
Wing venation and Distal-less expression in Heliconius butterfly wing pattern development

Functional Ecology
2007
21
, 28–40
28
© 2006 The Authors.Journal compilation© 2006 British Ecological Society
Blackwell Publishing Ltd
Leaf shape and venation pattern alter the support investments within leaf lamina in temperate species: a neglected source of leaf physiological differentiation?
Ü. NIINEMETS,*†‡ A. PORTSMUTH* and M. TOBIAS*
*
Department of Plant Physiology, University of Tartu, Riia 23, Tartu 51010, Estonia, and
‡
Institute of Agricultural and Environmental Sciences, Estonian University of Life Sciences, Kreutzwaldi 64, Tartu 51014, Estonia
Summary
1.
A trade-off between the investments in functional and support structures is a majordeterminant of leaf physiological activities. A variety of leaf shapes and venation densitiesoccur in coexisting vegetation, but the costs and benefits of various leaf shapes andvenation architectures are poorly understood. As the lever arms (location of leaf masscentre) become effectively longer as the maximum of lamina mass distribution shiftsfarther from the lamina base, we hypothesized that the fraction of lamina biomass inthe mid-rib (
F
MR
) is larger in leaves in which the centroid of lamina mass is located at,or greater than, half-leaf length (‘elliptic’ leaves) compared with leaves having thecentroid of lamina mass located closer to the leaf base (‘ovate’ leaves). We furtherhypothesized that minor vein density (
ρ
V
) is larger in leaves with lower
F
MR
, compensatingfor lower investments in central support. Finally, we predicted that
ρ
V
is lower in parallel/palmate-veined than in pinnate-veined leaves, due to a more uniform distribution oflarge veins in parallel/palmate-veined leaves.
2.
F
MR
and
ρ
V
were studied in 44 herbs and woody seedlings with an overall variationin lamina fresh mass (
M
FL
) of more than five orders of magnitude, and a sixfold variationin leaf longevity. Species were separated between pinnate-veined elliptic and ovateleaves, and parallel- or palmate-veined elliptic and ovate leaves.
3.
Contrary to the hypothesis, support investment in the mid-rib was similar amongleaf shapes, and scaled positively with leaf size and negatively with leaf longevity. However,
F
MR
and
ρ
V
were negatively associated. Fractional biomass investment in the mid-ribscaled with lamina size (fresh and dry mass and area), but at a common lamina size
F
MR
was larger in pinnate-veined elliptic than in parallel/palmate-veined elliptic leaves. Inaddition,
ρ
V
was larger in pinnate than in parallel/palmate-veined leaves, and thedifferences in lamina carbon content further suggested an overall greater investment oflamina biomass in the minor veins of pinnate-veined leaves.
4.
These data demonstrate that the effect of leaf shape on biomass investments incentral support is less significant than predicted by biomechanical models, partlybecause of the trade-off between the biomass investments in central support and minor veins,which compensate for differences in lamina shape. These data collectively indicate that leafsize, longevity, shape and venation pattern can importantly modify the distribution of foliagebiomass between support and functional tissues, and thus can alter foliage physiologicalactivity and leaf functioning in environments with different resource availability.
Key-words
: biomass partitioning, economics of support, leaf life span, leaf shape, leaf size, leaf structure,nitrogen content, support costs
Functional Ecology
(2007)
21
, 28–40doi: 10.1111/j.1365-2435.2006.01221.x
Introduction
Plant species with widely varying leaf shape, size andvenation architecture co-occur in vegetation. Whilethe differences in foliar physiognomy have attractednumerous researchers, the significance of different leaf
†Author to whom correspondence should be addressed. E-mail: [email protected]

29
Leaf shape and support costs
© 2006 The Authors.Journal compilation© 2006 British Ecological Society,
Functional Ecology
,
21
, 28–40
shapes and sizes for species niche differentiation is stillnot entirely understood, partly because of a lack ofinformation on the costs and benefits of specific leafforms. Variations in leaf size are typically explained byoptimization of temperature responses of leaf gasexchange in different environments (for review seeGivnish 1978, 1987). Apart from gas exchange, large-leaved species appear to require less stem xylem tosupport the same leaf area (Pickup
et al
. 2005), suggestingan advantage for larger-leaved species in humid, non-stressful environments where the influence of temper-ature on carbon gain is minor. However, mechanicaltheory predicts that, for leaves of common thickness,the fraction of within-leaf biomass invested in supportshould be greater in larger leaves due to effectivelylonger lever arms (Givnish 1978; Gere & Timoshenko1997; Niinemets & Fleck 2002), possibly offsetting theadvantages of the low stem xylem requirement oflarger leaves (Pickup
et al
. 2005). Within-leaf supportinvestments have been investigated in a few species andgenerally demonstrate a scaling of biomass investmentin the mid-rib with leaf size, with several exceptions andlarge interspecific variability (Givnish 1986; Niinemets& Sack 2006).
One factor potentially altering the relationshipsbetween the fraction of leaf biomass in support andleaf size is leaf shape. Leaf shape is strongly linked withvascular patterns and biomass investments in vasculature
(Dengler & Kang 2001; Kessler & Sinha 2004). Inparticular, the way leaf lamina mass is distributed fromleaf base to apex determines the overall bending momentand the support investment required by leaves of commonarea and mass. The support requirements are apparentlyless for cordate and ovate leaves, in which most leafmass is located close to the leaf base, than for ellipticor obovate leaves, in which the bulk of leaf mass is locatedfarther from the leaf base. Optimization of water supplyand biomechanical support for a given biomass invest-ment in the mid-rib suggests that the optimal leaf formis wedge-shaped (Givnish 1978). Although these leavesare very efficient in terms of support, most angiospermleaves are not wedge-shaped, possibly partly becausewedge-shaped leaves cannot be packed efficiently on thestem (Givnish 1978). In addition, allocating most of theleaf mass and the light-intercepting area further fromthe point of axial support reduces self-shading and in-creases leaf light interception, potentially enhancing leafcarbon gain (Pearcy & Yang 1998; Pearcy
et al
. 2005).There are fundamental relationships between leaf
physiological and structural characteristics that areapparently valid for leaves of widely different physio-gnomy (Wright
et al
. 2004). Negative relationshipsbetween leaf photosynthetic capacity per dry mass andleaf dry mass per unit area arise as leaves with more robuststructures have a greater fraction of support tissues withinthe leaves, less efficient internal diffusion, and lowerfractions of nitrogen invested in photosynthetic enzymes(Reich
et al
. 1995; Poorter & Evans 1998; Niinemets1999b; Wright
et al
. 2004; Terashima
et al
. 2005;Niinemets & Sack 2006; Niinemets
et al
. 2006a). How-ever, leaves of similar thickness and lamina structure, butof different size, may have widely different investmentsof foliar biomass in the mid-rib. Since the mid-rib haslower photosynthetic activity compared with the rest ofthe lamina, the contrasting within-leaf support invest-ments among leaves of varying shape and size cansignificantly modify the scaling relationships betweenleaf structure and physiology at any specific pointalong the fundamental spectrum of leaf functioning.This study aimed to test the hypothesis that within-leafsupport costs are greater for leaves with larger area andmass; and that for leaves with equivalent area and mass,the support costs are larger for elliptic and obovateleaves than for ovate and cordate leaves (Fig. 1).
Apart from the mid-rib, a large fraction of leaf bio-mass can be invested in minor veins, which also playan important role in mechanical support and waterconduction. In many species, a major part of leafhydraulic resistance is in the minor veins (Sack
et al
.2004, 2005). As the density of major veins is larger forpalmate- and parallel-veined than for pinnate-veinedleaves, and veins with a larger cross-sectional lumenarea are hydraulically more efficient (Sack & Holbrook2006), within-leaf costs for production of minor andmajor veins may be inversely associated. In addition,palmate- and parallel-veined leaves may also be mechan-ically more efficient for a given biomass investment in
Fig. 1. Outline of working hypotheses of how leaf shape and venation affect biomassinvestment in support. For leaves of the same length (L) the location of mass centre(centroid, c) varies in dependence on the distribution of leaf mass along the leaf length.We hypothesize that the fraction of biomass in the mid-rib required for leaf self-support is larger for leaves with biomass located at a distance x(c) ≥ L/2 (hypothesis 1).As large veins are distributed more uniformly in the leaf lamina for palmate- andparallel-veined than for pinnate-veined leaves, we further assume that the density ofminor veins (ρV, length per unit area) is larger for pinnate-veined leaves (hypothesis 2).The distance of mass-centre (centroid) location, x(c), relative to leaf length is 0·34 forthe depicted cordate, 0·4 for the ovate, 0·52 for the elliptic and 0·61 for the obovate leaf.

30
Ü. Niinemets
et al.
© 2006 The Authors.Journal compilation© 2006 British Ecological Society,
Functional Ecology
,
21
, 28–40
major veins (Roth-Nebelsick
et al
. 2001), especiallypalmate- and parallel-veined leaves with cordate andovate shapes that have a major part of the leaf biomasslocated close to the base. We tested the hypotheses thatpinnate-veined leaves have a higher density of minor veinsthan parallel- or palmate-veined leaves; and that thereis an overall negative relationship between minor veindensity and biomass investment in the mid-rib (Fig. 1).
