
Distribution of neurotensin-immunoreactivity within baroreceptive portions of the nucleus of the...
10
THE JOURNAL OF COMPARATIVE NEUROLOGY 226: 155-164 (1984) Distribution of Neurotensin - Immunoreactivi ty Within Baroreceptive Portions of the Nucleus of the Tractus Solitarius and the Dorsal Vagal Nucleus of the Rat GERALD A. HIGGINS, GLORIA E. HOFFMAN, SUSAN WRAY, AND JAMES S. SCHWABER Central Research and Development Department, E. I. du Pont de Nemours and Company, Glenolden, Pennsylvania 19036 (G.A.H., J.S.S.), Department of Anatomy and Neurobiology, University of Vermont College of Medicine, Burlington, Vermont 05405 (G.A.H.), and Department of Anatomy, University of Rochester School of Medicine and Dentistry, Rochester, New York 14642 (G.E.H., S.W.) ABSTRACT We have examined the distribution of neurotensin immunoreactivity within subnuclear regions of the nucleus of the tractus solitarius (NTS) and the dorsal motor nucleus of the vagus nerve (DVN) in the rat. In order to determine which regions of the NTS were involved in the regulation of baroreceptor reflexes, we mapped the central distribution of the aortic branch of the vagus nerve using transganglionic transport of horseradish peroxi- dase. Comparison of the pattern of aortic nerve innervation with that of the distribution of neurotensin-immunoreactivecells and fibers shows the dor- somedial nucleus of the NTS both to be the primary site of aortic barorecep- tor termination and to contain the highest concentration of neurotensin- immunoreactive elements within the NTS. Neurotensin-immunoreactive fibers are also present in medial regions of the NTS adjacent to the area postrema where they may be involved in the modulation of vagal gastric afferents. Double-label experiments, in which, on the same tissue sections, neuro- tensin immunohistochemistry was combined with retrograde horseradish peroxidase labeling of DVN neurons, reveal a topographic innervation of vagal preganglionic motoneurons by neurotensin-immunoreactive fibers. The heaviest innervation is of lateral portions of the DVN and adjacent ventral portions of the NTS at the level of the obex, an area which may contain cardiac motoneurons. In this region neurotensin-immunoreactive fibers can be observed in close proximity to retrogradely labeled cells. The concentration of neurotensin elements in a region of the NTS which is involved in the control of baroreceptor reflexes provides a morphological basis for the cardiovascular effects produced by central administration of the peptide. Additional control may be exerted at the level of the motoneu- ron, as evidenced by apparent neurotensin fiber innervation of presumptive cardiac preganglionic neurons. Similarly, the distribution of neurotensin fibers suggests that the peptide may be acting in gastric regulatory areas of the NTS or on vagal secretomotor neurons to regulate gastric acid secretion. Key words: neurotensin immunoreactivity, tractus solitarius, dorsal motor nucleus, vagus nerve A growing body of evidence suggests that neurotensin- ergic neurons in the central nervous system are involved in the regulation of autonomic function (Prange and Nemer- off, '82). For example, intracerebroventricular (i.c.v.) ad- ministration of neurotensin produces changes in blood pres- 0 1984 ALAN R. LISS, INC. Accepted December 2, 1983. Address reprint requests to Dr. G.A. Higgins, Department of Molecular Biology, Research Institute of Scripps Clinic, La Jolla, CA 92037.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Distribution of neurotensin-immunoreactivity within baroreceptive portions of the nucleus of the...

THE JOURNAL OF COMPARATIVE NEUROLOGY 226: 155-164 (1984)
Distribution of Neurotensin - Immunoreactivi ty Within Baroreceptive Portions of the Nucleus of the Tractus
Solitarius and the Dorsal Vagal Nucleus of the Rat
GERALD A. HIGGINS, GLORIA E. HOFFMAN, SUSAN WRAY, AND
JAMES S. SCHWABER Central Research and Development Department, E. I. du Pont de Nemours and
Company, Glenolden, Pennsylvania 19036 (G.A.H., J.S.S.), Department of Anatomy and Neurobiology, University of Vermont College of Medicine, Burlington, Vermont 05405 (G.A.H.), and Department of Anatomy, University of Rochester School of Medicine and
Dentistry, Rochester, New York 14642 (G.E.H., S.W.)
ABSTRACT We have examined the distribution of neurotensin immunoreactivity
within subnuclear regions of the nucleus of the tractus solitarius (NTS) and the dorsal motor nucleus of the vagus nerve (DVN) in the rat. In order to determine which regions of the NTS were involved in the regulation of baroreceptor reflexes, we mapped the central distribution of the aortic branch of the vagus nerve using transganglionic transport of horseradish peroxi- dase. Comparison of the pattern of aortic nerve innervation with that of the distribution of neurotensin-immunoreactive cells and fibers shows the dor- somedial nucleus of the NTS both to be the primary site of aortic barorecep- tor termination and to contain the highest concentration of neurotensin- immunoreactive elements within the NTS. Neurotensin-immunoreactive fibers are also present in medial regions of the NTS adjacent to the area postrema where they may be involved in the modulation of vagal gastric afferents.
Double-label experiments, in which, on the same tissue sections, neuro- tensin immunohistochemistry was combined with retrograde horseradish peroxidase labeling of DVN neurons, reveal a topographic innervation of vagal preganglionic motoneurons by neurotensin-immunoreactive fibers. The heaviest innervation is of lateral portions of the DVN and adjacent ventral portions of the NTS at the level of the obex, an area which may contain cardiac motoneurons. In this region neurotensin-immunoreactive fibers can be observed in close proximity to retrogradely labeled cells.
The concentration of neurotensin elements in a region of the NTS which is involved in the control of baroreceptor reflexes provides a morphological basis for the cardiovascular effects produced by central administration of the peptide. Additional control may be exerted at the level of the motoneu- ron, as evidenced by apparent neurotensin fiber innervation of presumptive cardiac preganglionic neurons. Similarly, the distribution of neurotensin fibers suggests that the peptide may be acting in gastric regulatory areas of the NTS or on vagal secretomotor neurons to regulate gastric acid secretion.
