
Neurotensin and neuromedin N undergo distinct catabolic processes in murine astrocytes and primary...
10
Eur. J. Biochem. 221,297-306 (1994) 0 FEBS 1994 Neurotensin and neuromedin N undergo distinct catabolic processes in murine astrocytes and primary cultured neurons Bruno VINCENT, Jean-Pierre VINCENT and FrCdCric CHECLER Institut de Pharmacologie Moltculaire et Cellulaire, UPR 411, CNRS, UniversitC de Nice Sophia Antipolis, Valbonne, France (Received November 30, 1993Februaq 11, 1994) - EJB 93 1766/4 We examined the occurence of various endopeptidases and exopeptidases and their subcellular partition within soluble and membrane-associated compartments of 15-day-old astrocytes and 4-day-old primary cultured neurons. Peptidases were monitored with chromogenic or fluorimetric substrates and identified by means of specific inhibitors. We assessed the contribution of these peptidases in the catabolism of two related neuropeptides, neurotensin and neuromedin N. Metabo- lites were separated by HPLC and the identity of the proteolytic activities involved in their formation was established using specific inhibitors. Neuromedin N and neurotensin undergo both quantitative and qualitative differential proteolysis. Initial maximal rates of neuromedin N degradation were higher than those of neurotensin in both cell types. Furthermore, the two peptides were inactivated much more rapidly by the soluble than by the membrane-associated fractions prepared from both cell cultures. Neuromedin N was rapidly broken down by an aminopeptidase M/leucine aminopepti- dase attack, leading to the functionally silent Des-Lysl-neuromedin N metabolite. In the astrocytic membrane-associated fraction, neuromedin N underwent an additional minor endoproteolytic cleav- age at the Pro3-Tyr4 bond elicited by endopeptidase 24.11, as suggested by the protective effect of its blocking agent phosphoramidon. Unlike neuromedin N, neurotensin totally resisted hydrolysis by aminopeptidases. Primary inactivating cleavages detected in both cell types appeared mainly located at the Arg8-Arg9 and ProlO-Tyrll bonds, leading to the formations of neurotensin-(1 - 8) and neurotensin-( 1 - 10) as the major biologically inactive neurotensin catabolites. Endopeptidase 24.15 appeared mainly responsible for neurotensin-( 1 - 8) formation by the soluble fraction of neu- rons and astrocytes. In contrast, endopeptidase 24.16 was involved in neurotensin-(1 - 10) formation by both soluble and membrane-associated fractions of the two cell types. An additional cleavage leading to neurotensin-(1 - 11) formation and ascribed to endopeptidase 24.11 was detected mainly in the membrane-associated fraction from astrocytes. Finally, the secondary processing of neuroten- sin degradation products indicated that: (a) neurotensin-(1 - 11) was converted into neurotensin-(1 - 8) in the membrane fraction prepared from astrocytes; (b) neurotensin-(1 - 10) was transformed into neurotensin-(1-8) by an unidentified peptidase belonging to the class of metalloenzymes. The significance of distinct quantitative and qualitative catabolic fates of neuromedin N and neurotensin in cultured astrocytes and neurons is discussed. The cloning of the canine preproneurotensin precursor revealed that it encoded not only one copy of neurotensin but also one copy of neuromedin N (Dobner et al., 1987). The latter peptide that belonged to the neurotensin family was located besides neurotensin, at the C-terminal end of the pre- cursor (Dobner et al., 1987). The elucidation of the gene structure revealed that both peptides were encoded by the fourth exon (Kislauskis et al., 1988). Radioimmunoassays carried out with specific antibodies directed towards each peptide and devoid of cross-reactivity indicated that a virtu- ally constant ratio existed between the recoveries of neuro- medin N-like and neurotensin-like immunoreactivities in var- Correspondence to F. Checler Phone: +33 93 95 77 60. Fax: +33 93 95 77 08. Abbreviations. AMC, 7 amido-4-methylcoumarin; Hip, hippuric acid; CPP, carboxy-2-phenylpropyl ; pAB, p-aminobenzoate; Glut, glutaryl ; Z, benzyloxycarbonyl ; Mcc, 7-methoxy coumarin-3 ; Dnp, dinitrophenyl ious brain nuclei (De Nadai et al., 1989). It seemed, there- fore, unlikely that, at least in the central nervous system, differential maturation processes could lead to region-spe- cific variations of the neurotensinheuromedin N concentra- tion ratio. Thus, the neuromodulatory properties of the two peptides could be under the control of subsequent events such as metabolic processes that could modulate their life- times. In this respect, it is noticeable that the structure of neuromedin N renders it susceptible to rapid degradation by aminopeptidases (since its N-terminal residue is not blocked) while the pyroglutamate group of neurotensin theoretically precludes such a possibility. These structural features ex- plained our previous data demonstrating that neurotensin and neuromedin N underwent differential catabolic processes by rat brain membranes (Checler et al., 1991; Checler et al., 1986b) and by neuronal and plasma membranes from dog ileum mucosa (Barelli et al., 1993). Our previous work established that endopeptidase 24.16 was the only peptidase that always contributed to neurotensin
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Neurotensin and neuromedin N undergo distinct catabolic processes in murine astrocytes and primary...

Eur. J . Biochem. 221,297-306 (1994) 0 FEBS 1994
Neurotensin and neuromedin N undergo distinct catabolic processes in murine astrocytes and primary cultured neurons Bruno VINCENT, Jean-Pierre VINCENT and FrCdCric CHECLER Institut de Pharmacologie Moltculaire et Cellulaire, UPR 411, CNRS, UniversitC de Nice Sophia Antipolis, Valbonne, France
(Received November 30, 1993Februaq 11, 1994) - EJB 93 1766/4
We examined the occurence of various endopeptidases and exopeptidases and their subcellular partition within soluble and membrane-associated compartments of 15-day-old astrocytes and 4-day-old primary cultured neurons. Peptidases were monitored with chromogenic or fluorimetric substrates and identified by means of specific inhibitors. We assessed the contribution of these peptidases in the catabolism of two related neuropeptides, neurotensin and neuromedin N. Metabo- lites were separated by HPLC and the identity of the proteolytic activities involved in their formation was established using specific inhibitors. Neuromedin N and neurotensin undergo both quantitative and qualitative differential proteolysis. Initial maximal rates of neuromedin N degradation were higher than those of neurotensin in both cell types. Furthermore, the two peptides were inactivated much more rapidly by the soluble than by the membrane-associated fractions prepared from both cell cultures. Neuromedin N was rapidly broken down by an aminopeptidase M/leucine aminopepti- dase attack, leading to the functionally silent Des-Lysl-neuromedin N metabolite. In the astrocytic membrane-associated fraction, neuromedin N underwent an additional minor endoproteolytic cleav- age at the Pro3-Tyr4 bond elicited by endopeptidase 24.11, as suggested by the protective effect of its blocking agent phosphoramidon. Unlike neuromedin N, neurotensin totally resisted hydrolysis by aminopeptidases. Primary inactivating cleavages detected in both cell types appeared mainly located at the Arg8-Arg9 and ProlO-Tyrll bonds, leading to the formations of neurotensin-(1 - 8) and neurotensin-( 1 - 10) as the major biologically inactive neurotensin catabolites. Endopeptidase 24.15 appeared mainly responsible for neurotensin-( 1 - 8) formation by the soluble fraction of neu- rons and astrocytes. In contrast, endopeptidase 24.16 was involved in neurotensin-(1 - 10) formation by both soluble and membrane-associated fractions of the two cell types. An additional cleavage leading to neurotensin-(1 - 11) formation and ascribed to endopeptidase 24.11 was detected mainly in the membrane-associated fraction from astrocytes. Finally, the secondary processing of neuroten- sin degradation products indicated that: (a) neurotensin-(1 - 11) was converted into neurotensin-(1 - 8) in the membrane fraction prepared from astrocytes; (b) neurotensin-(1 - 10) was transformed into neurotensin-(1-8) by an unidentified peptidase belonging to the class of metalloenzymes. The significance of distinct quantitative and qualitative catabolic fates of neuromedin N and neurotensin in cultured astrocytes and neurons is discussed.
