisition trait ndash SLA ndash by the simultaneous
effects of climate soil fertility and fire Interactiveeffects of these three filters arose from two factors thephylogenetic and functional non-independence of SLA andanother critical trait (post-fire regeneration strategy) andthe non-independence of the three environmental filtersthemselves Our results provide a clear warning that incor-rect inferences about functional and phylogenetic commu-nity structure may be reached when either traits or filtersare considered in isolation
AcknowledgementsWe thank Naveed Davoodian for assistance in compiling relevantliterature and early discussions on the role of fire and substrateon reproductive strategies in the California flora and Tom Parkerand Tanner Harris for comments on an earlier version of thismanuscript
Notes on contributorsBrian Anacker is a post doctoral researcher at University ofCalifornia Davis His research addresses the historical and eco-logical controls of diversity in Californian plant communities
Nishanta Rajakaruna is a professor of botany at College of theAtlantic Bar Harbor ME USA His research focuses on thefactors and mechanisms driving plant speciation and communityassembly on serpentine and other edaphically harsh environments
David Ackerly is an associate professor at University ofCalifornia Berkeley His research addresses the ecological andevolutionary processes that shape the functional diversity ofterrestrial plants with a particular focus on the California flora
Susan Harrison is a professor at University of California DavisThe major theme of her research is understanding the processesthat shape and maintain plant species diversity at the landscapescale where small-scale forces such as competition and facilita-tion interact with large-scale forces such as niche evolution anddispersal
Jon Keeley is a research ecologist with the US Geological Surveyand adjunct professor at University of California Los AngelesHis research focuses on the ecological impacts and history ofwildfires in Mediterranean-climate ecosystems
Michael Vasey is a graduate student at University of CaliforniaSanta Cruz His research is focused on the importance of themarine layer (coastal fog) to the ecology and diversity of chaparralvegetation and manzanitas
ReferencesAckerly DD 2004 Functional strategies of chaparral shrubs in
relation to seasonal water deficit and disturbance EcologicalMonographs 7425ndash44
Ackerly D Knight C Weiss S Barton K Starmer K 2002Leaf size specific leaf area and microhabitat distribution ofchaparral woody plants contrasting patterns in species leveland community level analyses Oecologia 130449ndash457
Anacker BL 2011 Phylogenetic patterns of serpentine plantendemism and diversity In Harrison SP Rajakaruna Neditors Serpentine as a model in evolution and ecologyBerkeley California USA University of California Pressp 49ndash70
Bell DT 2001 Ecological response syndromes in the floraof southwestern Western Australia fire resprouters versusreseeders The Botanical Review 67417ndash440
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
Effects of climate soil and fire on specific leaf area 9
Bell TL Pate JS Dixon KW 1996 Relationships between fireresponse morphology root anatomy and starch distributionin south-west Australian Epacridaceae Annals of Botany77357
Bond WJ Midgley JJ 2001 The persistence niche ecology ofsprouting in woody plants Trends in Ecology and Evolution1645ndash51
Bond WJ Midgley JJ 2003 The evolutionary ecology of sprout-ing in woody plants International Journal of Plant Sciences164103ndash114
Burns K 2004 Patterns in specific leaf area and the structureof a temperate heath community Diversity and Distributions10105ndash112
Cavender-Bares J Kozak KH Fine PVA Kembel SW 2009 Themerging of community ecology and phylogenetic biologyEcology Letters 12693ndash715
Chew SJ Bonser SP 2009 The evolution of growth rate resourceallocation and competitive ability in seeder and resprouter treeseedlings Evolutionary Ecology 23723ndash735
Clarke PJ 2002a Habitat insularity and fire response traitsevidence from a sclerophyll archipelago Oecologia132582ndash591
Clarke PJ 2002b Habitat islands in fire-prone vegetation do land-scape features influence community composition Journal ofBiogeography 29677ndash684
Clarke PJ Knox KJE 2002 Post-fire response of shrubs inthe tablelands of eastern Australia do existing modelsexplain habitat differences Australian Journal of Botany 5053ndash62
Clarke PJ Knox KJE 2009 Trade offs in resource alloca-tion that favour resprouting affect the competitive ability ofwoody seedlings in grassy communities Journal of Ecology971374ndash1382
Clarke PJ Knox KJE Wills KE Campbell M 2005 Landscapepatterns of woody plant response to crown fire distur-bance and productivity influence sprouting ability Ecology93544ndash555
Cornelissen JHC Lavorel S Garnier E Diaz S Buchmann NGurvich DE Reich PB Steege H Morgan HD HeijdenMGA 2003 A handbook of protocols for standardisedand easy measurement of plant functional traits worldwideAustralian Journal of Botany 51335ndash380
Cornwell WK Ackerly DD 2009 Community assembly andshifts in plant trait distributions across an environmen-tal gradient in coastal California Ecological Monographs79109ndash126
Diamond JM 1975 Assembly of species communities In CodyML Diamond JM editors Ecology and evolution of com-munities Cambridge MA USA Harvard University Pressp 342ndash444
Diaz S Cabido M Casanoves F 1998 Plant functional traits andenvironmental filters at a regional scale Journal of VegetationScience 9113ndash122
Falster DS Westoby M 2005 Tradeoffs between height growthrate stem persistence and maximum height among plantspecies in a post fire succession Oikos 11157ndash66
Franklin J Syphard AD He HS Mladenoff DJ 2005 Altered fireregimes affect landscape patterns of plant succession in thefoothills and mountains of southern California Ecosystems8885ndash898
Griffin JR 1978 Maritime chaparral and endemic shrubs of theMonterey Bay region California Madrono 2565ndash81
Grime JP 1979 Plant strategies and vegetation processesChichester UK John Wiley amp Sons Ltd
Grubb PJ 1977 The maintenance of species-richness in plantcommunities the importance of the regeneration nicheBiological Reviews 52107ndash145
Hawkes CV Menges ES 1996 The relationship between openspace and fire for species in a xeric Florida shrubland Bulletinof the Torrey Botanical Club 12381ndash92
Hickman JC editor 1993 The Jepson Manual Higher plantsof California Berkeley California USA University ofCalifornia Press
Hoffmann WA Franco AC Moreira MZ Haridasan M 2005Specific leaf area explains differences in leaf traits betweencongeneric savanna and forest trees Functional Ecology19932ndash940
Hopper SD Brown AP Marchant NG 1997 Plants of WesternAustralian granite outcrops Journal of the Royal Society ofWestern Australia 80141ndash158
Keeley JE 1981 Reproductive cycles and fire regimes InProceedings of the Conference Fire Regimes and EcosystemProperties vol General Technical Report WO-26 USDAForest Service p 231ndash277
Keeley JE Fotheringham CJ Baer-Keeley M 2006 Demographicpatterns of postfire regeneration in Mediterranean-climate shrublands of California Ecological Monographs76235ndash255
Keeley JE Keeley SC 2000 Chaparral North American terres-trial vegetation 2204ndash253
Keeley JE Zedler PH 1978 Reproduction of chaparral shrubsafter fire a comparison of sprouting and seeding strategiesAmerican Midland Naturalist 99142ndash161
Knops JMH Reinhart K 2000 Specific leaf area along anitrogen fertilization gradient American Midland Naturalist265ndash272
Knox KJE Clarke PJ 2005 Nutrient availability induces contrast-ing allocation and starch formation in resprouting and obligateseeding shrubs Functional Ecology 19690ndash698
Lamont BB 1985 The comparative reproductive biology ofthree Leucospermum species (Proteaceae) in relation to fireresponses and breeding system Australian Journal of Botany33139ndash145
Lamont BB Markey A 1995 Biogeography of fire-killedand resprouting Banksia species in south-western AustraliaAustralian Journal of Botany 43283ndash303
Lamont BB Wiens D 2003 Are seed set and speciation ratesalways low among species that resprout after fire and whyEvolutionary Ecology 17277ndash292
Langley J Drake B Hungate B 2002 Extensive belowground car-bon storage supports roots and mycorrhizae in regeneratingscrub oaks Oecologia 131542ndash548
Lloret F Estevan H Vayreda J Terradas J 2005 Fire regenerativesyndromes of forest woody species across fire and climaticgradients Oecologia 146461ndash468
Lusk C Onoda Y Kooyman R Gutierrez Girun A 2010Reconciling species level vs plastic responses of evergreenleaf structure to light gradients shade leaves punch abovetheir weight New Phytologist 186429ndash438
McGill BJ Enquist BJ Weiher E Westoby M 2006 Rebuildingcommunity ecology from functional traits Trends in Ecologyamp Evolution 21178ndash185
Meinzer FC 2003 Functional convergence in plant responses tothe environment Oecologia 1341ndash11
Menges ES 2007 Integrating demography and fire managementan example from Florida scrub Australian Journal of Botany55261ndash272
Messier J McGill BJ Lechowicz MJ 2010 How do traits varyacross ecological scales A case for trait based ecologyEcology Letters 13838ndash848
Midgley JJ 1996 Why the worldrsquos vegetation is not totally dom-inated by resprouting plants because resprouters are shorterthan reseeders Ecography 1992ndash95
Niinemets U 2001 Global-scale climatic controls of leaf dry massper area density and thickness in trees and shrubs Ecology82453ndash469
Odion D Tyler C 2002 Are long fire-free periods needed tomaintain the endangered fire-recruiting shrub Arctostaphylosmorroensis (Ericaceae) Conservation Ecology 64 [online]URL httpwwwconsecolorgvol6iss2art4
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
10 B Anacker et al
Odion DC Davis FW 2000 Fire soil heating and the formationof vegetation patterns in chaparral Ecological Monographs70149ndash169
Ojeda F Brun FG Vergara JJ 2005 Fire rain and the selection ofseeder and resprouter life-histories in fire-recruiting woodyplants New Phytologist155ndash165
Pate JS Froend RH Bown BJ Hansen A Kuo J 1990 Seedlinggrowth and storage characteristics of seeder and resprouterspecies of Mediterranean-type ecosystems of SW AustraliaAnnals of Botany 65585
Paula S Pausas J 2006 Leaf traits and resprouting abil-ity in the Mediterranean basin Functional Ecology 20941ndash947
Pausas J Verdu M 2005 Plant persistence traits in fire-prone ecosystems of the Mediterranean basin a phylogeneticapproach Oikos 109196ndash202
Pausas JG 2001 Resprouting vs seeding ndash a Mediterraneanperspective Oikos 94193ndash194
Pausas JG Bradstock RA Keith DA Keeley JE 2004 Plantfunctional traits in relation to fire in crown-fire ecosystemsEcology 851085ndash1100
Pausas JG Keeley JE VerdUacute M 2006 Inferring differential evo-lutionary processes of plant persistence traits in NorthernHemisphere Mediterranean fire-prone ecosystems Ecology9431ndash39
Poorter H Niinemets Poorter L Wright IJ Villar R 2009Causes and consequences of variation in leaf mass perarea (LMA) a meta analysis New Phytologist 182565ndash588
Pratt RB Jacobsen AL Mohla R Ewers FW Davis SD 2008Linkage between water stress tolerance and life history typein seedlings of nine chaparral species (Rhamnaceae) Journalof Ecology 961252ndash1265
Price T 1997 Correlated evolution and independent contrastsTransactions of the Royal Society B Biological Sciences352519ndash529
Reich PB Walters MB Ellsworth DS 1997 From tropics to tun-dra global convergence in plant functioning Proceedings ofthe National Academy of Sciences of the United States ofAmerica 9413730ndash13734
Reich PB Wright IJ Cavender-Bares J Craine JM Oleksyn JWestoby M Walters MB 2003 The evolution of plant func-tional variation traits spectra and strategies InternationalJournal of Plant Sciences 164143ndash164
Russell-Smith J Whitehead P Cook G Hoare J 2003 Responseof Eucalyptus-dominated savanna to frequent fires lessonsfrom Munmarlary 1973ndash1996 Ecological Monographs73349ndash375
Safford HD Harrison SP 2004 Fire effects on plant diver-sity in serpentine vs sandstone chaparral Ecology 85539ndash548
Saura Mas S Shipley B Lloret F 2009 Relationship betweenpost fire regeneration and leaf economics spectrum inMediterranean woody species Functional Ecology 23103ndash110
Sawyer J Keeler-Wolf T Evans J 2009 A Manual ofCalifornia Vegetation Second Edition Sacramento CAUSA California Native Plant Society
Schmalzer PA 2003 Growth and recovery of oak-saw palmettoscrub through ten years after fire Natural Areas Journal235ndash13
Schwilk DW Ackerly DD 2005 Is there a cost to resproutingSeedling growth rate and drought tolerance in sprouting andnonsprouting Ceanothus (Rhamnaceae) 1 American Journalof Botany 92404ndash410
Shipley B Almeida-Cortez J 2003 Interspecific consistency andintraspecific variability of specific leaf area with respect toirradiance and nutrient availability Ecoscience 1074ndash79
Stearns S 1992 The evolution of life histories Oxford UKOxford University Press
Verduacute M 2000 Ecological and evolutionary differences betweenMediterranean seeders and resprouters Journal of VegetationScience 11265ndash268
Verduacute M Pausas JG 2007 Fire drives phylogenetic clusteringin Mediterranean Basin woody plant communities Ecology951316ndash1323
Vesk PA Westoby M 2004 Sprouting ability across diverse dis-turbances and vegetation types worldwide Journal of Ecology92310ndash320
Webb CO Ackerly DD McPeek MA Donoghue MJ 2002Phylogenies and community ecology Annual Review ofEcology and Systematics 33475ndash505
Weiher E Keddy PA 2001 Ecological assembly rules per-spectives advances retreats Cambridge UK CambridgeUniversity Press
Westerling AL Hidalgo HG Cayan DR Swetnam TW 2006Warming and earlier spring increase western US forest wild-fire activity Science 313940
Westoby M Falster DS Moles AT Vesk PA Wright IJ 2002Plant ecological strategies Some leading dimensions ofvariation between species Annual Review of Ecology andSystematics 33125ndash159
Westoby M Wright IJ 2006 Land-plant ecology on thebasis of functional traits Trends in Ecology amp Evolution21261ndash268
Willis CG Halina M Lehman C Reich PB Keen A McCarthyS Cavender-Bares J 2010 Phylogenetic community structurein Minnesota oak savanna is influenced by spatial extent andenvironmental variation Ecography 33565ndash577
Wisheu IC Rosenzweig ML Olsvig-Whittaker L Shmida A2000 What makes nutrient-poor Mediterranean heathlandsso rich in plant diversity Evolutionary Ecology Research2935ndash955
Wright IJ Reich PB Cornelissen JHC Falster DS GroomPK Hikosaka K Lee W Lusk CH Niinemets OleksynJ 2005 Modulation of leaf economic traits and trait rela-tionships by climate Global Ecology and Biogeography14411ndash422
Wright IJ Reich PB Westoby M Ackerly DD Baruch Z BongersF Cavender-Bares J Chapin T Cornelissen JHC DiemerM 2004 The worldwide leaf economics spectrum Nature428821ndash827
Wright IJ Westoby M Reich PB 2002 Convergence towardshigher leaf mass per area in dry and nutrient poor habitats hasdifferent consequences for leaf life span Journal of Ecology90534ndash543
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011

4 B Anacker et al
sandstoneserpentine
maritimeinterior
McLaughlin
San Francisco
Santa Cruz
Santa Barbara
0 64 128km
Figure 1 Locations of 115 chaparral study plots in westernCalifornia Eighty plots were surveyed at McLaughlin aUniversity of California Natural Reserve stratified by soil type(sandstone and serpentine) and burn history (recently burned andunburned) In addition 35 plots were surveyed on sandstone soilsin the Central Coast region these plots were stratified by climaticzone (maritime and interior)
To obtain data on fire regimes we intersected plotlocations with raster grids (250 m2) of lsquofuel rankrsquo andlsquofire rotationrsquo from the California Department of Forestryand Fire Protectionrsquos Fire and Resource AssessmentProgram (CDF 2003 Fire Regime and Condition ClassEdition 03_2 httpfrapcdfcagovdatafrapgisdataselectasptheme=5) Fuel rank is based on topography vegeta-tive fuel loads and climate and varied from 1 for lsquolittle orno fire hazardrsquo to 3 for lsquovery high fire hazardrdquo The fire rota-tion data are also given in classes and are based on annualacres burned vegetation type development status eleva-tion and the bioregion values range from 1 for lsquomoderatefrequencyrsquo to 3 for lsquohigh frequencyrsquo
Trait data ndash SLA and post-fire regeneration strategy
Mean SLA (mm2 mg1) was obtained for the 87 speciesfound in the plots (see Supplementary Table 1 inAppendix 1 available via the supplementary tab on the arti-clersquos online page at httpdxdoiorg101080175508742011633573) from several sources primarily from newfield measurements For 66 of the species SLA was mea-sured from new field collections where 10 leaves werecollected from each of 10 individuals Individuals were
sampled from at least three unique populations To accountfor phenotypic plasticity of SLA with respect to soil forspecies occurring on both serpentine and non-serpentine(34 of the 66 species) five individuals were sampled fromeach soil for the others all individuals were sampledfrom one soil type Mean SLA per species was determinedusing a standard protocol (Cornelissen et al 2003) Fornine species SLA data were available from The EcologicalFlora of California (httpucjepsberkeleyeduefc) so newfield measurements were not sought For an additional12 Arctostaphylos that were very geographically rare wemeasured SLA using our own dried herbarium specimensTo validate using SLA from dried leaves versus fresh leavesfor Arctostaphylos we measured SLA from both dry leavesand from fresh leaves for 20 additional more commonArctostaphylos taxa and found that the two measures werehighly correlated (r2 = 088 n = 20 b = 101 intercept= 009 P lt 0001) To examine intraspecific variation ofSLA by soil type soil type-specific SLA means were alsocalculated for the 34 species collected on two soils In allanalyses described below mean SLA was log-transformedto meet the assumptions of parametric analyses It is pos-sible that the SLA values we determined for these specieswould differ if measured in other environments than thoseselected because genotypes and environments interact todetermine phenotypes
For each of the 87 species we determined post-fireregeneration strategy using Hickman (1993) published lit-erature and expert opinion by the authors (primarily JK)We designated each species as an obligate resprouter obli-gate seeder or facultative resprouter (can resprout or recruitfrom seed following fire) Only 20 of our species can bothresprout and post-fire seed suggesting a negative trade-off between resprouting capacity and propagule persistence(Pausas 2001 Pausas and Verdu 2005)
Analyses
Correlated environmental filters We tested if field-measured edaphic variables remotely sensed macroclimatevariables andor remotely sensed fire attributes differedamong climatic zones (interior maritime) using one-wayANOVA Relationships of soil fertility fire frequency andintensity and recent burn history at McLaughlin describedabove are reported elsewhere (Safford and Harrison 2004)and thus new analyses were not conducted here
Correlated traits At the species level (n = 87) we testedwhether SLA differed by post-fire regeneration strategyusing one-way ANOVA and also repeated this analy-sis using a correction for phylogenetic non-independence(comparative analysis by independent contrasts (CAIC)details in Appendix 2 available online as before)
The percentage of obligate seeders in each plot wascalculated percentages were weighted by abundance todown-weight the relative influence of rare species on
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
Effects of climate soil and fire on specific leaf area 5