Key words: neurotensin immunoreactivity, tractus solitarius, dorsal motor nucleus, vagus nerve
A growing body of evidence suggests that neurotensin- ergic neurons in the central nervous system are involved in the regulation of autonomic function (Prange and Nemer- off, '82). For example, intracerebroventricular (i.c.v.) ad- ministration of neurotensin produces changes in blood pres-
0 1984 ALAN R. LISS, INC.
Accepted December 2, 1983.
Address reprint requests to Dr. G.A. Higgins, Department of Molecular Biology, Research Institute of Scripps Clinic, La Jolla, CA 92037.

156 G.A. HIGGINS ET AL.
sure (Quirion et al., '81; Rioux et al., '81; Sumners et al., '81) and inhibition of gastric acid secretion (Osumi et al., '78). Immunocytochemical (Jennes et al., '82; Uhl et al., '79) and radioimmunoassay (Emson et al., '82) studies show that the peptide is concentrated in autonomic regulatory struc- tures, which also contain receptors for neurotensin (Young and Kuhar, '79).
The nucleus of the tractus solitarius (NTS) and dorsal motor nucleus of the vagus nerve (DVN) contain abundant neurotensinergic cells and fibers (Jennes et al., '821, as well as a variety of other neuropeptides (Armstrong et al., '82; Maley and Elde, '82). Recent studies suggest that the NTS and DVN have complex viscerotopic organizations, with separate regions of the NTS involved in the regulation of cardiovascular, respiratory, and gastric function (Higgins and Schwaber, '82a; Loewy and Burton, '78). This subnu- clear heterogeneity makes accurate immunohistochemical localization a requirement for understanding the role of neuropeptides in the functions regulated by these nuclei.
In the present study we have attempted to relate neuro- tensin to the autonomic functions mediated by the NTS and DVN by mapping the distribution of neurotensin-immuno- reactive cells and fibers within these nuclei. To determine whether neurotensin may be involved in the regulation of baroreceptor reflex mechanisms within the NTS, we have compared its distribution to that of the aortic branch of the vagus nerve within the nucleus. Simultaneous neurotensin immunohistochemistry and retrograde cell labeling from the vagus nerve were used to examine the relationship of neurotensinergic fibers to preganglionic neurons of the DVN.
METHODS Labeling of the central connections of the
aortic nerve Adult male Wistar-Kyoto rats (Charles River) weighing
250-350 gm were used in this study. This strain has a separate, easily identifiable aortic branch of the vagus nerve (Helke et al., '80). Under chloral hydrate anesthesia (400 mgkg), the nerve was identified at its juncture with the superior laryngeal nerve, then carefully dissected out and sectioned, and the cut, central end was placed into a sealed polyethylene cuff containing a 30% solution of horseradish peroxidase (Boerhinger-Mannheim, grade one) for 1 hour. During incubation, a piece of polyethylene was placed un- der the cuff to prevent spread of the enzyme. Following application of the enzyme, the cuff was removed and the area was thoroughly cleaned to remove any traces of the horseradish peroxidase.
Survival times ranged from 24 to 48 hours, following which the animal was deeply anesthetized with sodium pentobarbital, perfused transaortically with isotonic saline, followed by a mixture of 1% paraformaldehyde and 1.25% glutaraldehyde at pH 7.4 (500 ml), and finally, by a postfix- ation rinse of chilled, buffered 10% sucrose. The medullas were removed and sectioned at 40 pm on a freezing micro- tome. Transverse sections were collected and reacted for the presence of transganglionic and retrograde horseradish peroxidase using tetramethylbenzidine as the chromagen.
Retrograde labeling of the vagus nerve combined with neurotensin immunohistochemistry
For studies of vagal connections, adult male Sprague- Dawley or Wistar-Kyoto rats weighing 250-350 gm were used. The animals were anesthetized with chloral hydrate and the right cervical vagus nerve was dissected out and
sectioned just distal to the nodose ganglion. Horseradish peroxidase was applied to the nerve as above.
In six animals, colchicine (75 pg/5 pl) was administered intracisternally (i.c.)24 hours after application of the tracer to block axonal transport, causing accumulation of the pep- tide in cell bodies. After survival times of 24 to 48 hours, the animals were deeply anesthetized with sodium pento- barbital and perfused with saline (150-300 ml). This was followed by 500 ml of Zamboni's fixative (buffered formalin and picric acid). Brains were sectioned on a vibrating micro- tome (Oxford Instruments or D.S.K.), and 25-pm transverse sections were collected.
Double staining for the presence of retrogradely trans- ported horseradish peroxidase and for neurotensin on the same tissue section was accomplished using a modification of the procedure of Hoffman et al. ('82). The tissue was developed for retrograde horseradish peroxidase using 3,3'- diaminobenzidine4HCl (50 pg/ml) as the chromagen. For immunocytochemical localization, the tissue was then rinsed in potassium phosphate buffered saline (KPBS; ph 7.2) and placed in a 1:1,000 dilution of antineurotensin (Immunonuclear Corp., Stillwater, MN) in KPBS with 0.4% Triton X-100 for 12-14 hours at 4°C. The tissue was next very thoroughly rinsed in KPBS with 0.02% Trition X-100 and placed, at room temperature, into a 1:40 dilution of goat antirabbit immunoglobulin (Bachem) in KPBS 0.02% Triton-X-100 for 1 hour followed by incubation in a 1 : l O O dilution of peroxidase-antiperoxidase complex (Miles-Yeda) for 1 hour. After rinsing, the complex was visualized using 4-chloro 1-naphthol (100 mg15 ml ethyl alcohol dissolved in 225 ml TRIS buffer, pH 7) as the chromagen to yield a blue reaction product. The tissue sections were then mounted onto subbed slides and coverslipped with a mixture of PBS and glycerin. This double-staining procedure produced brown, granular labeling of retrogradely transported horse- radish peroxidase and blue labeling of neurotensin immunoreactivity .
Specific staining was abolished by preincubation of the primary antiserum with a 50 nM concentration of synthetic neurotensin (Peninsula) for 1 hour prior to the immunohis- tochemical procedure. Replacement of the primary anti- serum with normal rabbit serum did not produce specific staining.