The cloning of the canine preproneurotensin precursor revealed that it encoded not only one copy of neurotensin but also one copy of neuromedin N (Dobner et al., 1987). The latter peptide that belonged to the neurotensin family was located besides neurotensin, at the C-terminal end of the pre- cursor (Dobner et al., 1987). The elucidation of the gene structure revealed that both peptides were encoded by the fourth exon (Kislauskis et al., 1988). Radioimmunoassays carried out with specific antibodies directed towards each peptide and devoid of cross-reactivity indicated that a virtu- ally constant ratio existed between the recoveries of neuro- medin N-like and neurotensin-like immunoreactivities in var-
Correspondence to F. Checler Phone: +33 93 95 77 60. Fax: +33 93 95 77 08. Abbreviations. AMC, 7 amido-4-methylcoumarin; Hip, hippuric
acid; CPP, carboxy-2-phenylpropyl ; pAB, p-aminobenzoate; Glut, glutaryl ; Z, benzyloxycarbonyl ; Mcc, 7-methoxy coumarin-3 ; Dnp, dinitrophenyl
ious brain nuclei (De Nadai et al., 1989). It seemed, there- fore, unlikely that, at least in the central nervous system, differential maturation processes could lead to region-spe- cific variations of the neurotensinheuromedin N concentra- tion ratio. Thus, the neuromodulatory properties of the two peptides could be under the control of subsequent events such as metabolic processes that could modulate their life- times. In this respect, it is noticeable that the structure of neuromedin N renders it susceptible to rapid degradation by aminopeptidases (since its N-terminal residue is not blocked) while the pyroglutamate group of neurotensin theoretically precludes such a possibility. These structural features ex- plained our previous data demonstrating that neurotensin and neuromedin N underwent differential catabolic processes by rat brain membranes (Checler et al., 1991; Checler et al., 1986b) and by neuronal and plasma membranes from dog ileum mucosa (Barelli et al., 1993).
Our previous work established that endopeptidase 24.16 was the only peptidase that always contributed to neurotensin

298
proteolysis in cell models and membrane preparations from neural and extraneural origins (Checler et al., 1988). This enzyme was recently shown at the light and electronic micro- scopic levels to be associated with neurons and astrocytes in adult rat mesencephalon (Woulfe et al., 1992). The present study was designed to establish the contribution of endopep- tidase 24.16 in neurotensin degradation in astrocytes and primary-cultured neurons. Furthermore, we fully charac- terized the proteolytic events (i.e the peptidases involved in the primary inactivating and secondary cleavages) responsi- ble for the catabolism of neurotensin and neuromedin N in the two cell types in order to assess whether quantitative or qualitative differences also took place in cultured neurons and astrocytes.
MATERIALS AND METHODS
Material Amastatin { [(2S, 3R)-3-amino-2-hydroxy-5-methyl-hexa-
noyll-valyl-valyl-aspartate} ; Arg-7-amido-4-methylcouma- rin (arginyl-7AMC), bestatin ((2S, 3R)-3 amino 2-hydroxy- 4-phenyl-butanoyl leucine), cytosine P-D-arabino-furanoside, leucine aminopeptidase (microsomale type 4), Gly-Pro- 7 AMC (glycyl-prolyl-7AMC), Hip-His-Leu (hippuryl-hist- idyl-leucine), Leu-7AMC (leucyl-7-AMC), phosphoramidon (N-(a-rhamno-pyranosyloxy-hydroxyphosphinyl)leucyl-tryp- tophan) and polylysine, were from Sigma. Benzyloxycarbon- yl-Gly-Pro-7AMC (Z-glycyl-prolyl-7-AMC), QFS (7-me- thoxy ~-coumar~n-pro~y~-~eucy~-g~ycy~-pro~y~-~-~ys~ne-d~n~- trophenyl), actinonin, diprotin A were from Novabiochem. Arphamenine B (2R, 5S)-5-amino-8-guanido-4-0~0-2-para- hydroxyphenylmethylethanoic acid was from Interchim. Fe- tal calf serum was from Boehringer. HAM-F,, medium was from GIBCO BRL. Neurotensin was from Neosystem. Z - Pro-prolinal (Z-prolyl prolinal), benzoyl-Gly-Ala-Ala-Phe- para-aminobenzoate (Bz-glycyl-alanyl-alanyl-phenylalanyl- pAB) and CPP-Ala-Ala-Tyr-pAB { N-[ 1 (R,S)-carboxyl-3- phenyl-propyll-alanyl-alanyl-tyrosyl-pAB} were generously given by Drs S. Wilk and M. Orlowski (Mount Sinai School of Medicine, New York). Captopril (2-D-methyl-3-mercapto- propanoyl-L-proline) was kindly given by Dr B. P. Roques (UniversitC Rent Descartes, Paris). Phosphodiepryl 03 (N- (phenylethylphosphony1)glycyl-prolyl-hexanoic acid) was synthetized and generously provided by Dr V. Dive (CEN Saclay, Gif/Yvette).
Cell cultures Primary cultures of neurons were prepared from cerebral
hemispheres of 14-day-old mouse embryos as previously de- scribed (Checler et al., 1986a). Briefly, cells were mechani- cally dissociated with a pipette in HAM F12 medium (Gibco) supplemented with 0.6% glucose and 10% fetal calf serum (Boehringer Manheim). Dissociated cells were then plated at a density of 3X lo6 cells in 35-mm plastic tissue-culture dishes precoated with polylysine (10 pg/ml) and grown in a humidified atmosphere of 5% C02, 95% air. At the begin- ning of the third day and every two days thereafter, cytosine arabinofuranoside (10 pM) was added in the medium for 24 h to prevent glial cell proliferation. According to this pro- cedure, we previously showed that growth cones could be distinguished as early as 1 - 3 hours after seeding. Until 24 hours, processes grew in length and number, then constituted a network of interconnecting neurites over 1-4 days after
plating (Chabry et a]., 1990). Neuronal cultures appeared vir- tually devoid of glial fibrillary acidic protein immunoreactiv- ity, a specific astrocytic marker.