the description of functional structure The abundance-weighted percentage for each plot was calculated as fol-lows (sum abundance obligate seeders sum abundanceof all species) 100 We calculated community-level post-fire strategy with obligate seeders as the numerator becauseboth facultative resprouters and obligate resprouters areecologically similar in the sense that they can both resproutFor McLaughlin we tested if the percentage of obligateseeders differed by soil type burn history and their interac-tion (two-way ANOVA) Prior to analysis the percentage ofobligate seeders was arc-sine transformed For the CentralCoast dataset we tested if the percentage of seeders differedamong climatic zones using one-way ANOVA
Environmental correlates of SLA At the community levelwe tested if the mean abundance-weighted SLA observedin each plot at McLaughlin differed by soil type burn his-tory and their interaction (two-way ANOVA) For each plotthe mean abundance-weighted SLA was calculated by sum-ming the product of each speciesrsquo SLA and its proportionof total cover in that plot and then dividing by the numberof species in that plot We used the proportion of a speciescover in each plot as our metric of abundance rather thanabundance alone To examine for intraspecific variation inSLA by soil type we compared the SLA of sandstonecollections with the SLA of serpentine collections for the34 species collected from both soil types using a pairedt-test For the Central Coast we tested if abundance-weighted mean SLA differed among climatic zones usingone-way ANOVA
Environmental filtering of SLA To test for filtering of SLAthe range of SLA for each community was calculated(rangeobs) and compared with a null expectation of therange of SLA at the observed level of richness (rangeexp)(sensu Cornwell and Ackerly 2009) We randomly sam-pled the species pool by data set 9999 times for eachlevel of observed species richness among the plots andcalculated the mean range of SLA values for each level ofspecies richness Communities in which a filter is operatingwill have rangeexp gt rangeobs We tested if the distribu-tion of rangeexp rangeobs was significantly greater thanzero using a one-tailed t-test We then tested if rangeexp rangeobs in each plot differed by soil type burn history andclimatic zone using a two-way ANOVA All analyses wereperformed in R version 120 (R Development Core Team2010)
Results
Correlated environmental filters
Serpentine plots had a significantly lower CaMg ratiohigher organic matter content and lower P and K levelsthan the sandstone plots The less fertile maritime plots hada significantly lower CaMg ratio and lower P level than theinterior plots (Table 2) There were higher fuel levels andalso more frequent fires in interior plots than in maritime
plots (Table 2) For the macroclimate variables interiorplots were shown to be significantly more arid than mar-itime plots (lower relative humidity higher potential evap-otranspiration higher summer temperature lower wintertemperature and higher temperature seasonality) althoughprecipitation did not differ by zone (Supplementary Table 2in Appendix 1 available online)
Correlated traits
At the species level mean SLA was significantly higher inobligate resprouters than in either facultative resproutersor obligate seeders (one-way ANOVA F2 84 = 164P lt 0001) (Figure 2) This relationship was not signif-icant after correction for phylogenetic non-independence(P = 0327) (Appendix 2 available online) demonstratingshared variation among higher taxa
The abundance-weighted percentage of seeders wassignificantly lower on sandstone soils compared to serpen-tine soils (Figure 3a) but this soil contrast disappeared inrecently burned plots (two-way ANOVA soil F1 76 = 262P lt 0001 burn history F1 76 = 219 P lt 0001 soil burn history F176 = 294 P = 009 Figure 3a) For theCentral Coast plots there was a higher abundance-weightedpercentage of obligate seeders in the low-fire maritime cli-matic zone compared with the high-fire interior climaticzone (Table 2 Figure 3c) The Central Coast compared withMcLaughlin had a lesser proportion of obligate resprouters(040 vs 054) more obligate seeders (038 vs 027)and roughly the same proportion of facultative seeders(022 vs 019)
Environmental correlates of SLA
In the McLaughlin plots abundance-weighted mean SLAwas significantly higher on sandstone soils compared withserpentine soils (Figure 3b) regardless of burn history(two-way ANOVA soil F1 76 = 203 P lt 0001 burn his-tory F1 76 = 10 P = 031 soil burn history F1 76 = 03P = 059) Plant abundances were significantly lower onserpentine soils but this difference was only present amongthe unburned plots (Figure 3c two-way ANOVA soil F1 76
= 61 P = 0015 burn history F1 76 = 6905 P lt 0001soil burn history F1 76 = 49 P = 003) In the CentralCoast plots abundance-weighted mean SLA was not sig-nificantly different between climatic zones (Figure 3e) norwas abundance (Figure 3f)
Conspecifics had 18 higher mean SLA on sandstonethan serpentine soil (paired t-test t = 231 df = 33P = 0027) At the community level in contrast meanunweighted SLA was even greater with values in sand-stone plots 32 higher than in serpentine Thus variationin SLA among communities was due to both compositionalturnover and intraspecific variation (ie phenotypic plastic-ity) but species replacement played a larger role similarto previous results for Californian woody plants (Cornwelland Ackerly 2009)
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
6 B Anacker et al
Table 2 Environmental differences by soil type (McLaughlin plots sandstone serpentine) and climatic zone (Central Coast plotsinterior maritime)
McLaughlin Central Coast
Sandstone Serpentine F P Interior Maritime F P
EdaphicCaMg ratio 34 plusmn 22 05 plusmn 05 691 lt 0001 69 plusmn 16 45 plusmn 34 72 0011Bray P (ppm) 70 plusmn 26 34 plusmn 18 530 lt 0001 376 plusmn 236 144 plusmn 335 55 0025K (ppm) 2278 plusmn 628 1285 plusmn 713 440 lt 0001 2211 plusmn 767 1726 plusmn 886 30 0094Organic
matter ()43 plusmn 09 52 plusmn 15 95 0003 50 plusmn 38 65 plusmn 18 22 0152
FireFuel rank1 na na na na 21 plusmn 05 16 plusmn 07 66 0015Fire rotation na na na na 26 plusmn 06 15 plusmn 11 153 0001
Notes Mean plusmn one standard deviation P values are reported from one-way ANOVA of each variable with soil type or climatic zone Degrees of freedomfor McLaughlin are 1 and 78 for the factor and the residuals respectively Degrees of freedom for Central Coast are 1 and 33 for the factor and theresiduals respectively1Higher fuel ranks correspond to higher potential fire hazard
10
15
20
25
30
35
40
All species (n = 87) found in plots
Post-fire regeneration strategy
log(
SLA
mm
2 m
g1)
Obligateresprouters
Facultativeresprouters
Obligateseeders
a
b
b
Figure 2 Relationship of SLA and post-fire regeneration strat-egy among the 87 species considered here Letters above box-plots indicate significant differences based on a post-hoc TukeyrsquosHSD test Open circles indicate observations considered to beoutliers
Environmental filtering of SLA
The range of SLA values for the McLaughlin plots wassignificantly lower than expected under the null modelindicating environmental filtering (54 of 80 had rangeexp rangeobs lt 0 P = 0004) (Figures 4a b) Filtering strength(rangeexp rangeobs) was significantly higher on sandstonethan serpentine (one-way ANOVA soil F1 78 = 157 P =00002 Table 2) Filtering strength was also related to burnhistory and soil type burn history (two-way ANOVAsoil F1 76 = 184 P lt 0001 burn history F1 76 = 116P = 0001 soil burn history F1 76 = 42 P = 0044) Theinteraction term reflected that filtering was significant forunburned plots on both soils and for recently burned plotson serpentine but not recently burned plots on sandstoneThe range of SLA values for the Central Coast plots was not
significantly lower than expected (17 of 35 had rangeexp rangeobs lt 0 P = 0378) indicating a lack of filtering byclimatic zone (Table 2 Figure 4c)
Discussion
Among Californian chaparral communities varying in cli-mate soil fertility and fire histories we found that SLAa critical indicator of resource-use strategy did not followthe simple patterns anticipated from considering individ-ual filters Within the McLaughlin data SLA was lower oninfertile serpentine soils as expected but fire also played arole in broadening the range of SLA present on more fer-tile sandstone soils (relaxing filtering) Within the CentralCoast data SLA did not follow any simple expectationsbeing higher neither in more equable maritime climatesnor in the interior where soils are more fertile and fires aremore frequent These results are most consistent with thehypothesis that SLA is affected by an interaction of climatesoil fertility and fire Key to understanding this interac-tion are the correlations we found at both the species andcommunity levels between SLA and post-fire regenerationstrategies and also the environmental correlations betweenclimate soils and fire frequency
Obligate post-fire seeders had significantly lower SLAthan obligate post-fire resprouters (Figure 2) This contra-dicts the notion that the reduced need for storage of theseeding strategy should lead to relatively higher growthrates and associated higher SLA in seeders (Table 1) Thecorrelation we observed may reflect a physiological trade-off in which obligate seeding imposes strong selection fordrought tolerance at the seedling stage (Ackerly 2004)Infertile soils may also select for drought-tolerant seed-ers due to imposing arid conditions (high rock contenthigh exposure to direct sun high rates of soil evapora-tion) (Ojeda et al 2005 Pratt et al 2008) While seedershave low allocation to roots and shallow rooting depth andthus a limited ability to avoid drought by reaching water inlower soil horizons they may tolerate soil aridity if they
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
Effects of climate soil and fire on specific leaf area 7
020406080
100120140
McLaughlin
Soil and Burn historyasin
(abu
ndan
cew
eigh
ted
o
blig
ate
seed
ers)
B U B USandstone Serpentine
a
aa
bA
0001020304050607
McLaughlin
Soil and burn historyA
bund
ance
wei
ghte
d lo
g(S
LA)
B U B USandstone Serpentine
a a
b b
B
0
50
100
150
McLaughlin
Soil and burn history
Abu
ndan
ce
B U B USandstone Serpentine
a
b
a
c
C
020406080
100120140
Central Coast
Climatic zoneasin
(abu
ndan
cew
eigh
ted
o
blig
ate
seed
ers)
Interior Maritime
Climatic zone
Interior Maritime
Climatic zone
Interior Maritime
a
bD
0001020304050607
Central Coast
Abu
ndan
cew
eigh
ted
log(
SLA
)
a
a
E
0
50
100
150
Central Coast
Abu
ndan
ce
aaF
Figure 3 Plant community attributes for serpentine and sandstone soils (andashc) and interior and maritime climatic zones (dndashf) Recentfire history is indicated for the McLaughlin data as follows B recently burned U unburned Letters above boxplots indicate significantdifferences based on a post-hoc Tukeyrsquos HSD test Open circles indicate observations considered to be outliers
have leaf-level strategies such as low SLA that providean advantage under low-resource conditions (Poorter et al2009 Saura Mas et al 2009) Post-fire resprouters in con-trast avoid drought by having deeper rooting systems (Bellet al 1996 Bell 2001)
The correlation between post-fire regeneration strategyand SLA was not significant in an analysis of phyloge-netically independent contrasts reflecting the influence ofspecies-rich lineages (ie Arctostaphylos and CeanothusAppendices 1 and 2 available online) that share particu-lar trait combinations (eg post-fire seeding and low SLA)through common inheritance This means that the obser-vation of low SLA and obligate seeding in a communityis in part due to common descent In other words anenvironmental filter may be directly selecting for obligateseeding and thereby indirectly selecting for the corre-lated low SLA value or vice versa Similarly a study of37 plants of the Iberian peninsula showed phylogeneticnon-independence among post-fire regeneration strategyand fruit type (Pausas and Verdu 2005) Such patterns ofphylogenetic non-independence in traits are well knownand form the basis of phylogenetic clustering in communitystructure (Webb et al 2002 Anacker 2011)
The observed patterns of post-fire regeneration strategyconfirmed that fire was more frequent on sandstone rela-tive to serpentine (obligate seeders were more common inserpentine chaparral) and that fire was more frequent in
the interior than on the coast (obligate seeders more com-mon in maritime chaparral) In the arid interior the infertileconditions of serpentine soil limit fuel accumulation cre-ating local fire refugia and favouring seeders Along thecoast cool and wet climates limit the length and severityof the fire season and the flammability of fuels leadingto low regional fire frequencies favouring seeders lowsoil fertility is likely important in determining the balancebetween woodland and chaparral (Griffin 1978 Sawyeret al 2009) rather than affecting fire regime Anotherfactor favouring obligate seeding in maritime chaparralis that longer time intervals between fires may increasethe severity of fires when they do occur lowering adultsurvivorship
Chaparral on serpentine soils at McLaughlin had sig-nificantly lower SLA than chaparral on sandstone evenimmediately following a fire (Figure 3) suggesting thatsoils play the dominant role in shaping SLA in this settingHowever our observation that low-SLA obligate seedersare much more prevalent on infrequently burning serpen-tine stands (Figure 3) suggests that fire does interact withsoil to determine community-level SLA over longer timeperiods In our Central Coast data SLA did not differbetween maritime and interior plots even though mesicmaritime climates should promote high SLA near the coastwhile both frequent fire and more fertile soils are predictedto lead to higher SLA in the interior Abundant evidence
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
8 B Anacker et al
5 10 15
00
05
10
15
20
25
30
McLaughlin sandstone
Richness
Ran
ge o
f lo
g(S
LA)
burnedunburned
expectedA
5 10 15
00
05
10
15
20
25
30
McLaughlin serpentine
Richness
Ran
ge o
f lo
g(S
LA)
burnedunburned
B
5 10 15
00
05
10
15
20
25
30
Central Coast sandstone
Richness
Ran
ge o
f lo
g(S
LA)
maritimeinterior
C
Figure 4 The range of SLA observed in chaparral plots versusnull expectation Points below the line have lower observed rangesthan expected by chance
from other studies shows that SLA tends to be higher inproductive habitats (reviewed in Cornelissen et al 2003Westoby and Wright 2006 Poorter et al 2009) such as ourmaritime plots where water availability is high Thus oneof our more striking findings is that the globally predomi-nant role of climate in shaping SLA can be nullified by themore localised effects of soil fertility and fire if these gra-dients happen to vary in the opposite direction with respectto their effects on SLA
Conclusions
We found that community structure in Californianchaparral is shaped by the environmental filtering of a keyresource-acquisition trait ndash SLA ndash by the simultaneous
effects of climate soil fertility and fire Interactiveeffects of these three filters arose from two factors thephylogenetic and functional non-independence of SLA andanother critical trait (post-fire regeneration strategy) andthe non-independence of the three environmental filtersthemselves Our results provide a clear warning that incor-rect inferences about functional and phylogenetic commu-nity structure may be reached when either traits or filtersare considered in isolation
AcknowledgementsWe thank Naveed Davoodian for assistance in compiling relevantliterature and early discussions on the role of fire and substrateon reproductive strategies in the California flora and Tom Parkerand Tanner Harris for comments on an earlier version of thismanuscript
Notes on contributorsBrian Anacker is a post doctoral researcher at University ofCalifornia Davis His research addresses the historical and eco-logical controls of diversity in Californian plant communities
Nishanta Rajakaruna is a professor of botany at College of theAtlantic Bar Harbor ME USA His research focuses on thefactors and mechanisms driving plant speciation and communityassembly on serpentine and other edaphically harsh environments
David Ackerly is an associate professor at University ofCalifornia Berkeley His research addresses the ecological andevolutionary processes that shape the functional diversity ofterrestrial plants with a particular focus on the California flora
Susan Harrison is a professor at University of California DavisThe major theme of her research is understanding the processesthat shape and maintain plant species diversity at the landscapescale where small-scale forces such as competition and facilita-tion interact with large-scale forces such as niche evolution anddispersal
Jon Keeley is a research ecologist with the US Geological Surveyand adjunct professor at University of California Los AngelesHis research focuses on the ecological impacts and history ofwildfires in Mediterranean-climate ecosystems
Michael Vasey is a graduate student at University of CaliforniaSanta Cruz His research is focused on the importance of themarine layer (coastal fog) to the ecology and diversity of chaparralvegetation and manzanitas
ReferencesAckerly DD 2004 Functional strategies of chaparral shrubs in
relation to seasonal water deficit and disturbance EcologicalMonographs 7425ndash44
Ackerly D Knight C Weiss S Barton K Starmer K 2002Leaf size specific leaf area and microhabitat distribution ofchaparral woody plants contrasting patterns in species leveland community level analyses Oecologia 130449ndash457
Anacker BL 2011 Phylogenetic patterns of serpentine plantendemism and diversity In Harrison SP Rajakaruna Neditors Serpentine as a model in evolution and ecologyBerkeley California USA University of California Pressp 49ndash70
Bell DT 2001 Ecological response syndromes in the floraof southwestern Western Australia fire resprouters versusreseeders The Botanical Review 67417ndash440
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
Effects of climate soil and fire on specific leaf area 9
Bell TL Pate JS Dixon KW 1996 Relationships between fireresponse morphology root anatomy and starch distributionin south-west Australian Epacridaceae Annals of Botany77357
Bond WJ Midgley JJ 2001 The persistence niche ecology ofsprouting in woody plants Trends in Ecology and Evolution1645ndash51
Bond WJ Midgley JJ 2003 The evolutionary ecology of sprout-ing in woody plants International Journal of Plant Sciences164103ndash114
Burns K 2004 Patterns in specific leaf area and the structureof a temperate heath community Diversity and Distributions10105ndash112
Cavender-Bares J Kozak KH Fine PVA Kembel SW 2009 Themerging of community ecology and phylogenetic biologyEcology Letters 12693ndash715
Chew SJ Bonser SP 2009 The evolution of growth rate resourceallocation and competitive ability in seeder and resprouter treeseedlings Evolutionary Ecology 23723ndash735
Clarke PJ 2002a Habitat insularity and fire response traitsevidence from a sclerophyll archipelago Oecologia132582ndash591
Clarke PJ 2002b Habitat islands in fire-prone vegetation do land-scape features influence community composition Journal ofBiogeography 29677ndash684
Clarke PJ Knox KJE 2002 Post-fire response of shrubs inthe tablelands of eastern Australia do existing modelsexplain habitat differences Australian Journal of Botany 5053ndash62
Clarke PJ Knox KJE 2009 Trade offs in resource alloca-tion that favour resprouting affect the competitive ability ofwoody seedlings in grassy communities Journal of Ecology971374ndash1382
Clarke PJ Knox KJE Wills KE Campbell M 2005 Landscapepatterns of woody plant response to crown fire distur-bance and productivity influence sprouting ability Ecology93544ndash555
Cornelissen JHC Lavorel S Garnier E Diaz S Buchmann NGurvich DE Reich PB Steege H Morgan HD HeijdenMGA 2003 A handbook of protocols for standardisedand easy measurement of plant functional traits worldwideAustralian Journal of Botany 51335ndash380
