
AT Homopolymer Strings in Salmonella enterica Subspecies I ...
Upload
independentCategory
view
1download
0
Diversification of a Salmonella virulence effector protein functionby ubiquitin-dependent differential localization
Jayesh C. Patel1, Karsten Hueffer1,3, Tukiet T. Lam2, and Jorge E. Galán1,*
1Section of Microbial Pathogenesis, Yale University School of Medicine, 295 Congress Av., New Haven, CT06536
2W. M. Keck Foundation Biotechnology Resource Laboratory, Yale University School of Medicine, 300George St., New Haven, CT06511
SUMMARYMany bacterial pathogens and symbionts utilize type III secretion systems to deliver bacterial effectorproteins into host cells. These effector proteins have the capacity to modulate a large variety ofcellular functions in a highly regulated manner. Here we report that the phosphoinositide phosphataseSopB, a Salmonella Typhimurium type III secreted effector protein, diversifies its function bylocalizing to different cellular compartments in a ubiquitin-dependent manner. We show that SopButilizes the same enzymatic activity to modulate actin-mediated bacterial internalization and Aktactivation at the plasma membrane, and vesicular traffic and intracellular bacterial replication at thephagosome. Thus by exploiting the host cellular machinery, Salmonella Typhimurium has evolvedthe capacity to broaden the functional repertoire of a virulence factor to maximize its ability tomodulate cellular functions.
Keywordsbacterial pathogenesis; phosphoinositide phosphatase; type III secretion; vesicular trafficking; actinremodeling
INTRODUCTIONBacterial pathogens that have co-existed with their hosts for extended periods of time haveevolved complex functional interfaces involving specific bacterial adaptations to modulatecellular functions and secure the pathogen’s survival and replication. One of these adaptationsis the type III secretion system (TTSS), a multiprotein nano-machine that mediates the deliveryof bacterial effector proteins into target host cells (Galan and Wolf-Watz, 2006). Salmonellaenterica serovar Typhimurium (S. Typhimurium) encodes two of these machines within itspathogenicity islands 1 and 2 (SPI-1 and SPI-2), which at different times during infection,deliver more than 60 proteins into host cells (Galán, 2001; Waterman and Holden, 2003). Theseeffector proteins have the capacity to modulate a variety of cellular processes, including actin
*For correspondence Phone: (203) 737-2404, Fax: (203) 737-2630, E-mail: E-mail: [email protected] address: Institute of Arctic Biology, University of Alaska Fairbanks, Fairbanks, AK 99775SUPPLEMENTAL DATA Supplemental data for this article include 7 figures and 3 videosPublisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customerswe are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resultingproof before it is published in its final citable form. Please note that during the production process errors may be discovered which couldaffect the content, and all legal disclaimers that apply to the journal pertain.
NIH Public AccessAuthor ManuscriptCell. Author manuscript; available in PMC 2010 April 17.
Published in final edited form as:Cell. 2009 April 17; 137(2): 283–294. doi:10.1016/j.cell.2009.01.056.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

dynamics, vesicular trafficking, and transcriptional responses (Galán, 2001; Waterman andHolden, 2003), and many do so by mimicking the activities of host cell proteins (Stebbins andGalán, 2001). It is thought that the activity of type III secreted effector proteins within targetcells must be regulated so that their function is exerted at the appropriate time and subcellularspace. However, little is known about mechanisms that control these critical aspects of bacterialeffector protein function. One of the effector proteins delivered by the S. Typhimurium SPI-1TTSS is the phosphoinositide phosphatase SopB (Galyov et al., 1997; Hong and Miller,1998; Norris et al., 1998). This effector protein mediates a diverse set of responses at differenttimes during infection. Through the activation of SGEF, an exchange factor for the Rho-familyGTPase RhoG, SopB mediates actin-dependent bacterial internalization (Patel and Galan,2006; Zhou et al., 2001). In addition, SopB modulates vesicular trafficking by altering themetabolism of phosphoinositides at the Salmonella phagosomal membrane (Hernandez et al.,2004; Mallo et al., 2008). Furthermore, SopB activates the serine protein kinase Akt (Steele-Mortimer et al., 2000) and stimulates the production of nitric oxide (Drecktrah et al., 2005) bypoorly understood mechanisms. Remarkably, all of these activities are strictly dependent onthe phosphatase activity of SopB. The mechanism by which this effector modulates such adiverse set of functions with the same enzymatic activity is not understood. We show here thatSopB diversifies its function by localizing to different cellular compartments at different timesduring infection. Early in infection, SopB localizes to the plasma membrane to mediatebacterial entry and Akt activation. Later in infection, SopB localizes to the Salmonellacontaining vacuole where it is required for bacterial replication. We also show that thetranslocation of SopB from the plasma membrane to the Salmonella-containing vacuolerequires its ubiquitination. Therefore by co-opting the host-cell ubiquitination machinery,SopB can modulate distinct functions during the infection process utilizing the same enzymaticactivity.
RESULTSUbiquitinated SopB persists during S. Typhimurium infection of cultured epithelial cells
The observation that SopB modulates different cellular processes at different times duringinfection prompted us to examine the levels of translocated SopB throughout the infectionprocess. We used a S. Typhimurium strain expressing carboxyl-terminus FLAG-tagged SopB,which had been introduced in its chromosome by allelic exchange (see Materials and Methods).Epitope-tagged SopB was found to behave in a manner that was indistinguishable from wildtype SopB (Fig. S1). SopB could be detected within infected cells from as early as a few minutesto up to several hours after infection (Fig. 1A), which is consistent with previous results(Drecktrah et al., 2005). Although a proportion of SopB detected late in infection representednewly synthesized protein, we found that a significant fraction represented protein deliveredearly in infection since it was detected even in the presence of a bacterial protein synthesisinhibitor (Fig. 1B). Shortly after infection, a significant proportion of translocated SopB wasrapidly modified resulting in a change of its mobility in SDS PAGE (Fig. 1A and B), anobservation previously attributed to its ubiquitination (Marcus et al., 2002;Rogers et al.,2008). To confirm that the change in SopB mobility was due to its ubiquitination, COS-2 cellstransiently expressing HA-tagged ubiquitin were infected with S. Typhimurium, and 60 minafter infection, SopB was immunoprecipitated with a specific monoclonal antibody.Immunoprecipitated proteins were separated by SDS-PAGE and the presence of ubiquitinconjugated to SopB was probed by Western immunoblotting using a specific antibody to theHA epitope tag. As shown in Fig. 1C, ubiquitinated SopB was readily detected in infected cellsconfirming that the observed change in its mobility upon translocation is due to itsubiquitination.
Patel et al. Page 2
Cell. Author manuscript; available in PMC 2010 April 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

S. Typhimurium encodes three E3 ubiquitin ligases, the Hect-like protein SopA (Zhang et al.,2006), as well as three other proteins, SspH1 (absent from some strains including the one usedin this study), SlrP, and SspH2, which belong to a novel family of E3 ligases (Quezada et al.,2008; Rohde et al., 2007). These effector proteins are also delivered by the SPI-1 and SPI-2-encoded type III secretion systems (Miao et al., 1999). We investigated the potentialinvolvement of threes E3 ligases and found that the pattern of SopB ubiquitination after itsdelivery by the S. Typhimurium ΔsspH2 ΔslrP ΔsopA triple mutant was indistinguishable fromthat observed after its delivery by wild type S. Typhimurium (Fig. 1D). These results indicatethat SopB ubiquitination does not require any of the bacterially-encoded E3 ubiquitin ligases.