Cultures of astrocytes were obtained from primary cul- tured neurons which were not treated with cytosine arabino- furanoside but maintained in a 10% fetal calf serum medium in order to prevent oligodendrocytes expansion. Astrocytes were used after 15 days in culture. At this time, immunola- belling with antibodies directed against GFAP revealed that more than 95% of the cells indeed corresponded to genuine astrocytes. These cultures did not display galactocerebroside imunoreactivity (an oligodendrocyte marker ; Coger, Paris), and were not labelled after exposure to an antiserum that recognizes a 160-kDa protein present in the membrane of macrophage (a marker of microglia; Serotec, Paris)
Preparation of cell homogenates and subcellular fractions Four-day-old primary cultured neurons and 15-day-old
astrocytes were washed twice with 8.5 mM sodium phos- phate, 140 mM NaC1, pH 7.5, then scraped in 5 mM Tris/ HCl, pH 7.5 and homogenized with a syringe. Homogenates were centrifuged (4"C, 35 min, 150000 g) and supernatants (soluble fractions) were collected. The pellets, referred to as membrane-associated fractions, were resuspended in 5 mM Tris/HCl, pH 7.5, and concentrations of proteins were ad- justed by dilution in the same buffer. The pellet displayed about 7% total lactate deshydrogenase activity that was mainly recovered in the soluble fraction (not shown).
Degradation experiments and HPLC analysis Peptides (2 nmol) were incubated for various times at
37°C in a final volume of 100 p1 5 mM Tris/HCl, pH 7.5, containing a final protein concentration of 0.05 -0.1 mg/ml, in the absence or in the presence of specific peptidase inhibi- tors (incubated with the enzymic source for 10 min at 37 "C). Incubations were stopped by acidification (10 pl, 1 M HCl) and centrifuged for 5 min at 13 000 g, then supernatants were submitted to HPLC analysis. Samples were applied to a re- verse-phase C,, lichrosorb column (Merck). Elutions were carried out at room temperature by means of a 42-min linear gradient of 0.1 % trifluoroacetic acid, 0.05 % triethylamine/ 0.1 % trifluoroacetic acid, 0.05 % triethylamine in acetonitrile for 90-10 (by vol.) to 60-40 (by vol.) at a flow rate of 1 ml/min as described (Barelli et al., 1991). Absorbance was monitored at 230 nm with a detector setting of 0.1 full scale.
Degradation of neuromedin N by purified endopeptidase 24.11
Neuromedin N (5 nmol) was incubated for 1 hour at 37°C in a final volume of 100 pl 5 mM Tris/HCI, pH 7.5, containing 0.5 pg purified endopeptidase 24.11 in absence or in the presence of 1 pM phosphoramidon. After acidification, the samples were submitted to HPLC analysis as described above.
Degradation of neurotensin 1-11 by purified endopeptidase 24.16
Neurotensin-(I - 11) (2 nmol) was incubated at 37°C for 5 hours with purified rat brain endopeptidase 24-16 (Checler et al., 1986c) in a final volume of 100 p1 5 mM Tris/HCl, pH 7.5, in the absence (control) or in the presence of various

299
Table 1. Peptidase contents in homogenates of astrocytes and primary-cultured neurons. Activities were measured as described in Materials and Methods by incubating the indicated substrates with homogenates of 4-day-old cultured neurons or 15-day-old cultured astrocytes. Inhibition studies were carried out by incubating the homogenates with the indicated specific inhibitors as described in Materials and Methods. Values which corresponded to the inhibitory-sensitive fractions of peptidase activities were the means +- SEM from 5-9 independent determinations. Concentrations of inhibitors were as follows : Cpp-Ala-Ala-Tyr-pAB, phosphoramidon, Z-pro-prolinal, capto- pnl, 10 pM; arphamenine B, 0.5 pM; bestatin, 50 pM; diprotin A, 10 pM; Pro-Ile, 10 mM. QFS = Mcc-Pro-Leu-Gly-Pro-D-Lys-Dnp; LAP, leucine aminopeptidase
Activity Substrate/Inhibitor Specific activity
neurons astrocytes
Endopeptidase 24.16 Endopeptidase 24.15 Endopeptidase 24.11 Proline endopeptidase Angiotensin-converting enzyme Post-proline dipeptidyl aminopeptidase (DAP IV) Aminopeptidase B Aminopeptidase MLAP
QFS/Pro-Ile Bz-Gly- Ala- Ala-Phe-p AB/CPP- Ala- Ala-Tyr-p AB Glut-Ala-Ala-Phe-pAB/phosphoramidon Z-Gly-Pro-7 AMCIZ-pro-prolinal Hip-His-Leu/captopril Gly-Pro-7AMC/diprotin A Arg-7 AMClarphamenine B Leu-7 AMChestatin
specific inhibitors. Peptides were separated by HPLC as de- scribed above.
Fluorimetric and chromogenic peptidase assays All the fluorimetric or chromogenic substrates were incu-
bated for various times at 37°C in a final volume of 100 pl 5 mM Tris/HCl, pH 7.5, containing a final concentration of 1-20 pg/ml protein, in the absence or presence of the corre- sponding specific inhibitor and quantifications were carried out as previously described (Dauch et al., 1993). We clearly established that, under these experimental conditions, even in a crude enzymic fraction, it was possible to measure a single peptidase activity (Dauch et al., 1993).
Protein concentration Protein concentrations were determined by the Bradford
method according to the manufacturer's recommendations with white egg lysozyme as the standard.
RESULTS Peptidases in astrocytes and neurons
Five endopeptidases and three exopeptidases were quan- tified in the whole homogenate of astrocytes and 4-day-old primary cultured neurons (Table l), and their partition be- tween soluble and membrane-associated compartments was examined (Fig. 1). Specific activities of proline endopepti- dase, aminopeptidase B and endopeptidase 24.11 were 4-5- fold higher in astrocytes than in neurons (Table 1). In both cell types, endopeptidase 24.11 was fully recovered in the membrane-associated fraction while proline endopeptidase and APB appeared mainly cytosoluble (Fig. 1) in agreement with the previously reported physicochemical properties of these three peptidases (for review see Checler, 1993). Dipep- tidylaminopeptidase IV and aminopeptidase M display simi- lar specific activities in astrocytes and neurons (Table 1) and were recovered in both soluble and membrane-associated fractions (Fig. 1) as expected from previous studies reporting
nmol . h-' . mg protein-'
1 9 t 2 51' 2 5 3 ? 5 2 8 2 6 6 2 2
1 7 t 5 1 6 8 % 5 73 t 10
4 0 2 2 2 1 t 3
245 +- 23 154 t 13 2 8 % 6 1 1 2 6
6 4 0 2 4 107 -t 10
Percent of total activity 100 50 0 0 50 100
A B
Fig. 1. Distribution of peptidases in soluble and membrane-asso- ciated fractions of astrocytes and primary-cultured neurons. Activities were measured with the substrates and inhibitors indicated in Table 1 with soluble (open bars) and membrane-associated (black bars) fractions of 4-day-old cultured neurons (A) or 15 day-old cul- tured astrocytes (B) prepared as described in Materials and Methods. Values are the means t SEM of four independent determinations and are expressed as the percent of total activity (soluble + mem- brane-associated activities) taken as 100. AP, aminopeptidase ; LAP, leucine aminopeptidase; DAP IV, post-proline dipeptidyl aminopep- tidase; E, endopeptidase ; PE, proline endopeptidase; ACE, angio- tensin-converting enzyme.
