
The Potassium Transporter Trk and External Potassium Modulate Salmonella enterica Protein Secretion...
10
Published Ahead of Print 10 November 2008. 2009, 77(2):667. DOI: 10.1128/IAI.01027-08. Infect. Immun. Jing Su, Hao Gong, Jeff Lai, Andrew Main and Sangwei Lu Protein Secretion and Virulence enterica Salmonella External Potassium Modulate The Potassium Transporter Trk and http://iai.asm.org/content/77/2/667 Updated information and services can be found at: These include: REFERENCES http://iai.asm.org/content/77/2/667#ref-list-1 at: This article cites 44 articles, 26 of which can be accessed free CONTENT ALERTS more» articles cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on July 17, 2014 by guest http://iai.asm.org/ Downloaded from on July 17, 2014 by guest http://iai.asm.org/ Downloaded from
Transcript of The Potassium Transporter Trk and External Potassium Modulate Salmonella enterica Protein Secretion...

Published Ahead of Print 10 November 2008. 2009, 77(2):667. DOI: 10.1128/IAI.01027-08. Infect. Immun.
Jing Su, Hao Gong, Jeff Lai, Andrew Main and Sangwei Lu
Protein Secretion and Virulence entericaSalmonellaExternal Potassium Modulate
The Potassium Transporter Trk and
http://iai.asm.org/content/77/2/667Updated information and services can be found at:
These include:
REFERENCEShttp://iai.asm.org/content/77/2/667#ref-list-1at:
This article cites 44 articles, 26 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on July 17, 2014 by guesthttp://iai.asm
.org/D
ownloaded from
on July 17, 2014 by guest
http://iai.asm.org/
Dow
nloaded from

INFECTION AND IMMUNITY, Feb. 2009, p. 667–675 Vol. 77, No. 20019-9567/09/$08.00�0 doi:10.1128/IAI.01027-08Copyright © 2009, American Society for Microbiology. All Rights Reserved.
The Potassium Transporter Trk and External Potassium ModulateSalmonella enterica Protein Secretion and Virulence�
Jing Su,1,2 Hao Gong,1 Jeff Lai,1 Andrew Main,1 and Sangwei Lu1*Program in Infectious Diseases and Immunity, School of Public Health, University of California, Berkeley, California 94720,1 andDepartment of Bioscience and Technology, School of Life Science, Nanjing University, Nanjing, Jiangsu, People’s Republic of China2
Received 18 August 2008/Returned for modification 17 September 2008/Accepted 31 October 2008
Potassium (K�) is the most abundant intracellular cation and is essential for many physiological functionsof all living organisms; however, its role in the pathogenesis of human pathogens is not well understood. In thisstudy, we characterized the functions of the bacterial Trk K� transport system and external K� in thepathogenesis of Salmonella enterica, a major food-borne bacterial pathogen. Here we report that Trk isimportant for Salmonella to invade and grow inside epithelial cells. It is also necessary for the full virulence ofSalmonella in an animal infection model. Analysis of proteins of Salmonella indicated that Trk is involved in theexpression and secretion of effector proteins of the type III secretion system (TTSS) encoded by Salmonellapathogenicity island 1 (SPI1) that were previously shown to be necessary for Salmonella invasion. In additionto the role of the Trk transporter in the pathogenesis of Salmonella, we discovered that external K� modulatesthe pathogenic properties of Salmonella by increasing the expression and secretion of effector proteins of theSPI1-encoded TTSS and by enhancing epithelial cell invasion. Our studies demonstrated that K� is activelyinvolved in the pathogenesis of Salmonella and indicated that Salmonella may take advantage of the high K�
content inside host cells and in the intestinal fluid during diarrhea to become more virulent.
Potassium (K�) is the most abundant intracellular cation inall living organisms, including bacteria (22). Previous studieson K� have focused mostly on its role in the physiology ofeukaryotic and prokaryotic organisms, while its role, if any, inthe pathogenesis of human pathogens has not been well char-acterized. Human pathogens often need to survive and grow inconditions with a wide range of K� concentrations, from en-vironmental niches such as rivers, soil, and sewage with low K�
concentrations to inside host cells with high K� concentrations(100 to 160 mM) (22). Therefore, it is important to understandhow pathogens adapt to various K� concentrations and howK� affects pathogens.
Bacteria maintain a relatively constant intracellular K� con-centration (300 to 500 mM) for many essential cellular func-tions, including maintenance of cell turgor and homeostasis,adaptation of cells to osmotic conditions, and activation ofcytoplasmic enzymes (8, 15). Since bacteria are exposed to awide range of external K� concentrations, they use a numberof transporters and efflux pumps to maintain their intracellularK� concentrations. The best-characterized K� transport sys-tems include the Trk, Kdp, and Kup K� transport systems ingram-negative bacteria. The Trk system is a low-affinity, rapid-transport system that is the main K� transporter at neutral oralkaline pH (7, 15, 43). It is a multiunit protein complexformed by gene products that are constitutively expressed. TheKdp system is a high-affinity K� transport system that is in-duced in low-K� environments (K� concentration, 5 mM orless) (15, 17). The Kup system has an affinity for K� similar tothat of the Trk system and is believed to be the major K�
transport system under acidic conditions (44, 48). Previousstudies showed that Salmonella trkA (sapG), which encodes anessential NAD� binding subunit of the Trk system, was nec-essary for resistance to antimicrobial peptides (37). TrkA ofVibrio vulnificus has been reported to be required for serum,protamine, and polymyxin B resistance (9). These results sug-gest that intracellular K� is important for the virulence char-acteristics of bacterial pathogens.
Salmonella enterica is a gram-negative bacterium and amajor human pathogen that causes significant mortality andmorbidity worldwide. In this study, we analyzed the role of theTrk K� transport system in the pathogenesis of Salmonella andinvestigated the influence of external K� on the virulencecharacteristics of Salmonella. Our results demonstrate thatboth intracellular K� and external K� modulate the virulencecharacteristics of Salmonella.
MATERIALS AND METHODS
Reagents. Growth media for bacteria and HeLa cells were purchased from BDDiagnostics (Sparks, MD) and Invitrogen (Carlsbad, CA), respectively. Chemi-cals and antibiotics were obtained from Sigma-Aldrich (St. Louis, MO) unlessotherwise indicated. Restriction and modifying enzymes for manipulation ofDNA were obtained from New England Biolabs (Ipswich, MA). Custom oligo-nucleotides were purchased from Sigma Genosys (The Woodlands, TX).
Bacterial strains and culture conditions. S. enterica serovar Enteritidis isolateSE2472 (a clinical isolate) was used as the wild-type parental strain in all exper-iments (Table 1) (30–32). Escherichia coli DH5� was used for construction ofrecombinant plasmid DNA (Table 1).
All bacteria were routinely cultured in Luria-Bertani (LB) broth at 37°C withshaking at 225 rpm (3). A minimal K� medium consisting of 6.78 g of Na2HPO4/liter, 2.9587 g of NaH2PO4 � H2O/liter, 10 g of NaCl/liter, 1 g of NH4Cl/liter, 4 gof glucose/liter, 2 mM MgSO4, and 0.2 mM CaCl2 (pH 7) was used to examinethe role of K� in bacterial growth. Although no K� was added to the medium,the medium contained approximately 0.2 mM K� from K� contaminants in thesalts used, as measured with a K�-selective electrode (Denver Instruments,Denver, CO). Antibiotics were added when necessary.
