
Dose-specific transcriptional responses in thyroid tissue in mice after 131I administration
6
Dose-specific transcriptional responses in thyroid tissue in mice after 131 I administration Nils Rudqvist a, ⁎, Emil Schüler a , Toshima Z. Parris b , Britta Langen a , Khalil Helou b , Eva Forssell-Aronsson a, c a Department of Radiation Physics, Institute of Clinical Sciences, Sahlgrenska Cancer Center, Sahlgrenska Academy, University of Gothenburg, Sahlgrenska University Hospital, SE-413 45 Gothenburg, Sweden b Department of Oncology, Institute of Clinical Sciences, Sahlgrenska Cancer Center, Sahlgrenska Academy, University of Gothenburg, Sahlgrenska University Hospital, SE-413 45 Gothenburg, Sweden c Department of Medical Physics and Medical Bioengineering, Sahlgrenska University Hospital, SE-413 45 Gothenburg, Sweden abstract article info Article history: Received 16 July 2014 Received in revised form 14 November 2014 Accepted 19 November 2014 Available online xxxx Keywords: Radiation biology Normal thyroid gland Radionuclide therapy 131 I Global transcriptional gene regulation Introduction: In the present investigation, microarray analysis was used to monitor transcriptional activity in thy- roids in mice 24 h after 131 I exposure. The aims of this study were to 1) assess the transcriptional patterns associated with 131 I exposure in normal mouse thyroid tissue and 2) propose biomarkers for 131 I exposure of the thyroid. Methods: Adult BALB/c nude mice were i.v. injected with 13, 130 or 260 kBq of 131 I and killed 24 h after injection (absorbed dose to thyroid: 0.85, 8.5, or 17 Gy). Mock-treated mice were used as controls. Total RNA was extracted from thyroids and processed using the Illumina platform. Results: In total, 497, 546, and 90 transcripts were regulated (fold change ≥1.5) in the thyroid after 0.85, 8.5, and 17 Gy, respectively. These were involved in several biological functions, e.g. oxygen access, inflammation and im- mune response, and apoptosis/anti-apoptosis. Approximately 50% of the involved transcripts at each absorbed dose level were dose-specific, and 18 transcripts were commonly detected at all absorbed dose levels. The Agpat9, Plau, Prf1, and S100a8 gene expression displayed a monotone decrease in regulation with absorbed dose, and further studies need to be performed to evaluate if they may be useful as dose-related biomarkers for 131I exposure. Conclusion: Distinct and substantial differences in gene expression and affected biological functions were detected at the different absorbed dose levels. The transcriptional profiles were specific for the different absorbed dose levels. We propose that the Agpat9, Plau, Prf1, and S100a8 genes might be novel potential absorbed dose-related bio- markers to 131 I exposure of thyroid. Advances in knowledge: During the recent years, genomic techniques have been developed; however, they have not been fully utilized in nuclear medicine and radiation biology. We have used RNA microarrays to investigate genome- wide transcriptional regulations in thyroid tissue in mice after low, intermediate, and high absorbed doses from 131 I exposure in vivo. Using this approach, we have identified novel biological responses and potential absorbed dose- related biomarkers to 131 I exposure. Our research shows the importance of embracing technological advances and multi-disciplinary collaboration in order to apply them in radiation therapy, nuclear medicine, and radiation biology. Implications on Patient Care: This work may contribute with new knowledge of potential normal tissue effects or complications that may occur after exposure to ionizing radiation in diagnostic and therapeutic nuclear medicine, and due to radioactive fallout or accident with radionuclide spread. © 2014 Elsevier Inc. All rights reserved. 1. Background Iodide is taken up by the thyroid gland and incorporated into thyroid hormones, crucial for fetal and neonatal central nervous system devel- opment and in the regulation of metabolic processes [1]. While 127 I is the only stable iodine isotope, several radioactive isotopes exist, and al- ready in 1941 131 I was reported to be suitable for treatment of Graves' disease [2]. Currently, 131 I is routinely used for treatment of thyroid can- cer and hyperthyroidism (as iodide), and for treatment of other tumour types (when bound to tumour-seeking agents). Cellular responses to irradiation are induced by e.g. a direct hit of the DNA molecule or other targets within a cell, radiation-induced free rad- icals, or as indirect (bystander) response by extracellular signaling [3–6]. These responses affect gene expression levels and cause tran- scriptional regulation. Transcriptional profiling with microarrays enables detection of global mRNA variation in tissues in a single exper- iment. The majority of studies using genome-wide approaches have been performed in vitro using gamma radiation and often at few absorbed dose levels or temporal points [7,8]. In contrast, studies per- formed in vivo have demonstrated complex and distinct dose- dependent cellular responses following irradiation. Transcriptional profiles of mouse kidneys and brain demonstrated that both tissues respond in a tissue-specific manner [9]. We recently demonstrated Nuclear Medicine and Biology xxx (2014) xxx–xxx ⁎ Corresponding author. Tel.: +46 31 3421000; fax: +46 31 3421378. E-mail address: [email protected] (N. Rudqvist). http://dx.doi.org/10.1016/j.nucmedbio.2014.11.006 0969-8051/© 2014 Elsevier Inc. All rights reserved. Contents lists available at ScienceDirect Nuclear Medicine and Biology journal homepage: www.elsevier.com/locate/nucmedbio Please cite this article as: Rudqvist N., et al, Dose-specific transcriptional responses in thyroid tissue in mice after 131 I administration, Nucl Med Biol (2014), http://dx.doi.org/10.1016/j.nucmedbio.2014.11.006
Transcript of Dose-specific transcriptional responses in thyroid tissue in mice after 131I administration

Nuclear Medicine and Biology xxx (2014) xxx–xxx
Contents lists available at ScienceDirect
Nuclear Medicine and Biology
j ourna l homepage: www.e lsev ie r .com/ locate /nucmedb io
Dose-specific transcriptional responses in thyroid tissue in miceafter 131I administration
Nils Rudqvist a,⁎, Emil Schüler a, Toshima Z. Parris b, Britta Langen a, Khalil Helou b, Eva Forssell-Aronsson a,c
a Department of Radiation Physics, Institute of Clinical Sciences, Sahlgrenska Cancer Center, Sahlgrenska Academy, University of Gothenburg,Sahlgrenska University Hospital, SE-413 45 Gothenburg, Swedenb Department of Oncology, Institute of Clinical Sciences, Sahlgrenska Cancer Center, Sahlgrenska Academy, University of Gothenburg,Sahlgrenska University Hospital, SE-413 45 Gothenburg, Swedenc Department of Medical Physics and Medical Bioengineering, Sahlgrenska University Hospital, SE-413 45 Gothenburg, Sweden
a b s t r a c ta r t i c l e i n f o
⁎ Corresponding author. Tel.: +46 31 3421000; fax: +E-mail address: [email protected] (N. Rudqvis
http://dx.doi.org/10.1016/j.nucmedbio.2014.11.0060969-8051/© 2014 Elsevier Inc. All rights reserved.