on their dual subcellular localization (Checler, 1993). It was likely that the soluble form of aminopeptidase M indeed rep- resented leucine aminopeptidase (Checler, 1993). Endopepti- dase 24.15 was more abundant in neurons than in astrocytes while the opposite situation was observed for endopeptidase 24.16 (Table 1). Unlike endopeptidase 24.15 that mainly be- haved as a soluble protein, endopeptidase 24.16 equally par-

300
Table 2. Maximal initial rates and half-lives of neuromedin N and neurotensin degradation by astrocytes and primary-cultured neurons. 2 nmol peptides were incubated for various times at 37 "C with homogenates, soluble and membrane-associated fractions of 4- day-old cultured neurons and 15-day-old cultured astrocytes as described in Materials and Methods. Incubations were stopped, the samples centrifuged and analyzed by HPLC as described in Materials and Methods. Maximal initial rates and half-lives were calculated from the linear parts of the disappearance curves of the peptides. Values were the means of four independent determinations.
Peptide Degradation rate Half-lives in homogenate
soluble fraction membrane-associated fraction
nmol . h.-' . mg protein-' min
Neuromedin N neurons 141 t 10 astrocytes 502 t 41
Neurotensin neurons 8 1 t 6 astrocytes 239 t 28
18 t 4 7 ? 1
38 t 6 17 +- 2
154 t 26 3 1 t 4
348 t 68 85 -C 13
loo r .- % a 8 0 1
0 1 2 3 4 5
Time in culture (day)
Fig.2. Degradation rates of neurotensin and neuromedin N at various differentiation times of primary-cultured neurons. Neu- rotensin and neuromedin N (2 nmol, 20 pM) were incubated with homogenates of primary cultured neurons (10 pg, 1 hour) obtained after the indicated times in culture. Incubations were acidified, the samples centrifuged and analyzed by HPLC as described in the Ma- terials and Methods. Values correspond to intact remaining peptides after HPLC and are the means of two independent determinations.
titioned between membrane-associated and soluble fractions in the two cell types (Fig. 1). Finally, angiotensin-converting enzyme was barely detectable in pure cultured neurons (Table 1). In astrocytes, the low enzyme activity was mostly detected in the membrane-associated compartment (Fig. 1).
A NN
NEURONS
B
. Des-Lysl NN
f Des-Lys' "
DecLysl NN ASTROCYTES
Des-Lvsl NN
Quantitative aspects of neurotensin and neuromedin N degradation in astrocytes and primary cultured neurons
Half-lives and maximal degradation rates of neurotensin and neuromedin N by homogenates of astrocytes and 4-day- old primary cultured neurons were deduced from HPLC analyses. Table 2 indicated that: (a) degradation rates of both peptides were about threefold higher in astrocytes than in neurons; (b) neuromedin N was more efficiently degraded than neurotensin by both cell types; (c) in the soluble fraction prepared from both cell cultures, neuromedin N half-lives were about twofold shorter than those of neurotensin; (d) The half-lives of neurotensin and neuromedin N were about 5- to %fold longer in the membrane-associated than in the soluble fractions in both cell types.
It was interesting to note that the degradation rates of neurotensin and neuromedin N appeared virtually identical at various times of the neuronal differentiation process (Fig. 2). This seemed to indicate that the peptidase activities involved
I I I I I
0 10 20 30 40 I I I I I 0 10 20 30 40
TIME (min)
Fig. 3. HPLC analyses of neuromedin N degradation by soluble and membrane-associated fractions of astrocytes and primary- cultured neurons. HPLC profiles corresponded to incubations of neuromedin N (2 nmol, 20 pM) with soluble (A) or membrane-asso- ciated (B) fractions of 4-day-old cultured neurons and 15-day-old cultured astrocytes. Incubations were stopped, the samples centri- fuged and analyzed by HPLC as described in Materials and Meth- ods. Arrows indicated the elution times of synthetic standards run in the same HPLC conditions.
in the catabolism of both peptides remained constant during neuronal differentiation. In agreement with this hypothesis, we established that aminopeptidases M and B and endopep- tidase 24-1 1 activities were not modified during neuronal maturation (not shown).
301
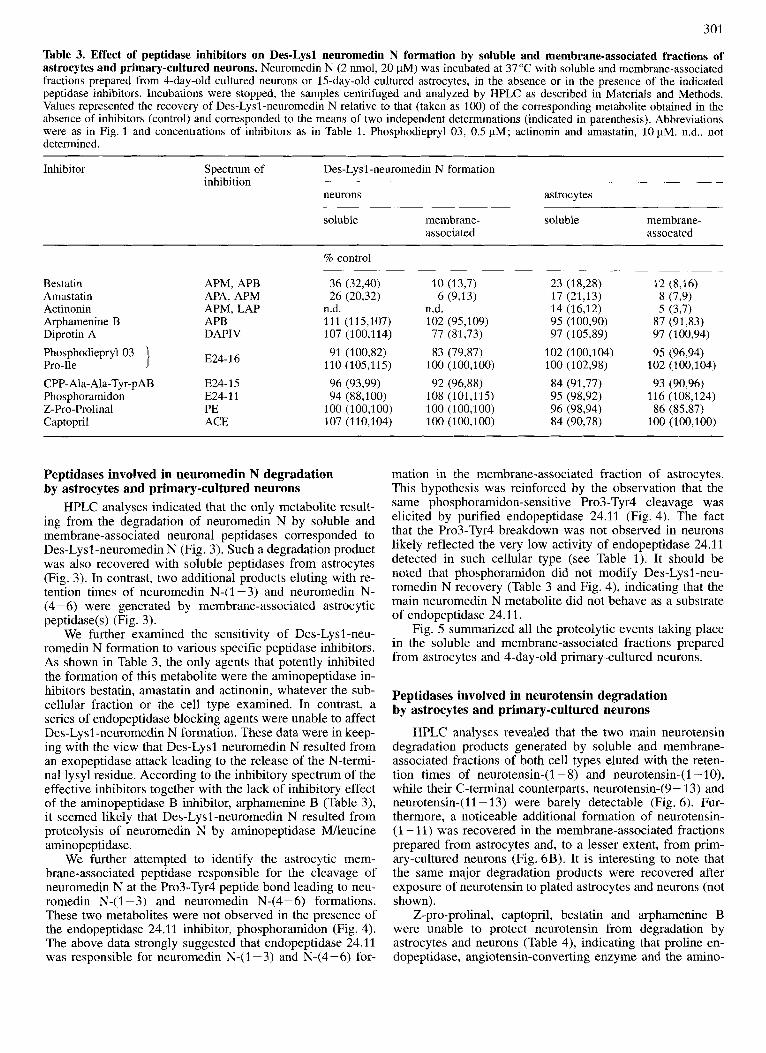
Table 3. Effect of peptidase inhibitors on Des-Lysl neuromedin N formation by soluble and membrane-associated fractions of astrocytes and primary-cultured neurons. Neuromedin N (2 nmol, 20 pM) was incubated at 37 "C with soluble and membrane-associated fractions prepared from 4-day-old cultured neurons or 15-day-old cultured astrocytes, in the absence or in the presence of the indicated peptidase inhibitors. Incubations were stopped, the samples centrifuged and analyzed by HPLC as described in Materials and Methods. Values represented the recovery of Des-Lysl-neuromedin N relative to that (taken as 100) of the corresponding metabolite obtained in the absence of inhibitors (control) and corresponded to the means of two independent determinations (indicated in parenthesis). Abbreviations were as in Fig. 1 and concentrations of inhibitors as in Table 1. Phosphodiepryl 03, 0.5 pM; actinonin and amastatin, 10 pM. n.d., not determined.