Microscopy and data analysis A camera lucida attached to a Nikon Optiphot microscope
equipped with darkfield illumination was used for the map- ping of aortic afferents. These drawings were matched with darkfield photomicrographs of the same tissue sections to insure accuracy of localization. Transganglionically trans- ported horseradish peroxidase was identified as extracellu- lar grains, present in linear arrays or as diffuse reaction product. Endogenous peroxidase, present in lateral portions of the area postrema and medial portions of the NTS, was clearly distinguished on the basis of its smaller, more punc- tate-appearing reaction product.
In the double-staining experiments, the distribution and relationship of neurotensin-immunoreative cells and fibers to retrogradely labeled vagal motoneurons was plotted us- ing a drawing tube attached to an Optiphot equipped with brightfield illumination.
In order to compare the distribution of neurotensin to that of the afferent nerves, matched rostrocaudal sections from the aortic afferent experiments were aligned with those from the double-staining experiments using a microprojector.

NT IMMUNOHISTOCHEMISTRY OF THE NTS AND DVN 157
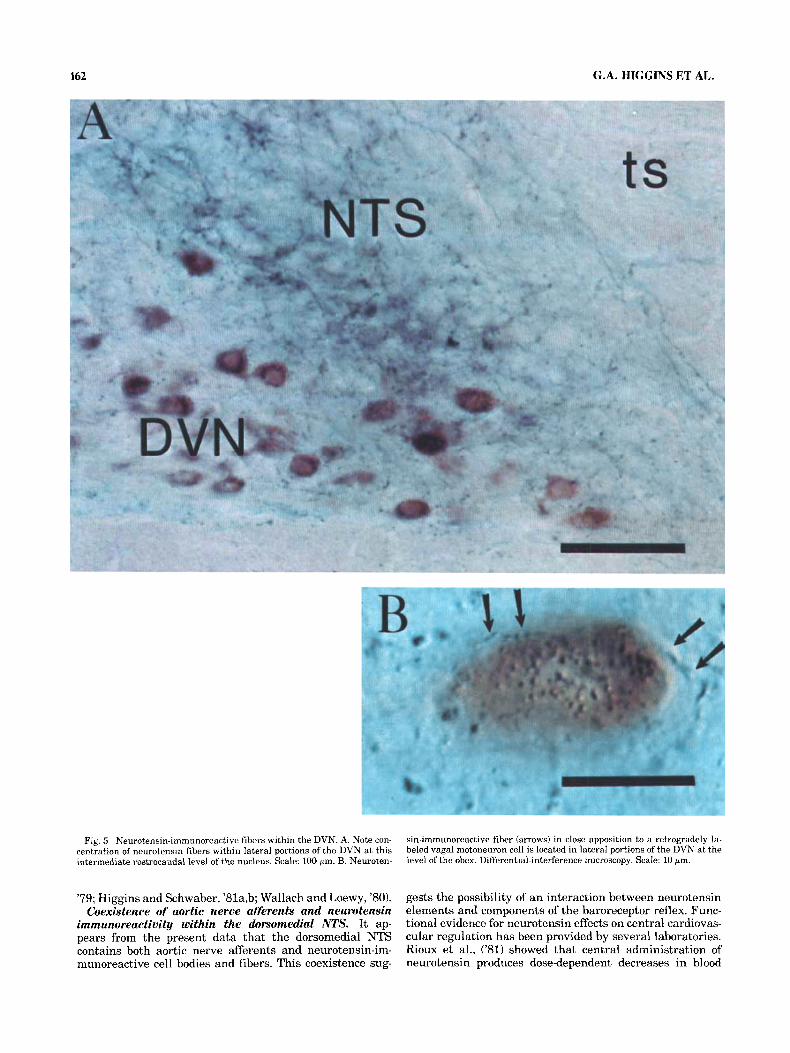
nucleus at the level of the obex (Fig. 5A), numerous stained fibers with varicosities could be observed in apposition to retrogradely labeled perikarya and primary dendritic pro- cesses (Fig. 5B). Although unequivocal identification of syn- aptic contacts depends upon electron microscopic exam- ination, these results demonstrate that neurotensin fibers are appropriately positioned to make direct contact with vagal motoneurons.
DISCUSSION By combining immunohistochemistry with retrograde la-
beling of the vagus nerve and transganglionic labeling of the aortic nerve, we have been able to demonstrate the close anatomical relationship of neurotensin-containing cells and fibers to vagal motoneurons and aortic afferents within the medulla. Retrogradely labeled preganglionic parasympathetic neurons of the DVN appear to be heavily innervated by neurotensin-immunoreactive fibers. The NTS contains neurotensin-immunoreactive cells and fibers which are located dorsomedial to the tractus solitarius, an area receiving aortic nerve afferents, and immunoreactive fi- bers, which are present in medial regions adjacent to the area postrema and in ventral regions lateral to the DVN.
The nucleus tractus of the solitarius (NTS) The most compelling evidence for the presence of a vis-
ceral topographic map within the NTS has come from trac- ing the central distribution of nerve branches arising from specific viscera using the transganglionic transport of horseradish peroxidase. Work from our laboratory and oth- ers in the rabbit and the cat have shown that cardiovascu- lar afferents terminate dorsally and medially within the nucleus (Berger, '79; Higgins and Schwaber, '81a,b; Kalia and Welles, '80a; Wallach and Loewy, '80), gastric afferents project to parvicellular portions of the nucleus adjacent to the area postrema (Leslie et al, '82) and pulmonary affer- ents end in ventrolateral regions (Kalia and Mesulam, '80b).
Central projections of the aortic nerve. In the present study we mapped the central distribution of the aortic branch of the vagus nerve in the rat in order to identify regions of the NTS that receive baroreceptor input. This nerve is easy to identify in the Wistar-Kyoto rat (Sapru and Krieger, '77), and, like the aortic depressor nerve in the rabbit, is probably composed entirely of baroreceptor affer- ents (Sapru et al., '81). Other baroreceptor afferents are conveyed in the carotid sinus and vagus nerves, which are mixed afferent nerves and thus are not as useful for ana- tomical studies of baroreceptor afferent projections. Our results confirm the recent report of Ciriello ('83).
