
Targeting IL-10 Family Cytokines for the Treatment of Human ...
Upload
independentCategory
view
3download
0
ORIGINAL PAPER
Dose-rate effects of protons on in vivo activation of nuclearfactor-kappa B and cytokines in mouse bone marrow cells
Kanokporn Noy Rithidech • Paiboon Reungpatthanaphong •
Louise Honikel • Adam Rusek • Sanford R. Simon
Received: 9 October 2009 / Accepted: 11 May 2010 / Published online: 28 May 2010
� Springer-Verlag 2010
Abstract The objective of this study was to determine the
kinetics of nuclear factor-kappa B (NF-jB) activation and
cytokine expression in bone marrow (BM) cells of exposed
mice as a function of the dose rate of protons. The cytokines
included in this study are pro-inflammatory [i.e., tumor
necrosis factor-alpha (TNF-a), interleukin-1beta (IL-1b),and IL-6] and anti-inflammatory cytokines (i.e., IL-4 and
IL-10). We gave male BALB/cJ mice a whole-body expo-
sure to 0 (sham-controls) or 1.0 Gy of 100 MeV protons,
delivered at 5 or 10 mGy min-1, the dose and dose rates
found during solar particle events in space. As a reference
radiation, groups of mice were exposed to 0 (sham-controls)
or 1 Gy of 137Cs c rays (10 mGy min-1). After irradiation,
BM cells were collected at 1.5, 3, 24 h, and 1 month for
analyses (five mice per treatment group per harvest time).
The results indicated that the in vivo time course of effects
induced by a single dose of 1 Gy of 100 MeV protons or137Cs c rays, delivered at 10 mGy min-1, was similar.
Although statistically significant levels of NF-jB activation
and pro-inflammatory cytokines in BM cells of exposed
mice when compared to those in the corresponding sham
controls (Student’s t-test, p\ 0.05 or\0.01) were induced
by either dose rate, these levels varied over time for each
protein. Further, only a dose rate of 5 mGy min-1 induced
significant levels of anti-inflammatory cytokines. The
results indicate dose-rate effects of protons.
Introduction
Galactic cosmic rays (GCRs) and solar particle events
(SPEs) are the greatest sources of radiation exposure to
astronauts venturing beyond Earth. Hence, it is vitally
important to protect astronauts in space environments. This
will require improvement in our knowledge of radiation-
induced cellular and molecular changes that may impact
the health of astronauts. Moreover, for research on radia-
tion-induced genetic damage to be useful for risk assess-
ment, it must be established in in vivo systems, especially
in cells that are known to be at risk for health problems
associated with radiation exposure such as bone marrow
(BM) cells, the known target cell for radiation-induced
leukemia. While the biological effects of GCR particles, in
particular those with large mass (Z) and high energy (E)
(known as HZE particles, such as 56Fe ions) have been
investigated extensively (Durante and Cucinotta 2008),
little is known about the biological effects of SPE
components.
The SPEs are unpredictable in terms of the time of
occurrence and the size of the event (Cucinotta 1999;
Kim et al. 2009). Further, SPEs may last for several hours
up to a few days (Shea and Smart 1994). Exposure to
radiation during large SPEs can be life threatening if the
This manuscript is based on a contribution given at the ‘‘Heavy Ions
in Therapy and Space Symposium 2009,’’ July 6–10, 2009, Cologne,
Germany.
K. N. Rithidech (&) � P. Reungpatthanaphong � L. Honikel �S. R. Simon
Pathology Department, Stony Brook University,
Stony Brook, NY 11794-8691, USA
e-mail: [email protected]
Present Address:P. Reungpatthanaphong
Radio-Isotope Department, Faculty of Sciences,
Kasetsart University, Chatuchak, Bangkok 10900, Thailand
A. Rusek
Accelerator Department, NASA Research Laboratory,
Brookhaven National Laboratory, Upton, NY 11973-5000, USA
123
Radiat Environ Biophys (2010) 49:405–419
DOI 10.1007/s00411-010-0295-z

astronauts are inadequately protected (Cucinotta 1999; Hu
et al. 2009). Various organ dose rates (0–1 Gy h-1)
(Parsons and Townsend 2000) and doses (0–2 Gy)
(Wilson et al. 2006) of radiation from SPEs have been
estimated. The majority of ions produced during SPEs are
energetic protons ([30 MeV up to hundred and occa-
sionally thousands of MeV), followed by helium nuclei
(Cleghorn et al. 2004; Zapp et al. 1999). The flux of these
energetic protons drops rapidly with increasing energy
(Smart and Shea 1989). A large SPE produces very
intense charged particles composed almost entirely of
protons below 150 MeV. For these reasons, the focus of
this study was on the biological effects of 100 MeV
protons, which are one of the most abundant types of
protons induced during SPEs (Simpson 1983; Smart and
Shea 2002) making them the ions of interest for space-
radiation risk assessment. Further, information on bio-
logical effects of 100 MeV protons is currently lacking.
In contrast, using varying endpoints (e.g., micronu-
cleus assay and cell transformation), a few in vitro or in
vivo studies have been conducted to examine biological
effects of protons with different energies, i.e., 55, 200,
232, or 250 MeV, delivered at different dose rates,
ranging from 20 mGy min-1, 7, and 1 Gy min-1 (Chang
et al. 2005; Elmore et al. 2005; Green et al. 2001;
Gueulette et al. 2000). The focus of this study was on
the dose-rate effects of 100 MeV protons on the in vivo
activation of NF-jB and the expression of NF-jB-relatedproteins. As mentioned above, protons with varying
energies below 150 MeV are produced during SPEs;
thus, the data presented here are part of a study series to
investigate the in vivo biological effects of protons with
varying energies below 150 MeV, i.e., 30, 50, and 100
MeV.
In the past, it was generally assumed that protons induce
the same level of biological effects as those produced by X
and c rays. However, results from an in vitro study indi-
cated that 250 MeV protons induced higher levels of
damage (e.g., micronuclei, cell killing) in exposed rat
thyroid cells than in cells exposed to 60Co c rays, while thecell cycle phase distribution was not statistically different
(Green et al. 2001). Moreover, an in silico study indicated
that 1 GeV protons produce a higher level of clustered
DNA damage (presumably leading to subsequent double-
strand breaks) when compared to X or c rays (Hada and
Sutherland 2006). This important finding suggests that
health risks from exposure to protons (*1 GeV) may be
higher than previously thought. Further, it also shows that
protons with varying energies (most likely with diverse
LET) induce different levels of biological effects. More-
over, these results were in accordance with previously
observed phenomenon on LET-dependent biological
effects of radiation (Barendsen 1968, 1994).
Dose-rate effects of c or X rays, but not protons, have
been studied extensively. It has been well established, both
in in vivo and in vitro systems, that the clastogenic,
mutagenic, and carcinogenic activities of c or X rays
decrease for a given dose as the dose rate is decreased
(BEIR 2005; Boreham et al. 2000; Cornforth et al. 2002;
Elmore et al. 2006; Geard and Chen 1990; Kato et al. 2006;
Kumar et al. 2006; Little and Muirhead 1997; Loucas et al.
2004; Preston et al. 2002). Currently, there is no informa-
tion on the dose/dose-rate effects of 100 MeV protons
which are more abundant than 1 GeV protons during SPEs.
To provide important information directly related to the
NASA mission, we conducted experiments describing the
differences in dose-rate effects of 1 Gy of 100 MeV pro-
tons. Groups of mice were exposed to 100 MeV protons of
one selected moderate dose (1 Gy) delivered at two dif-
ferent dose rates (5 or 10 mGy min-1). As a reference
radiation, groups of mice were exposed to 0 or 1 Gy of137Cs c rays (delivered at 10 mGy min-1). Only one dose
rate of 137Cs c rays was used because there is abundant
information on dose/dose-rate effects of 137Cs c rays.
Further, the lowest dose rate possible from the c-irradiator(located in the Biology Department, Brookhaven National
Laboratory, Upton, New York) used in this study is
10 mGy min-1. Mice included in this study were also part
of a study series conducted to examine the extent and the
mechanisms involved in in vivo induction of genomic
instability (expressed as late occurring chromosome insta-
bility) by 100 MeV protons in which BM cells were col-
lected from groups of mice for analysis at different times
post-exposure. The scope of this study was to determine the
kinetics of NF-jB activation and the expression of selected
NF-jB target proteins known to be involved in inflam-
matory response, i.e., pro-inflammatory cytokines (TNF-a,IL-1b, and IL-6) and anti-inflammatory cytokines (IL-4
and IL-10) in BM cells of exposed mice.
