
Reliability-based Analysis and Maintenance of Buried Pipes ...
Upload
independentCategory
view
1download
0
Tansley review
Drowned, buried and carried away: effects ofplant traits on thedistributionof native andalienspecies in riparian ecosystems
Author for correspondence:Roland Jansson
Tel: +46 90 786 9573
Email: [email protected]
Received: 10 March 2014Accepted: 19 June 2014
Jane A. Catford1,2,3* and Roland Jansson4*1School of Botany, The University of Melbourne, Melbourne, Vic. 3010, Australia; 2Fenner School of Environment and Society, The
Australian National University, Canberra, ACT 0200, Australia; 3Department of Ecology, Evolution and Behavior, University of
Minnesota, Saint Paul, MN 55108, USA; 4Department of Ecology and Environmental Science, Ume�a University, SE-901 87 Ume�a,
Sweden
Contents
Summary 19
I. Introduction 19
II. Functional traits and life history adaptations of riparian plants 20
III. Local and regional patterns in the distribution of riparian plants 24
IV. Alien plant invasion in riparian zones 26
V. Modification and management of riparian plant communities 30
VI. Conclusions and future research 32
Acknowledgements 33
References 33
New Phytologist (2014) 204: 19–36doi: 10.1111/nph.12951
Key words: community assembly,disturbance, exotic species, flooding regime,functional traits, hydrochory, inundation,non-native species.
Summary
Riparian vegetation is exposed to stress from inundation and hydraulic disturbance, and is often
rich innative andalienplant species.Wedescribe35 traits that enableplants to copewith riparian
conditions. These include traits for tolerating or avoiding anoxia and enabling underwater
photosynthesis, traits that confer resistance and resilience to hydraulic disturbance, and
attributes that facilitate dispersal, such as floating propagules. This diversity of life-history
strategies illustrates that there are many ways of sustaining life in riparian zones, which helps
to explain high riparian biodiversity. Using community assembly theory, we examine how
adaptations to inundation, disturbance and dispersal shape plant community composition along
key environmental gradients, and how human actions have modified communities. Dispersal-
relatedprocesses seemtoexplainmanypatterns, highlighting the influenceof regional processes
on local species assemblages. Using alien plant invasions like an (uncontrolled) experiment in
community assembly, we use an Australian and a global dataset to examine possible causes of
high degrees of riparian invasion.We found that high proportions of alien species in the regional
species pools have invaded riparian zones, despite not being riparian specialists, and that riparian
invaders disperse in more ways, including by water and humans, than species invading other
ecosystems.
I. Introduction
Riparian zones, defined as areas bordering runningwaters and lakesthat are temporarily flooded and influenced by elevated water
tables, are challenging environments for vascular plants. Riparianspecies must be able to cope with waterlogging, fluctuating waterlevels and physical disturbance from floods and, in cold areas, ice(Lytle & Poff, 2004; Bornette et al., 2008; Merritt et al., 2010b;Lind et al., 2014). The hydrology of many rivers, for example indesert environments, is highly unpredictable, requiring that*These authors contributed equally to this work.
� 2014 The Authors
New Phytologist� 2014 New Phytologist Trust
New Phytologist (2014) 204: 19–36 19www.newphytologist.com
Review

riparian species are able to withstand long periods between suitableenvironmental conditions, which may themselves be of shortduration (Capon, 2003). Riparian zones often have distinct speciesassemblages (Sabo et al., 2005) and are oftenmore species-rich thansurrounding ecosystems (Brown&Peet, 2003). Riparian zones alsoexperience high degrees of invasion by alien species (i.e. speciesintroduced beyond their native range) (Stohlgren et al., 1999;Richardson et al., 2007). This raises something of a puzzle: why areriparian zones often so rich in species, given that riparian speciesneed one or several specialized traits to cope with the environment?
High native species richness could result from large proportionsof regional species pools being able to colonize riparian zones(Mouw & Alaback, 2003) or high rates of diversification (highspeciation and/or low extinction rates) among clades of riparianspecialists. High invasion of alien species suggests the former is true(Stohlgren et al., 1999; Richardson et al., 2007). Both high speciesrichness and high degrees of invasion are facilitated by spatial andtemporal environmental heterogeneity (Chesson &Huntly, 1997;Melbourne et al., 2007). Fluctuations in environmental condi-tions, often from disturbance, can result in successional mosaics,facilitating local species coexistence (Chesson &Huntly, 1997). Inriparian zones, floods remove plant biomass and provide newsubstrate for colonization. With time, these resource-rich, earlysuccessional patches transition into communities dominated bysuperior competitors; this usually occurs on infrequently floodedsurfaces (Dahlskog, 1966; Kalliola&Puhakka, 1988). Coexistenceis also facilitated by differences in seed production, dispersal,germination and establishment among species (Grubb, 1977).Overlaid on this gradient of flood disturbance is the ecophysiolog-ical stress caused by inundation. The high diversity of environ-mental conditions in riparian zones necessitates a diversity ofphysiological, morphological or life-history strategies, but the roleof these in determining patterns in riparian plant composition atvarious spatial and temporal scales is poorly understood.
Here,we review traits that enable plant species to inhabit riparianzones and examine the ways in which these adaptations determinewhere species occur in space and time. Specifically, we ask:(1) Which functional and life-history traits allow plant species toinhabit riparian environments, and howhave these traits evolved, asdemonstrated by their phylogenetic distribution?(2) How do plant adaptations affect the composition of riparianplant communities at different spatial scales?(3) What plant traits and environmental conditions facilitate highdegrees of alien plant invasion in riparian zones?(4) How do human activities modify the process of communityassembly and the functional composition of riparian plantcommunities?
We address these questions by reviewing the literature on plantadaptations to inundation stress, physical disturbance from floods,and dispersal along watercourses. We use the concept of commu-nity assembly to examine ways in which species’ traits affect localriparian assemblages by enabling certain species to pass thedispersal, environmental and biotic filters of a site. Treating alienplant invasions like an (uncontrolled) experiment in communityassembly, we use riparian invasions to illuminate the ripariancommunity assembly process.We test whether invading species are
better able to disperse into, tolerate local environmental conditionsof, and compete in riparian zones as compared with otherecosystems. We discuss the processes by which human actionscan change the composition of riparian flora and how such changescan be ameliorated, and conclude by outlining research needs. Ourdiscussion is limited to plants that inhabit the wet–dry ecotone,excluding plants with a fully aquatic habit unless they are found inriparian zones. We focus on riparian zones along running watersand lakes (including floodplains and floodplain wetlands), but donot consider coastal wetlands.
II. Functional traits and life-history adaptations ofriparian plants
1. Key structuring forces of the riparian environment
Despite occupying relatively small areas, riparian ecosystems play acrucial role in the landscape.Occurring at the land–water interface,they filter nutrients and other compounds that flow from uplandecosystems (Lowrance et al., 1984). Riparian vegetation stabilizesriverbanks (Hubble et al., 2010), buffers temperature variation instreams through shading (Barton et al., 1985), increases habitatheterogeneity (e.g. roots, logs) and provides energy and nutrients toaquatic ecosystems (Wallace et al., 1997).
The key characteristic that distinguishes riparian zones fromadjacent aquatic and terrestrial environments is the occurrence ofperiodic flooding andwaterlogging. Thewater regime is consideredthe most important disturbance factor in freshwater ecosystems(Puckridge et al., 1998), and numerous studies have evaluated howplants respond to variation in the flooding regime (Poff et al., 1997;Lytle & Poff, 2004; Merritt et al., 2010b). Here, we focus on traitsthat allow plants to cope with, and benefit from, physical andchemical effects of flooding and flows. They can be organized intothree main categories:(1) stress from inundation andwaterlogging,which can limit plantgrowth and development;(2) physical disturbance from floods, where high-velocity flowsand sediment erosion and deposition (exacerbated by ice action incold climates) can result in plant mortality and biomass loss, andincreases in resource availability;(3) waterborne dispersal through stream networks, where watertypically flows in a single direction down a river (Grant et al., 2007)and laterally in riparian zones in response to water-level fluctua-tions.
Building on previous reviews of riparian plant adaptations (Lytle&Poff, 2004), strategies (Bornette et al., 2008) and response guilds(Merritt et al., 2010b), we concentrate on traits that enable someplants in the regional species pool to colonize and thrive in riparianzones. We also consider the evolution of traits: if a trait occurs inmany clades, it implies that new, unrelated species potentially ableto colonize riparian zones have evolved relatively frequently.Conversely, if traits are found in few clades, this suggests thepresence of barriers that inhibit adaptation to riparian environ-ments, so that specialists, whose clade evolution is restricted toriparian zones, dominate the riparian flora. The review is notexhaustive, but is intended to highlight the diversity of traits
New Phytologist (2014) 204: 19–36 � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Review Tansley reviewNewPhytologist20

possessed by riparian plant species. All traits identified are listed inTable 1. Note that references to traits are only found in the table.
2. Inundation
Vascular plant species have evolved several traits to deal with life onland (e.g. gamete transfer through air, tissue for structural support,and vessels for internal fluid transport), whichmeans that toleratingperiods of inundation can be challenging, especially for seedlings.Gas diffusion is considerably slower underwater than in air (CO2
diffusion being 104 times slower underwater; Armstrong et al.,1994) and low underwater light availability limits photosynthesis(Vervuren et al., 2003). Underwater photosynthesis is enhanced byhaving thin or finely dissected leaves with thin cuticle facilitatingunderwater gas exchange, and chloroplasts in or close to theepidermis (Table 1). Some amphibious genera, such as Iso€etes,Crassula and Littorella, possess CO2-concentrating mechanisms tofacilitate photosynthesis. However, internal transport of mineralnutrients and photosynthetates becomes difficult in the absence ofleaf evapotranspiration, which is only possible in air (Kozlowski,1984). Inundation may also disrupt sexual reproduction (Craw-ford, 1987) and can lead to anoxic and toxic root environments(Jackson & Drew, 1984). By inhibiting respiration, oxygenshortage can lead to a lack of energy (Voesenek et al., 2006). Evenamong flood-tolerant species, roots cannot tolerate anaerobicconditions formore than a fewdays (Vartapetian&Jackson, 1997).Plant adaptations to oxygen shortage either reduce the need foroxygen and/or increase the tolerance of low oxygen conditions, oravoid low oxygen conditions (Table 1; Bailey-Serres & Voesenek,2008).
The physiological tolerance of anoxia can be increased, andoxygen requirements reduced, for short time periods by minimiz-ing energy use, producing ATP by glycolysis and fermentation, andsynthesizing compounds that protect cells from toxins producedduring anoxia (Table 1). Species adapted to prolonged inundationavoid anoxia by extending out of the water (shoot elongation andhyponastic growth) or by transporting oxygen to oxygen-deficientparts. Interconnected gas-filled spaces known as aerenchyma allowoxygen diffusion from shoots to roots, where oxygen helps todetoxify the rhizosphere. Adventitious roots form at shoot bases toreplace roots killed by anoxia. Anoxia can also be avoided throughdormancy, during which metabolic activity is low, or if the timingof life cycle events (e.g. flowering and seed release) correspondswithperiods of receding or low water levels. Many plants can toleratelonger periods of inundation when dormant. Accordingly, thelower elevational limits of species seem to be primarily determinedby the duration of summer floods (Vervuren et al., 2003; van Ecket al., 2006). Several adaptations can facilitate photosynthesis insubmerged conditions.
When analysing the number of times adaptations to inundationhave evolved, it is difficult to make a distinction between aquaticand riparian species, as the fully aquatic habit represents one end inthe range of flooding tolerance amongst angiosperms (Jacksonet al., 2009). Thewater lilies,Nymphaeales, are entirely aquatic anddiverged at the base of the angiosperm tree (APG III, 2009),showing that adaptations to submergence evolved early. Similarly,
the aquatic and wetland clades Acorales and Alismatales, alsodiverged at the base of the monocot clade (APG II, 2003).Evolution of inundation tolerance from terrestrial ancestors hasoccurred 205–245 separate times among seed plants (Cook, 1999),and traits such as aerenchyma, thin cuticles and finely dissectedleaves are found in many clades. Thus, overcoming the stress ofinundation – and passing that particular environmental filter toenable occupancy of riparian conditions – does not seem to havebeen a major evolutionary hurdle.
3. Physical disturbance from water-level variation
Riparian plants may cope with physical disturbance from floodsand ice through traits that enable survival of an individual(‘resistance’, sensu Townsend & Hildrew, 1994), or traits thatfacilitate rapid population growth and (re)colonization of dis-turbed patches or recently deposited substrate (‘resilience’, sensuTownsend & Hildrew, 1994).
Resistance strategies include stem, leaf and root flexibility,reduction of plant size, increased plant density, brittle twigsenabling self-thinning, and deep and widespread root systems(Table 1). Although some biomassmay be lost, these strategies helpindividuals to survive disturbance events. Flexible stems, wide-spread root systems and perennating organs below or close to theground protect against ice scour.
‘Resilient’ plants able to recover following the accumulation oflitter and sediment tend to have large seeds, persistent seed banksand capacity for lateral spread (Table 1). Cleared areas and newsurfaces can be rapidly (re)colonized by plant species producingmany seeds or vegetative propagules, or by recruitment from soilseedbanks. In some species, seed release is timed to themost suitableperiods, such as the recession of spring floods, which facilitatescolonization of exposed surfaces. Many resilient species are annualor have high growth rates once established.
Vegetative reproduction can confer both resistance and resilienceto flood disturbance. Many riparian species produce suckers, tillersor shoots from deeply buried, and thus protected, plant organs.Rhizomes, for example, facilitate recolonization of disturbedpatches and reduce the risk that floods destroy entire clones.Vegetative fragments can become detached and dispersed duringdisturbance events, further enhancing recolonization ability.Vegetative fragments are often buoyant and so aid in dispersal,and can remain viable for weeks after detachment. They can alsostart to produce roots shortly after detachment, facilitating rapidestablishment once stranded.
Being subject to water-level variation, riparian zones mayexperience long periods of drought. Plants can avoid (or ‘resist’)effects of drought by having traits such as phreatophytic roots thatextend to the water table, or can be resilient to drought throughtraits that enable rapid recolonization, such as those describedearlier (Capon, 2003; Stromberg et al., 2007).
Traits conferring resilience to floods anddroughts are not uniqueto riparian taxa, but can be found in many clades adapted toepisodic disturbance, suggesting that numerous disturbance-adapted species in regional species pools may colonize riparianzones. Many resistance traits, such as deep, wide root systems and
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014) 204: 19–36
www.newphytologist.com
NewPhytologist Tansley review Review 21