Please cite this article as: Rudqvist N., et al, DBiol (2014), http://dx.doi.org/10.1016/j.nucm
Article history:
Received 16 July 2014Received in revised form 14 November 2014Accepted 19 November 2014Available online xxxxKeywords:Radiation biologyNormal thyroid glandRadionuclide therapy131IGlobal transcriptional gene regulation
Introduction: In the present investigation, microarray analysis was used to monitor transcriptional activity in thy-roids inmice 24 h after 131I exposure. The aims of this studywere to 1) assess the transcriptional patterns associatedwith 131I exposure in normal mouse thyroid tissue and 2) propose biomarkers for 131I exposure of the thyroid.Methods: Adult BALB/c nude mice were i.v. injected with 13, 130 or 260 kBq of 131I and killed 24 h after injection(absorbed dose to thyroid: 0.85, 8.5, or 17 Gy). Mock-treated mice were used as controls. Total RNA was extractedfrom thyroids and processed using the Illumina platform.Results: In total, 497, 546, and 90 transcripts were regulated (fold change ≥1.5) in the thyroid after 0.85, 8.5, and17 Gy, respectively. These were involved in several biological functions, e.g. oxygen access, inflammation and im-mune response, and apoptosis/anti-apoptosis. Approximately 50% of the involved transcripts at each absorbeddose level were dose-specific, and 18 transcripts were commonly detected at all absorbed dose levels. The Agpat9,Plau, Prf1, and S100a8 gene expression displayed amonotone decrease in regulationwith absorbed dose, and further
studies need to be performed to evaluate if they may be useful as dose-related biomarkers for 131I exposure.Conclusion:Distinct and substantial differences in gene expression and affected biological functionswere detected atthe different absorbed dose levels. The transcriptional profiles were specific for the different absorbed dose levels.We propose that the Agpat9, Plau, Prf1, and S100a8 genes might be novel potential absorbed dose-related bio-markers to 131I exposure of thyroid.Advances in knowledge:During the recent years, genomic techniques have been developed; however, they have notbeen fully utilized in nuclearmedicine and radiationbiology.Wehave usedRNAmicroarrays to investigate genome-wide transcriptional regulations in thyroid tissue inmice after low, intermediate, and high absorbed doses from 131Iexposure in vivo. Using this approach, we have identified novel biological responses and potential absorbed dose-related biomarkers to 131I exposure. Our research shows the importance of embracing technological advances andmulti-disciplinary collaboration in order to apply them in radiation therapy, nuclearmedicine, and radiation biology.Implications on Patient Care: This work may contribute with new knowledge of potential normal tissue effects orcomplications that may occur after exposure to ionizing radiation in diagnostic and therapeutic nuclear medicine,and due to radioactive fallout or accident with radionuclide spread.© 2014 Elsevier Inc. All rights reserved.
1. Background
Iodide is taken up by the thyroid gland and incorporated into thyroidhormones, crucial for fetal and neonatal central nervous system devel-opment and in the regulation of metabolic processes [1]. While 127I isthe only stable iodine isotope, several radioactive isotopes exist, and al-ready in 1941 131I was reported to be suitable for treatment of Graves'disease [2]. Currently, 131I is routinely used for treatment of thyroid can-cer and hyperthyroidism (as iodide), and for treatment of other tumourtypes (when bound to tumour-seeking agents).
46 31 3421378.t).
ose-specific transcriptional redbio.2014.11.006
Cellular responses to irradiation are induced by e.g. a direct hit of theDNAmolecule or other targets within a cell, radiation-induced free rad-icals, or as indirect (bystander) response by extracellular signaling[3–6]. These responses affect gene expression levels and cause tran-scriptional regulation. Transcriptional profiling with microarraysenables detection of global mRNA variation in tissues in a single exper-iment. The majority of studies using genome-wide approaches havebeen performed in vitro using gamma radiation and often at fewabsorbed dose levels or temporal points [7,8]. In contrast, studies per-formed in vivo have demonstrated complex and distinct dose-dependent cellular responses following irradiation. Transcriptionalprofiles of mouse kidneys and brain demonstrated that both tissuesrespond in a tissue-specific manner [9]. We recently demonstrated
esponses in thyroid tissue in mice after 131I administration, Nucl Med

2 N. Rudqvist et al. / Nuclear Medicine and Biology xxx (2014) xxx–xxx
that 0.1–9.7 mGy from 131I, 0.24–1000 mGy from 211At, and variousabsorbed dose levels of 177Lu exposure induced distinctly different re-sponses in normal tissues in various mice tissues [10–13]. Furthermore,thyroid tissue inmice exposed to 0.05–32Gy 211At showed a substantialdifference in the gene expression profiles between the absorbed doselevels [14]. In the same study, 130 genes were regulated at all absorbeddose levels and may thus potential biomarkers for 211At exposure.
The aims of the current study were to 1) assess the transcriptionalpatterns associated with 131I exposure of thyroid tissue in adult miceand 2) to propose novel biomarkers for 131I exposure of the thyroid. Inthe present study, we identified distinct differences in global transcrip-tional levels and affected biological processes between the absorbeddose levels.