An inflammatory process is the first cellular defense
mechanism in response to stress-inducing agents, including
radiation, in which several pro- and anti-inflammatory
cytokines are produced. NF-jB is a key transcription factor
involved in regulating the expression of such cytokines
(Baldwin 1996; Ghosh et al. 1998). An imbalance between
levels of pro-inflammatory and anti-inflammatory cyto-
kines at early and late times post-irradiation may contribute
to perpetuating inflammation. If inflammation or the pro-
duction of pro-inflammatory cytokines persists (chronic
inflammation), it becomes detrimental to exposed cells or
tissues. Further, NF-jB activation is also an important
contributor to cell survival and protection from apoptosis
following exposure to DNA-damaging agents in vitro (Liu
et al. 1996; Weichselbaum et al. 1994) and in vivo (Wang
et al. 2004). NF-jB is a member of the ubiquitously
expressed family of the Rel-related transcription factors
406 Radiat Environ Biophys (2010) 49:405–419
123

(Verma et al. 1995). These are p65 (RelA), p68 (RelB), p50
(NF-jB1), p52 (NF-jB2), and c-Rel. NF-jB proteins
interact to form homo- or heterodimers that bind DNA with
different specificities. The most prevalent and well studied
is NF-jB/p65. Hence, the activation of NF-jB/p65 is the
focus of this study.
The results from several in vitro studies showed that
low LET radiation (at varying dose and dose-rate levels)
induces NF-jB activation in different cell types such as
human lymphoblastoid cells (Mohan and Meltz 1994;
Prasad et al. 1994), human KG-I myeloid leukemia cells
(Brach et al. 1991), human glioma cell lines (Basu et al.
1998), human B-cells (Rho et al. 2005), or mouse skin
epithelial cells (Fan et al. 2007). Depending on the cell
types, elevation of activated NF-jB was detected at early
time points (1–8-h post-irradiation), then the levels of
activated NF-jB declined toward the level detected in
non-irradiated cells within 24 h. In contrast, information
on radiation-induced NF-jB activation in vivo is scant
(Rithidech et al. 2005; Zhou et al. 1999a, 2001). Like-
wise, there is little information on NF-jB activation
induced by space radiation and the existing knowledge is
derived from in vitro studies only. These include NF-jBactivation in human monocytes by 1 GeV/n 56Fe ions,
delivered at the dose rate of 1 Gy min-1 (Natarajan et al.
2002) and in human embryonic kidney (293/HEK) cells
by 95 MeV/n 36Ar ions (Baumstark-Khan et al. 2005;
Hellweg et al. 2005). To date, there is no information,
either in vitro or in vivo, on the activation of NF-jB as
well as the expression of NF-jB-targeted cytokines by
100 MeV protons. This study is the first to study the dose-
rate effects of 100 MeV protons on in vivo activation of
NF-jB and cytokines in a total cell population of the BM
using a mouse model.
Materials and methods
Animals
All male BALB/cJ mice were purchased from the Jackson
Laboratory (Bar Harbor, ME) and were delivered directly
from the vendor to Brookhaven National Laboratory
(BNL), Upton, New York, where irradiation took place.
They were allowed 2 weeks to acclimate prior to their
irradiation (at 10–12 weeks old, with an average body
weight of 25 g). They were housed in a facility approved
by the Association for Assessment and Accreditation of
Laboratory Animal Care (AAALAC) at BNL. The animal
rooms were maintained with the light cycle of 12 h light/
12 h dark, 21 ± 2�C, with 10-15 hourly cycles of fresh air
and a relative humidity of 50 ± 10%. The protocols for
animal housing and care, including experimental design of
the study, were approved by both the BNL and the Stony
Brook University (SBU) Institutional Animal Care and Use
Committee (IACUC).
Irradiation of mice
For 100 MeV proton-irradiation, the exposure was done at
the NASA Research Laboratory (NSRL) located at BNL,
Upton, New York. Mice were exposed whole body to the
average total-body doses of 0 or 1 Gy, delivered at the dose
rates of 5 or 10 mGy min-1 by a 20 9 20 cm beam. Mice
exposed to 0 Gy served as sham controls. As previously
mentioned, the selected dose and dose rates of 100 MeV
protons are comparable to those found in space. Further,
only 1 Gy of protons was used to compare with 0 Gy
(sham-control) because the focus of this study was on the
influence of dose rate, not the dose–response effect. For
each dose rate, a total of 40 mice were used. The dose-
average LET at the sample position was 0.7 keV/lm. The
direction of the 100 MeV protons to the target was hori-
zontal. The uniformity of the beam was measured to ensure
even dose distribution across the field of exposure. Further,
the exposure chambers were rotated 180 degrees at 10-min
intervals. Exposure procedures described previously
(Rithidech et al. 2005) were followed. Groups of mice that
were used for sample collection at 1-month post-irradiation
(10 mice for each dose of each dose rate of 100 MeV
protons) were transported back to SBU in a climate-
controlled vehicle within 1-week post-exposure. The
animal facility of SBU is also approved by the Association
for Assessment and Accreditation of Laboratory Animal
Care (AAALAC), with the same light cycle (12 h light/
12 h dark), temperature (21 ± 2�C), 10–15 hourly cycles
of fresh air, and relative humidity (50 ± 10%) as those
of BNL. Sample collections at 1.5, 3, and 24 h were
performed at the BNL Animal Facility.
For c-irradiation (a reference radiation, LET = 0.662
keV/lm), two groups of 20 mice in each were given a
whole body dose of 0 and 1 Gy of 137Cs c rays (at the doserate of 10 mGy min-1) using a gamma-irradiator located at
the Biology Department, BNL. Exposure procedures used
for 100 MeV protons were followed. Further, the direction
of c rays to the target was also horizontal. The schedule of
sample collections was the same as in mice exposed to
100 MeV protons.
Collection of bone marrow cells
At each harvest time for each dose rate, we collected BM
cells from each mouse (five mice per radiation dose) by
flushing both femurs and tibiae with 10 mL of McCoy’s
5A medium. As mentioned in the ‘‘Introduction’’ section,
the measurement of NF-jB activation presented in this
Radiat Environ Biophys (2010) 49:405–419 407
123

study is part of a study series examining the induction of
genomic instability (expressed as late occurring chromo-
somal damage). Consequently, the majority of BM cells
collected from each mouse were used in molecular cyto-
genetics for the detection of chromosomal damage.
Availability of BM cells used for the measurements of
NF-jB activation and levels of cytokines from each
mouse was therefore limited. Approximately 3 9 106
cells per mouse were available for the measurement of
NF-jB or cytokines.
Measurement of NF-jB activation
We used the enzyme-linked immunosorbent assay (ELISA)
NF-jB/p65 kits from Active Motif North America Inc.
(Carlsbad, CA) for measuring NF-jB activation in nuclear
extracts obtained from BM cells (approximately 3 9 106
cells) collected from each mouse at different times post-
irradiation (Rithidech et al. 2005). Nuclear extract kits
(Active Motif, Carlsbad, CA) were used for obtaining
nuclear lysates. Activation of NF-jB was measured using a
microplate spectrophotometer (Molecular Devices, Sun-
nyvale, CA) at 450 nm. It also should be noted that the data
obtained from microplate spectrophotometer readings rep-
resented the corrected values of the rate of tetra-methyl
benzidine conversion (expressed as mOD/minute) in actual
samples (including sham controls) after subtraction of the
substrate conversion rates in reagent blank samples. The
assay was performed in duplicate wells for each BM
sample.
Measurement levels of TNF-a, IL-1b, IL-6, IL-4,and IL-10
We measured the expression levels of these selected
cytokines involved in inflammatory response and known to
be regulated by NF-jB activation in BM cells (the cyto-
solic fraction) using the specific ELISA kits for each
cytokine from Biosource (Invitrogen, Carlsbad, CA).
Commercially available extraction kits (Active Motif,
Carlsbad, CA) were used for obtaining the cytosolic frac-
tion of BM cells. The method of extraction suggested by
the manufacturer was followed. Serial dilutions of known
concentrations of each cytokine were used to create a
standard curve for each of the cytokines included in the
study. The concentrations of the cytokines in BM samples
were calculated from their respective standard curves. The
levels of each cytokine were measured using a microplate
spectrophotometer (Molecular Devices, Sunnyvale, CA) at
450 nm. Similar to the measurements of activated NF-jB,the assay for each cytokine was performed in duplicate
wells for each BM sample.
