
Dissociation equilibrium of human recombinant interferon gamma
Upload
independentCategory
view
3download
0
0041-1337/00/6906-1147/0TRANSPLANTATION Vol. 69, 1147–1155, No. 6, March 27, 2000Copyright © 2000 by Lippincott Williams & Wilkins, Inc. Printed in U.S.A.
EARLY EXPRESSION OF INTERFERON-g INDUCIBLE PROTEIN 10AND MONOKINE INDUCED BY INTERFERON-g IN CARDIAC
ALLOGRAFTS IS MEDIATED BY CD81 T CELLS1
ANIL KAPOOR,2,3 KEN MORITA,2,3 TARA M. ENGEMAN,4 SHOJI KOGA,3 EVAN M. VAPNEK,3
MICHAEL G. HOBART,3 AND ROBERT L. FAIRCHILD3–6
Departments of Urology and Immunology, Cleveland Clinic Foundation, Cleveland, OH 44195and Department of Pathology, Case Western Reserve University School of Medicine, Cleveland, OH
Background. Our goal was to test the intragraftmRNA expression and production of two chemokinesthat are potent chemoattractants for antigen-primedT cells, interferon-g inducible protein 10 (IP-10) andmonokine induced by IFN-g, (Mig), in allogeneic heartgrafts.
Methods. Syngeneic or allogeneic A/J (H-2a) heartswere heterotopically transplanted to wild-type,CD42/2, CD8a2/2, or IFN-g2/2 C57BL/6 (H-2b) recipi-ents. To test expression of IP-10 and Mig, grafts wereremoved 1–8 days posttransplant for RNA isolationand Northern blot analysis. To test the potential recip-ient leukocyte populations mediating intraallograftexpression of IP-10 and Mig, recipients were treatedwith anti-NK 1.1, anti-CD4, and/or anti-CD8 monoclo-nal antibodies before transplantation.
Results. Allogeneic heart grafts transplanted towild-type, but not IFN-g2/2, recipients expressed IP-10and Mig at day 12 posttransplant that increasedthereafter until rejection was completed. Expressionof IP-10 and Mig in isografts was low or undetectable.Cardiac allografts from CD81 T cell depleted, but notNK cell or CD41 T cell depleted, recipients had low toundetectable expression of IP-10 and Mig on day 12posttransplant. Similarly, cardiac allografts fromCD82/2, but not CD42/2, recipients had low to unde-tectable expression of IP-10 and Mig on day 12 post-transplant.
Conclusions. Early intraallograft expression of Migand IP-10 during primary rejection of cardiac allo-grafts is dependent on the activities of recipient CD81
T cells.
Allogeneic organ transplantation has become a commonlyused therapy for end-stage heart and renal disease (1). Withthe use of current immunosuppressive strategies, the loss ofallografts due to T cell-mediated acute rejection has de-
creased significantly. The development of chronic rejectionhas become the leading cause of solid organ allograft loss (2,3). It has become increasingly clear, however, that earlyacute rejection episodes are a critical risk factor for thesubsequent development of chronic rejection (4). Acute rejec-tion of allografts is mediated by the coordinated infiltrationand effector functions of T cells with specificity for alloanti-gens (5, 6). The surgical trauma of organ transplantationinduces a low level intragraft inflammatory response thatincludes the up-regulation of adhesion molecules on vascularendothelium and production of cytokines such as tumor ne-crosis factor-a and interleukin-1 (7–10). The critical compo-nents of this inflammation that mediate recruitment of al-loantigen-specific T cell into allografts during acute rejectionepisodes remain unclear.
The chemokines constitute a superfamily of low molecularweight proteins that direct the trafficking of cells to tissuesites of inflammation (11, 12). Leukocyte infiltration andtissue pathology is inhibited by neutralization of specificchemokines in experimental models of inflammation, indicat-ing a critical role of chemokines in at least some forms ofinflammation (13–16). Whether or not chemokines direct re-cipient leukocytes to allograft tissue is not known. Severalgroups of investigators have approached this problem bytesting the presence of various chemokines in allografts dur-ing rejection (17–23). Elucidation of the temporal aspects andmechanisms mediating chemokine production in allograftswill be important for designing chemokine neutralizationstrategies to prevent alloantigen-primed T cell recruitmentto grafts and prolonging allograft survival.
Several chemokines are specifically chemoattractant for Tcells and are attractive candidates for facilitating T cell re-cruitment to allografts. Two of these chemokines, interfer-on-g (IFN-g*) inducible protein 10 (IP-10) and monokineinduced by IFN-g (Mig), are induced by IFN-g and are potentchemoattractants for antigen-primed T cells (24, 25). Previ-ous studies indicated the expression of IP-10 mRNA in het-erotopically transplanted murine cardiac allografts but notisografts as early as day 3 posttransplant (26). These resultsindicated the ability of the recipient to quickly respond to thepresence of allogeneic heart tissue by inducing the expression
1 Supported by NIH grant AI40459 (RLF), the Ferdinand C. Val-entine Fellowship for Research in Urology from the New York Acad-emy of Medicine (EMV), and a gift to the Renal Transplant ResearchProgram at the Cleveland Clinic Foundation from the State of Qatar.
2 AK and KM each contributed equally to this work and should beconsidered principal authors.
3 Department of Urology, Cleveland Clinic Foundation.4 Department of Immunology, Cleveland Clinic Foundation5 Department of Pathology, Case Western Reserve University
School of Medicine.6 Address correspondence to: Robert L. Fairchild, PhD, Depart-
ments of Urology and Immunology, NB3–79, Cleveland Clinic Foun-dation, 9500 Euclid Avenue, Cleveland, OH 44195.
* Abbreviations: ; IFN-g, interferon-g; FITC, fluorescein isothio-cyanate; FSC, forward scatter; GAPDH, glyceraldehyde-3-phosphatedehydrogenase; IP-10, interferon-g inducible protein 10; mAb, mono-clonal antibody; MCP, monocyte chemotactic protein; Mig, monokineinduced by IFN-g; MIP, macrophage inflammatory protein; PE, phy-coerythrin; SSC, side scatter.
1147

of the IFN-g-induced chemokine at early times posttrans-plant. Whether Mig is also induced in these heart grafts andthe cellular mechanisms mediating the expression of these Tcell chemoattractants has not been tested. The goal of ourstudy was to compare the temporal expression of Mig andIP-10 in heart allografts and to test the relationship betweenintraallograft expression of IP-10 and Mig with recipientleukocyte populations. The results indicate that intraal-lograft expression of Mig and IP-10 occurs as early as day 2posttransplant during primary rejection of cardiac allografts.This early intraallograft expression is dependent on IFN-gand the activities of recipient CD81 T cells. We propose thatthis early production of IP-10 and Mig may facilitate thesubsequent recruitment of alloantigen-primed T cells to theallograft.
MATERIALS AND METHODS
Mice. A/J (H-2a) and C57BL/6 (H-2b) mice were obtained throughDr. Clarence Reeder at the National Cancer Institute (Frederick,MD). C57BL/6 mice with a targeted deletion in the gene encodingCD4 (C57BL/6-Cd4tm1 Mak), CD8a (C57BL/6-Cd8atm1 Mak), and IFN-g(C57BL/6-Ifngtm1Ts) were purchased from The Jackson Laboratory(Bar Harbor, ME). Adult males of 6–10 weeks old were used through-out this study.