Cornwell WK Ackerly DD 2009 Community assembly andshifts in plant trait distributions across an environmen-tal gradient in coastal California Ecological Monographs79109ndash126
Diamond JM 1975 Assembly of species communities In CodyML Diamond JM editors Ecology and evolution of com-munities Cambridge MA USA Harvard University Pressp 342ndash444
Diaz S Cabido M Casanoves F 1998 Plant functional traits andenvironmental filters at a regional scale Journal of VegetationScience 9113ndash122
Falster DS Westoby M 2005 Tradeoffs between height growthrate stem persistence and maximum height among plantspecies in a post fire succession Oikos 11157ndash66
Franklin J Syphard AD He HS Mladenoff DJ 2005 Altered fireregimes affect landscape patterns of plant succession in thefoothills and mountains of southern California Ecosystems8885ndash898
Griffin JR 1978 Maritime chaparral and endemic shrubs of theMonterey Bay region California Madrono 2565ndash81
Grime JP 1979 Plant strategies and vegetation processesChichester UK John Wiley amp Sons Ltd
Grubb PJ 1977 The maintenance of species-richness in plantcommunities the importance of the regeneration nicheBiological Reviews 52107ndash145
Hawkes CV Menges ES 1996 The relationship between openspace and fire for species in a xeric Florida shrubland Bulletinof the Torrey Botanical Club 12381ndash92
Hickman JC editor 1993 The Jepson Manual Higher plantsof California Berkeley California USA University ofCalifornia Press
Hoffmann WA Franco AC Moreira MZ Haridasan M 2005Specific leaf area explains differences in leaf traits betweencongeneric savanna and forest trees Functional Ecology19932ndash940
Hopper SD Brown AP Marchant NG 1997 Plants of WesternAustralian granite outcrops Journal of the Royal Society ofWestern Australia 80141ndash158
Keeley JE 1981 Reproductive cycles and fire regimes InProceedings of the Conference Fire Regimes and EcosystemProperties vol General Technical Report WO-26 USDAForest Service p 231ndash277
Keeley JE Fotheringham CJ Baer-Keeley M 2006 Demographicpatterns of postfire regeneration in Mediterranean-climate shrublands of California Ecological Monographs76235ndash255
Keeley JE Keeley SC 2000 Chaparral North American terres-trial vegetation 2204ndash253
Keeley JE Zedler PH 1978 Reproduction of chaparral shrubsafter fire a comparison of sprouting and seeding strategiesAmerican Midland Naturalist 99142ndash161
Knops JMH Reinhart K 2000 Specific leaf area along anitrogen fertilization gradient American Midland Naturalist265ndash272
Knox KJE Clarke PJ 2005 Nutrient availability induces contrast-ing allocation and starch formation in resprouting and obligateseeding shrubs Functional Ecology 19690ndash698
Lamont BB 1985 The comparative reproductive biology ofthree Leucospermum species (Proteaceae) in relation to fireresponses and breeding system Australian Journal of Botany33139ndash145
Lamont BB Markey A 1995 Biogeography of fire-killedand resprouting Banksia species in south-western AustraliaAustralian Journal of Botany 43283ndash303
Lamont BB Wiens D 2003 Are seed set and speciation ratesalways low among species that resprout after fire and whyEvolutionary Ecology 17277ndash292
Langley J Drake B Hungate B 2002 Extensive belowground car-bon storage supports roots and mycorrhizae in regeneratingscrub oaks Oecologia 131542ndash548
Lloret F Estevan H Vayreda J Terradas J 2005 Fire regenerativesyndromes of forest woody species across fire and climaticgradients Oecologia 146461ndash468
Lusk C Onoda Y Kooyman R Gutierrez Girun A 2010Reconciling species level vs plastic responses of evergreenleaf structure to light gradients shade leaves punch abovetheir weight New Phytologist 186429ndash438
McGill BJ Enquist BJ Weiher E Westoby M 2006 Rebuildingcommunity ecology from functional traits Trends in Ecologyamp Evolution 21178ndash185
Meinzer FC 2003 Functional convergence in plant responses tothe environment Oecologia 1341ndash11
Menges ES 2007 Integrating demography and fire managementan example from Florida scrub Australian Journal of Botany55261ndash272
Messier J McGill BJ Lechowicz MJ 2010 How do traits varyacross ecological scales A case for trait based ecologyEcology Letters 13838ndash848
Midgley JJ 1996 Why the worldrsquos vegetation is not totally dom-inated by resprouting plants because resprouters are shorterthan reseeders Ecography 1992ndash95
Niinemets U 2001 Global-scale climatic controls of leaf dry massper area density and thickness in trees and shrubs Ecology82453ndash469
Odion D Tyler C 2002 Are long fire-free periods needed tomaintain the endangered fire-recruiting shrub Arctostaphylosmorroensis (Ericaceae) Conservation Ecology 64 [online]URL httpwwwconsecolorgvol6iss2art4
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
10 B Anacker et al
Odion DC Davis FW 2000 Fire soil heating and the formationof vegetation patterns in chaparral Ecological Monographs70149ndash169
Ojeda F Brun FG Vergara JJ 2005 Fire rain and the selection ofseeder and resprouter life-histories in fire-recruiting woodyplants New Phytologist155ndash165
Pate JS Froend RH Bown BJ Hansen A Kuo J 1990 Seedlinggrowth and storage characteristics of seeder and resprouterspecies of Mediterranean-type ecosystems of SW AustraliaAnnals of Botany 65585
Paula S Pausas J 2006 Leaf traits and resprouting abil-ity in the Mediterranean basin Functional Ecology 20941ndash947
Pausas J Verdu M 2005 Plant persistence traits in fire-prone ecosystems of the Mediterranean basin a phylogeneticapproach Oikos 109196ndash202
Pausas JG 2001 Resprouting vs seeding ndash a Mediterraneanperspective Oikos 94193ndash194
Pausas JG Bradstock RA Keith DA Keeley JE 2004 Plantfunctional traits in relation to fire in crown-fire ecosystemsEcology 851085ndash1100
Pausas JG Keeley JE VerdUacute M 2006 Inferring differential evo-lutionary processes of plant persistence traits in NorthernHemisphere Mediterranean fire-prone ecosystems Ecology9431ndash39
Poorter H Niinemets Poorter L Wright IJ Villar R 2009Causes and consequences of variation in leaf mass perarea (LMA) a meta analysis New Phytologist 182565ndash588
Pratt RB Jacobsen AL Mohla R Ewers FW Davis SD 2008Linkage between water stress tolerance and life history typein seedlings of nine chaparral species (Rhamnaceae) Journalof Ecology 961252ndash1265
Price T 1997 Correlated evolution and independent contrastsTransactions of the Royal Society B Biological Sciences352519ndash529
Reich PB Walters MB Ellsworth DS 1997 From tropics to tun-dra global convergence in plant functioning Proceedings ofthe National Academy of Sciences of the United States ofAmerica 9413730ndash13734
Reich PB Wright IJ Cavender-Bares J Craine JM Oleksyn JWestoby M Walters MB 2003 The evolution of plant func-tional variation traits spectra and strategies InternationalJournal of Plant Sciences 164143ndash164
Russell-Smith J Whitehead P Cook G Hoare J 2003 Responseof Eucalyptus-dominated savanna to frequent fires lessonsfrom Munmarlary 1973ndash1996 Ecological Monographs73349ndash375
Safford HD Harrison SP 2004 Fire effects on plant diver-sity in serpentine vs sandstone chaparral Ecology 85539ndash548
Saura Mas S Shipley B Lloret F 2009 Relationship betweenpost fire regeneration and leaf economics spectrum inMediterranean woody species Functional Ecology 23103ndash110
Sawyer J Keeler-Wolf T Evans J 2009 A Manual ofCalifornia Vegetation Second Edition Sacramento CAUSA California Native Plant Society
Schmalzer PA 2003 Growth and recovery of oak-saw palmettoscrub through ten years after fire Natural Areas Journal235ndash13
Schwilk DW Ackerly DD 2005 Is there a cost to resproutingSeedling growth rate and drought tolerance in sprouting andnonsprouting Ceanothus (Rhamnaceae) 1 American Journalof Botany 92404ndash410
Shipley B Almeida-Cortez J 2003 Interspecific consistency andintraspecific variability of specific leaf area with respect toirradiance and nutrient availability Ecoscience 1074ndash79
Stearns S 1992 The evolution of life histories Oxford UKOxford University Press
Verduacute M 2000 Ecological and evolutionary differences betweenMediterranean seeders and resprouters Journal of VegetationScience 11265ndash268
Verduacute M Pausas JG 2007 Fire drives phylogenetic clusteringin Mediterranean Basin woody plant communities Ecology951316ndash1323
Vesk PA Westoby M 2004 Sprouting ability across diverse dis-turbances and vegetation types worldwide Journal of Ecology92310ndash320
Webb CO Ackerly DD McPeek MA Donoghue MJ 2002Phylogenies and community ecology Annual Review ofEcology and Systematics 33475ndash505
Weiher E Keddy PA 2001 Ecological assembly rules per-spectives advances retreats Cambridge UK CambridgeUniversity Press
Westerling AL Hidalgo HG Cayan DR Swetnam TW 2006Warming and earlier spring increase western US forest wild-fire activity Science 313940
Westoby M Falster DS Moles AT Vesk PA Wright IJ 2002Plant ecological strategies Some leading dimensions ofvariation between species Annual Review of Ecology andSystematics 33125ndash159
Westoby M Wright IJ 2006 Land-plant ecology on thebasis of functional traits Trends in Ecology amp Evolution21261ndash268
Willis CG Halina M Lehman C Reich PB Keen A McCarthyS Cavender-Bares J 2010 Phylogenetic community structurein Minnesota oak savanna is influenced by spatial extent andenvironmental variation Ecography 33565ndash577
Wisheu IC Rosenzweig ML Olsvig-Whittaker L Shmida A2000 What makes nutrient-poor Mediterranean heathlandsso rich in plant diversity Evolutionary Ecology Research2935ndash955
Wright IJ Reich PB Cornelissen JHC Falster DS GroomPK Hikosaka K Lee W Lusk CH Niinemets OleksynJ 2005 Modulation of leaf economic traits and trait rela-tionships by climate Global Ecology and Biogeography14411ndash422
Wright IJ Reich PB Westoby M Ackerly DD Baruch Z BongersF Cavender-Bares J Chapin T Cornelissen JHC DiemerM 2004 The worldwide leaf economics spectrum Nature428821ndash827
Wright IJ Westoby M Reich PB 2002 Convergence towardshigher leaf mass per area in dry and nutrient poor habitats hasdifferent consequences for leaf life span Journal of Ecology90534ndash543
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011

Effects of climate soil and fire on specific leaf area 5
the description of functional structure The abundance-weighted percentage for each plot was calculated as fol-lows (sum abundance obligate seeders sum abundanceof all species) 100 We calculated community-level post-fire strategy with obligate seeders as the numerator becauseboth facultative resprouters and obligate resprouters areecologically similar in the sense that they can both resproutFor McLaughlin we tested if the percentage of obligateseeders differed by soil type burn history and their interac-tion (two-way ANOVA) Prior to analysis the percentage ofobligate seeders was arc-sine transformed For the CentralCoast dataset we tested if the percentage of seeders differedamong climatic zones using one-way ANOVA
Environmental correlates of SLA At the community levelwe tested if the mean abundance-weighted SLA observedin each plot at McLaughlin differed by soil type burn his-tory and their interaction (two-way ANOVA) For each plotthe mean abundance-weighted SLA was calculated by sum-ming the product of each speciesrsquo SLA and its proportionof total cover in that plot and then dividing by the numberof species in that plot We used the proportion of a speciescover in each plot as our metric of abundance rather thanabundance alone To examine for intraspecific variation inSLA by soil type we compared the SLA of sandstonecollections with the SLA of serpentine collections for the34 species collected from both soil types using a pairedt-test For the Central Coast we tested if abundance-weighted mean SLA differed among climatic zones usingone-way ANOVA
Environmental filtering of SLA To test for filtering of SLAthe range of SLA for each community was calculated(rangeobs) and compared with a null expectation of therange of SLA at the observed level of richness (rangeexp)(sensu Cornwell and Ackerly 2009) We randomly sam-pled the species pool by data set 9999 times for eachlevel of observed species richness among the plots andcalculated the mean range of SLA values for each level ofspecies richness Communities in which a filter is operatingwill have rangeexp gt rangeobs We tested if the distribu-tion of rangeexp rangeobs was significantly greater thanzero using a one-tailed t-test We then tested if rangeexp rangeobs in each plot differed by soil type burn history andclimatic zone using a two-way ANOVA All analyses wereperformed in R version 120 (R Development Core Team2010)
Results
Correlated environmental filters
Serpentine plots had a significantly lower CaMg ratiohigher organic matter content and lower P and K levelsthan the sandstone plots The less fertile maritime plots hada significantly lower CaMg ratio and lower P level than theinterior plots (Table 2) There were higher fuel levels andalso more frequent fires in interior plots than in maritime
plots (Table 2) For the macroclimate variables interiorplots were shown to be significantly more arid than mar-itime plots (lower relative humidity higher potential evap-otranspiration higher summer temperature lower wintertemperature and higher temperature seasonality) althoughprecipitation did not differ by zone (Supplementary Table 2in Appendix 1 available online)
Correlated traits
At the species level mean SLA was significantly higher inobligate resprouters than in either facultative resproutersor obligate seeders (one-way ANOVA F2 84 = 164P lt 0001) (Figure 2) This relationship was not signif-icant after correction for phylogenetic non-independence(P = 0327) (Appendix 2 available online) demonstratingshared variation among higher taxa
The abundance-weighted percentage of seeders wassignificantly lower on sandstone soils compared to serpen-tine soils (Figure 3a) but this soil contrast disappeared inrecently burned plots (two-way ANOVA soil F1 76 = 262P lt 0001 burn history F1 76 = 219 P lt 0001 soil burn history F176 = 294 P = 009 Figure 3a) For theCentral Coast plots there was a higher abundance-weightedpercentage of obligate seeders in the low-fire maritime cli-matic zone compared with the high-fire interior climaticzone (Table 2 Figure 3c) The Central Coast compared withMcLaughlin had a lesser proportion of obligate resprouters(040 vs 054) more obligate seeders (038 vs 027)and roughly the same proportion of facultative seeders(022 vs 019)
Environmental correlates of SLA
In the McLaughlin plots abundance-weighted mean SLAwas significantly higher on sandstone soils compared withserpentine soils (Figure 3b) regardless of burn history(two-way ANOVA soil F1 76 = 203 P lt 0001 burn his-tory F1 76 = 10 P = 031 soil burn history F1 76 = 03P = 059) Plant abundances were significantly lower onserpentine soils but this difference was only present amongthe unburned plots (Figure 3c two-way ANOVA soil F1 76
= 61 P = 0015 burn history F1 76 = 6905 P lt 0001soil burn history F1 76 = 49 P = 003) In the CentralCoast plots abundance-weighted mean SLA was not sig-nificantly different between climatic zones (Figure 3e) norwas abundance (Figure 3f)
Conspecifics had 18 higher mean SLA on sandstonethan serpentine soil (paired t-test t = 231 df = 33P = 0027) At the community level in contrast meanunweighted SLA was even greater with values in sand-stone plots 32 higher than in serpentine Thus variationin SLA among communities was due to both compositionalturnover and intraspecific variation (ie phenotypic plastic-ity) but species replacement played a larger role similarto previous results for Californian woody plants (Cornwelland Ackerly 2009)
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
6 B Anacker et al
Table 2 Environmental differences by soil type (McLaughlin plots sandstone serpentine) and climatic zone (Central Coast plotsinterior maritime)
McLaughlin Central Coast
Sandstone Serpentine F P Interior Maritime F P
EdaphicCaMg ratio 34 plusmn 22 05 plusmn 05 691 lt 0001 69 plusmn 16 45 plusmn 34 72 0011Bray P (ppm) 70 plusmn 26 34 plusmn 18 530 lt 0001 376 plusmn 236 144 plusmn 335 55 0025K (ppm) 2278 plusmn 628 1285 plusmn 713 440 lt 0001 2211 plusmn 767 1726 plusmn 886 30 0094Organic
matter ()43 plusmn 09 52 plusmn 15 95 0003 50 plusmn 38 65 plusmn 18 22 0152
FireFuel rank1 na na na na 21 plusmn 05 16 plusmn 07 66 0015Fire rotation na na na na 26 plusmn 06 15 plusmn 11 153 0001
Notes Mean plusmn one standard deviation P values are reported from one-way ANOVA of each variable with soil type or climatic zone Degrees of freedomfor McLaughlin are 1 and 78 for the factor and the residuals respectively Degrees of freedom for Central Coast are 1 and 33 for the factor and theresiduals respectively1Higher fuel ranks correspond to higher potential fire hazard
10
15
20
25
30
35
40
All species (n = 87) found in plots
Post-fire regeneration strategy
log(
SLA
mm
2 m
g1)
Obligateresprouters
Facultativeresprouters
Obligateseeders
a
b
b
Figure 2 Relationship of SLA and post-fire regeneration strat-egy among the 87 species considered here Letters above box-plots indicate significant differences based on a post-hoc TukeyrsquosHSD test Open circles indicate observations considered to beoutliers
Environmental filtering of SLA
The range of SLA values for the McLaughlin plots wassignificantly lower than expected under the null modelindicating environmental filtering (54 of 80 had rangeexp rangeobs lt 0 P = 0004) (Figures 4a b) Filtering strength(rangeexp rangeobs) was significantly higher on sandstonethan serpentine (one-way ANOVA soil F1 78 = 157 P =00002 Table 2) Filtering strength was also related to burnhistory and soil type burn history (two-way ANOVAsoil F1 76 = 184 P lt 0001 burn history F1 76 = 116P = 0001 soil burn history F1 76 = 42 P = 0044) Theinteraction term reflected that filtering was significant forunburned plots on both soils and for recently burned plotson serpentine but not recently burned plots on sandstoneThe range of SLA values for the Central Coast plots was not
significantly lower than expected (17 of 35 had rangeexp rangeobs lt 0 P = 0378) indicating a lack of filtering byclimatic zone (Table 2 Figure 4c)
Discussion
Among Californian chaparral communities varying in cli-mate soil fertility and fire histories we found that SLAa critical indicator of resource-use strategy did not followthe simple patterns anticipated from considering individ-ual filters Within the McLaughlin data SLA was lower oninfertile serpentine soils as expected but fire also played arole in broadening the range of SLA present on more fer-tile sandstone soils (relaxing filtering) Within the CentralCoast data SLA did not follow any simple expectationsbeing higher neither in more equable maritime climatesnor in the interior where soils are more fertile and fires aremore frequent These results are most consistent with thehypothesis that SLA is affected by an interaction of climatesoil fertility and fire Key to understanding this interac-tion are the correlations we found at both the species andcommunity levels between SLA and post-fire regenerationstrategies and also the environmental correlations betweenclimate soils and fire frequency
Obligate post-fire seeders had significantly lower SLAthan obligate post-fire resprouters (Figure 2) This contra-dicts the notion that the reduced need for storage of theseeding strategy should lead to relatively higher growthrates and associated higher SLA in seeders (Table 1) Thecorrelation we observed may reflect a physiological trade-off in which obligate seeding imposes strong selection fordrought tolerance at the seedling stage (Ackerly 2004)Infertile soils may also select for drought-tolerant seed-ers due to imposing arid conditions (high rock contenthigh exposure to direct sun high rates of soil evapora-tion) (Ojeda et al 2005 Pratt et al 2008) While seedershave low allocation to roots and shallow rooting depth andthus a limited ability to avoid drought by reaching water inlower soil horizons they may tolerate soil aridity if they