Inhibitor Spectrum of inhibition
Des-Lysl -neuromedin N formation
neurons astrocytes
soluble membrane- soluble membrane- associated assocated
% control
Bestatin Amastatin Actinonin Arphamenine B Diprotin A PhosphodieprylO3 Pro-Ile CPP- Ala- Ala-Tyr-p AB Phosphoramidon Z-Pro-Prolinal Captopril
APM, APB APA, APM APM, LAP APB DAPIV
E24-16
E24- 1 5 E24-11 PE ACE
36 (32,40) 26 (20,32)
n.d. 111 (115,107) 107 (100,114) 91 (100,82)
110 (105,115) 96 (93,99) 94 (88,100)
100 (100,100) 107 (110,104)
10 (13,7) 6 (9,131
n.d. 102 (95,109) 77 (81,73) 83 (79,87)
100 (100,100) 92 (96,88)
108 (101,115) 100 (100,100) 100 (100,100)
23 (18,28) 17 (21,13) 14 (16,12) 95 (100,90) 97 (105,89)
102 (100,104) 100 (102,98) 84 (91,77) 95 (98,92) 96 (98,94) 84 (90,78)
12 (8,16) 8 (7,9) 5 (3,7)
87 (9133) 97 (100,94) 95 (96,94)
102 (100,104) 93 (90,96)
116 (108,124) 86 (85,87)
100 (100,100)
Peptidases involved in neuromedin N degradation by astrocytes and primary-cultured neurons
HPLC analyses indicated that the only metabolite result- ing from the degradation of neuromedin N by soluble and membrane-associated neuronal peptidases corresponded to Des-Lysl-neuromedin N (Fig. 3). Such a degradation product was also recovered with soluble peptidases from astrocytes (Fig. 3). In contrast, two additional products eluting with re- tention times of neuromedin N-( 1 - 3) and neuromedin N- (4-6) were generated by membrane-associated astrocytic peptidase(s) (Fig. 3).
We further examined the sensitivity of Des-Lysl-neu- romedin N formation to various specific peptidase inhibitors. As shown in Table 3, the only agents that potently inhibited the formation of this metabolite were the aminopeptidase in- hibitors bestatin, amastatin and actinonin, whatever the sub- cellular fraction or the cell type examined. In contrast, a series of endopeptidase blocking agents were unable to affect Des-Lysl-neuromedin N formation. These data were in keep- ing with the view that Des-Lysl neuromedin N resulted from an exopeptidase attack leading to the release of the N-termi- nal lysyl residue. According to the inhibitory spectrum of the effective inhibitors together with the lack of inhibitory effect of the aminopeptidase B inhibitor, arphamenine B (Table 3), it seemed likely that Des-Lysl -neuromedin N resulted from proteolysis of neuromedin N by aminopeptidase MAeucine aminopeptidase.
We further attempted to identify the astrocytic mem- brane-associated peptidase responsible for the cleavage of neuromedin N at the Pro3-Tyr4 peptide bond leading to neu- romedin N-(I -3) and neuromedin N-(4-6) formations. These two metabolites were not observed in the presence of the endopeptidase 24.11 inhibitor, phosphoramidon (Fig. 4). The above data strongly suggested that endopeptidase 24.11 was responsible for neuromedin N-(1-3) and N-(4-6) for-
mation in the membrane-associated fraction of astrocytes. This hypothesis was reinforced by the observation that the same phosphoramidon-sensitive Pro3-Tyr4 cleavage was elicited by purified endopeptidase 24.11 (Fig. 4). The fact that the Pro3-Tyr4 breakdown was not observed in neurons likely reflected the very low activity of endopeptidase 24.11 detected in such cellular type (see Table 1). It should be noted that phosphoramidon did not modify Des-Lysl -neu- romedin N recovery (Table 3 and Fig. 4), indicating that the main neuromedin N metabolite did not behave as a substrate of endopeptidase 24.11.
Fig. 5 summarized all the proteolytic events taking place in the soluble and membrane-associated fractions prepared from astrocytes and 4-day-old primary-cultured neurons.
Peptidases involved in neurotensin degradation by astrocytes and primary-cultured neurons
HPLC analyses revealed that the two main neurotensin degradation products generated by soluble and membrane- associated fractions of both cell types eluted with the reten- tion times of neurotensin-(I - 8) and neurotensin-(I -lo), while their C-terminal counterparts, neurotensin-(9 - 13) and neurotensin-(ll-13) were barely detectable (Fig. 6). Fur- thermore, a noticeable additional formation of neurotensin- (1 - 11) was recovered in the membrane-associated fractions prepared from astrocytes and, to a lesser extent, from prim- ary-cultured neurons (Fig. 6B). It is interesting to note that the same major degradation products were recovered after exposure of neurotensin to plated astrocytes and neurons (not shown).