We have previously shown that two other effectors delivered by the SPI-1 TTSS, SopE andSptP, are also ubiquitinated upon translocation (Kubori and Galan, 2003). In this case,ubiquitination serves as a degradation signal since these effectors are degraded upontranslocation. In contrast, ubiquitinated SopB persisted within cells for extended periods oftime even in the presence of a bacterial protein synthesis inhibitor (Fig. 1B). Furthermore,ubiquitinated SopB could be readily detected in the absence of proteasome inhibitors (Fig. 1Aand B), and addition of a proteasome inhibitor did not significantly alter the levels ofubiquitinated SopB (Fig. S2). These results suggest that, in the case of SopB, ubiquitinationmay play a role other than that of a degradation signal. Consistent with this hypothesis, themobility shift observed in SopB subsequent to its translocation into mammalian cells indicatesthat monoubiquitination is the most common modification observed in this effector (Fig. 1A-D). In fact, SopB may undergo multi-monoubiquitination since in immunoprecipitationexperiments conducted in cells over-expressing lysine-less ubiquitin, which cannot formubiquitin chains although it can be conjugated to substrates, we still observed laddering in theSopB mobility (Fig. 1E). We also observed the presence of ubiquitinated SopB in infected cellsoverexpressing a ubiquitin mutant in which its lysine 63 (K63-ubiquitin) was the only lysineavailable for conjugation (Fig. 1E). Like monoubiquitination, K63-linked ubiquitin is notusually associated with degradation (Urbe, 2005). However, these results do not rule out thatthe laddering observed under these conditions may also be the result of multi-monoubiquitination by K63 ubiquitin. Together, these results further support the hypothesisthat ubiquitination of SopB plays a role other than that of a degradation signal.
Identification of the SopB ubiquitination sitesTo evaluate the role of ubiquitination in SopB function we mapped its ubiquitination sites. Weisolated translocated SopB from infected cells by affinity purification and subjected it to massspectrometric analysis to identify ubiquitinated peptides. This analysis identified ubiquitinatedSopB fragments consisting of amino acids 14-23, and 24-41 (Fig. S3). These results, inconjunction with the ubiquitin laddering observed in infected cells expressing lysine-lessubiquitin (see above), suggested that several lysine residues located at the amino terminus ofSopB could serve as potential ubiquitination sites. Consequently, we focused our analysis onthe amino terminus of SopB, which comprises 9 lysine residues (Fig. 2A). We generated aseries of FLAG-tagged point mutants, replacing each of these lysine residues, eitherindividually or in combination, in the context of full-length SopB. The different SopB mutantswere expressed in S. Typhimurium, tested for their secretion from the bacteria, and assayedfor ubiquitination upon TTSS-mediated translocation into cultured epithelial cells. Allconstructs were expressed, secreted, and translocated into cultured cells at levelsindistinguishable from those of wild type SopB (Fig. 2B). Surprisingly, individual substitutionof each one of all the lysine residues located within the first 120 amino acids of SopB did notprevent its ubiquitination (Fig. 2B). Furthermore, even pair-wise substitution of lysine residueswithin this region had a negligible effect on SopB ubiquitination (Fig. 2B). Only when all thelysine residues contained within this region were substituted was SopB ubiquitinationabrogated, resulting in its migration as a predominantly single species at the predicted
Patel et al. Page 3
Cell. Author manuscript; available in PMC 2010 April 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

molecular weight of non-ubiquitinated SopB (Fig. 2B). Moreover, additional mutation of alysine residue located at the carboxy terminus (K541), which has previously been shown to beubiquitinated upon transient over expression of SopB in cultured cells (Rogers et al., 2008),had no effect on the ubiquitination profile of SopB (Fig. S4). In addition, the mutant in whichall the amino terminal lysines had been substituted did not co-immunoprecipitate with epitopetagged ubiquitin (data not shown). These results revealed a high degree of complexity in thepattern of SopB ubiquitination, with different lysines potentially serving as targets forubiquitination. Similar observations have been made for the ubiquitination sites of theepidermal growth factor receptor (Haglund et al., 2003a;Haglund et al., 2003b;Huang et al.,2006).
The non-ubiquitinated SopB mutant accumulates at the plasma membrane of infected cellsfollowing its TTSS-mediated delivery
To investigate the role of SopB and its ubiquitination during infection we compared thelocalization of the non-ubiquitinated (K1-9 substituted, see Fig. 2) SopB mutant (from hereforth referred to as SopBΔub) with that of wild type SopB. Cultured epithelial cells were infectedwith S. Typhimurium strains expressing a chromosomally encoded FLAG tagged wild-typeSopB or its SopBΔub mutant derivative constructed by allelic exchange, and the localizationof these effector proteins was examined by confocal microscopy. Ten minutes after infection,wild-type SopB was observed localized at the plasma membrane, tightly around enteringbacteria (Fig. S5). Following internalization, wild-type SopB was seen decorating theSalmonella containing vacuoles, and its association with this compartment was maintained forup to 3 hrs (Fig. 3A, supplementary Video S1, and data not shown). In contrast, the SopBΔub
mutant revealed a strikingly different localization at the plasma membrane in the form ofsurface puncta radiating from the site of infection (Fig. 3B, Supplementary Video S2, and datanot shown). Within 30 minutes of infection, diffuse SopBΔub staining over the entire plasmamembrane was readily evident in x-z and y-z projections following reconstruction of confocalz-stacks (Supplementary Video S2). Despite the significant differences in localization,biochemical fractionation of lysates of host cells infected with bacteria expressing the differentconstructs indicated that the levels of translocated SopBΔub mutant protein were equivalent tothose of wild type SopB (Fig. 3C). The half-life within host cells of the non-ubiquinated SopBmutant was equivalent to wild type SopB (data not shown), further demonstrating that, in thecase of this effector, ubiquitination does not serve as a degradation signal. Consistent with therole of ubiquitination in SopB localization, mutants in which lysine substitutions (individualor in combination) did not result in changes in its ubiquitination pattern (Fig. 2B) localized ina manner indistinguishable to wild type SopB (Fig. S6).
To further explore the role of ubiquitination in the localization of SopB, we examined thedistribution of the SopBΔub mutant upon delivery by bacteria competent for TTSS-mediatedprotein translocation but defective for internalization. We reasoned that this would allow us toexamine the distribution of this effector protein without the potential influence of actinremodeling and bacterial entry. To render the bacteria unable to induce its own internalization,we constructed catalytic mutant derivatives of the wild type (SopBC460S) and its non-ubiquitinated mutant (SopBΔub C460S), and expressed them in a strain lacking the effectorsSopE and SopE2, which are functionally redundant with SopB in mediating bacterial entry.When delivered in the context of this non-invasive strain, SopBC460S was seen in smallintracellular vesicle-like compartments that could be labeled with endocytic tracers (Fig. 3D).In addition, mixed infection experiments showed that SopB delivered by non-invasive bacteriacould be observed surrounding phagosomes of invasive bacteria, demonstrating that thisdelivery pathway can result in the transfer of translocated SopB to Salmonella-containingphagosomes (supplementary video S3). SopBC460S was readily ubiquitinated upon deliveryinto cells by non-invasive bacteria, indicating that its phosphatase activity is dispensable for
Patel et al. Page 4
Cell. Author manuscript; available in PMC 2010 April 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

this modification (Fig. 3E). In contrast, SopBΔubC460S delivered by non-invasive bacteria wasseen localized exclusively at the plasma membrane, did not co-localize with endocytic tracers(Fig. 3D) or bacterial-containing phagosomes after mixed infections, and as expected, was notubiquitinated upon its delivery to infected cells (Fig. 3E). These results indicate that thelocalization of SopB to an intracellular vesicular compartment such as the Salmonella-containing vacuole is not the consequence of its passive trapping in the phagosomal membraneduring bacterial uptake. Rather, these results indicate that ubiquitination may serve as a signalnecessary for the removal of SopB from the host cell plasma membrane and its delivery tointracellular vesicular compartments such as the Salmonella-containing vacuole. Thismechanism is reminiscent of the ubiquitination–dependent internalization mechanisms ofgrowth factors receptors after stimulation by their ligands (Dikic, 2003). However, our resultscannot rule out that during bacterial internalization, ubiquitination may also serve as a signalto localize SopB to the nascent phagosome and contribute to its targeting to the bacterialphagosome.