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
Effects of climate soil and fire on specific leaf area 7
020406080
100120140
McLaughlin
Soil and Burn historyasin
(abu
ndan
cew
eigh
ted
o
blig
ate
seed
ers)
B U B USandstone Serpentine
a
aa
bA
0001020304050607
McLaughlin
Soil and burn historyA
bund
ance
wei
ghte
d lo
g(S
LA)
B U B USandstone Serpentine
a a
b b
B
0
50
100
150
McLaughlin
Soil and burn history
Abu
ndan
ce
B U B USandstone Serpentine
a
b
a
c
C
020406080
100120140
Central Coast
Climatic zoneasin
(abu
ndan
cew
eigh
ted
o
blig
ate
seed
ers)
Interior Maritime
Climatic zone
Interior Maritime
Climatic zone
Interior Maritime
a
bD
0001020304050607
Central Coast
Abu
ndan
cew
eigh
ted
log(
SLA
)
a
a
E
0
50
100
150
Central Coast
Abu
ndan
ce
aaF
Figure 3 Plant community attributes for serpentine and sandstone soils (andashc) and interior and maritime climatic zones (dndashf) Recentfire history is indicated for the McLaughlin data as follows B recently burned U unburned Letters above boxplots indicate significantdifferences based on a post-hoc Tukeyrsquos HSD test Open circles indicate observations considered to be outliers
have leaf-level strategies such as low SLA that providean advantage under low-resource conditions (Poorter et al2009 Saura Mas et al 2009) Post-fire resprouters in con-trast avoid drought by having deeper rooting systems (Bellet al 1996 Bell 2001)
The correlation between post-fire regeneration strategyand SLA was not significant in an analysis of phyloge-netically independent contrasts reflecting the influence ofspecies-rich lineages (ie Arctostaphylos and CeanothusAppendices 1 and 2 available online) that share particu-lar trait combinations (eg post-fire seeding and low SLA)through common inheritance This means that the obser-vation of low SLA and obligate seeding in a communityis in part due to common descent In other words anenvironmental filter may be directly selecting for obligateseeding and thereby indirectly selecting for the corre-lated low SLA value or vice versa Similarly a study of37 plants of the Iberian peninsula showed phylogeneticnon-independence among post-fire regeneration strategyand fruit type (Pausas and Verdu 2005) Such patterns ofphylogenetic non-independence in traits are well knownand form the basis of phylogenetic clustering in communitystructure (Webb et al 2002 Anacker 2011)
The observed patterns of post-fire regeneration strategyconfirmed that fire was more frequent on sandstone rela-tive to serpentine (obligate seeders were more common inserpentine chaparral) and that fire was more frequent in
the interior than on the coast (obligate seeders more com-mon in maritime chaparral) In the arid interior the infertileconditions of serpentine soil limit fuel accumulation cre-ating local fire refugia and favouring seeders Along thecoast cool and wet climates limit the length and severityof the fire season and the flammability of fuels leadingto low regional fire frequencies favouring seeders lowsoil fertility is likely important in determining the balancebetween woodland and chaparral (Griffin 1978 Sawyeret al 2009) rather than affecting fire regime Anotherfactor favouring obligate seeding in maritime chaparralis that longer time intervals between fires may increasethe severity of fires when they do occur lowering adultsurvivorship
Chaparral on serpentine soils at McLaughlin had sig-nificantly lower SLA than chaparral on sandstone evenimmediately following a fire (Figure 3) suggesting thatsoils play the dominant role in shaping SLA in this settingHowever our observation that low-SLA obligate seedersare much more prevalent on infrequently burning serpen-tine stands (Figure 3) suggests that fire does interact withsoil to determine community-level SLA over longer timeperiods In our Central Coast data SLA did not differbetween maritime and interior plots even though mesicmaritime climates should promote high SLA near the coastwhile both frequent fire and more fertile soils are predictedto lead to higher SLA in the interior Abundant evidence
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
8 B Anacker et al
5 10 15
00
05
10
15
20
25
30
McLaughlin sandstone
Richness
Ran
ge o
f lo
g(S
LA)
burnedunburned
expectedA
5 10 15
00
05
10
15
20
25
30
McLaughlin serpentine
Richness
Ran
ge o
f lo
g(S
LA)
burnedunburned
B
5 10 15
00
05
10
15
20
25
30
Central Coast sandstone
Richness
Ran
ge o
f lo
g(S
LA)
maritimeinterior
C
Figure 4 The range of SLA observed in chaparral plots versusnull expectation Points below the line have lower observed rangesthan expected by chance
from other studies shows that SLA tends to be higher inproductive habitats (reviewed in Cornelissen et al 2003Westoby and Wright 2006 Poorter et al 2009) such as ourmaritime plots where water availability is high Thus oneof our more striking findings is that the globally predomi-nant role of climate in shaping SLA can be nullified by themore localised effects of soil fertility and fire if these gra-dients happen to vary in the opposite direction with respectto their effects on SLA
Conclusions
We found that community structure in Californianchaparral is shaped by the environmental filtering of a keyresource-acquisition trait ndash SLA ndash by the simultaneous
effects of climate soil fertility and fire Interactiveeffects of these three filters arose from two factors thephylogenetic and functional non-independence of SLA andanother critical trait (post-fire regeneration strategy) andthe non-independence of the three environmental filtersthemselves Our results provide a clear warning that incor-rect inferences about functional and phylogenetic commu-nity structure may be reached when either traits or filtersare considered in isolation
AcknowledgementsWe thank Naveed Davoodian for assistance in compiling relevantliterature and early discussions on the role of fire and substrateon reproductive strategies in the California flora and Tom Parkerand Tanner Harris for comments on an earlier version of thismanuscript
Notes on contributorsBrian Anacker is a post doctoral researcher at University ofCalifornia Davis His research addresses the historical and eco-logical controls of diversity in Californian plant communities
Nishanta Rajakaruna is a professor of botany at College of theAtlantic Bar Harbor ME USA His research focuses on thefactors and mechanisms driving plant speciation and communityassembly on serpentine and other edaphically harsh environments
David Ackerly is an associate professor at University ofCalifornia Berkeley His research addresses the ecological andevolutionary processes that shape the functional diversity ofterrestrial plants with a particular focus on the California flora
Susan Harrison is a professor at University of California DavisThe major theme of her research is understanding the processesthat shape and maintain plant species diversity at the landscapescale where small-scale forces such as competition and facilita-tion interact with large-scale forces such as niche evolution anddispersal
Jon Keeley is a research ecologist with the US Geological Surveyand adjunct professor at University of California Los AngelesHis research focuses on the ecological impacts and history ofwildfires in Mediterranean-climate ecosystems
Michael Vasey is a graduate student at University of CaliforniaSanta Cruz His research is focused on the importance of themarine layer (coastal fog) to the ecology and diversity of chaparralvegetation and manzanitas
ReferencesAckerly DD 2004 Functional strategies of chaparral shrubs in
relation to seasonal water deficit and disturbance EcologicalMonographs 7425ndash44
Ackerly D Knight C Weiss S Barton K Starmer K 2002Leaf size specific leaf area and microhabitat distribution ofchaparral woody plants contrasting patterns in species leveland community level analyses Oecologia 130449ndash457
Anacker BL 2011 Phylogenetic patterns of serpentine plantendemism and diversity In Harrison SP Rajakaruna Neditors Serpentine as a model in evolution and ecologyBerkeley California USA University of California Pressp 49ndash70
Bell DT 2001 Ecological response syndromes in the floraof southwestern Western Australia fire resprouters versusreseeders The Botanical Review 67417ndash440
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
Effects of climate soil and fire on specific leaf area 9
Bell TL Pate JS Dixon KW 1996 Relationships between fireresponse morphology root anatomy and starch distributionin south-west Australian Epacridaceae Annals of Botany77357
Bond WJ Midgley JJ 2001 The persistence niche ecology ofsprouting in woody plants Trends in Ecology and Evolution1645ndash51
Bond WJ Midgley JJ 2003 The evolutionary ecology of sprout-ing in woody plants International Journal of Plant Sciences164103ndash114
Burns K 2004 Patterns in specific leaf area and the structureof a temperate heath community Diversity and Distributions10105ndash112
Cavender-Bares J Kozak KH Fine PVA Kembel SW 2009 Themerging of community ecology and phylogenetic biologyEcology Letters 12693ndash715
Chew SJ Bonser SP 2009 The evolution of growth rate resourceallocation and competitive ability in seeder and resprouter treeseedlings Evolutionary Ecology 23723ndash735
Clarke PJ 2002a Habitat insularity and fire response traitsevidence from a sclerophyll archipelago Oecologia132582ndash591
Clarke PJ 2002b Habitat islands in fire-prone vegetation do land-scape features influence community composition Journal ofBiogeography 29677ndash684
Clarke PJ Knox KJE 2002 Post-fire response of shrubs inthe tablelands of eastern Australia do existing modelsexplain habitat differences Australian Journal of Botany 5053ndash62
Clarke PJ Knox KJE 2009 Trade offs in resource alloca-tion that favour resprouting affect the competitive ability ofwoody seedlings in grassy communities Journal of Ecology971374ndash1382
Clarke PJ Knox KJE Wills KE Campbell M 2005 Landscapepatterns of woody plant response to crown fire distur-bance and productivity influence sprouting ability Ecology93544ndash555
Cornelissen JHC Lavorel S Garnier E Diaz S Buchmann NGurvich DE Reich PB Steege H Morgan HD HeijdenMGA 2003 A handbook of protocols for standardisedand easy measurement of plant functional traits worldwideAustralian Journal of Botany 51335ndash380
Cornwell WK Ackerly DD 2009 Community assembly andshifts in plant trait distributions across an environmen-tal gradient in coastal California Ecological Monographs79109ndash126
Diamond JM 1975 Assembly of species communities In CodyML Diamond JM editors Ecology and evolution of com-munities Cambridge MA USA Harvard University Pressp 342ndash444
Diaz S Cabido M Casanoves F 1998 Plant functional traits andenvironmental filters at a regional scale Journal of VegetationScience 9113ndash122
Falster DS Westoby M 2005 Tradeoffs between height growthrate stem persistence and maximum height among plantspecies in a post fire succession Oikos 11157ndash66
Franklin J Syphard AD He HS Mladenoff DJ 2005 Altered fireregimes affect landscape patterns of plant succession in thefoothills and mountains of southern California Ecosystems8885ndash898
Griffin JR 1978 Maritime chaparral and endemic shrubs of theMonterey Bay region California Madrono 2565ndash81
Grime JP 1979 Plant strategies and vegetation processesChichester UK John Wiley amp Sons Ltd
Grubb PJ 1977 The maintenance of species-richness in plantcommunities the importance of the regeneration nicheBiological Reviews 52107ndash145
Hawkes CV Menges ES 1996 The relationship between openspace and fire for species in a xeric Florida shrubland Bulletinof the Torrey Botanical Club 12381ndash92
Hickman JC editor 1993 The Jepson Manual Higher plantsof California Berkeley California USA University ofCalifornia Press
Hoffmann WA Franco AC Moreira MZ Haridasan M 2005Specific leaf area explains differences in leaf traits betweencongeneric savanna and forest trees Functional Ecology19932ndash940
Hopper SD Brown AP Marchant NG 1997 Plants of WesternAustralian granite outcrops Journal of the Royal Society ofWestern Australia 80141ndash158
Keeley JE 1981 Reproductive cycles and fire regimes InProceedings of the Conference Fire Regimes and EcosystemProperties vol General Technical Report WO-26 USDAForest Service p 231ndash277
Keeley JE Fotheringham CJ Baer-Keeley M 2006 Demographicpatterns of postfire regeneration in Mediterranean-climate shrublands of California Ecological Monographs76235ndash255
Keeley JE Keeley SC 2000 Chaparral North American terres-trial vegetation 2204ndash253
Keeley JE Zedler PH 1978 Reproduction of chaparral shrubsafter fire a comparison of sprouting and seeding strategiesAmerican Midland Naturalist 99142ndash161
Knops JMH Reinhart K 2000 Specific leaf area along anitrogen fertilization gradient American Midland Naturalist265ndash272
Knox KJE Clarke PJ 2005 Nutrient availability induces contrast-ing allocation and starch formation in resprouting and obligateseeding shrubs Functional Ecology 19690ndash698
Lamont BB 1985 The comparative reproductive biology ofthree Leucospermum species (Proteaceae) in relation to fireresponses and breeding system Australian Journal of Botany33139ndash145
Lamont BB Markey A 1995 Biogeography of fire-killedand resprouting Banksia species in south-western AustraliaAustralian Journal of Botany 43283ndash303
Lamont BB Wiens D 2003 Are seed set and speciation ratesalways low among species that resprout after fire and whyEvolutionary Ecology 17277ndash292
Langley J Drake B Hungate B 2002 Extensive belowground car-bon storage supports roots and mycorrhizae in regeneratingscrub oaks Oecologia 131542ndash548
Lloret F Estevan H Vayreda J Terradas J 2005 Fire regenerativesyndromes of forest woody species across fire and climaticgradients Oecologia 146461ndash468
Lusk C Onoda Y Kooyman R Gutierrez Girun A 2010Reconciling species level vs plastic responses of evergreenleaf structure to light gradients shade leaves punch abovetheir weight New Phytologist 186429ndash438
McGill BJ Enquist BJ Weiher E Westoby M 2006 Rebuildingcommunity ecology from functional traits Trends in Ecologyamp Evolution 21178ndash185
Meinzer FC 2003 Functional convergence in plant responses tothe environment Oecologia 1341ndash11
Menges ES 2007 Integrating demography and fire managementan example from Florida scrub Australian Journal of Botany55261ndash272
Messier J McGill BJ Lechowicz MJ 2010 How do traits varyacross ecological scales A case for trait based ecologyEcology Letters 13838ndash848
Midgley JJ 1996 Why the worldrsquos vegetation is not totally dom-inated by resprouting plants because resprouters are shorterthan reseeders Ecography 1992ndash95
Niinemets U 2001 Global-scale climatic controls of leaf dry massper area density and thickness in trees and shrubs Ecology82453ndash469
Odion D Tyler C 2002 Are long fire-free periods needed tomaintain the endangered fire-recruiting shrub Arctostaphylosmorroensis (Ericaceae) Conservation Ecology 64 [online]URL httpwwwconsecolorgvol6iss2art4
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
10 B Anacker et al
Odion DC Davis FW 2000 Fire soil heating and the formationof vegetation patterns in chaparral Ecological Monographs70149ndash169
Ojeda F Brun FG Vergara JJ 2005 Fire rain and the selection ofseeder and resprouter life-histories in fire-recruiting woodyplants New Phytologist155ndash165
Pate JS Froend RH Bown BJ Hansen A Kuo J 1990 Seedlinggrowth and storage characteristics of seeder and resprouterspecies of Mediterranean-type ecosystems of SW AustraliaAnnals of Botany 65585
Paula S Pausas J 2006 Leaf traits and resprouting abil-ity in the Mediterranean basin Functional Ecology 20941ndash947
Pausas J Verdu M 2005 Plant persistence traits in fire-prone ecosystems of the Mediterranean basin a phylogeneticapproach Oikos 109196ndash202
Pausas JG 2001 Resprouting vs seeding ndash a Mediterraneanperspective Oikos 94193ndash194
Pausas JG Bradstock RA Keith DA Keeley JE 2004 Plantfunctional traits in relation to fire in crown-fire ecosystemsEcology 851085ndash1100
Pausas JG Keeley JE VerdUacute M 2006 Inferring differential evo-lutionary processes of plant persistence traits in NorthernHemisphere Mediterranean fire-prone ecosystems Ecology9431ndash39
Poorter H Niinemets Poorter L Wright IJ Villar R 2009Causes and consequences of variation in leaf mass perarea (LMA) a meta analysis New Phytologist 182565ndash588
Pratt RB Jacobsen AL Mohla R Ewers FW Davis SD 2008Linkage between water stress tolerance and life history typein seedlings of nine chaparral species (Rhamnaceae) Journalof Ecology 961252ndash1265
Price T 1997 Correlated evolution and independent contrastsTransactions of the Royal Society B Biological Sciences352519ndash529
Reich PB Walters MB Ellsworth DS 1997 From tropics to tun-dra global convergence in plant functioning Proceedings ofthe National Academy of Sciences of the United States ofAmerica 9413730ndash13734
Reich PB Wright IJ Cavender-Bares J Craine JM Oleksyn JWestoby M Walters MB 2003 The evolution of plant func-tional variation traits spectra and strategies InternationalJournal of Plant Sciences 164143ndash164
Russell-Smith J Whitehead P Cook G Hoare J 2003 Responseof Eucalyptus-dominated savanna to frequent fires lessonsfrom Munmarlary 1973ndash1996 Ecological Monographs73349ndash375
Safford HD Harrison SP 2004 Fire effects on plant diver-sity in serpentine vs sandstone chaparral Ecology 85539ndash548
Saura Mas S Shipley B Lloret F 2009 Relationship betweenpost fire regeneration and leaf economics spectrum inMediterranean woody species Functional Ecology 23103ndash110
Sawyer J Keeler-Wolf T Evans J 2009 A Manual ofCalifornia Vegetation Second Edition Sacramento CAUSA California Native Plant Society
Schmalzer PA 2003 Growth and recovery of oak-saw palmettoscrub through ten years after fire Natural Areas Journal235ndash13
Schwilk DW Ackerly DD 2005 Is there a cost to resproutingSeedling growth rate and drought tolerance in sprouting andnonsprouting Ceanothus (Rhamnaceae) 1 American Journalof Botany 92404ndash410
Shipley B Almeida-Cortez J 2003 Interspecific consistency andintraspecific variability of specific leaf area with respect toirradiance and nutrient availability Ecoscience 1074ndash79
Stearns S 1992 The evolution of life histories Oxford UKOxford University Press
Verduacute M 2000 Ecological and evolutionary differences betweenMediterranean seeders and resprouters Journal of VegetationScience 11265ndash268
Verduacute M Pausas JG 2007 Fire drives phylogenetic clusteringin Mediterranean Basin woody plant communities Ecology951316ndash1323
Vesk PA Westoby M 2004 Sprouting ability across diverse dis-turbances and vegetation types worldwide Journal of Ecology92310ndash320
Webb CO Ackerly DD McPeek MA Donoghue MJ 2002Phylogenies and community ecology Annual Review ofEcology and Systematics 33475ndash505
Weiher E Keddy PA 2001 Ecological assembly rules per-spectives advances retreats Cambridge UK CambridgeUniversity Press
Westerling AL Hidalgo HG Cayan DR Swetnam TW 2006Warming and earlier spring increase western US forest wild-fire activity Science 313940
Westoby M Falster DS Moles AT Vesk PA Wright IJ 2002Plant ecological strategies Some leading dimensions ofvariation between species Annual Review of Ecology andSystematics 33125ndash159
Westoby M Wright IJ 2006 Land-plant ecology on thebasis of functional traits Trends in Ecology amp Evolution21261ndash268
Willis CG Halina M Lehman C Reich PB Keen A McCarthyS Cavender-Bares J 2010 Phylogenetic community structurein Minnesota oak savanna is influenced by spatial extent andenvironmental variation Ecography 33565ndash577
Wisheu IC Rosenzweig ML Olsvig-Whittaker L Shmida A2000 What makes nutrient-poor Mediterranean heathlandsso rich in plant diversity Evolutionary Ecology Research2935ndash955