We show here that the aortic nerve of the rat projects to commissural and medial portions of the NTS, and to a region of the nucleus which is located just dorsomedial to the tractus solitarius. This pattern is very similar to the
RESULTS Distribution of aortic nerve afferents within
the NTS Afferents from the right aortic branch of the vagus nerve
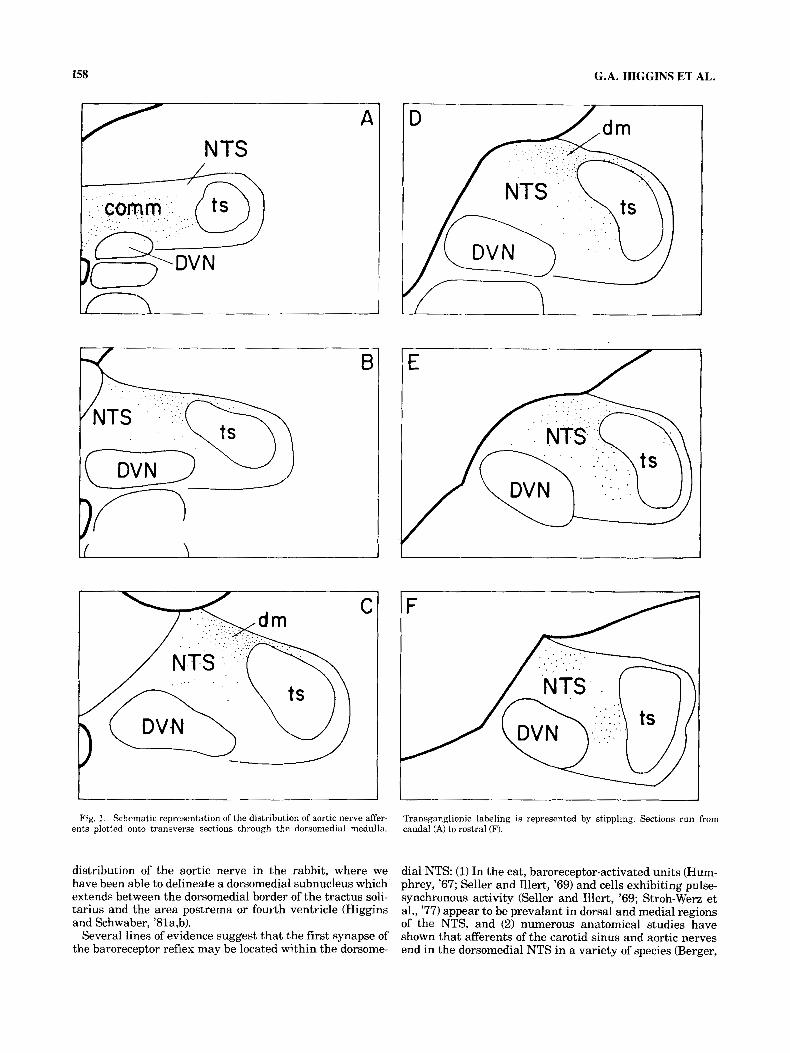
enter the rostral medulla and travel caudally within lateral portions of the TS. The commissural subdivision of the NTS receives an abundance of aortic fibers, and a few axons appear to decussate into the contralateral nucleus (Fig. 1A). The DVN also appears to receive a few fibers at these caudal levels. The aortic nerve projects most heavily to a region of the ipsilateral NTS which is located just dorso- medial to the TS at rostrocaudal levels surrounding the obex (Fig. 1B-D, Fig. 2). At these levels a few fibers also appear to be present within lateral portions of the nucleus (Fig. lB,C). Rostral to the obex, as the NTS begins to move laterally in the medulla, the innervation shifts to a more ventral position in the nucleus (Fig. lE,F).
Localization of neurotensin-immunoreactive cells and fibers within the NTS and DVN and their
relationship to vagal preganglionic motoneurons and afferents
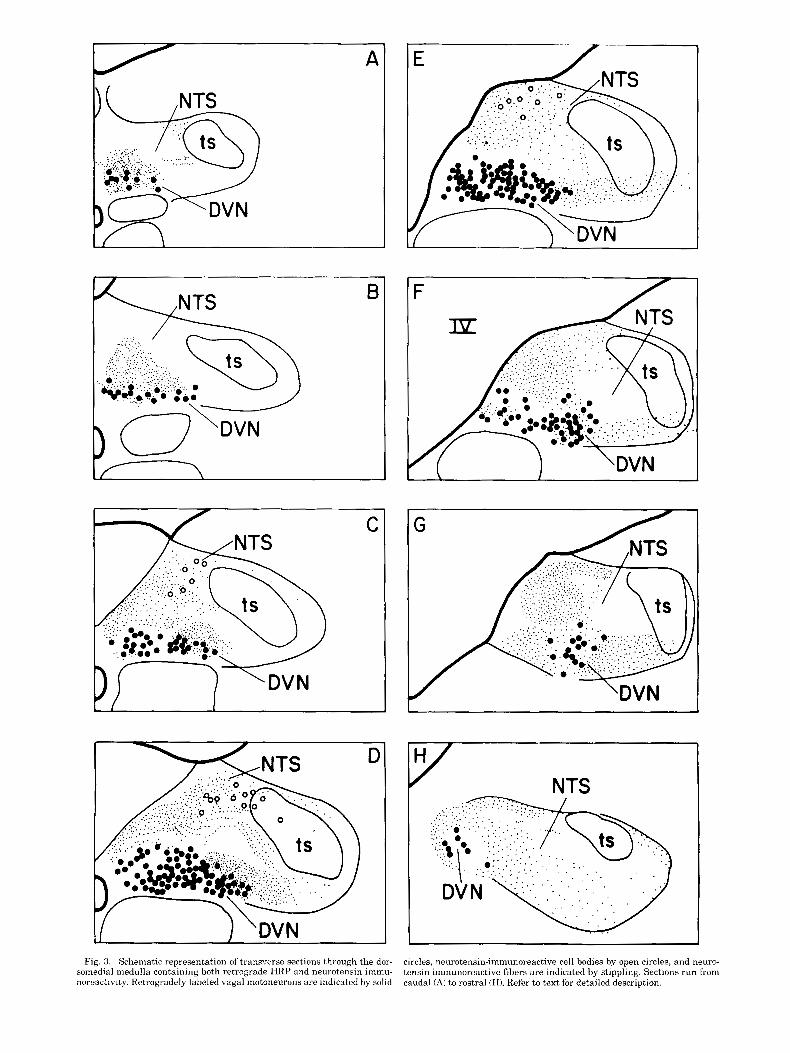
Neurotensin-immunoreactive fibers are present within the DVN as far caudally as the retrogradely labeled moto- neurons extend. At these caudal levels, the immunoreac- tive fibers are concentrated among the small cluster of DVN cells which are adjacent to the central canal (Fig. 3A). More rostrally, as the DVN expands laterally, the neuroten- sinergic fibers are located within medial portions of the DVN and dorsally, within the commissural subdivision of the NTS (Fig. 3B). At levels caudal to the obex and in which the caudal area postrema (AP) is present, neurotensin fi- bers are present throughout the DVN and extend into the medial NTS (Fig. 3C). Little or no immunoreactivity is found within lateral portions of the NTS at these levels.