Antibodies. Monoclonal antibodies (mAb) from the culture super-natant of the IgG-producing hybridomas GK1.5 (anti-mouse CD4),YTS 169.4.2.1 (anti-mouse CD8), and PK 136 (anti-mouse NK 1.1)were purified by protein G chromatography. Phycoerythrin (PE)-conjugated antibodies specific for NK cells and CD41 and CD81 Tcells for use in monitoring antibody-mediated depletion of cell pop-ulations by flow cytometry were purchased from PharMingen (SanDiego, CA). Polyclonal goat antiserum to mouse Mig was purchasedfrom R & D Systems (Minneapolis, MN) and polyclonal goat anti-serum to mouse IP-10 was purchased from Santa Cruz Biotechnology(Santa Cruz, CA). Fluorescein isothiocyanate (FITC)-conjugateddonkey antibody specific for goat IgG was purchased from JacksonImmunoResearch Laboratories, Inc. (West Grove, PA).
Heterotopic cardiac transplant. Cardiac transplants were per-formed according to the method of Corry and co-workers (27). Briefly,donor and recipient mice were anesthetized with chloral hydrate/phenobarbital. Donor hearts were harvested and placed in chilledlactated Ringer’s solution during which the recipient mice wereprepared. The donor heart was anastomosed to the recipient abdom-inal aorta and vena cava using microsurgical techniques. On com-pletion of the anastomosis and organ perfusion, the transplantedhearts resumed spontaneous contraction. The strength and qualityof cardiac impulses were graded each day as previously described(28). Rejection of cardiac grafts was considered complete by thecessation of impulse and confirmed visually by laparotomy. C57BL/6recipients rejected A/J cardiac allografts between 8 and 10 days aftertransplantation. Cardiac isografts in C57BL/6 recipients functionedfor more than 100 days. In some experiments, recipient C57BL/6mice were given 150 mg of GK1.5, YTS 169.4.2.1 or PK136 i.p. on 3consecutive days and were then transplanted with a cardiac allograftwithin 2 days of the final mAb treatment. Antibody staining and flowcytometry analyses of spleen cells from mAb treated recipients ondays 2–7 posttransplant indicated .90% depletion of the target cellpopulation (data not shown).
Northern blot analysis. Whole-cell RNA was isolated from trans-planted and native heart tissue using Trizol (Gibco BRL, GrandIsland, NY). Briefly, transplanted and naive hearts were excisedfrom recipients, snap frozen, and homogenized in Trizol. After ex-traction, precipitation and resuspension in diethylpyrocarbonate-treated dH2O, 10-mg aliquots of RNA were electrophoresed in 1%agarose formaldehyde-denaturing gels and analyzed by Northernblot analysis, as described previously (29). Blots were hybridized
with 32P-labeled oligonucleotide probes specific for IP-10 and Mig.After hybridization and exposure with one chemokine oligonucleo-tide probe, the filter was stripped of the probe by washing the blotthree times in 0.4% sodium dodecyl sulfate at 90°C and then hybrid-ized with the next test chemokine probe. After hybridization with thetest chemokine probes was completed, the blot was stripped andprobed with a rat GAPDH-specific oligonucleotide. Densitometryusing Storage Phosphor Screen analyzer (Molecular Dynamics,Sunnyview, CA) was utilized to comparatively measure the chemo-kine signal and the GAPDH signal for each sample of the blot. Thechemokine signals for each sample of the blot were then normalizedby expressing the density of the chemokine signal as a ratio to thesignal of the GAPDH signal for each RNA sample. The mean ratio foreach group (i.e., time point) was determined and differences betweenthe means was tested using unpaired Student’s t test. A P,0.01 wasconsidered a significant difference between group means.
Isolation and analysis of graft infiltrating cells by flow cytometry.Isografts and A/J cardiac allografts were retrieved from C57BL/6recipients and rinsed in Hanks’ balanced salt solution. The graftswere incubated for 1 hr at 37°C in Hanks’ balanced salt solutioncontaining 1.5 mg/ml of collagenase (type 4, Sigma Chemical Co., St.Louis, MO), 1.5 mg/ml hyaluronidase (type IV-S, Sigma), and 0.2mg/ml DNAse (type I, Boehringer Mannheim, Indianapolis, IN). Thegrafts were then placed in kidney dialysis tubing and mashed with asyringe piston. The cell suspension was collected, washed once, andcentrifuged through a Lympholyte M (Cederlane Laboratories Ltd.,Hornby, Ontario, Canada) gradient. The interface cells were col-lected and washed three times with fluorescence activated cell sorterstaining buffer (Dulbecco’s phosphate-buffered saline with 2% fetalcalf serum/0.2% NaN3) and 105 cell aliquots were incubated on ice in100 ml rat serum (Rockland, Gilbertsville, PA) diluted 1023 in thestaining buffer. After 30 min, the cells were washed twice andstained with FITC conjugated antibodies for mouse CD69 and PEconjugated antibodies for mouse NK1.1, CD4, or CD8. After 30 minon ice, the cells were washed 5 times, resuspended in staining buffer,and analyzed by two-color flow cytometry using a FACScan andCellQuest software (Becton Dickinson, San Jose, CA). Each experi-ment was performed three times and results from a single represen-tative experiment are shown.
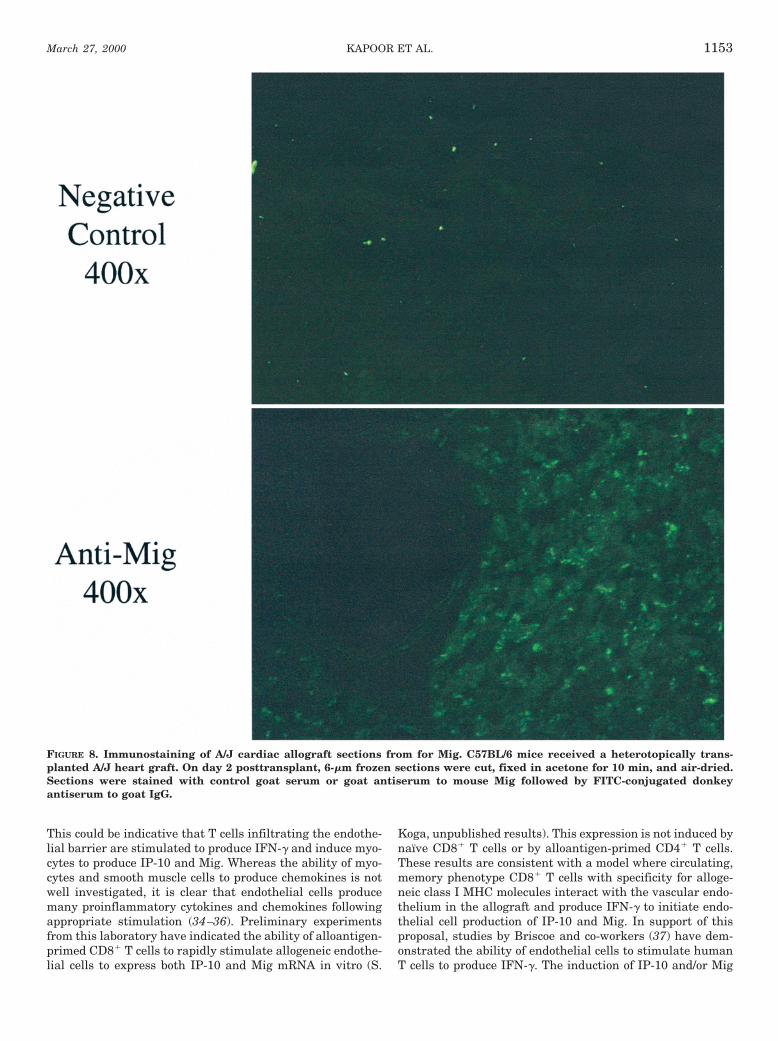
Tissue staining for immunofluorescence. For immunohistology,8-mm frozen sections of cardiac allografts were cut, fixed in acetonefor 10 min, air dried, and stained overnight with goat anti-mouseIP-10 or Mig, diluted 50–100 mg/ml. Control slides were incubatedwith goat IgG as the primary antibody. After three washes in phos-phate-buffered saline for 5 min each, slides were incubated withFITC-conjugated donkey anti-goat IgG, diluted to 7.5 mg/ml. Afterstaining, slides were washed in phosphate-buffered saline and a dropof Vectashield (Vector Laboratories, Inc., Burlingame, CA) was usedto reduce fluorescence photobleaching. The slides were viewed undera fluorescent microscope and the images captured using Adobe Pho-toshop 4.0 (Mountain View, CA).