Wright IJ Reich PB Cornelissen JHC Falster DS GroomPK Hikosaka K Lee W Lusk CH Niinemets OleksynJ 2005 Modulation of leaf economic traits and trait rela-tionships by climate Global Ecology and Biogeography14411ndash422
Wright IJ Reich PB Westoby M Ackerly DD Baruch Z BongersF Cavender-Bares J Chapin T Cornelissen JHC DiemerM 2004 The worldwide leaf economics spectrum Nature428821ndash827
Wright IJ Westoby M Reich PB 2002 Convergence towardshigher leaf mass per area in dry and nutrient poor habitats hasdifferent consequences for leaf life span Journal of Ecology90534ndash543
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011

6 B Anacker et al
Table 2 Environmental differences by soil type (McLaughlin plots sandstone serpentine) and climatic zone (Central Coast plotsinterior maritime)
McLaughlin Central Coast
Sandstone Serpentine F P Interior Maritime F P
EdaphicCaMg ratio 34 plusmn 22 05 plusmn 05 691 lt 0001 69 plusmn 16 45 plusmn 34 72 0011Bray P (ppm) 70 plusmn 26 34 plusmn 18 530 lt 0001 376 plusmn 236 144 plusmn 335 55 0025K (ppm) 2278 plusmn 628 1285 plusmn 713 440 lt 0001 2211 plusmn 767 1726 plusmn 886 30 0094Organic
matter ()43 plusmn 09 52 plusmn 15 95 0003 50 plusmn 38 65 plusmn 18 22 0152
FireFuel rank1 na na na na 21 plusmn 05 16 plusmn 07 66 0015Fire rotation na na na na 26 plusmn 06 15 plusmn 11 153 0001
Notes Mean plusmn one standard deviation P values are reported from one-way ANOVA of each variable with soil type or climatic zone Degrees of freedomfor McLaughlin are 1 and 78 for the factor and the residuals respectively Degrees of freedom for Central Coast are 1 and 33 for the factor and theresiduals respectively1Higher fuel ranks correspond to higher potential fire hazard
10
15
20
25
30
35
40
All species (n = 87) found in plots
Post-fire regeneration strategy
log(
SLA
mm
2 m
g1)
Obligateresprouters
Facultativeresprouters
Obligateseeders
a
b
b
Figure 2 Relationship of SLA and post-fire regeneration strat-egy among the 87 species considered here Letters above box-plots indicate significant differences based on a post-hoc TukeyrsquosHSD test Open circles indicate observations considered to beoutliers
Environmental filtering of SLA
The range of SLA values for the McLaughlin plots wassignificantly lower than expected under the null modelindicating environmental filtering (54 of 80 had rangeexp rangeobs lt 0 P = 0004) (Figures 4a b) Filtering strength(rangeexp rangeobs) was significantly higher on sandstonethan serpentine (one-way ANOVA soil F1 78 = 157 P =00002 Table 2) Filtering strength was also related to burnhistory and soil type burn history (two-way ANOVAsoil F1 76 = 184 P lt 0001 burn history F1 76 = 116P = 0001 soil burn history F1 76 = 42 P = 0044) Theinteraction term reflected that filtering was significant forunburned plots on both soils and for recently burned plotson serpentine but not recently burned plots on sandstoneThe range of SLA values for the Central Coast plots was not
significantly lower than expected (17 of 35 had rangeexp rangeobs lt 0 P = 0378) indicating a lack of filtering byclimatic zone (Table 2 Figure 4c)
Discussion
Among Californian chaparral communities varying in cli-mate soil fertility and fire histories we found that SLAa critical indicator of resource-use strategy did not followthe simple patterns anticipated from considering individ-ual filters Within the McLaughlin data SLA was lower oninfertile serpentine soils as expected but fire also played arole in broadening the range of SLA present on more fer-tile sandstone soils (relaxing filtering) Within the CentralCoast data SLA did not follow any simple expectationsbeing higher neither in more equable maritime climatesnor in the interior where soils are more fertile and fires aremore frequent These results are most consistent with thehypothesis that SLA is affected by an interaction of climatesoil fertility and fire Key to understanding this interac-tion are the correlations we found at both the species andcommunity levels between SLA and post-fire regenerationstrategies and also the environmental correlations betweenclimate soils and fire frequency
Obligate post-fire seeders had significantly lower SLAthan obligate post-fire resprouters (Figure 2) This contra-dicts the notion that the reduced need for storage of theseeding strategy should lead to relatively higher growthrates and associated higher SLA in seeders (Table 1) Thecorrelation we observed may reflect a physiological trade-off in which obligate seeding imposes strong selection fordrought tolerance at the seedling stage (Ackerly 2004)Infertile soils may also select for drought-tolerant seed-ers due to imposing arid conditions (high rock contenthigh exposure to direct sun high rates of soil evapora-tion) (Ojeda et al 2005 Pratt et al 2008) While seedershave low allocation to roots and shallow rooting depth andthus a limited ability to avoid drought by reaching water inlower soil horizons they may tolerate soil aridity if they
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
Effects of climate soil and fire on specific leaf area 7
020406080
100120140
McLaughlin
Soil and Burn historyasin
(abu
ndan
cew
eigh
ted
o
blig
ate
seed
ers)
B U B USandstone Serpentine
a
aa
bA
0001020304050607
McLaughlin
Soil and burn historyA
bund
ance
wei
ghte
d lo
g(S
LA)
B U B USandstone Serpentine
a a
b b
B
0
50
100
150
McLaughlin
Soil and burn history
Abu
ndan
ce
B U B USandstone Serpentine
a
b
a
c
C
020406080
100120140
Central Coast
Climatic zoneasin
(abu
ndan
cew
eigh
ted
o
blig
ate
seed
ers)
Interior Maritime
Climatic zone
Interior Maritime
Climatic zone
Interior Maritime
a
bD
0001020304050607
Central Coast
Abu
ndan
cew
eigh
ted
log(
SLA
)
a
a
E
0
50
100
150
Central Coast
Abu
ndan
ce
aaF
Figure 3 Plant community attributes for serpentine and sandstone soils (andashc) and interior and maritime climatic zones (dndashf) Recentfire history is indicated for the McLaughlin data as follows B recently burned U unburned Letters above boxplots indicate significantdifferences based on a post-hoc Tukeyrsquos HSD test Open circles indicate observations considered to be outliers
have leaf-level strategies such as low SLA that providean advantage under low-resource conditions (Poorter et al2009 Saura Mas et al 2009) Post-fire resprouters in con-trast avoid drought by having deeper rooting systems (Bellet al 1996 Bell 2001)
The correlation between post-fire regeneration strategyand SLA was not significant in an analysis of phyloge-netically independent contrasts reflecting the influence ofspecies-rich lineages (ie Arctostaphylos and CeanothusAppendices 1 and 2 available online) that share particu-lar trait combinations (eg post-fire seeding and low SLA)through common inheritance This means that the obser-vation of low SLA and obligate seeding in a communityis in part due to common descent In other words anenvironmental filter may be directly selecting for obligateseeding and thereby indirectly selecting for the corre-lated low SLA value or vice versa Similarly a study of37 plants of the Iberian peninsula showed phylogeneticnon-independence among post-fire regeneration strategyand fruit type (Pausas and Verdu 2005) Such patterns ofphylogenetic non-independence in traits are well knownand form the basis of phylogenetic clustering in communitystructure (Webb et al 2002 Anacker 2011)
The observed patterns of post-fire regeneration strategyconfirmed that fire was more frequent on sandstone rela-tive to serpentine (obligate seeders were more common inserpentine chaparral) and that fire was more frequent in
the interior than on the coast (obligate seeders more com-mon in maritime chaparral) In the arid interior the infertileconditions of serpentine soil limit fuel accumulation cre-ating local fire refugia and favouring seeders Along thecoast cool and wet climates limit the length and severityof the fire season and the flammability of fuels leadingto low regional fire frequencies favouring seeders lowsoil fertility is likely important in determining the balancebetween woodland and chaparral (Griffin 1978 Sawyeret al 2009) rather than affecting fire regime Anotherfactor favouring obligate seeding in maritime chaparralis that longer time intervals between fires may increasethe severity of fires when they do occur lowering adultsurvivorship
Chaparral on serpentine soils at McLaughlin had sig-nificantly lower SLA than chaparral on sandstone evenimmediately following a fire (Figure 3) suggesting thatsoils play the dominant role in shaping SLA in this settingHowever our observation that low-SLA obligate seedersare much more prevalent on infrequently burning serpen-tine stands (Figure 3) suggests that fire does interact withsoil to determine community-level SLA over longer timeperiods In our Central Coast data SLA did not differbetween maritime and interior plots even though mesicmaritime climates should promote high SLA near the coastwhile both frequent fire and more fertile soils are predictedto lead to higher SLA in the interior Abundant evidence
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
8 B Anacker et al
5 10 15
00
05
10
15
20
25
30
McLaughlin sandstone
Richness
Ran
ge o
f lo
g(S
LA)
burnedunburned
expectedA
5 10 15
00
05
10
15
20
25
30
McLaughlin serpentine
Richness
Ran
ge o
f lo
g(S
LA)
burnedunburned
B
5 10 15
00
05
10
15
20
25
30
Central Coast sandstone
Richness
Ran
ge o
f lo
g(S
LA)
maritimeinterior
C
Figure 4 The range of SLA observed in chaparral plots versusnull expectation Points below the line have lower observed rangesthan expected by chance
from other studies shows that SLA tends to be higher inproductive habitats (reviewed in Cornelissen et al 2003Westoby and Wright 2006 Poorter et al 2009) such as ourmaritime plots where water availability is high Thus oneof our more striking findings is that the globally predomi-nant role of climate in shaping SLA can be nullified by themore localised effects of soil fertility and fire if these gra-dients happen to vary in the opposite direction with respectto their effects on SLA
Conclusions
We found that community structure in Californianchaparral is shaped by the environmental filtering of a keyresource-acquisition trait ndash SLA ndash by the simultaneous
effects of climate soil fertility and fire Interactiveeffects of these three filters arose from two factors thephylogenetic and functional non-independence of SLA andanother critical trait (post-fire regeneration strategy) andthe non-independence of the three environmental filtersthemselves Our results provide a clear warning that incor-rect inferences about functional and phylogenetic commu-nity structure may be reached when either traits or filtersare considered in isolation
AcknowledgementsWe thank Naveed Davoodian for assistance in compiling relevantliterature and early discussions on the role of fire and substrateon reproductive strategies in the California flora and Tom Parkerand Tanner Harris for comments on an earlier version of thismanuscript
Notes on contributorsBrian Anacker is a post doctoral researcher at University ofCalifornia Davis His research addresses the historical and eco-logical controls of diversity in Californian plant communities
Nishanta Rajakaruna is a professor of botany at College of theAtlantic Bar Harbor ME USA His research focuses on thefactors and mechanisms driving plant speciation and communityassembly on serpentine and other edaphically harsh environments
David Ackerly is an associate professor at University ofCalifornia Berkeley His research addresses the ecological andevolutionary processes that shape the functional diversity ofterrestrial plants with a particular focus on the California flora
Susan Harrison is a professor at University of California DavisThe major theme of her research is understanding the processesthat shape and maintain plant species diversity at the landscapescale where small-scale forces such as competition and facilita-tion interact with large-scale forces such as niche evolution anddispersal
Jon Keeley is a research ecologist with the US Geological Surveyand adjunct professor at University of California Los AngelesHis research focuses on the ecological impacts and history ofwildfires in Mediterranean-climate ecosystems
Michael Vasey is a graduate student at University of CaliforniaSanta Cruz His research is focused on the importance of themarine layer (coastal fog) to the ecology and diversity of chaparralvegetation and manzanitas
ReferencesAckerly DD 2004 Functional strategies of chaparral shrubs in
relation to seasonal water deficit and disturbance EcologicalMonographs 7425ndash44
Ackerly D Knight C Weiss S Barton K Starmer K 2002Leaf size specific leaf area and microhabitat distribution ofchaparral woody plants contrasting patterns in species leveland community level analyses Oecologia 130449ndash457
Anacker BL 2011 Phylogenetic patterns of serpentine plantendemism and diversity In Harrison SP Rajakaruna Neditors Serpentine as a model in evolution and ecologyBerkeley California USA University of California Pressp 49ndash70
Bell DT 2001 Ecological response syndromes in the floraof southwestern Western Australia fire resprouters versusreseeders The Botanical Review 67417ndash440
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
Effects of climate soil and fire on specific leaf area 9
Bell TL Pate JS Dixon KW 1996 Relationships between fireresponse morphology root anatomy and starch distributionin south-west Australian Epacridaceae Annals of Botany77357
Bond WJ Midgley JJ 2001 The persistence niche ecology ofsprouting in woody plants Trends in Ecology and Evolution1645ndash51
Bond WJ Midgley JJ 2003 The evolutionary ecology of sprout-ing in woody plants International Journal of Plant Sciences164103ndash114
Burns K 2004 Patterns in specific leaf area and the structureof a temperate heath community Diversity and Distributions10105ndash112
Cavender-Bares J Kozak KH Fine PVA Kembel SW 2009 Themerging of community ecology and phylogenetic biologyEcology Letters 12693ndash715
Chew SJ Bonser SP 2009 The evolution of growth rate resourceallocation and competitive ability in seeder and resprouter treeseedlings Evolutionary Ecology 23723ndash735
Clarke PJ 2002a Habitat insularity and fire response traitsevidence from a sclerophyll archipelago Oecologia132582ndash591
Clarke PJ 2002b Habitat islands in fire-prone vegetation do land-scape features influence community composition Journal ofBiogeography 29677ndash684
Clarke PJ Knox KJE 2002 Post-fire response of shrubs inthe tablelands of eastern Australia do existing modelsexplain habitat differences Australian Journal of Botany 5053ndash62
Clarke PJ Knox KJE 2009 Trade offs in resource alloca-tion that favour resprouting affect the competitive ability ofwoody seedlings in grassy communities Journal of Ecology971374ndash1382
Clarke PJ Knox KJE Wills KE Campbell M 2005 Landscapepatterns of woody plant response to crown fire distur-bance and productivity influence sprouting ability Ecology93544ndash555
Cornelissen JHC Lavorel S Garnier E Diaz S Buchmann NGurvich DE Reich PB Steege H Morgan HD HeijdenMGA 2003 A handbook of protocols for standardisedand easy measurement of plant functional traits worldwideAustralian Journal of Botany 51335ndash380
Cornwell WK Ackerly DD 2009 Community assembly andshifts in plant trait distributions across an environmen-tal gradient in coastal California Ecological Monographs79109ndash126
Diamond JM 1975 Assembly of species communities In CodyML Diamond JM editors Ecology and evolution of com-munities Cambridge MA USA Harvard University Pressp 342ndash444
Diaz S Cabido M Casanoves F 1998 Plant functional traits andenvironmental filters at a regional scale Journal of VegetationScience 9113ndash122
Falster DS Westoby M 2005 Tradeoffs between height growthrate stem persistence and maximum height among plantspecies in a post fire succession Oikos 11157ndash66
Franklin J Syphard AD He HS Mladenoff DJ 2005 Altered fireregimes affect landscape patterns of plant succession in thefoothills and mountains of southern California Ecosystems8885ndash898
Griffin JR 1978 Maritime chaparral and endemic shrubs of theMonterey Bay region California Madrono 2565ndash81
Grime JP 1979 Plant strategies and vegetation processesChichester UK John Wiley amp Sons Ltd
Grubb PJ 1977 The maintenance of species-richness in plantcommunities the importance of the regeneration nicheBiological Reviews 52107ndash145
Hawkes CV Menges ES 1996 The relationship between openspace and fire for species in a xeric Florida shrubland Bulletinof the Torrey Botanical Club 12381ndash92
Hickman JC editor 1993 The Jepson Manual Higher plantsof California Berkeley California USA University ofCalifornia Press
Hoffmann WA Franco AC Moreira MZ Haridasan M 2005Specific leaf area explains differences in leaf traits betweencongeneric savanna and forest trees Functional Ecology19932ndash940
Hopper SD Brown AP Marchant NG 1997 Plants of WesternAustralian granite outcrops Journal of the Royal Society ofWestern Australia 80141ndash158
Keeley JE 1981 Reproductive cycles and fire regimes InProceedings of the Conference Fire Regimes and EcosystemProperties vol General Technical Report WO-26 USDAForest Service p 231ndash277
Keeley JE Fotheringham CJ Baer-Keeley M 2006 Demographicpatterns of postfire regeneration in Mediterranean-climate shrublands of California Ecological Monographs76235ndash255
Keeley JE Keeley SC 2000 Chaparral North American terres-trial vegetation 2204ndash253
Keeley JE Zedler PH 1978 Reproduction of chaparral shrubsafter fire a comparison of sprouting and seeding strategiesAmerican Midland Naturalist 99142ndash161
Knops JMH Reinhart K 2000 Specific leaf area along anitrogen fertilization gradient American Midland Naturalist265ndash272
Knox KJE Clarke PJ 2005 Nutrient availability induces contrast-ing allocation and starch formation in resprouting and obligateseeding shrubs Functional Ecology 19690ndash698
Lamont BB 1985 The comparative reproductive biology ofthree Leucospermum species (Proteaceae) in relation to fireresponses and breeding system Australian Journal of Botany33139ndash145
Lamont BB Markey A 1995 Biogeography of fire-killedand resprouting Banksia species in south-western AustraliaAustralian Journal of Botany 43283ndash303
Lamont BB Wiens D 2003 Are seed set and speciation ratesalways low among species that resprout after fire and whyEvolutionary Ecology 17277ndash292
Langley J Drake B Hungate B 2002 Extensive belowground car-bon storage supports roots and mycorrhizae in regeneratingscrub oaks Oecologia 131542ndash548
Lloret F Estevan H Vayreda J Terradas J 2005 Fire regenerativesyndromes of forest woody species across fire and climaticgradients Oecologia 146461ndash468
Lusk C Onoda Y Kooyman R Gutierrez Girun A 2010Reconciling species level vs plastic responses of evergreenleaf structure to light gradients shade leaves punch abovetheir weight New Phytologist 186429ndash438
McGill BJ Enquist BJ Weiher E Westoby M 2006 Rebuildingcommunity ecology from functional traits Trends in Ecologyamp Evolution 21178ndash185
Meinzer FC 2003 Functional convergence in plant responses tothe environment Oecologia 1341ndash11
Menges ES 2007 Integrating demography and fire managementan example from Florida scrub Australian Journal of Botany55261ndash272
Messier J McGill BJ Lechowicz MJ 2010 How do traits varyacross ecological scales A case for trait based ecologyEcology Letters 13838ndash848
Midgley JJ 1996 Why the worldrsquos vegetation is not totally dom-inated by resprouting plants because resprouters are shorterthan reseeders Ecography 1992ndash95
Niinemets U 2001 Global-scale climatic controls of leaf dry massper area density and thickness in trees and shrubs Ecology82453ndash469
Odion D Tyler C 2002 Are long fire-free periods needed tomaintain the endangered fire-recruiting shrub Arctostaphylosmorroensis (Ericaceae) Conservation Ecology 64 [online]URL httpwwwconsecolorgvol6iss2art4
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
10 B Anacker et al