Around the obex, neurotensin-containing fibers occupy the dorsomedial portion of the NTS and also extend into the most medial portions of the NTS adjacent to the AP (Figs. 3 C,D, 4). Immunoreactive fibers are also present in ventral portions of the NTS adjacent to the DVN. At these levels, the lateral portion of the DVN is heavily innervated but few fibers are present in other portions of the nucleus (Figs. 3-5).
Rostral to the obex, neurotensin-immunoreactive fibers are concentrated within lateral portions of the DVN and the lateral NTS ventral to the TS (Fig. 3E). Additional fibers are present within the NTS dorsal and medial to the DVN (Fig. 3F). Figure 3G,H shows a characteristic pattern of immnoreactive staining present within the most rostral aspects of the NTS and DVN. Fibers are located lateral and medial to retrogradely labeled cells of the DVN and in the medial part of the NTS.
At levels surrounding the obex, neurotensinergic cells are located in the same dorsomedial region which receives the bulk of the aortic nerve afferents (Figs. 2-4). Rostral to the obex, this cell group shifts to a more medial position within the nucleus (Fig. 3E). These neurons are observable without colchicine treatment; however, their numbers increase by approximately 50% with pretreatment.
The DVN was examined under high magnification ( x 1,000) differential-interference microscopy to determine whether neurotensin-immunoreactive fibers may make contacts with retrogradely labeled vagal motoneurons. In all portions of the DVN, especially in lateral portions of the
AP
comm dm DVN NCU NGR NTS ts rv
cc
Abbreviations
Area postrema Central canal Commissural nucleus of the NTS Dorsomedial nucleus of the NTS Dorsal motor nucleus of the vagus nerve Nucleus cuneatus Nucleus gracilis Nucleus of the tractus solitarius Tractus solitarius Fourth ventricle
158 G.A. HIGGINS ET AL.
A NTS
7-
\
C
Fig. 1. Schematic representation of the distribution of aortic nerve affer- ents plotted onto transverse sections through the dorsomedial medulla.
distribution of the aortic nerve in the rabbit, where we have been able to delineate a dorsomedial subnucleus which extends between the dorsomedial border of the tractus soli- tarius and the area postrema or fourth ventricle (Higgins and Schwaber, '81a,b).
Several lines of evidence suggest that the first synapse of the baroreceptor reflex may be located within the dorsome-
Transganglionic labeling is represented by stippling. Sections run from caudal (A) to rostra1 (F).
dial NTS: (1) In the cat, baroreceptor-activated units (Hum- phrey, '67; Seller and Illert, '69) and cells exhibiting pulse- synchronous activity (Seller and Illert, '69; Stroh-Werz et al., "77) appear to be prevalant in dorsal and medial regions of the NTS, and (2) numerous anatomical studies have shown that afferents of the carotid sinus and aortic nerves end in the dorsomedial NTS in a variety of species (Berger,
NT IMMUNOHISTOCHEMISTRY OF THE NTS AND DVN 159
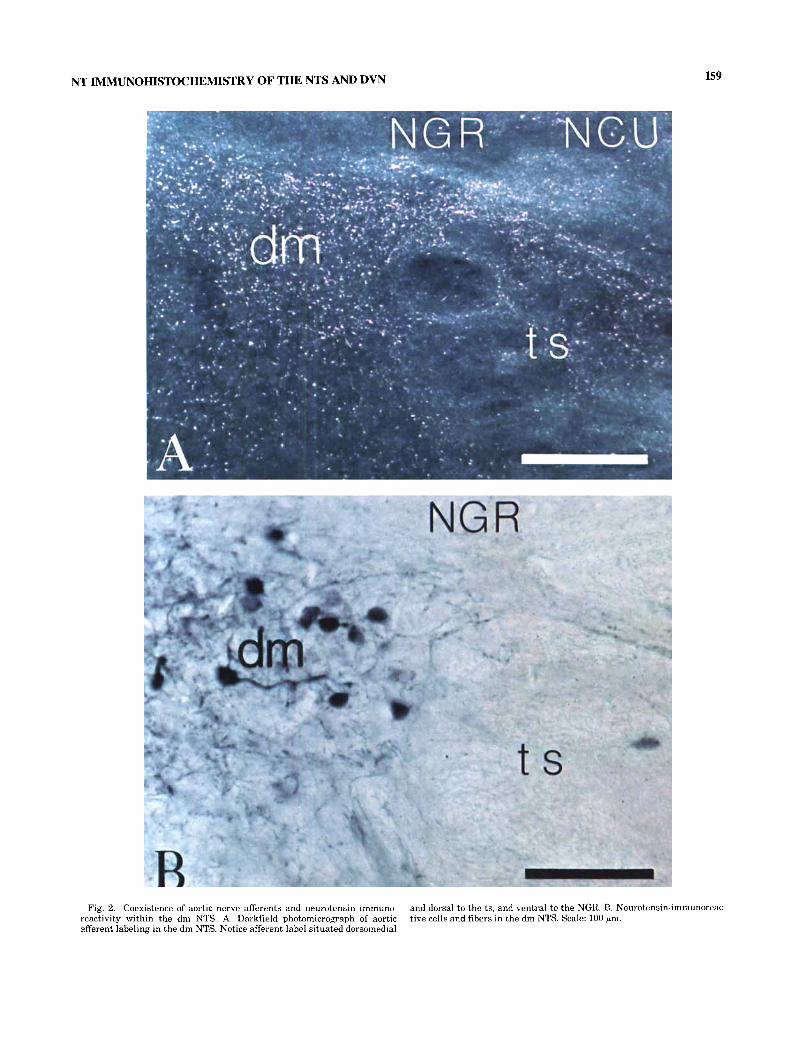
Fig. 2. Coexistence of aortic nerve afferents and neurotensin immuno- reactivity within the dm NTS. A. Darkfield photomicrograph of aortic afferent labeling in the dm NTS. Notice afferent label situated dorsomedial
and dorsal to the ts, and ventral to the NGII. B. Neurotensin-immunoreac- tive cells and fibers in the dm NTS. Scale: 100 pm.