RESULTS
Intragraft expression of IP-10 and Mig mRNA during re-jection of cardiac allografts. To begin to examine the appear-ance of the T cell chemoattractants IP-10 and Mig in cardiacallografts during acute rejection, the temporal expression ofMig and IP-10 mRNA in isogeneic and allogeneic heart graftswas tested. C57BL/6 (H-2b) mice were heterotopically trans-planted with a heart graft from a syngeneic donor or from anA/J (H-2a) donor. The A/J allografts were rejected betweendays 8 and 10 posttransplant. From day 1 to day 8 posttrans-plant, the iso- and allografts were retrieved for RNA isolationand analysis by Northern blot hybridization. Intragraft ex-pression levels of both IP-10 and Mig were low or undetect-able in the native recipient heart and in isografts at each day
TRANSPLANTATION1148 Vol. 69, No. 6
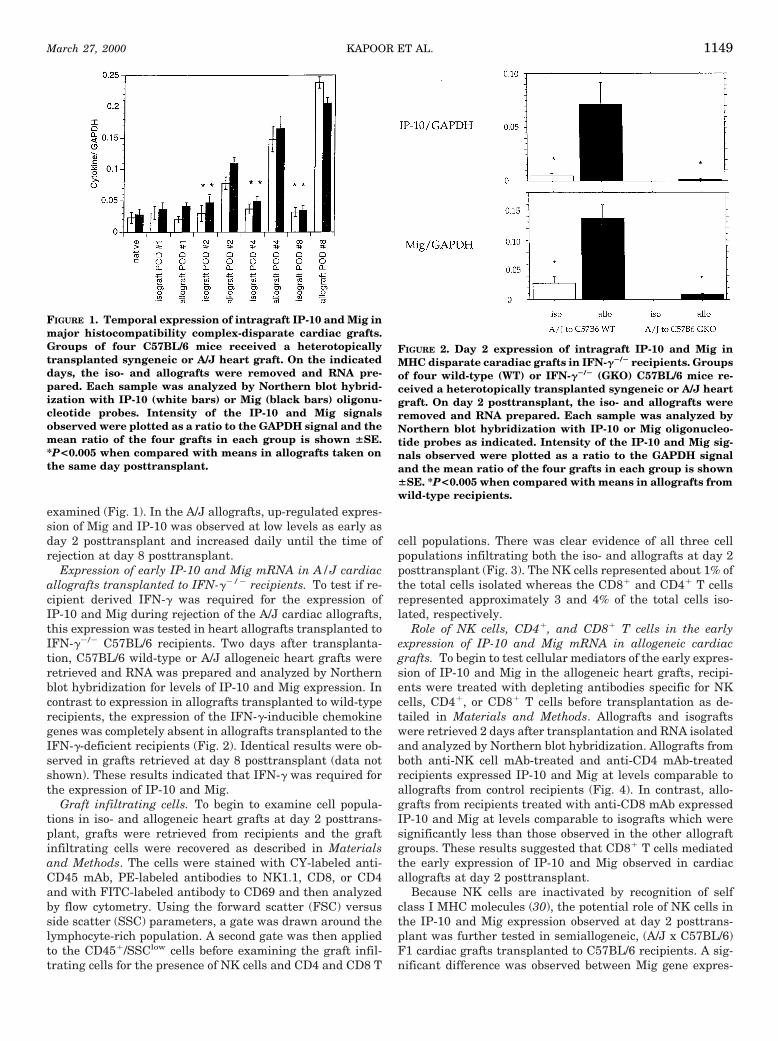
examined (Fig. 1). In the A/J allografts, up-regulated expres-sion of Mig and IP-10 was observed at low levels as early asday 2 posttransplant and increased daily until the time ofrejection at day 8 posttransplant.
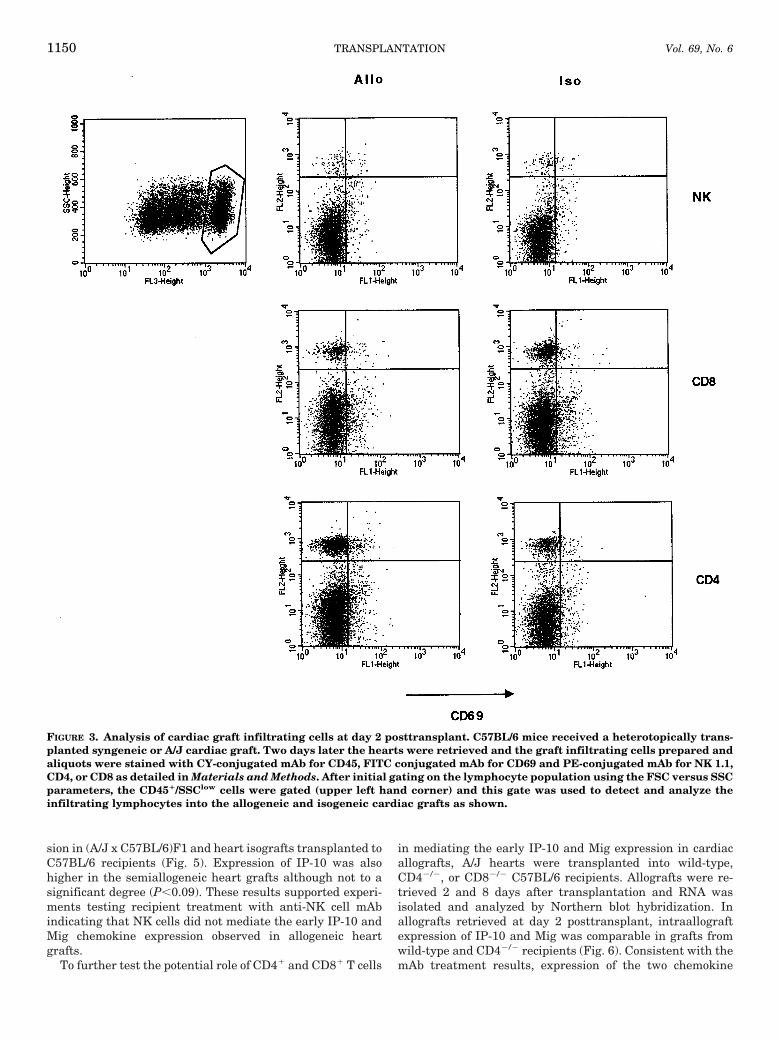
Expression of early IP-10 and Mig mRNA in A/J cardiacallografts transplanted to IFN-g2/2 recipients. To test if re-cipient derived IFN-g was required for the expression ofIP-10 and Mig during rejection of the A/J cardiac allografts,this expression was tested in heart allografts transplanted toIFN-g2/2 C57BL/6 recipients. Two days after transplanta-tion, C57BL/6 wild-type or A/J allogeneic heart grafts wereretrieved and RNA was prepared and analyzed by Northernblot hybridization for levels of IP-10 and Mig expression. Incontrast to expression in allografts transplanted to wild-typerecipients, the expression of the IFN-g-inducible chemokinegenes was completely absent in allografts transplanted to theIFN-g-deficient recipients (Fig. 2). Identical results were ob-served in grafts retrieved at day 8 posttransplant (data notshown). These results indicated that IFN-g was required forthe expression of IP-10 and Mig.