Statistical analysis
We expressed NF-jB and cytokine data as mean ± SEM
and performed statistical analysis with the Student’s t-test.
Differences in these values between the exposed group and
the corresponding sham-control mice were considered
statistically significant at p-values\ 0.05.
Results
The summaries of the results are shown in Figs. 1, 2, 3, 4,
5, and 6. Each data point shown in each figure represents
mean values from 10 measurements (5 mice per group, 2
measurements per mouse) ± standard error of the mean
(SE). Additionally, the fold changes in levels of expression
of each protein in BM cells after exposure of mice to
radiation relative to the levels in their corresponding sham
controls at each time point were also presented in the fig-
ures adjacent to the relative amount of protein for each
radiation treatment protocol. The details of each measure-
ment are presented below.
NF-jB activation
The level of activated NF-jB in BM cells of male BALB/
cJ mice collected at 1.5 h after exposure to 1 Gy of
100 MeV protons delivered at 5 mGy min-1 did not differ
from that in sham controls (Fig. 1a). However, highly
significant levels (p\ 0.05) of activated NF-jB in BM
cells of exposed mice were found at 3- and 24-h post-
irradiation, and persisted up to 1-month post-exposure
(p\ 0.01). Such increases were 1.3-, 1.8-, and 3.8-fold
higher than those in their corresponding sham control
levels at 3-h, 24-h, and 1-month post-irradiation, respec-
tively (Fig. 1a.1).
In contrast to the exposure at the dose rate of
5 mGy min-1, the level of activated NF-jB in BM cells
collected at 1.5 h after exposure of male BALB/cJ mice to
1 Gy of either 100 MeV protons delivered at 10 mGy
min-1 or 137Cs c rays was significantly increased
(p\0.01) above that in the sham control samples (Fig. 1b, c).
Subsequently, at 3- and 24-h post-exposure, there was a
reduction in the levels of activated NF-jB in BM cells of
both sham control and exposed mice. However, the levels
of activated NF-jB in BM cells of exposed mice were still
higher (but not significantly) than those in their corre-
sponding sham controls. At late time (1 month) post-
exposure, a significant increase (p\ 0.05) in the level of
activated NF-jB was detected in BM cells of exposed
mice when compared to that in the BM cells of sham
controls. The results also indicate that a single dose of
408 Radiat Environ Biophys (2010) 49:405–419
123

1 Gy of 100 MeV protons exhibited a similar pattern of
effects to those induced by a single dose of 1 Gy of 137Cs crays, delivered at the same dose rate (10 mGy min-1). It
also should be noted that the levels of NF-jB activation in
BM cells of proton-exposed mice relative to those in their
corresponding sham controls were slightly higher than
those in the c-exposed mice compared to their corre-
sponding sham controls (Fig. 1b.1, c.1).
TNF-a
At 1.5 and 24 h (but not at 3 h) after exposure, the levels of
TNF-a in BM cells were significantly increased (p\ 0.01)
above those in their corresponding sham controls after
exposure of male BALB/cJ mice to 1.0 Gy of 100 MeV
protons delivered at 5 mGy min-1 (Fig. 2a). Further, the
highly significant level of TNF-a in BM cells of exposed
Fig. 1 Time course of NF-jB/p65 activation, measured by the
ELISA Trans-AM kits (Active
Motif North America Inc.,
Carlsbad, CA), in BM cells
collected from male BALB/cJ
mice following in vivo exposure
to 0 or 1 Gy of 100 MeV
protons delivered at
5 mGy min-1 (a) or at10 mGy min-1 (b). c Shows theeffects of 1 Gy of 137Cs c rays
(a reference radiation) delivered
at 10 mGy min-1. Each point
represents mean ± SE of five
mice (two measurements per
mouse). Fold changes in levels
of activated NF-jB/p65 induced
by each exposure regimen
relative to the corresponding
sham-control levels are shown
in a.1, b.1, and c.1, respectively.Significant differences in the
levels of activated NF-jB/p65are shown at *
Radiat Environ Biophys (2010) 49:405–419 409
123
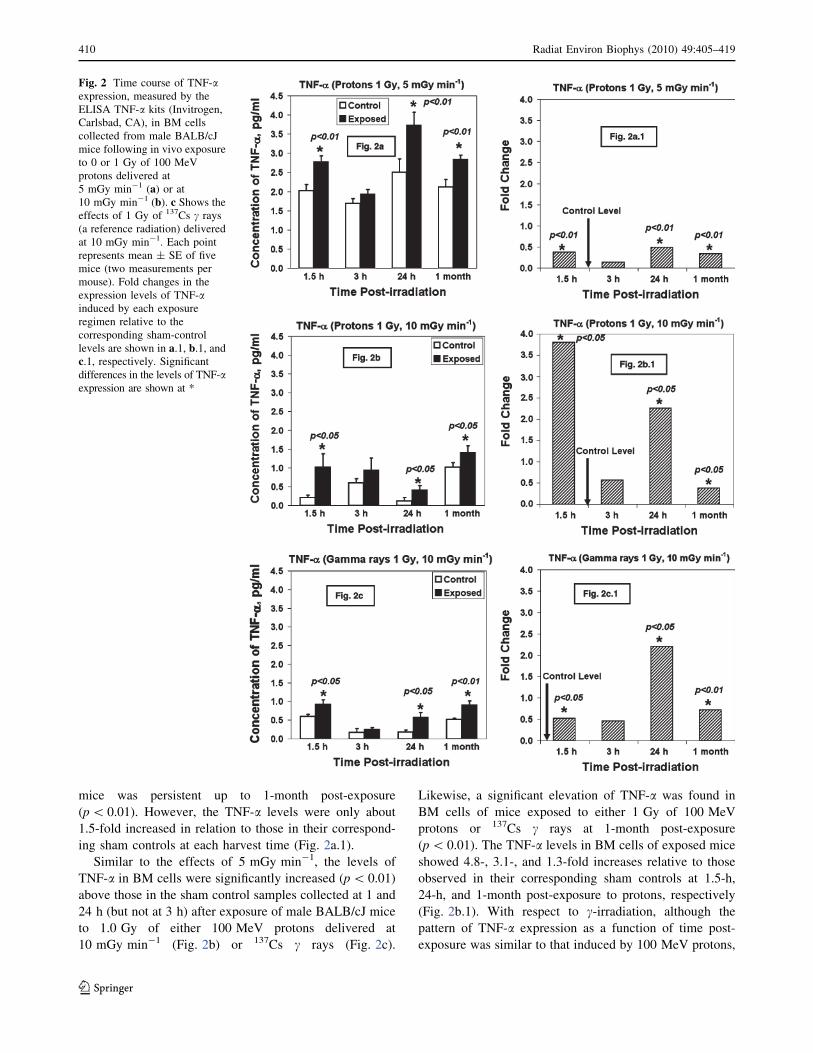
mice was persistent up to 1-month post-exposure
(p\ 0.01). However, the TNF-a levels were only about
1.5-fold increased in relation to those in their correspond-
ing sham controls at each harvest time (Fig. 2a.1).
Similar to the effects of 5 mGy min-1, the levels of
TNF-a in BM cells were significantly increased (p\ 0.01)
above those in the sham control samples collected at 1 and
24 h (but not at 3 h) after exposure of male BALB/cJ mice
to 1.0 Gy of either 100 MeV protons delivered at
10 mGy min-1 (Fig. 2b) or 137Cs c rays (Fig. 2c).
Likewise, a significant elevation of TNF-a was found in
BM cells of mice exposed to either 1 Gy of 100 MeV
protons or 137Cs c rays at 1-month post-exposure
(p\ 0.01). The TNF-a levels in BM cells of exposed mice
showed 4.8-, 3.1-, and 1.3-fold increases relative to those
observed in their corresponding sham controls at 1.5-h,
24-h, and 1-month post-exposure to protons, respectively
(Fig. 2b.1). With respect to c-irradiation, although the
pattern of TNF-a expression as a function of time post-
exposure was similar to that induced by 100 MeV protons,
Fig. 2 Time course of TNF-aexpression, measured by the
ELISA TNF-a kits (Invitrogen,
Carlsbad, CA), in BM cells
collected from male BALB/cJ
mice following in vivo exposure
to 0 or 1 Gy of 100 MeV
protons delivered at
5 mGy min-1 (a) or at10 mGy min-1 (b). c Shows theeffects of 1 Gy of 137Cs c rays
(a reference radiation) delivered
at 10 mGy min-1. Each point
represents mean ± SE of five
mice (two measurements per
mouse). Fold changes in the
expression levels of TNF-ainduced by each exposure
regimen relative to the
corresponding sham-control
levels are shown in a.1, b.1, andc.1, respectively. Significantdifferences in the levels of TNF-aexpression are shown at *
410 Radiat Environ Biophys (2010) 49:405–419
123
the increases in TNF-a levels in exposed mice relative to
the corresponding sham control levels were 1.5-, 3.2-, and
1.6-fold at 1.5-h, 24-h, and 1-month-post-exposure,
respectively (Fig. 2c.1).