Table 1 Plant traits and means by which they enable occupancy and facilitate growth in riparian environments
Trait Function References
Adaptations to inundation and submergence in waterFacilitation of photosynthesisThin cuticle Enhances gas exchange to counteract slow diffusion in water Mommer et al. (2005), Mommer &
Visser (2005), Voesenek et al. (2006)Chloroplasts in, or close to,epidermis
Increase access to light and promote carbon uptake fromwater
Mommer et al. (2005), Mommer &Visser (2005), Voesenek et al. (2006)
Thin or finely dissected leaves Increase leaf area to maximize carbon and nutrient uptakefrom water
Mommer et al. (2005), Mommer &Visser (2005), Voesenek et al. (2006)
CAM or C4 physiology toconcentrate CO2
Enables carbon uptake from sediment Madsen & Sand-Jensen (1991),Keeley (1998)
Anoxia toleranceEconomic use of energy (ATP) Reduction in the use of ATP when in short supply; fast recovery
of protein synthesis at reoxygenationBailey-Serres & Voesenek (2008),Colmer & Voesenek (2009)
Biochemical pathways for ATPproduction in anoxic conditions(glycolysis and fermentation)
Energy production in the absence of oxygen Bailey-Serres & Voesenek (2008),Colmer & Voesenek (2009)
Synthesis of compounds (e.g.phytoglobins and antioxidants)that reduce harmful and toxiceffects of anoxia in cells
Protects against toxic compounds such as nitric oxide andreactive oxygen species (ROS) produced during anoxia
Visser et al. (2003)
Anoxia avoidanceShoot elongation Enables shoots and leaves to extend out of the water to
facilitate gas exchange, flowering and pollinationRidge (1987), Voesenek et al. (2004)
Hyponastic growth Differential cell elongation in for example petioles of leaves tomake organs more upright, which enables plants to reachwater surface and/or acquire more light and helps limitsedimentation on leaves
Ridge (1987)
Aerenchyma Hollow areas allowing internal oxygen diffusion from shoots toroots
Seago et al. (2005), Voesenek et al.(2006)
Oxygenation of rhizosphere Detoxifies the rhizosphere from reduced forms of iron,manganese and hydrogen sulphide
Laan et al. (1989)
Suberin barriers in the epidermisof basal roots
Limits loss of oxygen from roots and ensures that roots act as a‘pipeline’ for oxygen transport to root tips
Jackson & Drew (1984)
Adventitious roots New roots in aerated soils replacing roots killed by anoxia, or soilssupplied with oxygen from roots
Blom et al. (1994)
Lenticels on stems and roots Allow gas diffusion and entrance of oxygen through the barkof woody species
Kozlowski (1997)
Low metabolic activity Limits negative effects of energy deficit Geigenberger (2003)Timing of life cycle events Ensures that events such as flowering, seed release and
germination occur at appropriate times, such as duringperiods of low water levels
Blom & Voesenek (1996)
Adaptations to disturbance by floods and iceResistanceHigh stem, root or leaf flexibility Decreases the risk of biomass loss as a result of high-velocity
flowsKarrenberg et al. (2002)
Reduction in plant size Smaller and more compact plants are more resistant tomechanical damage from flow
Boeger & Poulson (2003), Puijalon &Bornette (2004)
Reduced internode or spacerlength
Smaller and more compact plants are more resistant tomechanical damage from flow
Pollux et al. (2007)
Twig bases with preformedbreaking points
Allows self-thinning and hence reduces the drag force of flowingwater
Beismann et al. (2000)
Deep and wide root systems Resistance to scour from flow and ice, and ability to acquireresources over a large area
Karrenberg et al. (2002)
Budding parts at or below theground
Protects the perennating organs of herbs from ice scourand being frozen into ice
Engstr€om et al. (2011)
ResilienceLarge seeds Seedlings more likely to penetrate upwards through layers of
sediment or litterXiong et al. (2001)
Lateral spread with runners, tillersor rhizomes
Faster recolonization of disturbed patches; less likely that an entireclone is affected by a physical disturbance
Xiong et al. (2001)
New Phytologist (2014) 204: 19–36 � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Review Tansley reviewNewPhytologist22

resting buds at, or close to, the ground are found in a large numberof clades. By contrast, several of the resistance traits are primarilyfound in the family Salicaceae (Table 1), with the genera Salix andPopulus dominating active floodplains in the Northern Hemi-sphere (Karrenberg et al., 2002), in which the riparian habit isprimitive.
4. Dispersal by water
Compared with terrestrial plants, plants that border streams andrivers may disperse their propagules (diaspores) via two additionalpathways: by water (hydrochory) or via fish (ichtyochory).Although the prevalence of hydrochory in riparian floras is notwell quantified and is likely to vary among biogeographic regions,evidence suggests that a large proportion of riparian plant speciesdisperse by hydrochory (Andersson et al., 2000a; Pettit & Froend,2001; Boedeltje et al., 2004; Jansson et al., 2005; Merritt &Wohl,2006).
Hydrochores can be dispersed floating on the water surface orunderwater (including along the stream bed; Parolin, 2006).Although the former arguably receives more attention, largenumbers of propagules of many species are transported underwater(Goodson et al., 2003; Gurnell et al., 2008). In a study of seedtransport along two Rocky Mountain rivers, Merritt & Wohl(2006) found that 31 and 56% of the seeds were transported on thesurface of the two streams, whereas the remaining seeds were
transported in suspension or along the channel bed. Hydrochory isusually a secondary dispersalmechanism; propagules typically droponto the groundbefore being carried away during floods (Schneider& Sharitz, 1988; Parolin, 2006). Some riparian plants, however,such as orchids of the ‘Disa uniflora type’ in southern Africa(Kurzweil, 1993), lean over and drop their seeds directly intostreams. Buoyancy of viable seeds can be achieved through corkytissue and seed coats that prevent imbibition, and adaptations likeaerenchyma may enable vegetative propagules to float (Table 1).Propagules lacking buoyancy can still be transported by water bysticking to floating objects, such as logs, branches and leaf-litterpacks (Nilsson et al., 2010).
Hydrochory offers five main advantages to riparian plants. First,a large proportion of hydrochores, especially buoyant hydrochores,can disperse long distances compared with other dispersal vectors(Andersson et al., 2000b; Pollux et al., 2009). Secondly, unlikewind-dispersed seeds, the dispersal distance of buoyant hydroch-ores is largely independent of their mass (Ikeda & Itoh, 2001).Accordingly, hydrochores tend to be heavier than wind-dispersedseeds (Moles et al., 2005), and heavier seeds have higher seedlingsurvival (Moles & Westoby, 2004). Thirdly, by preventingdesiccation, immersion in water increases the longevity of vegeta-tive propagules. Fourthly, being bound to stream networks,hydrochory increases the probability that propagules are depositedin sites suitable for germination and growth, such as bare,waterlogged soil (van der Valk, 1981; Schneider & Sharitz,
Table 1 (Continued)
Trait Function References
Persistent seed bank More likely that viable seeds are present and germinate followingdisturbance or when suitable environmental conditions occur(e.g. desert systems); external sources of propagules are lessimportant
Xiong et al. (2001), Bossuyt &Honnay (2008)
Vegetative reproduction Rapid recolonization following disturbance Barrat-Segretain et al. (1999), Riis &Sand-Jensen (2006)
Timing of reproduction and seedrelease
Arrival of seeds when conditions are suitable for germination andestablishment, such as exposed surfaces saturated with water
Mahoney & Rood (1998)
High growth rates Increase ability to capitalize on suitable environmental conditions,which may be transient
Karrenberg et al. (2002)
Early reproduction and largeinvestment in seeds
Increase likelihood of reproducing and increasing population sizewhile environmental conditions are suitable; high investment inreproduction increases probability of species persisting inspace and time
Pettit & Froend (2001), Bagstad et al.(2005)
Traits enhancing dispersal along streams and riversPropagule floating ability Enables long-distance dispersal and effective colonization of
riparian zonesNilsson et al. (2010)
Corky or spongy tissue of low density Increases propagule buoyancy van der Pijl (1972)Waxy, cuticularized epidermis Prevents imbibition and sinking of propagules Sculthorpe (1967)Surface with furrows, pits or hairs Traps air bubbles, which increases the buoyancy of propagules Rivadavia et al. (2009)Floating vegetative propagules Enables long-distance dispersal and effective colonization of
riparian zonesBoedeltje et al. (2004), Riis &Sand-Jensen (2006)
Timing of propagule release More likely that propagules are released when conditions aresuitable for dispersal and germination
Kubitzki & Ziburski (1994), Pettit &Froend (2001), Karrenberg et al.
(2002)Tolerance to passing the digestivesystem of fish
Enables dispersal by riverine fish Horn et al. (2011)
Seeds with hard coat and toleranceof anoxic conditions
Abrasion resistance and tolerance of burial, which enables seedtransport with sediment and litter and enhances longevity in thesoil seedbank
Goodson et al. (2003), Gurnell et al.(2008)
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014) 204: 19–36
www.newphytologist.com
NewPhytologist Tansley review Review 23