Growth curves and stress resistance assays. The growth and survival of bac-teria in various media and with various chemicals were determined as previously
* Corresponding author. Mailing address: 16 Barker Hall, Univer-sity of California, Berkeley, CA 94720-7354. Phone: (510) 643-4986.Fax: (510) 643-9955. E-mail: [email protected].
� Published ahead of print on 10 November 2008.
667
on July 17, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

described (32). The following media and conditions were used to determine thegrowth and stress resistance of bacteria: Dulbecco’s modified Eagle’s medium(DMEM) supplemented with 10% fetal bovine serum, mouse serum, LB broth,LB broth containing 5% sodium dodecyl sulfate (SDS) or 1 M NaCl, and LBbroth at pH 4.0. Survival of bacteria at 50°C was determined in LB brothincubated at 50°C without shaking. Potassium chloride was added as a K�
supplement when appropriate. In each assay, bacterial concentrations were de-termined by plating appropriately diluted aliquots onto LB agar plates.
Invasion of HeLa cells by Salmonella and intracellular growth of Salmonella inHeLa cells. A HeLa cell invasion assay was performed as described previously(32). Briefly, all Salmonella strains were cultured in LB broth overnight at 37°Cwithout shaking. Antibiotics were added when appropriate. An overnight cultureof bacteria was added to HeLa cells at multiplicities of infection of 5:10 to 10:1,and intracellular bacteria were quantified after 1 or 2 h of incubation for inva-sion, which was followed by incubation in the presence of 50 �g/ml of gentamicinto kill extracellular bacteria. The invasiveness of Salmonella was measured bydetermining the ratio of intracellular bacteria, which was calculated as follows:(number of intracellular bacteria/number of input bacteria) � 100.
The intracellular growth of Salmonella in HeLa cells was determined by in-fecting HeLa cells with Salmonella for 1 h. The HeLa cells were then washed withphosphate-buffered saline to remove extracellular bacteria and incubated infresh medium containing 50 �g/ml of gentamicin for 1 h to kill extracellularbacteria. A set of cells was washed and lysed to quantify the intracellular bacteriaand used as the zero-time sample. Additional sets of cells were harvested after anadditional 4 or 8 h of incubation to quantify intracellular Salmonella (4- and 8-hsamples). The growth of Salmonella inside HeLa cells was measured by deter-mining the increase in the number of intracellular Salmonella cells, which wascalculated by dividing the quantity of intracellular bacteria in a 4- or 8-h sampleby the quantity of intracellular bacteria in the zero-time sample.
Epitope tagging of Salmonella proteins and analysis of tagged proteins. Pro-tein tagging was carried out as described by Uzzau et al. (45), except that bothsix-His and FLAG tags were used. First, plasmid pUC-H1PF1 was constructed asa template. This plasmid was designed to contain a kanamycin resistance cassette(Kanr) and the DNA sequence encoding the six-His and FLAG small epitopetags (45). Kanr in plasmid pKD4 (Table 1) was amplified by PCR using primersH1PF1(fwd) and H1PF1(rev) (Table 2). Primer H1PF1(fwd) contained a BamHIlinker, the sequence encoding the six-His and FLAG epitopes, and the sequencecorresponding to nucleotides 31 to 49 of plasmid pKD4. Primer H1PF1(rev)contained an EcoRI linker and the sequence corresponding to nucleotides 1485to 1507 of plasmid pKD4. The PCR product was then cloned into plasmidpUC19 (New England Biolabs, Ipswich, MA) between the BamHI and EcoRIsites. The sequence of the resulting plasmid, pUC-H1PF1, was verified, and this
plasmid was used as template in PCRs with SipA-, SipC-, or SopB-specificprimers listed in Table 2. Each PCR produced a DNA module consisting of the3� end of the target gene, FLAG and six-His tag nucleotides, the stop codon,Kanr, and the target gene sequence immediately following the stop codon. TheDNA module was introduced into the Salmonella genome, the sequence wasverified, and the module was transduced into fresh SE2472 (12). Kanr wassubsequently removed using plasmid pCP20 (12), and the sequence of the re-sulting strain was verified. When necessary, mutations were transduced intostrains with epitope tags by general transduction with phage P22, and phage-freecolonies were used for analysis of expression (35).
The levels of the tagged proteins in bacterial lysates and culture supernatantwere determined by Western blot analysis. Bacterial lysates were prepared bycentrifuging bacterial cultures, resuspending the bacterial pellets in sodium do-decyl sulfate (SDS) sample buffer (50 mM Tris-HCl [pH 6.8], 100 mM dithio-threitol, 2% SDS, 10% glycerol, 0.1% bromophenol blue), and boiling the prep-arations for 10 min (42). Secreted proteins were prepared as described byKomoriya et al. (23). For Western blot analysis, a monoclonal anti-FLAG anti-body (Sigma-Aldrich, St. Louis, MO) was used at a 1:1,000 dilution as theprimary antibody, and horseradish peroxidase-conjugated sheep anti-mouse im-munoglobulin G (GE Healthcare, Piscataway, NJ) was used at a 1:2,000 dilutionas the secondary antibody. The signal was visualized by the enhanced chemilu-minescence method (GE Healthcare, Piscataway, NJ). Coomassie blue-stainedgels were used for comparison of protein bands.
Mutagenesis of trkA and complementation of the mutant. A trkA deletion(�trkA) mutant of SE2472 was constructed using the one-step mutagenesismethod of Datsenko and Wanner, as described previously (12, 30). PrimersTrkA5KO and TrkA3KO were used to replace the trkA sequence betweennucleotides 91 and 1270 with Kanr (Table 2). The deletion mutation was subse-quently transduced into fresh SE2472 by general transduction using phage P22(35), and phage-free colonies were used for further analysis. A nonpolar trkAmutant was also generated as described by Datsenko and Wanner (12).
To complement the �trkA mutant of SE2472, we cloned the wild-type allele oftrkA into plasmid pRB3-273C (6). Primers TrkA5H3 and TrkA3H3 were used toamplify trkA from SE2472 (Table 2), and the PCR product was digested withHindIII and cloned into pRB3-273C at the HindIII site. The sequence of theresulting plasmid, pRB3-trkA, was confirmed, and the plasmid was transformedinto the �trkA mutant of SE2472 to complement the deletion mutation (30).Vector pRB3-273C was also transformed into the �trkA mutant of SE2472 as acontrol for any possible effect of the plasmid transformation alone (30).