2. Methods
2.1. Radionuclide and radioactivity measurements
131I was obtained as NaI from GE Healthcare (Braunschweig,Germany). A gamma counter (Wallac 1480 Wizard® 3"; Wallac Oy,Turku, Finland) was used to determine injected activity by measure-ment of aliquots of the stem solutions.
2.2. Study design
Eight BALB/c nude mice aged 6–8 months were divided into fourgroups (n = 2). Animals from 3 groups were injected in the tail veinwith 0.15 ml of 13, 130 or 260 kBq 131I diluted in phosphate-buffered sa-line (pH 7). The fourth group was mock-treated in the tail vein withempty syringes. Mice had access to water and standard mouse food adlibitum. Animals were killed by cardiac puncture under anesthesia withpentobarbitalnatrium (APL; Kungens Kurva, Sweden) 24 h after injection,and thyroidswere excised and immediately snap-frozen in liquid nitrogenand stored at−80 °C until analysis. The study designwas approved by theEthical Committee on Animal Experiments in Gothenburg, Sweden.
2.3. Dosimetric calculations
Themedical internal radiation dose (MIRD) [15] formalismwas usedto calculate the thyroid mean absorbed dose,
Dthyroid ¼ Ae thyroid�X
niEi � фi
mthyroid
:
Ãthyroid is then the cumulated activity in the thyroid, ni is the radia-tion yield, Ei is the energy, фi is the absorbed fraction for radiation i inthe thyroid, and mthyroid is the thyroid mass. Previously published dataon activity concentration in thyroids were used to determine Ãthyroid
[16]. The data consist of radioactivity measurements at t = 0, 1, 2, 4, 6,and 24 h on thyroid in mice, and the trapezoidal rule was used to deter-mine Ãthyroid in the thyroid gland. Only electrons emitted by 131I locatedin the thyroid gland were considered, and the фi was set to 0.742 [17].The absorbed dose to thyroid was 0.85, 8.5, and 17 Gy after injectionsof 13, 130 and 260 kBq 131I, respectively.
2.4. Transcriptional analysis
Total RNAwas extracted frompooled thyroids from each group usingthe RNeasy Lipid Tissue Mini Kit (Qiagen; Hilden, Germany) and proc-essed at the Swegene Center for Integrative Biology at Lund University(SCIBLU) using the MouseRef-8 Whole-Genome Expression BeadChips(Illumina; San Diego, CA, USA). Raw signal intensities and images wereacquired using the Illumina BeadScan 3.5.31.17122 (Illumina; SanDiego, CA) image analysis software and Illumina BeadArray Reader scan-ner, respectively. BioArray Software Environment system (BASE) were
Please cite this article as: Rudqvist N., et al, Dose-specific transcriptional rBiol (2014), http://dx.doi.org/10.1016/j.nucmedbio.2014.11.006
used to preprocess data and for quantile normalization of the raw signalintensities [18]. Further data processing was performed with Nexus Ex-pression 2.0 (BioDiscovery; El Segundo, CA) to identify regulated tran-scripts (fold change N1.5) with an FDR-corrected p-value b0.01 usingthe Benjamini-Hochberg method [19]. Raw data have been depositedat NCBI Gene Expression Omnibus (GEO no. GSE54594).
Results from statistical analyses on regulated genes are presented inthe Supplementary Table 1. All genes in the text are considered statisticallysignificant, but p-values are not included in the text to enhance readability.
2.5. Gene ontology analysis
A categorization of detected gene ontology (GO) termsbased on ances-tor charts that link transcripts to biological function was established(http://www.geneontology.org) [20]. Eight main categories (withcorresponding subcategories) were used: 1) DNA integrity; 2) geneexpression integrity; 3) cell communication; 4) organismic regulation; 5)cell cycle and differentiation; 6) metabolic processes; 7) cellular integrity;8) stress responses.
Statistical analyses results of enriched gene ontology terms arefound in Supplementary Table 3. All terms in the text are considered sta-tistically significant with a p-value b0.05, but p-values are not includedin the text to enhance readability.
3. Results
3.1. Regulated transcripts
The number of statistically significant transcripts regulated in the thy-roid at 24 h after exposure to 0.85, 8.5, and 17 Gy from i.v. injected 131Iwas 497, 546, and 90, respectively (Fig. 1A, Supplementary Table 1).Results from statistical analysis (p-values) are presented in Supplementa-ry Table 1. The fraction of up-regulated transcripts increased withabsorbed dose, particularly between 8.5 and 17 Gy. Approximately 50%of the regulated transcripts detected at each absorbed dose level weredose-specific (Fig. 1B, Table 1). Among the 852 regulated transcripts(758 genes), 18 genes were commonly detected at 0.85, 8.5, and 17 Gy(Table 2). Of these, down-regulation (12 genes) was more prevalentthan up-regulation (4 genes), and two genes (Clec2d and Ccl9) weredown-regulated at 0.85 and 8.5 Gy, but up-regulated at 17 Gy.
3.2. Gene ontology analysis
Regulated transcripts were classified according to enriched GOterms and further categorized (Fig. 2, Supplementary Tables 2 and 3).Results of statistical analysis (p-values) are given in SupplementaryTable 3. The number of affected biological processes was 41 for 17 Gy,followed by 36 and 31 processes for 0.85 and 8.5 Gy, respectively.Only one GO term (cell adhesion) was affected at all exposure levels.