Z-pro-prolinal, captopril, bestatin and arphamenine B were unable to protect neurotensin from degradation by astrocytes and neurons (Table 4), indicating that proline en- dopeptidase, angiotensin-converting enzyme and the amino-

302
A
ASTROCYTES Des-Lysl NN
4- E 24-11
5 4
L
NK(1-3)
7
NN
1 li NN-(4-6)
I I I I I 0 10 20 30 40
B
L NN
../
I I I 1 I 0 10 20 30 40
TIME (min)
Fig. 4. Degradation of neuromedin N by membrane-associated fractions of astrocytes and by purified E 24.11: effect of phos- phoramidon. The membrane-associated fraction of 15 day-old cul- tured astrocytes and purified rat kidney E 24.11 (0.5 pg) were incu- bated with neuromedin N (2 and 5 nmoles respectively) in the ab- sence (A) or in the presence (B) of 1 pM phosphoramidon. Samples were acidified and HPLC analyzed as described in the Methods.
I Des-Lysi NN I
Neurons strocytes
E 24-1 1
NN-(1-3) t NN-(4-6)
Fig.5. Model for neuromedin N degradation by neurons and astrocytes. Solid arrows indicated cleavages by soluble peptidases and dashed arrows corresponded to breakdowns elicited by mem- brane-associated proteolytic activities. NN = Lysl-Ile2-Pro3-Tyr4- Ile5-Leu6-OH.
peptidases M and B, respectively, did not participate in neu- rotensin catabolism by these cell cultures. The inhibitors of endopeptidases 24.16 and 24.15 potently protected neuroten- sin from degradation by the soluble fractions from astrocytes and neurons. In the membrane-associated compartments, phosphodiepryl 03 also induced an inhibition of neurotensin degradation while the endopeptidase 24.15 blocking agent CPP-Ala-Ala-Tyr-pAB appeared ineffective (Table 4). This was in keeping with the very low endopeptidase 24-15 activ- ity recovered in the membrane-associated fractions of both
NEURONS
0 r)
2
4
ASTROCYTES
I
c L + NT a
NT
NT
I I I I J 0 10 20 30 40
1 I I I I 0 10 20 30 40
TIME (min)
Fig. 6. HPLC analyses of neurotensin degradation by soluble and membrane-associated fractions of astrocytes and primary-cul- tured neurons. HPLC profiles corresponded to incubations of neu- rotensin with soluble (A) and membrane-associated (B) fractions of 4-day-old cultured neurons and 15-day-old astrocytes. Incubations were stopped, the samples centrifuged and analyzed by HPLC as described in Materials and Methods. Arrows indicated the elution times of neurotensin partial sequences run in the same HPLC condi- tions.
cell types (see Fig. 1). The endopeptidase 24.11 inhibitor, phosphoramidon, only inhibited neurotensin degradation in the membrane associated fraction derived from astrocytes (Table 4). Such effect was not observed in neurons (Table 4) in agreement with the faint endopeptidase 24.11 activity in this cell type (Table 1).
The effect of the various specific inhibitors on the forma- tion of the major catabolites neurotensin-(1 - 8) and neuro- tensin-(1-10) is presented in Table 5. In the astrocytic and neuronal soluble fractions, the endopeptidase 24.16 inhibitor phosphodiepryl 03 was the only agent that potently blocked the neurotensin-(1 - 10) formation (Table 5). This was in agreement with the reported specificity of rat endopeptidase 24.16 towards neurotensin that led to the formations of neu- rotensin-(I -10) and neurotensin-(11-13) (Checler et al., 1986~) . In the membrane-associated fractions, an additional phosphoramidon-sensitive neurotensin-( 1 - 10) formation was evidenced (Table 5) . This indicated that, besides endo- peptidase 24.16, endopeptidase 24.11 also participated in the production of this metabolite. This correlated well with: (a) the protective effect of phosphoramidon on the recovery of neurotensin (Table 4); (b) the detection of neurotensin-(1-
303
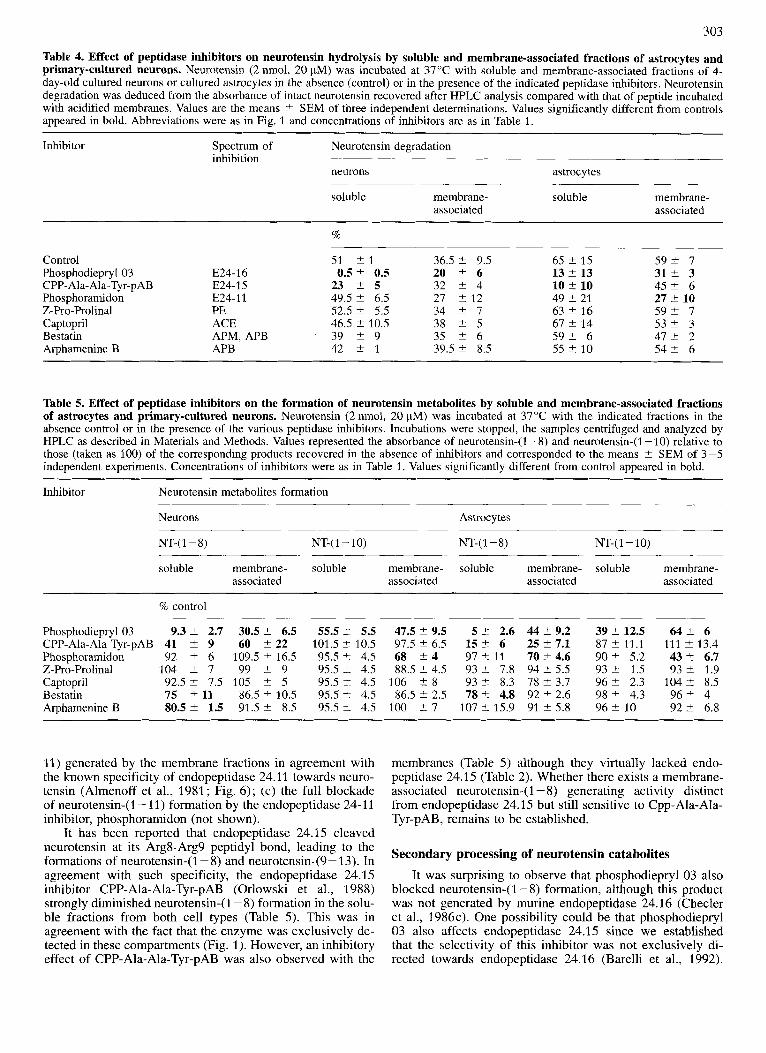
Table 4. Effect of peptidase inhibitors on neurotensin hydrolysis by soluble and membrane-associated fractions of astrocytes and primary-cultured neurons. Neurotensin (2 nmol, 20 FM) was incubated at 37°C with soluble and membrane-associated fractions of 4- day-old cultured neurons or cultured astrocytes in the absence (control) or in the presence of the indicated peptidase inhibitors. Neurotensin degradation was deduced from the absorbance of intact neurotensin recovered after HPLC analysis compared with that of peptide incubated with acidified membranes. Values are the means t SEM of three independent determinations. Values significantly different from controls appeared in bold. Abbreviations were as in Fig. 1 and concentrations of inhibitors are as in Table 1.