Infection of mice with Salmonella. Salmonella cultured overnight in LB brothat 37°C with shaking at 225 rpm was used in all animal infection experiments.Mice were infected intragastrically or intraperitoneally, and the 50% lethal dose
TABLE 1. Bacterial strains and plasmids
Strain or plasmid Characteristics Source or reference
E. coli DH5� F� �80dlacZ�M15 �(lacZYA-argF)U169 deoR recA1 endA1 hsdR17(rk� mk
�)phoA supE44 � thi-1 gyrA96 relA1
Invitrogen
S. enterica serovar Enteritidis strainsSE2472 Clinical isolate 32�trkA SE2472 trkA::kan This study�trkA-comp SE2472 trkA::kan transformed with pRB3-trkA This studySipA(HF) SE2472 sipA::His-FLAG, SipA� This studySipC(HF) SE2472 sipC::His-FLAG, SipC� This studySopB(HF) SE2472 sopB::His-FLAG, SopB� This studySipA(HF)�trkA SipA(HF) trkA::kan This studySipC(HF)�trkA SipC(HF) trkA::kan This studySopB(HF)�trkA SopB(HF) trkA::kan This studySipA(HF)�trkA-comp SipA(HF)�trkA transformed with pRB3-trkA This studySipC(HF)�trkA-comp SipC(HF)�trkA transformed with pRB3-trkA This studySopB(HF)�trkA-comp SopB(HF)�trkA transformed with pRB3-trkA This study
PlasmidspUC-H1PF1 Apr, derivative of pUC19 containing Kanr, FLAG, and His6 This studypKD4 Apr Kanr oriR 12pKD46 Apr, contains the Red recombinase of phage 12pCP20 Apr cat cl857 Pr flp pSC101 oriTS 12pRB3-273C Apr, low- to medium-copy-number plasmid for Salmonella 12pRB3-trkA Derivative of pRB3-273C containing trkA This study
668 SU ET AL. INFECT. IMMUN.
on July 17, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

(LD50) was determined by infecting groups of five mice with 10-fold dilutions ofbacteria in phosphate-buffered saline (32, 41).
RESULTS
Construction of a Salmonella �trkA mutant and analysis ofits growth properties. To determine the roles of K� transportsystems in the pathogenesis of Salmonella, we constructed adeletion mutant with a mutation in the low-affinity Trk K�
transport system of S. enterica serovar Enteritidis SE2472(�trkA) (32). In the �trkA mutant of SE2472, the trkA openreading frame, which encodes a subunit essential for the Trkfunction, was replaced by Kanr (12, 30). The �trkA mutantformed smaller colonies on LB agar but otherwise had thesame colony morphology as wild-type strain SE2472 (data notshown).
To determine if TrkA is necessary for the growth of bacteria,we constructed growth curves for the �trkA mutant and wild-type strain SE2472 grown in LB broth, minimal K� medium,and minimal K� medium supplemented with various concen-trations of potassium chloride (KCl) as the source of K�.Using a K�-selective electrode (Denver Instruments, Denver,CO), the concentration of K� in LB broth was determined tobe approximately 8 mM (data not shown), a concentration atwhich Trk is expected to be the main K� transporter. MinimalK� medium was prepared without any K� salt; however, itcontained approximately 0.2 mM K� due to K� contaminationin the salts used for the medium (data not shown). In LB broth,minimal K� medium, or minimal K� medium supplementedwith less than 10 mM K�, the growth of the �trkA mutant wassimilar to the growth of wild-type strain SE2472 (data notshown). When more than 10 mM supplemental K� was used,the �trkA mutant displayed a small but consistent growth delaycompared to the wild-type strain between 4 to 8 h of growth(Fig. 1). The �trkA mutant appeared to have the same lagphase as wild-type strain SE2472, but it had a lower growthrate in the log phase of growth (Fig. 1). The growth delay wasobserved at all K� concentrations that we tested (10 to 100
mM), and the delay was rescued by transforming the �trkAmutant with plasmid pRB3-trkA, which contained a wild-typeallele of trkA in plasmid vector pRB3-273C (Fig. 1) (6). Trans-formation with the pRB3-273C vector alone had no effect(data not shown). These results indicate that TrkA is involvedin optimal growth of Salmonella at relatively high concentra-tions of K�.
The resistance of the �trkA mutant to general stress condi-tions was tested to determine the overall fitness of this mutant.The �trkA mutant was exposed to salt (1 M NaCl), detergent(5% SDS), heat (incubation at 50°C), and acidic medium (pH4). No difference in bacterial growth or survival was observedbetween the �trkA mutant and wild-type strain SE2472 (datanow shown). Since the TrkA protein of V. vulnificus was re-ported to be necessary for serum resistance (9), we tested the
TABLE 2. Oligonucleotide primers
Primer Sequence
H1PF1(fwd) ...................................................5�-CGCGGATCCGATTACAAGGATGACGACGATAAGGAGAATTTGTATTTTCAGGGACACCACCACCACCACCACTGATGTGTAGGCTGGAGCTGCTT-3�
H1PF1(rev) ....................................................5�-CCGGAATTCCATATGAATATCCTCCTTAGTTC-3�SipA(fwd) .......................................................5�-GCCCGGCTTACGAGTCATACCTGAATAAACCTGGCGTGGATCGGGTTATTACTACCGT
TGATGGCTTGCACATGCAGCGTGATTACAAGGATGACGACGA-3�SipA(rev) ........................................................5�-ACATCAACGGCAATACAGGAGGTGATCACTTTTTTGACTCTTGCTTCAATATCCATATT
CATCGCATCTTTCCCGGTTAACATATGAATATCCTCCTTAGTT-3�SipC(fwd) .......................................................5�-AGGAAATGCTGAAAACAATGGAGAGCATTAACCAGTCGAAAGCATCCGCACTTGCTG
CTATCGCAGGCAATATTCGCGCTGATTACAAGGATGACGACGA-3�SipC(rev) ........................................................5�-TATTAAGCATAATATCCCCAGTTCGCCATCAGGAGCGCGATTAAATCACACCCATGAT
GGCGTATAGATGACCTTTCAGACATATGAATATCCTCCTTAGTT-3�SopB(fwd) ......................................................5�-TCAATCTTTCCTATCAAAAACGAGTTGGGGATGAAAATATTTGGCAGTCAGTAAAAG
GCATTTCTTCATTAATCACATCTGATTACAAGGATGACGACGA-3�SopB(rev) .......................................................5�-GTGGCTCATCTTCTGGCGCATCCAGGCCTAACGCGTCATATAAACGATTTAATAGACT
TTCCATATAGTTACCTCAAGACCATATGAATATCCTCCTTAGTT-3�TrkA5KO .......................................................5�-TCATACTGGG CGCAGGGCAA GTTGGCGGAA CACTGGCGGA AAACCTGGTC GGCG
AAAACAACGACATCAC GGTGGTTGAT CATATGAATATCCTCCTTAG-3�TrkA3KO .......................................................5�-CAGACGTTCGACGTCGGTAATAAACTTTTTATCGGTCAAAAACATAATGACGTGATCG
CCTTGTTCAATGCGCAAATTGT GTGTAGGCTG GAGCTGCTTC-3�TrkA5H3 ........................................................5�-AGACCAGGAAGCTTGATGAAAATTATCATACTGGG-3�TrkA3H3 ........................................................5�-ATAGCAACAAGCTTTTAGAGGAAGAAAGGACTTG-3�
FIG. 1. Growth of the �trkA mutant of Salmonella with a highconcentration of potassium. Wild-type strain SE2472, the �trkA mu-tant, and the complemented mutant (�trkA-comp) were cultured inminimal K� medium supplemented with 50 mM KCl. Bacterial con-centrations were determined by plating. At least three experimentswere performed, and the results of a representative experiment per-formed in triplicate are shown. The error bars indicate standard devi-ations.