Odion DC Davis FW 2000 Fire soil heating and the formationof vegetation patterns in chaparral Ecological Monographs70149ndash169
Ojeda F Brun FG Vergara JJ 2005 Fire rain and the selection ofseeder and resprouter life-histories in fire-recruiting woodyplants New Phytologist155ndash165
Pate JS Froend RH Bown BJ Hansen A Kuo J 1990 Seedlinggrowth and storage characteristics of seeder and resprouterspecies of Mediterranean-type ecosystems of SW AustraliaAnnals of Botany 65585
Paula S Pausas J 2006 Leaf traits and resprouting abil-ity in the Mediterranean basin Functional Ecology 20941ndash947
Pausas J Verdu M 2005 Plant persistence traits in fire-prone ecosystems of the Mediterranean basin a phylogeneticapproach Oikos 109196ndash202
Pausas JG 2001 Resprouting vs seeding ndash a Mediterraneanperspective Oikos 94193ndash194
Pausas JG Bradstock RA Keith DA Keeley JE 2004 Plantfunctional traits in relation to fire in crown-fire ecosystemsEcology 851085ndash1100
Pausas JG Keeley JE VerdUacute M 2006 Inferring differential evo-lutionary processes of plant persistence traits in NorthernHemisphere Mediterranean fire-prone ecosystems Ecology9431ndash39
Poorter H Niinemets Poorter L Wright IJ Villar R 2009Causes and consequences of variation in leaf mass perarea (LMA) a meta analysis New Phytologist 182565ndash588
Pratt RB Jacobsen AL Mohla R Ewers FW Davis SD 2008Linkage between water stress tolerance and life history typein seedlings of nine chaparral species (Rhamnaceae) Journalof Ecology 961252ndash1265
Price T 1997 Correlated evolution and independent contrastsTransactions of the Royal Society B Biological Sciences352519ndash529
Reich PB Walters MB Ellsworth DS 1997 From tropics to tun-dra global convergence in plant functioning Proceedings ofthe National Academy of Sciences of the United States ofAmerica 9413730ndash13734
Reich PB Wright IJ Cavender-Bares J Craine JM Oleksyn JWestoby M Walters MB 2003 The evolution of plant func-tional variation traits spectra and strategies InternationalJournal of Plant Sciences 164143ndash164
Russell-Smith J Whitehead P Cook G Hoare J 2003 Responseof Eucalyptus-dominated savanna to frequent fires lessonsfrom Munmarlary 1973ndash1996 Ecological Monographs73349ndash375
Safford HD Harrison SP 2004 Fire effects on plant diver-sity in serpentine vs sandstone chaparral Ecology 85539ndash548
Saura Mas S Shipley B Lloret F 2009 Relationship betweenpost fire regeneration and leaf economics spectrum inMediterranean woody species Functional Ecology 23103ndash110
Sawyer J Keeler-Wolf T Evans J 2009 A Manual ofCalifornia Vegetation Second Edition Sacramento CAUSA California Native Plant Society
Schmalzer PA 2003 Growth and recovery of oak-saw palmettoscrub through ten years after fire Natural Areas Journal235ndash13
Schwilk DW Ackerly DD 2005 Is there a cost to resproutingSeedling growth rate and drought tolerance in sprouting andnonsprouting Ceanothus (Rhamnaceae) 1 American Journalof Botany 92404ndash410
Shipley B Almeida-Cortez J 2003 Interspecific consistency andintraspecific variability of specific leaf area with respect toirradiance and nutrient availability Ecoscience 1074ndash79
Stearns S 1992 The evolution of life histories Oxford UKOxford University Press
Verduacute M 2000 Ecological and evolutionary differences betweenMediterranean seeders and resprouters Journal of VegetationScience 11265ndash268
Verduacute M Pausas JG 2007 Fire drives phylogenetic clusteringin Mediterranean Basin woody plant communities Ecology951316ndash1323
Vesk PA Westoby M 2004 Sprouting ability across diverse dis-turbances and vegetation types worldwide Journal of Ecology92310ndash320
Webb CO Ackerly DD McPeek MA Donoghue MJ 2002Phylogenies and community ecology Annual Review ofEcology and Systematics 33475ndash505
Weiher E Keddy PA 2001 Ecological assembly rules per-spectives advances retreats Cambridge UK CambridgeUniversity Press
Westerling AL Hidalgo HG Cayan DR Swetnam TW 2006Warming and earlier spring increase western US forest wild-fire activity Science 313940
Westoby M Falster DS Moles AT Vesk PA Wright IJ 2002Plant ecological strategies Some leading dimensions ofvariation between species Annual Review of Ecology andSystematics 33125ndash159
Westoby M Wright IJ 2006 Land-plant ecology on thebasis of functional traits Trends in Ecology amp Evolution21261ndash268
Willis CG Halina M Lehman C Reich PB Keen A McCarthyS Cavender-Bares J 2010 Phylogenetic community structurein Minnesota oak savanna is influenced by spatial extent andenvironmental variation Ecography 33565ndash577
Wisheu IC Rosenzweig ML Olsvig-Whittaker L Shmida A2000 What makes nutrient-poor Mediterranean heathlandsso rich in plant diversity Evolutionary Ecology Research2935ndash955
Wright IJ Reich PB Cornelissen JHC Falster DS GroomPK Hikosaka K Lee W Lusk CH Niinemets OleksynJ 2005 Modulation of leaf economic traits and trait rela-tionships by climate Global Ecology and Biogeography14411ndash422
Wright IJ Reich PB Westoby M Ackerly DD Baruch Z BongersF Cavender-Bares J Chapin T Cornelissen JHC DiemerM 2004 The worldwide leaf economics spectrum Nature428821ndash827
Wright IJ Westoby M Reich PB 2002 Convergence towardshigher leaf mass per area in dry and nutrient poor habitats hasdifferent consequences for leaf life span Journal of Ecology90534ndash543
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011

Effects of climate soil and fire on specific leaf area 7
020406080
100120140
McLaughlin
Soil and Burn historyasin
(abu
ndan
cew
eigh
ted
o
blig
ate
seed
ers)
B U B USandstone Serpentine
a
aa
bA
0001020304050607
McLaughlin
Soil and burn historyA
bund
ance
wei
ghte
d lo
g(S
LA)
B U B USandstone Serpentine
a a
b b
B
0
50
100
150
McLaughlin
Soil and burn history
Abu
ndan
ce
B U B USandstone Serpentine
a
b
a
c
C
020406080
100120140
Central Coast
Climatic zoneasin
(abu
ndan
cew
eigh
ted
o
blig
ate
seed
ers)
Interior Maritime
Climatic zone
Interior Maritime
Climatic zone
Interior Maritime
a
bD
0001020304050607
Central Coast
Abu
ndan
cew
eigh
ted
log(
SLA
)
a
a
E
0
50
100
150
Central Coast
Abu
ndan
ce
aaF
Figure 3 Plant community attributes for serpentine and sandstone soils (andashc) and interior and maritime climatic zones (dndashf) Recentfire history is indicated for the McLaughlin data as follows B recently burned U unburned Letters above boxplots indicate significantdifferences based on a post-hoc Tukeyrsquos HSD test Open circles indicate observations considered to be outliers
have leaf-level strategies such as low SLA that providean advantage under low-resource conditions (Poorter et al2009 Saura Mas et al 2009) Post-fire resprouters in con-trast avoid drought by having deeper rooting systems (Bellet al 1996 Bell 2001)
The correlation between post-fire regeneration strategyand SLA was not significant in an analysis of phyloge-netically independent contrasts reflecting the influence ofspecies-rich lineages (ie Arctostaphylos and CeanothusAppendices 1 and 2 available online) that share particu-lar trait combinations (eg post-fire seeding and low SLA)through common inheritance This means that the obser-vation of low SLA and obligate seeding in a communityis in part due to common descent In other words anenvironmental filter may be directly selecting for obligateseeding and thereby indirectly selecting for the corre-lated low SLA value or vice versa Similarly a study of37 plants of the Iberian peninsula showed phylogeneticnon-independence among post-fire regeneration strategyand fruit type (Pausas and Verdu 2005) Such patterns ofphylogenetic non-independence in traits are well knownand form the basis of phylogenetic clustering in communitystructure (Webb et al 2002 Anacker 2011)
The observed patterns of post-fire regeneration strategyconfirmed that fire was more frequent on sandstone rela-tive to serpentine (obligate seeders were more common inserpentine chaparral) and that fire was more frequent in
the interior than on the coast (obligate seeders more com-mon in maritime chaparral) In the arid interior the infertileconditions of serpentine soil limit fuel accumulation cre-ating local fire refugia and favouring seeders Along thecoast cool and wet climates limit the length and severityof the fire season and the flammability of fuels leadingto low regional fire frequencies favouring seeders lowsoil fertility is likely important in determining the balancebetween woodland and chaparral (Griffin 1978 Sawyeret al 2009) rather than affecting fire regime Anotherfactor favouring obligate seeding in maritime chaparralis that longer time intervals between fires may increasethe severity of fires when they do occur lowering adultsurvivorship
Chaparral on serpentine soils at McLaughlin had sig-nificantly lower SLA than chaparral on sandstone evenimmediately following a fire (Figure 3) suggesting thatsoils play the dominant role in shaping SLA in this settingHowever our observation that low-SLA obligate seedersare much more prevalent on infrequently burning serpen-tine stands (Figure 3) suggests that fire does interact withsoil to determine community-level SLA over longer timeperiods In our Central Coast data SLA did not differbetween maritime and interior plots even though mesicmaritime climates should promote high SLA near the coastwhile both frequent fire and more fertile soils are predictedto lead to higher SLA in the interior Abundant evidence
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
8 B Anacker et al
5 10 15
00
05
10
15
20
25
30
McLaughlin sandstone
Richness
Ran
ge o
f lo
g(S
LA)
burnedunburned
expectedA
5 10 15
00
05
10
15
20
25
30
McLaughlin serpentine
Richness
Ran
ge o
f lo
g(S
LA)
burnedunburned
B
5 10 15
00
05
10
15
20
25
30
Central Coast sandstone
Richness
Ran
ge o
f lo
g(S
LA)
maritimeinterior
C
Figure 4 The range of SLA observed in chaparral plots versusnull expectation Points below the line have lower observed rangesthan expected by chance
from other studies shows that SLA tends to be higher inproductive habitats (reviewed in Cornelissen et al 2003Westoby and Wright 2006 Poorter et al 2009) such as ourmaritime plots where water availability is high Thus oneof our more striking findings is that the globally predomi-nant role of climate in shaping SLA can be nullified by themore localised effects of soil fertility and fire if these gra-dients happen to vary in the opposite direction with respectto their effects on SLA
Conclusions
We found that community structure in Californianchaparral is shaped by the environmental filtering of a keyresource-acquisition trait ndash SLA ndash by the simultaneous
effects of climate soil fertility and fire Interactiveeffects of these three filters arose from two factors thephylogenetic and functional non-independence of SLA andanother critical trait (post-fire regeneration strategy) andthe non-independence of the three environmental filtersthemselves Our results provide a clear warning that incor-rect inferences about functional and phylogenetic commu-nity structure may be reached when either traits or filtersare considered in isolation
AcknowledgementsWe thank Naveed Davoodian for assistance in compiling relevantliterature and early discussions on the role of fire and substrateon reproductive strategies in the California flora and Tom Parkerand Tanner Harris for comments on an earlier version of thismanuscript
Notes on contributorsBrian Anacker is a post doctoral researcher at University ofCalifornia Davis His research addresses the historical and eco-logical controls of diversity in Californian plant communities
Nishanta Rajakaruna is a professor of botany at College of theAtlantic Bar Harbor ME USA His research focuses on thefactors and mechanisms driving plant speciation and communityassembly on serpentine and other edaphically harsh environments
David Ackerly is an associate professor at University ofCalifornia Berkeley His research addresses the ecological andevolutionary processes that shape the functional diversity ofterrestrial plants with a particular focus on the California flora
Susan Harrison is a professor at University of California DavisThe major theme of her research is understanding the processesthat shape and maintain plant species diversity at the landscapescale where small-scale forces such as competition and facilita-tion interact with large-scale forces such as niche evolution anddispersal
Jon Keeley is a research ecologist with the US Geological Surveyand adjunct professor at University of California Los AngelesHis research focuses on the ecological impacts and history ofwildfires in Mediterranean-climate ecosystems
Michael Vasey is a graduate student at University of CaliforniaSanta Cruz His research is focused on the importance of themarine layer (coastal fog) to the ecology and diversity of chaparralvegetation and manzanitas
ReferencesAckerly DD 2004 Functional strategies of chaparral shrubs in
relation to seasonal water deficit and disturbance EcologicalMonographs 7425ndash44
Ackerly D Knight C Weiss S Barton K Starmer K 2002Leaf size specific leaf area and microhabitat distribution ofchaparral woody plants contrasting patterns in species leveland community level analyses Oecologia 130449ndash457
Anacker BL 2011 Phylogenetic patterns of serpentine plantendemism and diversity In Harrison SP Rajakaruna Neditors Serpentine as a model in evolution and ecologyBerkeley California USA University of California Pressp 49ndash70
Bell DT 2001 Ecological response syndromes in the floraof southwestern Western Australia fire resprouters versusreseeders The Botanical Review 67417ndash440
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
Effects of climate soil and fire on specific leaf area 9
Bell TL Pate JS Dixon KW 1996 Relationships between fireresponse morphology root anatomy and starch distributionin south-west Australian Epacridaceae Annals of Botany77357
Bond WJ Midgley JJ 2001 The persistence niche ecology ofsprouting in woody plants Trends in Ecology and Evolution1645ndash51
Bond WJ Midgley JJ 2003 The evolutionary ecology of sprout-ing in woody plants International Journal of Plant Sciences164103ndash114
Burns K 2004 Patterns in specific leaf area and the structureof a temperate heath community Diversity and Distributions10105ndash112
Cavender-Bares J Kozak KH Fine PVA Kembel SW 2009 Themerging of community ecology and phylogenetic biologyEcology Letters 12693ndash715
Chew SJ Bonser SP 2009 The evolution of growth rate resourceallocation and competitive ability in seeder and resprouter treeseedlings Evolutionary Ecology 23723ndash735
Clarke PJ 2002a Habitat insularity and fire response traitsevidence from a sclerophyll archipelago Oecologia132582ndash591
Clarke PJ 2002b Habitat islands in fire-prone vegetation do land-scape features influence community composition Journal ofBiogeography 29677ndash684
Clarke PJ Knox KJE 2002 Post-fire response of shrubs inthe tablelands of eastern Australia do existing modelsexplain habitat differences Australian Journal of Botany 5053ndash62
Clarke PJ Knox KJE 2009 Trade offs in resource alloca-tion that favour resprouting affect the competitive ability ofwoody seedlings in grassy communities Journal of Ecology971374ndash1382
Clarke PJ Knox KJE Wills KE Campbell M 2005 Landscapepatterns of woody plant response to crown fire distur-bance and productivity influence sprouting ability Ecology93544ndash555
Cornelissen JHC Lavorel S Garnier E Diaz S Buchmann NGurvich DE Reich PB Steege H Morgan HD HeijdenMGA 2003 A handbook of protocols for standardisedand easy measurement of plant functional traits worldwideAustralian Journal of Botany 51335ndash380
Cornwell WK Ackerly DD 2009 Community assembly andshifts in plant trait distributions across an environmen-tal gradient in coastal California Ecological Monographs79109ndash126
Diamond JM 1975 Assembly of species communities In CodyML Diamond JM editors Ecology and evolution of com-munities Cambridge MA USA Harvard University Pressp 342ndash444
Diaz S Cabido M Casanoves F 1998 Plant functional traits andenvironmental filters at a regional scale Journal of VegetationScience 9113ndash122
Falster DS Westoby M 2005 Tradeoffs between height growthrate stem persistence and maximum height among plantspecies in a post fire succession Oikos 11157ndash66
Franklin J Syphard AD He HS Mladenoff DJ 2005 Altered fireregimes affect landscape patterns of plant succession in thefoothills and mountains of southern California Ecosystems8885ndash898
Griffin JR 1978 Maritime chaparral and endemic shrubs of theMonterey Bay region California Madrono 2565ndash81
Grime JP 1979 Plant strategies and vegetation processesChichester UK John Wiley amp Sons Ltd
Grubb PJ 1977 The maintenance of species-richness in plantcommunities the importance of the regeneration nicheBiological Reviews 52107ndash145
Hawkes CV Menges ES 1996 The relationship between openspace and fire for species in a xeric Florida shrubland Bulletinof the Torrey Botanical Club 12381ndash92
Hickman JC editor 1993 The Jepson Manual Higher plantsof California Berkeley California USA University ofCalifornia Press
Hoffmann WA Franco AC Moreira MZ Haridasan M 2005Specific leaf area explains differences in leaf traits betweencongeneric savanna and forest trees Functional Ecology19932ndash940
Hopper SD Brown AP Marchant NG 1997 Plants of WesternAustralian granite outcrops Journal of the Royal Society ofWestern Australia 80141ndash158
Keeley JE 1981 Reproductive cycles and fire regimes InProceedings of the Conference Fire Regimes and EcosystemProperties vol General Technical Report WO-26 USDAForest Service p 231ndash277
Keeley JE Fotheringham CJ Baer-Keeley M 2006 Demographicpatterns of postfire regeneration in Mediterranean-climate shrublands of California Ecological Monographs76235ndash255
Keeley JE Keeley SC 2000 Chaparral North American terres-trial vegetation 2204ndash253
Keeley JE Zedler PH 1978 Reproduction of chaparral shrubsafter fire a comparison of sprouting and seeding strategiesAmerican Midland Naturalist 99142ndash161
Knops JMH Reinhart K 2000 Specific leaf area along anitrogen fertilization gradient American Midland Naturalist265ndash272
Knox KJE Clarke PJ 2005 Nutrient availability induces contrast-ing allocation and starch formation in resprouting and obligateseeding shrubs Functional Ecology 19690ndash698
Lamont BB 1985 The comparative reproductive biology ofthree Leucospermum species (Proteaceae) in relation to fireresponses and breeding system Australian Journal of Botany33139ndash145
Lamont BB Markey A 1995 Biogeography of fire-killedand resprouting Banksia species in south-western AustraliaAustralian Journal of Botany 43283ndash303
Lamont BB Wiens D 2003 Are seed set and speciation ratesalways low among species that resprout after fire and whyEvolutionary Ecology 17277ndash292
Langley J Drake B Hungate B 2002 Extensive belowground car-bon storage supports roots and mycorrhizae in regeneratingscrub oaks Oecologia 131542ndash548
Lloret F Estevan H Vayreda J Terradas J 2005 Fire regenerativesyndromes of forest woody species across fire and climaticgradients Oecologia 146461ndash468
Lusk C Onoda Y Kooyman R Gutierrez Girun A 2010Reconciling species level vs plastic responses of evergreenleaf structure to light gradients shade leaves punch abovetheir weight New Phytologist 186429ndash438
McGill BJ Enquist BJ Weiher E Westoby M 2006 Rebuildingcommunity ecology from functional traits Trends in Ecologyamp Evolution 21178ndash185
Meinzer FC 2003 Functional convergence in plant responses tothe environment Oecologia 1341ndash11
Menges ES 2007 Integrating demography and fire managementan example from Florida scrub Australian Journal of Botany55261ndash272
Messier J McGill BJ Lechowicz MJ 2010 How do traits varyacross ecological scales A case for trait based ecologyEcology Letters 13838ndash848
Midgley JJ 1996 Why the worldrsquos vegetation is not totally dom-inated by resprouting plants because resprouters are shorterthan reseeders Ecography 1992ndash95