1988). Finally, contact with water often acts as a phenological cue(Kubitzki & Ziburski, 1994). In temperate climates, immersion ofseeds in water tends to break dormancy in spring and inducedormancy in autumn, ensuring that germination only occurs inspring (Boedeltje et al., 2004). Indeed, some riparian species timeseed release to periods suitable for hydrochory or germination(Table 1).
For many species, traits enabling hydrochory have probablyevolved for other reasons (Johansson et al., 1996; Andersson et al.,2000b); their role in facilitating water dispersal has been asecondary effect. For example, traits enhancingwinddispersal, suchas wings, hairs or plumes, can facilitate hydrochory wherepropagules ‘float’ on water surface tension. For a number of treespecies, floating distances can be several times longer than typicalwind-related transport (S€aumel & Kowarik, 2013). Likewise,barbs, hooks and other structures that facilitate animal dispersal canreduce water surface tension and enhance buoyancy. Hence, it isnot only species that have evolved in riparian habitats that candisperse well by water.
Ichtyochory represents a special case of hydrochory (Horn et al.,2011). There are at least 275 fruit-eating fish species,most of whichare tropical. Frugivorous fish can be effective seed dispersers bydispersing large and nonbuoyant seeds, and contributing to long-distance, as well as upstream, dispersal (Horn et al., 2011). As withhydrochory, it is doubtful whether plants have adapted toichtyochory by selection. Although fish disperse many seeds, fruitsmay have evolved to be consumed by a range of animals, not justfish.
5. Phylogenetic patterns
Plants found in riparian zones are scattered across the angiospermphylogeny (APG II, 2003; APG III, 2009). For example, in theriparian floras of northern Sweden and Alberta, Canada, all themajor families present in the regional flora are also represented inriparian zones (Dynesius et al., 2004). This indicates that littleevolutionary adaptation is required for (formerly) terrestrial-onlyspecies to start occupying riparian zones as well, or that manyspecies from the regional species pool can inhabit riparianecosystems without the need for specific adaptations.
To better understand how adaptations to riparian environmentshave evolved, studies where multiple traits are mapped ontophylogenies are required. There are examples of single riparianrepresentatives in otherwise terrestrial clades, as well as clades thatare entirely aquatic/riparian. One might hypothesize that thegreater the differences between terrestrial and riparian environ-mental conditions, the greater the adaptations required byterrestrial clades to occupy riparian or aquatic environments.Amphibious and truly aquatic species should therefore occur infewer lineages than species occupying higher riparian elevations.This pattern is observed in the genus Ranunculus, which consists ofc. 600 herbaceous species (H€orandl & Emadzade, 2012). Theaquatic and wetland Ranunculus species are found in two clades:sections Hecatonia/Batrachium and Flammula, which possessnumerous adaptations to flood tolerance and hydrochory, such asfinely dissected leaves, aerenchyma, physiological tolerance of
anoxia and hydrochoric seeds (H€orandl & Emadzade, 2012). Inaddition to these two clades, there are many riparian Ranunculusspecies in predominantly terrestrial clades. These species have feweradaptations to cope with inundation, and are accordingly found athigher riparian elevations (He et al., 1999).
III. Local and regional patterns in the distribution ofriparian plants
1. Community assembly
The framework of assembly rules can be used to understand theways in which plant adaptations to inundation, flood disturbanceand dispersal shape riparian plant community composition.Community assembly can be conceived as a series of sieves, ornested species pools, that are determined by dispersal, environ-mental and biotic constraints (Fig. 1, Belyea & Lancaster, 1999;G€otzenberger et al., 2012). These filters vary in space and time, andeach can be modified (Fig. 1). Although this conceptual modelrepresents community assembly as a sequential, one-way process(Fig. 1), this is primarily for clarity of presentation and compre-hension. The filters, and processes governing them, necessarilyinteract (see examples in Section V.5).
The first filter relates to dispersal. Acting on the regional speciespool, species’ fecundity and dispersal ability (such as capacity forhydrochory) determine the pool of potential colonists for a givensite at a particular point in time (Fig. 1). Once present in thegeographic species pool (sensu Belyea & Lancaster, 1999), a speciesmust pass the environmental filters of that site. Depending on thelocal conditions, a species may need to be able to tolerate prolongedinundation, desiccation or scour to become a member of theecological species pool (Fig. 1). Interspecific differences in toler-ance to physical disturbance, flooding and drawdown may lead tothe classic zonation of vegetation observed in riparian systems(Fig. 2). By grouping species with similar environmental niches,environmental filtering typically results in coexisting species havingecophysiological traits that are more similar than expected bychance (i.e. trait underdispersion or convergence; Weiher &Keddy, 1995b; de Bello, 2012). The third and final filter relates tobiotic interactions, representing the outcome of interactions(including competition, mutualism and predation) among indi-viduals in that community. In the following section, we illustrateways in which community assembly filtering can influence thespatial distribution of riparian plants at various spatial scales, fromlocal to regional, retaining our focus on inundation, flooddisturbance and hydrochory (Table 2).
2. Spatial patterns in riparian plant community composition
As mentioned earlier, riparian vegetation often forms zones alongan elevation gradient (Fig. 2), transitioning from terrestrial speciestolerant of only short periods of inundation, to amphibious species,to obligate aquatic species at the lowest elevations (Blom et al.,1994). This zonationmay be driven by species’ increasing toleranceof submergence (Vervuren et al., 2003; van Eck et al., 2006) or byspecies’ niche differences that relate to species’ competitive abilities
New Phytologist (2014) 204: 19–36 � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Review Tansley reviewNewPhytologist24

(Grace, 1988) and tradeoffs between tolerance of soil drying vstolerance of inundation (Silvertown et al., 1999; Table 2). As wellas differences in species’ characteristics, there are also differences in
the number of species along the elevation gradient, with speciesrichness generally declining towards lower elevations (Fig. 3),presumably reflecting that fewer species in the geographic speciespool are tolerant of inundation.
Changes in species’ occupancy with increasing distance from thechannel may be caused by other factors, such as probability ofpropagules arriving at a site or successional processes (Table 2).Merritt&Wohl (2006) found that the probability of encounteringhydrochoric species decreased from 90% at the edge of the streamchannel to 1% in areas that were 10 m from the stream channel.This could be because hydrochores are more likely to be depositedalong stream margins (dispersal filter) or because hydric speciestolerating conditions close to the channel tend to be water-dispersed (environmental filter).
Differences in community composition among patches or alonga reach can reflect variation in time since disturbance, where eachpatch undergoes succession with species being replaced viafacilitation or competition (biotic interactions). Likewise, distur-bance frequencymay affect species richness, withmaximum speciesrichness at intermediate disturbance frequency and site productiv-ity (Pollock et al., 1998).However, flood duration and intensity areusually highly correlated with frequency of disturbance (Hupp &Osterkamp, 1985), making it difficult to distinguish their effects.In a rare experimental test of community assembly, Weiher &Keddy (1995a) were able to replicate the segregation of plantcommunities found in riverine wetlands by manipulating floodduration and productivity.
Dispersal can result in differences in community compositionamong riparian zones at the reach scale. Surveys (Andersson et al.,2000b) and experiments (Jansson et al., 2005;Merritt et al., 2010a)show that species richness is higher on riverbanks that receive manyhydrochores. There are also more species with long-floatingpropagules in riparian than in terrestrial communities (Johansson
ECOSYSTEM CHARACTERISTICS
(a) Not applicable
(b) High rates of introduction because of: large edge effects; close proximity to human activities, e.g. agriculture; hydrochory as an additional dispersal opportunity.
(c) Wide range of environmental filters because of high temporal and spatial environmental heterogeneity.
(d) Frequent disturbance reduces strength of competition for resources.
INVADER CHARACTERISTICS
(a) Bias towards riparian specialists in the exotic species pool.
(b) Riparian invaders have higher propagule availability and greater dispersal capacity, e.g. because of a close association with humans.
(c) Riparian invaders can pass through a broader range of filters than other types of invaders.
(d) Riparian invaders more invasive than species that invade other ecosystems.
Regional species pool
Dispersal filter
Environmental filter
Biotic filter
Geographic species pool
Ecological species pool
Actual species pool
Local community assemblage
Fig. 1 Possible explanations for high degrees of invasion in riparian ecosystems. Explanations are based on characteristics of riparian ecosystems andcharacteristics of the invadingspecies (seeSection V).Explanation types: a, regional speciespool; b,dispersalfilter; c, environmental filter;d,bioticfilter.Thefiltersinteract and determine the composition of the geographic, ecological and actual species pools (sensu Belyea & Lancaster, 1999). Explanations are not mutuallyexclusive. Native species are white symbols and alien species grey symbols; circles, triangles and stars represent three hypothetical species’ functional groups.
(a)
(b)
Fig. 2 Zonationof riparianvegetationalonganelevationgradient. Eachzoneis characterized by a different type of vegetation, as illustrated by variation invegetation texture and colour with increasing wetland depth. (Source: J. A.Catford, River Murray wetlands, Victoria, Australia.)
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014) 204: 19–36
www.newphytologist.com
NewPhytologist Tansley review Review 25
et al., 1996) and more on riverbanks along tranquil than alongturbulent reaches (Nilsson et al., 2002), potentially because short-floating propagules are more likely to sink before reaching riparianzones.
Longitudinal patterns in riparian species richness have beenextensively studied, with variation in both patterns found andexplanations invoked. Several studies have documented peaks inspecies richness in mid-reaches (Nilsson et al., 1989; D�ecamps &Tabacchi, 1994), which has been interpreted both as an outcome ofintermediate disturbance and as dispersal along the river (Nilssonet al., 1994). The complexity of the gradient is demonstrated by thefact that diversity patterns have been observed to vary depending onthe type of riparian habitat studied and the time since major flooddisturbance (Ren€of€alt et al., 2005).
IV. Alien plant invasion in riparian zones
1. Expectations
It is well established that riparian zones experience high degrees ofalien plant invasion (Stohlgren et al., 1999; Richardson et al.,2007), whether that be in relation to the frequency of occurrence ofalien species (Vil�a et al., 2007), or absolute (Stohlgren et al., 1998)or relative (Catford et al., 2012b) alien species richness and cover.The reasons for high degrees of riparian invasion are usuallyinvestigated from an ecosystem perspective, where their geographyand environmental characteristics are primarily considered (Fig. 1,left-hand side). Common explanations include effective dispersalprovided by hydrochory, the high edge-to-area ratio of riparian
Table 2 Potential ways that inundation, disturbance and dispersal affect the distribution of riparian plants at different spatial scales
Scale Pattern Factor Hypothesized mechanismCommunityassembly filter References
Local Species separated alongelevation gradients
Inundation Differences among species in theirtolerance of inundation depth andduration
Environmentaland bioticfilters
Vervuren et al. (2003),van Eck et al. (2006)
Segregation of species as a result oftradeoffs between tolerance of soildrying and waterlogging
Biotic filter Silvertown et al. (1999)
Local Decreased speciesrichness with increasingduration of inundation
Inundation Fewer species can tolerate longperiods of inundation
Environmentalfilter
Fig. 3
Local More hydrochorousspecies close to streamchannels
Dispersal Hydrochorous propagules more likely tobe deposited close to the channel
Dispersal filter Merritt & Wohl (2006)
Reach Successional changes invegetation followingdisturbance
Disturbance Tradeoffs between species’ colonizationand competitive abilities mean that earlycolonizers are replaced by superiorcompetitors with increasing time sincedisturbance
Biotic filter Dahlskog (1966),Kalliola & Puhakka(1988)
Reach toLandscape
Higher species richness insites that receivehydrochorous propagules
Dispersal Hydrochory results in greater numbersof colonizing species
Dispersal filter Andersson et al. (2000b),Jansson et al. (2005)
Reach toLandscape
Maximum species richnessat intermediate floodfrequency and productivity
Disturbance Maximum species richness at intermediatedisturbance frequency and vegetationgrowth rates
Biotic filter Pollock et al. (1998)
Reach toLandscape
More species with long-floating propagules alongtranquil than turbulentreaches
Dispersal Species with short-floating propagules aremore likely to sink before being depositedalong tranquil reaches
Dispersal filter Nilsson et al. (2002)
Landscape More species have propaguleswith floating capacity in ripariancommunities than uplandcommunities
Dispersal Hydrochorous species more likely tocolonize riparian communities
Dispersal filter Johansson et al. (1996)
More species with capacity for hydrochoryin riparian zones
Environmentalfilter
Regional Peaks in species richness inthe mid-reaches of rivers
Disturbance Intermediate disturbance in mid-reaches Environmentalfilter
Nilsson et al. (1989),D�ecamps & Tabacchi(1994)
Dispersal;disturbance
Diversity of hydrochorous propagulesincreases downstream, but higherdisturbance intensity limits speciesestablishment in lower reaches
Dispersal andenvironmentalfilters
Nilsson et al. (1994)
Monotonic decrease inspecies richness towardslower reaches
Disturbance Increasing disturbance intensity fromfloods
Environmentalfilter
Ren€of€alt et al. (2005)
We list likelymechanisms andassociated community assemblyfilters (Fig. 1) for the spatial patternsobserved.References relate to theobservedpatterns, not tothe hypothesized mechanisms, factors or community filters.
New Phytologist (2014) 204: 19–36 � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Review Tansley reviewNewPhytologist26