GO terms related to DNA and gene expression integrity,e.g., maintenance of telomeres and transcription, were only detectedat 0.85 Gy. Various growth factor receptor signaling pathways were af-fected at 0.85 and 8.5 Gy. The SMAD pathway was affected by down-regulation of Spnb2 at 0.85 Gy and up-regulation of Tob1 at both 0.85and 17 Gy. Furthermore, the TGF-β receptor signaling pathway was af-fected at 8.5 Gy, via up-regulation of DOH48114 and down-regulation ofLtbp4. Exposure to all absorbed dose levels resulted in similar numbersof detected GO terms for the category cell cycle and differentiation (sub-categories: apoptotic cell death, cell cycle regulation, and differentiationand aging).While 0.85 and 8.5 Gy had an impact on negative regulationof apoptosis, cell cycle regulation was affected at all absorbed doselevels. Additionally, GO terms related to differentiation and aging weredetected at all exposure levels with regulation of cell growth at 0.85and 8.5 Gy and negative regulation of cell differentiation at 17 Gy. Var-ious GO terms related to metabolic processes were detected at allabsorbed doses. More GO terms related to cellular integrity were
esponses in thyroid tissue in mice after 131I administration, Nucl Med
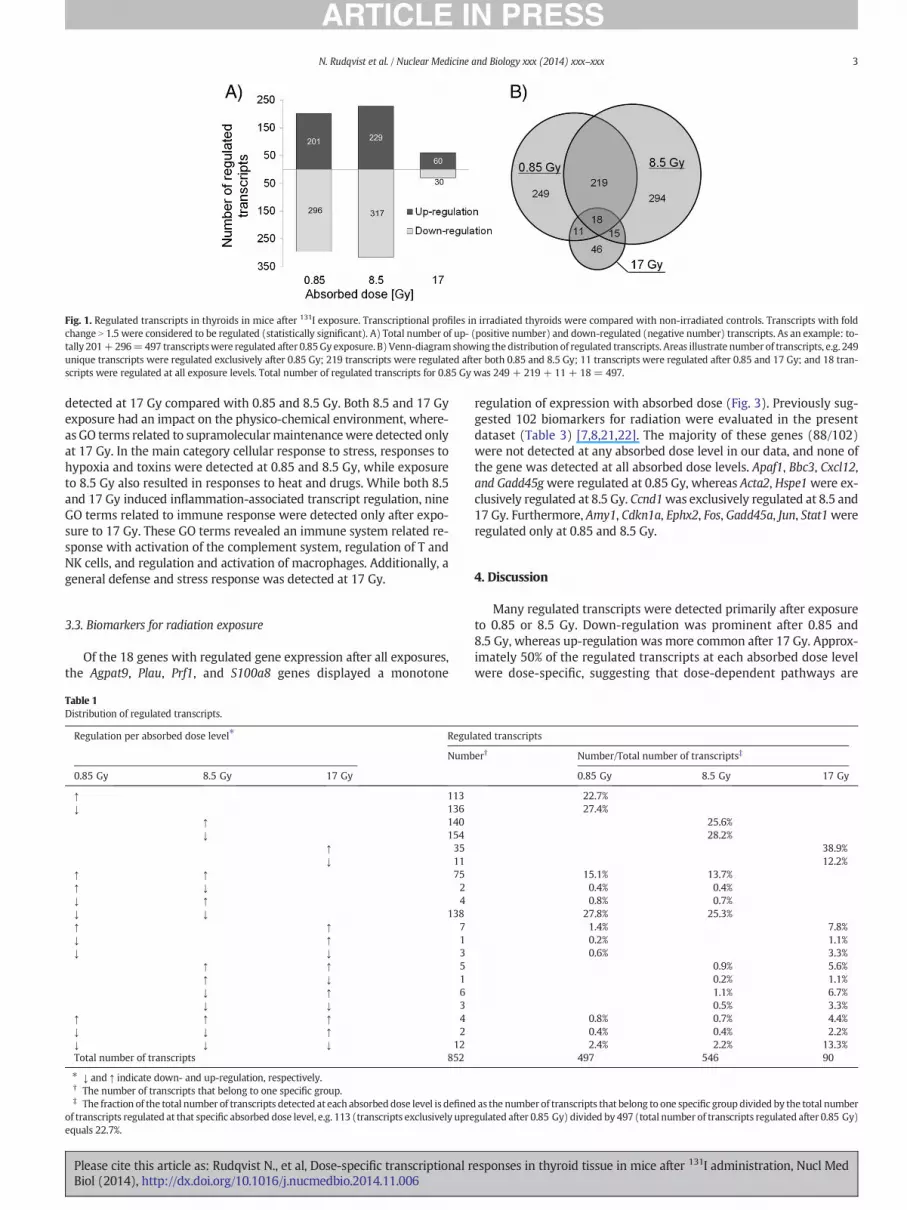
Fig. 1. Regulated transcripts in thyroids in mice after 131I exposure. Transcriptional profiles in irradiated thyroids were compared with non-irradiated controls. Transcripts with foldchange N 1.5 were considered to be regulated (statistically significant). A) Total number of up- (positive number) and down-regulated (negative number) transcripts. As an example: to-tally 201+296=497 transcriptswere regulated after 0.85Gy exposure. B) Venn-diagram showing the distribution of regulated transcripts. Areas illustrate number of transcripts, e.g. 249unique transcripts were regulated exclusively after 0.85 Gy; 219 transcripts were regulated after both 0.85 and 8.5 Gy; 11 transcripts were regulated after 0.85 and 17 Gy; and 18 tran-scripts were regulated at all exposure levels. Total number of regulated transcripts for 0.85 Gy was 249 + 219 + 11 + 18= 497.
3N. Rudqvist et al. / Nuclear Medicine and Biology xxx (2014) xxx–xxx
detected at 17 Gy compared with 0.85 and 8.5 Gy. Both 8.5 and 17 Gyexposure had an impact on the physico-chemical environment, where-as GO terms related to supramolecularmaintenancewere detected onlyat 17 Gy. In the main category cellular response to stress, responses tohypoxia and toxins were detected at 0.85 and 8.5 Gy, while exposureto 8.5 Gy also resulted in responses to heat and drugs. While both 8.5and 17 Gy induced inflammation-associated transcript regulation, nineGO terms related to immune response were detected only after expo-sure to 17 Gy. These GO terms revealed an immune system related re-sponse with activation of the complement system, regulation of T andNK cells, and regulation and activation of macrophages. Additionally, ageneral defense and stress response was detected at 17 Gy.
3.3. Biomarkers for radiation exposure
Of the 18 genes with regulated gene expression after all exposures,the Agpat9, Plau, Prf1, and S100a8 genes displayed a monotone
Table 1Distribution of regulated transcripts.