In many plant species, turgor (the pressure exertedby cellular water on cell walls) contributes significantlyto leaf support, in particular in species with thin cellwalls and high leaf water content (Niklas 1986; Niklas1989), which also tend to have a shorter life span andgreater physiological activity (Reich
et al
. 1999; Wright
et al
. 2004). As species with higher leaf water content areexpected to rely more strongly on turgor for mechanicalsupport, we hypothesized further that among leaves ofcommon mass, area and shape, dry matter investmentin the mid-rib is less for short-lived leaves with relativelyhigher leaf water content.
Materials and methods
The study was conducted at the end of June 2000 inEstonia. Within-leaf and whole-plant biomass distributionwas studied in 26 herb species and in seedlings of sixwoody species (Table 1) in a broad-leaved deciduousforest in Ülenurme (58
°
18
′
N, 26
°
42
′
E, elevation60 m). The soil is a sandy-loam glossic gleysol (FAO,ISRIC & ISSS 1998; pseudopodzolic soil, cf. Kõlli &Ellermäe 2003 for a comparison between FAO andEstonian soil classifications). The forest overstorey isdominated by 16–19-m-tall trees of
Populus tremula
L.and
Fraxinus excelsior
L., while the shrub layer isdominated by
Corylus avellana
L. and the herb layer by
Calamagrostis arundinacea
(L.) Roth,
Fragaria vesca
L.,
Hepatica nobilis
Gras.,
Melampyrum nemorosum
L.and
Oxalis acetosella
L. A detailed description of thestudy site is provided by Niinemets & Kull (1998).
Above-ground plant material was collected foranalysis from at least two individuals of every species(average
±
SE = 3·0
±
0·7). Plants were sampled frommedium-sized gaps, and average (
±
SE) integrated dailyquantum flux density for May and June, determinedfrom hemispherical photographs (cf. Niinemets
et al
. 2004for details of light estimation), was 8·0
±
0·8 mol m
−
2
day
−
1
corresponding to
≈
25% canopy openness. Ingeneral, in herb species, light-driven foliage structuraland morphological plasticity saturates at a relativeirradiance of 20–30% of full light (Winn & Evans 1991;Olff 1992; Niinemets 2004).
The maximum average leaf fresh mass (FM) was
≈
20 g in the forest species. To achieve a larger range in leafFM and increase the generality of our results, within-leafbiomass partitioning was studied in 11 herb species withaverage leaf FM varying between
≈
4 and 320 g(Table 1) at the Botanical Garden of the University of
Tartu (58
°
22
′
N, 26
°
43
′
E, elevation 20 m). This sitesupported an open, deciduous, woody plantation on asandy-loam mollic cambisol (brown soil), and the average(
±
SE) integrated daily quantum flux density was 18·1
±
2·2 mol m
−
2
day
−
1
, corresponding to
≈
50% of full light.In all cases, the sampled material was enclosed in
plastic bags containing wet filter paper and was imme-diately transported to the laboratory for further analyses.
Species were selected for the study to achieve a re-presentative range of leaves with different size,shape and venation. Of the 44 species sampled (Table 1),33 were simple-leaved and 11 were compound-leaved (
Aegopodium podagraria
L.,
Angelica sylvestris
L.,
Anthriscus sylvestris
(L.) Hoffm.,
F. vesca
,
F. excelsior
,
Geranium palustre
L.,
O. acetosella
,
Paris quadrifolia
L.,
Rubus idaeus
L.,
Rubus saxatilis
L.,
Sorbus aucuparia
L.).For the entire data set, variation in average leaf FMwas
≈
15 000-fold (Table 1); for total within-leaf biomassfraction in support, the variation was approximatelyeightfold.
Most of the compound-leaved species sampled weresimple pinnate, but
A. sylvestris
was bipinnate;
F. vesca
and
O. acetosella
were trifoliate; and
G. palustre
waspalmately compound. In the analyses of leaf shapeand venation, leaflets of compound-leaved species wereconsidered as functional analogues of leaves in simple-leaved species (Givnish 1984). The species were dividedinto two groups (ovate
vs
elliptic) depending on thelocation of the centroid (mass centre) of the laminafrom the point of attachment (Fig. 1; Table 1). In‘ovate’-leaved species, the bulk of the leaf mass islocated close to the leaf attachment i.e., the laminamass centre is at a distance
a
<
L
/2 from the leafattachment to the petiole (
L
is total leaf length). In‘elliptic’-leaved species, the lamina centroid is locatedat a distance
a
≥
L
/2 from the leaf attachment to the pet-iole (see Table 2 for explanation of variables). Cordate,ovate or palmate shapes were considered ‘ovate’; elliptic,obcordate, oblong, obovate, orbicular, oval or reniformshapes were considered ‘elliptic’ (Table 1).
Species were grouped according to the venationpattern as pinnate-veined (higher-order veins stronglyassociated with central vein) and parallel/palmate-veined(higher-order veins less strongly linked to the mid-rib).Leaf venation was pinnate for 28 species, while six specieshad a parallel venation and 10 species a palmate venationpattern (Table 1). In all comparisons we pooled parallel-and palmate-veined species. Although the parallel andpalmate venations have their specific architecturalfeatures, the current comparison is justified from abiomechanical perspective as the major veins aredistributed more uniformly across the lamina in bothparallel- and palmate-veined leaves than in pinnate-veined leaves (Sack
et al
. 2003a).The 44 species studied belonged to 20 different
families (Table 1); each shape/venation group consisted

31
Leaf shape and
support costs
© 2006 T
he Authors.
Journal compilation
© 2006 B
ritish E
cological Society,
Functional E
cology
,
21
, 28–
40
Table 1.
Leaf habit, life span and average (
±
SE) lamina fresh mass (
M
FL
)*, dry mass per unit area (
M
A
), dry to fresh mass ratio (
D
F
) and nitrogen content per dry mass (
N
L
) in 33 species sampled from the broad-leaved mixed deciduous forest (F) and in 11 species sampled from the botanical garden (G)
Species Family Site Leaf/leaflet shape Venation typeLeaf life span (months)
M
FL
(g)*
M
A
(g m
−
2
)
D
F
(g g
−
1
) NL (%)
Aegopodium podagraria L. Apiaceae F Ovate Pinnate 3·5 0·159 ± 0·012 29·2 ± 1·7 0·293 ± 0·014 2·02 ± 0·19Angelica sylvestris L. Apiaceae G Ovate Pinnate 3·5 0·15 ± 0·08 19·1 ± 1·9 0·255 ± 0·029 2·61 ± 0·21Anthriscus sylvestris (L.) Hoffm. Apiaceae F Ovate Pinnate 3·5 0·0129 ± 0·001 16·2 ± 1·9 0·256 ± 0·029 2·72 ± 0·34Arctium tomentosum Mill. Compositae G Cordate Pinnate 3·5 6·0 ± 2·3 41 ± 7 0·175 ± 0·045 2·5 ± 0·7Armoracia rusticana (Lam.) Gaer. Cruciferae G Elliptic Pinnate 2 46 ± 20 24·8 ± 3·5 0·18 ± 0·05 3·43 ± 0·48Asarum europaeum L. Aristolochiaceae F Reniform Parallel 13 0·413 ± 0·047 44·4 ± 2·9 0·231 ± 0·014 2·24 ± 0·17Cirsium oleraceum (L.) Scop. Compositae F Ovate Pinnate 3·5 10·0 ± 3·5 22·5 ± 2·3 0·101 ± 0·029 3·04 ± 0·35Convallaria majalis L. Liliaceae F Elliptic Parallel 3·5 0·42 ± 0·08 44·2 ± 3·7 0·31 ± 0·05 2·15 ± 0·42Corylus avellana L. Corylaceae F Oval/obovate Pinnate 7 0·23 ± 0·10 29 ± 5 0·36 ± 0·06 2·14 ± 0·25Crepis paludosa (L.) Moench Compositae F Obovate Pinnate 3·5 0·37 ± 0·35 13·8 ± 1·3 0·165 ± 0·014 1·97 ± 0·22Cucurbita pepo L. Cucurbitaceae G Orbicular Palmate 2 57 ± 16 27·7 ± 4·5 0·083 ± 0·012 6·6 ± 0·5Echinops sphaerocephalus L. Compositae G Obovate Pinnate 3·5 7·8 ± 1·2 47 ± 7 0·189 ± 