zones, high habitat heterogeneity, frequent flood disturbance, highhuman use and close proximity to human activities (Richardsonet al., 2007; Catford et al., 2011; Eschtruth & Battles, 2011). Eachof these factors can facilitate invasion by modifying communityassembly (Section III.1, Fig. 1). Structured around communityassembly processes and considering riparian invasion from theperspective of the invaders, we proffer four explanations for highdegrees of riparian invasion (Fig. 1, right-hand side):
Hypothesis 1.Ahigh proportion of alien species in the regional species pool
preferentially invade riparian zones. Species specifically adapted to
riparian ecosystems may be overrepresented in the alien species pool
because of an introduction bias, leading to higher alien species richness in
riparian ecosystems.
Hypothesis 2. Alien species are more likely to invade riparian ecosystems
because: dispersal is more effective in river networks than in other parts of
landscapes; species tend to have dispersal characteristics conducive to
dispersal in river networks, namely hydrochory; or riparian invaders have
a stronger association with humans than with other species. By planting
and transporting certain plant species in high numbers, humans can
increase the dispersal opportunities of species in a way that is independent
of their traits (Catford et al., 2012a). Regardless of the cause, increased
dispersal may allowmore alien species to invade riparian zones (i.e. higher
colonization pressure, sensu Lockwood et al., 2009) or it may increase the
frequency and abundance of particular species that havemany propagules
(i.e. propagule pressure, sensu Lockwood et al., 2009).
Hypothesis 3. More alien species may be able to pass through the
environmental filters and invade riparian ecosystems compared with
other ecosystem types, as a result of, for example, the high environmental
heterogeneity of riparian ecosystems.
Hypothesis 4. Species that invade riparian ecosystems may reach higher
abundance than species that invade other ecosystems. If the degree of
invasion is based on total or per capita alien abundance (Catford et al.,
2012b), higher occurrence of these species will result in riparian zones
having higher degrees of invasion.
Using plant community composition data collected acrossalmost 30 000 quadrats in Victoria, Australia (SupportingInformation, Table S1), and information on 458 higher plantspecies in the Global Invasive Species Database (GISD; ISSG,2013), we assess the likelihood of these four explanations(Table S2). A description of the datasets and analysis is providedin Notes S1.
2. Findings
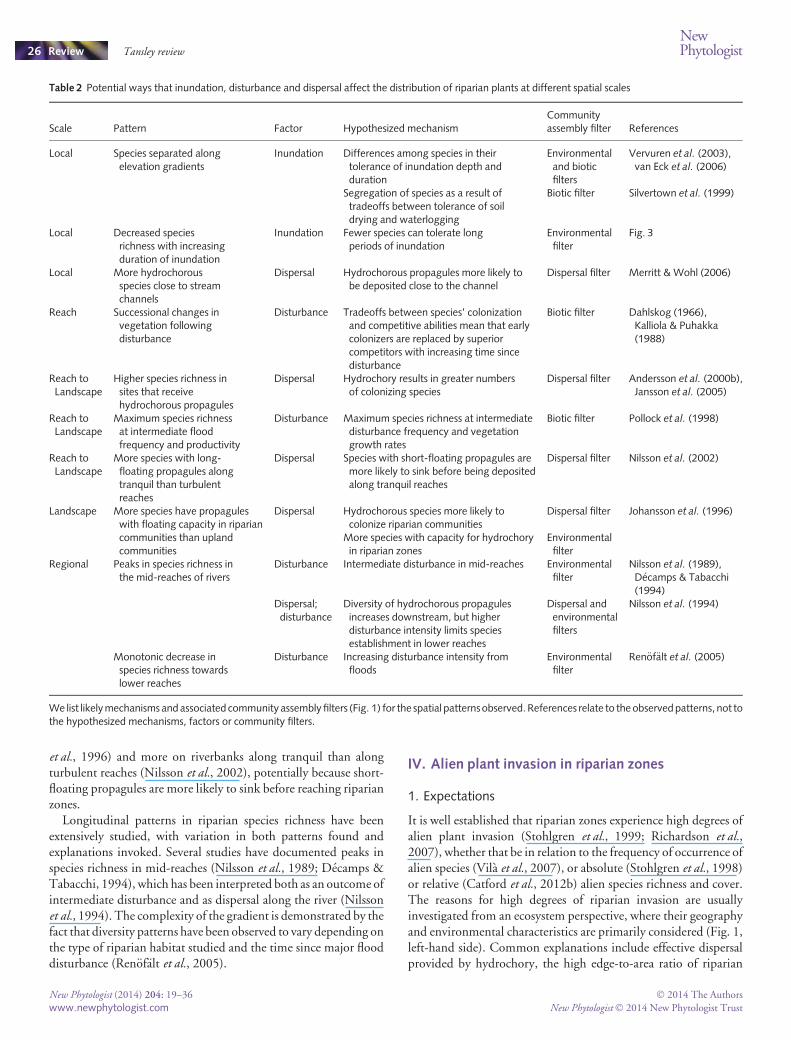
Hypothesis 1 Few alien species exclusively invade riparianecosystems in either of the datasets examined. Species specificallyadapted to riparian environments do not appear to be overrepre-sented in the pool of potential invaders (the regional species pool,Fig. 1). Across the 458 higher plants listed in theGISD, two speciesinvade riparian zones and wetlands exclusively. In Victoria, 10 of1036 alien species exclusively invade theRiparian Scrubs ecosystem(24 species exclusively invade the Broad Riparian category, whichcombines three ecosystem types, Notes S1).
Hypothesis 2 Information about the modes of dispersal used byinvaders in the GISD suggests that riparian invaders disperse morewidelyandinhighernumbersthanothertypesofinvaders(TableS2).They may therefore have higher colonization and propagulepressure than species that invade other ecosystems. Of 11 possiblemodes of dispersal, riparian invaders usemore dispersalmodes thanspecies that do not invade riparian ecosystems (Fig. 4, Table S3).
Compared with species that do not invade riparian zones, agreater proportion of riparian invaders are dispersed by hydrochory(riparian invaders (mean� SE), 0.41 � 0.04; non-riparian invad-ers, 0.24� 0.03; P < 0.001, Table S4) and humans (riparianinvaders, 0.59� 0.038; non-riparian invaders, 0.42� 0.029;P < 0.001, Table S5). Similar trends were foundwhen comparisonswere made using species that invade riparian zones and wetlands(i.e. Broad Riparian category).
Hypothesis 3 Across both datasets, a high proportion of the alienor invasive species pool invades riparian zones. Based on theproportion of the invasive species pool invading any givenecosystem type (excluding Broad Riparian), riparian zones wereranked fourth in the GISD (Fig. 5a) and third in the Victorian data(Fig. 5b; second and first, respectively, if Broad Riparian was used).For the GISD, 37% of invaders are known to invade riparian zonesand 47% to invade riparian zones or wetlands. In the Victoriandataset, 50%of the alien species recordedwere observed inRiparian
Amphibianspecies(22 sp.)
Riparianforest
(60 sp.)
Willows
(64 sp.)
Gram-inoids(36 sp.)
Upland
(46 sp.)
20
15
10
100500–50–100–150–200–250–300–350
5
0
Elevation (cm)
Num
ber o
f spe
cies
per
plo
t
Fig. 3 Species richness in riparian zones declines at lower elevations as floodfrequency and duration of inundation increase. The number of riparian plantspecies was recorded in 334 quadrats (0.25m2, open circles and trendline),and the cumulative number of species per vegetation belt (hatched lines andnamesat the topof thefigure) areplottedagainst theelevation in the riparianzone. 0 indicates the elevation of the annual high-water mark. Data werecollected along a 200-m-long slow-flowing reach (Lappselet) of the free-flowing Vindel River in northern Sweden. Details of the sampling design canbe found in Str€om et al. (2012). The trendline is drawn with LOWESSregression in SPSS v. 21 (IBM Corp., Armonk, NY, USA), using a movingwindow that includes 50% of the points.
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014) 204: 19–36
www.newphytologist.com
NewPhytologist Tansley review Review 27

Scrubs, and 69% of the alien species occur across the threeecosystems that make up the Broad Riparian category (Fig. 5b).
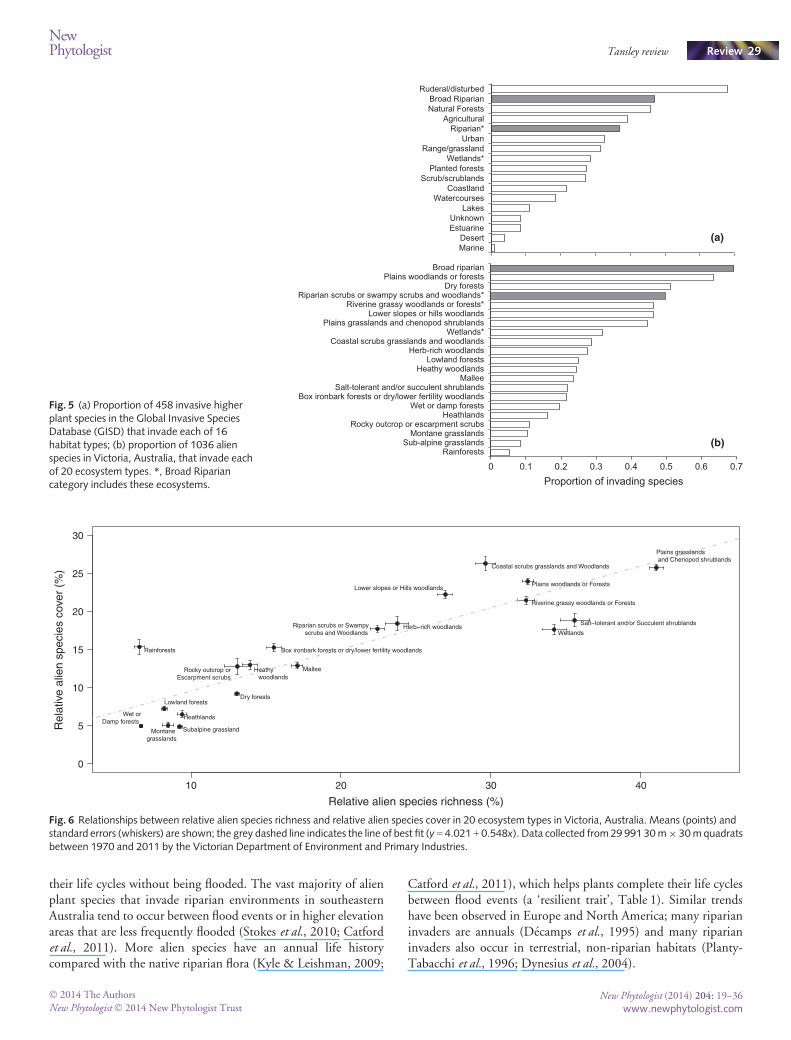
Hypothesis 4 As an indication of invasion success, we used theVictorian data to determine whether alien species in riparian zonescontribute more cover relative to their richness than invaders inother types of ecosystems. Across the 20 ecosystem types, alienspecies that invade Riparian Scrubs contribute slightly more thanaverage to total vegetation cover (i.e. they fall above the trendline inFig. 6), although Wetlands and Riverine Grassy Woodlands (theother vegetation classes that can be considered riparian) do not. Thisevidence suggests that high degrees of invasion in these riparianecosystems are not caused by riparian invaders being more invasiveand attaining higher cover than species that invade other ecosystems.
3. Synopsis
Based on our examination of the GISD and the Victoria data,species that invade riparian ecosystems may have higher coloniza-tion and propagule pressure (Hypothesis 2,H2) and are able to passthrough a broader range of environmental filters (H3) than species
that invade other ecosystems. Although additional data on speciestraits and environmental conditions are required to ascertainsupport for H3, H2 and H3 seem more likely to explain highriparian invasion than those based on a bias towards riparianspecialists in the alien regional species pool (H1) and riparianinvaders being more invasive than invaders of other ecosystems(H4).We recognize that this analysis is limited to two datasets, butthese findings appear to be in agreement with observed trends.
Although there are exceptions, observations suggest thatrelatively few alien invaders occur inmid-elevation areas of riparianzones. In River Murray wetlands, Australia, only six of 57 alienspecies observed were classified as amphibious, inhabiting mid-elevations, compared with 38 out of 87 native species (Catfordet al., 2011). Characterized by frequent flood disturbance andpotentially long periods of both inundation and drying, this part ofthe riparian zone may represent both high stress and highdisturbance, arguably an unusual combination of conditionsrequiring specialized adaptations (Grime, 1977; Westoby, 1998).
Rather than having specialized adaptations to cope withfluctuating water levels and flood disturbance, most riparianinvaders may be temporally or spatially opportunistic, completing
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7
Contaminated
Forestry*
Agriculture*
Wind
Unspecified
Vehicles*
Vegetative
Hikers*
Ornamental*
Garden escape*
Water currents
Animals
Human-mediated
Proportion of invasive species
Riparian invadersNonriparian invaders
Num
ber o
f dis
pers
al m
odes
9
8
7
6
5
4
3
2
1
0(a)
(b)
Fig. 4 (a) Number of dispersal modes used byinvasive higher plant species in the GlobalInvasive Species Database (GISD) that invaderiparian ecosystems and species that do not(P < 0.001; Table S3; box shows interquartilerange with the median as a horizontal line;whiskers showthehighest and lowest values inthe dataset). (b) Proportion of riparianinvaders (grey) and non-riparian invaders(white) that use different dispersal modes.*, dispersal modes that are included as a formof human-mediated dispersal.
New Phytologist (2014) 204: 19–36 � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Review Tansley reviewNewPhytologist28
their life cycles without being flooded. The vast majority of alienplant species that invade riparian environments in southeasternAustralia tend to occur between flood events or in higher elevationareas that are less frequently flooded (Stokes et al., 2010; Catfordet al., 2011). More alien species have an annual life historycompared with the native riparian flora (Kyle & Leishman, 2009;
Catford et al., 2011), which helps plants complete their life cyclesbetween flood events (a ‘resilient trait’, Table 1). Similar trendshave been observed in Europe and North America; many riparianinvaders are annuals (D�ecamps et al., 1995) and many riparianinvaders also occur in terrestrial, non-riparian habitats (Planty-Tabacchi et al., 1996; Dynesius et al., 2004).
MarineDesert
EstuarineUnknown
LakesWatercourses
CoastlandScrub/scrublands
Planted forestsWetlands*
Range/grasslandUrban
Riparian*Agricultural
Natural ForestsBroad Riparian
Ruderal/disturbed
Proportion of invading species0 0.1 0.2 0.3 0.4 0.5 0.6 0.7
RainforestsSub-alpine grasslands
Montane grasslandsRocky outcrop or escarpment scrubs
HeathlandsWet or damp forests
Box ironbark forests or dry/lower fertility woodlandsSalt-tolerant and/or succulent shrublands
MalleeHeathy woodlands
Lowland forestsHerb-rich woodlands
Coastal scrubs grasslands and woodlandsWetlands*
Plains grasslands and chenopod shrublandsLower slopes or hills woodlands
Riverine grassy woodlands or forests*Riparian scrubs or swampy scrubs and woodlands*
Dry forestsPlains woodlands or forests
Broad riparian
(a)
(b)
Fig. 5 (a) Proportion of 458 invasive higherplant species in the Global Invasive SpeciesDatabase (GISD) that invade each of 16habitat types; (b) proportion of 1036 alienspecies in Victoria, Australia, that invade eachof 20 ecosystem types. *, Broad Ripariancategory includes these ecosystems.
10 20 30 40
0
5
10
15
20
25
30
Relative alien species richness (%)
Rel
ativ
e al
ien
spec
ies
cove
r (%
)
Box ironbark forests or dry/lower fertility woodlands
Coastal scrubs grasslands and Woodlands
Heathlands
Plains woodlands or Forests
Riverine grassy woodlands or Forests
Salt−tolerant and/or Succulent shrublands
Subalpine grassland
Heathy woodlands
Plains grasslands and Chenopod shrublands
Lowland forests
Wet or Damp forests
Montane grasslands
Lower slopes or Hills woodlands
Herb−rich woodlands
Rocky outcrop or Escarpment scrubs
Wetlands
Mallee
Dry forests
Rainforests
Riparian scrubs or Swampyscrubs and Woodlands
Fig. 6 Relationships between relative alien species richness and relative alien species cover in 20 ecosystem types in Victoria, Australia. Means (points) andstandard errors (whiskers) are shown; the grey dashed line indicates the line of best fit (y = 4.021 + 0.548x). Data collected from 29 991 30m9 30mquadratsbetween 1970 and 2011 by the Victorian Department of Environment and Primary Industries.
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014) 204: 19–36
www.newphytologist.com
NewPhytologist Tansley review Review 29