Identification of a domain required for SopB association with the host cell membraneWe found that following translocation into host cells SopB partitioned primarily to a membranefraction, and could not be extracted by high salt concentrations (Fig. 4A). Similar observationshave been previously made following transient over-expression of SopB in epithelial cells(Marcus et al., 2002). SopBΔub also partitioned to the membrane fraction and was equallyresistant to extraction by salt, which indicates that ubiquitination is not required for SopB toassociate with host-cell membranes after TTSS-mediated translocation (Fig. 4A). Theassociation of SopB with the host-cell membrane in a manner resistant to extraction with highsalt treatment suggests a strong interaction equivalent to that of a hydrophobic domain thatmay be intimately associated with the membrane. We examined the SopB sequence for thepresence of such a domain and identified a region between amino acids 288 and 309 whosefeatures would be consistent with those of a transmembrane domain as indicated by the TMpredprediction algorithm (data not shown). However, this domain does not appear to be a bonafide transmembrane domain since other prediction algorithms did not detect it (data not shown).To investigate the potential contribution of this domain to the membrane localization of SopB,we constructed a S. Typhimurium strain expressing a SopB mutant lacking this hydrophobicregion (SopBΔ288-309) and examined its phenotype. SopBΔ288-309 was secreted from thebacteria and delivered into host cells by the SPI-1 TTSS, albeit at slightly lower levels thanthose of the wild-type protein (Fig. 4B). Importantly, translocated SopBΔ288-309 migrated as asingle species on SDS-PAGE with the predicted molecular weight of the unmodified protein,which indicates that, unlike wild-type SopB, this mutant form cannot be ubiquitinated (Fig.4B). Furthermore, translocated SopBΔ288-309 exhibited a cytoplasmic localization (Fig. 4C),indicating that the hydrophobic domain is essential for the localization of SopB to host cellmembranes. A S. Typhimurium ΔsopE ΔsopE2 mutant strain expressing SopBΔ288-309 wasdefective for entry (Fig. 4D) and Akt activation (Fig. 4E), indicating that SopB must be ableto associate with host cell membranes to exert these functions. The phenotype of theSopBΔ288-309 mutant is unlikely to be due to gross conformational changes caused by thedeletion of the hydrophobic domain since purified SopBΔ288-309 exhibited almost wild-typephosphatase activity in vitro (Fig. 4F). However, this experiment cannot rule out the possibilitythat loss of function of this SopB mutant may be due to localized conformational changesinduced by introduction of the small deletion. Taken together, however, these results indicatethat the association of SopB with host cell membranes requires a discrete hydrophobic domainand is critical for its ubiquitination and function.
Patel et al. Page 5
Cell. Author manuscript; available in PMC 2010 April 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

S. Typhimurium expressing SopBΔub promotes increased actin remodeling,macropinocytosis, and Akt activation
To investigate the functional significance of SopB ubiquitination we examined the SopB-dependent phenotypes of a S. Typhimurium strain expressing the ubiquitination-deficientSopBΔub mutant. Since SopB is functionally redundant with the Rho-family GTPase exchangefactors SopE and SopE2 in mediating bacterial internalization (Zhou et al., 2001) (Patel andGalan, 2006), we expressed the SopBΔub mutant in a strain lacking these exchange factors sothat signaling for entry was exclusively delivered through SopBΔub. We found that a S.Typhimurium strain expressing SopBΔub was able to enter cells in a manner indistinguishablefrom that of a strain expressing wild type SopB (Fig. 5A). Interestingly, the S. TyphimuriumsopBΔub mutant strain induced more profuse actin cytoskeletal rearrangements than the strainexpressing wild type SopB. While the strain expressing wild-type SopB induced localized actinremodeling, the strain expressing SopBΔub induced diffuse membrane ruffling along the cellperiphery that coincided with a loss in cellular stress fibers (Fig. 5B and 5C). Moreover, incontrast with the transient actin remodeling induced by wild-type SopB, the actinrearrangements stimulated by the S. Typhimurium sopBΔub mutant strain persisted for up to 3hours after infection (data not shown). These results suggest that SopB triggers signaling forentry when it is localized at the plasma membrane and that its persistence at this location resultsin increased signaling to the actin cytoskeleton. These results also suggest that the removal ofSopB from the plasma membrane by an ubiquitination-mediated mechanism may contributeto the downregulation of this signaling event after bacterial entry.
We investigated whether ubiquitination of SopB influences the magnitude and temporalkinetics of macropinocytosis, a phenotype also dependent on SopB (Hernandez et al., 2004;Mallo et al., 2008). Henle-407 cells infected with S. Typhimurium expressing wild-type SopBor the SopBΔub mutant were analyzed by live microscopy for their ability to inducemacropinocytosis. In comparison to a wild type strain, cells infected with the S. TyphimuriumsopBΔub mutant exhibited increased and more prolonged macropinocytic activity (Fig. 6A and6B). These results suggest that the persistence of SopBΔub at the plasma membrane enhancesits ability to stimulate macropinocytosis.
We also compared the activation of Akt over time in cells infected with S. Typhimuriumexpressing wild type SopB or the SopBΔub mutant. Although at 30 minutes after infection thelevels of Akt activation were equivalent in cells infected with either bacterial strain, cellsinfected with the S. Typhimurium sopBΔub mutant showed more sustained kinase activation(Fig. 6C). Three hours after infection the levels of phosphorylated (activated) Akt were higherin cells infected with the S. Typhimurium sopBΔub mutant (Fig. 6C), which correlates with theprolonged presence of SopBΔub at the plasma membrane. These results suggest that SopBactivates Akt at the plasma membrane.
In summary, persistence of SopB at the plasma membranes resulted in enhanced ability tostimulate actin remodeling, macropinocytosis, and Akt activation, suggesting that theseactivities require the presence of SopB at this location.
S. Typhimurium expressing SopBΔub is defective for intracellular replicationIt has been previously shown that SopB is required for the efficient intracellular replication ofS. Typhimurium by modulating the vesicular trafficking of the Salmonella-containing vacuole(SCV) (Hernandez et al., 2004). It has also been recently shown that recruitment of Rab5 tothe SCV is dependent on the presence of SopB, and it has been postulated that this recruitmentis necessary for efficient S. Typhimurium intracellular replication (Mallo et al., 2008). Wereasoned that these phenotypes must depend on the presence of SopB at the SCV, in whichcase a S. Typhimurium expressing SopBΔub, which is retained at the plasma membrane, may
Patel et al. Page 6
Cell. Author manuscript; available in PMC 2010 April 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

exhibit a defect in these phenotypes. To test this hypothesis, we first compared the ability ofS. Typhimurium expressing either wild-type SopB or the SopBΔub mutant with a ΔsopB strainfor their ability to recruit Rab5 to the SCV. Both the ΔsopB and the strain expressing theSopBΔub mutant were equally defective in their ability to recruit Rab5 to the SCV (Fig. 7Aand Supplementary Fig. S7). To further evaluate the importance of ubiquitin-mediated SopBlocalization to the SCV, we compared the ability of a S. Typhimurium ΔsopE ΔsopE2 mutantstrain expressing either wild-type SopB or the SopBΔub mutant to grow within epithelial cells.Although both strains were internalized at equivalent levels, 24 hs after infection a significantlylower number of colony forming units were recovered from cells infected with the strainexpressing the SopBΔub mutant (Fig. 7B). We also compared the S. typhimurium ΔsopB mutantwith wild type S. Typhimurium or an isogenic mutant strain expressing SopBΔub for their abilityto survive within primary bone-marrow-derived macrophages. As shown in Fig. 7C, whencompared to wild type S. Typhimurium, the ΔsopB and the strain expressing SopBΔub wereequally defective in their ability to survive within macrophages. Introduction of a plasmidencoding wild type sopB readily complemented the S. Typhimurium strain expressing theSopBΔub mutant for its ability to survive within macrophages. Combined, these results areconsistent with the hypothesis that SopB modulation of bacterial intracellular replicationrequires the localization of SopB at the SCV. In addition, together with the results presentedabove, these findings indicate that the differential localization of SopB at different sites allowsit to modulate different cellular processes and hence diversify its function.