0·037 3·25 ± 0·36Epilobium montanum L. Onagraceae F Ovate Pinnate 3·5 0·17 ± 0·07 22·3 ± 4·6 0·112 ± 0·032 4·4 ± 0·6Fragaria vesca L. Rosaceae F Oval Pinnate 3·5 0·056 ± 0·009 32 ± 8 0·414 ± 0·029 1·92 ± 0·04Fraxinus excelsior L. Oleaceae F Elliptic Pinnate 7 0·029 ± 0·009 18·0 ± 2·6 0·266 ± 0·018 1·60 ± 0·27Galeobdolon luteum Huds. Labiatae F Ovate Pinnate 3·5 0·101 ± 0·047 35 ± 10 0·262 ± 0·036 2·45 ± 0·12Geranium palustre L. Geraniaceae F Palmate Palmate 3·5 0·63 ± 0·06 23 ± 6 0·226 ± 0·024 2·16 ± 0·28Helianthus annuus L. Compositae G Cordate Palmate 2 38 ± 10 35·3 ± 2·7 0·111 ± 0·029 4·7 ± 1·1Hepatica nobilis Gars. Ranunculaceae F Orbicular Palmate 13 0·370 ± 0·029 46·8 ± 2·2 0·320 ± 0·010 1·91 ± 0·09Heracleum sosnowskyi Manden Apiaceae G Orbicular/ovate Pinnate 2 30 ± 12 31 ± 5 0·167 ± 0·045 2·4 ± 0·5Ligularia wilsoniana (Hems.) Green. Compositae G Cordate Pinnate 3·5 26 ± 14 36·4 ± 2·9 0·19 ± 0·05 1·53 ± 0·19Lonicera xylosteum L. Caprifoliaceae F Oval Pinnate 7 0·12 ± 0·05 47 ± 17 0·329 ± 0·017 1·97 ± 0·45Maianthemum bifolium (L.) Schm. Liliaceae F Cordate Parallel 3·5 0·18 ± 0·05 35·8 ± 2·9 0·251 ± 0·035 1·33 ± 0·04Melampyrum nemorosum L. Scrophulariaceae F Ovate Pinnate 3·5 0·0565 ± 0·045 29·3 ± 4·5 0·237 ± 0·015 2·93 ± 0·45Oxalis acetosella L. Oxalidaceae F Obcordate Parallel 6 0·033 ± 0·026 12·9 ± 3·0 0·103 ± 0·047 2·39 ± 0·48Padus avium Mill. Rosaceae F Elliptic Pinnate 7 0·20 ± 0·06 41 ± 11 0·41 ± 0·09 1·98 ± 0·35Paris quadrifolia L. Liliaceae F Obovate Parallel 3·5 0·18 ± 0·05 16·3 ± 3·6 0·158 ± 0·024 2·04 ± 0·18Polygonatum multiflorum (L.) All. Liliaceae F Oval Parallel 3·5 0·46 ± 0·09 28·4 ± 2·8 0·21 ± 0·06 2·42 ± 0·15Polygonum cuspidatum Sieb. et Zucc. Polygonaceae G Ovate Pinnate 7 4·0 ± 1·3 56 ± 9 0·32 ± 0·07 1·70 ± 0·50Populus tremula L. Salicaceae F Orbicular Pinnate 7 0·19 ± 0·06 26 ± 5 0·254 ± 0·039 3·5 ± 0·8Prunella vulgaris L. Labiatae F Ovate Pinnate 3·5 0·063 ± 0·015 16·7 ± 1·9 0·181 ± 0·027 1·82 ± 0·43Ranunculus cassubicus L. Ranunculaceae F Reniform Palmate 3·5 0·87 ± 0·21 44·6 ± 1·6 0·281 ± 0·049 2·37 ± 0·41Rheum rhabarbarum L. Polygonaceae G Cordate Palmate 2 208 ± 92 34·3 ± 1·8 0·066 ± 0·002 3·74 ± 0·23Ribes alpinum L. Grossulariaceae F Ovate Palmate 7 0·136 ± 0·019 42 ± 11 0·276 ± 0·021 2·0 ± 0·7Ribes nigrum L. Grossulariaceae F Ovate Palmate 7 0·253 ± 0·045 34·5 ± 3·5 0·360 ± 0·011 1·32 ± 0·43Rubus idaeus L. Rosaceae F Ovate Pinnate 7 0·043 ± 0·018 37 ± 7 0·28 ± 0·07 2·98 ± 0·49

32Ü. Niinemets et al.
© 2006 The Authors.Journal compilation© 2006 British Ecological Society, Functional Ecology, 21, 28–40 R
ubus
sax
atili
s L
.R
osac
eae
FO
vate
Pin
nate
3·5
0·20
± 0
·06
22·9
± 3
·90·
261
± 0·
043
1·49
± 0
·34
Rum
ex c
risp
us L
.Po
lygo
nace
aeG
Ova
teP
inna
te3·
58·
3 ±
4·2
45·6
± 2
·60·
280
± 0·
034
2·20
± 0
·15
Sor
bus
aucu
pari
a L
. R
osac
eae
FE
llipt
icP
inna
te7
0·02
2 ±
0·01
830
± 1
00·
416
± 0·
032
2·11
± 0
·33
Ste
llari
a ho
lost
ea L
.C
aryo
phyl
lace
aeF
Ova
te/la
nceo
late
Pin
nate
3·5
0·04
8 ±
0·02
733
·0 ±
4·2
0·24
1 ±
0·01
21·
97 ±
0·4
2T
arax
acum
offi
cina
le W
eb. e
x W
igg.
Com
posi
tae
FO
bova
teP
inna
te2
0·72
± 0
·37
20·2
± 3
·40·
088
± 0·
014
2·44
± 0
·33
Tus
sila
go fa
rfar
a L
.C
ompo
sita
eF
Cor
date
Pal
mat
e3·
52·
2 ±
2·0
33·2
± 4
·10·
180
± 0·
043
3·22
± 0
·43
Ver
basc
um n
igru
m L
.Sc
roph
ular
iace
aeF
Ova
teP
inna
te3·
55·
2 ±
2·4
30·9
± 4
·90·
115
± 0·
038
3·6
± 0·
5V
eron
ica
cham
aedr
ys L
.Sc
roph
ular
iace
aeF
Ova
teP
inna
te3·
50·
034
± 0·
010
19·3
± 3
·20·
207
± 0·
023
1·98
± 0
·09
Cor
ylus
ave
llana
, Lon
icer
a xy
lost
eum
, Rib
es a
lpin
um, R
ibes
nig
rum
and
Rub
us id
aeus
are
shr
ubs;
Fra
xinu
s ex
cels
ior,
Pad
us a
vium
, Pop
ulus
tre
mul
a an
d S
orbu
s au
cupa
ria
are
tree
s; a
ll ot
her
spec
ies
are
herb
s.*L
amin
a F
M in
clud
es m
id-r
ib a
nd th
e re
st o
f th
e la
min
a. F
or c
ompo
und-
leav
ed s
peci
es (A
egop
odiu
m p
odag
rari
a, A
ngel
ica
sylv
estr
is, A
nthr
iscu
s sy
lves
tris
, Fra
gari
a ve
sca,
Fra
xinu
s ex
cels
ior,
Ger
aniu
m p
alus
tre,
O
xalis
ace
tose
lla, P
aris
qua
drif
olia
, Rub
us id
aeus
, Rub
us s
axat
ilis,
Sor
bus
aucu
pari
a), a
vera
ge la
min
a F
M o
f le
aflet
s is
pro
vide
d.
Spec
ies
Fam
ilySi
teL
eaf/
leafl
et s
hape
Ven
atio
n ty
peL
eaf
life
span
(m
onth
s)M
FL (
g)*
MA (
g m
−2)
DF (
g g−1
)N
L (
%)
Tab
le 1
.C
onti
nued
of species belonging at least to five different families,and all groups contained species belonging to the samefamilies. Thus the species distribution among groupswas phylogenetically heterogeneous and the groupdifferences demonstrated in our study are not drivenby phylogenetic differences among the groups (sensuAckerly & Reich 1999).
Seven of the species sampled have been introducedto Estonia from southern Europe (Echinops sphaero-cephalus L., Heracleum sosnowskyi Manden, Rheumrhabarbarum L.); temperate East Asia [Japan, China:Ligularia wilsoniana (Hems.) Green, Polygonum cuspida-tum Sieb. ex Zucc.]; North America (Helianthus annuusL.) and Central America (Cucurbita pepo L.). Except forC. pepo and H. annuus, all foreign species disperse nat-urally in Estonia, and may be invasive in suitable sites.
Our data set included deciduous herbs and woodyspecies as well as evergreen herbs (Asarum europaeumL., H. nobilis, O. acetosella; Table 1). Leaf life span (Λ)was estimated using the GLOPNET database, currentlythe most comprehensive source of leaf life span (Wrightet al. 2004), and our own knowledge of species biology.Literature data for deciduous species were adjustedto the length of growing season at our latitudes. Thuswoody deciduous species were assigned Λ = 7 months,and most perennial herbs Λ = 3·5 months (7.5–8·5months for woody deciduous and 4–4·5 months fordeciduous perennial herbs in the original GLOPNETdatabase, Wright et al. 2004). Short-lived cultural plants[Armoracia rusticana (Lam.) Gaer., C. pepo, H. annuus,R. rhabarbarum] and invasive herbs (H. sosnowskyi,Taraxacum officinale Web. ex Wigg.) with rapid leafturnover were assigned a life span of 2 months, whilethe perennial shrub-like herb P. cuspidatum, whichproduces only one leaf flush in our climate, wasassigned Λ = 7 months. The evergreen herbs A. europaeumand H. nobilis have a life span of 13 months. AlthoughO. acetosella is also a wintergreen species, it continuouslyreplaces leaves during the growing season (Tessier2004), and the average leaf longevity of this speciesis therefore ≈6 months in our climate. As there canbe important variation in leaf longevity withinspecies groups (Diemer 1998), our estimates of Λ arecrude. Nevertheless, we suggest that even these roughestimates allow insights into the correlations between leaflongevity, leaf structure and investment in support.
Representative leaves from each plant were selectedfor detailed analyses. In compound-leaved species,a representative leaflet was further chosen. Theselected leaf lamina or leaflet lamina was dividedbetween the mid-rib and the rest of the lamina. After

33Leaf shape and support costs
© 2006 The Authors.Journal compilation© 2006 British Ecological Society, Functional Ecology, 21, 28–40
determination of the fresh mass of the mid-rib andthe rest of the lamina, all leaves were photocopied forarea measurements.
