
Ecological management of a Mediterranean mountainous reserve (Pindos National Park, Greece) using...
13
Journal for Nature Conservation 17 (2009) 47—59 Ecological management of a Mediterranean mountainous reserve (Pindos National Park, Greece) using the bird community as an indicator Vassiliki Kati a, , Panayotis Dimopoulos a , Haritakis Papaioannou a,b , Kostas Poirazidis c a Department of Environmental and Natural Resources Management, University of Ioannina, Seferi 2, 30100 Agrinio, Greece b Center of Biological and Cultural Diversity, 44004 Papingo, Greece c Department of Forestry, Environment and Natural Resources, Democritus University of Thrace, 68200 Orestiada, Greece Received 23 April 2008; accepted 5 December 2008 KEYWORDS Ecological structure; Forest management; Indicators; NATURA 2000; Passerines; Protected area Summary We used the community of passerines and woodpeckers as a target group for the conservation management of Pindos National Park (NW Greece). We conducted bird point counts twice during springtime in 72 plots that represented the main vegetation types (16 sites). We recorded 56 species (14 of conservation concern-SPEC). The montane grasslands were the most important habitats in terms of species of conservation concern, whereas the agricultural mosaics were the most species-rich habitats. The mixed pine-beech woods were significantly richer than the pinewoods, whereas pinewoods and broad-leaved woods did not differ significantly between them. The bird diversity was significantly correlated with the number of tree layers, the vertical structural complexity and the maximum height of trees. The presence of grassland, forest and agricultural habitat type, as well as the altitude and the vegetation structural complexity were the main environmental parameters determining species composition (Canonical Correspon- dence Analysis). We identified a set of 17 typical species (IndVal analysis) to be used in the monitoring scheme of the Park, which were characteristic of the main bird habitat types distinguished by Ward’s hierarchical clustering. Conservation measures should involve maintenance of the traditional agricultural practices, montane ARTICLE IN PRESS www.elsevier.de/jnc 1617-1381/$ - see front matter & 2008 Elsevier GmbH. All rights reserved. doi:10.1016/j.jnc.2008.12.002 Corresponding author. Tel.: +3026510 70993; fax: +302641033716. E-mail addresses: [email protected] , [email protected] (V. Kati).
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Ecological management of a Mediterranean mountainous reserve (Pindos National Park, Greece) using...

ARTICLE IN PRESS
Journal for Nature Conservation 17 (2009) 47—59
1617-1381/$ - sdoi:10.1016/j.
�CorrespondE-mail addr
www.elsevier.de/jnc
Ecological management of a Mediterraneanmountainous reserve (Pindos National Park,Greece) using the bird community as an indicator
Vassiliki Katia,�, Panayotis Dimopoulosa, Haritakis Papaioannoua,b,Kostas Poirazidisc
aDepartment of Environmental and Natural Resources Management, University of Ioannina,Seferi 2, 30100 Agrinio, GreecebCenter of Biological and Cultural Diversity, 44004 Papingo, GreececDepartment of Forestry, Environment and Natural Resources,Democritus University of Thrace, 68200 Orestiada, Greece
Received 23 April 2008; accepted 5 December 2008
KEYWORDSEcological structure;Forest management;Indicators;NATURA 2000;Passerines;Protected area
ee front matter & 2008jnc.2008.12.002
ing author. Tel.: +30 265esses: [email protected],
SummaryWe used the community of passerines and woodpeckers as a target group for theconservation management of Pindos National Park (NW Greece). We conductedbird point counts twice during springtime in 72 plots that represented themain vegetation types (16 sites). We recorded 56 species (14 of conservationconcern-SPEC). The montane grasslands were the most important habitats interms of species of conservation concern, whereas the agricultural mosaics were themost species-rich habitats. The mixed pine-beech woods were significantlyricher than the pinewoods, whereas pinewoods and broad-leaved woods did notdiffer significantly between them. The bird diversity was significantly correlatedwith the number of tree layers, the vertical structural complexity and the maximumheight of trees. The presence of grassland, forest and agricultural habitat type, aswell as the altitude and the vegetation structural complexity were the mainenvironmental parameters determining species composition (Canonical Correspon-dence Analysis). We identified a set of 17 typical species (IndVal analysis) to be usedin the monitoring scheme of the Park, which were characteristic of the main birdhabitat types distinguished by Ward’s hierarchical clustering. Conservation measuresshould involve maintenance of the traditional agricultural practices, montane
Elsevier GmbH. All rights reserved.
10 70993; fax: +30 [email protected] (V. Kati).

ARTICLE IN PRESS
V. Kati et al.48
grasslands, old growth woods, as well as the vertical vegetation complexity and hightrees in forest stands.& 2008 Elsevier GmbH. All rights reserved.
Introduction
Birds exhibit a diverse range of ecologicalfunctions, and benefit humans by providing a widerange of ecosystem services (Sekercioglu, 2006).However, birds also constitute one of the mostvulnerable groups of organisms, with many speciescurrently undergoing significant global populationdeclines, leading to detrimental consequencesfor total biodiversity and ecosystem function(Sekercioglu et al., 2004).
Because birds are, especially in Europe, amongthe best-studied organisms, they are often used toevaluate the impact of human activities (BirdLifeInternational, 2004; Heath et al., 2000). Onaccount of their ubiquitousness, terrestrial passer-ine communities in particular have been used asindicators of environmental change (Bani et al.,2005; Gregory et al., 2004), or as ecologicalsubstitutes for other, more difficult to quantifygroups of organisms (Howard et al., 1998; Katiet al., 2004a; Lombard, 1995; Prendergast et al.,1993). Although their use as indicator taxa has beensometimes criticised for its potentially low extra-polative value to other groups of organisms(Lindenmayer et al., 2006; Simberloff, 1998), smallterrestrial birds are considered as an importantcomponent of biodiversity and have been inte-grated into studies evaluating its conservation(Dobson et al., 1997; Kati et al., 2004b; Lawtonet al., 1998; Thiollay, 2002; Vessby et al., 2002).Thus, small terrestrial birds have often been usedto assess and guide ongoing forest managementpractices (e.g. Donald et al., 1998; Gil-Tena et al.,2007; Johnson & Freedman, 2002; King & DeGraaf,2000; Laiolo et al., 2003; Lance & Phinney, 2001;Muller et al., 2007; Sekercioglu, 2002) or currentagricultural practices (e.g. Chamberlain et al.,2000; Donald et al., 2001; Pain & Pienkowski,1997). In this applied context, the European Unionhas recently adopted, for the first time, anecological index that considers the conservationstatus of common farmland birds (Farmland BirdIndex) as a formal indicator of sustainable devel-opment in Europe (structural-sustainable develop-ment indicator). A prerequisite step before usingsmall terrestrial birds as an indicator group is toobtain a deeper understanding of the underlyingpatterns in diversity and the ecological structure ofthis community.