DISCUSSIONType III secretion systems have the capacity to mediate a very diverse array of cellularfunctions. They do so by delivering a variety of effector proteins, which in many cases functionas “mimics” of eukaryotic cell proteins (Galan, 2007; Stebbins and Galán, 2001). In order toexert their function, the activity of these effector proteins must be precisely coordinated andregulated in time and space. For example, in S. Typhimurium, two effector proteins, SopE andSptP, modulate Rho-family GTPases through opposing biochemical activities. SopE activatesRho-family members functioning as a guanine nucleotide exchange factor (GEF) for theseGTPases (Hardt et al., 1998) while SptP down-modulates the activity of the same GTPases byfunctioning as a GTPase-activating protein (GAP) (Fu and Galán, 1999). These effectors ensureactivation of the GTPases to allow bacterial internalization while preserving cellularhomeostasis, which is transiently altered during bacterial entry. The opposing activities of thesetwo effectors are coordinately regulated by their ubiquitin-dependent differential degradation(Kubori and Galan, 2003). We have shown here another example of ubiquitin-mediatedregulation of effector function. Rather than degradation, the mechanism described hereimplicates ubiquitination in the differential localization of the effector protein SopB during theinfection cycle, resulting in the diversification of its function.
Upon translocation from the bacteria, SopB localizes to the plasma membrane of infected cellsfrom where it mediates actin cytoskeleton reorganization, macropinocytosis, bacterial entry,and Akt activation (Fig. 7D). We found that this localization depends on a hydrophobic regionof SopB, which tightly tethers this effector protein to the host-cell plasma membrane. However,membrane localization may involve additional determinants since previous transienttransfection studies have identified an additional region of SopB capable of mediating itsputative membrane association or affecting its solubility under transient over expressionconditions (Marcus et al., 2002). Shortly after its translocation, SopB is multi-monoubiquitinated. This modification acts as a signal required for its removal from the host-cell plasma membrane and its subsequent delivery to internal vesicular compartments and/orfor its “trapping” on the nascent bacterial phagosome. At this location, SopB modulates vesicletraffic and facilitates intracellular bacterial replication. Consistent with this model, anubiquitination-defective SopB mutant remained localized to the plasma membrane throughout
Patel et al. Page 7
Cell. Author manuscript; available in PMC 2010 April 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

bacterial infection, and a S. Typhimurium strain expressing this mutant exhibited an increasedand prolonged ability to stimulate actin cytoskeleton rearrangements and Akt activation. Incontrast, this strain was defective for intracellular survival, which is dependent on theappropriate vesicle trafficking of the SCV. Therefore, ubiquitination is central not only for thedown modulation of the early activities of SopB but also for the diversification of its function.In this context, the parallels with the role of ubiquitination in the modulation and diversificationof growth factor receptor function are striking (Dikic, 2003; Haglund et al., 2003a). Uponstimulation by its ligand, the EGFR is monoubiquitinated and removed from the plasmamembrane to be delivered to an endocytic compartment where it can further stimulatedownstream signaling, or it can be sorted for recycling or degradation. Like SopB, in theabsence of ubiquitination, EGFR remains at the plasma membrane resulting in constitutivesignaling. Furthermore, the similarities extend to the highly redundant nature of the potentialsites for ubiquitination. We found that several lysines within the amino terminus of SopB canbe targets for monoubiquitination. Likewise, several lysines clustered around the kinasedomain of EGFR can be alternatively monoubiquitinated (Haglund et al., 2003a;Haglund etal., 2003b;Huang et al., 2006). However, despite these striking similarities, there are significantdifferences in the cellular machinery utilized for the removal and transport of SopB and EGFRfrom the plasma membrane to endocytic compartments. The ubiquitination-dependent removalof the EGFR from the plasma membrane is mediated by a complex mechanism that involvesproteins belonging to the endosomal sorting complexes required for transport (ESCRT)(Saksena et al., 2007), as well as dynamin and Epsin (Bache et al., 2006;Chen et al.,1998;Damke et al., 1994;Dikic, 2003;Malerød et al., 2007;Raiborg et al., 2008). In contrast,depletion of or interference with dynamin, clathrin, Epsin, or members of the ESCRT complexdid not affect the location of SopB (Supplementary Fig. S8) implying that an as yet unidentifiedbut potentially different cellular machinery is involved in SopB’s removal from the plasmamembrane and translocation to endocytic compartments.
In summary, we have described here a remarkable mechanism by which, through differentiallocalization, a virulence factor diversifies its function to modulate rather distinct cellularprocesses with the same biochemical activity. This is another example of the extraordinarycomplexity of the pathogen/host cell functional interface sculpted by evolutionary forcesdirected at securing pathogen and host survival.
EXPERIMENTAL PROCEDURESPlasmids, Bacterial Strains and Cell Lines
Full-length wild-type SopB and its derivatives (including truncations and deletions), tagged atthe C-terminus by a FLAG epitope were cloned into the arabinose inducible vector pBAD24(Guzman et al., 1995) by standard recombinant DNA procedures. SopB mutants harboringsingle or combinatorial Lys (K) to Glu (Q) substitutions at positions 13, 19, 23, 37, 41, 67, 93,109 and 111 were generated by site-directed mutagenesis and verified by DNA sequencing.Eukaryotic expression vectors encoding Epsin UIM and K44A Dynamin were provided byPietro de Camilli (Yale University), YFP tagged Vps4a derivatives (pEYFP-Vps4a andpEYFP-E228QVps4a) were provided by W. Sandquist (University of Utah), and HA taggedwild-type, K63, and K0 ubiquitin were provided by Yihong Ye (NIH). siGENOMESMARTpool (M-016835-00, Dharmacon) at a final concentration of 50nM was used to knockdown expression of endogenous HRS. RNAi efficacy was verified by qRT-PCR, using HRSprimers as previously described (Patel and Galan, 2006). Cells were transfected usingLipofectamine 2000 (Invitrogen) according to the manufacturers instructions.
The wild-type strain of S. Typhimurium SL1344 (Hoiseth and Stocker, 1981) and its isogenicderivatives used in this study, ΔsopB (SB1120) (Hernandez et al., 2004), ΔsopE ΔsopE2(SB1301), and ΔsopE ΔsopE2 ΔsopB (SB1302) (Zhou et al., 2001), have been previously
Patel et al. Page 8
Cell. Author manuscript; available in PMC 2010 April 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

described. Strains expressing chromosomally encoded FLAG tagged wild-type SopB and itsderivatives were constructed by standard recombinant DNA and allelic exchange proceduresas previously described (Kaniga et al., 1994). All bacterial strains were cultured underconditions that stimulate the expression of the Salmonella Pathogenicity Island-1-encoded typeIII secretion system (Galán and Curtiss III, 1990). Where appropriate, 0.05% l-arabinose wasadded to cultures at the early logarithmic phase of growth (OD600nm of 0.4) to induceexpression of genes under the control of the para BAD promoter.