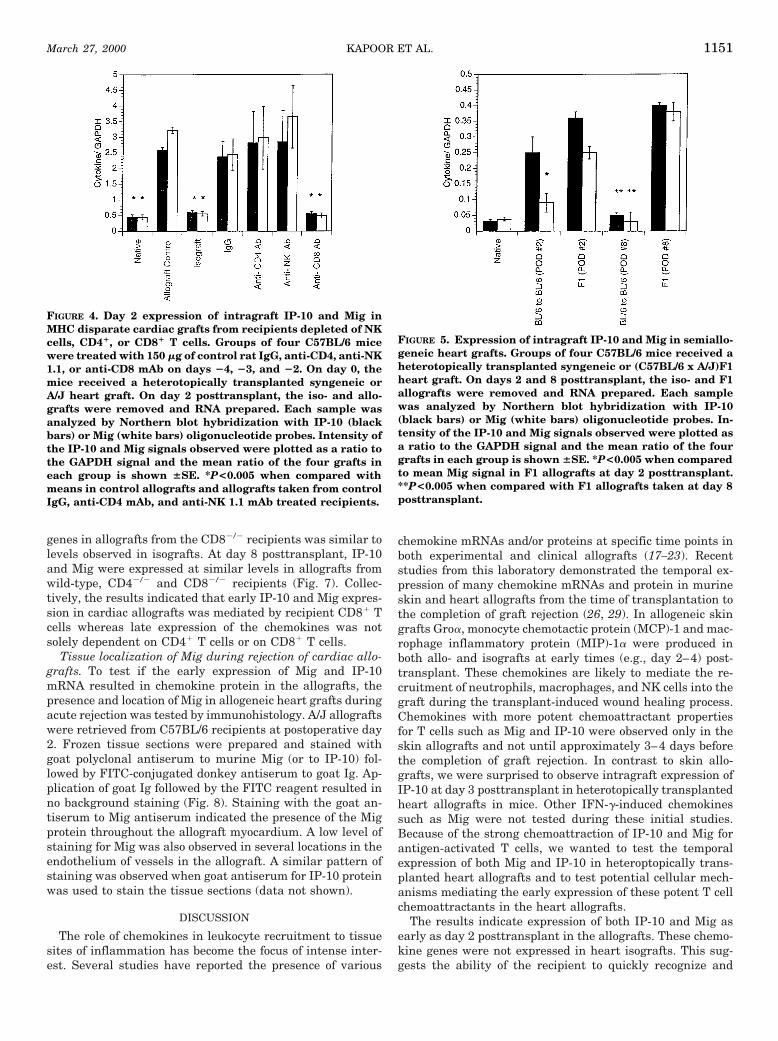
Graft infiltrating cells. To begin to examine cell popula-tions in iso- and allogeneic heart grafts at day 2 posttrans-plant, grafts were retrieved from recipients and the graftinfiltrating cells were recovered as described in Materialsand Methods. The cells were stained with CY-labeled anti-CD45 mAb, PE-labeled antibodies to NK1.1, CD8, or CD4and with FITC-labeled antibody to CD69 and then analyzedby flow cytometry. Using the forward scatter (FSC) versusside scatter (SSC) parameters, a gate was drawn around thelymphocyte-rich population. A second gate was then appliedto the CD451/SSClow cells before examining the graft infil-trating cells for the presence of NK cells and CD4 and CD8 T
cell populations. There was clear evidence of all three cellpopulations infiltrating both the iso- and allografts at day 2posttransplant (Fig. 3). The NK cells represented about 1% ofthe total cells isolated whereas the CD81 and CD41 T cellsrepresented approximately 3 and 4% of the total cells iso-lated, respectively.
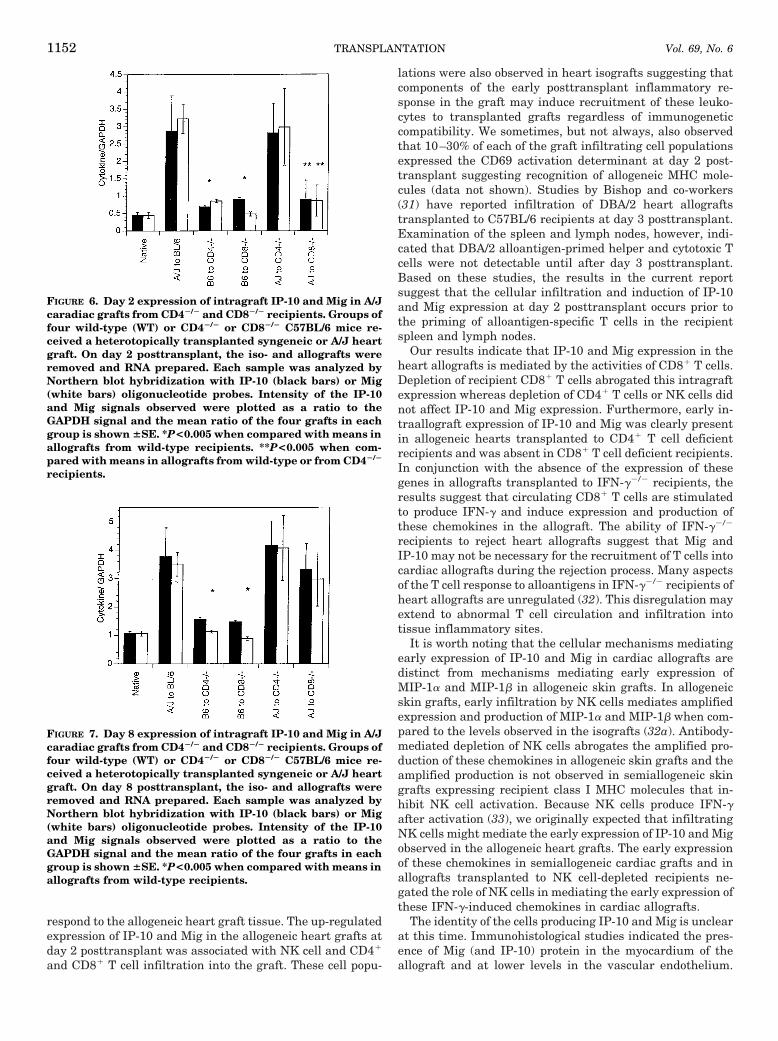
Role of NK cells, CD41, and CD81 T cells in the earlyexpression of IP-10 and Mig mRNA in allogeneic cardiacgrafts. To begin to test cellular mediators of the early expres-sion of IP-10 and Mig in the allogeneic heart grafts, recipi-ents were treated with depleting antibodies specific for NKcells, CD41, or CD81 T cells before transplantation as de-tailed in Materials and Methods. Allografts and isograftswere retrieved 2 days after transplantation and RNA isolatedand analyzed by Northern blot hybridization. Allografts fromboth anti-NK cell mAb-treated and anti-CD4 mAb-treatedrecipients expressed IP-10 and Mig at levels comparable toallografts from control recipients (Fig. 4). In contrast, allo-grafts from recipients treated with anti-CD8 mAb expressedIP-10 and Mig at levels comparable to isografts which weresignificantly less than those observed in the other allograftgroups. These results suggested that CD81 T cells mediatedthe early expression of IP-10 and Mig observed in cardiacallografts at day 2 posttransplant.