In the present study we conducted an ecologicalstudy, considering the land bird community (passerineand woodpecker species) as a target group, in orderto provide a guideline for the ecological managementof a remote area, surrounding the mountainousPindos (or Valia Calda) National Park in NW Greece.The Park was created in 1966 and is considered to beone of the most important protected areas for themaintenance of mountainous biodiversity and eco-system integrity at national scale. However, existingknowledge on the biological diversity of the arearemains fragmentary (Mertzanis, 1999; Tsounis et al.,1985). The Park is under the administrative authorityof a recently created (2003) Management Body, whichis responsible for the ecological management of amuch larger region that includes eight protectedareas of the NATURA 2000 network. This authoritytogether with the local Forest Service is responsiblefor regulating human activities and maintaining theecological value of Pindos National Park (NP).However, this target remains elusive, due to a lackof expertise and of a science-based management planfor the NP. Thus a great need exists to conductconservation-oriented research that can be trans-lated directly into practical management proposals.The present research attempts to fill this gap inrelation to the bird community. Specifically, ourobjectives are: (a) to assess the relative importanceof different habitat types for avian conservation; (b)to test whether tree composition, vertical structuralcomplexity and tree height affect bird community inforest stands; (c) to analyse the ecological structureof bird communities and to detect the main environ-mental factors regulating species diversity anddistribution; and, (d) to identify species that arecharacteristic for the main bird habitat typesdistinguished. On the applied level, we attempt totranslate these findings into practical monitoring andmanagement schemes for the NP. Our findings couldprovide a management guideline for other nationalparks throughout the Greek mountains and in theMediterranean area.
Methods
Study area and site selection
The study area is located in north-westernGreece (391540N, 211070E) and covers a surface of
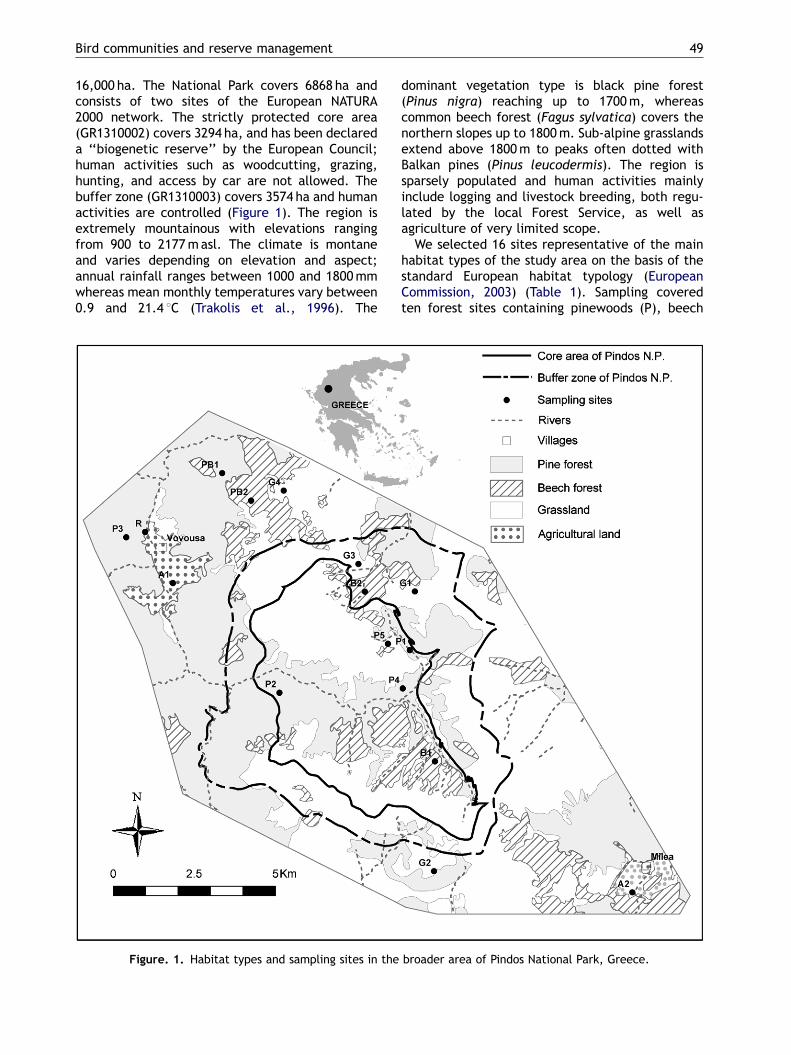
ARTICLE IN PRESS
Bird communities and reserve management 49
16,000 ha. The National Park covers 6868 ha andconsists of two sites of the European NATURA2000 network. The strictly protected core area(GR1310002) covers 3294 ha, and has been declareda ‘‘biogenetic reserve’’ by the European Council;human activities such as woodcutting, grazing,hunting, and access by car are not allowed. Thebuffer zone (GR1310003) covers 3574 ha and humanactivities are controlled (Figure 1). The region isextremely mountainous with elevations rangingfrom 900 to 2177masl. The climate is montaneand varies depending on elevation and aspect;annual rainfall ranges between 1000 and 1800mmwhereas mean monthly temperatures vary between0.9 and 21.4 1C (Trakolis et al., 1996). The
Figure. 1. Habitat types and sampling sites in the
dominant vegetation type is black pine forest(Pinus nigra) reaching up to 1700m, whereascommon beech forest (Fagus sylvatica) covers thenorthern slopes up to 1800m. Sub-alpine grasslandsextend above 1800m to peaks often dotted withBalkan pines (Pinus leucodermis). The region issparsely populated and human activities mainlyinclude logging and livestock breeding, both regu-lated by the local Forest Service, as well asagriculture of very limited scope.
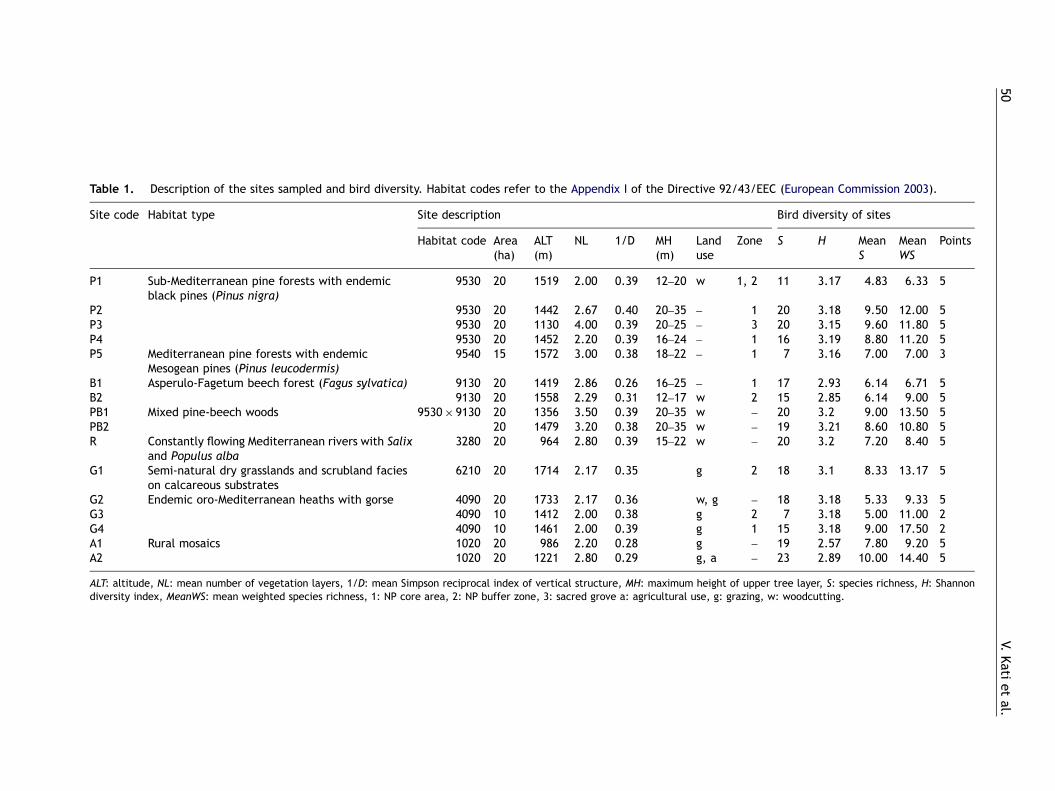
We selected 16 sites representative of the mainhabitat types of the study area on the basis of thestandard European habitat typology (EuropeanCommission, 2003) (Table 1). Sampling coveredten forest sites containing pinewoods (P), beech
broader area of Pindos National Park, Greece.