Dry masses of the mid-rib and lamina were estimatedafter oven-drying at 70 °C for at least 48 h, and dry tofresh mass ratios were calculated for the leaf fractions.The partitioning in support within leaf lamina (FMR) wascharacterized as the ratio of mid-rib dry mass (MMR) tothe sum of MMR and the rest of the lamina (ML).
The photocopied leaf images were scanned with aresolution of 300 dpi, and area, perimeter and elonga-tion (ratio of leaf major and minor semi-axes, ε; Baxes1994) were estimated separately for the selected leavesand leaflets with UTHSCSA imagetool 2·00 alpha (C.Donald Wilcox, S. Brent Dove, W. Doss McDavid andDavid B. Greer, Department of Dental DiagnosticScience, The University of Texas Health Science Center,San Antonio, TX, USA; http://ddsdx.uthscsa.edu). Leafdry mass per unit area (MAW) and lamina (withoutmid-rib) dry mass per unit area (MA), were also calcu-lated for the leaves sampled.
Leaf elongation characterizes the overall slender-ness of the leaves. To characterize the complexity of thelamina outline, we calculated the adimensional ratio oflamina (perimeter)2/area (γ) (Sack et al. 2003b). Leafperimeter-to-area ratio (m−1) has also been used toevaluate the degree of lamina dissection, but in contrastto γ, it is negatively correlated with leaf size (Sack et al.2003b). For all data pooled in our data set, we observedr 2 = 0·85, P < 0·001 for a correlation between perimeter/area and lamina fresh mass. Thus we prefer γ overperimeter/area ratio in our analysis.
Leaves and leaflets sampled for detailed analyses ofbiomass partitioning were also used to determine thedensity of veins. Digital images from the lower leaf
surface were made in bright field using a Nikon EclipseE-600 microscope (Nikon Corporation, Tokyo, Japan)equipped with a Nikon Coolpix 990 digital camera.The effective magnification was 30–120× (size of sampledarea, 4–60 mm2). Three to five images were taken indifferent positions on the same leaf. The length of sec-ondary, tertiary and quaternary veins was measured fromevery image using UTHSCSA imagetool 2·00 alpha, andthe average density of veins was calculated (ρV, mmmm−2). Leaf-specific mean values were further averagedto obtain a species-specific average value of vein density.
Lamina and mid-rib C and N contents were deter-mined for every selected leaf and leaflet using a PerkinElmer series II CHNS/O Analyzer 2400 (Perkin ElmerLife and Analytical Sciences, Inc., Boston, MA, USA).Values of C and N content were averaged for allindividual leaves sampled for a given species to determinespecies-specific means.
We used linear and non-linear regression analyses inthe form y = a log(x) + b and y = axb to test for statisticalrelationships between leaf characteristics. To improvethe linearity and normality of these relationships,lamina fresh (MFL) and dry (MDL) mass and leaf longevity(Λ) were log-transformed before statistical analysis.All regressions were considered significant at P < 0·05.Multiple linear regressions were employed to examinethe effect of leaf dry to fresh mass ratio on the fractionalbiomass investment in support, in leaves of varying size.
Means of different characteristics were separatedamong species groups (pinnate- vs parallel/palmate-veined species; ovate- vs elliptic-leaved species) using. In these comparisons, only species with leafelongation (ε, ratio of leaf length to width) <4 wereincluded, to avoid the bias due to species with narrowleaves that were present in some leaf shape/venationcombinations. The statistical relationships betweenspecies groups (pinnate-veined vs parallel/palmate-veined;ovate vs elliptic leaves; compound vs simple leaves) werecompared with covariance analyses () includingall species. For non-linear relationships, the independentvariable was log-transformed before the . Aseparate-slope was used first to test for theintercept differences among the groups. Whenever theinteraction term (grouping variable) × covariate wasstatistically not significant (P > 0·05), the analysis wascontinued according to a common-slope model to testfor the intercept differences (Sokal & Rohlf 1995).
As the species were sampled from two sites that dif-fered in edaphic conditions, in over- and understoreydominants and in light availability, we also tested forsite effect on leaf structure and biomass partitioningusing ; and on leaf structure and biomasspartitioning vs log(MFL) and log(MDL) relationships
Table 2. Variables used in the text
Variable Definition
ε (–) Lamina elongation (ratio of lamina major to minor semi-axes)γ (–) Lamina (perimeter)2/areaΛ (months) Leaf life spanρV (mm mm−2) Vein densitya (m) Distance from leaf attachmentCL (%) Lamina (without mid-rib) carbon content per dry massDF (g g−1) Lamina (without mid-rib) dry to fresh mass ratioFMR (g g−1) Mass fraction of mid-rib in lamina, MMR/(ML + MMR)L (m) Leaf lengthMA (g m−2) Lamina (without mid-rib) dry mass per unit areaMAW (g m−2) Total lamina (with mid-rib) dry mass per unit areaMDL (g) Total lamina (with mid-rib) dry massMFL (g) Total lamina (with mid-rib) fresh massML (g) Lamina (without mid-rib) dry massMMR (g) Mid-rib dry massNL (%) Lamina (without mid-rib) nitrogen content per dry massNMR (%) Mid-rib N content per dry massx(c) (m) Location of lamina mass centre from petiole attachment

34Ü. Niinemets et al.
© 2006 The Authors.Journal compilation© 2006 British Ecological Society, Functional Ecology, 21, 28–40
using covariation analyses. According to , andseparate-slope and common-slope models,site × covariate interactions and site effects alone werenot significant in any of the statistical relationshipsanalysed (P > 0·3), suggesting that size-dependentmodifications in leaf characteristics and biomassallocation between support and physiological structuresoccurred similarly in both sites. Thus data from bothsites were pooled in the following analyses. Allsymbols are explained in Table 2.
Results
-
The average density of minor veins (ρV) varied from0·23 to 0·91 mm mm−2 for 16 species with parallel orpalmate venation, while the range of variation was0·15–0·95 mm mm−2 for 28 species with pinnate venation.For leaves of similar elongation (ε < 4), ρV was largerfor pinnate-veined (average ± SE = 0·62 ± 0·05) thanfor parallel/palmate-veined species (0·48 ± 0·07; P < 0·02).This overall difference was due to larger ρV in pinnate-veined leaves with elliptic shape (Table 3). In ovate leaves,ρV was not significantly different among pinnate- andparallel/palmate-veined species (Table 3).
The fraction of lamina biomass in the mid-rib[FMR = MMR/(ML + MMR)] varied from 0·017 to 0·39 g g−1
for parallel/palmate-veined species, and from 0·048 to0·40 g g−1 for pinnate-veined species. The mid-rib drymass fraction did not depend on leaf venation andshape (Table 3).
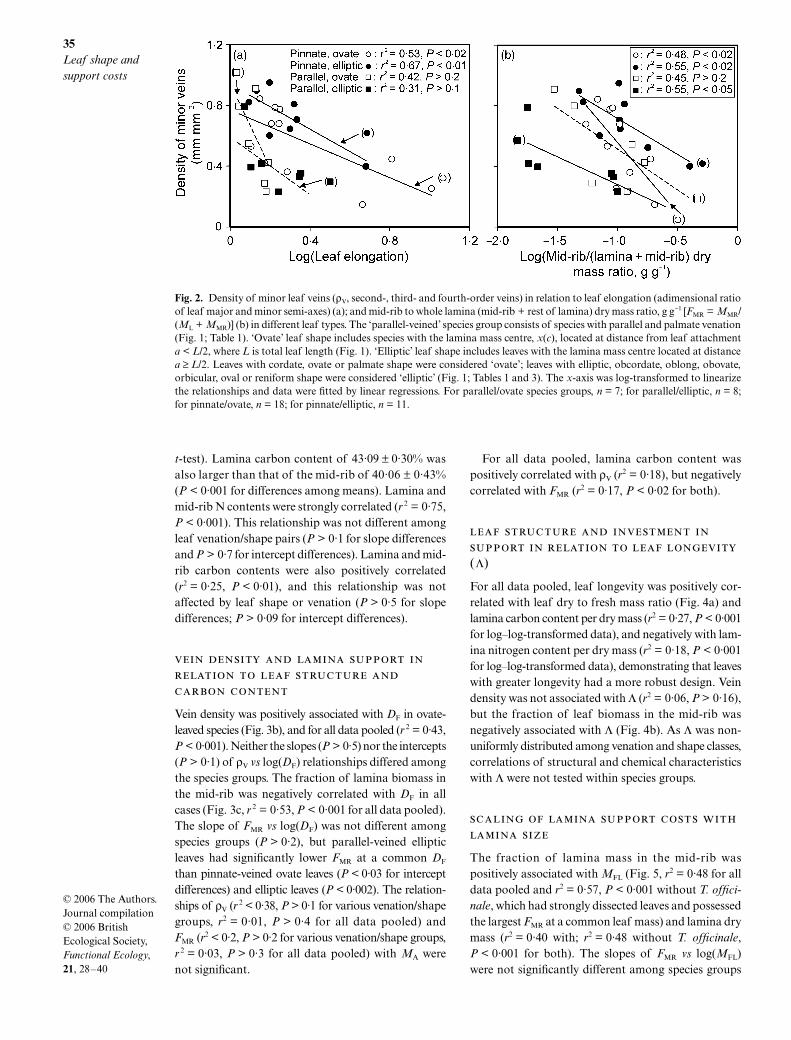
Vein density scaled negatively with leaf elongation inpinnate-veined species (Fig. 2a), while ρV was negativelyassociated with (perimeter)2/area (γ) only in pinnate-veined elliptic leaves (r 2 = 0·45, P < 0·05 for a correla-tion in the form y = ax b), but not for other venation/shape combinations (P > 0·1). The fraction of laminamass in the mid-rib was positively related to laminaelongation (ε) (r2 = 0·82, P < 0·001) and γ (r 2 = 0·65,
P < 0·01) in pinnate-veined elliptic leaves. For othervenation/shape combinations, r 2 < 0·20, P > 0·3 forboth ε and γ.