Type III protein secretion and translocation assayBacterial protein secretion and translocation into host cells were examined by biochemicalfractionation and western immunoblot as previously described (Collazo and Galán, 1997;Kubori and Galan, 2003). Where appropriate, the proteasome inhibitor MG132 (5μM,Calbiochem) or its vehicle solvent DMSO, was added to cells 30 min prior to infection andmaintained throughout the infection and lysis procedure. In bacterial protein synthesisinhibition experiments, chloramphenicol was added at a final concentration of 100μg/ml atdifferent time points during infection.
SopB immunoprecipitationCOS-2 cells seeded in 10 cm dishes and transfected with a plasmid encoding HA-taggedubiquitin were infected with S. Typhimurium expressing chromosomally encoded SopB FLAG(m.o.i. 30) at 37°C, 5% CO2. At different time points following infection, cells were harvestedon ice by scraping into 1ml modified RIPA lysis buffer (50mM Tris pH7.6, 150mM NaCl, 1%NP40, 0.5% sodium deoxycholate, 0.1% SDS, 20mM NEM, 1x protease cocktail (Roche),1mM PMSF) per well. Lysates were centrifuged at 20,000 × g at 4°C for 20 min to removeinsoluble material. An aliquot of the supernatant was saved (~ 100μl) to monitor levels of SopBFLAG translocation and HA-ubiquitin expression whilst the remainder (~900μl) was incubatedwith 15μl pre-washed anti-FLAG M2 agarose (50% slurry, Sigma), rotating for 1 hr at 4°C toprecipitate translocated SopB. Immune complexes were collected by centrifugation at 1,000 ×g for 5 min at 4°C, washed twice with 1ml cold lysis buffer and boiled in SDS loading bufferto dissociate proteins from beads. Samples were analyzed by SDS PAGE (8% gel) and westernblotting using antibodies directed against HA and FLAG.
Subcellular fractionation and extractionHenle-407 cells (10 cm plate) were infected (m.o.i. 60) for 1 hr as described above, washedand harvested by scraping gently into homogenization buffer (20mM Hepes, pH 7.2, 200mMsucrose, 0.5mM EGTA, 1mM PMSF, 1x protease inhibitor cocktail). Cells were disrupted bymechanical lysis using a dounce homogeniser (25x passage) and lysates were centrifuged twiceat 20,000g, 4°C for 30 min to remove bacteria and debris. The resulting supernatant wascentrifuged at 100,000g 4°C to separate the membrane (pellet) from the cytoplasmic(supernatant) fractions. Extraction of SopB from the pellet fraction was performed byresuspending in buffer containing 1M NaCl, 100mM sodium bicarbonate (pH11) or 1% TritonX-100 and incubating on ice for 30 min. Samples were centrifuged at 100,000g to separatemembrane and cytoplasmic fractions. The presence of SopB in the supernatant fraction wasdetermined by TCA precipitation and western blot analysis.
S. Typhimurium induced ruffling, macropinocytosis assays, and Rab5 recruitmentS. Typhimurium-induced ruffling and macropinocytosis assays were performed in Ref-52 andHenle-407 cells as previously described (Chen et al., 1996; Patel and Galán, 2008; Patel andGalan, 2006). Rab5 recruitment was examined in Henle-407 cells following transfection ofGFP-Rab5 as previously described (Mallo et al., 2008).
Patel et al. Page 9
Cell. Author manuscript; available in PMC 2010 April 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Extracellular delivery of SopBTo monitor the fate of SopB delivered into cells from extracellular bacteria we infectedHenle-407 cells with a DsopE ΔsopE2 ΔsopB S. Typhimurium mutant strain expressing acatalytically inactive FLAG-epitope tagged SopB mutant (SopBC460S) at an m.o.i. of 25. Thisstrain is non-invasive but it is fully competent for type III secretion and protein delivery. Priorto infection, cultured cells were loaded with alexa 594 dextran (100,00MW, 1mg/ml) to labelthe endocytic pathway. One hour after infection, cells were washed three times with cold HBSS,fixed and processed for immunofluorescence. To determine whether SopB delivered fromextracellular bacteria could associate with phagosomes derived from invasive S. Typhimuriumwe performed mixed infection experiments. Henle-407 cells were co-infected with wild typeS. Typhimurium expressing DsRED as a marker and a non-invasive a DsopE ΔsopE2 ΔsopBS. Typhimurium mutant strain expressing a catalytically inactive FLAG-epitope tagged SopBmutant (SopBC460S) at a ratio of 1:5. Cells were infected with a combined m.o.i of 40 for 1 hrbefore fixing and processing for immunofluoresence as described above.
S. Typhimurium-induced Akt activationS. Typhimurium induced Akt activation after infection was monitored in soluble cell lysatesby probing with antibodies directed to phospho-specific Ser473Akt and Akt (Cell Signalling.Cells were lysed in 50mM Tris pH7.6, 150mM NaCl, 10% glycerol, 1% Triton X100, 1mMEDTA, 2mM MgCl2, 100mM NaF, 200μM NaVO4, 1mM PMSF, 1x protease inhibitorcocktail.
S. Typhimurium replication assayS. Typhimurium replication assays were preformed in Henle-407 cells and bone marrow-derived macrophages from caspase-1-deficient mice (which are resistant to to rapid S.Typhimurium-induced killing) as previously described (Hernandez et al., 2004).
SopB in vitro phosphatase assayGST tagged derivatives of SopB and SopB288-309 were expressed and purified by standardaffinity purification procedures using glutathione-coupled Sepharose 4B beads (Pharmacia).SopB protein phosphatase activity was measured using a malachite green chromogenic assay,as previously described (Lee et al., 2004).
AcknowledgementsWe thank Walther Mothes for providing reagents, Christopher Case and Catarina Nogueira for caspase-1 deficientbone marrow derived macrophages, and members of the Galán laboratory for critical review of this manuscript. K. H.was supported by a fellowship from the Damon Runyon Cancer Research Foundation. This work was supported byPublic Health Research Grant AI055472 to J. E. G.
ReferencesBache KG, Stuffers S, Malerod L, Slagsvold T, Raiborg C, Lechardeur D, Walchli S, Lukacs GL, Brech
A, Stenmark H. The ESCRT-III subunit hVps24 is required for degradation but not silencing of theepidermal growth factor receptor. Mol Biol Cell 2006;17:2513–2523. [PubMed: 16554368]
Chen H, Fre S, Slepnev V, Capua M, Takei K, Butler M, Di Fiore P, De Camilli P. Epsin is an EH-domain-binding protein implicated in clathrin-mediated endocytosis. Nature 1998;394:793–797.[PubMed: 9723620]
Chen LM, Hobbie S, Galan JE. Requirement of CDC42 for Salmonella-induced cytoskeletal and nuclearresponses. Science 1996;274:2115–2118. [PubMed: 8953049]
Collazo C, Galán JE. The invasion-associated type III system of Salmonella typhimurium directs thetranslocation of Sip proteins into the host cell. Mol Microbiol 1997;24:747–756. [PubMed: 9194702]
Patel et al. Page 10
Cell. Author manuscript; available in PMC 2010 April 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Damke H, Baba T, Warnock D, Schmid S. Induction of mutant dynamin specifically blocks endocyticcoated vesicle formation. J Cell Biol 1994;127:915–934. [PubMed: 7962076]
Dikic I. Mechanisms controlling EGF receptor endocytosis and degradation. Biochem Soc Trans2003;31:1178–1181. [PubMed: 14641021]
Drecktrah D, Knodler L, Galbraith K, Steele-Mortimer O. The Salmonella SPI1 effector SopB stimulatesnitric oxide production long after invasion. Cell Microbiol 2005:105–113. [PubMed: 15617527]
Fu Y, Galán JE. A salmonella protein antagonizes Rac-1 and Cdc42 to mediate host-cell recovery afterbacterial invasion. Nature 1999;401:293–297. [PubMed: 10499590]
Galan JE. SnapShot: effector proteins of type III secretion systems. Cell 2007;130:192. [PubMed:17632065]
Galán JE. Salmonella interaction with host cells: Type III Secretion at Work. Annu Rev Cell Dev Biol2001;17:53–86. [PubMed: 11687484]
Galán JE, Curtiss R III. Expression of Salmonella typhimurium genes required for invasion is regulatedby changes in DNA supercoiling. Infect Immun 1990;58:1879–1885. [PubMed: 2160435]
Galan JE, Wolf-Watz H. Protein delivery into eukaryotic cells by type III secretion machines. Nature2006;444:567–573. [PubMed: 17136086]
Galyov EE, Wood MW, Rosqvist R, Mullan PB, Watson PR, Hedges S, Wallis TS. A secreted effectorprotein of Salmonella dublin is translocated into eukaryotic cells and mediates inflammation andfluid secretion in infected ileal mucosa. Mol Microbiol 1997;25:1903–1912.