ARTIC
LEIN
PRES
S
Table 1. Description of the sites sampled and bird diversity. Habitat codes refer to the Appendix I of the Directive 92/43/EEC (European Commission 2003).
Site code Habitat type Site description Bird diversity of sites
Habitat code Area(ha)
ALT(m)
NL 1/D MH(m)
Landuse
Zone S H MeanS
MeanWS
Points
P1 Sub-Mediterranean pine forests with endemicblack pines (Pinus nigra)
9530 20 1519 2.00 0.39 12–20 w 1, 2 11 3.17 4.83 6.33 5
P2 9530 20 1442 2.67 0.40 20–35 – 1 20 3.18 9.50 12.00 5P3 9530 20 1130 4.00 0.39 20–25 – 3 20 3.15 9.60 11.80 5P4 9530 20 1452 2.20 0.39 16–24 – 1 16 3.19 8.80 11.20 5P5 Mediterranean pine forests with endemic
Mesogean pines (Pinus leucodermis)9540 15 1572 3.00 0.38 18–22 – 1 7 3.16 7.00 7.00 3
B1 Asperulo-Fagetum beech forest (Fagus sylvatica) 9130 20 1419 2.86 0.26 16–25 – 1 17 2.93 6.14 6.71 5B2 9130 20 1558 2.29 0.31 12–17 w 2 15 2.85 6.14 9.00 5PB1 Mixed pine-beech woods 9530� 9130 20 1356 3.50 0.39 20–35 w – 20 3.2 9.00 13.50 5PB2 20 1479 3.20 0.38 20–35 w – 19 3.21 8.60 10.80 5R Constantly flowing Mediterranean rivers with Salix
and Populus alba3280 20 964 2.80 0.39 15–22 w – 20 3.2 7.20 8.40 5
G1 Semi-natural dry grasslands and scrubland facieson calcareous substrates
6210 20 1714 2.17 0.35 g 2 18 3.1 8.33 13.17 5
G2 Endemic oro-Mediterranean heaths with gorse 4090 20 1733 2.17 0.36 w, g – 18 3.18 5.33 9.33 5G3 4090 10 1412 2.00 0.38 g 2 7 3.18 5.00 11.00 2G4 4090 10 1461 2.00 0.39 g 1 15 3.18 9.00 17.50 2A1 Rural mosaics 1020 20 986 2.20 0.28 g – 19 2.57 7.80 9.20 5A2 1020 20 1221 2.80 0.29 g, a – 23 2.89 10.00 14.40 5
ALT: altitude, NL: mean number of vegetation layers, 1/D: mean Simpson reciprocal index of vertical structure, MH: maximum height of upper tree layer, S: species richness, H: Shannondiversity index, MeanWS: mean weighted species richness, 1: NP core area, 2: NP buffer zone, 3: sacred grove a: agricultural use, g: grazing, w: woodcutting.
V.Kati
etal.
50

ARTICLE IN PRESS
Bird communities and reserve management 51
woods (B), mixed woods (PB) and riverine vegeta-tion (R), as well as four grassland sites (G) and twoagricultural ones (A) (Table 1). Most of the sitessampled were located inside areas protected by theNATURA 2000 network, and one site (P3) wasstrictly protected as a sacred grove.
Bird sampling
We surveyed the bird communities at 72 pointlocations, recording all passerine and woodpeckerspecies seen or heard within a circle of a 100mradius of each point. Points were located at adistance of 200m between them and at least 100mfrom site edge, in order to avoid double countingand edge effects. All sites (20 ha) were representedby five points with the exception of three smallersites (10–15 ha) that were covered with 2–3 points.To cover both sedentary species and late-arrivingmigrants, we sampled every site twice, once in latespring (30 April–30 May 2003) and once in earlysummer (5 June–5 July 2003). In our bird countmethodology (see Bibby et al., 1992; Blondel et al.,1970), a singing, territorial male represented abreeding pair and was therefore counted as twoindividuals. Every call different from the malebreeding song was counted as one individual. Eachcount lasted for 10min and took place between30min before and four hours after sunrise.
Measurement of environmental variables
We located a smaller quadrate (50m� 50m) atthe centre of the each one of the 72 bird surveypoints where we sampled the four main vegetationlayers: (1) upper tree layer (47m); (2) lower treelayer (4.1–7m); (3) upper shrub layer (2.1–4m); (4)lower shrub layer (0.5–2m) (Kent & Coker, 1994).We visually determined the percentage cover(relative area occupied by the vertical projectionof all aerial parts of woody plants as a percentageof the surface area of the sample plot) for theseparate layers (van der Maarel, 2005), using thefollowing vegetation cover classes (Kuchler, 1988)1 ¼ 1–5%, 2 ¼ 6–25%, 3 ¼ 26–50%, 4 ¼ 51–75%,5 ¼ 76–100%. For each quadrate we recorded thealtitude (ALT), the overall vegetation cover (COV),the maximum height of the upper tree layer (MH) inforest habitats, the number of vegetation layers(NL) and the reciprocal of Simpson’s diversity index(1/D) as a measure of vertical vegetation complex-ity (Magurran, 2004). We also noted the generalhabitat category the station referred to (agricul-ture, forest, and grassland).
Data analysis
We estimated the overall number of species ofthe study area using the software EstimateS(Colwell 2006) with the option of 1000 randomisa-tions. We opted for non-parametric species esti-mators, which consider mainly the presence of rarespecies, recorded by one or two individuals(Chao 1), or present in one or two point counts(Chao 2) (Magurran, 2004). Species diversity wasestimated in terms of species richness (S),weighted species richness (WS), and Shannon–Wei-ner index (H) (Magurran, 2004), using the maximumabundance recorded over the two seasons. We alsocalculated their mean values per sampling point(MeanS, Mean WS) and we compared them forthe pine, broad-leaved and mixed woods using theKruskal–Wallis non-parametric test and then theMann–Whitney tests with Bonferroni correction,with the help of SPSS software (Field, 2005).Weighted species richness is the species richnessof the site, with each species having a differentweight based on its conservation status (SPECcategory) (BirdLife International, 2004). We as-signed a standard weight (w ¼ 1) to the species offavourable conservation status (SPEC 4 category ornon-SPEC category). We assigned double weight(w ¼ 2) to the species of SPEC 3 category, which arenot concentrated in Europe but have an unfavour-able conservation status. Finally we assigned aquadruple weight (w ¼ 4) to the species in the SPEC2 category, which are concentrated in Europe andhave an unfavourable conservation status and tothe species listed in Annex I of the EuropeanDirective 79/409 EU, which includes all bird speciessubject to special conservation measures inEurope. We also calculated the bird diversityindices (S, WS, H) for each point count andtested their correlation with the five quantitativeenvironmental variables sampled using Spearmancorrelation coefficients (SPSS, vers 15.).