Negative relationships were observed between ρV
and lamina mass in the mid-rib for pinnate-leavedspecies, and for parallel-veined ovate-leaved species(Fig. 2b). According to (venation type as maineffect, log(ρV) as covariate), pinnate-leaved species hada larger ρV at a common fractional biomass investmentin the mid-rib (P < 0·001). This effect resulted fromlower ρV in parallel-veined elliptic leaves than in ovateand elliptic pinnate-veined leaves (P < 0·001).
Lamina dry mass per unit area (MA), dry to fresh massratio (DF), lamina N content per dry mass (NL, Table 3),and mid-rib N content per dry mass (NMR, P = 0·16)were not different among leaf shape/venation combi-nations. However, lamina carbon content per dry mass(CL) was significantly larger for pinnate-veined ellipticleaves than for parallel/palmate-veined ovate leaves(Table 3), and the average lamina carbon content of43·60 ± 0·33% of all pinnate-veined species pooled wasalso significantly larger (P < 0·02) than that of parallel/palmate-veined species pooled (42·1 ± 0·5%).
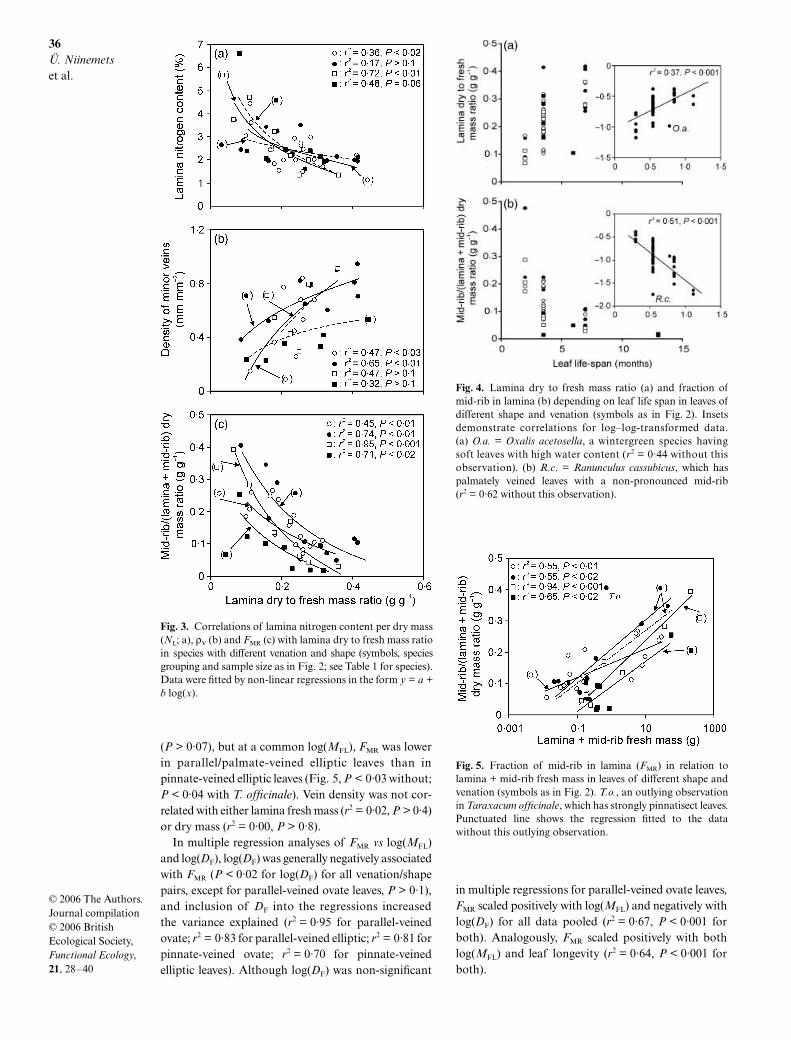
Lamina dry to fresh mass ratio was positively corre-lated with MA when all data were pooled (r2 = 0·18,P < 0·001), but among species groups this relationshipwas significant only for parallel/palmate-veined ellipticleaves (r2 = 0·63, P < 0·02). DF was negatively corre-lated with NL (Fig. 3a, r2 = 0·39, P < 0·001 for all datapooled), and positively correlated with CL (r2 = 0·31,P < 0·001 for all data pooled), while neither NL (r2 = 0·01,P > 0·6) nor CL (r2 = 0·07, P > 0·09) was correlatedwith MA. Neither the slopes (P > 0·09) nor the inter-cepts (P > 0·2) of NL vs log(DF) (P > 0·4) and CL vslog(DF) relationships were significantly differentamong the leaf venation/shape pairs.
Lamina N content was 1·8-fold larger than thatof mid-ribs (P < 0·001 according to a paired samples
Table 3. Average (± SE) density of minor veins (ρV), mid-rib to total lamina dry mass ratio FMR = MMR/(MMR + ML), lamina drymass per unit area (MA), lamina dry to fresh mass ratio (DF), and lamina nitrogen (NL) and carbon (CL) contents per dry massin species with different foliage venation and shape*
Venation type Leaf shape ρV (mm mm−2) FMR (g g−1) MA (g m−2) DF (g g−1) NL (%) CL (%) n
Parallel/palmate Ovate 0·59 ± 0·11ab 0·159 ± 0·051a 34·1 ± 2·1a 0·210 ± 0·038a 2·63 ± 0·48a 41·8 ± 0·8a 7Parallel/palmate Elliptic 0·39 ± 0·07a 0·089 ± 0·028a 33·2 ± 4·8a 0·212 ± 0·032a 2·8 ± 0·6a 42·4 ± 0·7ab 8Pinnate Ovate 0·55 ± 0·08ab 0·150 ± 0·016a 30·2 ± 2·5a 0·219 ± 0·015a 2·47 ± 0·18a 43·32 ± 0·41ab 15Pinnate Elliptic 0·71 ± 0·06b 0·165 ± 0·037a 30·3 ± 3·7a 0·277 ± 0·035a 2·39 ± 0·20a 44·1 ± 0·6b 10
*Species as described in Table 1. Three ovate-leaved and two elliptic-leaved species with leaf elongation (ratio of leaf length to width) >4 were removed from the statistical comparisons (Fig. 2a). Ovate-leaved group includes species with the centroid of lamina mass located at distance a from leaf attachment <L/2, where L is total leaf length. The elliptic-leaved group consists of species with the centre of lamina mass located at a ≥ L/2. In this analysis, leaves with cordate, ovate or palmate shape were considered ‘ovate’, while leaves with elliptic, obcordate, oblong, obovate, orbicular, oval or reniform shape were considered ‘elliptic’. The species groups were compared using and means were separated using α Bonferroni tests. Means with the same lower case letter are not significantly different among groups (P > 0·05).
35Leaf shape and support costs
© 2006 The Authors.Journal compilation© 2006 British Ecological Society, Functional Ecology, 21, 28–40
t-test). Lamina carbon content of 43·09 ± 0·30% wasalso larger than that of the mid-rib of 40·06 ± 0·43%(P < 0·001 for differences among means). Lamina andmid-rib N contents were strongly correlated (r 2 = 0·75,P < 0·001). This relationship was not different amongleaf venation/shape pairs (P > 0·1 for slope differencesand P > 0·7 for intercept differences). Lamina and mid-rib carbon contents were also positively correlated(r2 = 0·25, P < 0·01), and this relationship was notaffected by leaf shape or venation (P > 0·5 for slopedifferences; P > 0·09 for intercept differences).
Vein density was positively associated with DF in ovate-leaved species (Fig. 3b), and for all data pooled (r 2 = 0·43,P < 0·001). Neither the slopes (P > 0·5) nor the intercepts(P > 0·1) of ρV vs log(DF) relationships differed amongthe species groups. The fraction of lamina biomass inthe mid-rib was negatively correlated with DF in allcases (Fig. 3c, r 2 = 0·53, P < 0·001 for all data pooled).The slope of FMR vs log(DF) was not different amongspecies groups (P > 0·2), but parallel-veined ellipticleaves had significantly lower FMR at a common DF
than pinnate-veined ovate leaves (P < 0·03 for interceptdifferences) and elliptic leaves (P < 0·002). The relation-ships of ρV (r 2 < 0·38, P > 0·1 for various venation/shapegroups, r2 = 0·01, P > 0·4 for all data pooled) andFMR (r2 < 0·2, P > 0·2 for various venation/shape groups,r 2 = 0·03, P > 0·3 for all data pooled) with MA werenot significant.
For all data pooled, lamina carbon content waspositively correlated with ρV (r2 = 0·18), but negativelycorrelated with FMR (r2 = 0·17, P < 0·02 for both).