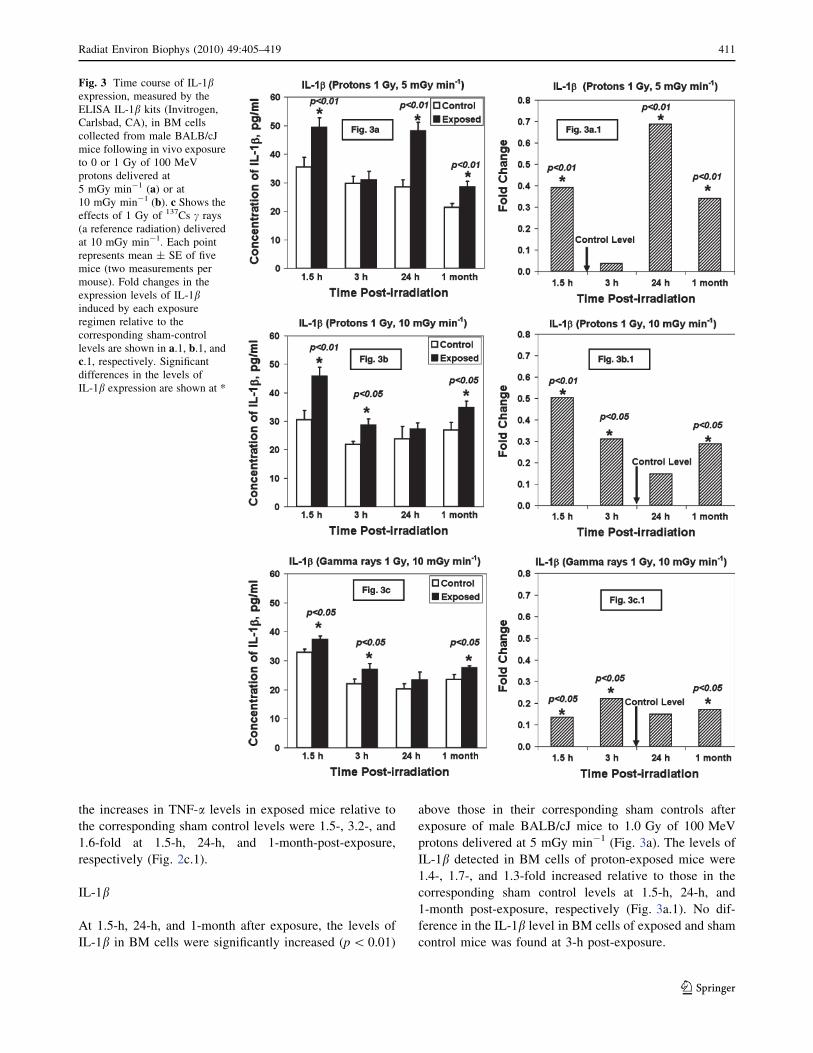
IL-1b
At 1.5-h, 24-h, and 1-month after exposure, the levels of
IL-1b in BM cells were significantly increased (p\ 0.01)
above those in their corresponding sham controls after
exposure of male BALB/cJ mice to 1.0 Gy of 100 MeV
protons delivered at 5 mGy min-1 (Fig. 3a). The levels of
IL-1b detected in BM cells of proton-exposed mice were
1.4-, 1.7-, and 1.3-fold increased relative to those in the
corresponding sham control levels at 1.5-h, 24-h, and
1-month post-exposure, respectively (Fig. 3a.1). No dif-
ference in the IL-1b level in BM cells of exposed and sham
control mice was found at 3-h post-exposure.
Fig. 3 Time course of IL-1bexpression, measured by the
ELISA IL-1b kits (Invitrogen,
Carlsbad, CA), in BM cells
collected from male BALB/cJ
mice following in vivo exposure
to 0 or 1 Gy of 100 MeV
protons delivered at
5 mGy min-1 (a) or at10 mGy min-1 (b). c Shows theeffects of 1 Gy of 137Cs c rays
(a reference radiation) delivered
at 10 mGy min-1. Each point
represents mean ± SE of five
mice (two measurements per
mouse). Fold changes in the
expression levels of IL-1binduced by each exposure
regimen relative to the
corresponding sham-control
levels are shown in a.1, b.1, andc.1, respectively. Significantdifferences in the levels of
IL-1b expression are shown at *
Radiat Environ Biophys (2010) 49:405–419 411
123

In contrast to the dose rate of 5 mGy min-1, the IL-1blevels in BM cells collected at 24 h after exposure of mice
to either 100 MeV protons delivered at 10 mGy min-1
(Fig. 3b) or 137Cs c rays (Fig. 3c) did not differ from those
of the corresponding sham control levels. However, sig-
nificant levels of IL-1b were detected in BM cells collected
at 1.5-h, 3-h, and 1-month after exposure of BABL/cJ mice
to either type of radiation, when compared to those in their
corresponding sham controls, with p-values ranging from
p\ 0.01 to \0.05. Figure 3b.1 and c.1 show the fold
changes in IL-1b levels detected in BM cells collected
from irradiated mice relative to those detected in the cor-
responding sham controls.
Il-6
After irradiation with 100 MeV protons at a dose rate of
5 mGy min-1, the expression levels of Il-6 in BM cells
Fig. 4 Time course of IL-6
expression, measured by the
ELISA IL-6 kits (Invitrogen,
Carlsbad, CA), in BM cells
collected from male BALB/cJ
mice following in vivo exposure
to 0 or 1 Gy of 100 MeV
protons delivered at
5 mGy min-1 (a) or at10 mGy min-1 (b). c Shows theeffects of 1 Gy of 137Cs c rays
(a reference radiation) delivered
at 10 mGy min-1. Each point
represents mean ± SE of five
mice (two measurements per
mouse). Fold changes in the
expression levels of IL-6
induced by each exposure
regimen relative to the
corresponding sham-control
levels are shown in a.1, b.1, andc.1, respectively. Significantdifferences in the levels of IL-6
expression are shown at *
412 Radiat Environ Biophys (2010) 49:405–419
123

collected from exposed mice were statistically higher than
those in the sham controls at 1.5-h (p\ 0.05), 24-h
(p\ 0.1), and 1-month (p\ 0.05) post-irradiation
(Fig. 4a). The levels of IL-6 in BM cells of exposed mice
were 1.4-, 1.7-, and 1.3-fold of those detected in the cor-
responding sham control levels (Fig. 4a.1). At 3 h after
exposure, however, IL-6 levels did not differ between
irradiated and sham control mice.
Expression levels of Il-6were highly significant inBMcells
of mice exposed to 1 Gy of 100 MeV protons delivered at
10 mGy min-1 (p\0.05, Fig. 4b) or 137Cs c rays (p\0.01,
Fig. 4c) only at 3-h post-exposure, when compared to
those of their corresponding sham controls. The highest
increase in IL-6 level related to that of the sham control
was found at 3-h post-irradiation, i.e., 1.8-fold for protons
and 2.3-fold for 137Cs c rays (Fig. 4b.1, c.1, respectively).
Fig. 5 Time course of IL-4
expression, measured by the
ELISA IL-4 kits (Invitrogen,
Carlsbad, CA), in BM cells
collected from male BALB/cJ
mice following in vivo exposure
to 0 or 1 Gy of 100 MeV
protons delivered at
5 mGy min-1 (a) or at10 mGy min-1 (b). c Shows theeffects of 1 Gy of 137Cs c rays
(a reference radiation) delivered
at 10 mGy min-1. Each point
represents mean ± SE of five
mice (two measurements per
mouse). Fold changes in the
expression levels of IL-4
induced by each exposure
regimen relative to the
corresponding sham-control
levels are shown in a.1, b.1, andc.1, respectively. Significantdifferences in the levels of IL-4
expression are shown at *
Radiat Environ Biophys (2010) 49:405–419 413
123
IL-4
The expression levels of IL-4 were significantly elevated in
BM cells collected at 1.5 h (p\ 0.05) and 24 h (p\ 0.01)
from mice exposed to 100 MeV protons at 5 mGy min-1
(Fig. 5a), when compared to their corresponding sham
controls. Such increases were 1.25- and 1.4-fold of the
corresponding sham control levels at 1.5 and 24 h,
respectively (Fig. 5a.1). At 3-h post-irradiation, the level of
IL-4 in BM cells of exposed mice was lower than that
detected in the sham control group. Further, there were no
differences in expression levels of IL-4 in BM cells of
exposed and sham control mice collected at 1-month post-
irradiation.