VOL. 77, 2009 ROLE OF POTASSIUM IN SALMONELLA PATHOGENESIS 669
on July 17, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

survival and growth of the �trkA mutant of SE2472 in mouseserum, and we detected no defect in the �trkA mutant (datanot shown).
TrkA is necessary for the invasion of epithelial cells bySalmonella. Since epithelial cell invasion is crucial in naturalinfection by Salmonella, we examined whether TrkA affects theability of Salmonella to invade epithelial cells. The invasion ofHeLa cells by the �trkA mutant and wild-type strain SE2472was assayed, and the ratio of the number of intracellular bac-teria to the number of input bacteria was determined after 1 or2 h of incubation. The ratio obtained for wild-type strainSE2472 intracellular bacteria at 1 h postinfection was arbi-trarily defined as 100%, and ratios of intracellular bacteria forother samples were expressed as relative values (Fig. 2A). Atboth 1 and 2 h postinfection, the ratios for the intracellular�trkA mutant were approximately 35 to 40% of those forwild-type strain SE2472 (Fig. 2A). The invasion defect of the�trkA mutant was rescued by transformation with plasmidpRB3-trkA (Fig. 2A), while transformation with the vectoralone had no effect (data not shown). The invasion defect ofthe �trkA mutant was not due to a growth defect in the me-dium used for the assay (DMEM with 10% fetal bovine se-rum), as the �trkA mutant grew as well as wild-type strainSE2472 in this medium (data not shown). DMEM containsapproximately 5 mM K� as measured with a K�-selectiveelectrode (data not shown); therefore, the normal growth ofthe �trkA mutant in DMEM is consistent with the previousobservation that the �trkA mutant had no growth defect whenthe K� concentration was less than 10 mM.
The �trkA mutant of Salmonella is defective for growth in-side HeLa cells. In addition to determining the ability of the�trkA mutant of SE2472 to invade, we also examined thegrowth of the �trkA mutant inside HeLa cells and compared itto the growth of wild-type strain SE2472. Since the �trkAmutant was less invasive than wild-type strain SE2472 (Fig.2A), the �trkA mutant and wild-type strain SE2472 were usedto infect HeLa cells at multiplicities of infection of 10:1 and2:1, respectively, in order to obtain similar numbers of intra-
cellular bacteria immediately after invasion. The infectedHeLa cells were incubated for up to 8 h in the presence ofgentamicin to stop further bacterial invasion, and the intracel-lular bacteria were quantified to determine the growth of the�trkA mutant and wild-type strain SE2472 inside HeLa cells.Compared to the initial levels of intracellular bacteria at zerotime, the levels of wild-type strain SE2472 were 13.4- � 2.2-and 16.4- � 3.1-fold higher at 4 and 8 h postinfection, respec-tively, while the levels of the �trkA mutant were only 4.3- �0.8- and 6.0- � 1.4-fold higher, respectively (Fig. 2B). Comple-mentation with plasmid pRB3-trkA partially rescued thegrowth defect of the �trkA mutant inside HeLa cells (Fig. 2B).The assay was terminated at 8 h, when the number of bacteriastarted to decrease, possibly because HeLa cells were beingdamaged by intracellular Salmonella and Salmonella was ex-posed to the gentamicin in the medium (data not shown). Thedefective intracellular growth of the �trkA mutant was not dueto increased susceptibility to the gentamicin used in the assay,as the �trkA mutant had the same susceptibility as the wild-type Salmonella strain to all of the antibiotics that we tested,including ampicillin, chloramphenicol, gentamicin, and tetra-cycline (data not shown). The observed intracellular growthdefect is consistent with the delayed growth the �trkA mutantobserved at K� concentrations higher than 10 mM, since hostcells have high concentrations of K�.
The �trkA mutant of Salmonella is less virulent in infectionof mice. To determine if TrkA is important for the virulence ofSalmonella, we infected mice with the �trkA mutant and wild-type strain SE2472 and determined the LD50 for intragastricand intraperitoneal infections. For both infection routes, the�trkA mutant had a higher LD50 and was less virulent thanwild-type strain SE2472 (P � 0.05, Student’s t test) (Table 3).The virulence defect was rescued in the complemented mutantby plasmid pRB3-trkA, and the difference in LD50 between thecomplemented �trkA mutant and wild-type strain SE2472 wasstatistically insignificant (P 0.4, Student’s t test) (Table 3),suggesting that deletion of trkA was responsible for the de-creased virulence of the �trkA Salmonella mutant.
FIG. 2. Epithelial cell invasion and intracellular growth of the wild-type strain and the �trkA mutant of Salmonella. (A) Invasion of HeLa cells.The ratio of the number of intracellular bacteria to the number of input bacteria was determined for wild-type strain SE2472 (WT), the �trkAmutant, and the complemented mutant (�trkA-comp). The ratio for wild-type strain SE2472 at 1 h postinfection was arbitrarily defined as 100%,and the ratios for other samples were expressed as relative values. (B) Growth inside HeLa cells. Intracellular wild-type strain SE2472, �trkAmutant, and complemented mutant (�trkA-comp) bacteria were quantified, and the results were compared to the initial number of bacteria at zerotime. The increase was calculated by dividing the number of intracellular bacteria at 4 or 8 h by the number of intracellular bacteria at zero time.The data are the averages of three experiments performed in triplicate. The error bars indicate standard deviations.
670 SU ET AL. INFECT. IMMUN.
on July 17, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

TrkA is important for expression and secretion of selectedeffector proteins of the SPI1-encoded TTSS of Salmonella. Wenext investigated how Trk contributes to the epithelial cellinvasion by and virulence of Salmonella. Since invasion ofepithelial cells by Salmonella is dependent on the effector pro-teins of the type III secretion system (TTSS) encoded by Sal-monella pathogenicity island 1 (SPI1), we reasoned that theexpression and secretion of the effector proteins of the SPI1-encoded TTSS could be altered in the �trkA mutant. We se-lected SipA, SipC, and SopB for this analysis because theseproteins have previously been shown to have important func-tions in host cell invasion and in the modulation of the hostcytoskeleton to facilitate Salmonella infection (20, 21, 40). Tomonitor the levels of these proteins, we introduced into thechromosome a tandem tag consisting of six histidines and aFLAG tag fused in frame to the C terminus of each protein(45) to generate tagged strains SipA(HF), SipC(HF), and Sop-B(HF) (Table 1). No difference between the tagged strains andparental strain SE2472 was observed for growth in vitro and forvirulence in mouse infections, suggesting that tagging did nothave deleterious effects (data not shown). The �trkA mutationwas transduced into each tagged strain, and the mutant strainswere complemented by transformation with plasmid pRB3-trkA. Equal quantities of proteins from both whole-cell lysateand culture supernatant were used for each strain. Since lessprotein was recovered from the culture supernatant of the�trkA mutant, proteins obtained from larger volumes of cul-tures were used to ensure that the total quantities of proteinsfrom each strain were equivalent (data not shown). The levelsof each tagged protein in the whole bacterial lysates and cul-ture supernatants of the corresponding set of tagged strainswith the wild-type, mutant, or complemented trkA allele wereanalyzed by Western hybridization using an anti-FLAG anti-body (Fig. 3).