Riparian zones may be highly invaded at the ecosystem level, butit does not seem that all areas (i.e. elevations) within riparian zonesare highly invaded. The high spatial and temporal heterogeneity ofriparian zones may thus help to explain the apparent conundrumbetween high degrees of invasion and adaptations required to live inriparian zones. Even if invading species are unable to tolerate someof the environmental conditions, the high diversity of niche space ofriparian ecosystems means that many different life strategies areviable, providing many opportunities for invasion.
V. Modification and management of riparian plantcommunities
As well as the addition of alien species to regional species pools,community composition can change through modification of anyof the three community assembly filters. Significant changes to thedispersal, environmental or biotic filters are likely to prompt achange in the composition of the associated species pools and,consequently, the resultant community (Fig. 7). In the following,we briefly discuss some ways in which these filters can be modifiedby human activities in ways that affect riparian ecosystems (Fig. 7).
1. Changes to the regional species pool
Species from other regions are often added to the regional speciespool through the actions of humans: alien species may be importedas a commodity, theymay arrive with transport, or theymay spreadfrom another invaded region (Hulme et al., 2008). Global patternssuggest that most alien plants escape into natural environmentsafter being intentionally introduced, usually for ornamentalpurposes (Hulme et al., 2008; Dehnen-Schmutz, 2011). Theconstruction of inland waterways and removal of dispersal barriers
can effectively provide new corridors for dispersal (Wilson et al.,2009), enabling species from different biogeographic regions tomix.
Loss of native species from the regional species pool implies thatthey are lost locally first (which would result from changes to thethree filters, discussed later). Species adapted to frequent distur-bance tend to have propagules with greater longevity (Bossuyt &Honnay, 2008), so riparian specialists may persist for longer,although that is not always the case, as the short seed viability of theendangered Japanese floodplain species, Aster kantoensis, attests(Washitani et al., 1997).
2. Changes to dispersal filters
Changes in habitat connectivity, the location and size of propagulesources and vector availability can alter species’ dispersal patternsand the composition of the geographic species pool (Belyea &Lancaster, 1999). The ability of species’ propagules to reach a givensite is crucial, as is the number of propagules that arrive because thiscan effectively increase a species’ chance of colonization anddominance (Pacala & Tilman, 1994; Simberloff, 2009). It istherefore of little surprise that dispersal and propagule pressurefacilitate invasions, as noted earlier.
Reflecting the importance of hydrochory for riparian plants, lossof hydrological connectivity can limit the dispersal ofmany species.Lateral connectivity (river floodplain) can be impeded through theconstruction of walls and levees. Dams, weirs and associatedimpoundments inhibit downstream transport of hydrochores(Andersson et al., 2000a; Merritt & Wohl, 2006), potentiallyaltering species composition andprompting the loss of some speciesbelow dams (Jansson et al., 2000). As well as reductions inconnectivity, some habitats may become more connected. Roads
Regional species pool
Dispersal filter
Environmental filter
Biotic filter
TYPES OF MODIFICATION
(a) Exotic species introduction and loss of native species.
(b) New dispersal barriers, e.g. dams, levees; changes in propagule sources and vectors, e.g. humans.
(c) Environmental modification, e.g. flow regulation, climate change, urbanization.
(d) Changes in competition, mutualisms and predation from changes in species occupancy and abundance.
REMEDIATION STRATEGIES
(a) Limit exotic introduction; assisted colonization to restock natives.
(b) Restore connectivity; limit human-mediated dispersal; maintain buffer between agriculture/gardens and riparian ecosystem.
(c) Amelioration of environmental change, e.g. environmental flows, salt interception, water-sensitive urban design.
(d) Limit native species harvest, e.g. logging, hunting; control of invasive species.
Geographic species pool
Ecological species pool
Actual species pool
Local community assemblage
Fig. 7 Ways in which human activities can alter community composition in riparian ecosystems through changes to the (a) regional species pool and the (b)dispersal, (c) environmental and (d) biotic filters. Thefilters interact anddetermine the compositionof thegeographic, ecological and actual species pools (sensuBelyea& Lancaster, 1999). Native species arewhite symbols and alien species are grey symbols; circles, triangles and stars represent three hypothetical species’functional groups.
New Phytologist (2014) 204: 19–36 � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Review Tansley reviewNewPhytologist30

and walking tracks can act as efficient corridors for dispersal (vonder Lippe & Kowarik, 2012) and may connect anthropogenichabitats with riparian zones. Humans disperse plant propagules byusing and travelling through riparian environments (Hodgkinson&Thompson, 1997). Species that frequently occur around areas ofhuman activity (e.g. weeds, crops, ornamentals) will be dispersedmore frequently and in greater numbers (vonder Lippe&Kowarik,2012).
Habitat transformation can reduce the input of nativeriparian species’ propagules whilst increasing the propaguleavailability of species normally absent or scarce in riparian zones,such as alien species (Niggemann et al., 2009). Floodplains areoften used for livestock grazing or crop production, and organicwaste from gardens and agriculture is frequently dumped inriparian zones. Centres of human activity thus provide a largeand ready source of propagules, the impact of which isheightened by the large edge effects typical of riparian zones,making them particularly susceptible to species invasion fromsurrounding environments. Propagule dispersal that coincideswith peak flows can be interrupted by changes in flow regimes,inhibiting dispersal and reducing seedling survival (Rood &Mahoney, 1990).
3. Changes to environmental filters
Abiotic conditions in riparian zones can be altered throughactivities like eutrophication, pollution, urbanization, salinization,climate change and flow regulation. Reflecting the focus of thispaper on inundation and flood disturbance, we concentrate on flowregime modification.
Damming of rivers has modified the amount, timing andtype of water flow altering river hydrology, hydraulics, geomor-phology and ecology (Rood et al., 1999). Given the stronginfluence of river flow regimes on riparian plants, hydrologicalmodification is a key explanation for changes in the compositionof riparian vegetation (Dynesius et al., 2004; Merritt et al.,2010b).
In its simplest form, community composition can change withenvironmental modification in two ways (Catford et al., 2011).First, native species specifically adapted to the historical regimemaybe unable to toleratemodified conditions (Evangelista et al., 2008),prompting a decline in the vigour and size of their populations.Abundance of alien generalists may increase with any abioticchange provided it is to the detriment of its competitors. Such ascenario may have occurred along the upper Rhine River, France,where a reduction in flooding facilitated invasion of generalistspecies (Deiller et al., 2001).
Secondly, if alien species are preadapted (or adapt rapidly;Whitney&Gabler, 2008) to the modified conditions, they may beable to outcompete native species that are poorly adapted to the newconditions. Invasion success will be greatestwhen a decline in nativespecies is coupled with environmental conditions that favour alienspecialists. For example, the success of alien Tamarix species alongregulated rivers in southwesternUSA is at least partially attributableto their superior adaptation to modified conditions (Stromberget al., 2007).
4. Changes to biotic filters
Biotic interactions within a community will change as theoccupancy and abundance of species change. Such changes canbe a result of human actions that directly affect biota and theirinteractions (e.g. logging, hunting, livestock grazing), but mostchanges probably result indirectly through the modification of theother filters. Working in the Eastern Sierra Nevada Mountains ofCalifornia, Elderd (2003) found that reduced flooding below damsprompted an increase in non-riparian herbaceous cover, grassthatch and shading, which led to lower germination success andhigher herbivory ofMimulus guttatus (common monkeyflower), acommon riparian species.
5. Feedbacks and interactions among filters
As well as changes that occur outside of a community (e.g. humanactivities), feedback effects from the resident community mayamplify changes to the three community assembly filters and,consequently, to community composition. Some plants canmodify abiotic characteristics, effectively changing environmentalfilters in their favour (Richardson et al., 2000). For example, innorthern Australia, invasive Urochloa mutica (para grass) has beenobserved to decrease stream channel capacity by up to 85% (Bunnet al., 1998), increasing the frequency of overbank flows, whichprobably help the species to spread.
6. Remediation and amelioration of impacts
Many of the changes to the structure and composition of riparianplant communities can be ameliorated through strategic manage-ment, as illustrated in the following sections.
Regional species pool Limiting the transport and introduction ofspecies through quarantine and screening would help to limit thenumber of alien species in the regional species pool. Assistedcolonization may be used to reintroduce native species that havebecome locally extinct, although it would be essential to firstaddress the cause of their initial decline.
Dispersal filters Ensuring sufficient habitat connectivity is crucialformaintainingviablepopulationsofnativespecies.Physicalstructuresthat fragment riverine ecosystems, including structures between theriverchannelandtheriparianzone,canberemovedortheirdesignandconstructionimproved(e.g.retrofittingdams(Bernhardtet al.,2005),leveebreachingandsetback(Palmeret al.,2005)).Thenumberofalienpropagules, and the frequency by which they are dispersed, can bereduced through preferential use of native pasture or ornamentalplants, appropriate disposal of organic waste, separation of urban oragricultural land fromriparianzoneswithbuffer strips (Catford et al.,2011),andcleaningvehicles, clothingandequipment before movinglocations (Pickering&Mount, 2010).
Environmental filters Recognizing the need to rectify some of thedamage caused by changes in river flow regimes, the use ofenvironmental flows (Arthington et al., 2006) and habitat
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014) 204: 19–36
www.newphytologist.com
NewPhytologist Tansley review Review 31