A
B
C
E r
NTS /
Fig. 3. Schematic representation of trans.rerse sections through the dor- somedial medulla containing both retrograde HRP and neurotensin immu- noreactivity. RetrohTadely laheled vagal motoneurons are indicated by solid
circles, neurotensin-immunoreactive cell bodies by open circles, and neuro- tensin-immunoreactive fibers are indicated by stippling. Sections run from caudal (A) to rostra1 (H). Refer to text for detailed description.

Fig. 4. Photomicrographs of transverse sectmns through the NTS and DVN which have been processed for the presence of retrograde HRP, which yields a brown reaction product, and neurotensin immunoreactivity, which aDpears a s a blue reaction product.. A. Section a t level of obex; notice
neurotensin immunoreactive cells in dm NTS. B. More rostra1 level; notice region medial to the ts, indicated by the asterisk, which is devoid of neuro- tensin immunoreactivity. Scale: 100 pm.
162 G.A. HIGGINS ET AL.
Fig. 5 Neurotensin-immunoreactive fibers within the DVN. A. Note con- centration of neurotensin fibers within lateral portions of the DVN at this intermediate rostrocaudal level of the nucleus. Scale: 100 pm. B. Neuroten-
sin-immunoreactive fiber (arrows) in close apposition to a retrogradely la- beled vagal motoneuron cell is located in lateral portions of the DVN at the ievel of the obex. Differential-interference microscopy. Scale: 10 pm.
'79; Higgins and Schwaber, '81a,b; Wallach and Loewy, '80). Coexistence of aortic nerve afferents and neurotensin
immunoreactivity within the dorsomedial NTS. It ap- pears from the present data that the dorsomedial NTS contains both aortic nerve afferents and neurotensin-im- munoreactive cell bodies and fibers. This coexistence sug-
gests the possibility of an interaction between neurotensin elements and components of the baroreceptor reflex. Func- tional evidence for neurotensin effects on central cardiovas- cular regulation has been provided by several laboratories. Rioux et al., ('81) showed that central administration of neurotensin produces dose-dependent decreases in blood

NT IMMUNOHISTOCHEMISTRY OF THE NTS AND DVN 163
pressure in the pentobarbital-anesthetized rat. Alterna- tively, Sumners et al. ('81) observed blood pressure in- creases following i.c.v. administration of the peptide in the conscious rat.
A characteristic of the cardiovascular response produced by centrally administered neurotensin is the lack of direct or reflexive changes in heart rate. The absence of reflex heart rate responses following neurotensin-induced blood pressure changes raises the possibility that the peptide may suppress the cardiac component of the baroreceptor reflex, and this suppression may occur within the dorsomedial NTS. Inhibition of reflex bradycardia following a peptide- stimulated increase in blood pressure is a characteristic effect of other neuropeptides including met-enkephalin and bradykinin (Unger et al., '81).
The dorsal motor nucleus of the vagus nerve (DVN) The differential distribution of neurotensin fibers within
the DVN may reflect an underlying "viscerotopic" organi- zation of preganglionic neurons. Several authors (Getz and Sirnes, '49; Molhant, '10) have suggested such an organiza- tion based on retrograde degeneration experiments, but more recent studies using retrograde horseradish peroxi- dase transport have yielded equivocal results (Coil and Norgren, '79; Dennison, et al., '81; Kalia and Mesulam, '80b; Nosaka et al., '79; Todo et al., '77). However, electro- physiological studies have demonstrated the presence of vagal cardiomotor neurons in lateral portions of the DVN at the level of the obex in rabbit (Jordan et al., '82; Schwa- ber and Schneiderman, '75), dog (Hopkins et al., '82), and pigeon (Schwaber and Cohen, '78), an area which has been shown to contain neurotensin-immunoreactive fibers in the present study.
In light of the profound inhibition of gastric acid secretion produced by i.c.v. administration of neurotensin (Osumi et al., '781, it is tempting to speculate that the neurotensin- immunoreactive fibers which surround vagal motoneurons may, in part, be responsible for inhibition of vagally in- duced gastric acid secretion. Physiological investigations have clearly established that the DVN is responsible for the gastric acid secretion induced by intravenous adminis- tration of insulin (Kerr and Preshaw, '69). Additionally, centrally administered neurotensin prevents the develop- ment of stress-induced gastric ulcers in the rat (Nemeroff et al., '811, an action which is consistent with suppression of vagal secretomotor activity.
Neurotensin-immunoreactive fibers also appear to be con- centrated within regions of the NTS that receive gastric afferents (Leslie et al., '82). These regions of the NTS, ventral portions of the area postrema, and possibly also medial regions of the DVN caudal to the obex may contain binding sites for insulin as demonstrated by autoradio- graphic methods (Van Houten et al., '79). Therefore, a mor- phological correspondence may exist between gastric affer- ents, vagal gastric secretomotor neurons, and neurotensin fibers within medial portions of the NTS and adjacent DVN. This neural circuitry may mediate central and reflex con- trol of gastric acid secretion, with neurotensin acting both at early stages of the reflex within the NTS and a t the final motor output in the DVN. The existence of other neuropep- tides which influence gastric acid secretion within this re- gion (Fuxe et al., '79; Moody et al., '81; Palkovits, '81) lends credence to this hypothesis. However, further functional studies are needed to test the significance of this relationship.