Because NK cells are inactivated by recognition of selfclass I MHC molecules (30), the potential role of NK cells inthe IP-10 and Mig expression observed at day 2 posttrans-plant was further tested in semiallogeneic, (A/J x C57BL/6)F1 cardiac grafts transplanted to C57BL/6 recipients. A sig-nificant difference was observed between Mig gene expres-
FIGURE 1. Temporal expression of intragraft IP-10 and Mig inmajor histocompatibility complex-disparate cardiac grafts.Groups of four C57BL/6 mice received a heterotopicallytransplanted syngeneic or A/J heart graft. On the indicateddays, the iso- and allografts were removed and RNA pre-pared. Each sample was analyzed by Northern blot hybrid-ization with IP-10 (white bars) or Mig (black bars) oligonu-cleotide probes. Intensity of the IP-10 and Mig signalsobserved were plotted as a ratio to the GAPDH signal and themean ratio of the four grafts in each group is shown 6SE.*P<0.005 when compared with means in allografts taken onthe same day posttransplant.
FIGURE 2. Day 2 expression of intragraft IP-10 and Mig inMHC disparate caradiac grafts in IFN-g2/2 recipients. Groupsof four wild-type (WT) or IFN-g2/2 (GKO) C57BL/6 mice re-ceived a heterotopically transplanted syngeneic or A/J heartgraft. On day 2 posttransplant, the iso- and allografts wereremoved and RNA prepared. Each sample was analyzed byNorthern blot hybridization with IP-10 or Mig oligonucleo-tide probes as indicated. Intensity of the IP-10 and Mig sig-nals observed were plotted as a ratio to the GAPDH signaland the mean ratio of the four grafts in each group is shown6SE. *P<0.005 when compared with means in allografts fromwild-type recipients.
KAPOOR ET AL.March 27, 2000 1149
sion in (A/J x C57BL/6)F1 and heart isografts transplanted toC57BL/6 recipients (Fig. 5). Expression of IP-10 was alsohigher in the semiallogeneic heart grafts although not to asignificant degree (P,0.09). These results supported experi-ments testing recipient treatment with anti-NK cell mAbindicating that NK cells did not mediate the early IP-10 andMig chemokine expression observed in allogeneic heartgrafts.
To further test the potential role of CD41 and CD81 T cells
in mediating the early IP-10 and Mig expression in cardiacallografts, A/J hearts were transplanted into wild-type,CD42/2, or CD82/2 C57BL/6 recipients. Allografts were re-trieved 2 and 8 days after transplantation and RNA wasisolated and analyzed by Northern blot hybridization. Inallografts retrieved at day 2 posttransplant, intraallograftexpression of IP-10 and Mig was comparable in grafts fromwild-type and CD42/2 recipients (Fig. 6). Consistent with themAb treatment results, expression of the two chemokine
FIGURE 3. Analysis of cardiac graft infiltrating cells at day 2 posttransplant. C57BL/6 mice received a heterotopically trans-planted syngeneic or A/J cardiac graft. Two days later the hearts were retrieved and the graft infiltrating cells prepared andaliquots were stained with CY-conjugated mAb for CD45, FITC conjugated mAb for CD69 and PE-conjugated mAb for NK 1.1,CD4, or CD8 as detailed in Materials and Methods. After initial gating on the lymphocyte population using the FSC versus SSCparameters, the CD451/SSClow cells were gated (upper left hand corner) and this gate was used to detect and analyze theinfiltrating lymphocytes into the allogeneic and isogeneic cardiac grafts as shown.
TRANSPLANTATION1150 Vol. 69, No. 6
genes in allografts from the CD82/2 recipients was similar tolevels observed in isografts. At day 8 posttransplant, IP-10and Mig were expressed at similar levels in allografts fromwild-type, CD42/2 and CD82/2 recipients (Fig. 7). Collec-tively, the results indicated that early IP-10 and Mig expres-sion in cardiac allografts was mediated by recipient CD81 Tcells whereas late expression of the chemokines was notsolely dependent on CD41 T cells or on CD81 T cells.
Tissue localization of Mig during rejection of cardiac allo-grafts. To test if the early expression of Mig and IP-10mRNA resulted in chemokine protein in the allografts, thepresence and location of Mig in allogeneic heart grafts duringacute rejection was tested by immunohistology. A/J allograftswere retrieved from C57BL/6 recipients at postoperative day2. Frozen tissue sections were prepared and stained withgoat polyclonal antiserum to murine Mig (or to IP-10) fol-lowed by FITC-conjugated donkey antiserum to goat Ig. Ap-plication of goat Ig followed by the FITC reagent resulted inno background staining (Fig. 8). Staining with the goat an-tiserum to Mig antiserum indicated the presence of the Migprotein throughout the allograft myocardium. A low level ofstaining for Mig was also observed in several locations in theendothelium of vessels in the allograft. A similar pattern ofstaining was observed when goat antiserum for IP-10 proteinwas used to stain the tissue sections (data not shown).
DISCUSSION
The role of chemokines in leukocyte recruitment to tissuesites of inflammation has become the focus of intense inter-est. Several studies have reported the presence of various
chemokine mRNAs and/or proteins at specific time points inboth experimental and clinical allografts (17–23). Recentstudies from this laboratory demonstrated the temporal ex-pression of many chemokine mRNAs and protein in murineskin and heart allografts from the time of transplantation tothe completion of graft rejection (26, 29). In allogeneic skingrafts Groa, monocyte chemotactic protein (MCP)-1 and mac-rophage inflammatory protein (MIP)-1a were produced inboth allo- and isografts at early times (e.g., day 2–4) post-transplant. These chemokines are likely to mediate the re-cruitment of neutrophils, macrophages, and NK cells into thegraft during the transplant-induced wound healing process.Chemokines with more potent chemoattractant propertiesfor T cells such as Mig and IP-10 were observed only in theskin allografts and not until approximately 3–4 days beforethe completion of graft rejection. In contrast to skin allo-grafts, we were surprised to observe intragraft expression ofIP-10 at day 3 posttransplant in heterotopically transplantedheart allografts in mice. Other IFN-g-induced chemokinessuch as Mig were not tested during these initial studies.Because of the strong chemoattraction of IP-10 and Mig forantigen-activated T cells, we wanted to test the temporalexpression of both Mig and IP-10 in heteroptopically trans-planted heart allografts and to test potential cellular mech-anisms mediating the early expression of these potent T cellchemoattractants in the heart allografts.