Regulation per absorbed dose level⁎ Regul
Numb
0.85 Gy 8.5 Gy 17 Gy
↑ 113↓ 136
↑ 140↓ 154
↑ 35↓ 11
↑ ↑ 75↑ ↓ 2↓ ↑ 4↓ ↓ 138↑ ↑ 7↓ ↑ 1↓ ↓ 3
↑ ↑ 5↑ ↓ 1↓ ↑ 6↓ ↓ 3
↑ ↑ ↑ 4↓ ↓ ↑ 2↓ ↓ ↓ 12Total number of transcripts 852
⁎ ↓ and ↑ indicate down- and up-regulation, respectively.† The number of transcripts that belong to one specific group.‡ The fraction of the total number of transcripts detected at each absorbeddose level is defined
of transcripts regulated at that specific absorbed dose level, e.g. 113 (transcripts exclusively upreequals 22.7%.
Please cite this article as: Rudqvist N., et al, Dose-specific transcriptional rBiol (2014), http://dx.doi.org/10.1016/j.nucmedbio.2014.11.006
regulation of expression with absorbed dose (Fig. 3). Previously sug-gested 102 biomarkers for radiation were evaluated in the presentdataset (Table 3) [7,8,21,22]. The majority of these genes (88/102)were not detected at any absorbed dose level in our data, and none ofthe gene was detected at all absorbed dose levels. Apaf1, Bbc3, Cxcl12,and Gadd45gwere regulated at 0.85 Gy, whereas Acta2, Hspe1were ex-clusively regulated at 8.5 Gy. Ccnd1was exclusively regulated at 8.5 and17 Gy. Furthermore, Amy1, Cdkn1a, Ephx2, Fos, Gadd45a, Jun, Stat1wereregulated only at 0.85 and 8.5 Gy.
4. Discussion
Many regulated transcripts were detected primarily after exposureto 0.85 or 8.5 Gy. Down-regulation was prominent after 0.85 and8.5 Gy, whereas up-regulation was more common after 17 Gy. Approx-imately 50% of the regulated transcripts at each absorbed dose levelwere dose-specific, suggesting that dose-dependent pathways are
ated transcripts
er† Number/Total number of transcripts‡
0.85 Gy 8.5 Gy 17 Gy
22.7%27.4%
25.6%28.2%
38.9%12.2%
15.1% 13.7%0.4% 0.4%0.8% 0.7%
27.8% 25.3%1.4% 7.8%0.2% 1.1%0.6% 3.3%
0.9% 5.6%0.2% 1.1%1.1% 6.7%0.5% 3.3%
0.8% 0.7% 4.4%0.4% 0.4% 2.2%2.4% 2.2% 13.3%
497 546 90
as the number of transcripts that belong to one specific group divided by the total numbergulated after 0.85 Gy) divided by 497 (total number of transcripts regulated after 0.85 Gy)
esponses in thyroid tissue in mice after 131I administration, Nucl Med

Table 2Genes differentially regulated at all absorbed dose levels (0.85, 8.5 and 17 Gy).
Fig. 3.Absorbed dose-related regulation of genes. Agpat9, Plau, Prf1, S100a8were differential-ly regulated at 0.85, 8.5, and 17 Gy with a monotone decrease in differential regulation withabsorbed dose. Agpat9, Plau, and Prf1were down-regulated while S100a8was up-regulated.
4 N. Rudqvist et al. / Nuclear Medicine and Biology xxx (2014) xxx–xxx
activated to manage different levels of radiation exposures. This alsomeans that approximately 50% of the detected transcripts at eachabsorbed dose level were found atmore than one dose level, suggestingsimilarities in the transcriptional response between absorbed doselevels even though only 18 unique transcripts were regulated at allabsorbed dose levels. It is not likely that themeasured response is an ef-fect from the chemical properties of iodine, because an injected amountof 13–260 kBq equals only 13–260×109 iodine atoms, whilemice have adietary intake in themagnitude of 1018 iodine atoms per day from stan-dard laboratory chow.
To our knowledge, no previous studies have been performed wherethe transcriptional response is measured in normal thyroid tissuein vivo after 131I exposure. However, comparison of the transcriptionalresponse between normal and cancer thyroid tissue have been per-formed in a Chernobyl cohort, but without using un-irradiated controls[22]. In a previous study where we investigated the thyroid responseafter 211At administration in a similar animal model, the fraction ofdose-specific transcripts decreasedwith increased absorbed dose, dem-onstrating one of the differences in response between 131I and 211At[14]. Additionally, of the 18 genes commonly regulated after 131I expo-sure, only Clec2d, Akam, Ccdc3,Dbp, and S100a8were detected at four ormore (out of five) absorbed dose levels in the previous 211At study. One
Fig. 2. Affected biological processes. Transcriptional changes associated with changes inbiological functions using gene ontology terms (statistically significant, p b 0.05). Theseterms were categorized using eight main categories (shown in the graph) and 22 subcat-egories (Supplementary Table 2).
Please cite this article as: Rudqvist N., et al, Dose-specific transcriptional rBiol (2014), http://dx.doi.org/10.1016/j.nucmedbio.2014.11.006
reason for differences in the transcriptional response between 131Iand 211At is the difference in radiation quality of themajor contributingparticles with high-LET alpha particles emitted by 211At and low-LETelectrons emitted by 131I. Another difference is themore heterogeneousenergy deposition delivered from 211At compared with 131I. Additional-ly, 211Atwill deposit the sameabsorbed dose butwith a higher dose-ratedue to shorter physical half-life compared with 131I.