(Λ)
For all data pooled, leaf longevity was positively cor-related with leaf dry to fresh mass ratio (Fig. 4a) andlamina carbon content per dry mass (r2 = 0·27, P < 0·001for log–log-transformed data), and negatively with lam-ina nitrogen content per dry mass (r2 = 0·18, P < 0·001for log–log-transformed data), demonstrating that leaveswith greater longevity had a more robust design. Veindensity was not associated with Λ (r2 = 0·06, P > 0·16),but the fraction of leaf biomass in the mid-rib wasnegatively associated with Λ (Fig. 4b). As Λ was non-uniformly distributed among venation and shape classes,correlations of structural and chemical characteristicswith Λ were not tested within species groups.
The fraction of lamina mass in the mid-rib waspositively associated with MFL (Fig. 5, r2 = 0·48 for alldata pooled and r2 = 0·57, P < 0·001 without T. offici-nale, which had strongly dissected leaves and possessedthe largest FMR at a common leaf mass) and lamina drymass (r2 = 0·40 with; r2 = 0·48 without T. officinale,P < 0·001 for both). The slopes of FMR vs log(MFL)were not significantly different among species groups
Fig. 2. Density of minor leaf veins (ρV, second-, third- and fourth-order veins) in relation to leaf elongation (adimensional ratioof leaf major and minor semi-axes) (a); and mid-rib to whole lamina (mid-rib + rest of lamina) dry mass ratio, g g−1 [FMR = MMR/(ML + MMR)] (b) in different leaf types. The ‘parallel-veined’ species group consists of species with parallel and palmate venation(Fig. 1; Table 1). ‘Ovate’ leaf shape includes species with the lamina mass centre, x(c), located at distance from leaf attachmenta < L/2, where L is total leaf length (Fig. 1). ‘Elliptic’ leaf shape includes leaves with the lamina mass centre located at distancea ≥ L/2. Leaves with cordate, ovate or palmate shape were considered ‘ovate’; leaves with elliptic, obcordate, oblong, obovate,orbicular, oval or reniform shape were considered ‘elliptic’ (Fig. 1; Tables 1 and 3). The x-axis was log-transformed to linearizethe relationships and data were fitted by linear regressions. For parallel/ovate species groups, n = 7; for parallel/elliptic, n = 8;for pinnate/ovate, n = 18; for pinnate/elliptic, n = 11.
36Ü. Niinemets et al.
© 2006 The Authors.Journal compilation© 2006 British Ecological Society, Functional Ecology, 21, 28–40
(P > 0·07), but at a common log(MFL), FMR was lowerin parallel/palmate-veined elliptic leaves than inpinnate-veined elliptic leaves (Fig. 5, P < 0·03 without;P < 0·04 with T. officinale). Vein density was not cor-related with either lamina fresh mass (r2 = 0·02, P > 0·4)or dry mass (r2 = 0·00, P > 0·8).
In multiple regression analyses of FMR vs log(MFL)and log(DF), log(DF) was generally negatively associatedwith FMR (P < 0·02 for log(DF) for all venation/shapepairs, except for parallel-veined ovate leaves, P > 0·1),and inclusion of DF into the regressions increasedthe variance explained (r2 = 0·95 for parallel-veinedovate; r2 = 0·83 for parallel-veined elliptic; r2 = 0·81 forpinnate-veined ovate; r2 = 0·70 for pinnate-veinedelliptic leaves). Although log(DF) was non-significant
in multiple regressions for parallel-veined ovate leaves,FMR scaled positively with log(MFL) and negatively withlog(DF) for all data pooled (r2 = 0·67, P < 0·001 forboth). Analogously, FMR scaled positively with bothlog(MFL) and leaf longevity (r2 = 0·64, P < 0·001 forboth).
Fig. 3. Correlations of lamina nitrogen content per dry mass(NL; a), ρV (b) and FMR (c) with lamina dry to fresh mass ratioin species with different venation and shape (symbols, speciesgrouping and sample size as in Fig. 2; see Table 1 for species).Data were fitted by non-linear regressions in the form y = a +b log(x).
Fig. 4. Lamina dry to fresh mass ratio (a) and fraction ofmid-rib in lamina (b) depending on leaf life span in leaves ofdifferent shape and venation (symbols as in Fig. 2). Insetsdemonstrate correlations for log–log-transformed data.(a) O.a. = Oxalis acetosella, a wintergreen species havingsoft leaves with high water content (r2 = 0·44 without thisobservation). (b) R.c. = Ranunculus cassubicus, which haspalmately veined leaves with a non-pronounced mid-rib(r2 = 0·62 without this observation).
Fig. 5. Fraction of mid-rib in lamina (FMR) in relation tolamina + mid-rib fresh mass in leaves of different shape andvenation (symbols as in Fig. 2). T.o., an outlying observationin Taraxacum officinale, which has strongly pinnatisect leaves.Punctuated line shows the regression fitted to the datawithout this outlying observation.

37Leaf shape and support costs
© 2006 The Authors.Journal compilation© 2006 British Ecological Society, Functional Ecology, 21, 28–40
Discussion
We hypothesized that the density of minor veins (ρV) islarger for pinnate-veined species, because the majorveins are less uniformly distributed within the leaf inpinnate-veined than in parallel- and palmate-veinedspecies (Fig. 1). All data collectively supported thishypothesis (Table 3). Vein density was further stronglyassociated with lamina carbon content (CL). As leafcarbon contents scale with the content of carbon-richchemicals such as lignin (Niinemets et al. 1999; Poorter1994), CL provides an alternative estimate of within-lamina investments in support, including minor veinsand cell walls. This positive correlation, and larger laminacarbon contents in pinnate-veined leaves, suggest anoverall greater biomass investment in within-leaf support(minor veins and cell walls) in these leaves (Table 3).
However, the difference in ρV and CL between leaveswith different venation types mainly reflected thedifference among leaves with the centre of lamina masslocated further than half the leaf length (‘elliptic’ leaves,Fig. 1), while the average values of ρV and CL didnot differ among pinnate- and parallel/palmate-veinedovate leaves. This may reflect the overall greatersupport requirements of mechanically less efficientelliptic leaves.
Modifications in venation architecture also affectleaf hydraulic characteristics, and can be driven partlyby requirements for efficient water conduction. Strongpositive correlations have been observed between veindensity and leaf hydraulic conductance (Sack & Frole2006). Thus optimization of leaf hydraulic architecturecan provide an alternative explanation for the observeddifferences in vein density. Because the hydraulicconductance of veins is not infinite, significant waterpotential gradients develop from the lamina basetowards the tip, resulting in particularly low laminawater potentials at the apical lamina parts (Zwienieckiet al. 2002). In elliptic leaves, in which the taperingmid-rib supplies a disproportionately large part of thelamina area, hydraulic limitations in apical portions ofpinnately veined leaves are expected to be especiallysevere. Minor veins constitute often the largestfraction of total leaf hydraulic resistance (Sack et al.2004, 2005), and increases in the number of parallelvascular paths through enhanced vein density mayprovide an important compensatory way to reducewater potential gradients along the lamina. Overall,the greater vein density in pinnate-veined leaves agreeswith the evidence that whole-leaf hydraulic conductanceis determined more strongly by the conductance ofminor veins in pinnate-veined than in palmate-veinedleaves (Sack et al. 2003b, 2004).
As leaf dissection, lobation and serration patternsfollow the distribution of secondary and tertiary veins,increases in the complexity of leaf perimeter have been
associated with increases in vein density (Sisó et al.2001). In our study, the ratio lamina (perimeter)2/areawas associated with vein density only in pinnate-veinedelliptic leaves, but this correlation was negative.Although, in our study, only a few species had stronglydissected laminas, this contrasting pattern suggeststhat the effects of lamina dissection are superimposedby overall modifications of venation architecture andlamina shape.
For pinnate-veined leaves, we further observed adecrease in vein density with leaf elongation (Fig. 2a),indicating that with decreasing the average distance ofleaf margins from the mid-rib, the requirement of minorveins for mechanical support or water conductionis lower. These data collectively indicate a significantinterspecific variation in vein density that is associatedwith venation architecture, lamina shape and elongation,and has important implications for lamina hydraulicefficiency and mechanical support.
- ,
Our study highlights a large fractional investment oflamina biomass in the mid-rib (FMR), often 0·1–0·2 g g−1
in medium-sized leaves and up to 0·3–0·4 g g−1 in thelargest dissected leaves. As larger leaves have effectivelylonger lever arms, biomechanical considerations forleaf self-support suggest that larger leaves should havea relatively larger fraction of biomass in the mid-rib(Givnish 1979, 1986). However, experimental data to testthis biomechanical principle have been scarce, especiallyfor a large set of species (for review see Niinemets &Sack 2006). Our study does indicate a strong scaling ofFMR with leaf size across a range of species withcontrasting shape and venation (Fig. 5).
Although we hypothesized that elliptic leaves witheffectively longer lever arms have greater FMR, thishypothesis was not supported by the data (Table 3;Fig. 5). In fact, we observed a broad trade-off betweenmid-rib investments and vein density (Fig. 2b). Thistrade-off highlights that lamina support requirementscan be met by either enhanced biomass investmentsin the mid-rib or in minor veins. This is possible asdifferent combinations of total support biomasspartitioning between minor and major veins can yieldsimilar lamina flexural rigidity. Unexpectedly, ellipticleaves were not significantly distinguished from ovateleaves according to this relationship (Fig. 2b), possiblyindicating a greater safety margin against mechanicalfailure in ovate leaves.
In our study, differences in venation architecturewere a more important determinant of differences insupport investments than leaf shape per se. Theoreticalstudies suggest that, for a common biomass investmentin major veins, palmate- and pinnate-veined leaves arehydraulically and mechanically more efficient thanpinnate-veined leaves (Jeje 1985; Kull & Herbig 1995).