To assess which environmental factors shape birdcommunity composition, we used the CanonicalCorrespondence Analysis (CCA) option from theordination program CANOCO (ter Braak & Smilauer,2002), because the length of the gradients ofDetrended Correspondence Analysis (DCA) indi-cated a heterogeneous dataset (Leps & Smilauer,2005). This method extracts the major gradients inthe data that are accounted for by the measuredvariables. The position of a species in the resultingplot indicates the characteristics of the ecologicaloptimum for this taxon; its abundance will de-crease with increasing distance from this point.Species encountered only once were not includedin the analysis and species abundances were log

ARTICLE IN PRESS
V. Kati et al.52
transformed. A forward selection procedure using aMonte-Carlo permutation test with 1000 iterationswas used to select and present in the CCA diagramonly the significant (po0.05) environmental vari-ables that explained variation in community struc-ture. The multi-collinearity of environmentalvariables was tested using the inflation factor.
We used Ward’s clustering method to grouphierarchically the point counts into clusters,expressing the different habitat types of birdcommunity. We also used the indicator valueprocedure (IndVal) (Dufrene & Legendre, 1997) toidentify the typical species characterising each ofthe clusters. IndVal is a percentage that rangesbetween 0 and 100 and takes its maximum valuewhen the species is present exclusively in all sitesof a single cluster. All calculations were carried outusing IndVal software (Dufrene, 1999). A speciesis considered to be a ‘‘symmetrical indicator’’(IndVal4 50%) for one cluster, when it is presentin 470% of the sites of the cluster and when 470%of its individuals occur in the cluster. A randomreallocation procedure (1000 iterations) of sitesamong site groups was used to test IndVal sig-nificance (alpha ¼ 0.05).
Table 2. Spearman correlation coefficients betweenbird diversity indices of point counts and environmentalparameters.
Diversity indices Environmental parameters
NL 1/D MH ALT
S 0.385� – 0.541� –0.305�
WS 0.219�� – 0.790� –
H – 0.637� –
ALT: altitude, NL: number of vegetation layers, 1/D: Simpsonreciprocal index of vertical structure, MH: maximum height ofupper tree layer, S: species richness, WS: weighted speciesrichness, H: Shannon diversity index.�Po0.01.��Po0.05.
Results
Point count sampling resulted in 56 species (14 ofconservation concern), whereas the overall birdspecies number recorded inside and outside pointcounts was 62 species, including six wood-peckers (Piciformes) and 56 passerine species(Passeriformes) (Appendix I). The ornithologicalimportance of the study area is high, given thatwe recorded six and ten species of unfavourableconservation status concentrated in Europe (SPEC 2)or not (SPEC 3), respectively (Appendix I). Theoverall estimated species richness using non-para-metric estimators was between 56 (Chao 1 estima-tor) and 66 species (Chao 2 estimator). We havetherefore sampled exhaustively the bird commu-nity of the area.
Bird diversity
The most important site for bird conservation, onthe basis of the criterion of mean weighted speciesrichness (MeanWS) was the montane grassland G4(Table 1). Although not very species-rich (15species), it held seven out of the 16 species ofconservation concern (SPEC 2: Carduelis cannabi-na, Emberiza hortulana, Lullula arborea; SPEC 3:Anthus campestris, Lanius collurio, Monticola
saxatilis, Oenanthe oenanthe). The second mostimportant site, which was also the richest inspecies number was a traditionally cultivated site(A2), including six out of the 16 species ofconservation concern (C. cannabina, E. hortulana,L. collurio, Milaria calandra, Muscicapa striata,Passer domesticus). The next sites in rank was amixed pine-beech forest (PB1), a montane grass-land (G1), a black pine forest located in the corearea of the NP that has not been harvested for atleast two centuries and is accessible exclusively onfoot (P2), and a sacred grove of black pines that hasalso not been harvested for at least two centuries(P3) (Table 1). The species richness and theweighted species richness were significantly higherin the mixed pine-beech woods than in the pine-woods (Kruskall Wallis test p ¼o0.05, Mann–Whitneypost-hoc tests po0.0160), whereas pinewoodsand broad-leaved woods did not differ significantlybetween them.
Community structure and typical species
Bird species richness in each point count (S)decreased significantly (po0.01) with increasingaltitude (Table 2). Bird species richness togetherwith the weighted species richness in each pointcount (WS) increased significantly (po0.01) withthe number of vegetation layers (NL) and with themaximum height of upper tree layer (MH) in theforested sites. Finally, the Shannon diversity indexfor birds in each point count (H) was significantly(po0.01) correlated with the vertical structuralcomplexity as expressed by the Simpson reciprocalindex 1/D (Table 2).
The CCA model was significant (po0.001) andrevealed clearly that the presence of grassland,forest or agricultural habitat type, as well asaltitude and vegetation structural complexity
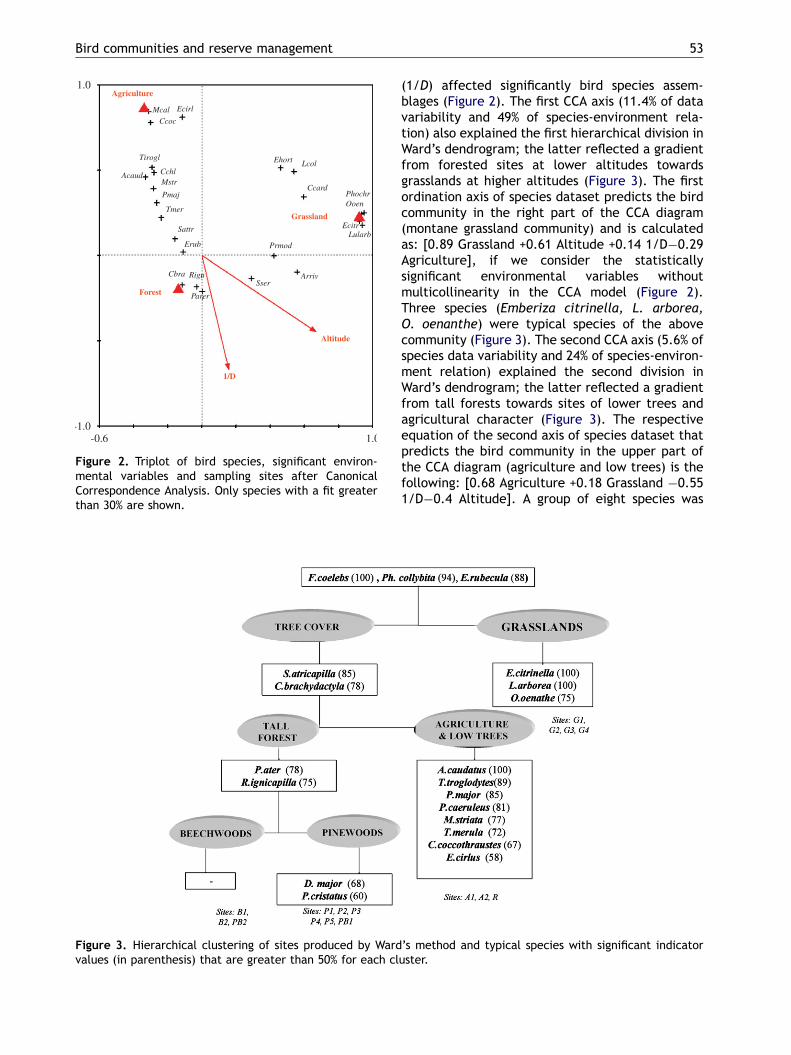
ARTICLE IN PRESS
Figure 3. Hierarchical clustering of sites produced by Wardvalues (in parenthesis) that are greater than 50% for each cl
1.0
-1.0-0.6 1.0
Agriculture
Mcal Ecirl
Tirogl
Acaud CchlMstr
Pmaj
Tmer
Sattr
Erub
Cbra Rign
ParerForest
1/D
Altitude
Grassland
SserArriv
EcitrLularb
PhochrOoen
Prmod
Ccard
LcolEhort
Ccoc
Figure 2. Triplot of bird species, significant environ-mental variables and sampling sites after CanonicalCorrespondence Analysis. Only species with a fit greaterthan 30% are shown.