The results indicate expression of both IP-10 and Mig asearly as day 2 posttransplant in the allografts. These chemo-kine genes were not expressed in heart isografts. This sug-gests the ability of the recipient to quickly recognize and
FIGURE 4. Day 2 expression of intragraft IP-10 and Mig inMHC disparate cardiac grafts from recipients depleted of NKcells, CD41, or CD81 T cells. Groups of four C57BL/6 micewere treated with 150 mg of control rat IgG, anti-CD4, anti-NK1.1, or anti-CD8 mAb on days 24, 23, and 22. On day 0, themice received a heterotopically transplanted syngeneic orA/J heart graft. On day 2 posttransplant, the iso- and allo-grafts were removed and RNA prepared. Each sample wasanalyzed by Northern blot hybridization with IP-10 (blackbars) or Mig (white bars) oligonucleotide probes. Intensity ofthe IP-10 and Mig signals observed were plotted as a ratio tothe GAPDH signal and the mean ratio of the four grafts ineach group is shown 6SE. *P<0.005 when compared withmeans in control allografts and allografts taken from controlIgG, anti-CD4 mAb, and anti-NK 1.1 mAb treated recipients.
FIGURE 5. Expression of intragraft IP-10 and Mig in semiallo-geneic heart grafts. Groups of four C57BL/6 mice received aheterotopically transplanted syngeneic or (C57BL/6 x A/J)F1heart graft. On days 2 and 8 posttransplant, the iso- and F1allografts were removed and RNA prepared. Each samplewas analyzed by Northern blot hybridization with IP-10(black bars) or Mig (white bars) oligonucleotide probes. In-tensity of the IP-10 and Mig signals observed were plotted asa ratio to the GAPDH signal and the mean ratio of the fourgrafts in each group is shown 6SE. *P<0.005 when comparedto mean Mig signal in F1 allografts at day 2 posttransplant.**P<0.005 when compared with F1 allografts taken at day 8posttransplant.
KAPOOR ET AL.March 27, 2000 1151
respond to the allogeneic heart graft tissue. The up-regulatedexpression of IP-10 and Mig in the allogeneic heart grafts atday 2 posttransplant was associated with NK cell and CD41
and CD81 T cell infiltration into the graft. These cell popu-
lations were also observed in heart isografts suggesting thatcomponents of the early posttransplant inflammatory re-sponse in the graft may induce recruitment of these leuko-cytes to transplanted grafts regardless of immunogeneticcompatibility. We sometimes, but not always, also observedthat 10–30% of each of the graft infiltrating cell populationsexpressed the CD69 activation determinant at day 2 post-transplant suggesting recognition of allogeneic MHC mole-cules (data not shown). Studies by Bishop and co-workers(31) have reported infiltration of DBA/2 heart allograftstransplanted to C57BL/6 recipients at day 3 posttransplant.Examination of the spleen and lymph nodes, however, indi-cated that DBA/2 alloantigen-primed helper and cytotoxic Tcells were not detectable until after day 3 posttransplant.Based on these studies, the results in the current reportsuggest that the cellular infiltration and induction of IP-10and Mig expression at day 2 posttransplant occurs prior tothe priming of alloantigen-specific T cells in the recipientspleen and lymph nodes.
Our results indicate that IP-10 and Mig expression in theheart allografts is mediated by the activities of CD81 T cells.Depletion of recipient CD81 T cells abrogated this intragraftexpression whereas depletion of CD41 T cells or NK cells didnot affect IP-10 and Mig expression. Furthermore, early in-traallograft expression of IP-10 and Mig was clearly presentin allogeneic hearts transplanted to CD41 T cell deficientrecipients and was absent in CD81 T cell deficient recipients.In conjunction with the absence of the expression of thesegenes in allografts transplanted to IFN-g2/2 recipients, theresults suggest that circulating CD81 T cells are stimulatedto produce IFN-g and induce expression and production ofthese chemokines in the allograft. The ability of IFN-g2/2
recipients to reject heart allografts suggest that Mig andIP-10 may not be necessary for the recruitment of T cells intocardiac allografts during the rejection process. Many aspectsof the T cell response to alloantigens in IFN-g2/2 recipients ofheart allografts are unregulated (32). This disregulation mayextend to abnormal T cell circulation and infiltration intotissue inflammatory sites.
It is worth noting that the cellular mechanisms mediatingearly expression of IP-10 and Mig in cardiac allografts aredistinct from mechanisms mediating early expression ofMIP-1a and MIP-1b in allogeneic skin grafts. In allogeneicskin grafts, early infiltration by NK cells mediates amplifiedexpression and production of MIP-1a and MIP-1b when com-pared to the levels observed in the isografts (32a). Antibody-mediated depletion of NK cells abrogates the amplified pro-duction of these chemokines in allogeneic skin grafts and theamplified production is not observed in semiallogeneic skingrafts expressing recipient class I MHC molecules that in-hibit NK cell activation. Because NK cells produce IFN-gafter activation (33), we originally expected that infiltratingNK cells might mediate the early expression of IP-10 and Migobserved in the allogeneic heart grafts. The early expressionof these chemokines in semiallogeneic cardiac grafts and inallografts transplanted to NK cell-depleted recipients ne-gated the role of NK cells in mediating the early expression ofthese IFN-g-induced chemokines in cardiac allografts.
The identity of the cells producing IP-10 and Mig is unclearat this time. Immunohistological studies indicated the pres-ence of Mig (and IP-10) protein in the myocardium of theallograft and at lower levels in the vascular endothelium.
FIGURE 6. Day 2 expression of intragraft IP-10 and Mig in A/Jcaradiac grafts from CD42/2 and CD82/2 recipients. Groups offour wild-type (WT) or CD42/2 or CD82/2 C57BL/6 mice re-ceived a heterotopically transplanted syngeneic or A/J heartgraft. On day 2 posttransplant, the iso- and allografts wereremoved and RNA prepared. Each sample was analyzed byNorthern blot hybridization with IP-10 (black bars) or Mig(white bars) oligonucleotide probes. Intensity of the IP-10and Mig signals observed were plotted as a ratio to theGAPDH signal and the mean ratio of the four grafts in eachgroup is shown 6SE. *P<0.005 when compared with means inallografts from wild-type recipients. **P<0.005 when com-pared with means in allografts from wild-type or from CD42/2
recipients.
FIGURE 7. Day 8 expression of intragraft IP-10 and Mig in A/Jcaradiac grafts from CD42/2 and CD82/2 recipients. Groups offour wild-type (WT) or CD42/2 or CD82/2 C57BL/6 mice re-ceived a heterotopically transplanted syngeneic or A/J heartgraft. On day 8 posttransplant, the iso- and allografts wereremoved and RNA prepared. Each sample was analyzed byNorthern blot hybridization with IP-10 (black bars) or Mig(white bars) oligonucleotide probes. Intensity of the IP-10and Mig signals observed were plotted as a ratio to theGAPDH signal and the mean ratio of the four grafts in eachgroup is shown 6SE. *P<0.005 when compared with means inallografts from wild-type recipients.