Inhibitor Spectrum of Neurotensin degradation inhibition
neurons
soluble membrane- associated
astrocytes
soluble membrane- associated
Control Phosphodiepryl 03 CPP- Ala-Ala-Tyr-pAB Phosphoramidon Z-Pro-Prolinal Captopril Bestatin Arphamenine B
%
51 t 1 36.5 2 9.5 65 2 15 5 9 2 7 E24-16 0.5 2 0.5 20 2 6 13 2 13 31% 3 E24-15 23 2 5 32 2 4 10 +. 10 4 5 2 6 E24-11 49.5 2 6.5 27 2 12 49 2 21 27 t 10 PE 52.5 2 5.5 34 2 7 63 2 16 5 9 2 7 ACE 46.5 2 10.5 38 t 5 67 2 14 5 3 2 3 APM, APB 39 2 9 35 2 6 5 9 2 6 4 7 t 2 APB 42 2 1 39.5 2 8.5 55 t 10 5 4 2 6
-
Table 5. Effect of peptidase inhibitors on the formation of neurotensin metabolites by soluble and membrane-associated fractions of astrocytes and primary-cultured neurons. Neurotensin (2 nmol, 20 pM) was incubated at 37°C with the indicated fractions in the. absence control or in the presence of the various peptidase inhibitors. Incubations were stopped, the samples centrifuged and analyzed by HPLC as described in Materials and Methods. Values represented the absorbance of neurotensin-(1-8) and neurotensin-(1 -10) relative to those (taken as 100) of the corresponding products recovered in the absence of inhibitors and corresponded to the means t SEM of 3-5 independent experiments. Concentrations of inhibitors were as in Table 1. Values significantly different from control appeared in bold.
Inhibitor Neurotensin metabolites formation
Neurons Astrocytes ~
NT-( 1 - 8) NT-( 1 - 10) NT-( 1 - 8) NT-( 1 - 10) ~~~ ~ -
soluble membrane- soluble membrane- soluble membrane- soluble membrane- associated associated associated associated
% control
Phosphodiepryl03 9.3 2 2.7 CPP-Ala-Ala-Tyr-pAB 41 2 9 Phosphoramidon 92 2 6 Z-Pro-Prolinal 104 2 7 Captopril 92.5 2 7.5 Bestatin 75 2 11 Arphamenine B 80.5 2 1.5
30.5 2 6.5 55.5 t 5.5 47.5 2 9.5 5 t 2.6 44 2 9.2 60 2 22 101.5 t 10.5 97.5 2 6.5 15 % 6 25 t 7.1
109.5 C 16.5 95.5 t 4.5 68 2 4 97 % 11 7 0 t 4.6 99 t 9 95.5 L 4.5 88.5 2 4.5 93 t 7.8 94 2 5.5
105 2 5 95.5 t 4.5 106 +. 8 93 t 8.3 78 ? 3.7 86.5 2 10.5 95.5 t 4.5 86.5 2 2.5 78 t 4.8 92 2 2.6 91.5 2 8.5 95.5 t 4.5 100 2 7 107 t 15.9 91 2 5.8
3 9 2 12.5 6 4 2 6 8 7 2 11.1 111 2 13.4 90 2 5.2 43 t 6.7 93 t 1.5 93 2 1.9 9 6 2 2.3 104 2 8.5 98 2 4.3 9 6 2 4
92 t 6.8 96 2 10
11) generated by the membrane fractions in agreement with the known specificity of endopeptidase 24.11 towards neuro- tensin (Almenoff et al., 1981; Fig. 6); (c) the full blockade of neurotensin-(1 - 11) formation by the endopeptidase 24-1 1 inhibitor, phosphoramidon (not shown).
It has been reported that endopeptidase 24.15 cleaved neurotensin at its Arg8-Arg9 peptidyl bond, leading to the formations of neurotensin-(1-8) and neurotensin-(9- 13). In agreement with such specificity, the endopeptidase 24.1 5 inhibitor CPP-Ala-Ala-Tyr-pAB (Orlowski et al., 1988) strongly diminished neurotensin-(1 - 8) formation in the solu- ble fractions from both cell types (Table 5). This was in agreement with the fact that the enzyme was exclusively de- tected in these compartments (Fig. 1). However, an inhibitory effect of CPP-Ala-Ala-Tyr-pAB was also observed with the
membranes (Table 5 ) although they virtually lacked endo- peptidase 24.1 5 (Table 2). Whether there exists a membrane- associated neurotensin-(1 - 8) generating activity distinct from endopeptidase 24.15 but still sensitive to Cpp-Ala-Ala- Tyr-pAB, remains to be established.
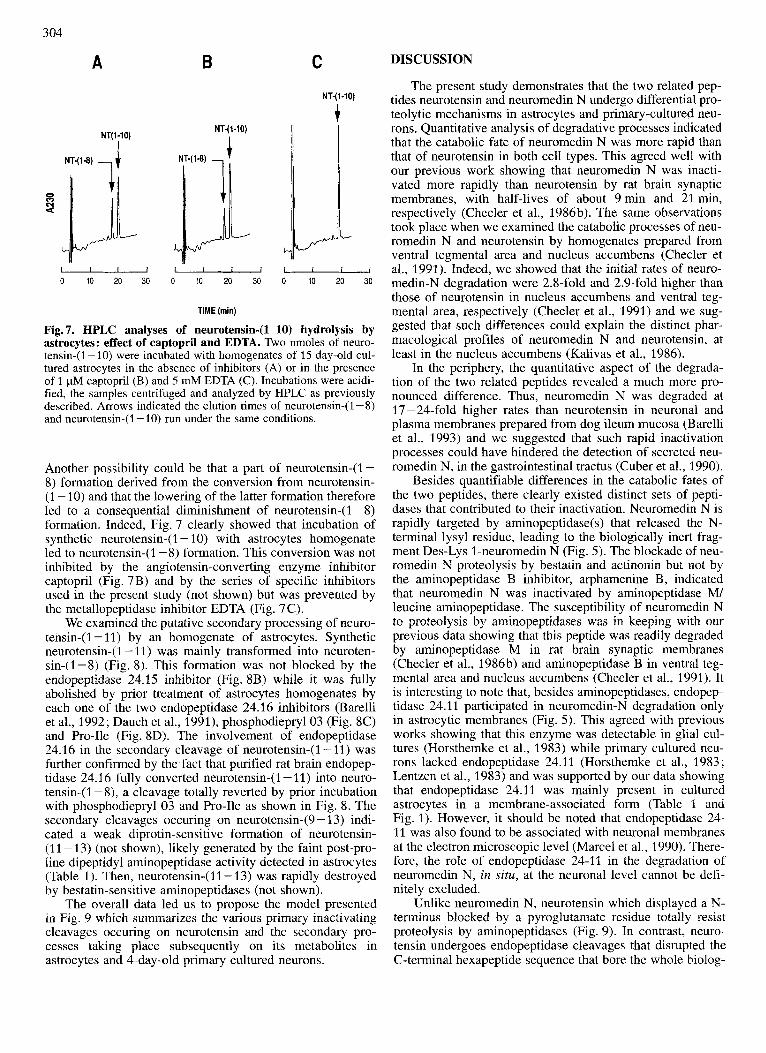
Secondary processing of neurotensin catabolites It was surprising to observe that phosphodiepryl 03 also
blocked neurotensin-( 1 - 8) formation, although this product was not generated by murine endopeptidase 24.16 (Checler et al., 1986~) . One possibility could be that phosphodiepryl 03 also affects endopeptidase 24.15 since we established that the selectivity of this inhibitor was not exclusively di- rected towards endopeptidase 24.16 (Barelli et al., 1992).