In contrast to the irradiation at the dose rate of
5 mGy min-1, there was a significant suppression of IL-4
levels in BM cells collected at 1.5 h after exposure of mice
to either 100 MeV protons delivered at 10 mGy min-1
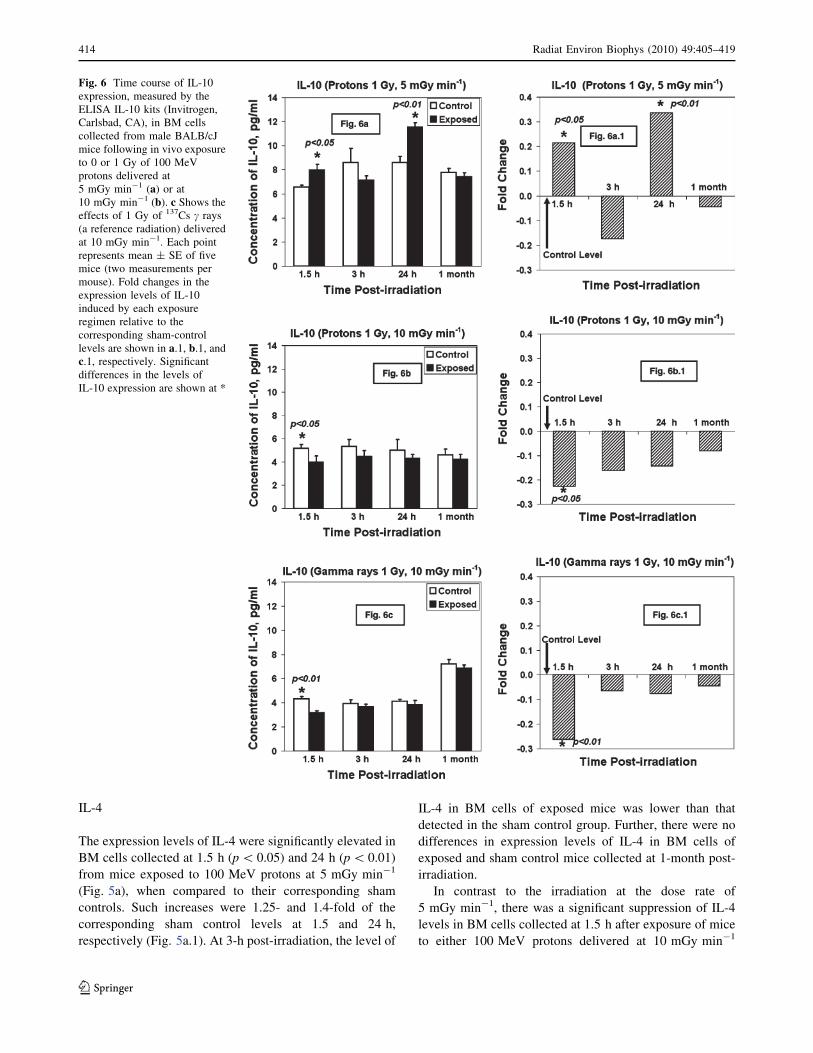
Fig. 6 Time course of IL-10
expression, measured by the
ELISA IL-10 kits (Invitrogen,
Carlsbad, CA), in BM cells
collected from male BALB/cJ
mice following in vivo exposure
to 0 or 1 Gy of 100 MeV
protons delivered at
5 mGy min-1 (a) or at10 mGy min-1 (b). c Shows theeffects of 1 Gy of 137Cs c rays
(a reference radiation) delivered
at 10 mGy min-1. Each point
represents mean ± SE of five
mice (two measurements per
mouse). Fold changes in the
expression levels of IL-10
induced by each exposure
regimen relative to the
corresponding sham-control
levels are shown in a.1, b.1, andc.1, respectively. Significantdifferences in the levels of
IL-10 expression are shown at *
414 Radiat Environ Biophys (2010) 49:405–419
123

(p\ 0.01, Fig. 5b, b.1) or 137Cs c rays (p\ 0.01, Fig. 5c,
c.1). Apparently, IL-4 levels in the exposed groups
remained lower than those in their corresponding sham
controls throughout the study (but not significant), although
there seemed to be some recovery of IL-4 production.
IL-10
Similar to the expression levels of IL-4, there were highly
significant levels of IL-10 in BM cells collected at 1.5 h
(p\ 0.05) and 24 h after exposure of mice (p\ 0.01) to
100 MeV protons at 5 mGy min-1 (Fig. 6a). There were
1.2- and 1.35-fold increases in IL-10 levels in exposed
groups relative to their corresponding sham controls at 1.5
and 24 h, respectively (Fig. 6a.1). At 3-h post-irradiation,
the level of IL-10 in BM cells of exposed mice was lower
than that detected in the sham control group. Further, the
IL-10 level in BM cells collected at 1 month from exposed
mice did not differ from that detected in their corre-
sponding sham controls.
There was a significant suppression of IL-10 levels in
BM cells collected at 1.5 h after exposure of mice to 1 Gy
of either 100 MeV protons delivered at 10 mGy min-1
(Fig. 6b) or 137Cs c rays (Fig. 6c). Subsequently, the pro-
duction of IL-10 was gradually increased in exposed mice,
but the levels of IL-10 remained lower than those detected
in the sham control groups (Fig. 6b, b.1, c, c.1).
Discussion
Our in vivo data demonstrated the impact of the dose rate
of 100 MeV protons on the pattern of NF-jB activation (as
well as the subsequent expression of proteins involved in
inflammatory response, i.e., cytokines known to be regu-
lated by activated NF-jB) in the total population of BM
cells of exposed mice. The cytokines included in this study
are pro-inflammatory cytokines (TNF-a, IL-1b, IL-6) andanti-inflammatory cytokines (IL-4 and IL-10). It is known
that the anti-inflammatory cytokines are necessary for
controlling and down-regulating inflammation (possibly by
counterbalancing the elevation of the pro-inflammatory
cytokines) and for maintaining homeostasis. In contrast,
the pro-inflammatory cytokines are primarily responsible
for initiating an effective inflammatory process. Our data
also demonstrated that a single dose of 1 Gy of 100 MeV
protons exhibited similar effects to those induced by a
single dose of 1 Gy of 137Cs c rays, delivered at the same
dose rate (10 mGy min-1). It also should be noted that
such a conclusion cannot be drawn between a dose rate of
5 mGy min-1 of 100 MeV protons and 137Cs c rays, sincethere were no data for the 5 mGy min-1 of 137Cs c rays forcomparison. However, it has been reported that increased
levels of pro-inflammatory cytokines were found with
increasing dose rates of 137Cs c rays (Ross et al. 1997).
We recognize that there are many other NF-jB-medi-
ated cytokines and that the production of these cytokines is
controlled by complex feedback mechanisms (Linard et al.
2003; Schreiber et al. 1998). The cytokines TNF-a, IL-1b,IL-4, IL-6, and IL-10 were selected for study because their
expressions, at the gene level, were elevated in human
mononuclear cells obtained from healthy adult individuals
who lived near the Chernobyl Nuclear Power Plant and
were chronically exposed to low-dose radiation ranging
from 0.18 to 49 mSv (Albanese et al. 2007). Further, these
cytokines were detected in the spleen or macrophages of
mice exposed to varying doses of radiation (ranging from
0.05 to 7.0 Gy) (Chang et al. 2000; Hosoi et al. 2001; Liu
et al. 2003). Additionally, these cytokines were detected in
the ileal muscular layer when a single dose of 10 Gy of60Co c rays (at a dose rate of 5 mGy min-1) was given to
the abdomen of male Wistar rats (Linard et al. 2004), or in
the bronchiolar epithelial cells when a single dose of 12 Gy
of 10 MV photons from a linear accelerator was given to
the thoracic region of C57BL/6J mice (Rube et al. 2005).
Exposure to 1 Gy of 100 MeV protons or 137Cs c rays
(delivered at 10 mGy min-1) rapidly activated NF-jB.Using this dose rate, a highly significant level of activated
NF-jB was detected as early as 1.5-h post-exposure.