TRANSPLANTATION1152 Vol. 69, No. 6
This could be indicative that T cells infiltrating the endothe-lial barrier are stimulated to produce IFN-g and induce myo-cytes to produce IP-10 and Mig. Whereas the ability of myo-cytes and smooth muscle cells to produce chemokines is notwell investigated, it is clear that endothelial cells producemany proinflammatory cytokines and chemokines followingappropriate stimulation (34–36). Preliminary experimentsfrom this laboratory have indicated the ability of alloantigen-primed CD81 T cells to rapidly stimulate allogeneic endothe-lial cells to express both IP-10 and Mig mRNA in vitro (S.
Koga, unpublished results). This expression is not induced bynaıve CD81 T cells or by alloantigen-primed CD41 T cells.These results are consistent with a model where circulating,memory phenotype CD81 T cells with specificity for alloge-neic class I MHC molecules interact with the vascular endo-thelium in the allograft and produce IFN-g to initiate endo-thelial cell production of IP-10 and Mig. In support of thisproposal, studies by Briscoe and co-workers (37) have dem-onstrated the ability of endothelial cells to stimulate humanT cells to produce IFN-g. The induction of IP-10 and/or Mig
FIGURE 8. Immunostaining of A/J cardiac allograft sections from for Mig. C57BL/6 mice received a heterotopically trans-planted A/J heart graft. On day 2 posttransplant, 6-mm frozen sections were cut, fixed in acetone for 10 min, and air-dried.Sections were stained with control goat serum or goat antiserum to mouse Mig followed by FITC-conjugated donkeyantiserum to goat IgG.
KAPOOR ET AL.March 27, 2000 1153

production by endothelial cells after T cell interaction orstimulation by IFN-g has not been previously reported. It isalso possible, however, that other cells in the allograft suchas smooth muscle cells and myocytes are stimulated to pro-duce IP-10 and Mig after interaction with CD81 T cellsand/or IFN-g produced by the infiltrating T cells.
The significance of the early production of IP-10 and Mig incardiac allografts is unclear at this time. As discussed above,T cell infiltration and graft rejection can occur in the absenceof the early IP-10 and Mig production. Heart allografts arerejected by CD82/2 recipients as efficiently as wild-type re-cipients (38). Although the early expression of IP-10 and Migis not observed in CD82/2 recipients these chemokines areexpressed at high levels late in the graft rejection process.The cells inducing this late expression and the role of thesechemokines in recruiting alloantigen-primed CD41 T cellinto the allograft in these recipients remains to be tested.Using a skin graft transplant model, we have recently shownthat an antiserum with specificity for Mig is able to inhibit Tcell infiltration into class II MHC disparate allografts andprolong survival (39). Based on the results presented in thecurrent report, IP-10 and/or Mig neutralization may alsoinhibit alloantigen-primed T cell infiltration into grafts andpromote survival of heart allografts.
REFERENCES
1. Suthanthiran M, Strom TB. Renal transplantation. N EnglJ Med 1994; 331: 365.
2. Paul LC, Fellstrom B. Chronic vascular rejection of the heartand the kidney-have rational treatment options emerged?Transplantation 1992; 53: 1169.
3. Paul L. Chronic renal transplant loss. Kidney Int 1995; 47: 1491.4. Matas AJ, Gillingham KJ, Payne WD, Najarian JS. The impact
of an acute rejection episode on long-term renal allograft sur-vival (t1/2). Transplantation 1994; 57: 857.
5. Hall B. Cells mediating allograft rejection. Transplantation1991; 51: 51.
6. Rosenberg AS, Singer A. Cellular basis of skin allograft rejec-tion: an in vivo model of immune-mediated tissue destruction.Annu Rev Immunol 1993; 10: 333.
7. Pizzaro TT, Malinowska K, Kovacs EJ, Clancy J, Robinson JA,Piccini LA. Induction of TNFa and TNFb gene expression inrat cardiac transplants during allograft rejection. Transplan-tation 1993; 56: 399.
8. Clausell N, Molossi S, Rabinovitch M. Increased interleukin-1band fibronectin expression are early features of the develop-ment of the postcardiac transplant coronary arteriopathy inpiglets. Am J Pathol 1993; 142: 1772.
9. Mannon RB, Sundar SK, Sanfillipo FP, Coffman TM. Alterationsin renal interleukin-1 production during kidney transplantrejection in the rat. The effects of high-dose methylpred-nisolone. Transplantation 1993; 56: 1157.
10. DeMeester SR, Rolfe MW, Kunkel SL, et al. The bimodal expres-sion of tumor necrosis factor-a in association with rat lungreimplantation and allograft rejection. J Immunol 1993; 150:2494.
11. Taub DD, Oppenheim JJ. Chemokines, inflammation, and im-munity. Ther Immunol 1994; 1: 229.
12. Rollins BJ. Chemokines. Blood 1997; 90: 909.13. Shanley TP, Schmal H, Friedl HP, Jones ML, Ward PA. Role of
macrophage inflammatory protein-1a (MIP-1a) in acute lunginjury in rats. J Immunol 1995; 154: 4793.
14. Smith RE, Strieter RM, Phan SH, et al. Production and functionof murine macrophage inflammatory protein-1a in bleomycin-induced lung injury. J Immunol 1994; 153: 4704.
15. Gong J-H, Ratkay LG, Waterfield JD, Clark-Lewis I. An antag-onist of monocyte chemoattractant protein 1 (MCP-1) inhibitsarthritis in the MRL-lpr mouse model. J Exp Med 1997; 186:131.
16. Lloyd CM, Minto AW, Dorf ME, et al. RANTES and monocytechemoattractant protein-1 (MCP-1) play an important role inthe inflammatory phase of crescentic nephritis, but onlyMCP-1 is involved in crescent formation and interstitial fibro-sis. J Exp Med 1997; 185: 1371.
17. Russell ME, Adams DH, Wyner LR, Yamashita Y, Halnon NJ,Karnovsky MJ. Early and persistent induction of monocytechemoattractant protein 1 in rat cardiac allografts. Proc NatlAcad Sci USA 1993; 90: 6086.
18. Wieder KJ, Hancock WW, Schmidbauer G, et al. Rapamycintreatment depresses intragraft expression of KC/MIP-2, Gran-zyme B, and IFN-g in rat recipients of cardiac allografts. J Im-munol 1993; 151: 1158.
19. Schmouder RL, Strieter RM, Walz A, Kunkel SL. Epithelial-derived neutrophil-activating factor-78 production in humanrenal tubule epithelial cells and in renal allograft rejection.Transplantation 1995; 59: 118.
20. Pattison J, Nelson PJ, Huie P, et al. RANTES chemokine expres-sion in cell-mediated transplant rejection of the kidney. Lancet1994; 1 209.
21. Pattison JM, Nelson PJ, Huie P, Sibley RK, Krensky AM. RANTESchemokine expression in transplant-associated acceleratedatherosclerosis. J Heart Lung Transplant 1996; 15: 1194.
22. Nadeau KC, Azuma H, Tilney NL. Sequential cytokine dynamicsin chronic rejection of rat renal allografts: roles for cytokinesRANTES and MCP-1. Proc Natl Acad Sci USA 1995; 92: 8729.
23. Nagano H, Nadeau KC, Takada M, Kusaka M, Tilney NL. Se-quential cellular and molecular kinetics in acutely rejectingrenal allografts in rats. Transplantation 1997; 63: 1101.