Niinemets U 2001 Global-scale climatic controls of leaf dry massper area density and thickness in trees and shrubs Ecology82453ndash469
Odion D Tyler C 2002 Are long fire-free periods needed tomaintain the endangered fire-recruiting shrub Arctostaphylosmorroensis (Ericaceae) Conservation Ecology 64 [online]URL httpwwwconsecolorgvol6iss2art4
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
10 B Anacker et al
Odion DC Davis FW 2000 Fire soil heating and the formationof vegetation patterns in chaparral Ecological Monographs70149ndash169
Ojeda F Brun FG Vergara JJ 2005 Fire rain and the selection ofseeder and resprouter life-histories in fire-recruiting woodyplants New Phytologist155ndash165
Pate JS Froend RH Bown BJ Hansen A Kuo J 1990 Seedlinggrowth and storage characteristics of seeder and resprouterspecies of Mediterranean-type ecosystems of SW AustraliaAnnals of Botany 65585
Paula S Pausas J 2006 Leaf traits and resprouting abil-ity in the Mediterranean basin Functional Ecology 20941ndash947
Pausas J Verdu M 2005 Plant persistence traits in fire-prone ecosystems of the Mediterranean basin a phylogeneticapproach Oikos 109196ndash202
Pausas JG 2001 Resprouting vs seeding ndash a Mediterraneanperspective Oikos 94193ndash194
Pausas JG Bradstock RA Keith DA Keeley JE 2004 Plantfunctional traits in relation to fire in crown-fire ecosystemsEcology 851085ndash1100
Pausas JG Keeley JE VerdUacute M 2006 Inferring differential evo-lutionary processes of plant persistence traits in NorthernHemisphere Mediterranean fire-prone ecosystems Ecology9431ndash39
Poorter H Niinemets Poorter L Wright IJ Villar R 2009Causes and consequences of variation in leaf mass perarea (LMA) a meta analysis New Phytologist 182565ndash588
Pratt RB Jacobsen AL Mohla R Ewers FW Davis SD 2008Linkage between water stress tolerance and life history typein seedlings of nine chaparral species (Rhamnaceae) Journalof Ecology 961252ndash1265
Price T 1997 Correlated evolution and independent contrastsTransactions of the Royal Society B Biological Sciences352519ndash529
Reich PB Walters MB Ellsworth DS 1997 From tropics to tun-dra global convergence in plant functioning Proceedings ofthe National Academy of Sciences of the United States ofAmerica 9413730ndash13734
Reich PB Wright IJ Cavender-Bares J Craine JM Oleksyn JWestoby M Walters MB 2003 The evolution of plant func-tional variation traits spectra and strategies InternationalJournal of Plant Sciences 164143ndash164
Russell-Smith J Whitehead P Cook G Hoare J 2003 Responseof Eucalyptus-dominated savanna to frequent fires lessonsfrom Munmarlary 1973ndash1996 Ecological Monographs73349ndash375
Safford HD Harrison SP 2004 Fire effects on plant diver-sity in serpentine vs sandstone chaparral Ecology 85539ndash548
Saura Mas S Shipley B Lloret F 2009 Relationship betweenpost fire regeneration and leaf economics spectrum inMediterranean woody species Functional Ecology 23103ndash110
Sawyer J Keeler-Wolf T Evans J 2009 A Manual ofCalifornia Vegetation Second Edition Sacramento CAUSA California Native Plant Society
Schmalzer PA 2003 Growth and recovery of oak-saw palmettoscrub through ten years after fire Natural Areas Journal235ndash13
Schwilk DW Ackerly DD 2005 Is there a cost to resproutingSeedling growth rate and drought tolerance in sprouting andnonsprouting Ceanothus (Rhamnaceae) 1 American Journalof Botany 92404ndash410
Shipley B Almeida-Cortez J 2003 Interspecific consistency andintraspecific variability of specific leaf area with respect toirradiance and nutrient availability Ecoscience 1074ndash79
Stearns S 1992 The evolution of life histories Oxford UKOxford University Press
Verduacute M 2000 Ecological and evolutionary differences betweenMediterranean seeders and resprouters Journal of VegetationScience 11265ndash268
Verduacute M Pausas JG 2007 Fire drives phylogenetic clusteringin Mediterranean Basin woody plant communities Ecology951316ndash1323
Vesk PA Westoby M 2004 Sprouting ability across diverse dis-turbances and vegetation types worldwide Journal of Ecology92310ndash320
Webb CO Ackerly DD McPeek MA Donoghue MJ 2002Phylogenies and community ecology Annual Review ofEcology and Systematics 33475ndash505
Weiher E Keddy PA 2001 Ecological assembly rules per-spectives advances retreats Cambridge UK CambridgeUniversity Press
Westerling AL Hidalgo HG Cayan DR Swetnam TW 2006Warming and earlier spring increase western US forest wild-fire activity Science 313940
Westoby M Falster DS Moles AT Vesk PA Wright IJ 2002Plant ecological strategies Some leading dimensions ofvariation between species Annual Review of Ecology andSystematics 33125ndash159
Westoby M Wright IJ 2006 Land-plant ecology on thebasis of functional traits Trends in Ecology amp Evolution21261ndash268
Willis CG Halina M Lehman C Reich PB Keen A McCarthyS Cavender-Bares J 2010 Phylogenetic community structurein Minnesota oak savanna is influenced by spatial extent andenvironmental variation Ecography 33565ndash577
Wisheu IC Rosenzweig ML Olsvig-Whittaker L Shmida A2000 What makes nutrient-poor Mediterranean heathlandsso rich in plant diversity Evolutionary Ecology Research2935ndash955
Wright IJ Reich PB Cornelissen JHC Falster DS GroomPK Hikosaka K Lee W Lusk CH Niinemets OleksynJ 2005 Modulation of leaf economic traits and trait rela-tionships by climate Global Ecology and Biogeography14411ndash422
Wright IJ Reich PB Westoby M Ackerly DD Baruch Z BongersF Cavender-Bares J Chapin T Cornelissen JHC DiemerM 2004 The worldwide leaf economics spectrum Nature428821ndash827
Wright IJ Westoby M Reich PB 2002 Convergence towardshigher leaf mass per area in dry and nutrient poor habitats hasdifferent consequences for leaf life span Journal of Ecology90534ndash543
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
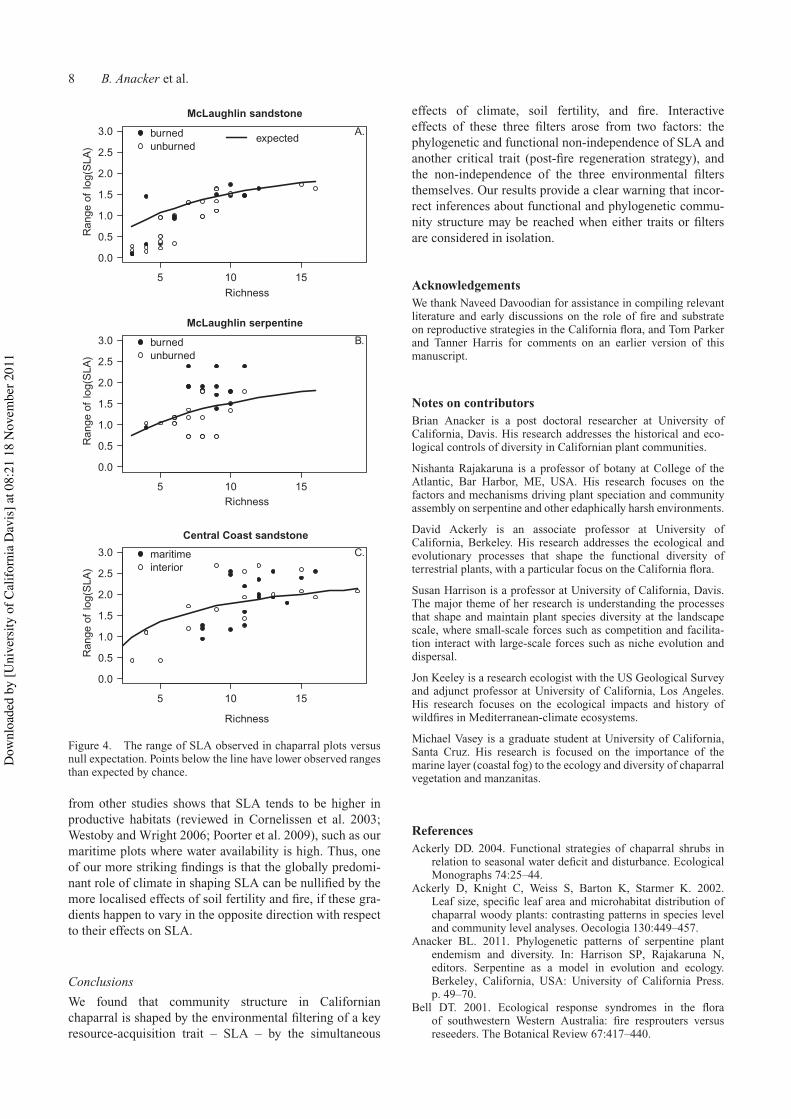
8 B Anacker et al
5 10 15
00
05
10
15
20
25
30
McLaughlin sandstone
Richness
Ran
ge o
f lo
g(S
LA)
burnedunburned
expectedA
5 10 15
00
05
10
15
20
25
30
McLaughlin serpentine
Richness
Ran
ge o
f lo
g(S
LA)
burnedunburned
B
5 10 15
00
05
10
15
20
25
30
Central Coast sandstone
Richness
Ran
ge o
f lo
g(S
LA)
maritimeinterior
C
Figure 4 The range of SLA observed in chaparral plots versusnull expectation Points below the line have lower observed rangesthan expected by chance
from other studies shows that SLA tends to be higher inproductive habitats (reviewed in Cornelissen et al 2003Westoby and Wright 2006 Poorter et al 2009) such as ourmaritime plots where water availability is high Thus oneof our more striking findings is that the globally predomi-nant role of climate in shaping SLA can be nullified by themore localised effects of soil fertility and fire if these gra-dients happen to vary in the opposite direction with respectto their effects on SLA
Conclusions
We found that community structure in Californianchaparral is shaped by the environmental filtering of a keyresource-acquisition trait ndash SLA ndash by the simultaneous
effects of climate soil fertility and fire Interactiveeffects of these three filters arose from two factors thephylogenetic and functional non-independence of SLA andanother critical trait (post-fire regeneration strategy) andthe non-independence of the three environmental filtersthemselves Our results provide a clear warning that incor-rect inferences about functional and phylogenetic commu-nity structure may be reached when either traits or filtersare considered in isolation
AcknowledgementsWe thank Naveed Davoodian for assistance in compiling relevantliterature and early discussions on the role of fire and substrateon reproductive strategies in the California flora and Tom Parkerand Tanner Harris for comments on an earlier version of thismanuscript
Notes on contributorsBrian Anacker is a post doctoral researcher at University ofCalifornia Davis His research addresses the historical and eco-logical controls of diversity in Californian plant communities
Nishanta Rajakaruna is a professor of botany at College of theAtlantic Bar Harbor ME USA His research focuses on thefactors and mechanisms driving plant speciation and communityassembly on serpentine and other edaphically harsh environments
David Ackerly is an associate professor at University ofCalifornia Berkeley His research addresses the ecological andevolutionary processes that shape the functional diversity ofterrestrial plants with a particular focus on the California flora
Susan Harrison is a professor at University of California DavisThe major theme of her research is understanding the processesthat shape and maintain plant species diversity at the landscapescale where small-scale forces such as competition and facilita-tion interact with large-scale forces such as niche evolution anddispersal
Jon Keeley is a research ecologist with the US Geological Surveyand adjunct professor at University of California Los AngelesHis research focuses on the ecological impacts and history ofwildfires in Mediterranean-climate ecosystems
Michael Vasey is a graduate student at University of CaliforniaSanta Cruz His research is focused on the importance of themarine layer (coastal fog) to the ecology and diversity of chaparralvegetation and manzanitas
ReferencesAckerly DD 2004 Functional strategies of chaparral shrubs in
relation to seasonal water deficit and disturbance EcologicalMonographs 7425ndash44
Ackerly D Knight C Weiss S Barton K Starmer K 2002Leaf size specific leaf area and microhabitat distribution ofchaparral woody plants contrasting patterns in species leveland community level analyses Oecologia 130449ndash457
Anacker BL 2011 Phylogenetic patterns of serpentine plantendemism and diversity In Harrison SP Rajakaruna Neditors Serpentine as a model in evolution and ecologyBerkeley California USA University of California Pressp 49ndash70
Bell DT 2001 Ecological response syndromes in the floraof southwestern Western Australia fire resprouters versusreseeders The Botanical Review 67417ndash440
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
Effects of climate soil and fire on specific leaf area 9
Bell TL Pate JS Dixon KW 1996 Relationships between fireresponse morphology root anatomy and starch distributionin south-west Australian Epacridaceae Annals of Botany77357
Bond WJ Midgley JJ 2001 The persistence niche ecology ofsprouting in woody plants Trends in Ecology and Evolution1645ndash51
Bond WJ Midgley JJ 2003 The evolutionary ecology of sprout-ing in woody plants International Journal of Plant Sciences164103ndash114
Burns K 2004 Patterns in specific leaf area and the structureof a temperate heath community Diversity and Distributions10105ndash112
Cavender-Bares J Kozak KH Fine PVA Kembel SW 2009 Themerging of community ecology and phylogenetic biologyEcology Letters 12693ndash715
Chew SJ Bonser SP 2009 The evolution of growth rate resourceallocation and competitive ability in seeder and resprouter treeseedlings Evolutionary Ecology 23723ndash735
Clarke PJ 2002a Habitat insularity and fire response traitsevidence from a sclerophyll archipelago Oecologia132582ndash591
Clarke PJ 2002b Habitat islands in fire-prone vegetation do land-scape features influence community composition Journal ofBiogeography 29677ndash684
Clarke PJ Knox KJE 2002 Post-fire response of shrubs inthe tablelands of eastern Australia do existing modelsexplain habitat differences Australian Journal of Botany 5053ndash62
Clarke PJ Knox KJE 2009 Trade offs in resource alloca-tion that favour resprouting affect the competitive ability ofwoody seedlings in grassy communities Journal of Ecology971374ndash1382
Clarke PJ Knox KJE Wills KE Campbell M 2005 Landscapepatterns of woody plant response to crown fire distur-bance and productivity influence sprouting ability Ecology93544ndash555
Cornelissen JHC Lavorel S Garnier E Diaz S Buchmann NGurvich DE Reich PB Steege H Morgan HD HeijdenMGA 2003 A handbook of protocols for standardisedand easy measurement of plant functional traits worldwideAustralian Journal of Botany 51335ndash380
Cornwell WK Ackerly DD 2009 Community assembly andshifts in plant trait distributions across an environmen-tal gradient in coastal California Ecological Monographs79109ndash126
Diamond JM 1975 Assembly of species communities In CodyML Diamond JM editors Ecology and evolution of com-munities Cambridge MA USA Harvard University Pressp 342ndash444
Diaz S Cabido M Casanoves F 1998 Plant functional traits andenvironmental filters at a regional scale Journal of VegetationScience 9113ndash122
Falster DS Westoby M 2005 Tradeoffs between height growthrate stem persistence and maximum height among plantspecies in a post fire succession Oikos 11157ndash66
Franklin J Syphard AD He HS Mladenoff DJ 2005 Altered fireregimes affect landscape patterns of plant succession in thefoothills and mountains of southern California Ecosystems8885ndash898
Griffin JR 1978 Maritime chaparral and endemic shrubs of theMonterey Bay region California Madrono 2565ndash81
Grime JP 1979 Plant strategies and vegetation processesChichester UK John Wiley amp Sons Ltd
Grubb PJ 1977 The maintenance of species-richness in plantcommunities the importance of the regeneration nicheBiological Reviews 52107ndash145
Hawkes CV Menges ES 1996 The relationship between openspace and fire for species in a xeric Florida shrubland Bulletinof the Torrey Botanical Club 12381ndash92
Hickman JC editor 1993 The Jepson Manual Higher plantsof California Berkeley California USA University ofCalifornia Press
Hoffmann WA Franco AC Moreira MZ Haridasan M 2005Specific leaf area explains differences in leaf traits betweencongeneric savanna and forest trees Functional Ecology19932ndash940
Hopper SD Brown AP Marchant NG 1997 Plants of WesternAustralian granite outcrops Journal of the Royal Society ofWestern Australia 80141ndash158
Keeley JE 1981 Reproductive cycles and fire regimes InProceedings of the Conference Fire Regimes and EcosystemProperties vol General Technical Report WO-26 USDAForest Service p 231ndash277
Keeley JE Fotheringham CJ Baer-Keeley M 2006 Demographicpatterns of postfire regeneration in Mediterranean-climate shrublands of California Ecological Monographs76235ndash255
Keeley JE Keeley SC 2000 Chaparral North American terres-trial vegetation 2204ndash253
Keeley JE Zedler PH 1978 Reproduction of chaparral shrubsafter fire a comparison of sprouting and seeding strategiesAmerican Midland Naturalist 99142ndash161
Knops JMH Reinhart K 2000 Specific leaf area along anitrogen fertilization gradient American Midland Naturalist265ndash272
Knox KJE Clarke PJ 2005 Nutrient availability induces contrast-ing allocation and starch formation in resprouting and obligateseeding shrubs Functional Ecology 19690ndash698
Lamont BB 1985 The comparative reproductive biology ofthree Leucospermum species (Proteaceae) in relation to fireresponses and breeding system Australian Journal of Botany33139ndash145
Lamont BB Markey A 1995 Biogeography of fire-killedand resprouting Banksia species in south-western AustraliaAustralian Journal of Botany 43283ndash303
Lamont BB Wiens D 2003 Are seed set and speciation ratesalways low among species that resprout after fire and whyEvolutionary Ecology 17277ndash292
Langley J Drake B Hungate B 2002 Extensive belowground car-bon storage supports roots and mycorrhizae in regeneratingscrub oaks Oecologia 131542ndash548
Lloret F Estevan H Vayreda J Terradas J 2005 Fire regenerativesyndromes of forest woody species across fire and climaticgradients Oecologia 146461ndash468
Lusk C Onoda Y Kooyman R Gutierrez Girun A 2010Reconciling species level vs plastic responses of evergreenleaf structure to light gradients shade leaves punch abovetheir weight New Phytologist 186429ndash438
McGill BJ Enquist BJ Weiher E Westoby M 2006 Rebuildingcommunity ecology from functional traits Trends in Ecologyamp Evolution 21178ndash185
Meinzer FC 2003 Functional convergence in plant responses tothe environment Oecologia 1341ndash11
Menges ES 2007 Integrating demography and fire managementan example from Florida scrub Australian Journal of Botany55261ndash272
Messier J McGill BJ Lechowicz MJ 2010 How do traits varyacross ecological scales A case for trait based ecologyEcology Letters 13838ndash848
Midgley JJ 1996 Why the worldrsquos vegetation is not totally dom-inated by resprouting plants because resprouters are shorterthan reseeders Ecography 1992ndash95
Niinemets U 2001 Global-scale climatic controls of leaf dry massper area density and thickness in trees and shrubs Ecology82453ndash469
Odion D Tyler C 2002 Are long fire-free periods needed tomaintain the endangered fire-recruiting shrub Arctostaphylosmorroensis (Ericaceae) Conservation Ecology 64 [online]URL httpwwwconsecolorgvol6iss2art4
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
10 B Anacker et al
Odion DC Davis FW 2000 Fire soil heating and the formationof vegetation patterns in chaparral Ecological Monographs70149ndash169
Ojeda F Brun FG Vergara JJ 2005 Fire rain and the selection ofseeder and resprouter life-histories in fire-recruiting woodyplants New Phytologist155ndash165
Pate JS Froend RH Bown BJ Hansen A Kuo J 1990 Seedlinggrowth and storage characteristics of seeder and resprouterspecies of Mediterranean-type ecosystems of SW AustraliaAnnals of Botany 65585
Paula S Pausas J 2006 Leaf traits and resprouting abil-ity in the Mediterranean basin Functional Ecology 20941ndash947
Pausas J Verdu M 2005 Plant persistence traits in fire-prone ecosystems of the Mediterranean basin a phylogeneticapproach Oikos 109196ndash202
Pausas JG 2001 Resprouting vs seeding ndash a Mediterraneanperspective Oikos 94193ndash194
Pausas JG Bradstock RA Keith DA Keeley JE 2004 Plantfunctional traits in relation to fire in crown-fire ecosystemsEcology 851085ndash1100
Pausas JG Keeley JE VerdUacute M 2006 Inferring differential evo-lutionary processes of plant persistence traits in NorthernHemisphere Mediterranean fire-prone ecosystems Ecology9431ndash39
Poorter H Niinemets Poorter L Wright IJ Villar R 2009Causes and consequences of variation in leaf mass perarea (LMA) a meta analysis New Phytologist 182565ndash588
Pratt RB Jacobsen AL Mohla R Ewers FW Davis SD 2008Linkage between water stress tolerance and life history typein seedlings of nine chaparral species (Rhamnaceae) Journalof Ecology 961252ndash1265
Price T 1997 Correlated evolution and independent contrastsTransactions of the Royal Society B Biological Sciences352519ndash529
Reich PB Walters MB Ellsworth DS 1997 From tropics to tun-dra global convergence in plant functioning Proceedings ofthe National Academy of Sciences of the United States ofAmerica 9413730ndash13734
Reich PB Wright IJ Cavender-Bares J Craine JM Oleksyn JWestoby M Walters MB 2003 The evolution of plant func-tional variation traits spectra and strategies InternationalJournal of Plant Sciences 164143ndash164
Russell-Smith J Whitehead P Cook G Hoare J 2003 Responseof Eucalyptus-dominated savanna to frequent fires lessonsfrom Munmarlary 1973ndash1996 Ecological Monographs73349ndash375
Safford HD Harrison SP 2004 Fire effects on plant diver-sity in serpentine vs sandstone chaparral Ecology 85539ndash548
Saura Mas S Shipley B Lloret F 2009 Relationship betweenpost fire regeneration and leaf economics spectrum inMediterranean woody species Functional Ecology 23103ndash110
Sawyer J Keeler-Wolf T Evans J 2009 A Manual ofCalifornia Vegetation Second Edition Sacramento CAUSA California Native Plant Society
Schmalzer PA 2003 Growth and recovery of oak-saw palmettoscrub through ten years after fire Natural Areas Journal235ndash13
Schwilk DW Ackerly DD 2005 Is there a cost to resproutingSeedling growth rate and drought tolerance in sprouting andnonsprouting Ceanothus (Rhamnaceae) 1 American Journalof Botany 92404ndash410
Shipley B Almeida-Cortez J 2003 Interspecific consistency andintraspecific variability of specific leaf area with respect toirradiance and nutrient availability Ecoscience 1074ndash79
Stearns S 1992 The evolution of life histories Oxford UKOxford University Press