Bird communities and reserve management 53
(1/D) affected significantly bird species assem-blages (Figure 2). The first CCA axis (11.4% of datavariability and 49% of species-environment rela-tion) also explained the first hierarchical division inWard’s dendrogram; the latter reflected a gradientfrom forested sites at lower altitudes towardsgrasslands at higher altitudes (Figure 3). The firstordination axis of species dataset predicts the birdcommunity in the right part of the CCA diagram(montane grassland community) and is calculatedas: [0.89 Grassland +0.61 Altitude +0.14 1/D�0.29Agriculture], if we consider the statisticallysignificant environmental variables withoutmulticollinearity in the CCA model (Figure 2).Three species (Emberiza citrinella, L. arborea,O. oenanthe) were typical species of the abovecommunity (Figure 3). The second CCA axis (5.6% ofspecies data variability and 24% of species-environ-ment relation) explained the second division inWard’s dendrogram; the latter reflected a gradientfrom tall forests towards sites of lower trees andagricultural character (Figure 3). The respectiveequation of the second axis of species dataset thatpredicts the bird community in the upper part ofthe CCA diagram (agriculture and low trees) is thefollowing: [0.68 Agriculture +0.18 Grassland �0.551/D�0.4 Altitude]. A group of eight species was
’s method and typical species with significant indicatoruster.

ARTICLE IN PRESS
V. Kati et al.54
identified as typical for the above community,increasing in abundance along the second axis(Aegithaulus caudatus, Troglodytes troglodytes,Parus major, Parus caerulaeus, M. striata, Turdusmerula, Coccothraustes coccothraustes, Emberizacirlus) (Figure 3). On the other hand, the relativeabundance of two typical forest species (Parus ater,Regulus ignicapilla), decreased along this axis.Furthermore, two typical species for pinewoodswere identified (Dendrocopos major, Paruscristatus) whereas no typical species existedfor beech woods. Finally, we found that somespecies were generalists, being common through-out the study area and presenting their highestindicator value for all the sites sampled (i.e.Fringilla coelebs, Phylloscopus collybita, Erithacusrubecula), whereas two forest species were generalindicators for all sites of forest character (Sylviaatricapilla, Certhia brachydactyla) (Figure 3).
Discussion
Community structure
The presence of agricultural land, grasslands orforests, as well as altitude and vegetation verticalcomplexity were the main statistically significantenvironmental gradients that influence the distri-bution of bird community in the broader area ofPindos National Park. The bird community wasdistributed along a gradient from forested habitatsat lower altitudes towards montane grasslands athigher altitudes and along a gradient from foreststands with high trees and greater vertical struc-tural complexity towards lower trees and culti-vated land. These results are in agreement withother studies in the Mediterranean area (e.g. Dıaz,2006; Kati & Sekercioglu, 2006; Prodon & Lebreton,1981). Birds distinguish three general bird habitattypes in the study area: grasslands; shrubbyhabitats of agricultural land or riverine vegetation;and forest habitats of pinewoods or beech woods,thus perceiving the environment at a relativelycoarse scale.
Typical species
Our study offers a valuable conservation tool forthe management and monitoring plan of the studyarea, as it reveals a set of 17 typical species of thedifferent habitat types, occurring almost exclu-sively in the relevant habitats and with greatabundances. Some of the species recorded weregeneralists having a broad ecological niche and
distribution range in the study area with noindicator value. Several species were encounteredwith increased abundances in the montane grass-lands and three of them were revealed as grasslandindicators (Figures 2 and 3). In the same context,eight bird species were typical of the agricul-tural sites and the bushy habitats of riverinevegetation, whereas another six bird specieswere typical for the forest conditions (Figures. 2and 3). If we compare our results with the typicalspecies found in a Mediterranean reserve atlow altitude (Dadia NP, less than 600m), we findsome similarities, concerning the generalist species(e.g. F. coelebs, E. rubecula) and the indicatorspecies of forest conditions (e.g. S. atricapilla,C. brachydactyla, typical for broad-leaved woods).However, we found that L. arborea is typical ofmontane grasslands in Pindos NP but it is alsotypical for lowland heaths in Dadia NP in easternGreece (Kati & Sekercioglu 2006). Special attentionshould be paid to the typical species that have anunfavourable conservation status in Europe at thistime (SPEC 2, 3). For our study area, these were:L. arborea and O. oenanthe (typical for montanegrasslands); M. striata (typical farmland species);and P. cristatus (typical pine forest species). Allindicator species as well as the species withunfavourable conservation status should be inte-grated into the future monitoring program of theNP, in order to evaluate their own conservationstatus as well as their habitat condition, on a long-term basis.
Bird diversity
Bird species richness was inversely correlatedwith altitude in our study area, but the weightedspecies richness, which also considers the conser-vation status of the species, was not. This wellknown pattern of decreased species richness withincreasing altitude is explained by the theory ofisland biogeography and has been shown for birdcommunities in a variety of climatic zones (Kattan& Franco, 2004; Prodon et al., 2002). However, wefound that the bird community of montane grass-lands, which is a quite widespread habitat type inthe Mediterranean mountains, comprises fewer butimportant species with unfavourable conservationstatus in Europe (SPEC 2, 3) and therefore should beconsidered as a primary habitat for conservation inthe NP.