Subsequently, there was a reduction in levels of activated
NF-jB, although higher levels (but not significant) were
found in BM cells of mice exposed to either 100 MeV
protons or 137Cs c rays, when compared to those of their
corresponding sham controls. At 1-month post-exposure,
there was another wave of a significantly high level of
activated NF-jB. In groups of mice exposed to 1 Gy of
100 MeV protons (delivered at 5 mGy min-1), however,
NF-jB activation was gradually increased. A significant
increase in the level of activated NF-jB was first observed
at 3-h (not at 1.5 h) post-irradiation in BM cells of exposed
mice. The mechanisms potentially involved in the delay of
NF-jB activation are currently unknown.
Our studies are the first in which the measurements of
NF-jB activation are done in BM cells collected at late
time (1 month) after exposure of mice to different dose
rates of radiation, specifically those found in space. Among
the three radiation treatment protocols, the highest increase
in the level of activated NF-jB was found in BM cells
collected at 1 month after exposure of mice to a single dose
of 1 Gy of 100 MeVn protons delivered at 5 mGy min-1
(i.e., a 3.8-fold increase when compared to the corre-
sponding control level, as shown in Fig. 1a.1, while there
were 2.2- and 1.5-fold increases in mice exposed to protons
or c rays with the dose rate of 10 mGy min-1 (Fig. 1b.1,
c.1, respectively). The finding of persistent increases in
the level of activated NF-jB at this late time (1 month)
Radiat Environ Biophys (2010) 49:405–419 415
123

post-exposure among mice exposed to either type of radi-
ation suggests that chronic inflammation might occur and
warrants further investigation. As mentioned above, the
NF-jB-mediated cytokine production is a complex system
leading to a loop of feedback mechanism. It has been
shown that pro-inflammatory cytokines [such as TNF-alpha
(Pikarsky et al. 2004; Todorov et al. 2004), or IL-1b(Catley et al. 2003; Lancel et al. 2008)] can activate NF-
jB. Hence, the persistence of significantly increased NF-
jB activation detected at 1-month post-irradiation may
reflect the feedback mechanisms induced by these cyto-
kines. Thus, the feedback loop can further enhance the
inflammation process. Whether or not the persistence of
NF-jB found in this study will be linked to the induction of
genomic instability (assessed by the presence of late
occurring chromosomal damage) in BM cells of exposed
mice remains unclear. Studies directed toward the deter-
mination of such an association are being investigated in
our laboratory.
In the past, the in vivo studies were carried out to
determine only the level of NF-jB activation at early time
(\24-h post-irradiation). Further, the earlier in vivo studies
used low LET radiation and high dose rates (Rithidech
et al. 2005; Zhou et al. 1999a). In a study previously
conducted in our laboratory, we measured the levels of
NF-jB activation in BM cells only at 1 and 4 h after
exposure of BALB/cJ mice to 0, 0.05, 0.1 or 1 Gy of 137Cs
c rays (delivered at 7.5 mGy min-1) (Rithidech et al.
2005). We found that 0.1 and 1.0 Gy (but not 0.05 Gy) of137Cs c rays were capable of activating NF-jB in BM cells
collected at 1-h (but not at 4 h) post-irradiation from
BALB/cJ mice (Rithidech et al. 2005). In that study, we
also observed no increase in the frequencies of chromo-
some aberrations in the group of mice without the signal of
early-activated NF-jB (i.e., mice exposed to 0.05 Gy of137Cs c rays) at any time point included in our study up to
6-month post-irradiation (unpublished data). Overall,
although the levels of activated NF-jB were determined at
early time points after exposure only, the findings suggest a
link between radiation-induced NF-jB activation and
radiation-induced genomic instability.
Further, tissue specificity of NF-jB activation by a high
dose of 137Cs c rays (8.5 Gy) has been reported after
exposure of the BALB/c mouse (Zhou et al. 1999b). In that
study, the investigators observed the highest level of NF-
jB activation in the BM, followed by the spleen and
mesenteric lymph nodes of the BALB/c mouse. In contrast,
there was no activated NF-jB signal in liver, lung, colon,
or brain of BALB/c mice exposed to a single dose of
8.5 Gy of 137Cs c rays. This high dose of radiation induced
a rapid increase in NF-jB activation, reaching a peak
within 1-h post-irradiation. Subsequently, the level of NF-
jB activation was decreased at 2.5 h, reaching background
levels at 5 h, and was less than the background level after
5-h post-irradiation. The same group of investigators also
found high levels of activated NF-jB in intestinal epithelial
cells of the small intestine after a whole-body exposure of
mice to varying doses of 137Cs c rays (0.5–12 Gy, at the
dose rate of 2.4 Gy min-1) (Wang et al. 2004). The authors
suggested that in vivo activation of NF-jB selectively
protects the small intestine against radiation exposure.
Additionally, it was shown that NF-jB activation is
directly involved in the expression of genes coding for pro-
inflammatory cytokines (i.e., TNF-a, IL-1b, and IL-6) in
the BM tissues of exposed mice (Zhou et al. 2001).
However, there was no information on NF-jB-mediated
expression of anti-inflammatory cytokines. Since NF-jBactivation regulates the production of both pro- and anti-
inflammatory cytokines, it is imperative to determine the
levels of both types of cytokines.
Our study also is the first in that the measurements of
pro- and anti-inflammatory cytokine levels (at the protein
level) are performed in BM cells of the same individual
mouse after a whole-body exposure to 100 MeV protons
delivered at the dose rates comparable to those found in
space. Similar to NF-jB activation, the highly significant
levels of expression of pro-inflammatory cytokines TNF-aand IL1-b in BM cells of exposed mice were found at early
time post-irradiation and persisted up to 1-month post-
irradiation. This phenomenon may also reflect the feedback
loop of NF-jB-mediated cytokine production, as observed
in the lung tissue of mice after thoracic irradiation (Rube
et al. 2005). It also has previously been suggested that a
high level of pro-inflammatory cytokines may contribute to
bystander-mediated effects and to genomic instability
(Lorimore et al. 2003; Moore et al. 2005).
With respect to IL-6, our data showed that a significant
level was persistent at 1-month post-irradiation only in BM
cells collected from mice exposed to 1 Gy of 100 MeV
protons, delivered at 5 mGy min-1. When a higher dose
rate (10 mGy min-1) was used, however, the IL-6 level in
BM cells of mice exposed to either 100 MeV protons or137Cs c rays did not differ from those detected in their
corresponding sham controls. Mechanisms involved in this
difference are currently unknown. However, it should be
noted that IL-6 (unlike TNF-a or IL-1b) has both pro- and
anti-inflammatory properties (Barton 1997; Opal and
DePalo 2000). Our data demonstrated that the time course
in the fold change of expression for IL-6 found in BM cells
of mice exposed to 1 Gy of 100 MeV protons, delivered at
5 mGy min-1 (Fig. 4a.1), is similar to those of the two
anti-inflammatory cytokines selected for study, i.e., IL-4
(Fig. 5a.1) or IL-10 (Fig. 6a.1).
IL-4 and IL-10 are known to be the important anti-
inflammatory cytokines (Opal and DePalo 2000). For each
radiation treatment protocol, the patterns of expression of
416 Radiat Environ Biophys (2010) 49:405–419
123

these two anti-inflammatory cytokines are similar. Highly
significant increases in levels of both cytokines were
detected in BM cells collected from mice exposed to 1 Gy
of 100 MeV protons delivered at 5 mGy min-1 only. In
contrast, a higher dose rate (10 mGy min-1) of 100 MeV
protons or 137Cs c rays significantly suppressed the pro-
duction of these two anti-inflammatory cytokines from
1.5 h up to 1-month post-exposure. Although there was
some recovery, the levels of IL-4 or IL-10 did not reach the
levels detected in their corresponding sham controls within
1-month post-irradiation (Figs. 5b, c, 6b, and c). It has been
established that both anti-inflammatory cytokines can
inhibit the production of pro-inflammatory cytokines TNF-
a and IL-1b (Cassatella et al. 1994; Linard et al. 2003; Opal
and DePalo 2000). It is thus possible that only a small
increase in TNF-a levels (about 1.3 fold increase,
Fig. 2a.1) observed in BM cells collected from mice
exposed to 100 MeV protons (delivered at 5 mGy min-1)
reflects the counterbalance action of anti-inflammatory
cytokines. In contrast, several fold increases in TNF-alevels were detected in BM cells collected from mice
exposed to a higher dose rate (10 mGy min-1) of radiation
(Fig. 2b.1, c.1), in which the levels of IL-4 and IL-10 were
significantly suppressed. Our data, however, showed that
production of IL-1b seems to be unaffected by the pro-
duction of IL-4 and Il-10. With the radiation dose used in
our study, it is possible that the production of IL-4 and
IL-10 is inadequate resulting in insufficient control over
pro-inflammatory cytokines TNF-a or IL-1b.In summary, we have detected differential patterns of in
vivo NF-jB activation and the expression of a subset of
cytokines in mouse BM cells as a function of the dose rate
of 1 Gy of 100 MeV protons. We also have found that the
in vivo time course of effects induced by a single dose of
1 Gy of 100 MeV protons or 137Cs c rays, delivered at the
same dose rate (10 mGy min-1), was similar. Further, the
persistence of NF-jB activation and pro-inflammatory
cytokines was detected at late time (1 month) post-exposure.