Several genes were detected at 0.85 Gy and play a role in the regula-tion of DNA and gene expression integrity, aswell as in response to hyp-oxia, toxin, drug, and heat. On the other hand, an acute response relatedto the immune system (17 Gy: 13 genes) and inflammation (8.5 Gy: 8genes; 17 Gy: 3 genes) were observed at 0.85 and 17 Gy. Studies thatmainly used high absorbed doses from external radiation sources haveshown that ionizing radiation causes inflammation and activation ofthe immune system [23]. It is not clear how and if the inflammatoryand immune system machinery is affected at low absorbed dosesor how properties of the radiation source matters, e.g. if thereare differences in response between external radiation exposure(e.g., radiation therapy and diagnostic radiology) and internal exposure(e.g., radionuclides used for diagnostics or therapy in nuclearmedicine).However, studies have shown that the inflammatory and immune re-sponse to irradiation is tissue specific, and since radionuclides usuallyare heterogeneously distributed in an organism, large differences in in-flammatory and immune response can be anticipated [24]. In the pres-ent study, no inflammation-associated GO terms were detected after0.85 Gy, and no immune system related responses were detected after0.85 and 8.5 Gy. This suggests that high absorbed doses are more likelyto affect inflammatory and immunological processes on the transcrip-tional level or that these processes may function without de novoprotein synthesis. Interestingly, we have previously shown that few in-flammatory and immunological responses are activated in mouse thy-roids in vivo following 211At exposure (0.05–32 Gy) [14]. Thesefindings indicate that mouse thyroid tissue responds differently to 131Iand 211At, maybe partly due to differences in radiation quality anddose distribution.
In the present investigation, the Edn1, Angptl4, Casp1 and Mb genesinvolved in hypoxia were regulated after 0.85 and 8.5 Gy. The Edn1encoded protein is a potent vasoconstrictor while theMb gene encodesfor the myoglobin protein, an O2 storage molecule and intracellular O2
transporter with observed expression in striated muscular tissue [25].The mRNA levels of Edn1 and Mb were down-regulated after exposureto 0.85Gy. If themRNA levels reflect protein levels, these results suggesta reduction in vasoconstriction and less available O2 transporters andstorage molecules in the analyzed tissue. Knock-out mice lacking theMb gene responded to a hypoxic condition by widening blood vesselsin heart tissue [25]. Further studies should test the hypothesis that at
esponses in thyroid tissue in mice after 131I administration, Nucl Med

Table 3Validation of 102 previously proposed radiation-related biomarkers.
Absorbed dose Gene
0.85 Gy Apaf1, Bbc3, Cxcl12, Gadd45g8.5 Gy Acta2, Hspe10.85 and 8.5 Gy Amy1, Cdkn1a, Ephx2, Fos, Gadd45a, Jun8.5 and 17 Gy Ccnd10.85, 8.5 and 17 Gy -Genes not detectedin the present data
Amy2, Apaf1, Atf3, Bag-1, Batf3, Bax, Bbc3,Btg2, Ccl26, Ccna2, Ccnb1, Ccnb2, Ccnd1,Ccne1, Ccng, Ccng1, Cdc20, Cdc25, Cdh5,Cdkn1a, Csf1, Cx3cl1, Cxcl10, Cxcl11,Cxcl12, Cxcl5, Cxcl9, Dd2, Ddb2, Ephx2,Fas, Fcgr1a, Fdxr, Fen1, Fhl2, Flt3, Fos,Gadd45a, Gadd45g, Gcgr1a, Gja1, Gjb2,Hiap, Hrad23b, Hspe1, Iap3, Icsbp1, Ifna2,Il1b, IL-8, Jun (Junb), Lep, Lif, Mak10,Makp8pi, Mapk1, Mapk3, Mdk, Mdm2,Myc, Noxa, Ogg1, Osm, Pcna, Pcsk1,Plcg2, Ppmid, Prkch, Rad51l1, Relb,Rhoa, Serpine1,Tbfsf4, Tbfrsf10b,Tgfb3, Tgfbr2, Tnf, Tnfaip8, Tnfrsf10b,Tnfrsf19, Tnfrsf21, Tnfsf4, Tp53, Tp53i3,Trp53inp1, Trp53inp2, Ttf, Ube2s, Wee,Vegfb, Xab2, X-iap, Xpc, Xrcc1, Xrcc6
5N. Rudqvist et al. / Nuclear Medicine and Biology xxx (2014) xxx–xxx
0.85 Gy, blood vessels are widened as a counter-measure to lower oxy-gen levels. TheMb genewas up-regulated after exposure to 8.5 Gy, indi-cating a high amount of oxygen in the analyzed tissue. This is supportedby down-regulation of the Angptl4 gene expression which is inducedduring hypoxia [26]. Furthermore, no response to hypoxia was seenafter 17 Gy. Additionally, the Angptl4 gene expression was recurrentlyregulated in liver, lungs, kidney cortex and medulla, and spleen 24 hafter injection of 131I or 211At into mice (0.1–1000 mGy) [10,11].
Effects on cell communication and signal transduction through reg-ulation of several growth factor receptor signaling pathways were seenafter 0.85 and 8.5 Gy. The SMAD pathway was affected by regulation ofthe Spnb2 and the Tob1 genes at 0.85 Gy and at 0.85 and 17 Gy, respec-tively. Furthermore, TGF-β receptor signaling was affected at 8.5 Gy. In-volvement of signal transduction via the SMAD- and the TGF-βpathways indicates that several important cell cycle control mecha-nisms are affected at different absorbed dose levels after 131I exposure.Negative regulation of apoptosis was seen after 0.85 and 8.5 Gy, andanti-apoptotic processes were affected at 0.85 Gy but not detectedafter 17 Gy. Together, this suggests a change in cellular fate, with regardto apoptosis, when the absorbed dose increases.