Verduacute M 2000 Ecological and evolutionary differences betweenMediterranean seeders and resprouters Journal of VegetationScience 11265ndash268
Verduacute M Pausas JG 2007 Fire drives phylogenetic clusteringin Mediterranean Basin woody plant communities Ecology951316ndash1323
Vesk PA Westoby M 2004 Sprouting ability across diverse dis-turbances and vegetation types worldwide Journal of Ecology92310ndash320
Webb CO Ackerly DD McPeek MA Donoghue MJ 2002Phylogenies and community ecology Annual Review ofEcology and Systematics 33475ndash505
Weiher E Keddy PA 2001 Ecological assembly rules per-spectives advances retreats Cambridge UK CambridgeUniversity Press
Westerling AL Hidalgo HG Cayan DR Swetnam TW 2006Warming and earlier spring increase western US forest wild-fire activity Science 313940
Westoby M Falster DS Moles AT Vesk PA Wright IJ 2002Plant ecological strategies Some leading dimensions ofvariation between species Annual Review of Ecology andSystematics 33125ndash159
Westoby M Wright IJ 2006 Land-plant ecology on thebasis of functional traits Trends in Ecology amp Evolution21261ndash268
Willis CG Halina M Lehman C Reich PB Keen A McCarthyS Cavender-Bares J 2010 Phylogenetic community structurein Minnesota oak savanna is influenced by spatial extent andenvironmental variation Ecography 33565ndash577
Wisheu IC Rosenzweig ML Olsvig-Whittaker L Shmida A2000 What makes nutrient-poor Mediterranean heathlandsso rich in plant diversity Evolutionary Ecology Research2935ndash955
Wright IJ Reich PB Cornelissen JHC Falster DS GroomPK Hikosaka K Lee W Lusk CH Niinemets OleksynJ 2005 Modulation of leaf economic traits and trait rela-tionships by climate Global Ecology and Biogeography14411ndash422
Wright IJ Reich PB Westoby M Ackerly DD Baruch Z BongersF Cavender-Bares J Chapin T Cornelissen JHC DiemerM 2004 The worldwide leaf economics spectrum Nature428821ndash827
Wright IJ Westoby M Reich PB 2002 Convergence towardshigher leaf mass per area in dry and nutrient poor habitats hasdifferent consequences for leaf life span Journal of Ecology90534ndash543
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011

Effects of climate soil and fire on specific leaf area 9
Bell TL Pate JS Dixon KW 1996 Relationships between fireresponse morphology root anatomy and starch distributionin south-west Australian Epacridaceae Annals of Botany77357
Bond WJ Midgley JJ 2001 The persistence niche ecology ofsprouting in woody plants Trends in Ecology and Evolution1645ndash51
Bond WJ Midgley JJ 2003 The evolutionary ecology of sprout-ing in woody plants International Journal of Plant Sciences164103ndash114
Burns K 2004 Patterns in specific leaf area and the structureof a temperate heath community Diversity and Distributions10105ndash112
Cavender-Bares J Kozak KH Fine PVA Kembel SW 2009 Themerging of community ecology and phylogenetic biologyEcology Letters 12693ndash715
Chew SJ Bonser SP 2009 The evolution of growth rate resourceallocation and competitive ability in seeder and resprouter treeseedlings Evolutionary Ecology 23723ndash735
Clarke PJ 2002a Habitat insularity and fire response traitsevidence from a sclerophyll archipelago Oecologia132582ndash591
Clarke PJ 2002b Habitat islands in fire-prone vegetation do land-scape features influence community composition Journal ofBiogeography 29677ndash684
Clarke PJ Knox KJE 2002 Post-fire response of shrubs inthe tablelands of eastern Australia do existing modelsexplain habitat differences Australian Journal of Botany 5053ndash62
Clarke PJ Knox KJE 2009 Trade offs in resource alloca-tion that favour resprouting affect the competitive ability ofwoody seedlings in grassy communities Journal of Ecology971374ndash1382
Clarke PJ Knox KJE Wills KE Campbell M 2005 Landscapepatterns of woody plant response to crown fire distur-bance and productivity influence sprouting ability Ecology93544ndash555
Cornelissen JHC Lavorel S Garnier E Diaz S Buchmann NGurvich DE Reich PB Steege H Morgan HD HeijdenMGA 2003 A handbook of protocols for standardisedand easy measurement of plant functional traits worldwideAustralian Journal of Botany 51335ndash380
Cornwell WK Ackerly DD 2009 Community assembly andshifts in plant trait distributions across an environmen-tal gradient in coastal California Ecological Monographs79109ndash126
Diamond JM 1975 Assembly of species communities In CodyML Diamond JM editors Ecology and evolution of com-munities Cambridge MA USA Harvard University Pressp 342ndash444
Diaz S Cabido M Casanoves F 1998 Plant functional traits andenvironmental filters at a regional scale Journal of VegetationScience 9113ndash122
Falster DS Westoby M 2005 Tradeoffs between height growthrate stem persistence and maximum height among plantspecies in a post fire succession Oikos 11157ndash66
Franklin J Syphard AD He HS Mladenoff DJ 2005 Altered fireregimes affect landscape patterns of plant succession in thefoothills and mountains of southern California Ecosystems8885ndash898
Griffin JR 1978 Maritime chaparral and endemic shrubs of theMonterey Bay region California Madrono 2565ndash81
Grime JP 1979 Plant strategies and vegetation processesChichester UK John Wiley amp Sons Ltd
Grubb PJ 1977 The maintenance of species-richness in plantcommunities the importance of the regeneration nicheBiological Reviews 52107ndash145
Hawkes CV Menges ES 1996 The relationship between openspace and fire for species in a xeric Florida shrubland Bulletinof the Torrey Botanical Club 12381ndash92
Hickman JC editor 1993 The Jepson Manual Higher plantsof California Berkeley California USA University ofCalifornia Press
Hoffmann WA Franco AC Moreira MZ Haridasan M 2005Specific leaf area explains differences in leaf traits betweencongeneric savanna and forest trees Functional Ecology19932ndash940
Hopper SD Brown AP Marchant NG 1997 Plants of WesternAustralian granite outcrops Journal of the Royal Society ofWestern Australia 80141ndash158
Keeley JE 1981 Reproductive cycles and fire regimes InProceedings of the Conference Fire Regimes and EcosystemProperties vol General Technical Report WO-26 USDAForest Service p 231ndash277
Keeley JE Fotheringham CJ Baer-Keeley M 2006 Demographicpatterns of postfire regeneration in Mediterranean-climate shrublands of California Ecological Monographs76235ndash255
Keeley JE Keeley SC 2000 Chaparral North American terres-trial vegetation 2204ndash253
Keeley JE Zedler PH 1978 Reproduction of chaparral shrubsafter fire a comparison of sprouting and seeding strategiesAmerican Midland Naturalist 99142ndash161
Knops JMH Reinhart K 2000 Specific leaf area along anitrogen fertilization gradient American Midland Naturalist265ndash272
Knox KJE Clarke PJ 2005 Nutrient availability induces contrast-ing allocation and starch formation in resprouting and obligateseeding shrubs Functional Ecology 19690ndash698
Lamont BB 1985 The comparative reproductive biology ofthree Leucospermum species (Proteaceae) in relation to fireresponses and breeding system Australian Journal of Botany33139ndash145
Lamont BB Markey A 1995 Biogeography of fire-killedand resprouting Banksia species in south-western AustraliaAustralian Journal of Botany 43283ndash303
Lamont BB Wiens D 2003 Are seed set and speciation ratesalways low among species that resprout after fire and whyEvolutionary Ecology 17277ndash292
Langley J Drake B Hungate B 2002 Extensive belowground car-bon storage supports roots and mycorrhizae in regeneratingscrub oaks Oecologia 131542ndash548
Lloret F Estevan H Vayreda J Terradas J 2005 Fire regenerativesyndromes of forest woody species across fire and climaticgradients Oecologia 146461ndash468
Lusk C Onoda Y Kooyman R Gutierrez Girun A 2010Reconciling species level vs plastic responses of evergreenleaf structure to light gradients shade leaves punch abovetheir weight New Phytologist 186429ndash438
McGill BJ Enquist BJ Weiher E Westoby M 2006 Rebuildingcommunity ecology from functional traits Trends in Ecologyamp Evolution 21178ndash185
Meinzer FC 2003 Functional convergence in plant responses tothe environment Oecologia 1341ndash11
Menges ES 2007 Integrating demography and fire managementan example from Florida scrub Australian Journal of Botany55261ndash272
Messier J McGill BJ Lechowicz MJ 2010 How do traits varyacross ecological scales A case for trait based ecologyEcology Letters 13838ndash848
Midgley JJ 1996 Why the worldrsquos vegetation is not totally dom-inated by resprouting plants because resprouters are shorterthan reseeders Ecography 1992ndash95
Niinemets U 2001 Global-scale climatic controls of leaf dry massper area density and thickness in trees and shrubs Ecology82453ndash469
Odion D Tyler C 2002 Are long fire-free periods needed tomaintain the endangered fire-recruiting shrub Arctostaphylosmorroensis (Ericaceae) Conservation Ecology 64 [online]URL httpwwwconsecolorgvol6iss2art4
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011
10 B Anacker et al
Odion DC Davis FW 2000 Fire soil heating and the formationof vegetation patterns in chaparral Ecological Monographs70149ndash169
Ojeda F Brun FG Vergara JJ 2005 Fire rain and the selection ofseeder and resprouter life-histories in fire-recruiting woodyplants New Phytologist155ndash165
Pate JS Froend RH Bown BJ Hansen A Kuo J 1990 Seedlinggrowth and storage characteristics of seeder and resprouterspecies of Mediterranean-type ecosystems of SW AustraliaAnnals of Botany 65585
Paula S Pausas J 2006 Leaf traits and resprouting abil-ity in the Mediterranean basin Functional Ecology 20941ndash947
Pausas J Verdu M 2005 Plant persistence traits in fire-prone ecosystems of the Mediterranean basin a phylogeneticapproach Oikos 109196ndash202
Pausas JG 2001 Resprouting vs seeding ndash a Mediterraneanperspective Oikos 94193ndash194
Pausas JG Bradstock RA Keith DA Keeley JE 2004 Plantfunctional traits in relation to fire in crown-fire ecosystemsEcology 851085ndash1100
Pausas JG Keeley JE VerdUacute M 2006 Inferring differential evo-lutionary processes of plant persistence traits in NorthernHemisphere Mediterranean fire-prone ecosystems Ecology9431ndash39
Poorter H Niinemets Poorter L Wright IJ Villar R 2009Causes and consequences of variation in leaf mass perarea (LMA) a meta analysis New Phytologist 182565ndash588
Pratt RB Jacobsen AL Mohla R Ewers FW Davis SD 2008Linkage between water stress tolerance and life history typein seedlings of nine chaparral species (Rhamnaceae) Journalof Ecology 961252ndash1265
Price T 1997 Correlated evolution and independent contrastsTransactions of the Royal Society B Biological Sciences352519ndash529
Reich PB Walters MB Ellsworth DS 1997 From tropics to tun-dra global convergence in plant functioning Proceedings ofthe National Academy of Sciences of the United States ofAmerica 9413730ndash13734
Reich PB Wright IJ Cavender-Bares J Craine JM Oleksyn JWestoby M Walters MB 2003 The evolution of plant func-tional variation traits spectra and strategies InternationalJournal of Plant Sciences 164143ndash164
Russell-Smith J Whitehead P Cook G Hoare J 2003 Responseof Eucalyptus-dominated savanna to frequent fires lessonsfrom Munmarlary 1973ndash1996 Ecological Monographs73349ndash375
Safford HD Harrison SP 2004 Fire effects on plant diver-sity in serpentine vs sandstone chaparral Ecology 85539ndash548
Saura Mas S Shipley B Lloret F 2009 Relationship betweenpost fire regeneration and leaf economics spectrum inMediterranean woody species Functional Ecology 23103ndash110
Sawyer J Keeler-Wolf T Evans J 2009 A Manual ofCalifornia Vegetation Second Edition Sacramento CAUSA California Native Plant Society
Schmalzer PA 2003 Growth and recovery of oak-saw palmettoscrub through ten years after fire Natural Areas Journal235ndash13
Schwilk DW Ackerly DD 2005 Is there a cost to resproutingSeedling growth rate and drought tolerance in sprouting andnonsprouting Ceanothus (Rhamnaceae) 1 American Journalof Botany 92404ndash410
Shipley B Almeida-Cortez J 2003 Interspecific consistency andintraspecific variability of specific leaf area with respect toirradiance and nutrient availability Ecoscience 1074ndash79
Stearns S 1992 The evolution of life histories Oxford UKOxford University Press
Verduacute M 2000 Ecological and evolutionary differences betweenMediterranean seeders and resprouters Journal of VegetationScience 11265ndash268
Verduacute M Pausas JG 2007 Fire drives phylogenetic clusteringin Mediterranean Basin woody plant communities Ecology951316ndash1323
Vesk PA Westoby M 2004 Sprouting ability across diverse dis-turbances and vegetation types worldwide Journal of Ecology92310ndash320
Webb CO Ackerly DD McPeek MA Donoghue MJ 2002Phylogenies and community ecology Annual Review ofEcology and Systematics 33475ndash505
Weiher E Keddy PA 2001 Ecological assembly rules per-spectives advances retreats Cambridge UK CambridgeUniversity Press
Westerling AL Hidalgo HG Cayan DR Swetnam TW 2006Warming and earlier spring increase western US forest wild-fire activity Science 313940
Westoby M Falster DS Moles AT Vesk PA Wright IJ 2002Plant ecological strategies Some leading dimensions ofvariation between species Annual Review of Ecology andSystematics 33125ndash159
Westoby M Wright IJ 2006 Land-plant ecology on thebasis of functional traits Trends in Ecology amp Evolution21261ndash268
Willis CG Halina M Lehman C Reich PB Keen A McCarthyS Cavender-Bares J 2010 Phylogenetic community structurein Minnesota oak savanna is influenced by spatial extent andenvironmental variation Ecography 33565ndash577
Wisheu IC Rosenzweig ML Olsvig-Whittaker L Shmida A2000 What makes nutrient-poor Mediterranean heathlandsso rich in plant diversity Evolutionary Ecology Research2935ndash955
Wright IJ Reich PB Cornelissen JHC Falster DS GroomPK Hikosaka K Lee W Lusk CH Niinemets OleksynJ 2005 Modulation of leaf economic traits and trait rela-tionships by climate Global Ecology and Biogeography14411ndash422
Wright IJ Reich PB Westoby M Ackerly DD Baruch Z BongersF Cavender-Bares J Chapin T Cornelissen JHC DiemerM 2004 The worldwide leaf economics spectrum Nature428821ndash827
Wright IJ Westoby M Reich PB 2002 Convergence towardshigher leaf mass per area in dry and nutrient poor habitats hasdifferent consequences for leaf life span Journal of Ecology90534ndash543
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011

10 B Anacker et al
Odion DC Davis FW 2000 Fire soil heating and the formationof vegetation patterns in chaparral Ecological Monographs70149ndash169
Ojeda F Brun FG Vergara JJ 2005 Fire rain and the selection ofseeder and resprouter life-histories in fire-recruiting woodyplants New Phytologist155ndash165
Pate JS Froend RH Bown BJ Hansen A Kuo J 1990 Seedlinggrowth and storage characteristics of seeder and resprouterspecies of Mediterranean-type ecosystems of SW AustraliaAnnals of Botany 65585
Paula S Pausas J 2006 Leaf traits and resprouting abil-ity in the Mediterranean basin Functional Ecology 20941ndash947
Pausas J Verdu M 2005 Plant persistence traits in fire-prone ecosystems of the Mediterranean basin a phylogeneticapproach Oikos 109196ndash202
Pausas JG 2001 Resprouting vs seeding ndash a Mediterraneanperspective Oikos 94193ndash194
Pausas JG Bradstock RA Keith DA Keeley JE 2004 Plantfunctional traits in relation to fire in crown-fire ecosystemsEcology 851085ndash1100
Pausas JG Keeley JE VerdUacute M 2006 Inferring differential evo-lutionary processes of plant persistence traits in NorthernHemisphere Mediterranean fire-prone ecosystems Ecology9431ndash39
Poorter H Niinemets Poorter L Wright IJ Villar R 2009Causes and consequences of variation in leaf mass perarea (LMA) a meta analysis New Phytologist 182565ndash588
Pratt RB Jacobsen AL Mohla R Ewers FW Davis SD 2008Linkage between water stress tolerance and life history typein seedlings of nine chaparral species (Rhamnaceae) Journalof Ecology 961252ndash1265
Price T 1997 Correlated evolution and independent contrastsTransactions of the Royal Society B Biological Sciences352519ndash529
Reich PB Walters MB Ellsworth DS 1997 From tropics to tun-dra global convergence in plant functioning Proceedings ofthe National Academy of Sciences of the United States ofAmerica 9413730ndash13734
Reich PB Wright IJ Cavender-Bares J Craine JM Oleksyn JWestoby M Walters MB 2003 The evolution of plant func-tional variation traits spectra and strategies InternationalJournal of Plant Sciences 164143ndash164
Russell-Smith J Whitehead P Cook G Hoare J 2003 Responseof Eucalyptus-dominated savanna to frequent fires lessonsfrom Munmarlary 1973ndash1996 Ecological Monographs73349ndash375
Safford HD Harrison SP 2004 Fire effects on plant diver-sity in serpentine vs sandstone chaparral Ecology 85539ndash548
Saura Mas S Shipley B Lloret F 2009 Relationship betweenpost fire regeneration and leaf economics spectrum inMediterranean woody species Functional Ecology 23103ndash110
Sawyer J Keeler-Wolf T Evans J 2009 A Manual ofCalifornia Vegetation Second Edition Sacramento CAUSA California Native Plant Society
Schmalzer PA 2003 Growth and recovery of oak-saw palmettoscrub through ten years after fire Natural Areas Journal235ndash13
Schwilk DW Ackerly DD 2005 Is there a cost to resproutingSeedling growth rate and drought tolerance in sprouting andnonsprouting Ceanothus (Rhamnaceae) 1 American Journalof Botany 92404ndash410
Shipley B Almeida-Cortez J 2003 Interspecific consistency andintraspecific variability of specific leaf area with respect toirradiance and nutrient availability Ecoscience 1074ndash79
Stearns S 1992 The evolution of life histories Oxford UKOxford University Press
Verduacute M 2000 Ecological and evolutionary differences betweenMediterranean seeders and resprouters Journal of VegetationScience 11265ndash268
Verduacute M Pausas JG 2007 Fire drives phylogenetic clusteringin Mediterranean Basin woody plant communities Ecology951316ndash1323
Vesk PA Westoby M 2004 Sprouting ability across diverse dis-turbances and vegetation types worldwide Journal of Ecology92310ndash320
Webb CO Ackerly DD McPeek MA Donoghue MJ 2002Phylogenies and community ecology Annual Review ofEcology and Systematics 33475ndash505
Weiher E Keddy PA 2001 Ecological assembly rules per-spectives advances retreats Cambridge UK CambridgeUniversity Press
Westerling AL Hidalgo HG Cayan DR Swetnam TW 2006Warming and earlier spring increase western US forest wild-fire activity Science 313940
Westoby M Falster DS Moles AT Vesk PA Wright IJ 2002Plant ecological strategies Some leading dimensions ofvariation between species Annual Review of Ecology andSystematics 33125ndash159
Westoby M Wright IJ 2006 Land-plant ecology on thebasis of functional traits Trends in Ecology amp Evolution21261ndash268
Willis CG Halina M Lehman C Reich PB Keen A McCarthyS Cavender-Bares J 2010 Phylogenetic community structurein Minnesota oak savanna is influenced by spatial extent andenvironmental variation Ecography 33565ndash577
Wisheu IC Rosenzweig ML Olsvig-Whittaker L Shmida A2000 What makes nutrient-poor Mediterranean heathlandsso rich in plant diversity Evolutionary Ecology Research2935ndash955
Wright IJ Reich PB Cornelissen JHC Falster DS GroomPK Hikosaka K Lee W Lusk CH Niinemets OleksynJ 2005 Modulation of leaf economic traits and trait rela-tionships by climate Global Ecology and Biogeography14411ndash422
Wright IJ Reich PB Westoby M Ackerly DD Baruch Z BongersF Cavender-Bares J Chapin T Cornelissen JHC DiemerM 2004 The worldwide leaf economics spectrum Nature428821ndash827
Wright IJ Westoby M Reich PB 2002 Convergence towardshigher leaf mass per area in dry and nutrient poor habitats hasdifferent consequences for leaf life span Journal of Ecology90534ndash543
Dow
nloa
ded
by [U
nive
rsity
of C
alifo
rnia
Dav
is] a
t 08
21 1
8 N
ovem
ber 2
011




![Erratum: Secretome weaponries of Cochliobolus lunatus interacting with potato leaf at different temperature regimes reveal a CL[xxxx]LHM - motif](https://static.fdokumen.com/doc/165x107/633be2bafca68fa67503bbd6/erratum-secretome-weaponries-of-cochliobolus-lunatus-interacting-with-potato-leaf.jpg)