Such findings suggest the occurrence of the persistence of
cytokine production, presumably due to the feedback
mechanisms. The balance or imbalance of the anti- and
pro-inflammatory cytokines after radiation exposure may
play an important role in determining the extent of the
immune response and the health outcome. Our data provide
an important initial basis for risk assessment of in vivo
exposure to protons that are abundant in space during
SPEs. However, only a subset of cytokines was studied.
There are many other NF-jB-mediated cytokines (both
pro- and anti-inflammatory cytokines) such as interferon-
alpha (IFN-a) and IL-8, as well as their receptors. To
improve our understanding of the biological effects of
protons, it will be important in subsequent studies to
determine the levels of these cytokines and their receptors
after exposure to protons. This is particularly important
because the ultimate activities of cytokines are also regu-
lated by their receptors.
Acknowledgments We thank Dr. Peter Guida and his team for
logistic support, MaryAnn Petry and her BLAF staff for their assis-
tance in animal handling. We also thank Dr. Michael Sivertz for
dosimetry support. This work was supported by the National Aero-
nautics and Space Administration (NASA) Grant #NNX07AP88G.
References
Albanese J, Martens K, Karkanitsa LV, Dainiak N (2007) Multivar-
iate analysis of low-dose radiation-associated changes in cyto-
kine gene expression profiles using microarray technology. Exp
Hematol 35:47–54
Baldwin AS Jr (1996) The NF-kappa B and I kappa B proteins: new
discoveries and insights. Annu Rev Immunol 14:649–683
Barendsen GW (1968) Responses of cultured cells, tumours and
normal tissues to radiations of different linear enerygy transfer.
Curr Top Radiat Res 4:293–356
Barendsen GW (1994) RBE-LET relationships for different types of
lethal radiation damage in mammalian cells: comparison with
DNA dsb and an interpretation of differences in radiosensitivity.
Int J Radiat Biol 66:433–436
Barton BE (1997) IL-6: insights into novel biological activities. Clin
Immunol and Immunopathol 85:16–20
Basu S, Rosenzweig KR, Youmell M, Price BD (1998) The DNA-
dependent protein kinase participates in the activation of
NF[kappa]B following DNA damage. Biochem Biophys Res
Commun 247:79–83
Baumstark-Khan C, Hellweg CE, Arenz A, Meie MA (2005) Cellular
monitoring of the nuclear factor jB pathway for assessment of
space environmental radiation. Radiat Res 164:527–530
BEIR (2005) Health risks from exposure to low levels of ionizing
radiation: BEIR VII Phase 2 vol BEIR VII. The National
Academies
Boreham DR, Dolling JA, Maves SR, Siwarungsun N, Mitchel RE
(2000) Dose-rate effects for apoptosis and micronucleus forma-
tion in gamma-irradiated human lymphocytes. Radiat Res
153:579–586
Brach MA, Hass R, Sherman ML, Gunji H, Weichselbaum R, Kufe D
(1991) Ionizing radiation induces expression and binding activity
of the nuclear factor kappa B. J Clin Invest 88:691–695
Cassatella MA, Meda L, Gasperini S, Calzetti F, Bonora S (1994)
Interleukin 10 (IL-10) upregulates IL-1 receptor antagonist
production from lipopolysaccharide-stimulated human polymor-
phonuclear leukocytes by delaying mRNA degradation. J Exp
Med 179:1695–1699
Catley MC, Chivers JE, Cambridge LM, Holden N, Slater DM,
Staples KJ, Bergmann MW, Loser P, Barnes PJ, Newton R
(2003) IL-1[beta]-dependent activation of NF-[kappa]B medi-
ates PGE2 release via the expression of cyclooxygenase-2 and
microsomal prostaglandin E synthase. FEBS Lett 547:75–79
Chang CM, Elliott TB, Dobson ME, Jackson WE, Ledney GD (2000)
Ionizing radiation and bacterial challenge alter splenic cytokine
gene expression. J Radiat Res 41:259–277
Chang PY, Bjornstad KA, Rosen CJ, McNamara MP, Mancini R,
Goldstein LE, Chylack LT, Blakely EA (2005) Effects of iron
ions, protons and X rays on human lens cell differentiation.
Radiat Res 164:531–539
Cleghorn TF, Saganti PB, Zeitlin CJ, Cucinotta FA (2004) Solar
particle events observed at Mars: dosimetry measurements and
model calculations. Adv Space Res 33:2215–2218
Radiat Environ Biophys (2010) 49:405–419 417
123

Cornforth MN, Bailey SM, Goodwin EH (2002) Dose responses for
chromosome aberrations produced in noncycling primary human
fibroblasts by alpha particles, and by gamma rays delivered at
sublimiting low dose rates. Radiat Res 158:43–53
Cucinotta FA (1999) Issues in risk assessment from solar particle
events. Radiat Meas 30:261–268
Durante M, Cucinotta FA (2008) Heavy ion carcinogenesis and
human space exploration. Nat Rev Cancer 8:465–472
Elmore E, Lao XY, Ko M, Rightnar S, Nelson G, Redpath J (2005)
Neoplastic transformation in vitro induced by low doses of
232 MeV protons. Int J Radiat Biol 81:291–297
Elmore E, Lao XY, Kapadia R, Redpath JL (2006) The effect of dose
rate on radiation-induced neoplastic transformation in vitro by
low doses of low-LET radiation. Radiat Res 166:832–838
Fan M, Ahmed KM, Coleman MC, Spitz DR, Li JJ (2007) Nuclear
factor-{kappa}B and manganese superoxide dismutase mediate
adaptive radioresistance in low-dose irradiated mouse skin
epithelial cells. Cancer Res 67:3220–3228
Geard CR, Chen CY (1990) Micronuclei and clonogenicity following
low- and high-dose-rate gamma irradiation of normal human
fibroblasts. Radiat Res 124:S56–S61
Ghosh S, May MJ, Kopp EB (1998) NF-kappa B and Rel proteins:
evolutionarily conserved mediators of immune responses. Annu
Rev Immunol 16:225–260
Green LM, Murray DK, Bant AM, Kazarians G, Moyers MF, Nelson
GA, Tran DT (2001) Response of thyroid follicular cells to
gamma irradiation compared to proton irradiation. I. Initial
characterization of DNA damage, micronucleus formation,
apoptosis, cell survival, and cell cycle phase redistribution.