The source of medullary neurotensin fibers is unknown. They may originate from neurotensinergic cells of the dor- somedial NTS, although in the present investigation we have not been able to observe a continuity between these cells and fibers located in other regions of the NTS and DVN. In this study we have not observed neurotensin- immunoreactive fiber staining within the tractus solitar- ius, suggesting that neurotensin fibers in the dorsomedial medulla do not originate from the vagus nerve. In immu- nohistochemical studies of neurons of the paraventricular nucleus of the hypothalamus which project to the NTSi DVN, Sawchenko and Swanson ('82) did not observe any cells which were neurotensin-immunoreactive. Similarly, we have not found neurotensin immunoreactivity within neurons of the central nucleus of the amygdala which proj- ect to the NTS and DVN (Higgins and Schwaber, '831, although we have frequently observed neurotensin-immu- noreactive neurons within the central nucleus adjacent to cells retrogradely labeled from the vagal nuclei (unpub- lished observations). Other autonomic structures which project to the NTS, e.g., the parabrachial nucleus, may provide the source of neurotensin innervation. Double-label studies are currently underway in our laboratory to iden- tify the origin of neurotensin fibers to the NTS and DVN.
The close association of neurotensin-containing struc- tures with parasympathetic regulatory nuclei is consistent with the "trophotrophic" profile of the peptide as one which is involved in the maintenance of visceral activity, hypoth- ermia, antinociception, muscle relaxation, and stress reduc- tion (Prange and Nemeroff, '82). In a recent review, Nemeroff et al, ('81) emphasize that "it is apparent that the anatomical distribution of neurotensin underlies many of the effects observed after neurotensin administration." In this context, it is interesting to speculate that the profound cardiovascular and gastric effects produced by central neu- rotensin may be mediated directly through vagal motoneu- rons or indirectly through vagal reflex mechanisms by the neurotensin circuitry present within defined portions of the NTS and the DVN.
ACKNOWLEDGMENTS We thank Judy Vanlare and Marie Moke for their expert
histological assistance and Tammie McHugh for her tire- less help in preparing the manuscript.
LITERATURE CITED Armstrong, D.M., V.M. Pickel, D.J. Reis, and R.J. Miller (1981) Immunocy-
tochemical localization of catecholamine synthesizing enzymes and n e u ~ ropeptides in area postrema and medial nucleus tractus solitarius of ra t brain. J . Comp. Neurol. 196:505-517.
Berger, A.J. (1979) Distribution of carotid sinus nerve afferent fibers to solitary tract nuclei of the cat using transganglionic transport of horse^ radish peroxidase. Neurosci. Lett. 14:153-158.
Ciriello, J. (1983) Brainstem projections of aortic baroreceptor afferents in the rat. Neurosci. Lett. 36:37-42.
Coil, J.D., and R. Norgren (1979) Cells of origin of motor axons in the subdiaphragmatic vagus of the rat. J. Auton. Nerv. Syst. 1:203-210.
Dennison, S.J., V.E. Merritt, M.H. Aprison, and D.L. Felton (1981) Redefi- nition of the location of the dorsal motor nucleus of the vagus in the rat . Brain Res. Bull. 6: 77-81.
Emson, P.C., M. Goedert, B. Williams, M. Ninkovic, and S.P. Hunt (1982) Neurotensin: Regional distribution, characterization and inactivation. In C.B. Nemeroff and A.J. Prange (eds): Neurotensin, A. Brain and Gastrointestinal Peptide. New York: Annals of the N.Y.A.S., Vol. 400, pp. 198-215.
Fuxe, K., P. Bolme, G. Jonsson, L.F. Agnati, M. Golstein, T. Hokfelt, R. Schwartz, and J. Engel (1979) On the cardiovascular role of noradrena- line, adrenaline and peptide-containing neuron systems in the brain. In

164 G.A. HIGGINS ET AL.
P. Meyer and H. Schmitt (eds): Nervous System and Hypertension Paris: Wiley-Flammarion.
Getz, B., and T. Sirnes (1949) The localization within the dorsal motor vagal neurons-an experimental investigation. J. Comp. Neurol. 90:95-110.
Helke, C.J., W. Goldman, and D.M. Jacobowitz (1980) Demonstration of substance P in aortic nerve afferent fibers by combined use of fluores- cent retrograde neuronal labeling and immunocytochemistry. Peptides 1:354-364.
Higgins, G.A., and J.S. Schwaber (1981aj Afferent organization of the nu- cleus tractus solitarius of the rabbit: Evidence for overlap of forebrain and primary afferent inputs. Anat. Rec. 199:144a.
Higgins, G.A., and J.S. Schwaber (1981b) Subnuclear organization and topography of forebrain inputs to the nucleus tractus solitarius in the rabbit. Neurosci. Abstr. 7:116.
Higgins, G.A., and J.S. Schwaber (1983) Somatostatinergic projections from the central nucleus of the aniygdala to the vagal nuclei. Peptides 4: 657-662.
Hoffman, G.E., S. Wray, and M. Goldstein (1982) Relationship of catechol- amines and LHRH: Light microscopic study. Brain Res. Bull. 9,417-430,
Hopkins, D.A., and J.A. Armour (1982) Medullary cells of origin of physio- logically identified cardiac nerves in the dog. E h i n Res. Bull. 8:359- 365.
Humphrey, D.R. (1967) Neuronal activity in the medulla oblongata of cat evoked by stimulation of the carotid sinus nerve. P. Kezdi (ed). In: Baroreceptors and Hypertension. Oxford: Pergamon, pp. 131-178,
Jennes, L., W.E. Stumpf, and P.W. Kalivas (1982) Neurotensin: Topograpb- ical distribution in ra t brain by immunohistocbemistry. J. Comp. Neu- ral. 210:211-224.
Jordan, D., M.E.M. Khalid, N. Schneiderman, and K.M. Spyer (1982) The locations and properties of preganglionic cardiomotor neurons in the rabbit. Pfliigers Arch. 395:224-250.
Kalia, M., and R.V. Welles (1980a) Brain stem project.ions ofthe aortic nerve in the cat: A study using tetraniethyl benzidine as the substrate for horseradish peroxidase. Brain Res. 188:523-553.