restoration (Jungwirth et al., 2002) have received growing support.Flows that restore crucial elements of the historical flooding regimeshould help redress the decline in native species and the increase inaliens observed in many river systems (Rood et al., 2003; Catfordet al., 2011). Determining the characteristics of optimal environ-mental flows remains a challenge (Arthington et al., 2006) andenvironmental flows are not without risk (e.g. risk of invasivespecies spread; Howell & Benson, 2000).
Biotic filters Biotic interactions can be directly addressed throughthe management of species populations. Limiting the decline ofnative species populations could involve: ensuring healthy popu-lations of mutualists (e.g. pollinators, endophytes); limitingcompetition from, and predation by, alien species; and reducingspecies harvest, which would help to maintain ‘natural’ food webstructures and trophic interactions.
Invasive plant species populations can be directly managedthrough physical removal and biological and chemical control.Integrated approaches are often more effective than using a singletechnique (Rea&Storrs, 1999). Suitable controlmeasures will varydepending on the target species, the stage of invasion, the resourcesavailable and the characteristics of the site. For example,mechanicalcontrol can be difficult in dense vegetation with poor access, andherbicide use is often restricted around waterways. As with otherforms of population management, indirect impacts of alien speciescontrol should be considered in decision-making (Flory & Clay,2009).
VI. Conclusions and future research
1. Plant traits and vegetation strategies
At least 35 traits enable plants to live in riparian ecosystems(Table 1), many of which have not evolved specifically for riparianconditions. The fact that there are numerous ways of sustaining lifein riparian zones (e.g. tolerating or avoiding stress) helps to explainwhy riparian environments can be rich in plant species despitebeing challenging environments to inhabit.
The high heterogeneity of riparian zonesmakes them suitable fornative and alien species with many different life strategies,representing a range of niches (Grubb, 1977). In the terminologyof Grime’s (1977) CSR model, which classifies plant species intocompetitors, stress tolerators and ruderals, riparian zones areinhabited by stress tolerators coping with inundation, ruderal speciesadapted to frequent disturbance, and competitors that are able tooutcompete other species where (and when) disturbance andinundation stress are low. However, many riparian species seem topossess traits that enable survival of both high stress and highdisturbance, a combination of conditions that Grime (1977) saw asdifficult to combine. Plant taxa such as Typha, Carex andPhragmites avoid anoxic stress by having aerenchyma and oxygen-ation of the rhizosphere, but they also have traits associated withdisturbance adaptation, like deep and wide root systems, andvegetative reproduction. Moreover, species with high growth ratesthat form tall, monospecific stands (such as Typha and Phragmites)possess characteristics typical of competitors (Grace, 1988).
Clearly, further research into traits of riparian species is required:to identify physiological tradeoffs pertinent to riparian environ-ments; to determine whether there are combinations, or suites, oftraits associated with different riparian plant strategies; and to testwhether there are differences in the prevalence of certain plant traitsand strategies among stream types and regions. Given that nativeand alien species are likely to come from different clades,comparing traits possessed by natives and aliens may help todistinguish between traits that facilitate survival from traits that areprimitive and shared by all species in a clade.
Westoby’s (1998) leaf-height-seed scheme provides an easy andtractable way to ascertain the prevalence of different CSR strategiesin terrestrial environments, but a suitable scheme is not yet availablefor riparian zones. Because amphibious and aquatic plants requireless structural support than terrestrial plants, theway inwhich plantheight or specific leaf area should be interpreted is unclear. Howshould plant height be understood for species that are prostrate interrestrial conditions, but floating when flooded? Heterophyllyprovides similar challenges for leaf ecophysiologicalmeasurements,as does the lack of tradeoff between seedmass and dispersal distancein hydrochores (Ikeda & Itoh, 2001). Identifying plant traits thatare comparable across the different conditions and elevationswithin riparian ecosystems, as well between riparian and terrestrialecosystems, is an area that is ripe for research.
2. Evolution of riparian flora
The distribution of riparian traits have been mapped ontophylogenies in only few clades, so differences and similarities intrait evolution among clades remain unknown. For example, havesome traits evolved more often than others? In clades with multipleadaptations, have traits evolved in a specific sequence, wherepossession of one trait enables acquisition of another? Are theredifferences in the proportion of species in regional species pools thatcan occupy riparian zones? In regions with humid climates, wherebogs, swamps and other wet habitats are common, large propor-tions of the regional species pools are represented in riparian zones(Dynesius et al., 2004). Although quantitative comparisons arelacking, in more arid regions, where conditions in riparian zonesdiffer markedly from matrix habitats, it seems that a smallerproportion of the regional species pool is able to cope with riparianconditions and these taxa tend to be confined to specific clades.Comparing the traits and phylogenetic relatedness of native andalien species will provide deeper insights into the factors that shaperiparian communities.
3. Community assembly processes
The distribution of plants within riparian zones, and the traits thoseplants possess, reflects processes that occur across all levels of thecommunity assembly hierarchy, from the composition of theregional species pool down to local community dynamics. Moststudies of community assembly processes focus on the effects ofenvironmental filtering and biotic interactions (Weiher et al.,1998) and ways in which to disentangle them (de Bello, 2012).Despite the widely acknowledged importance of dispersal filters
New Phytologist (2014) 204: 19–36 � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Review Tansley reviewNewPhytologist32

(G€otzenberger et al., 2012), relatively few studies specificallyexamine their effects on community assembly and the trait-basedpatterns that might emerge (Middleton, 2000). In this review, wefound many patterns that could be explained by dispersal-relatedprocesses, highlighting the importance of regional- and landscape-level processes in determining local species composition in riparianecosystems. Similar to environmental filtering, dispersal filterspresumably lead to underdispersion of traits related to dispersal andreproductive output. Given the prominent use of hydrochory andthe strong structuring forces of flooding and inundation in riparianecosystems, riparian zones seem to be suitable systems for studiesthat aim to disentangle the effects of dispersal and environmentalfilters.
4. Alien plant invasion and ecosystem management
Relatively few alien species appear to be specifically adapted toriparian conditions, but many alien species are still able to invaderiparian ecosystems. In general, a high proportion of introducedand invasive species can be classified as ruderals (Catford et al.,2012a), a suitable life strategy in ecosystems with frequentdisturbance. Because early succession is not limited by competitionfor resources, an infinite number of species can theoretically coexistprovided they are able to cope with disturbance (Catford et al.,2012a). However, as our analyses have shown, high alien speciesrichness does not necessarily equate with high alien abundance ordominance. This is presumably because aliens are largely restricted– in space and time – to early successional patches that are usuallydry (i.e. flooding and inundation in these areas would usually resetsuccession). Whether it is accurate to describe riparian ecosystemsas highly invaded, as is often the case, depends upon themetric usedand the community characteristics of interest. Hence, it is essentialthat suitable metrics are selected and clearly reported in studies.
Understanding species invasions through the prism of commu-nity assembly can highlight the various factors that may facilitate,drive and inhibit invasion. Determining the relative importance ofdifferent filters in facilitating or limiting species invasions will helpto guide suitable management responses to invasive plants, as wellas other ecological problems.However, identifying the chief driversof riparian invasion can be difficult because inferences are oftenbased on surveys where important environmental variables areconfounded (Catford et al., 2014). Hierarchical approaches thatcombine information about species traits with data about speciesdistributions along environmental gradients can help to increasethe diagnostic power of surveys (Catford et al., 2014).
Acknowledgements
We thank Alistair Hetherington for encouraging us to write thisreview,SamanthaDawsonforcommentsonanearlierdraft,ShyamaPagad and the IUCN SSC Invasive Species Specialist Group whogave us access to some of the data in the GISD,MattWhite and theVictorian Department of Environment and Primary Industries forpermission to use their data, Mary Gardner for helping process theGISD data and Est�ıbaliz Palma for helping to analyse the Victoriandata. We are grateful for feedback provided by the editor and three
anonymous reviewers. This work was supported by grants from theSwedish Research Council Formas (to R.J.) and the AustralianResearch Council (DE120102221 to J.A.C.).
References
Andersson E, Nilsson C, Johansson ME. 2000a. Effects of river fragmentation on
plant dispersal and riparian flora.Regulated Rivers – Research andManagement 16:83–89.
Andersson E, NilssonC, JohanssonME. 2000b. Plant dispersal in boreal rivers and
its relation to the diversity of riparian flora. Journal of Biogeography 27: 1095–1106.
APG II. 2003.An update of the Angiosperm PhylogenyGroup classification for the
orders and families of flowering plants: APG II. Botanical Journal of the LinneanSociety 141: 399–436.
APGIII. 2009.Anupdate of theAngiospermPhylogenyGroup classification for the
orders and families of flowering plants: APG III. Botanical Journal of the LinneanSociety 161: 105–121.
Armstrong W, Br€andle R, Jackson MB. 1994.Mechanisms of flood tolerance in
plants. Acta Botanica Neerlandica 43: 307–358.Arthington AH, Bunn SE, Poff NL, Naiman RJ. 2006.The challenge of providing
environmental flow rules to sustain river ecosystems. Ecological Applications 16:1311–1318.
BagstadKJ, Stromberg JC, Lite SJ. 2005.Response of herbaceous riparian plants to
rain and flooding on the San Pedro River, Arizona, USA.Wetlands 25: 210–223.Bailey-Serres J, Voesenek LACJ. 2008. Flooding stress: acclimations and genetic
diversity. Annual Review of Plant Biology 59: 313–339.Barrat-Segretain MH, Henry CP, Bornette G. 1999. Regeneration and
colonization of aquatic plant fragments in relation to the disturbance frequency of
their habitats. Archiv f€ur Hydrobiologie 145: 111–127.Barton DR, Taylor WD, Biette RM. 1985. Dimensions of riparian buffer strips
required to maintain trout habitat in southern Ontario (Canada) streams. NorthAmerican Journal of Fisheries Management 5: 364–378.
BeismannH,WilhelmiH, Bailleres H, Spatz HC, Bogenrieder A, Speck T. 2000.
Brittleness of twig bases in the genus Salix: fracture mechanics and ecological
relevance. Journal of Experimental Botany 51: 617–633.de Bello F. 2012. The quest for trait convergence and divergence in community
assembly: are null-models the magic wand? Global Ecology and Biogeography 21:312–317.
Belyea LR, Lancaster J. 1999.Assembly rules within a contingent ecology.Oikos 86:402–416.
Bernhardt ES, Palmer MA, Allan JD, Alexander G, Barnas K, Brooks S, Carr J,
Clayton S, Dahm C, Follstad-Shah J et al. 2005. Synthesizing U.S. riverrestoration efforts. Science 308: 636–637.
Blom CWPM, Voesenek LACJ. 1996. Flooding: the survival strategies of plants.
Trends in Ecology and Evolution 11: 290–295.Blom CWPM, Voesenek LACJ, Banga M, Engelaar WMHG, Rijnders JHGM,
Vandesteeg HM, Visser EJW. 1994. Physiological ecology of riverside
species: adaptive responses of plants to submergence. Annals of Botany 74:
253–263.BoedeltjeG,Bakker JP, TenBrinkeA,VanGroenendael JM, SoesbergenM. 2004.
Dispersal phenology of hydrochorous plants in relation to discharge, seed release
time and buoyancy of seeds: the flood pulse concept supported. Journal of Ecology92: 786–796.
Boeger MRT, Poulson ME. 2003.Morphological adaptations and photosynthetic
rates of amphibious Veronica anagallis-aquatica L. (Scrophulariaceae) underdifferent flow regimes. Aquatic Botany 75: 123–135.
Bornette G, Tabacchi E, Hupp C, Puijalon S, Rostan JC. 2008. A model of plant
strategies in fluvial hydrosystems. Freshwater Biology 53: 1692–1705.Bossuyt B, Honnay O. 2008.Can the seed bank be used for ecological restoration?
An overview of seed bank characteristics in European communities. Journal ofVegetation Science 19: 875–884.
BrownRL, Peet RK. 2003.Diversity and invasibility of southernAppalachian plant
communities. Ecology 84: 32–39.Bunn SE, Davies PM, Kellaway DM, Prosser IP. 1998. Influence of invasive
macrophytes on channel morphology and hydrology in an open tropical lowland
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014) 204: 19–36
www.newphytologist.com
NewPhytologist Tansley review Review 33