One of the major functions of the thyroid is to produce thyroid hor-mones containing iodine. The SLC5A5 gene encodes for the sodium-iodide symporter (NIS) and is partly responsible for transport of iodineand, to some extent, astatine into the thyroid follicle cells [27]. Previousstudies have demonstrated NISmRNA down-regulation in cultured thy-roid epithelial cells one and five days after 0.5 Gy exposure to 131I and211At, respectively [28]. Additionally, in the same system, iodide trans-port was immediately elevated and then reduced at 2–7 days after0.5 Gy of exposure to after 99mTc, 125I, 131I, and 211At with most signifi-cant changes after 211At exposure [28]. No deregulation of Slc5a5 wasseen in the present investigation or after 211At exposure in a previousstudy; however, the Slc5a8 gene which shares 70% sequence homologywith SLC5A5 in humanswas down-regulated at 1.4 Gy but up-regulatedafter 211At exposure of 0.05, 0.5, 11, and 32Gy [14]. In the present inves-tigation, regulation of previously published thyroid hormone synthesisassociated genes were not detected (Tshr, Slc5a5/NIS, Slc26a4/Pendrin,Tg,Duox2, Tpo, Slc16a2/Mct8, Clc5, Slc5a8/Smct1, andCftr) [29]. The iden-tified transcriptional response in the thyroid is generated from a combi-nation of different cell types. However, it is estimated that 70–80%of thecells in thyroid tissue are thyrocytes [30]. Together, these results
Please cite this article as: Rudqvist N., et al, Dose-specific transcriptional rBiol (2014), http://dx.doi.org/10.1016/j.nucmedbio.2014.11.006
indicate that the thyroid function is less affected at the transcriptionallevel 24 hours after administration of 131I than 211At.
Few of the 102 genes previously proposed as radiation exposure bio-markers found in the literaturewere detected in the present investigation[7,8,21,22]. None of the 102 genes was regulated at all absorbed doses,and 88 genes were not regulated at any absorbed dose level. Among thepreviously proposed biomarkers, the Acta2, Bbc3, Cdkn1a, and Gadd45agenes were also regulated in the present study, however not at allabsorbed doses levels [21]. The 102 genes were derived primarily fromin vitro and/or ex vivo studies, which may explain the discordance be-tween the previous studies and the present study in addition to the differ-ences in radiation quality. In the present study, the transcription factorStat3 was down-regulated at 0.85 and 8.5 Gy. This gene is involved inthe regulation of cell proliferation and cell death and was down-regulated in thyroid tumor patients exposed to 0.3–1 Gy from 131I re-leased in conjunction with the Chernobyl incident [22].
The Agpat9, Plau, Prf1, and S100a8 genes were monotonously regu-lated in the present investigation, i.e. increasing or decreasing gene ex-pression with absorbed dose throughout the entire absorbed doserange. The Prf1 gene, involved in T-cell mediated cytolysis, belongs tothe autoimmune thyroid disease signaling pathway, and the S100a8gene product may induce apoptosis under inflammatory condi-tions [31,32]. These genes are interesting biomarker candidate genesthatmight be useful to predict absorbed dose to thyroid inmice injectedwith 131I, at least in the interval 0.85–17 Gy. Their usefulness to predictabsorbed dose should, however, be further evaluated and validated inlarger studies. Further work is also needed to validate their function inthe response to 131I exposure.
5. Conclusion
In the present investigation, transcriptional microarray analysis wasperformed to evaluate changes in transcriptional activity in mouse thy-roid tissues in vivo 24 h after i.v. administration of 131I. Distinct and sub-stantial differences in gene expression and affected biological functionswere detected at the different absorbed dose levels. Furthermore, themajority of detected genes were dose-specific, and few genes werecommonly detected at all absorbed doses. We propose that theAgpat9, Plau, Prf1, and S100a8 genes might be novel potential absorbeddose-related biomarkers for 131I exposure. In addition, few previouslyproposed radiation-related biomarkers were detected in the presentdataset, most probably since they were primarily identified in in vitrostudies performed on several cell types, not only thyrocytes.
Competing interest
The authors declare that they have no competing interests.
Authors' contributions
NR and EFA designed the study, and NR and TZP carried out the an-imal trial. TZP did the extraction of RNA and pre-processing of the data.NR performed the first analysis of the data along with KH. All authorscontributed to the scientific and intellectual discussion and final inter-pretation of the data. NR drafted the manuscript, and all authors partic-ipated with substantial input and revision of the manuscript.
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.nucmedbio.2014.11.006.
Acknowledgments
The authors thank Lilian Karlsson and AnnWikström for their skilledtechnical assistance. This study was supported by grants from theEuropean Commission FP7 Collaborative Project TARCC HEALTH-F2-2007-201962, Swedish Research Council (grant no. 21073), SwedishCancer Society (grant no. 3427), BioCARE—a National Strategic Research
esponses in thyroid tissue in mice after 131I administration, Nucl Med

6 N. Rudqvist et al. / Nuclear Medicine and Biology xxx (2014) xxx–xxx
Program at University of Gothenburg, Swedish Radiation Safety Author-ity, King Gustav V Jubilee Clinic Cancer Research Foundation,SahlgrenskaUniversity Hospital Research Funds, Assar Gabrielsson Can-cer Research Foundation, and EC COST Action BM0607.
References
[1] Chan S, Kilby MD. Thyroid hormone and central nervous system development.J Endocrinol 2000;165(1):1–8.
[2] Hertz S, Roberts A, Salter WT. Radioactive iodine as an indicator in thyroid phys-iology. Iv. The metabolism of iodine in Graves' Disease. J Clin Invest 1942;21(1):25–9.
[3] Hall EJ, Giaccia AJ. Radiobiology for the radiologist. 6th ed. Philadelphia: LippincottWilliams & Wilkins; 2006[ix, 546 pp.].
[4] Prise KM, Schettino G, FolkardM, Held KD. New insights on cell death from radiationexposure. Lancet Oncol 2005;6(7):520–8.
[5] Morgan WF. Non-targeted and delayed effects of exposure to ionizing radiation:I. Radiation-induced genomic instability and bystander effects in vitro. Radiat Res2003;159(5):567–80.
[6] Mothersill C, Seymour CB. Radiation-induced bystander effects and the DNAparadigm: an "out of field" perspective. Mutat Res 2006;597(1–2):5–10.
[7] Chaudhry MA. Biomarkers for human radiation exposure. J Biomed Sci 2008;15(5):557–63.
[8] Snyder AR, Morgan WF. Gene expression profiling after irradiation: clues to under-standing acute and persistent responses? Cancer Metastasis Rev 2004;23(3–4):259–68.