Guzman LM, Belin D, Carson MJ, Beckwith J. Tight regulation, modulation, and high-level expressionby vectors containing the arabinose PBAD promoter. J Bacteriol 1995;177:4121–4130. [PubMed:7608087]
Haglund K, Di Fiore P, Dikic I. Distinct monoubiquitin signals in receptor endocytosis. Trends BiochemSci 2003a;28:598–603. [PubMed: 14607090]
Haglund K, Sigismund S, Polo S, Szymkiewicz I, Di Fiore P, Dikic I. Multiple monoubiquitination ofRTKs is sufficient for their endocytosis and degradation. Nat Cell Biol 2003b;5:461–466. [PubMed:12717448]
Hardt W-D, Chen L-M, Schuebel KE, Bustelo XR, Galán JE. Salmonella typhimurium encodes anactivator of Rho GTPases that induces membrane ruffling and nuclear responses in host cells. Cell1998;93:815–826. [PubMed: 9630225]
Hernandez LD, Hueffer K, Wenk MR, Galán JE. A Salmonella protein modulates vesicular traffickingby altering phosphoinositide metabolism. Science 2004;18:1805–1807. [PubMed: 15205533]
Hoiseth SK, Stocker BA. Aromatic-dependent Salmonella typhimurium are non-virulent and effective aslive vaccines. Nature 1981;291:238–239. [PubMed: 7015147]
Hong KH, Miller VL. Identification of a novel Salmonella invasion locus homologous to Shigella ipgDE.J Bacteriol 1998;180:1793–1802. [PubMed: 9537377]
Huang F, Kirkpatrick D, Jiang X, Gygi S, Sorkin A. Differential regulation of EGF receptor internalizationand degradation by multiubiquitination within the kinase domain. Mol Cell 2006;21:737–748.[PubMed: 16543144]
Kaniga K, Bossio JC, Galán JE. The Salmonella typhimurium invasion genes invF and invG encodehomologues to the PulD and AraC family of proteins. Mol Microbiol 1994;13:555–568. [PubMed:7997169]
Kubori T, Galan JE. Temporal regulation of salmonella virulence effector function by proteasome-dependent protein degradation. Cell 2003;115:333–342. [PubMed: 14636560]
Lee SY, Wenk MR, Kim Y, Nairn AC, De Camilli P. Regulation of synaptojanin 1 by cyclin-dependentkinase 5 at synapses. Proc Natl Acad Sci U S A 2004;101:546–551. [PubMed: 14704270]
Malerød L, Stuffers S, Brech A, Stenmark H. Vps22/EAP30 in ESCRT-II mediates endosomal sortingof growth factor and chemokine receptors destined for lysosomal degradation. Traffic 2007;8:1617–1629. [PubMed: 17714434]
Mallo G, Espina M, Smith A, Terebiznik M, Alemán A, Finlay B, Rameh L, Grinstein S, Brumell J. SopBpromotes phosphatidylinositol 3-phosphate formation on Salmonella vacuoles by recruiting Rab5and Vps34. J Cell Biol 2008;182:741–752. [PubMed: 18725540]
Patel et al. Page 11
Cell. Author manuscript; available in PMC 2010 April 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Marcus S, Knodler L, Finlay B. Salmonella enterica serovar Typhimurium effector SigD/SopB ismembrane-associated and ubiquitinated inside host cells. Cell Microbiol 2002;4:435–446. [PubMed:12102689]
Miao EA, Scherer CA, Tsolis RM, Kingsley RA, Adams LG, Baumler AJ, Miller SI. Salmonellatyphimurium leucine-rich repeat proteins are targeted to the SPI1 and SPI2 type III secretion systems.Mol Microbiol 1999;34:850–864. [PubMed: 10564523]
Norris FA, Wilson MP, Wallis TS, Galyov EE, Majerus PW. SopB, a protein required for virulence ofSalmonella dublin, is an inositol phosphate phosphatase. Proc Natl Acad Sc U S A 1998;95:14057–14059. [PubMed: 9826652]
Patel J, Galán J. Investigating the function of Rho family GTPases during Salmonella/host cellinteractions. Methods Enzymol 2008;439:145–158. [PubMed: 18374162]
Patel JC, Galan JE. Differential activation and function of Rho GTPases during Salmonella-host cellinteractions. J Cell Biol 2006;175:453–463. [PubMed: 17074883]
Quezada C, Hicks S, Galán J, Stebbins E. A Salmonella Virulence Factor is a Novel E3 Ligase Mimic.2008submitted for publication
Raiborg C, Malerod L, Pedersen NM, Stenmark H. Differential functions of Hrs and ESCRT proteins inendocytic membrane trafficking. Exp Cell Res 2008;314:801–813. [PubMed: 18031739]
Rogers L, Kristensen A, Boyle E, Robinson D, Ly R, Finlay B, Foster L. Identification of cognate hosttargets and specific ubiquitylation sites on the Salmonella SPI-1 effector SopB/SigD. J Proteomics2008;71:97–108. [PubMed: 18541478]
Rohde J, Breitkreutz A, Chenal A, Sansonetti P, Parsot C. Type III Secretion Effectors of the IpaH FamilyAre E3 Ubiquitin Ligases. Cell Host Microbe 2007;1:77–83. [PubMed: 18005683]
Saksena S, Sun J, Chu T, Emr S. ESCRTing proteins in the endocytic pathway. Trends Biochem Sci2007;32:561–573. [PubMed: 17988873]
Stebbins CE, Galán JE. Structural mimicry in bacterial virulence. Nature 2001;412:701–705. [PubMed:11507631]
Steele-Mortimer O, Knodler L, Marcus S, Scheid M, Goh B, Pfeifer C, Duronio V, Finlay B. Activationof Akt/protein kinase B in epithelial cells by the Salmonella typhimurium effector sigD. J Biol Chem2000;275:37718–37724. [PubMed: 10978351]
Urbe S. Ubiquitin and endocytic protein sorting. Essays Biochem 2005;41:81–98. [PubMed: 16250899]Waterman SR, Holden DW. Functions and effectors of the Salmonella pathogenicity island 2 type III
secretion system. Cell Microbiol 2003;5:501–511. [PubMed: 12864810]Zhang Y, Higashide W, McCormick B, Chen J, Zhou D. The inflammation-associated Salmonella SopA
is a HECT-like E3 ubiquitin ligase. Mol Microbiol 2006;62:786–793. [PubMed: 17076670]Zhou D, Chen LM, Hernandez L, Shears SB, Galán JE. A Salmonella inositol polyphosphatase acts in
conjunction with other bacterial effectors to promote host cell actin cytoskeleton rearrangements andbacterial internalization. Mol Microbiol 2001;39:248–259. [PubMed: 11136447]
Patel et al. Page 12
Cell. Author manuscript; available in PMC 2010 April 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 1. SopB is ubiquitinated upon bacterial delivery and persists during infectionA. SopB persists during infection. Henle-407 cells were infected with a S. Typhimurium strainexpressing FLAG epitope-tagged SopB for 1 hr and its presence in the protein translocatedfraction was examined by western immunoblot. B. Effect of bacterial protein synthesisinhibitors on the levels of translocated SopB at different times after infection. Henle-407 cellswere infected with a S. Typhimurium strain expressing FLAG epitope-tagged SopB and at theindicated times after infection, chloramphenicol was added. Infected cells were lysed 6 hoursafter infection and the presence of SopB in the translocated fraction was examined by westernimmunoblot. C. SopB is ubiquitinated upon translocation into epithelial cells. COS-2 cellstransfected with a plasmid encoding HA epitope tagged ubiquitin, were infected with a S.Typhimurium strain expressing FLAG-epitope-tagged SopB for 1 hr. The presence ofubiquitinated SopB in infected cells was analyzed by immunoprecipitation and western blotanalysis. D. SopB ubiquitination does not require the bacterially-encoded E3 ubiquitin ligases.Henle-407 cells were infected for 1 hr with S. Typhimurium strains expressing FLAG epitope-tagged SopB and lacking the bacterially-encoded E3 ubiquitin ligases SlrP, SspH2, or SopA,as indicated. The presence of SopB in the protein translocated fraction was examined bywestern immunoblot. E. SopB is conjugated to lysineless and K63 ubiquitin. Cultured cellstransfected with a plasmid encoding HA-tagged lysineless (K0) or K63 ubiquitin were infectedwith a S. Typhimurium strain expressing FLAG-epitope-tagged SopB. The presence ofubiquitinated SopB in infected cells was analyzed by immunoprecipitation and western blotanalysis. In all panels the star denotes the predicted mobility of unmodified SopB.