Another important site for bird diversity con-servation in the study area was the agriculturalsite A2. It was a rural mosaic, situated in theforested zone, alternating small cultivated plots

ARTICLE IN PRESS
Bird communities and reserve management 55
with pastures and orchards, separated by naturalvegetation of hedges and tree lines. Site A1 wasless species-rich, where former agricultural plotswere not cultivated any more but were periodicallygrazed by livestock (Table 1). These rural mosaicswere the rarest habitat in our study area, becausesmall-scale farming using traditional agriculturalpractices has become financially unprofitable overthe last decades and has therefore led to landabandonment in the broader area of Pindos NP,as in other mountainous areas in Europe (Gellrich &Zimmermann, 2007; MacDonald et al., 2000).Farmland bird diversity is subject to two majorthreats in Europe: agricultural intensificationmostly in the northern lowlands (Chamberlainet al., 2000; Donald et al., 2001; Kati & Sekercioglu,2006; Pain & Pienkowski, 1997); and agro-pastoralland abandonment mostly in the Mediterraneanmountains. The latter results in forest encroachment,landscape heterogeneity decline, and agriculturalhabitat loss negatively affecting farmland birddiversity (Farina, 1997; Laiolo et al., 2004; Preisset al., 1997; Suarez-Seoane et al., 2002). Our resultsprovide additional evidence supporting the need toenhance the agri-environmental measures of theEuropean Common Agricultural Policy (CAP) againstland abandonment in less favoured mountainousareas, so as to maintain the traditional agriculturalpractices and the associated, biologically richagricultural mosaics.
We found that the mixed pine-beech woods werericher and more important for bird conservationrather than the pinewoods. However, the mixedwoods did not form a separate cluster in hierarch-ical clustering analysis and did not hold a specia-lised avifauna (no indicator species for mixedwoods). Mixed stands can be either more species-rich or not of unmixed stands, but they are knownto hold an intermediate bird species communitybetween those of broadleaved and coniferousstands (Archaux & Bakkaus, 2007; Dıaz, 2006;Donald et al., 1998; Willson & Comet, 1996).Besides, we found no statistically significant differ-ence when comparing the species richness (and theweighted species richness) of the unmixednatural pine and broad-leaved woods. We questiontherefore the general admission that broad-leavedstands hold richer bird communities than pureconiferous stands, when natural native standsare compared. This relationship could be attributedto the reduced ecological value of introducedconifer plantations compared to the natural nativecharacter of broad-leaved woods (Archaux &Bakkaus, 2007). These results may have a goodextrapolative value to other Mediterranean naturalforest ecosystems, since the forest habitat types
sampled in Pindos NP, such as the pinewoods(P. nigra) and the beech woods (F. sylvatica)are encountered in other mountains in theMediterranean (Italy, Corsica, Spain, Portugal,France) with the exception of the Balkanpinewoods, encountered only in Greece andsouthern Italy.
Bird diversity was strongly correlated withvertical structural vegetation complexity (NL,1/D). Vertical stratification in forest vegetationhas a positive influence over forest avifauna, as thepresence of shrub layers offers more ecologicalniches, foraging opportunities and breeding re-sources especially for undergrowth-dependent birdspecies (e.g. Camprodon & Brotons, 2006; Dıaz,2006; James & Wamer, 1982; Prodon & Lebreton,1981; Wesolowski, 2007). This relationship wasdetected for the first time in Greek islandecosystems (Watson, 1964), but no other studyhas been conducted to quantify it in differentGreek forest ecosystems since then.
Although tree height was not a statisticallysignificant environmental parameter to predict birdspecies in our CCA model, we found that thepresence of high trees was positively correlatedwith the number of species (S) and also with thenumber of important bird species (WS) in foresthabitats. We also found that two tall old growthpinewoods that have not been harvested forcenturies (P2, P3) were richer than those lowerpinewoods (P1, P4, P5). Forest bird species richnessincreases with tree height (Donald et al., 1998;Helle & Monkkonen, 1990). If we assume that hightree stands are also more mature, we couldattribute this positive relationship to the fact thatolder forest stands provide more tree holes, as wellas a higher amount of dead wood as breeding andfeeding habitat for a variety of specialist taxa (e.g.Dıaz, 2006; Gil-Tena et al., 2007; Hobson & Bayne,2000; Keller et al., 2003; Laiolo et al., 2003;Sekercioglu, 2002; Wesolowski, 2007).
Conservation management
In the applied context, our study proves theimportance and the need for maintenance of themontane grasslands in their present status. Sec-ondly, it suggests that the enhancement of tradi-tional agricultural practices in the broader areasurrounding the Pindos NP could have beneficialeffects on local bird communities. Thirdly, weemphasise the need for protection of the naturalremaining old growth forests in the core area.Although timber extraction is not allowed in thecore area since the creation of the NP (1966),

ARTICLE IN PRESS
V. Kati et al.56
logging of old growth pine trees for commercialreasons seems to have occurred in 80s, facilitatedby the existing road network. Fourthly, our resultsindicate that sustainable forestry practices need tomaintain the mixed character of forest standswhere they occur, the vertical structural complex-ity as well as a number of high and mature trees inmanaged forest stands. Finally, we argue that the14 species of unfavourable conservation status(SPEC 2 and 3) together with the typical speciesfound per bird habitat type should be integratedinto the future monitoring scheme of PindosNational Park, as a tool for forest management(Kati & Sekercioglu 2006; Muller 2005). The aboveproposals can have an extrapolative value in othermountainous reserves across the country and in theMediterranean mountains.
Table A1
Code Species w SPEC 2004 Cod
Piciformes PhcPvir Picus viridis 4 2 RreDmart Dryocopus martius 4a RigDmaj Dendrocopos major 1 MstDsyr Dendrocopos syriacus 4a 4 PpaDmed Dendrocopos medius 4a 4 PluDmin Dendrocopos minor 1 Pm
Passeriformes Pcr— Galerida cristata 2 3 PatLarb Lullula arborea 4a 2 Pca— Alauda arvensis 2 3 Pm— Hirundo daurica 1 AcaDurb Delichon urbica 2 3 SeuAcamp Anthus campestris 4a 3 CfaAtriv Anthus trivialis 1 CbrMflav Motacilla flava 1 Lco— Motacilla cinerea 1 GglMalb Motacilla alba 1 CcoCcincl Cinclus cinclus 1 CcoTtrogl Troglodytes troglodytes 1 PdoPmod Prunella modularis 1 4 FcoErub Erithacus rubecula 1 4 SseLmeg Luscinia megarhynchos 1 4 CchPhocrh Phoenicurus ochrurus 1 CcaOoen Oenanthe oenanthe 2 3 CcaMontsax Monticola saxatilis 2 3 LcuTmer Turdus merula 1 4 PyrTphil Turdus philomelos 1 4 CcoTvisc Turdus viscivorus 1 4 Eci— Sylvia melanocephala 1 4 Eci— Sylvia curruca 1 EhoSatr Sylvia atricapilla 1 4 Mca
aSpecies of Annex I of 79/409EU. SPEC 2: concentrated in Europe andin Europe but with unfavourable conservation status, SPEC 4: concen
Acknowledgements
This research was partially funded by theHellenic Ministry of Environment and Public Worksin the frame of the Sustainable DevelopmentProject (ETERPS funds) and from Pindos Perivallon-tiki NGO. We are grateful to I. Leonardos from theUniversity of Ioannina for support, to J. Foufopou-los and to S. Sfenthourakis for helpful comments onthe manuscript.