38Ü. Niinemets et al.
© 2006 The Authors.Journal compilation© 2006 British Ecological Society, Functional Ecology, 21, 28–40
Although such theoretical studies provide importantinsight into the optimality of venation architecture,such simulations are valid only for specific leaf size/shape combinations, and need to consider higher-orderveins as well.
We observed that parallel/palmate-veined ellipticleaves had a lower vein density at common FMR thanovate and elliptic pinnate-veined leaves (Fig. 2a). As inpinnate-leaved species, the mid-rib also appears tobear the major bending stresses in elongated parallel-and palmate-veined species (Moulia et al. 1994; Moulia& Fournier 1997). However, in our study, parallel/palmate-veined species were generally also less extendedthan pinnate-leaved species (Fig. 2a). Given that theconductance of veins is inversely proportional to thefourth power of their diameter (for review see Tyree &Jarvis 1982), a central vein and high density of minorveins is not apparently an optimal strategy for wide leaves.In fact, as the distance of the lamina margin fromthe central support element increases, leaf hydrauliccharacteristics can be optimized if the biomass investedin major veins is distributed between several veinsradiating from the base, instead of a single central vein.Thus the overall greater density of major veins in lesselongated palmate/parallel-veined species is probablyanother part of the general trade-off between theinvestments in minor and major veins observed for alldata pooled (Fig. 2b).
, ,
Previous analysis suggests that larger leaves have asignificant disadvantage because of disproportionatelylarger support costs. Although support tissues also containchlorophyll and photosynthetic enzymes (Niinemets1999a; Hibberd & Quick 2001), the photosyntheticactivity of support structures is low (Niinemets 1999a).Significantly lower N contents of mid-ribs relative tolamina found in our study also support this suggestion.
Extending a larger fraction of leaf area further fromstem and leaf attachment can compensate for thisdrawback by decreased self-shading and enhancedlight-capture efficiency (Pearcy & Yang 1998). In addition,we observed a negative correlation between the fractionof lamina biomass in mid-rib and lamina dry to freshmass ratio (DF, Fig. 3c). As multiple regression analy-ses demonstrated, DF affected the fractional biomassinvestment in the mid-rib independently of leaf size. Asincreases in DF result mainly from a greater fraction ofcell walls, DF is negatively correlated with leaf physiologicalactivity (Garnier & Laurent 1994). The uniform negativecorrelation between DF and lamina nitrogen content(Fig. 3a) further underscores this pattern, and suggeststhat a larger mid-rib mass fraction is combined withgreater physiological activity of the rest of the lamina.In addition, DF was positively, and the biomass fractionin the mid-rib negatively, associated with leaf longevity(Fig. 4). These relationships are consistent with the
large body of information of lower functional activity oflong-lasting leaves (Reich et al. 1992, 1999; Wright et al.2004). The negative relationship between longevity andFMR suggests that lifetime support tissue investment islarger in short-lived leaves. This pattern is apparentlycounterintuitive, but emphasizes the large supportbiomass cost of high functional leaf activity.
Turgor (the pressure exerted by cellular water on cellwalls) can contribute strongly to leaf support in specieswith thin cell walls and high leaf water content (Niklas1986, 1989). As species with lower DF are expected to relymore strongly on turgor for mechanical support, wesuggested that, among the leaves of common mass, areaand shape, dry mass investment in the mid-rib is lowerfor leaves with lower DF. However, the negativecorrelation between DF and the biomass fraction in themid-rib suggests that reliance on turgor for mechanicalstability does not necessarily mean lower biomassinvestments in support. Given that leaves with highphysiological activity also have greater water loss viatranspiration than plants with lower physiological activity(for correlations between leaf structure and stomatalconductance see Schulze et al. 1994; Reich et al. 1997),rapid water loss must be accompanied by rapid watersupply for turgor maintenance. Although plants withhigher water content can rely more heavily on turgorfor mechanical stability, our data suggest that mechan-ical support through turgor is not necessarily a cheapsolution in terms of biomass investment in support.
The negative relationship between FMR and DF issurprising at first glance, but may provide a majormeans by which large and structurally expensive leavesoffset the high cost of leaf construction. In this regard,it is interesting that there are well known positiverelationships between site nutrient availability andaverage leaf size of dominant species (Givnish 1978).Our data suggest that this general worldwide relationshipmay reflect an enhanced within-leaf support cost oflarge-leaved species, and accordingly inherent require-ments for high soil nutrient availabilities to achievehigh foliage physiological capacities that allow theleaves to pay back the costs of their construction.
Previously, it has been demonstrated that large-leavedspecies generally have relatively lower stem xylem cross-sectional areas for a common leaf area supported (Pickupet al. 2005). If so, this could provide an additionalstrategy to compensate for high within-leaf supportcosts. Although we investigated mostly herbs in ourstudy, it is interesting that in several of the large-leavedspecies studied, the petioles and stems are hollow (e.g.Arctium tomentosum, C. pepo, H. sosnowskyi, L. wilsoniana,Rumex crispus, Tussilago farfara), suggesting that theremay be a general trade-off between within-leaf and within-plant support investments (Niinemets et al. 2006b).
Conclusions
There is extensive variation in within-plant supportinvestments among plant species (Valladares et al.

39Leaf shape and support costs
© 2006 The Authors.Journal compilation© 2006 British Ecological Society, Functional Ecology, 21, 28–40
2002; Niinemets et al. 2006b). Our study further highlightsa dramatic, more than eightfold variation in within-leaf support costs among leaves of different shape, sizeand venation, and an important trade-off betweeninvestments in minor and major veins, suggesting thatleaves of different size and shape are characterized bydifferent combinations of mid-rib investment and minorvein density. This quantitative gradient in investmentsin minor vs major veins has important implications forleaf physiological activity and hydraulic efficiency. Whilethere appear to be fundamental scaling relationshipsamong leaf physiological and structural characteristics(Wright et al. 2004), we suggest that, among leaves ofcommon integrated characteristics such as leaf Ncontent and DM per unit area, partitioning of leafhydraulic conductance between minor and major veinsand overall investments in support may lead to a gradationof foliar physiological activity at specific points alongthe continuous spectrum of leaf functioning. Thisspectrum of vein density vs biomass investment inmid-rib may have important implications for nichedifferentiation of coexisting species.
Acknowledgements
We thank the Estonian Science Foundation (grant4584), the Estonian Ministry of Education and Science(grant 0182468As03), and the Estonian Academy ofSciences for financial support.
References
Ackerly, D.D. & Reich, P.B. (1999) Convergence and correlationsamong leaf size and function in seed plants: a comparativetest using independent contrasts. American Journal ofBotany 86, 1272–1281.
Baxes, G. (1994) Digital Image Processing: Principles andApplications. John Wiley and Sons, New York.
Dengler, N.G. & Kang, J. (2001) Vascular patterning and leafshape. Current Opinion in Plant Biology 4, 50–56.
Diemer, M. (1998) Life span and dynamics of leavesof herbaceous perennials in high-elevation environments:‘news from the elephant’s leg’. Functional Ecology 12, 413–425.
FAO, ISRIC & ISSS (1998) World Reference Base for SoilResources. Food and Agriculture Organization of the UN(with International Society of Soil Science and InternationalSoil Reference and Information Centre), Rome.
Garnier, E. & Laurent, G. (1994) Leaf anatomy, specific massand water content in congeneric annual and perennialgrass species. New Phytologist 128, 725–736.
Gere, J.M. & Timoshenko, S.P. (1997) Mechanics of Materials,4th edn. PWS Publishing, Boston, MA, USA.
Givnish, T.J. (1978) Ecological aspects of plant morphology:leaf form in relation to environment. Acta Biotheoretica 27,83–142.
Givnish, T. (1979) On the adaptive significance of leaf form.Topics in Plant Population Biology (eds O.T. Solbrig, S.Jain, G.B. Johnson & P.H. Raven), pp. 375–407. ColumbiaUniversity Press, New York.
Givnish, T.J. (1984) Leaf and canopy adaptations in tropicalforests. Physiological Ecology of Plants of the Wet Tropics.(eds E. Medina, H.A. Mooney, C. Vásquez-Yánes &
H.A. Mooney), pp. 51–84. Proceedings of an InternationalSymposium Held in Oxatepec and Los Tuxtlas, Mexico, 29June–6 July 1983. Dr W. Junk, The Hague.
Givnish, T.J. (1986) Biomechanical constraints on crowngeometry in forest herbs. On the Economy of Plant Formand Function (ed. T.J. Givnish), pp. 525–583. Proceedingsof the Sixth Maria Moors Cabot Symposium, ‘EvolutionaryConstraints on Primary Productivity: Adaptive Patterns ofEnergy Capture in Plants’, Harvard Forest, August 1983.Cambridge University Press, Cambridge, UK.
Givnish, T.J. (1987) Comparative studies of leaf form: assessingthe relative roles of selective pressures and phylogeneticconstraints. New Phytologist 106, 131–160.
Hibberd, J.M. & Quick, W.P. (2001) Characteristics of C4
photosynthesis in stems and petioles of C3 flowering plants.Nature 415, 451–454.
Jeje, A.A. (1985) The flow and dispersion of water in the vascularnetwork of dicotyledonous leaves. Biorheology 22, 285–302.