We analyzed SipA, SipC, and SopB, and SipA was detectedin both the whole bacterial lysate and the culture supernatant,while SipC and SopB were detected only in the culture super-natant (Fig. 3). Compared to the results for the parental strainswith the wild-type trkA allele, the level of SipA was lower in thewhole bacterial lysate of the �trkA mutant (Fig. 3A) and thelevels of SipA, SipC, and SopB were lower in the culturesupernatant of the �trkA mutant (Fig. 3B). In each comple-mented �trkA mutant strain, the level of SipA, SipC, or SopBwas restored to the level in the parental strain (Fig. 3).
Potassium and sodium enhance the invasion of epithelialcells by Salmonella. We showed in the experiments describedabove that the Trk K� transport system is necessary for Sal-monella to invade and grow in epithelial cells and for expres-sion and secretion of effector proteins of the SPI1-encodedTTSS. Next we examined the effect of external K� on Salmo-
nella and whether external K� modulated the invasion of Sal-monella and the expression of effector proteins of the TTSS.The invasion of HeLa cells by bacteria cultured with variousconcentrations of KCl was analyzed, and the results were com-pared with the results for organisms cultured in LB brothalone. We chose moderate K� concentrations (50 and 100mM) that are in the physiological range that Salmonella mayencounter in its hosts. When cultured in the presence of eitherconcentration of supplemental KCl, both the wild type and the�trkA mutant showed enhanced invasion of HeLa cells (Fig.4A). The level of invasion of HeLa cells by the �trkA mutantcultured with supplemental KCl increased to the level of inva-sion by wild-type strain SE2472 cultured without the supple-ment, although the �trkA mutant was consistently less invasivethan wild-type strain SE2472 cultured in the same medium(Fig. 4A). To determine if the enhanced invasion of HeLa cellsby Salmonella could also be modulated by other cations thathave physiological relevance, we determined the effect of sup-plemental NaCl on the invasiveness of Salmonella. Na� is themost abundant extracellular cation, and the plasma level of thiscation in humans is 135 to 145 mM. As shown in Fig. 4B, thepresence of supplemental NaCl increased the invasiveness ofboth wild-type SE2472 and the �trkA mutant of SE2472, al-though the increase in invasion after addition of supplementalNaCl was not as pronounced as the increase in invasion afteraddition of KCl (Fig. 4A and 4B). This suggests that Salmo-nella responds to physiological concentrations of both Na� andK� and that K� is likely to have a greater effect on the invasionof HeLa cells by Salmonella.
FIG. 3. Expression and secretion of selected effector proteins ofthe SPI1-encoded TTSS by the wild-type strain and �trkA mutantof Salmonella. Epitope-tagged strains SipA(HF), SipC(HF), andSopB(HF) with the wild-type (WT) or �trkA mutant allele and the�trkA mutant transformed with plasmid pRB3-trkA (�trkA-comp)were cultured in LB broth. The bacterial lysates or cultural superna-tants were analyzed to determine the levels of SipA, SipC, and SopB inthe tagged strains. (A) Western blot analysis of the SipA levels inwhole-cell lysates. (B) Western blot analysis of the levels of SipA,SipC, and SopB in culture supernatants. The trkA allele in each sampleis indicated above the lane. The effector proteins analyzed in the blotsare indicated on the right.
TABLE 3. LD50 of Salmonella wild-type strain SE2472 and the �trkAmutant in intragastric and intraperitoneal infections of mice
StrainLD50 (CFU)
Intragastric infection Intraperitoneal infection
SE2472 4.5 � 103 1.0 � 101
�trkA 3.7 � 104 1.5 � 102
�trkA-comp 2.7 � 103 1.3 � 101
VOL. 77, 2009 ROLE OF POTASSIUM IN SALMONELLA PATHOGENESIS 671
on July 17, 2014 by guesthttp://iai.asm
.org/D
ownloaded from
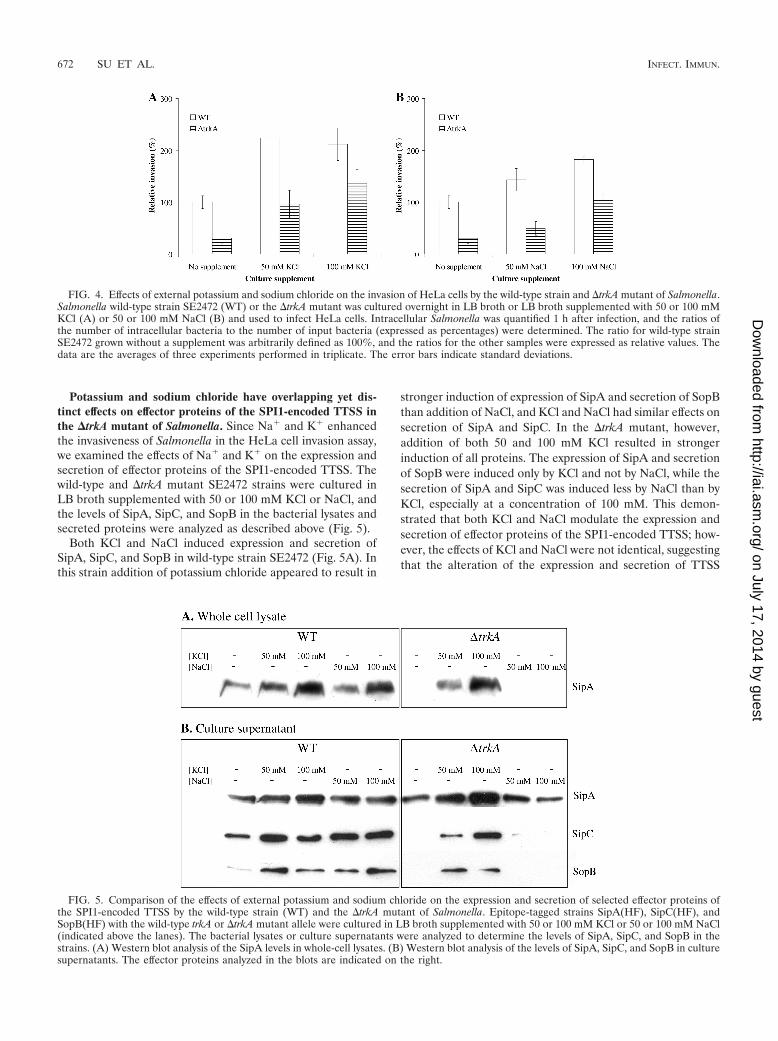
Potassium and sodium chloride have overlapping yet dis-tinct effects on effector proteins of the SPI1-encoded TTSS inthe �trkA mutant of Salmonella. Since Na� and K� enhancedthe invasiveness of Salmonella in the HeLa cell invasion assay,we examined the effects of Na� and K� on the expression andsecretion of effector proteins of the SPI1-encoded TTSS. Thewild-type and �trkA mutant SE2472 strains were cultured inLB broth supplemented with 50 or 100 mM KCl or NaCl, andthe levels of SipA, SipC, and SopB in the bacterial lysates andsecreted proteins were analyzed as described above (Fig. 5).