Patel et al. Page 13
Cell. Author manuscript; available in PMC 2010 April 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 2. Identification of the SopB ubiquitination sitesA. Diagram indicating the location of lysine residues within the amino terminus of SopB. B.Effect of mutations in SopB lysine residues on its secretion, translocation, and ubiquitination.Bacterial strains were examined for their ability to express and secrete the different mutantconstructs (left panels). In addition, mutants were examined for their ubiquitination upontranslocation into cultured cells (right panels). Numbers correspond to the lysine residuesindicated in the diagram shown in A. The star denotes the predicted mobility of unmodifiedSopB.
Patel et al. Page 14
Cell. Author manuscript; available in PMC 2010 April 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 3. Subcellular localization of wild-type SopB and its ubiquitination-deficient mutantA and B. Henle-407 cells were infected with a S. Typhimurium strain expressing wild typeepitope-tagged SopB (A) or its ubiquination-deficient SopBΔub mutant (B) and examined byfluorescence confocal microscopy after staining with antibodies directed to the epitope tag(green) or to S. Typhumrium (red) Bar = 10 μM. C. Levels of SopB or SopBΔub in infectedcells. Henle-407 cells were infected with S. Typhimurium strains expressing FLAG epitope-tagged wild type SopB or the SopBΔub mutant. At the indicated times after infection, cells wereprobed for the presence of SopB in the translocated fraction by western immunoblot. Blotswere reprobed with antibodies directed to SipB (another SPI-1 TTSS effector protein) or actinto control for loading. The star denotes the predicted mobility of unmodified SopB. D.Localization of wild-type SopB or its ubiquitination deficient mutant derivative after theirdelivery by a non-invasive S. Typhimurium mutant. Cultured epithelial cells pre-loaded withAlexa 594-dextran to label endosomal compartments were infected for 1 hr with the invasion-defective ΔsopE ΔsopE2 ΔsopB S. Typhimurium mutant strain expressing FLAG epitope-tagged catalytic-inactive SopBC460S or its ubiquitination defective derivative SopBC460S Δub.Cells were fixed, stained for SopB (green) and DAPI (blue) and examined by fluorescencemicroscopy. Insets show enlarged detail area. Bar = 10 μM. E. Levels of SopBC460S or itsderivative SopBC460S Δub after their TTS-mediated delivery by the non-invasive S.Typhimurium mutant strain. Henle-407 cells were infected with ΔsopE ΔsopE2 ΔsopB S.Typhimurium mutant strains expressing FLAG epitope-tagged wild type SopB or its notubiquitinitable mutant (SopBΔub) and 1 hr after infection, cells were probed for the presence
Patel et al. Page 15
Cell. Author manuscript; available in PMC 2010 April 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

of SopB in the translocated fraction by western immunoblot. The star denotes the predictedmobility of unmodified SopB.
Patel et al. Page 16
Cell. Author manuscript; available in PMC 2010 April 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
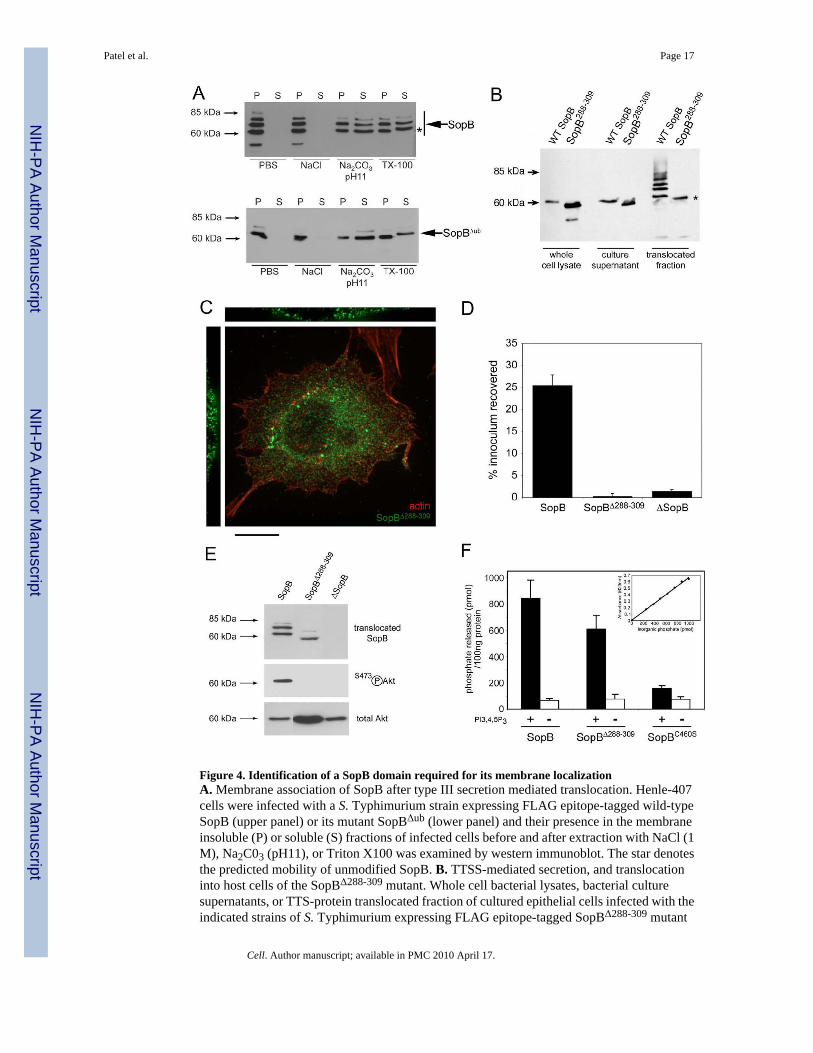
Figure 4. Identification of a SopB domain required for its membrane localizationA. Membrane association of SopB after type III secretion mediated translocation. Henle-407cells were infected with a S. Typhimurium strain expressing FLAG epitope-tagged wild-typeSopB (upper panel) or its mutant SopBΔub (lower panel) and their presence in the membraneinsoluble (P) or soluble (S) fractions of infected cells before and after extraction with NaCl (1M), Na2C03 (pH11), or Triton X100 was examined by western immunoblot. The star denotesthe predicted mobility of unmodified SopB. B. TTSS-mediated secretion, and translocationinto host cells of the SopBΔ288-309 mutant. Whole cell bacterial lysates, bacterial culturesupernatants, or TTS-protein translocated fraction of cultured epithelial cells infected with theindicated strains of S. Typhimurium expressing FLAG epitope-tagged SopBΔ288-309 mutant
Patel et al. Page 17
Cell. Author manuscript; available in PMC 2010 April 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

or wild type SopB (as control) were analyzed by western immunoblot. The star denotes thepredicted mobility of unmodified SopB. C. Subcellular localization of the SopBΔ288-309
mutant. Henle-407 cells, were infected with a S. Typhimurium strain expressing an epitope-tagged SopBΔ288-309 mutant and examined by immunofluorescence confocal microscopy withan antibody directed to the epitope tag (green) or actin (red). Bar = 10 μM. D. Ability of wildtype SopB and the SopB288-309 mutant to mediate bacterial entry into cultured epithelial cells.Henle-407 cells were infected with a S. Typhimurium ΔsopE ΔsopE2 ΔsopB mutant strain (ascontrol indicated “ΔsopE” in the panel) or a S. Typhimurium ΔsopE ΔsopE2 mutant expressingwild type SopB, or SopBΔ288-309 (as indicated), and the levels of internalized bacteria weredetermined as indicated in the Material and Methods. Values are the mean ± standard deviationof three independent experiments and represent the percentage of the original bacterialinoculum that survived the antibiotic treatment due to internalization. E. Ability of wild typeSopB and the SopB288-309 mutant to activate Akt. Henle-407 cells were infected with a S.Typhimurium expressing FLAG epitope tagged wild-type SopB, or the SopBΔ288-309 mutant,and the levels of phosphorylated (activated) or total Akt 1 hr after infection were examined bywestern immunoblot analysis. The levels of translocated epitope-tagged SopB, orSopBΔ288-309 were determined by western immunoblot analysis of the fraction of infected cellscontaining the translocated protein (upper panel). F. Comparison of the in-vitrophosphoinositide phosphatase activity of purified wild type SopB, the SopBΔ288-309 mutant,and the phosphatase-inactive SopBC460S mutant. The phosphatase activity was calculatedbased on the standard curve (shown in inset) for inorganic phosphate standards using amalachite green chromogenic assay.
Patel et al. Page 18