Appendix I
Inventory of all 62 bird species recorded in thestudy area and weighted index (w) according totheir conservation status (SPEC category 2004).
e Species w SPEC 2004
ol Phylloscopus collybita 1g Regulus regulus 1 4n Regulus ignicapilla 1 4r Muscicapa striata 2 3l Parus palustris 2 3g Parus lugubris 1 4ont Parus montanus 1ist Parus cristatus 4 2er Parus ater 1er Parus caeruleus 1 4aj Parus major 1ud Aegithalos caudatus 1r Sitta europaea 1m Certhia familiaris 1a Certhia brachydactyla 1 4l Lanius collurio 4a 3and Garrulus glandarius 1r Corvus corone 1rax Corvus corax 1m Passer domesticus 2 3el Fringilla coelebs 1 4r Serinus serinus 1 4l Chloris chloris 1 4rd Carduelis carduelis 1n Carduelis cannabina 4 2rv Loxia curvirostra 1pyr Pyrrhula pyrrhyla 1c Coccothraustes coccothraustes 1tr Emberiza citrinella 1 4rl Emberiza cirlus 1 4rt Emberiza hortulana 4 2l Milaria calandra 4 2
with unfavourable conservation status, SPEC 3: not concentratedtrated in Europe and with favourable conservation status.

ARTICLE IN PRESS
Bird communities and reserve management 57
Species codes are given only for the speciesrecorded during the point counts. See Table A1
References
Archaux, F., & Bakkaus, N. (2007). Relative impact ofstand structure, tree composition and climate onmountain bird communities. Forest Ecology andManagement, 247, 72–79.
Bani, L., Massimino, D., Bottoni, L., & Massa, R. A.(2005). Multiscale method for selecting indicatorspecies and priority conservation areas: A case studyfor broadleaved forests in Lombardy, Italy. Conserva-tion Biology, 20, 512–526.
Bibby, C. J., Burgess, N. D., & Hill, D. (1992). Bird censustechniques. London: Academy Press.
BirdLife International (2004). Birds in Europe: Populationestimates, trends and conservation status. Cam-bridge, UK: BirdLife International (BirdLife Conserva-tion Series No 12).
Blondel, J., Ferry, F., & Frochot, B. (1970). La methodedes indices ponctuels d‘abondance (I.P.A.) ou desreleves d‘ avifauna par ‘‘stations d‘ecoute’’. Alauda,38, 55–71.
Camprodon, J., & Brotons, L. (2006). Effects of under-growth clearing on the bird communities of theNorthwestern Mediterranean Coppice Holm oak for-ests. Forest Ecology and Management, 221, 72–82.
Chamberlain, D. E., Fuller, R. J., Bunce, R. G. H., &Duckworth, J. C. (2000). Changes in the abundance offarmland birds in relation to the timing of agriculturalintensification in England and Wales. Journal ofApplied Ecology, 37, 771–778.
Colwell, R. K. (2006). Estimate S: Statistical Estimationof species richness and shared species from sample.Version 8. User’s Guide and application /http://viceroy.eeb.unconn.edu/EstimateSS.
Dıaz, L. (2006). Influences of forest type and foreststructure on bird communities in oak and pine wood-lands in Spain. Forest Ecology and Management, 223,54–65.
Dobson, A. P., Rodrigue, J. P., Roberts, W. M., &Wilcove, D. S. (1997). Geographic distribution ofendangered species in the United States. Science,275, 550–553.
Donald, P. F., Fuller, R. J., Evans, A. D., & Gough, S. J.(1998). Effects of forest management and grazing onbreeding bird communities in plantations of broad-leaved and coniferous trees in western England.Biological Conservation, 85, 183–197.
Donald, P. F., Green, R. E., & Heath, M. F. (2001).Agricultural intensification and the collapse of Eur-ope’s farmland bird populations. Proceedings of theRoyal Society of London B, 268, 25–29.
Dufrene, M. (1999). IndVal or how to identify indicatorspecies of a sample typology. /http://mrw.wallonie.be/dgrne/sibw/outils/indval/home.htmlS.
Dufrene, M., & Legendre, P. (1997). Species assemblagesand indicator species: The need for a flexible
asymmetrical approach. Ecological Monographs, 67,345–366.
European Commission (2003). Interpretation manual ofEuropean Union habitats (EUR25). Brussels: EuropeanCommission.
Farina, A. (1997). Landscape structure and breeding birddistribution in a sub-Mediterranean agro-ecosystem.Landscape Ecology, 12, 365–378.
Field, A. (2005). Discovering statistics using SPSS. UK:SAGE publications.
Gellrich, M., & Zimmermann, N. E. (2007). Investigatingthe regional-scale pattern of agricultural land aban-donment in the Swiss mountains: A spatial statisticalmodelling approach. Landscape and Urban Planning,79, 65–76.
Gil-Tena, A., Saura, S., & Brotons, L. (2007). Effects offorest composition and structure on bird speciesrichness in a Mediterranean context: Implications forforest ecosystem management. Forest Ecology andManagement, 242, 470–476.
Gregory, R. D., Noble, D. G., & Custance, J. (2004). Thestate of play of farmland birds: Population trends andconservation status of lowland farmland birds in theUnited Kingdom. Ibis, 146, 1–13.
Heath, M. E., Evans, M. I., Hoccom, D. G., Payne, A. J., &Peet, N. B. (2000). Important bird areas in Europe:Priority sites for conservation. Cambridge: BirdLifeInternational.
Helle, P., & Monkkonen, M. (1990). Forest succession andbird communities: Theoretical aspects and practicalimplications. In A. Keast (Ed.), Biogeography andEcology of Forest Bird Communities (pp. 299–318).The Hague: SPB Academic Publishing.
Hobson, K., & Bayne, E. (2000). The effects of stand ageon avian communities in aspen dominated forests ofcentral Saskatchewan, Canada. Forest Ecology andManagement, 136, 121–134.
Howard, P. C., Viskanic, P., Davenport, T. R. B.,Kigenyi, F. W., Baltzer, M., Dickinson, C. J., et al.(1998). Complementarity and the use of indicatorgroups for reserve selection in Uganda. Nature, 394,275–472.
James, F. C., & Wamer, N. O. (1982). Relationshipsbetween temperate forest bird communities andvegetation structure. Ecology, 63, 159–171.
Johnson, G. A. M., & Freedman, B. (2002). Breeding birdsin forestry plantations and natural forest in thevicinity of Fundy National Park, New Brunswick.Canadian Field-Naturalist, 116, 475–487.
Kati, V., & Sekercioglu, C. H. (2006). Diversity, ecologicalstructure, and conservation of the landbird commu-nity of Dadia reserve, Greece. Diversity & Distribu-tion, 12, 620–629.
Kati, V., Devillers, P., Dufrene, M., Legakis, A., Vokou, D.,& Lebrun, Ph. (2004a). Testing the value of sixtaxonomic groups as biodiversity indicators at a localscale. Conservation Biology, 18, 667–675.