Both KCl and NaCl induced expression and secretion ofSipA, SipC, and SopB in wild-type strain SE2472 (Fig. 5A). Inthis strain addition of potassium chloride appeared to result in
stronger induction of expression of SipA and secretion of SopBthan addition of NaCl, and KCl and NaCl had similar effects onsecretion of SipA and SipC. In the �trkA mutant, however,addition of both 50 and 100 mM KCl resulted in strongerinduction of all proteins. The expression of SipA and secretionof SopB were induced only by KCl and not by NaCl, while thesecretion of SipA and SipC was induced less by NaCl than byKCl, especially at a concentration of 100 mM. This demon-strated that both KCl and NaCl modulate the expression andsecretion of effector proteins of the SPI1-encoded TTSS; how-ever, the effects of KCl and NaCl were not identical, suggestingthat the alteration of the expression and secretion of TTSS
FIG. 4. Effects of external potassium and sodium chloride on the invasion of HeLa cells by the wild-type strain and �trkA mutant of Salmonella.Salmonella wild-type strain SE2472 (WT) or the �trkA mutant was cultured overnight in LB broth or LB broth supplemented with 50 or 100 mMKCl (A) or 50 or 100 mM NaCl (B) and used to infect HeLa cells. Intracellular Salmonella was quantified 1 h after infection, and the ratios ofthe number of intracellular bacteria to the number of input bacteria (expressed as percentages) were determined. The ratio for wild-type strainSE2472 grown without a supplement was arbitrarily defined as 100%, and the ratios for the other samples were expressed as relative values. Thedata are the averages of three experiments performed in triplicate. The error bars indicate standard deviations.
FIG. 5. Comparison of the effects of external potassium and sodium chloride on the expression and secretion of selected effector proteins ofthe SPI1-encoded TTSS by the wild-type strain (WT) and the �trkA mutant of Salmonella. Epitope-tagged strains SipA(HF), SipC(HF), andSopB(HF) with the wild-type trkA or �trkA mutant allele were cultured in LB broth supplemented with 50 or 100 mM KCl or 50 or 100 mM NaCl(indicated above the lanes). The bacterial lysates or culture supernatants were analyzed to determine the levels of SipA, SipC, and SopB in thestrains. (A) Western blot analysis of the SipA levels in whole-cell lysates. (B) Western blot analysis of the levels of SipA, SipC, and SopB in culturesupernatants. The effector proteins analyzed in the blots are indicated on the right.
672 SU ET AL. INFECT. IMMUN.
on July 17, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

effector proteins was not simply due to a change in the osmoticpressure, especially in the �trkA mutant of Salmonella.
DISCUSSION
The importance of K� for bacterial physiology has beenstudied by several investigators (for a review, see reference 15);however, the function of intracellular and external K� in thepathogenicity of bacteria has not been well characterized. Inthis report, we provide direct evidence that maintaining aproper intracellular K� level is important for the pathogenicityof Salmonella and that external K� regulates the virulencecharacteristics of Salmonella. Salmonella lacking the low-affin-ity K� transporter Trk invaded epithelial cells less, was lessvirulent in mouse infections, and expressed and secretedsmaller amounts of effector proteins of the SPI1-encodedTTSS (Fig. 2 and 3, and Table 3), while addition of K� im-proved epithelial cell invasion and the expression and secretionof the same set of the effector proteins (Fig. 4 and 5). To ourknowledge, this is the first study demonstrating that both in-tracellular K� and external K� influence the virulence char-acteristics of a pathogenic bacterium. The only other exampleof the effect of K� on human pathogens is a recent report byKumar et al., who demonstrated that exposure of Plasmodiumsporozoites to the intracellular concentration of K� enhancesinfectivity and reduces cell passage activity (24).
The effect of K� on human pathogens is not unexpected,since K� is the most abundant cation inside human cells, wherethe concentrations are more than 100 mM, while the extracel-lular concentrations are usually less than 10 mM (22). The K�
concentration inside Salmonella-containing vacuoles has notbeen reported; however, the K� concentration inside Myco-bacterium-containing vacuoles inside macrophages is approxi-mately 20 to 50 mM and is in the range of K� concentrationsthat we used in this analysis (47). In the human small intestinewhere Salmonella infection occurs, the K� concentration hasbeen reported to be approximately 6 mM (16). However, dur-ing diarrhea, the K� concentration is likely to be much higherdue to electrolytes released from damaged intestinal mucosa.Although results of in vitro assays may not be directly appli-cable to natural infections, our results indicate that it is possi-ble that Salmonella benefits from the increased K� concentra-tion in the intestinal fluid when diarrhea occurs and becomesmore invasive. In addition to the results for K�, we have alsoshown that Na�, the most abundant extracellular cation, in-creased the invasion of epithelial cells by Salmonella and theexpression and secretion of effector proteins of the SPI1-en-coded TTSS (Fig. 4B and 5). The concentrations of Na� thatwe used are within the range of the normal plasma concentra-tion of Na� (135 mM), suggesting that Na� may modulate theinfectivity of Salmonella during systemic infection. It has beenreported previously that invasion by Salmonella is modulatedby oxygen, acids (acetic and butyric acids), bile, and formate inthe distal ileum (19, 27, 39, 46). Taken together, these resultssuggest that Salmonella responds to multiple signals in vivo inorder to regulate its invasion and pathogenesis in differentorgans, and the ability to respond to a wide spectrum of ex-ternal signals is likely a factor that contributes to Salmonella’ssuccess as a human pathogen.
In gram-negative bacteria, the intracellular concentration of
K� is regulated by multiple K� transport systems, includingthe Trk and Kup low-affinity systems and the Kdp high-affinitysystem (for a review, see reference 15). Since the �trkA mutanthas normal growth properties except at high concentrations ofK�, we believe that the �trkA mutant has relatively normalphysiology and that other K� transport systems can compen-sate for the lack of Trk. We do not know yet which K� trans-port system(s) compensates for the function of Trk in the�trkA mutant of Salmonella or the extent of the compensation.At high concentrations of K� (10 mM or higher), Kdp isrepressed, which probably explains why the �trkA mutant grewmore slowly in the log phase (2, 17, 25, 34) (Fig. 1). Both Kupand Kdp likely contribute to maintaining the K� homeostasisin the absence of Trk at lower K� concentrations. In E. coliand Salmonella, the kdp operon is normally repressed at a K�
concentration of 5 mM or higher; however, this operon can beexpressed at K� concentrations around 10 mM in the absenceof Trk (2, 17, 25, 34). Our preliminary analysis of the Kuptransport system indicated that a �kup �trkA double mutantwas more defective in growth, protein secretion, and patho-genesis than either single mutant (unpublished results). Athorough analysis of mutants with deletions in multiple K�
transport systems should provide insight into the specific func-tions of each K� transport system.