[9] Zhao W, Chuang EY, Mishra M, Awwad R, Bisht K, Sun L, et al. Distinct effects ofionizing radiation on in vivo murine kidney and brain normal tissue geneexpression. Clin Cancer Res 2006;12(12):3823–30.
[10] Schuler E, Parris TZ, Rudqvist N, Helou K, Forssell-Aronsson E. Effects of internallow-dose irradiation from 131I on gene expression in normal tissues in Balb/cmice. EJNMMI Res 2011;1(29):1–14.
[11] Langen B, Rudqvist N, Parris TZ, Schuler E, Helou K, Forssell-Aronsson E. Comparativeanalysis of transcriptional gene regulation indicates similar physiologic response inmouse tissues at low absorbed doses from intravenously administered 211At.J Nucl Med 2013;54(6):990–8.
[12] Schuler E, Rudqvist N, Parris TZ, Langen B, Helou K, Forssell-Aronsson E. Transcrip-tional response of kidney tissue after 177Lu-octreotate administration in mice.Nucl Med Biol 2014;41(3):238–47.
[13] Schuler E, Rudqvist N, Parris TZ, Langen B, Spetz J, Helou K, et al. Time- and doserate-related effects of internal (177)Lu exposure on gene expression in mousekidney tissue. Nucl Med Biol 2014;41(10):825–32.
[14] Rudqvist N, Parris TZ, Schuler E, Helou K, Forssell-Aronsson E. Transcriptionalresponse of BALB/c mouse thyroids following in vivo astatine-211 exposure revealsdistinct gene expression profiles. EJNMMI Res 2012;2(32):1–12.
Please cite this article as: Rudqvist N., et al, Dose-specific transcriptional rBiol (2014), http://dx.doi.org/10.1016/j.nucmedbio.2014.11.006
[15] Bolch WE, Eckerman KF, Sgouros G, Thomas SR. MIRD pamphlet No. 21: a general-ized schema for radiopharmaceutical dosimetry–standardization of nomenclature.J Nucl Med 2009;50(3):477–84.
[16] Garg PK, Harrison CL, Zalutsky MR. Comparative tissue distribution in mice of thealpha-emitter 211At and 131I as labels of a monoclonal antibody and F(ab')2fragment. Cancer Res 1990;50(12):3514–20.
[17] Flynn AA, Green AJ, Pedley RB, Boxer GM, Boden R, Begent RH. A mouse model forcalculating the absorbed beta-particle dose from (131)I- and (90)Y-labeledimmunoconjugates, including a method for dealing with heterogeneity in kidneyand tumor. Radiat Res 2001;156(1):28–35.
[18] Saal LH, Troein C, Vallon-Christersson J, Gruvberger S, Borg A, Peterson C. BioArraySoftware Environment (BASE): a platform for comprehensive management andanalysis of microarray data. Genome Biol 2002;3(8) [p. SOFTWARE0003].
[19] Benjamini Y, Hochberg Y. Controlling the false discovery rate - a practical andpowerful approach to multiple testing. J R Statist Soc B 1995;57(1):289–300.
[20] AshburnerM,Ball CA,Blake JA,BotsteinD,ButlerH,Cherry JM,et al.Geneontology: tool forthe unification of biology. The Gene Ontology Consortium. Nat Genet 2000;25(1):25–9.
[21] Filiano AN, Fathallah-Shaykh HM, Fiveash J, Gage J, Cantor A, Kharbanda S, et al.Gene expression analysis in radiotherapy patients and C57BL/6 mice as a measureof exposure to ionizing radiation. Radiat Res 2011;176(1):49–61.
[22] Abend M, Pfeiffer RM, Ruf C, Hatch M, Bogdanova TI, Tronko MD, et al. Iodine-131dose-dependent gene expression: alterations in both normal and tumour thyroidtissues of post-Chernobyl thyroid cancers. Br J Cancer 2013;109:2286–94.
[23] Bentzen SM. Preventing or reducing late side effects of radiation therapy:radiobiology meets molecular pathology. Nat Rev Cancer 2006;6(9):702–13.
[24] Dent P, Yacoub A, Contessa J, Caron R, Amorino G, Valerie K, et al. Stress andradiation-induced activation of multiple intracellular signaling pathways. RadiatRes 2003;159(3):283–300.
[25] Mammen PP, Kanatous SB, Yuhanna IS, Shaul PW, Garry MG, Balaban RS, et al.Hypoxia-induced left ventricular dysfunction in myoglobin-deficient mice. Am JPhysiol Heart Circ Physiol 2003;285(5):H2132–41.
[26] Zhu P, Goh YY, Chin HF, Kersten S, Tan NS. Angiopoietin-like 4: a decade of research.Biosci Rep 2012;32(3):211–9.
[27] Lindencrona U, Nilsson M, Forssell-Aronsson E. Similarities and differences betweenfree 211At and 125I- transport in porcine thyroid epithelial cells cultured in bicam-eral chambers. Nucl Med Biol 2001;28(1):41–50.
[28] Lundh C, Lindencrona U, Postgard P, Carlsson T, Nilsson M, Forssell-Aronsson E.Radiation-induced thyroid stunning: differential effects of (123)I, (131)I, (99 m)Tc, and (211)At on iodide transport and NIS mRNA expression in cultured thyroidcells. J Nucl Med 2009;50(7):1161–7.
[29] Fong P. Thyroid iodide efflux: a team effort? J Physiol 2011;589(Pt 24):5929–39.[30] Malone JF. The radiation biology of the thyroid. Curr Top Radiat Res Q 1975;10(4):
263–368.[31] Yui S, Nakatani Y, Mikami M. Calprotectin (S100A8/S100A9), an inflammatory pro-
tein complex from neutrophils with a broad apoptosis-inducing activity. Biol PharmBull 2003;26(6):753–60.
[32] Wu Z, Podack ER, McKenzie JM, Olsen KJ, Zakarija M. Perforin expression by thyroid-infiltrating T cells in autoimmune thyroid disease. Clin Exp Immunol 1994;98(3):470–7.
esponses in thyroid tissue in mice after 131I administration, Nucl Med




