Kalia, M., and M.-M. Mesulam (1980b) Brain stem projections of sensory and motor components of the vagus complex in the cat. 11. Laryngeal, tracheobronchial, pulmonary, cardiac, and gastrointestinal branches. J . Comp. Neurol. 193:467-503.
Kerr, F.W.L. and R.M. Preshaw (1969) Secrctomotor function of the dorsal motor nucleus of the vagus. J. Physiol 205:405-415.
Leslie, R.A., D.G. Gwyn, and D.A. Hopkins (1982) The central distribution of the cervical vagus nerve and gastic afferent and efferent projections in the rat. Brain Res. Bull. 8:37-43.
Loewy, A.D., and H. Burton (1973 Nuclei of the solitary tract: Efferent projections to the lower brain stem and spinal cord of the cat. J. Comp. Neurol. 181:421-450.
Maley, B., and R. Elde (1982) Immunohistochemical localization of putative neurotransmitters within the feline nucleus tractus solitarius. Neuro- science 79469-2490.
Molhant, M. (1910) Les connexions anatomiques et la valeui functionelle du noyau dorsal du vague. Nevraxe 12221-236.
Moody, T.W., T.1,. O'Donohue, and D.M. Jacobowitz (1981) Biochemical lo- ealization and characterization of bombesin-like peptides in discrete regions of ra t brain. Peptides 275-79.
Nemeroff, C.B., D. Luttinger, and A.J. Prange (1981) Neurotensin and bombesin. In L.L. Iverson, S.D. Iverson, and S.H. Snyder (eds): The Handbook of Psychopharmacology. New York: Pknum, pp. 24-64.
Nosaka, S., T. Yamamoto, and K. Yasunga (1979) Localization of vagal preganglionic neurons within rat brain stem. J. Comp. Neurol. 186:79- 92.
Osumi, Y., L. Nagasaka, H. Wang, and M. Fujiwara (1978) Inhibition of gastric acid secretion and mucosal blood flow induced by intraventricu- larly applied neurotensin in rats. Life Sci. 23:2275-2280.
Palkovits, M. (1981) Neuropeptides and biogenic amines in central cardio- vascular control mechanisms. In J.P. Buckley and C.M. Ferrario (eds): Central Nervous System Mechanisms in Hypertension. New York: Raven Press, pp. 73-87.
Prange, A.J., and C.B. Nemeroff (1982) The manifold actions of neurotensin: A first synthesis. In C.B. Nemeroff and A.J. Prange (edsj: Neurotensin, A Brain and Gastrointestinal Peptide. New York: Annals of the N.Y.A.S., Vol. 400, pp. 198-215.
Quirion, R., F. Rioux, S. %Pierre, F. Belanger, F.B. Joligoeur, and A. Barbeau (1981) Hypotensive effects of centrally and peripherally admin- istered neurotensin and neurotensin derivatives in rats. Neuropeptides 1:253-259.
Rioux, F., R. Quirion, S. St-Pierre, D. Regoui, F.B. Jolicoeur, F. Belanger, and A. Barbeau (1981) The hypotensive effects of centrally administered neurotensin in rats. Eur. J . Pharmacol. 69:241-247.
Sapru, H.N., E. Gonzalez, and A.J. Krieger (1981) Aortic nerve stimulation in the rat: Cardiovascular and respiratory responses. Brain Res. Bull. 6:393-398.
Sapru, H.N., and A.J. Krieger (1977) Carotid and aortic chemoreceptor function in the rat . J. Appl. Physiol. 42:344-348.
Sawchenko, P.E., and L.W. Swanson (1982) Immunohistochemical identifi- cation of neurons in the paraventricular nucleus that project to the medulla or to the spinal cord in the rat . J. Comp. Neurol. 105: 260-272.
Schwaher, J.S., and D.H. Cohen (1978) Field potential and single unit analysis of the avian dorsal motor nucleus of the vagus and criteria for identifying vagal cardiac cells of origin. Brain Res. f47: 79-90.
Schwaber. J., and N. Schneiderman (1975) Aortic nerve-activated cardioin- hibitory neurons and interneurons. Am. J. Physiol. 229:783-789.
Seller, H., and M. Illert (1969) The localization of the first synapse in the carotid sinus baroreceptor reflex pathway and its alterations of the afferent input. Pfliigers Arch. 306:l-19.
Stroh-Werz, M., P. Langhorst, andd H. Camerer (1977) Neuronal activity in the nucleus of the solitary tract in cats and dogs. 11. Activity modulation in relation to the respiratory cycle. Brain Res. 133%-93.
Sumners, C., M.I. Phillips, andd A. Camacho (1981) Central pressor and peripheral depressor effects of neurotensin in conscious rats. SOC. Neu- rosci. Abstr. 7.505.
Todo, K., T. Yamamoto, H. Satomi, H. Ise, H. Takatama, and K. Takahashi (1977) Origins of vagal preganglionic fibers to the sinio-atrial and atrio- ventricular node regions in the cat heart as studied by the horseradish peroxidase method. Brain Res. 136:545-550.
Uhl, G.R., R.R. Goodman, and S.H. Snyder (1979) Neurotensin-con- taining cell bodies, fibers and nerve terminals in the brain stem of the rat: Immunohistochemical staining. Brain Res. 167:77-91.
Unger, T, R.W. Rockhold, T. Yukimura, R. Rettig, W. Rascher, and D. Ganten (1981) Role of kinins and substance P in the central blood pressure regulation of normotensive and spontaneously hypertensive rats. In J.P. Buckley and C.M. Ferrario (eds): Central Nervous System Mechanisms in Hypertension. New York: Raven Press, pp. 115-127.
Van Houten, M., B.T. Posner, B.M. Kopriwa, and J.R. Brawer (1979) Insulin- binding sites in the r a t brain: In uiuo localization to the circumventri- cular organs by quantitative radioautography. Endocrinology 105:666- 673.
Wallach, J.H., and A.D. Loewy (1980) Projections of the aortic nerve to the nucleus tractus solitarius in the rabbit. Brain Res. 188:247-251.
Young, W.S., 111, and M. J . Kuhar (1979) Neurotensin receptors: Autoradio- gaphic localization in the rat CNS. Eur. J. Pharmacol. 59:161-163.




