The SPI1-encoded TTSS is regulated by multiple signals in ahighly complex regulatory network (for a review, see reference14). Factors that have been reported to directly regulate theSPI-encoded TTSS are HilA (1, 4, 13, 28, 29) and InvF (10, 11,13). Factors that indirectly regulate SPI1 through HilA andInvF include PhoQ/PhoP, HilE, HilC, HilD, RtsA, FimZY,PhoR/PhoB, Fur, BarA/SirA, and EnvZ/OmpR (for a review,see reference 14). Perhaps more relevant for the pathogenesisof Salmonella are the environmental conditions that regulategenes of the SPI1-encoded TTSS, including pH, oxygen, osmo-larity, growth phase, and bile (5, 18, 26–28, 33, 38, 39), some ofwhich have been shown to directly affect the invasiveness ofSalmonella. For example, low oxygen tension and acetic acidwere shown to enhance Salmonella invasion, while bile andbutyric acid reduce invasion (27, 39, 46). Our direct compari-son of the expression and secretion of the SipA, SipC, andSopB proteins in the presence of KCl and NaCl and the inva-siveness of Salmonella cultured with supplemental KCl orNaCl indicated that both KCl and NaCl regulated the effectorproteins of the SPI1-encoded TTSS; however, the effects werenot identical (Fig. 4 and 5). Addition of potassium chlorideresulted in stronger induction of the proteins overall, especiallyin the �trkA mutant, whereas addition of NaCl had little or noeffect on the expression of SipA and the secretion of SipC andSopB (Fig. 4 and 5). This suggests that the signaling pathwaysinvolved were not identical for KCl and NaCl, and the induc-tion of the effector proteins of the SPI1-encoded TTSS byNaCl was at least partially dependent on Trk. However, themolecular mechanisms that determine the different effects thatKCl and NaCl have on the TTSS of Salmonella lacking the Trksystem have not been characterized yet. One hypothesis toexplain the difference between supplemental KCl and supple-mental NaCl is that they may affect the alternative K� trans-porter(s) that Salmonella uses in the absence of the major K�
transporter Trk. Without Trk, the high-affinity Kdp transporterand the low-affinity Kup transporter likely substitute for Trk,
VOL. 77, 2009 ROLE OF POTASSIUM IN SALMONELLA PATHOGENESIS 673
on July 17, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

and K� downregulates Kdp but not Kup. As a result, Kup maybe more active with supplemental K� than with supplementalNa�, leading to a state more similar to that of the wild-typebacteria.
After we performed the experiments reported here, Mi-zusaki et al. reported that the secretion of Sips was induced bysucrose and several salts, including NaCl and KCl, and thatNaCl-induced secretion of Sips was due to induction of hilAexpression mediated by BarA/SirA but not by EnvZ (36). Wedo not know yet how the Trk system affects the regulation ofSPI1 and why the expression and secretion of effector proteinsof the SPI1-encoded TTSS were defective in the �trkA mutantof Salmonella. The defect did not occur because the �trkAmutant expressed less hilA and invF, which encode direct reg-ulators of SPI1 expression. In fact, we found that the expres-sion of both hilA and invF was elevated in the �trkA mutant(unpublished results). An expression profile analysis of thewild-type strain and �trkA mutant of Salmonella and an anal-ysis of physiological changes caused by mutations in K� trans-porters and external K� should provide more insight into howpotassium transporters and external K� regulate the growthand pathogenicity of Salmonella.
ACKNOWLEDGMENTS
We thank Hiroshi Nikaido for insightful discussions and C. S.Shashikant for critical comments on the manuscript.
This work was supported by USDA grant CALR-2005-01892 to S.L.
REFERENCES
1. Ahmer, B. M., J. van Reeuwijk, P. R. Watson, T. S. Wallis, and F. Heffron.1999. Salmonella SirA is a global regulator of genes mediating enteropatho-genesis. Mol. Microbiol. 31:971–982.
2. Asha, H., and J. Gowrishankar. 1993. Regulation of kdp operon expressionin Escherichia coli: evidence against turgor as signal for transcriptional con-trol. J. Bacteriol. 175:4528–4537.
3. Ausubel, F., R. Brent, R. Kingston, D. Moore, J. Smith, and K. Struhl. 1997.Current protocols in molecular biology. John Wiley & Sons, New York, NY.
4. Bajaj, V., C. Hwang, and C. A. Lee. 1995. hilA is a novel ompR/toxR familymember that activates the expression of Salmonella typhimurium invasiongenes. Mol. Microbiol. 18:715–727.
5. Bajaj, V., R. L. Lucas, C. Hwang, and C. A. Lee. 1996. Co-ordinate regulationof Salmonella typhimurium invasion genes by environmental and regulatoryfactors is mediated by control of hilA expression. Mol. Microbiol. 22:703–714.
6. Berggren, R. E., A. Wunderlich, E. Ziegler, M. Schleicher, R. C. Duke, D.Looney, and F. C. Fang. 1995. HIV gp120-specific cell-mediated immuneresponses in mice after oral immunization with recombinant Salmonella. J.Acquir. Immune Defic. Syndr. Hum. Retrovirol. 10:489–495.
7. Bossemeyer, D., A. Borchard, D. C. Dosch, G. C. Helmer, W. Epstein, I. R.Booth, and E. P. Bakker. 1989. K�-transport protein TrkA of Escherichia coliis a peripheral membrane protein that requires other trk gene products forattachment to the cytoplasmic membrane. J. Biol. Chem. 264:16403–16410.
8. Bossemeyer, D., A. Schlosser, and E. P. Bakker. 1989. Specific cesium trans-port via the Escherichia coli Kup (TrkD) K� uptake system. J. Bacteriol.171:2219–2221.
9. Chen, Y. C., Y. C. Chuang, C. C. Chang, C. L. Jeang, and M. C. Chang. 2004.A K� uptake protein, TrkA, is required for serum, protamine, and polymyxinB resistance in Vibrio vulnificus. Infect. Immun. 72:629–636.
10. Darwin, K. H., and V. L. Miller. 1999. InvF is required for expression ofgenes encoding proteins secreted by the SPI1 type III secretion apparatus inSalmonella typhimurium. J. Bacteriol. 181:4949–4954.
11. Darwin, K. H., and V. L. Miller. 2000. The putative invasion protein chap-erone SicA acts together with InvF to activate the expression of Salmonellatyphimurium virulence genes. Mol. Microbiol. 35:949–960.
12. Datsenko, K. A., and B. L. Wanner. 2000. One-step inactivation of chromo-somal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad.Sci. USA 97:6640–6645.
13. Eichelberg, K., and J. E. Galan. 1999. Differential regulation of Salmonellatyphimurium type III secreted proteins by pathogenicity island 1 (SPI-1)-encoded transcriptional activators InvF and HilA. Infect. Immun. 67:4099–4105.
14. Ellermeier, J. R., and J. M. Slauch. 2007. Adaptation to the host environ-
ment: regulation of the SPI1 type III secretion system in Salmonella entericaserovar Typhimurium. Curr. Opin. Microbiol. 10:24–29.
15. Epstein, W. 2003. The roles and regulation of potassium in bacteria. Prog.Nucleic Acid Res. Mol. Biol. 75:293–320.
16. Fordtran, J. S., and T. W. Locklear. 1966. Ionic constituents and osmolalityof gastric and small-intestinal fluids after eating. Am. J. Dig. Dis. 11:503–521.
17. Frymier, J. S., T. D. Reed, S. A. Fletcher, and L. N. Csonka. 1997. Charac-terization of transcriptional regulation of the kdp operon of Salmonellatyphimurium. J. Bacteriol. 179:3061–3063.
18. Galan, J. E., and R. Curtiss III. 1990. Expression of Salmonella typhimuriumgenes required for invasion is regulated by changes in DNA supercoiling.Infect. Immun. 58:1879–1885.
19. Huang, Y., M. Suyemoto, C. D. Garner, K. M. Cicconi, and C. Altier. 2008.Formate acts as a diffusible signal to induce Salmonella invasion. J. Bacteriol.190:4233–4241.