Kati, V., Devillers, P., Dufrene, M., Legakis, A., Vokou, D.,& Lebrun, Ph. (2004b). Hotspots, complementarity orrepresentativeness? Designing optimal small-scale

ARTICLE IN PRESS
V. Kati et al.58
reserves for biodiversity conservation. BiologicalConservation, 120, 471–480.
Kattan, G. H., & Franco, P. (2004). Bird diversity alongelevational gradients in the Andes of Colombia: Areaand mass effects. Global Ecology and Biogeography,13, 451–458.
Keller, J. K., Richmond, M. E., & Smith, C. R. (2003). Anexplanation of patterns of breeding bird speciesrichness and density following clearcutting in north-eastern USA forests. Forest Ecology and Management,541–564.
Kent, M., & Coker, P. (1994). Vegetation descriptionand analysis: A practical approach. Chirchester:Wiley.
King, D. I., & DeGraaf, R. M. (2000). Bird species diversityand nesting success in mature, clearcut and shelter-wood forest in northern New Hampshire, USA. ForestEcology and Management, 129, 227–235.
Kuchler, A. W. (1988). A physiognomic and structuralanalysis of vegetation. In A. W. Kuchler, & I. S.Zonneveld (Eds.), Vegetation Mapping. Handbook ofVegetation Science (pp. 37–50). Dordrecht: KluwerAcademic Publishers.
Laiolo, P., Caprio, E., & Rolando, A. (2003). Effects oflogging and non-native tree proliferation on the birdsoverwintering in the upland forests of north-westernItaly. Forest Ecology and Management, 179, 441–451.
Laiolo, P., Dondero, R., Ciliento, E., & Rolando, A.(2004). Consequences of pastoral abandonment forthe structure and diversity of the alpine avifauna.Journal of Applied Ecology, 41, 294–304.
Lance, A. N., & Phinney, M. (2001). Bird responses topartial retention timber harvesting in central interiorBritish Columbia. Forest Ecology and Management,142, 267–280.
Lawton, J. H., Bignell, D. E., Bolton, B., Bloemers, G. F.,Eggleston, P., Hammond, P. M., et al. (1998).Biodiversity inventories, indicator taxa and effects ofhabitat modification on tropical forest. Nature, 391,72–76.
Leps, J., & Smilauer, P. (2005). Multivariate analysis ofecological data using CANOCO. UK: Cambridge Uni-versity Press.
Lindenmayer, D. B., Franklin, J. F., & Fischer, J. (2006).General management principles and a checklist ofstrategies to guide forest biodiversity conservation.Biological Conservation, 131, 433–445.
Lombard, A. (1995). The problem with multi-speciesconservation: Do hotspots, ideal reserves and existingreserves coincide? South African Journal of Zoology,30, 145–163.
MacDonald, D., Crabtree, J. R., Wiesinger, G., Dax, T.,Stamou, N., Fleury, P., et al. (2000). Agriculturalabandonment in mountain areas of Europe: Environ-mental consequences and policy response. Journal ofEnvironmental Management, 59, 47–69.
Magurran, A. E. (2004). Measuring biological diversity.USA: Blackwell Publishing.
Mertzanis, G. (1999). Special Environmental study ofPindos-Zagori. Life 96 NAT/GR/3222. Project ARCTOS:
Conservation of brown bear and its habitats in Greece(phase b). Thessaloniki: ARCTOUROS.
Muller, J. (2005). Bird communities as indicators forwoodland structures in oak woods. OrnithologischeBeobachter, 102, 15–32.
Muller, J., Hothorn, T., & Pretzsch, H. (2007). Long-termeffects of logging intensity on structures, birds,saproxylic beetles and wood-inhabiting fungi in standsof European beech Fagus sylvatica L. Forest Ecologyand Management, 242, 297–305.
Pain, D. J., & Pienkowski, M. W. (1997). Farming andbirds in Europe. The common agricultural policy andits implications for bird conservation. London: Aca-demic Press.
Preiss, E., Martin, J.-L., & Debussche, M. (1997). Ruraldepopulation and recent landscape changes in aMediterranean region: Consequences to the breedingavifauna. Landscape Ecology, 12, 51–61.
Prendergast, J. R., Quinn, R. M., Lawton, J. H.,Eversham, B. C., & Gibbon, D. W. (1993). Rare species,the coincidence of diversity hotsposts and conserva-tion strategies. Nature, 365, 335–337.
Prodon, R., & Lebreton, J-D. (1981). Breeding avifauna ofa Mediterranean succession: The holm oak andcork oak series in the eastern Pyrenees, 1. Analysisand modelling of the structure gradient. Oikos, 37,21–38.
Prodon, R., Thibault, J.-C., & Dejaifve, P.-A. (2002).Expansion vs. compression of bird altitudinalranges on a Mediterranean island. Ecology, 83,1294–1306.
Sekercioglu, C. H. (2002). Effects of forestry practices onvegetation structure and birds community of KibaleNational Park, Uganda. Biological Conservation, 107,229–240.
Sekercioglu, C. H. (2006). Increasing awareness of avianecological function. Trends in Ecology & Evolution, 21,464–471.
Sekercioglu, C. H., Daily, G. C., & Ehrlich, P. R. (2004).Ecosystem consequences of bird declines. Proceedingsof the National Academy of Science USA, 101,18042–18047.
Simberloff, D. (1998). Flagships, umbrellas and key-stones: A single species management passe in thelandscape era? Biological Conservation, 83, 247–257.
Suarez-Seoane, S., Osborne, P. E., & Baudry, J. (2002).Responses of birds of different biogeographic originsand habitat requirements to agricultural land aban-donment in northern Spain. Biological Conservation,105, 333–344.
ter Braak, C. J. F. & Smilauer, P. (2002). CANOCOReference Manual and Canoco Draw for WindowsUser’s Guide: Software for Canonical CommunityOrdination (version 4.5). Ithaca, NY, USA: Microcom-puter Power.
Thiollay, J.-M. (2002). Bird diversity and selection ofprotected areas in a large neotropical forest tract.Biodiversity and Conservation, 11, 1377–1395.
Trakolis, D., Platis, P., Tsiontsis, A., Spanos, K., Meliadis,I. & Malamidis, G. (1996). Management Plan of Pindos

ARTICLE IN PRESS
Bird communities and reserve management 59
National Park. Thessaloniki, Greece: Forest Service ofGrevena, Ioannina, and Metsovo.
Tsounis, G., Malakou, M. & Ravasini, M. (1985).Study of the fauna of Pindos National Park(in greek). Athens: Hellenic Society for NatureProtection.
van der Maarel, E. (2005). Vegetation ecology. Oxford:Blackwell Publishing.
Vessby, K., Soderstrom, B., Glimskar, A., & Svensson, B.(2002). Species-richness correlations of six different
taxa in Swedish seminatural grasslands. ConservationBiology, 16, 430–439.
Watson, G. E. (1964). Ecology and evolution of passerinebirds on the islands of the Aegean Sea. Ph.D. Thesis,Yale University.
Wesolowski, T. (2007). Primeval conditions—what can welearn from them? Ibis, 149, 64–77.
Willson, M. F., & Comet, T. A. (1996). Bird communities ofnorthern forests: Patterns of diversity and abundance.Condor, 98, 337–349.




