Kessler, S. & Sinha, N. (2004) Shaping up: the genetic controlof leaf shape. Current Opinion in Plant Biology 7, 65–72.
Kõlli, R. & Ellermäe, O. (2003) Humus status of postlithogenicarable mineral soils. Agronomy Research 1, 161–174.
Kull, U. & Herbig, A. (1995) Das Blattadersystem derAngiospermen: Form und Evolution. Naturwissenschaften82, 441–451.
Moulia, B. & Fournier, M. (1997) Mechanics of the maizeleaf: a composite beam model of the midrib. Journal ofMaterials Science 32, 2771–2780.
Moulia, B., Fournier, M. & Guitard, D. (1994) Mechanicsand form of the maize leaf: in vivo qualification of flexuralbehaviour. Journal of Materials Science 29, 2359–2366.
Niinemets, Ü. (1999a) Differences in chemical compositionrelative to functional differentiation between petioles andlaminas of Fraxinus excelsior. Tree Physiology 19, 39–45.
Niinemets, Ü. (1999b) Research review. Components of leafdry mass per area – thickness and density – alter leafphotosynthetic capacity in reverse directions in woodyplants. New Phytologist 144, 35–47.
Niinemets, Ü. (2004) Adaptive adjustments to light in foliageand whole-plant characteristics depend on relative age inthe perennial herb Leontodon hispidus. New Phytologist162, 683–696.
Niinemets, Ü. & Fleck, S. (2002) Petiole mechanics, leafinclination, morphology, and investment in support inrelation to light availability in the canopy of Liriodendrontulipifera. Oecologia 132, 21–33.
Niinemets, Ü. & Kull, O. (1998) Stoichiometry of foliarcarbon constituents varies along light gradients in temperatewoody canopies: implications for foliage morphologicalplasticity. Tree Physiology 18, 467–479.
Niinemets, Ü. & Sack, L. (2006) Structural determinantsof leaf light-harvesting capacity and photosynthetic poten-tials. Progress in Botany Vol. 67 (eds K. Esser, U.E. Lüttge,W. Beyschlag & J. Murata), pp. 385–419. Springer Verlag,Berlin.
Niinemets, Ü., Kull, O. & Tenhunen, J.D. (1999) Variabilityin leaf morphology and chemical composition as a functionof canopy light environment in co-existing trees. InternationalJournal of Plant Sciences 160, 837–848.
Niinemets, Ü., Sonninen, E. & Tobias, M. (2004) Canopygradients in leaf intercellular CO2 mole fractions revisited:interactions between leaf irradiance and water stress needconsideration. Plant, Cell & Environment 27, 569–583.
Niinemets, Ü., Cescatti, A., Rodeghiero, M. & Tosens, T.(2006a) Complex adjustments of photosynthetic capacityand internal mesophyll conductance to current andprevious light availabilities and leaf age in Mediterraneanevergreen species Quercus ilex. Plant, Cell & Environment29, 1159–1178.

40Ü. Niinemets et al.
© 2006 The Authors.Journal compilation© 2006 British Ecological Society, Functional Ecology, 21, 28–40
Niinemets, Ü., Portsmuth, A. & Tobias, M. (2006b) Leaf sizemodifies support biomass distribution between stems,petioles and mid-ribs in temperate plants. New Phytologist171, 91–104.
Niklas, K.J. (1986) Biomechanical responses of chive (Alliumschoenoprasum var. shoenoprasum) leaves to changes inwater potential. American Journal of Botany 73, 636–637.
Niklas, K.J. (1989) Mechanical behavior of plant tissues asinferred from the theory of pressurized cellular solids.American Journal of Botany 76, 929–937.
Olff, H. (1992) Effects of light and nutrient availability on drymatter and N allocation in six successional grasslandspecies. Testing for resource ratio effects. Oecologia 89,412–421.
Pearcy, R.W. & Yang, W. (1998) The functional morphologyof light capture and carbon gain in the redwood forestunderstorey plant, Adenocaulon bicolor Hook. FunctionalEcology 12, 543–552.
Pearcy, R.W., Muraoka, H. & Valladares, F. (2005) Crownarchitecture in sun and shade environments: assessingfunction and trade-offs with a three-dimensional simulationmodel. New Phytologist 166, 791–800.
Pickup, M., Westoby, M. & Basden, A. (2005) Dry mass costsof deploying leaf area in relation to leaf size. FunctionalEcology 19, 88–97.
Poorter, H. (1994) Construction costs and payback time ofbiomass: a whole plant perspective. A Whole Plant Perspectiveon Carbon–Nitrogen Interactions (eds J. Roy & E. Garnier),pp. 111–127. SPB Academic Publishing, The Hague.
Poorter, H. & Evans, J.R. (1998) Photosynthetic nitrogen-useefficiency of species that differ inherently in specific leafarea. Oecologia 116, 26–37.
Reich, P.B., Walters, M.B. & Ellsworth, D.S. (1992) Leaf life-span in relation to leaf, plant, and stand characteristicsamong diverse ecosystems. Ecological Monographs 62,365–392.
Reich, P.B., Kloeppel, B.D., Ellsworth, D.S. & Walters, M.B.(1995) Different photosynthesis–nitrogen relations indeciduous hardwood and evergreen coniferous tree species.Oecologia 104, 24–30.
Reich, P.B., Walters, M.B. & Ellsworth, D.S. (1997) Fromtropics to tundra: global convergence in plant functioning.Proceedings of the National Academy of Sciences, USA 94,13730–13734.
Reich, P.B., Ellsworth, D.S., Walters, M.B., Vose, J.M.,Gresham, C., Volin, J.C. & Bowman, W.D. (1999) Generalityof leaf trait relationships: a test across six biomes. Ecology80, 1955–1969.
Roth-Nebelsick, A., Uhl, D., Mosbrugger, V. & Kerp, H.(2001) Evolution and function of leaf venation architecture:a review. Annals of Botany 87, 553–566.
Sack, L. & Frole, K. (2006) Leaf structural diversity is relatedto hydraulic capacity in tropical rain forest trees. Ecology87, 483–491.
Sack, L. & Holbrook, N.M. (2006) Leaf hydraulics. AnnualReview in Plant Biology 57, 361–381.
Sack, L., Cowan, P.D. & Holbrook, N.M. (2003a) The majorveins of mesomorphic leaves revisited: testing for conductiveoverload in Acer saccharum (Aceraceae) and Quercus rubra(Fagaceae). American Journal of Botany 90, 32–39.
Sack, L., Cowan, P.D., Jaikumar, N. & Holbrook, N.M.(2003b) The ‘hydrology’ of leaves: co-ordination of structureand function in temperate woody species. Plant, Cell &Environment 26, 1343–1356.
Sack, L., Streeter, C.M. & Holbrook, N.M. (2004) Hydraulicanalysis of water flow through leaves of sugar maple andred oak. Plant Physiology 134, 1824–1833.
Sack, L., Tyree, M.T. & Holbrook, N.M. (2005) Leaf hydraulicarchitecture correlates with regeneration irradiance intropical rainforest trees. New Phytologist 167, 403–413.
Schulze, E.D., Kelliher, F.M., Körner, C., Lloyd, J. &Leuning, R. (1994) Relationships among maximum stomatalconductance, ecosystem surface conductance, carbonassimilation rate, and plant nitrogen nutrition: a globalecology scaling exercise. Annual Review of Ecology andSystematics 25, 629–660.
Sisó, S., Camarero, J.J. & Gil-Pelegrín, E. (2001) Relation-ship between hydraulic resistance and leaf morphology inbroadleaf Quercus species: a new interpretation of leaflobation. Trees: Structure and Function 15, 341–345.
Sokal, R.R. & Rohlf, F.J. (1995) Biometry. The Principles andPractice of Statistics in Biology Research, 3rd edn. W.H.Freeman, New York.
Terashima, I., Araya, T., Miyazawa, S.-I., Sone, K. & Yano, S.(2005) Construction and maintenance of the optimal photo-synthetic systems of the leaf, herbaceous plant and tree:an eco-developmental treatise. Annals of Botany 95, 507–519.
Tessier, J.T. (2004) Leaf longevity of Oxalis acetosella(Oxalidaceae) in the Catskill Mountains, New York, USA.American Journal of Botany 91, 1371–1377.
Tyree, M.T. & Jarvis, P.G. (1982) Water in tissues and cells.Physiological Plant Ecology, Vol. II: Water Relations andCarbon Assimilation (eds P.S. Nobel, C.B. Osmond &H. Ziegler), pp. 35–77. Springer-Verlag, Berlin.
Valladares, F., Skillman, J.B. & Pearcy, R.W. (2002)Convergence in light capture efficiencies among tropicalforest understory plants with contrasting crown architectures:a case of morphological compensation. American Journalof Botany 89, 1275–1284.
Winn, A.A. & Evans, A.S. (1991) Variation among populationsof Prunella vulgaris L. in plastic responses to light.Functional Ecology 5, 562–571.
Wright, I.J., Reich, P.B., Westoby, M. et al. (2004) The world-wide leaf economics spectrum. Nature 428, 821–827.
Zwieniecki, M.A., Melcher, P.J., Boyce, C.K., Sack, L. &Holbrook, N.M. (2002) Hydraulic architecture of leafvenation in Laurus nobilis L. Plant, Cell & Environment 25,1445–1450.
Received 27 July 2006; accepted 29 September 2006 Editor: Ken Thompson
Copyright © 2022 FDOKUMEN




