Cell. Author manuscript; available in PMC 2010 April 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 5. The ubiquitination-deficient SopB mutant induces efficient entry and distinct actinreorganizationA. Henle-407 cells were infected with a S. Typhimurium ΔsopE ΔsopE2 mutant strainexpressing wild type SopB, or the SopBΔub mutant (as indicated), and the levels of internalizedbacteria were determined by the gentamicin resistance assay. Values are the mean ± standarddeviation of three independent experiments and represent the percentage of the originalbacterial inoculum that survived the antibiotic treatment due to internalization. B and C. Ref-52cells were infected with a ΔsopE ΔsopE2 S. Typhimurium strain expressing either wild-typeSopB or its ubiquitination deficient mutant derivative (SopBΔub). Cells were stained withrhodamine-phalloidin to visualize the actin cytoskeleton, with a FITC-labeled antibody to
Patel et al. Page 19
Cell. Author manuscript; available in PMC 2010 April 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

visualize S. Typhimurium, and examined by fluorescence microscopy. Cells exhibiting eitherlocalized or peripheral actin cytoskeletal changes were quantified. Values (shown in B)represent the percentage of infected cells showing the indicated changes and are the mean ±standard deviation of 3 experiments in which a minimum of 200 cells were examined for eachcategory. The values obtained in cells infected with the SopBΔub mutant were statisticallysignificant (p < 0.02) when compared to those of wild type with the student t test.
Patel et al. Page 20
Cell. Author manuscript; available in PMC 2010 April 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 6. The ubiquitination-deficient SopB mutant induces prolonged macropinocytosis and AktactivationA and B. Henle-407 cells were infected with a S. Typhimurium strain expressing either wild-type SopB or its ubiquitination deficient mutant derivative (SopBΔub) and visualized by livephase contrast microscopy (A). Cells exhibiting macropinosomes were quantified at theindicated times (B). Values represent the percentage of infected cells showing macropinosomesand are the mean ± standard deviation of 3 experiments in which a minimum of 200 cells wereexamined for each category and time point. * indicates statistically significant (p < 0.05)differences when compared to those of wild type with the student t test. C. Henle-407 cellswere infected with a S. Typhimurium expressing either wild-type SopB or its ubiquitination
Patel et al. Page 21
Cell. Author manuscript; available in PMC 2010 April 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

deficient mutant derivative (SopBΔub) and the levels of phosphorylated (activated) Akt andtotal Akt at different times (as indicated) after infection were examined by western immunoblotanalysis.
Patel et al. Page 22
Cell. Author manuscript; available in PMC 2010 April 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 7. The ubiquitination-deficient SopB mutant is defective for Rab5 recruitment andintracellular growthA. Rab5 recruitment by different strains of S. Typhimurium. Henle-407 cells that had beentransfected with a plasmid expressing Rab5-GFP were infected with a ΔsopB S. Typhimuriummutant strain (ΔsopB) or a strain expressing either wild-type SopB (WT SopB) or itsubiquitination deficient mutant derivative (SopBΔub). At the indicated times the percentagesof the different S. Typhimurium strains showing co-localization with Rab5-GFP weredetermined. Values are mean ± standard deviation of three independent determinations inwhich the indicated number of bacteria (n) were examined. * indicates statistically significant(p < 0.05) differences when compared to the values of cells infected with wild type bacteriausing the student t test. B. S. Typhimurium expressing SopBΔub is defective for intracellulargrowth in cultured epithelial cells. Henle-407 cells were infected with a ΔsopE ΔsopE2 S.Typhimurium strain expressing either wild type SopB or expressing SopBΔub. At the indicatedtimes after infection, the levels of intracellular c. f. u. of the different strains were determined.Values represent the mean ± standard deviation of three independent experiments. * indicatesstatistically significant (p < 0.05) differences when compared to the values of cells infectedwith wild type bacteria using the student t test. C. S. Typhimurium expressing sopBΔub isdefective for survival within primary macrophages. Primary bone marrow-derivedmacrophages were infected with wild type S. Typhimurium (WT), a ΔsopB mutant (ΔsopB), astrain expressing expressing sopBΔub (sopBΔub) or a derivative of the same straincomplemented with a plasmid expressing wild type sopB (sopBΔub + WT sopB). At theindicated times after infection, the levels of intracellular c. f. u. of the different strains weredetermined. Values are normalized to those of wild type and represent the mean ± standarddeviation of three independent experiments. * indicates statistically significant (p < 0.05)differences when compared to the values of cells infected with wild type bacteria using thestudent t test. D. Model for ubiquitin-mediated diversification of SopB function. Dx Upondelivery by the S. Typhimurium TTSS, SopB localizes to the plasma membrane through ismembrane localization domain (I). While at the plasma membrane SopB mediates entry andmacropinocytosis by activating SGEF, a RhoG exchange factor, and Akt activation byunknown mechanisms (I). Membrane-localized SopB is ubiquitinated (mono ubiquitinated orlinked via K63), which results in its trapping in the incoming phagosome or its removal fromthe plasma membrane and its subsequent translocation to the Salmonella- containing vacuole,where it modulates vesicle trafficking by altering phosphoinositide metabolism at that site thusavoiding the delivery of S. Typhimurium to lysosomes (II). See text for further details.
Patel et al. Page 23
Cell. Author manuscript; available in PMC 2010 April 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Copyright © 2022 FDOKUMEN




