20. Jepson, M. A., B. Kenny, and A. D. Leard. 2001. Role of sipA in the earlystages of Salmonella typhimurium entry into epithelial cells. Cell. Microbiol.3:417–426.
21. Kaniga, K., D. Trollinger, and J. E. Galan. 1995. Identification of two targetsof the type III protein secretion system encoded by the inv and spa loci ofSalmonella typhimurium that have homology to the Shigella IpaD and IpaAproteins. J. Bacteriol. 177:7078–7085.
22. Kem, D. C., and D. Trachewsky. 1983. Potassium metabolism, p. 25–35. In R.Whang (ed.), Potassium: its biologic significance. CRC Press, Inc., BocaRaton, FL.
23. Komoriya, K., N. Shibano, T. Higano, N. Azuma, S. Yamaguchi, and S. I.Aizawa. 1999. Flagellar proteins and type III-exported virulence factors arethe predominant proteins secreted into the culture media of Salmonellatyphimurium. Mol. Microbiol. 34:767–779.
24. Kumar, K. A., C. R. Garcia, V. R. Chandran, N. Van Rooijen, Y. Zhou, E.Winzeler, and V. Nussenzweig. 2007. Exposure of Plasmodium sporozoites tothe intracellular concentration of potassium enhances infectivity and reducescell passage activity. Mol. Biochem. Parasitol. 156:32–40.
25. Laimins, L. A., D. B. Rhoads, and W. Epstein. 1981. Osmotic control of kdpoperon expression in Escherichia coli. Proc. Natl. Acad. Sci. USA 78:464–468.
26. Leclerc, G. J., C. Tartera, and E. S. Metcalf. 1998. Environmental regulationof Salmonella typhi invasion-defective mutants. Infect. Immun. 66:682–691.
27. Lee, C. A., and S. Falkow. 1990. The ability of Salmonella to enter mamma-lian cells is affected by bacterial growth state. Proc. Natl. Acad. Sci. USA87:4304–4308.
28. Lee, C. A., B. D. Jones, and S. Falkow. 1992. Identification of a Salmonellatyphimurium invasion locus by selection for hyperinvasive mutants. Proc.Natl. Acad. Sci. USA 89:1847–1851.
29. Lostroh, C. P., and C. A. Lee. 2001. The HilA box and sequences outside itdetermine the magnitude of HilA-dependent activation of PprgH from Sal-monella pathogenicity island 1. J. Bacteriol. 183:4876–4885.
30. Lu, S., P. B. Killoran, F. C. Fang, and L. W. Riley. 2002. The global regulatorArcA controls resistance to reactive nitrogen and oxygen intermediates inSalmonella enterica serovar Enteritidis. Infect. Immun. 70:451–461.
31. Lu, S., P. B. Killoran, and L. W. Riley. 2003. Association of Salmonellaenterica serovar Enteritidis yafD with resistance to chicken egg albumen.Infect. Immun. 71:6734–6741.
32. Lu, S., A. R. Manges, Y. Xu, F. C. Fang, and L. W. Riley. 1999. Analysis ofvirulence of clinical isolates of Salmonella enteritidis in vivo and in vitro.Infect. Immun. 67:5651–5657.
33. Lundberg, U., U. Vinatzer, D. Berdnik, A. von Gabain, and M. Baccarini.1999. Growth phase-regulated induction of Salmonella-induced macrophageapoptosis correlates with transient expression of SPI-1 genes. J. Bacteriol.181:3433–3437.
34. Malli, R., and W. Epstein. 1998. Expression of the Kdp ATPase is consistentwith regulation by turgor pressure. J. Bacteriol. 180:5102–5108.
35. Maloy, S. R., V. J. Stewart, and R. K. Taylor. 1996. Genetic analysis ofpathogenic bacteria. Cold Spring Harbor Laboratory Press, Cold SpringHarbor, NY.
36. Mizusaki, H., A. Takaya, T. Yamamoto, and S. I. Aizawa. 2008. Signalpathway in salt-activated expression of the Salmonella pathogenicity island 1type III secretion system in Salmonella enterica serovar Typhimurium. J.Bacteriol. 190:4624–4631.
37. Parra-Lopez, C., R. Lin, A. Aspedon, and E. A. Groisman. 1994. A Salmo-nella protein that is required for resistance to antimicrobial peptides andtransport of potassium. EMBO J. 13:3964–3972.
38. Prouty, A. M., I. E. Brodsky, J. Manos, R. Belas, S. Falkow, and J. S. Gunn.2004. Transcriptional regulation of Salmonella enterica serovar Typhimuriumgenes by bile. FEMS Immunol. Med. Microbiol. 41:177–185.
39. Prouty, A. M., and J. S. Gunn. 2000. Salmonella enterica serovar Typhi-murium invasion is repressed in the presence of bile. Infect. Immun. 68:6763–6769.
40. Raffatellu, M., R. P. Wilson, D. Chessa, H. Andrews-Polymenis, Q. T. Tran,S. Lawhon, S. Khare, L. G. Adams, and A. J. Baumler. 2005. SipA, SopA,SopB, SopD, and SopE2 contribute to Salmonella enterica serotype Typhi-murium invasion of epithelial cells. Infect. Immun. 73:146–154.
674 SU ET AL. INFECT. IMMUN.
on July 17, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

41. Reed, L. J., and H. Muench. 1938. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 27:493–497.
42. Sambrook, J., and D. W. Russell. 2001. Molecular cloning: a laboratory manual,3rd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
43. Trchounian, A., and H. Kobayashi. 2000. K� uptake by fermenting Esche-richia coli cells: pH dependent mode of the TrkA system operating. Biosci.Rep. 20:277–288.
44. Trchounian, A., and H. Kobayashi. 1999. Kup is the major K� uptake systemin Escherichia coli upon hyper-osmotic stress at a low pH. FEBS Lett.447:144–148.
45. Uzzau, S., N. Figueroa-Bossi, S. Rubino, and L. Bossi. 2001. Epitope taggingof chromosomal genes in Salmonella. Proc. Natl. Acad. Sci. USA 98:15264–15269.
46. Van Immerseel, F., J. De Buck, I. De Smet, F. Pasmans, F. Haesebrouck, andR. Ducatelle. 2004. Interactions of butyric acid- and acetic acid-treatedSalmonella with chicken primary cecal epithelial cells in vitro. Avian Dis.48:384–391.
47. Wagner, D., J. Maser, B. Lai, Z. Cai, C. E. Barry III, K. Honer ZuBentrup, D. G. Russell, and L. E. Bermudez. 2005. Elemental analysis ofMycobacterium avium-, Mycobacterium tuberculosis-, and Mycobacteriumsmegmatis-containing phagosomes indicates pathogen-induced microen-vironments within the host cell’s endosomal system. J. Immunol. 174:1491–1500.
48. Zakharyan, E., and A. Trchounian. 2001. K� influx by Kup in Escherichiacoli is accompanied by a decrease in H� efflux. FEMS Microbiol. Lett.204:61–64.
Editor: A. J. Baumler
VOL. 77, 2009 ROLE OF POTASSIUM IN SALMONELLA PATHOGENESIS 675
on July 17, 2014 by guesthttp://iai.asm
.org/D
ownloaded from




