stream, and potential control by riparian shading. Freshwater Biology 39: 171–178.
Capon SJ. 2003. Plant community responses to wetting and drying in a large arid
floodplain. River Research and Applications 19: 509–520.Catford JA, Daehler CC, Murphy HT, Sheppard AW, Hardesty BD, Westcott
DA, Rejm�anek M, Bellingham PJ, Pergl J, Horvitz CC et al. 2012a. Theintermediate disturbance hypothesis and plant invasions: implications for species
richness and management. Perspectives in Plant Ecology, Evolution and Systematics14: 231–241.
Catford JA, Downes BJ, Gippel CJ, Vesk PA. 2011. Flow regulation reduces native
plant cover and facilitates exotic invasion in riparian wetlands. Journal of AppliedEcology 48: 432–442.
Catford JA, Morris WK, Vesk PA, Gippel CJ, Downes BJ. 2014. Species and
environmental characteristics point to flow regulation and drought as drivers of
riparian plant invasion. Diversity and Distributions. doi: 10.1111/ddi.12225.Catford JA, Vesk PA, Richardson DM, Py�sek P. 2012b.Quantifying levels of
biological invasion: towards the objective classification of invaded and invasible
ecosystems. Global Change Biology 18: 44–62.Chesson P, Huntly N. 1997. The roles of harsh and fluctuating conditions in the
dynamics of ecological communities. The American Naturalist 150: 519–553.Colmer TD, Voesenek LACJ. 2009. Flooding tolerance: suites of plant traits in
variable environments. Functional Plant Biology 36: 665–681.Cook CD. 1999. The number and kinds of embryo-bearing plants which have
become aquatic: a survey.Perspectives in Plant Ecology, Evolution and Systematics 2:79–102.
Crawford RMM. 1987. Plant life in aquatic and amphibious habitats. Oxford, UK:
Blackwell.
Dahlskog S. 1966. Sedimentation and vegetation in a Lapland mountain delta.
Geografiska Annaler. Series A, Physical Geography 48: 86–101.D�ecampsH, Planty-Tabacchi AM, Tabacchi E. 1995.Changes in the hydrological
regime and invasions by plant species along riparian systems of the Adour River,
France. Regulated Rivers – Research and Management 11: 23–33.D�ecamps H, Tabacchi E. 1994. Species richness in vegetation along river margins.
In: Giller PS, Hildrew AG, Rafaelli DG, eds. Aquatic ecology. Scale, pattern andprocess. Oxford, UK: Blackwell Scientific Publishers, 1–20.
Dehnen-Schmutz K. 2011.Determining non-invasiveness in ornamental plants to
build green lists. Journal of Applied Ecology 48: 1374–1380.Deiller A-F, Walter J-MN, Tr�emoli�eres M. 2001. Effects of flood interruption on
species richness, diversity and florsistic composition of woody regeneration in the
upper Rhine alluvial hardwood forest. Regulated Rivers – Research andManagement 17: 393–405.
Dynesius M, Jansson R, Johansson ME, Nilsson C. 2004. Intercontinental
similarities in riparian-plant diversity and sensitivity to river regulation.EcologicalApplications 14: 173–191.
van Eck WHJM, Lenssen JPM, van de Steeg HM, Blom CWPM, de Kroon H.
2006. Seasonal dependent effects of flooding on plant species survival and
zonation: a comparative study of 10 terrestrial grassland species. Hydrobiologia565: 59–69.
Elderd BD. 2003. The impact of changing flow regimes on riparian vegetation
and the riparian species Mimulus guttatus. Ecological Applications 13: 1610–1625.
Engstr€om J, Jansson R, Nilsson C, Weber C. 2011. Effects of river ice on riparian
vegetation. Freshwater Biology 56: 1095–1105.Eschtruth AK, Battles JJ. 2011. The importance of quantifying propagule pressure
to understand invasion: an examination of riparian forest invasibility. Ecology 92:1314–1322.
Evangelista PH, Kumar S, Stohlgren TJ, Jarnevich CS, Crall AW,Norman JB III,
Barnett DT. 2008.Modelling invasion for a habitat generalist and a specialist
plant species. Diversity and Distributions 14: 808–817.Flory SL, Clay K. 2009. Invasive plant removal method determines native plant
community responses. Journal of Applied Ecology 46: 434–442.Geigenberger P. 2003. Response of plant metabolism to too little oxygen. CurrentOpinion in Plant Biology 6: 247–256.
Goodson JM, Gurnell AM, Angold PG, Morrissey IP. 2003. Evidence for
hydrochory and the deposition of viable seeds within winter flow-deposited
sediments: the River Dove, Derbyshire, UK. River Research and Applications 19:317–334.
G€otzenberger L, de Bello F, Br�athen KA, Davison J, Dubuis A, Guisan A, Lep�s J,
Lindborg R, Moora M, P€artel M et al. 2012. Ecological assembly rules in plant
communities – approaches, patterns and prospects. Biological Reviews 87: 111–127.
Grace JB. 1988. The effects of nutrient additions on mixures of Typha latifolia L.and Typha domingensis Pers. along a water-depth gradient. Aquatic Botany 31:83–92.
Grant EHC, Lowe WH, Fagan WF. 2007. Living in the branches: population
dynamics and ecological processes in dendritic networks. Ecology Letters 10: 165–175.
Grime JP. 1977. Evidence for the existence of three primary strategies in plants and
its relevance to ecological and evolutionary theory. The American Naturalist 111:1169–1194.
Grubb PJ. 1977. The maintenance of species-richness in plant communities: the
importance of the regeneration niche. Biological Reviews 52: 107–145.Gurnell A, Thompson K, Goodson J, Moggridge H. 2008. Propagule deposition
along river margins: linking hydrology and ecology. Journal of Ecology 96: 553–565.
He JB, B€ogemannGM, van de SteegHG, Rijnders JGHM,Voesenek LACJ, Blom
CWPM. 1999. Survival tactics of Ranunculus species in river floodplains.
Oecologia 118: 1–8.Hodgkinson DJ, Thompson K. 1997. Plant dispersal: the role of man. Journal ofApplied Ecology 34: 1484–1496.
H€orandl E, Emadzade K. 2012. Evolutionary classification: a case study on the
diverse plant genus Ranunculus L. (Ranunculaceae). Perspectives in Plant Ecology,Evolution and Systematics 14: 310–324.
HornMH, Correa SB, Parolin P, Pollux BJA, Anderson JT, Lucas C,Widmann
P, Tjiu A, Galetti M, Goulding M. 2011. Seed dispersal by fishes in tropical
and temperate fresh waters: the growing evidence. Acta Oecologica 37: 561–577.
Howell J, BensonD. 2000. Predicting potential impacts of environmental flows on
weedy riparian vegetation of the Hawkesbury-Nepean River, south-eastern
Australia. Austral Ecology 25: 463–475.Hubble TCT, Docker BB, Rutherfurd ID. 2010. The role of riparian trees in
maintaining riverbank stability: a review of Australian experience and practice.
Ecological Engineering 36: 292–304.HulmePE,Bacher S,KenisM,Klotz S,K€uhn I,MinchinD,NentwigW,OleninS,
Panov V, Pergl J et al. 2008. Grasping at the routes of biological invasions: a
framework for integrating pathways into policy. Journal of Applied Ecology 45:403–414.
Hupp CR, Osterkamp WR. 1985. Bottomland vegetation distribution along
Passage Creek, Virginia, in relaton to fluvial landforms. Ecology 66: 670–681.
Ikeda H, Itoh K. 2001.Germination and water dispersal of seeds from a threatened
plant species Penthorum chinense. Ecological Research 16: 99–106.ISSG 2013. Global invasive species database (GISD) invasive species specialist group,IUCN (The World Conservation Union). [WWW document] URL http://www.
issg.org/database [accessed 6 November 2013].
Jackson MB, Drew MC. 1984. Effects of flooding on growth and metabolism of
herbaceous plants. In: Kozlowski TT, ed. Flooding and plant growth. New York,
NY, USA: Academic Press, 47–128.JacksonMB, IshizawaK, ItoO. 2009.Evolution andmechanisms of plant tolerance
to flooding stress. Annals of Botany 103: 137–142.Jansson R, Nilsson C, Ren€of€alt B. 2000. Fragmentation of riparian floras in rivers
with multiple dams. Ecology 81: 899–903.Jansson R, Zinko U,Merritt DM,Nilsson C. 2005.Hydrochory increases riparian
plant species richness: a comparison between a free-flowing and a regulated river.
Journal of Ecology 93: 1094–1103.JohanssonME,NilssonC,NilssonE.1996.Dorivers function as corridors for plant
dispersal? Journal of Vegetation Science 7: 593–598.Jungwirth M,Muhar S, Schmutz S. 2002. Re-establishing and assessing ecological
integrity in riverine landscapes. Freshwater Biology 47: 867–887.Kalliola R, Puhakka M. 1988. River dynamics and vegetation mosaicism – acase-study of the river Kamajohka, northernmost Finland. Journal of Biogeography15: 703–719.
Karrenberg S, EdwardsPJ,Kollmann J. 2002.The life history of Salicaceae living in
the active zone of floodplains. Freshwater Biology 47: 733–748.
New Phytologist (2014) 204: 19–36 � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Review Tansley reviewNewPhytologist34