24. Luster AD, Ravetch JV. Biochemical characterization of a g
interferon-inducible cytokine (IP-10). J Exp Med 1987; 166:1084.
25. Liao F, Rabin RL, Yannelli JR, Koniaris LG, Vanguri P, FarberJM. Human Mig chemokine: biochemical and functional char-acterization. J Exp Med 1995; 182: 1301.
26. Fairchild RL, VanBuskirk AM, Kondo T, Wakely ME, Orosz CG.Expression of chemokine genes during rejection and long-termacceptance of cardiac allografts. Transplantation 1997; 63:1807.
27. Corry RJ, Winn HJ, Russell PS. Primarily vascularized allo-grafts of hearts in mice. Transplantation 1973; 16: 343.
28. Orosz CG, Wakely E, Bergese SD, et al. Prevention of murinecardiac allograft rejection with gallium nitrate. Transplanta-tion 1996; 61: 783.
29. Kondo T, Novick AC, Toma H, Fairchild RL. Induction of che-mokine gene expression during allogeneic skin graft rejection.Transplantation 1996; 61: 1750.
30. Hoglund P, Sundback J, Olsson-Alheim MY, et al. Host MHCclass I gene control of NK-cell specificity in the mouse. Immu-nol Rev 1997; 155: 11.
31. Bishop DK, Shelby J, Eichwald EJ. Mobilization of T lympho-cytes following cardiac transplantation. Evidence that CD4-positive cells are required for cytotoxic T lymphocyte activa-tion, inflammatory endothelial development, graft infiltration,and acute allograft rejection. Transplantation 1992; 53: 849.
32. Konieczny BT, Dai Z, Elwood ET, et al. IFN-g is critical forlong-term allograft survival induced by blocking the CD28 andCD40 ligand T cell costimulation pathways. J Immunol 1998;160: 2059.
32a. Kondo T, Morita K, Watarai Y, et al. Early increased chemo-kine expression and production in murine allogeneic skingrafts is mediated by natural killer cells. Transplantation2000; 69: 969.
33. Cuturi MC, Anegon I, Sherman F, et al. Production of hemato-
TRANSPLANTATION1154 Vol. 69, No. 6

poietic colony-stimulating factors by human natural killercells. J Exp Med 1989; 169: 569.
34. Strieter RM, Kunkel SL, Showell HJ, et al. Endothelial cell geneexpression of a neutrophil chemotactic factor by TNFa, LPSand IL-1b. Science 1989; 243: 1467.
35. Mantovani A, Bussolino G, Dejana E. Cytokine regulation ofendothelial cell function. FASEB J 1992; 6: 2591.
36. Robson SC, Candinas D, Hancock WW, Wrighton C, Winkler H,Bach FH. Role of endothelial cells in transplantation. Int ArchAllergy Immunol 1995; 106: 305.
37. Briscoe DM, Henault LE, Geehan C, Alexander SI, Lichtman
AH. Human endothelial cell costimulation of T cell IFN-gproduction. J Immunol 1997; 159: 3247.
38. Krieger NR, Yin DP, Fathman CG. CD41 but not CD81 cells areessential for allorejection. J Exp Med 1996; 184: 2013.
39. Koga S, Auerbach MB, Engeman TM, Novick AC, Toma H,Fairchild RL. T cell infiltration into class II MHC disparateallografts and acute rejection is dependent on the IFN-g in-duced chemokine Mig. J Immunol 1999; 163: 4878.
Received 27 July 1999.Accepted 12 October 1999
0041-1337/00/6906-1155/0TRANSPLANTATION Vol. 69, 1155–1159, No. 6, March 27, 2000Copyright © 2000 by Lippincott Williams & Wilkins, Inc. Printed in U.S.A.
INFLUENCE OF COLD ISCHEMIA TIME, PRETRANSPLANT ANTI-PORCINE ANTIBODIES, AND DONOR/RECIPIENT SIZE
MATCHING ON HYPERACUTE GRAFT REJECTION AFTERDISCORDANT PORCINE TO CYNOMOLGUS KIDNEY
TRANSPLANTATION
M. LOSS,1 R. KUNZ,1 M. PRZEMECK,2 J. SCHMIDTKO,1 H, ARENDS,1 A. JALALI,1 R. LORENZ,1
S. PIEPENBROCK,2 J. KLEMPNAUER,1 AND M. WINKLER1,3
Klinik fur Viszeral- und Transplantationschirurgie, Zentrum Anasthesiologie, Medizinische Hochschule Hannover,30625 Hannover, Germany
Organs transplanted between phylogenetically dis-parate species, such as from the pig into the primate,are subject to hyperacute rejection (HAR). This formof xenograft rejection is mediated by preformed natu-ral antibodies and is believed to occur invariably indiscordant xenografts thus leading to rapid destruc-tion and complete thrombosis of the graft. Recentdata, however, have shown that in the porcine to cyno-molgus monkey setting, HAR is not inevitably seenafter porcine kidney transplantation. The influence ofpreoperative antiporcine antibody levels in the recip-ient, cold ischemia time, and donor organ weight onthe onset of HAR was investigated by using unmodi-fied large white pigs (aged 3–12 weeks) as organ do-nors and adult cynomolgus monkeys (aged 1.5–3.5years) as recipients. Porcine kidney xenotransplanta-tion was performed in either a non-life-supportingmodel (n57) or in a life-supporting model (n58). Inboth models, no correlation was found between coldischemia time and HAR. When preoperative anti-por-cine antibody levels were investigated, a significantincrease in incidence of HAR was observed in animals
with elevated anti-porcine IgM (P<0.05) but not IgGlevels (P5NS). Interestingly, although 5 of 12 graftswith an organ weight of less than 50 g underwent HAR,none of three grafts with a donor organ weight of morethan 70 g showed signs of HAR. In addition, all threelarger grafts showed intraoperative and postoperativeurine production, although only in 1 (48 g) of the 12grafts weighing less than 50 g primary graft functionwas observed. In one animal, a second porcine kidney(23 g) was successfully transplanted (without HAR)immediately after HAR and subsequent removal of afirst porcine kidney (20 g). These results indicate thatin the porcine to cynomolgus monkey setting anti-porcine IgM rather than IgG anti-porcine antibodylevels seem to be of predominant importance for theinduction of HAR. By increasing the donor organ sizeand weight the frequency of the onset of HAR can be atleast reduced. This is most likely due to immunoab-sorption of the recipients preformed antibodies in theporcine kidney without lethal damage for the graft.
After transplantation of Gala1–3Gal positive solid organsinto Gala1–3Gal negative recipients, a rapid destruction ofthe donor organ mediated by preformed recipient antibodieswith subsequent complement activation may occur withinminutes to hours after reperfusion (1, 2). This reaction hasbeen classified as hyperacute xenograft rejection (HAR) (3,4). Although there was general consensus that HAR almost
1 Klinik fur Viszeral- und Transplantationschirurgie.2 Zentrum Anasthesiologie.3 Address correspondence to: Michael Winkler, M.D., Klinik fur
Viszeral- und Transplantationschirurgie, Medizinische Hochschule,Carl Neuberg Str. 1, 30625 Hannover, Germany.
LOSS ET AL.March 27, 2000 1155
Copyright © 2022 FDOKUMEN




