Radiat Res 155:32–42
Gueulette J, Bohm L, Slabbert JP, De Coster BM, Rutherfoord GS,
Ruifrok A, Octave-Prignot M, Binns PJ, Schreuder AN, Symons
JE, Scalliet P, Jones DTL (2000) Proton relative biological
effectiveness (RBE) for survival in mice after thoracic irradia-
tion with fractionated doses. Int J Radiat Oncol Biol Phys
47:1051–1058
Hada M, Sutherland BM (2006) Spectrum of complex DNA damages
depends on the incident radiation. Radiat Res 165:223–230
Hellweg CE, Arenz A, Meier MM, Baumstark-Khan C (2005)
Cellular monitoring systems for the assessment of space
environmental factors. Adv Space Res 36:1673–1679
Hosoi Y, Miyachi H, Matsumoto Y, Enomoto A, Nakagawa K,
Suzuki N, Ono T (2001) Induction of interleukin-1beta and
interleukin-6 mRNA by low doses of ionizing radiation in
macrophages. Int J Cancer 96:270–276
Hu S, Kim M-HY, McClellan GE, Cucinotta FA (2009) Modeling the
acute health effects of astronauts from exposure to large solar
particle events. Health Phys 96:465–476
Kato TA, Nagasawa H, Weil MM, Genik PC, Little JB, Bedford JS
(2006) gamma-H2AX foci after low-dose-rate irradiation reveal
atm haploinsufficiency in mice. Radiat Res 166:47–54
Kim M-HY, Hayat MJ, Feiveson AH, Cucinotta FA (2009) Prediction
of frequency and exposure level of solar particle events. Health
Phys 97:68–81. doi:10.1097/01.HP.0000346799.65001.9c
Kumar PR, Mohankumar MN, Hamza VZ, Jeevanram RK (2006)
Dose-rate effect on the induction of HPRT mutants in human G0
lymphocytes exposed in vitro to gamma radiation. Radiat Res
165:43–50
Lancel S, Bachschmid MM, Kirber MT, Weinberg EO (2008)
Abstract 3378: IL-33 translocates to the nucleus and has NF-kB
transcriptional repressor function following treatment with IL-
1beta in human endothelial cells. Circulation 118:S_415-a
Linard C, Ropenga A, Vozenin-Brotons MC, Chapel A, Mathe D
(2003) Abdominal irradiation increases inflammatory cytokine
expression and activates NF-{kappa}B in rat ileal muscularis
layer. Am J Physiol Gastrointest Liver Physiol 285:G556–G565
Linard C, Marquette C, Mathieu J, Pennequin A, Clarencon D, Mathe
D (2004) Acute induction of inflammatory cytokine expression
after [gamma]-irradiation in the rat: effect of an NF-[kappa]B
inhibitor. Int J Radiat Oncol Biol Phys 58:427–434
Little MP, Muirhead CR (1997) Curvilinearity in the dose-response
curve for cancer in Japanese atomic bomb survivors. Environ
Health Perspect 105(Suppl 6):1505–1509
Liu Z-g, Hsu H, Goeddel DV, Karin M (1996) Dissection of TNF
receptor 1 effector functions: JNK activation is not linked to
apoptosis while NF-[kappa]B activation prevents cell death. Cell
87:565–576
Liu XD, Ma SM, Liu SZ (2003) Effects of 0.075 Gy x-ray irradiation
on the expression of IL-10 and IL-12 in mice. Phys Med Biol
48:2041–2049
Lorimore SA, Coates PJ, Wright EG (2003) Radiation-induced
genomic instability and bystander effects: inter-related nontar-
geted effects of exposure to ionizing radiation. Oncogene
22:7058–7069
Loucas BD, Eberle R, Bailey SM, Cornforth MN (2004) Influence of
dose rate on the induction of simple and complex chromosome
exchanges by gamma rays. Radiat Res 162:339–349
Mohan N, Meltz ML (1994) Induction of nuclear factor kappa B after
low-dose ionizing radiation involves a reactive oxygen interme-
diate signaling pathway. Radiat Res 140:97–104
Moore SR, Marsden S, Macdonald D, Mitchell S, Folkard M, Michael
B, Goodhead DT, Prise KM, Kadhim MA (2005) Genomic
instability in human lymphocytes irradiated with individual
charged particles: involvement of tumor necrosis factor alpha in
irradiated cells but not bystander cells. Radiat Res 163:183–190
Natarajan M, Aravinda N, Meltz M, Herman T (2002) Post-
translational modification of I-kappa B alpha activates NF-?B
in human monocytes exposed to 56Fe ions. Radiat Environ
Biophys 41:139–144
Opal SM, DePalo VA (2000) Anti-inflammatory cytokines. Chest
117:1162–1172
Parsons JL, Townsend LW (2000) Interplanetary crew dose rates for
the August 1972 solar particle event. Radiat Res 153:729–733
Pikarsky E, Porat RM, Stein I, Abramovitch R, Amit S, Kasem S,
Gutkovich-Pyest E, Urieli-Shoval S, Galun E, Ben-Neriah Y
(2004) NF-[kappa]B functions as a tumour promoter in inflam-
mation-associated cancer. Nature 431:461–466
Prasad AVM, Mohan N, Meltz ML (1994) Activation of nuclear
factor kappa B in human lymphoblastoid cells by low-dose
ionizing radiation. Radiat Res 138:367–372
Preston DL, Mattsson A, Holmberg E, Shore R, Hildreth NG, Boice
JD Jr (2002) Radiation effects on breast cancer risk: a pooled
analysis of eight cohorts. Radiat Res 158:220–235
Rho H-S, Kim S-H, Lee C-E (2005) Mechanism of NF-kB activation
induced by gamma-irradiation in B-lymphoma cells: role of ras.
J Toxicol Environ Health 68:2019–2031
Rithidech K, Tungjai M, Arbab E, Simon SR (2005) Activation of
NF-kappa B in bone marrow cells of BALB/cJ mice following
exposure in vivo to low doses of 137Cs gamma rays. Radiat Env
Biophys 44:139–143
Ross HJ, Canada AL, Antoniono RJ, Redpath JL (1997) High and low
dose rate irradiation have opposing effects on cytokine gene
expression in human glioblastoma cell lines. Eur J Cancer
33:144–152
Rube C, Uthe D, Wilfert F, Ludwig D, Yang K, Konig J, Palm J,
Schuck A, Willich N, Remberger K, Rube C (2005) The
bronchiolar epithelium as a prominent source of pro-inflamma-
tory cytokines after lung irradiation. Int J Radiat Oncol Biol Phys
61:1482–1492
Schreiber S, Nikolaus S, Hampe J (1998) Activation of nuclear factor
kappaB in inflammatory bowel disease. Gut 42:477–484
Shea MA, Smart DF (1994) Recent and historical solar proton events
418 Radiat Environ Biophys (2010) 49:405–419
123

Simpson JA (1983) Elemental and isotopic composition of the
galactic cosmic rays. Annu Rev Nucl Part Sci 33:323
Smart DF, Shea M (1989) Solar proton events during the past three
solar cycles. J Spacecr Rockets 26:403–415
Smart DF, Shea MA (2002) A review of solar proton events during
the 22nd solar cycle. Adv Space Res 30:1033–1044
Todorov VT, Volkl S, Muller M, Bohla A, Klar J, Kunz-Schughart
LA, Hehlgans T, Kurtz A (2004) Tumor Necrosis Factor-alpha
activates NF kappa B to inhibit renin transcription by targeting
cAMP-responsive element. J Biol Chem 279:1458–1467
Verma IM, Stevenson JK, Schwarz EM, Van Antwerp D, Miyamoto S
(1995) Rel/NF-kappa B/I kappa B family: intimate tales of
association and dissociation. Genes Dev 9:2723–2735
Wang Y, Meng A, Lang H, Brown SA, Konopa JL, Kindy MS,
Schmiedt RA, Thompson JS, Zhou D (2004) Activation of
nuclear factor kappaB in vivo selectively protects the murine
small intestine against ionizing radiation-induced damage.
Cancer Res 64:6240–6246
Weichselbaum RR, Hallahan D, Fuks Z, Kufe D (1994) Radiation
induction of immediate early genes: effectors of the radiation-
stress response. Int J Radiat Oncol Biol Phys 30:229–234
Wilson JW, Anderson BM, Cucinotta FA, Ware J, Zeitlin CJ (2006,
July) Spacesuit radiation shield design methods. Paper presented
at the International Conference on Environmental Systems,
Norfolk
Zapp EN, Ramsey CR, Townsend LW, Badhwar GD (1999) Solar
particle event dose and dose-rate distributions: parameterization
of dose-time profiles, with subsequent dose-rate analysis. Radiat
Meas 30:393–400
Zhou D, Brown S, Yu T, Chen G, Barve S, Kang BC (1999a) A high
dose of ionizing radiation induces tissue-specific activation of
nuclear factor-kappaB in vivo. Rad Res 151:703–709
Zhou D, Brown SA, Yu T, Chen G, Barve S, Kang BC, Thompson JS
(1999b) A high dose of ionizing radiation induces tissue-specific
activation of nuclear factor-kappaB in vivo. Radiat Res 151:703–
709
Zhou D, Yu T, Chen G, Brown SA, Yu Z, Mattson MP, Thompson JS
(2001) Effects of NF-kappaB1 (p50) targeted gene disruption on
ionizing radiation-induced NF-kappaB activation and TNFalpha,
IL-1alpha,[ IL-1beta and IL-6 mRNA expression in vivo. Int J
Radiat Biol 77:763–772
Radiat Environ Biophys (2010) 49:405–419 419
123
Copyright © 2022 FDOKUMEN




