Keeley JE. 1998. CAM photosynthesis in submerged aquatic plants. BotanicalReview 64: 121–175.
Kozlowski TT, ed. 1984. Flooding and plant growth. Orlando, FL, USA: Academic
Press.
Kozlowski TT. 1997. Responses of woody plants to flooding and salinity. TreePhysiology Monograph 1: 1–29.
Kubitzki K, Ziburski A. 1994. Seed dispersal in flood-plain forests of Amazonia.
Biotropica 26: 30–43.Kurzweil H. 1993. Seed morphology in Southern African Orchidoideae(Orchidaceae). Plant Systematics and Evolution 185: 229–247.
Kyle G, Leishman MR. 2009. Functional trait differences between extant
exotic, native and extinct native plants in the Hunter River, NSW: a
potential tool in riparian rehabilitation. River Research and Applications 25:892–903.
Laan P, Berrevoets MJ, Lythe S, Armstrong W, Blom CWPM. 1989. Root
morphology and aerenchyma formation as indicators of the flood-tolerance of
Rumex species. Journal of Ecology 77: 693–703.Lind L, Nilsson C, Polvi LE, Weber C. 2014. The role of ice dynamics in shaping
vegetation in flowing waters. Biological Reviews. doi: 10.1111/brv.12077.von der Lippe M, Kowarik I. 2012. Interactions between propagule pressure and
seed traits shape human-mediated seed dispersal along roads. Perspectives in PlantEcology, Evolution and Systematics 14: 123–130.
Lockwood JL, Cassey P, Blackburn TM. 2009. The more you introduce the more
you get: the role of colonization pressure and propagule pressure in invasion
ecology. Diversity and Distributions 15: 904–910.LowranceR,ToddRL,Fail J,HendricksonO,LeonardR,AsmussenL.1984.Riparian
forests as nutrient filters in agricultural watersheds.BioScience 34: 374–377.LytleDA, Poff NL. 2004.Adaptation to natural flow regimes.Trends in Ecology andEvolution 19: 94–100.
Madsen TV, Sand-Jensen K. 1991. Photosynthetic carbon assimilation in aquatic
macrophytes. Aquatic Botany 41: 5–40.Mahoney JM, Rood SB. 1998. Streamflow, requirements for cottonwood seedling
recruitment – an interactive model.Wetlands 18: 634–645.Melbourne BA, Cornell HV, Davies KF, Dugaw CJ, Elmendorf S, Freestone AL,
Hall RJ, Harrison S, Hastings A, Holland M et al. 2007. Invasion in a
heterogeneous world: resistance, coexistence or hostile takeover? Ecology Letters10: 77–94.
Merritt DM, Nilsson C, Jansson R. 2010a. Consequences of propagule dispersal
and river fragmentation for riparian plant community diversity and turnover.
Ecological Monographs 80: 609–626.Merritt DM, Scott ML, Poff NL, Auble GT, Lytle DA. 2010b. Theory, methods
and tools for determining environmental flows for riparian vegetation: riparian
vegetation-flow response guilds. Freshwater Biology 55: 206–225.MerrittDM,WohlEE. 2006.Plant dispersal along rivers fragmented bydams.RiverResearch and Applications 22: 1–26.
Middleton B. 2000.Hydrochory, seed banks, and regeneration dynamics along the
landscape boundaries of a forested wetland. Plant Ecology 146: 167–181.Moles AT, Ackerly DD,Webb CO, Tweddle JC, Dickie JB, Pitman AJ, Westoby
M. 2005. Factors that shape seed mass evolution. Proceedings of the NationalAcademy of Sciences, USA 102: 10540–10544.
Moles AT, Westoby M. 2004. Seedling survival and seed size: a synthesis of the
literature. Journal of Ecology 92: 372–383.Mommer L, Pons TL, Wolters-Arts M, Venema JH, Visser EJW. 2005.
Submergence-induced morphological, anatomical, and biochemical responses in
a terrestrial species affect gas diffusion resistance and photosynthetic performance.
Plant Physiology 139: 497–508.Mommer L, Visser EJW. 2005. Underwater photosynthesis in flooded terrestrial
plants: a matter of leaf plasticity. Annals of Botany 96: 581–589.Mouw JEB, Alaback PB. 2003. Putting floodplain hyperdiversity in a regional
context: an assessment of terrestrial-floodplain connectivity in a montane
environment. Journal of Biogeography 30: 87–103.Niggemann M, Jetzkowitz J, Brunzel S, Wichmann MC, Bialozyt R. 2009.
Distribution patterns of plants explained by human movement behaviour.
Ecological Modelling 220: 1339–1346.NilssonC,AnderssonE,MerrittDM, JohanssonME.2002.Differences in riparian
flora between riverbanks and river lakeshores explained by dispersal traits. Ecology83: 2878–2887.
Nilsson C, Brown RL, Jansson R, Merritt DM. 2010. The role of hydrochory
in structuring riparian and wetland vegetation. Biological Reviews 85:837–858.
Nilsson C, Ekblad A, Dynesius M, Backe S, Gardfjell M, Carlberg B, Hellqvist
S, Jansson R. 1994. Comparison of species richness and traits of riparian
plants between a main river channel and its tributaries. Journal of Ecology 82:281–295.
Nilsson C, Grelsson G, Johansson M, Sperens U. 1989. Patterns of plant species
richness along riverbanks. Ecology 70: 77–84.Pacala SW, TilmanD. 1994. Limiting similarity in mechanistic and spatial models
of plant competition in heterogeneous environments. The American Naturalist143: 222–257.
Palmer MA, Bernhardt ES, Allan JD, Lake PS, Alexander G, Brooks S, Carr J,
Clayton S, Dahm CN, Follstad Shah J et al. 2005. Standards for ecologicallysuccessful river restoration. Journal of Applied Ecology 42: 208–217.
Parolin P. 2006.Ombrohydrochory: rain-operated seed dispersal in plants – withspecial regard to jet-action dispersal in Aizoaceae. Flora 201: 511–518.
Pettit NE, Froend RH. 2001. Availability of seed for recruitment of riparian
vegetation: a comparisonof a tropical and a temperate river ecosystem inAustralia.
Australian Journal of Botany 49: 515–528.Pickering C, Mount A. 2010.Do tourists disperse weed seed? A global review of
unintentional human-mediated terrestrial seed dispersal on clothing, vehicles and
horses. Journal of Sustainable Tourism 18: 239–256.van der Pijl L. 1972. Principles of dispersal in higher plants. Berlin, Germany:
Springer.
Planty-Tabacchi AM, Tabacchi E, Naiman RJ, DeFerrari C, Decamps H. 1996.
Invasibility of species rich communities in riparian zones.ConservationBiology10:598–607.
Poff NL, Allan JD, Bain MB, Karr JR, Prestegaard KL, Richter BD, Sparks RE,
Stromberg JC. 1997. The natural flow regime. BioScience 47: 769–784.Pollock MM, Naiman RJ, Hanley TA. 1998. Plant species richness in riparian
wetlands – a test of biodiversity theory. Ecology 79: 94–105.Pollux BJA, Jong MDE, Steegh A, Verbruggen E, Van Groenendael JM, Ouborg
NJ. 2007. Reproductive strategy, clonal structure and genetic diversity in
populations of the aquatic macrophyte Sparganium emersum in river systems.
Molecular Ecology 16: 313–325.Pollux BJA, Luteijn A, Van Groenendael JM, Ouborg NJ. 2009. Gene flow and
genetic structure of the aquatic macrophyte Sparganium emersum in a linear
unidirectional river. Freshwater Biology 54: 64–76.Puckridge JT, Sheldon F, Walker KF, Boulton AJ. 1998. Flow variability and the
ecology of large rivers.Marine and Freshwater Research 49: 55–72.PuijalonS,BornetteG.2004.Morphological variationof two taxonomically distant
plant species along a natural flowvelocity gradient.NewPhytologist163: 651–660.ReaN, StorrsMJ. 1999.Weed invasions inwetlands ofAustralia’s TopEnd: reasons
and solutions.Wetlands Ecology and Management 7: 47–62.Ren€of€alt BM,Nilsson C, Jansson R. 2005. Spatial and temporal patterns of species
richness in a riparian landscape. Journal of Biogeography 32: 2025–2037.RichardsonDM,Holmes PM, Esler KJ, Galatowitsch SM, Stromberg JC, Kirkman
SP, Py�sek P, Hobbs RJ. 2007. Riparian vegetation: degradation, alien plant
invasions, and restoration prospects. Diversity and Distributions 13: 126–139.Richardson DM, Py�sek P, Rejm�anek M, Barbour MG, Panetta FD, West CJ.
2000. Naturalization and invasion of alien plants: concepts and definitions.
Diversity and Distributions 6: 93–107.Ridge I. 1987. Ethylene and growth control in amphibious plants. In: Crawford
RMM, ed. Plant life in aquatic and amphibious habitats. Oxford, UK: Blackwell,
53–76.Riis T, Sand-Jensen K. 2006. Dispersal of plant fragments in small streams.
Freshwater Biology 51: 274–286.Rivadavia F, Vicentini A, Fleischmann A. 2009. A new species of sundew (Drosera,Droseraceae), with water-dispersed seed, from the floodplains of the northern
Amazon basin, Brazil. Ecotropica 15: 13–21.Rood SB,GourleyCR,AmmonEM,Heki LG,Klotz JR,MorrisonML,MosleyD,
Scoppettone GG, Swanson S, Wagner PL. 2003. Flows for floodplain forests: a
successful riparian restoration. BioScience 53: 647–656.Rood SB,Mahoney JM. 1990.Collapse of riparian poplar forests downstream from
dams in western prairies: probable causes and prospects for mitigation.
Environmental Management 14: 451–464.
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014) 204: 19–36
www.newphytologist.com
NewPhytologist Tansley review Review 35

Rood SB, Taboulchanas K, Bradley CE, Kalischuk AR. 1999. Influence of flow
regulation on channel dynamics and riparian cottonwoods along the Bow River,
Alberta. Rivers 7: 33–48.Sabo JL, Sponseller R, Dixon M, Gade K, Harms T, Heffernan J, Jani A, Katz G,
SoykanC,Watts J et al.2005.Riparian zones increase regional species richness byharboring different, not more, species. Ecology 86: 56–62.
S€aumel I, Kowarik I. 2013. Propagule morphology and river characteristics shape
secondary water dispersal in tree species. Plant Ecology 214: 1257–1272.Schneider RL, Sharitz RR. 1988.Hydrochory and regeneration in a Bald
Cypress-Water Tupelo Swamp Forest. Ecology 69: 1055–1063.Sculthorpe CD. 1967. The biology of squatic vascular plants. London, UK: EdwardArnold.
Seago JL, Marsh LC, Stevens KJ, Soukup A, Votrubov�a O, Enstone DE. 2005. A
re-examination of the root cortex in wetland flowering plants with respect to
aerenchyma. Annals of Botany 96: 565–579.Silvertown J, Dodd ME, Gowing DJG, Mountford JO. 1999.Hydrologically
definedniches reveal a basis for species richness in plant communities.Nature400:61–63.
Simberloff D. 2009. The role of propagule pressure in biological invasions. AnnualReview of Ecology, Evolution, and Systematics 40: 81–102.
StohlgrenTJ,BinkleyD,ChongGW,KalkhanMA, Schell LD,BullKA,Otsuki Y,
Newman G, Bashkin M, Son Y. 1999. Exotic plant species invade hot spots of
native plant diversity. Ecological Monographs 69: 25–46.StohlgrenTJ,BullKA,OtsukiY,VillaCA,LeeM.1998.Riparian zones as havens for
exotic plant species in the central grasslands. Plant Ecology 138: 113–125.Stokes K, Ward K, Colloff M. 2010. Alterations in flood frequency increase exotic
and native species richness of understorey vegetation in a temperate floodplain
eucalypt forest. Plant Ecology 211: 219–233.Str€omL, JanssonR,NilssonC. 2012. Projected changes in plant species richness and
extent of riparian vegetation belts as a result of climate-driven hydrological change
along the Vindel River in Sweden. Freshwater Biology 57: 49–60.Stromberg JC, Lite SJ, Marler R, Paradzick C, Shafroth PB, Shorrock D, White
JM,WhiteMS. 2007.Altered stream-flow regimes and invasive plant species: the
Tamarix case. Global Ecology and Biogeography 16: 381–393.TownsendCR,HildrewAG. 1994. Species traits in relation to a habitat templet for
river systems. Freshwater Biology 31: 265–275.van der Valk AG. 1981. Succession in wetlands: a Gleasonian approach. Ecology 62:688–696.
Vartapetian BB, JacksonMB. 1997. Plant adaptations to anaerobic stress. Annals ofBotany 79: 3–20.
Vervuren PJA, Blom CWPM, de Kroon H. 2003. Extreme flooding events on the
Rhine and the survival and distribution of riparian plant species. Journal of Ecology91: 135–146.
Vil�aM,Pino J, FontX. 2007.Regional assessment of plant invasions across different
habitat types. Journal of Vegetation Science 18: 35–42.Visser EJW, Voesenek L, Vartapetian BB, Jackson MB. 2003. Flooding and plant
growth. Annals of Botany 91: 107–109.Voesenek LACJ, Colmer TD, Pierik R, Millenaar FF, Peeters AJM. 2006.How
plants cope with complete submergence. New Phytologist 170: 213–226.Voesenek LACJ, Rijnders JHGM, Peeters AJM, Van de SteegHMV,DeKroonH.
2004. Plant hormones regulate fast shoot elongation under water: from genes to
communities. Ecology 85: 16–27.Wallace JB, Eggert SL, Meyer JL, Webster JR. 1997.Multiple trophic levels of a
forest stream linked to terrestrial litter inputs. Science 277: 102–104.Washitani I, Takenaka A, Kuramoto N, Inoue K. 1997. Aster kantoensisKitam., an
endangered flood plain endemic plant in Japan: its ability to form persistent soil
seed banks. Biological Conservation 82: 67–72.
Weiher E, Clarke GDP, Keddy PA. 1998. Community assembly rules,
morphological dispersion, and the coexistence of plant species. Oikos 81: 309–322.
Weiher E, Keddy PA. 1995a. The assembly of experimental wetland plant
communities. Oikos 73: 323–335.Weiher E, Keddy PA. 1995b.Assembly rules, null models, and trait dispersion: new
questions from old patterns. Oikos 74: 159–164.Westoby M. 1998. A leaf-height-seed (LHS) plant ecology strategy scheme. Plantand Soil 199: 213–227.
Whitney KD, Gabler CA. 2008. Rapid evolution in introduced species, ‘invasive
traits’ and recipient communities: challenges for predicting invasive potential.
Diversity and Distributions 14: 569–580.Wilson JRU, Dormontt EE, Prentis PJ, Lowe AJ, Richardson DM. 2009.
Something in theway youmove: dispersal pathways affect invasion success.Trendsin Ecology & Evolution 24: 136–144.
Xiong S, Nilsson C, JohanssonME, Jansson R. 2001. Responses of riparian plants
to accumulation of silt and plant litter: the importance of plant traits. Journal ofVegetation Science 12: 481–490.
Supporting Information
Additional supporting information may be found in the onlineversion of this article.
Table S1Number of quadrats surveyed for each ecosystem type (orecological vegetation class) in Victoria between 1970 and 2011
Table S2 Our approach for testing four hypotheses proposed toexplain high degrees of riparian invasion based on characteristics ofinvading species
Table S3 Results of a one-way ANOVA comparing the number ofdispersal modes used by species that invade and do not invaderiparian zones
Table S4 Results of a one-way ANOVA comparing the prevalenceof dispersal using water currents among species that invade and donot invade riparian zones
Table S5 Results of a one-way ANOVA comparing the prevalenceof human-mediated dispersal among species that invade and do notinvade riparian zones
Notes S1Description of datasets and analyses regarding the degreeof invasion of riparian zones
Please note: Wiley Blackwell are not responsible for the content orfunctionality of any supporting information supplied by theauthors. Any queries (other than missing material) should bedirected to the New Phytologist Central Office.
New Phytologist (2014) 204: 19–36 � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Review Tansley reviewNewPhytologist36
Copyright © 2022 FDOKUMEN




