304
A C DISCUSSION
NT-(l-lO) +
--- 0 10 20 30 0 10 20 30 0 10 20 30
TIME (min)
Fig. 7. HPLC analyses of neurotensin-(1-10) hydrolysis by astrocytes: effect of captopril and EDTA. Two nmoles of neuro- tensin-(1 - 10) were incubated with homogenates of 15 day-old cul- tured astrocytes in the absence of inhibitors (A) or in the presence of 1 pM captopril (B) and 5 mM EDTA (C). Incubations were acidi- fied, the samples centrifuged and analyzed by HPLC as previously described. Arrows indicated the elution times of neurotensin-( 1 - 8) and neurotensin-(1 -10) run under the same conditions.
Another possibility could be that a part of neurotensin-(1 - 8) formation derived from the conversion from neurotensin- (1 - 10) and that the lowering of the latter formation therefore led to a consequential diminishment of neurotensin-( 1 - 8) formation. Indeed, Fig. 7 clearly showed that incubation of synthetic neurotensin-( 1 - 10) with astrocytes homogenate led to neurotensin-( 1 - 8) formation. This conversion was not inhibited by the angiotensin-converting enzyme inhibitor captopril (Fig. 7B) and by the series of specific inhibitors used in the present study (not shown) but was prevented by the metallopeptidase inhibitor EDTA (Fig. 7C).
We examined the putative secondary processing of neuro- tensin-(1 - 11) by an homogenate of astrocytes. Synthetic neurotensin-( 1 - 11) was mainly transformed into neuroten- sin-(1-8) (Fig. 8). This formation was not blocked by the endopeptidase 24.15 inhibitor (Fig. 8B) while it was fully abolished by prior treatment of astrocytes homogenates by each one of the two endopeptidase 24.16 inhibitors (Barelli et al., 1992; Dauch et al., 1991), phosphodieprylO3 (Fig. 8C) and Pro-Ile (Fig. SD). The involvement of endopeptidase 24.16 in the secondary cleavage of neurotensin-(1-11) was further confirmed by the fact that purified rat brain endopep- tidase 24.16 fully converted neurotensin-(1 - 11) into neuro- tensin-(1 - 8), a cleavage totally reverted by prior incubation with phosphodiepryl 03 and Pro-Ile as shown in Fig. 8. The secondary cleavages occuring on neurotensin-(9 - 13) indi- cated a weak diprotin-sensitive formation of neurotensin- (11 -13) (not shown), likely generated by the faint post-pro- line dipeptidyl aminopeptidase activity detected in astrocytes (Table 1). Then, neurotensin-(11- 13) was rapidly destroyed by bestatin-sensitive aminopeptidases (not shown).
The overall data led us to propose the model presented in Fig. 9 which summarizes the various primary inactivating cleavages occuring on neurotensin and the secondary pro- cesses taking place subsequently on its metabolites in astrocytes and 4-day-old primary cultured neurons.
The present study demonstrates that the two related pep- tides neurotensin and neuromedin N undergo differential pro- teolytic mechanisms in astrocytes and primary-cultured neu- rons. Quantitative analysis of degradative processes indicated that the catabolic fate of neuromedin N was more rapid than that of neurotensin in both cell types. This agreed well with our previous work showing that neuromedin N was inacti- vated more rapidly than neurotensin by rat brain synaptic membranes, with half-lives of about 9 min and 21 min, respectively (Checler et al., 1986b). The same observations took place when we examined the catabolic processes of neu- romedin N and neurotensin by homogenates prepared from ventral tegmental area and nucleus accumbens (Checler et al., 1991). Indeed, we showed that the initial rates of neuro- medin-N degradation were 2.8-fold and 2.9-fold higher than those of neurotensin in nucleus accumbens and ventral teg- mental area, respectively (Checler et al., 1991) and we sug- gested that such differences could explain the distinct phar- macological profiles of neuromedin N and neurotensin, at least in the nucleus accumbens (Kalivas et al., 1986).
In the periphery, the quantitative aspect of the degrada- tion of the two related peptides revealed a much more pro- nounced difference. Thus, neuromedin N was degraded at 17 - 24-fold higher rates than neurotensin in neuronal and plasma membranes prepared from dog ileum mucosa (Barelli et al., 1993) and we suggested that such rapid inactivation processes could have hindered the detection of secreted neu- romedin N, in the gastrointestinal tractus (Cuber et al., 1990).
Besides quantifiable differences in the catabolic fates of the two peptides, there clearly existed distinct sets of pepti- dases that contributed to their inactivation. Neuromedin N is rapidly targeted by aminopeptidase(s) that released the N- terminal lysyl residue, leading to the biologically inert frag- ment Des-Lys 1-neuromedin N (Fig. 5). The blockade of neu- romedin N proteolysis by bestatin and actinonin but not by the aminopeptidase B inhibitor, arphamenine B, indicated that neuromedin N was inactivated by aminopeptidase M/ leucine aminopeptidase. The susceptibility of neuromedin N to proteolysis by aminopeptidases was in keeping with our previous data showing that this peptide was readily degraded by aminopeptidase M in rat brain synaptic membranes (Checler et al., 1986b) and aminopeptidase B in ventral teg- mental area and nucleus accumbens (Checler et al., 1991). It is interesting to note that, besides aminopeptidases, endopep- tidase 24.11 participated in neuromedin-N degradation only in astrocytic membranes (Fig. 5). This agreed with previous works showing that this enzyme was detectable in glial cul- tures (Horsthemke et al., 1983) while primary cultured neu- rons lacked endopeptidase 24.11 (Horsthemke et al., 1983; Lentzen et al., 1983) and was supported by our data showing that endopeptidase 24.1 1 was mainly present in cultured astrocytes in a membrane-associated form (Table 1 and Fig. 1). However, it should be noted that endopeptidase 24- 11 was also found to be associated with neuronal membranes at the electron microscopic level (Marcel et al., 1990). There- fore, the role of endopeptidase 24-11 in the degradation of neuromedin N, in situ, at the neuronal level cannot be defi- nitely excluded.
Unlike neuromedin N, neurotensin which displayed a N- terminus blocked by a pyroglutamate residue totally resist proteolysis by aminopeptidases (Fig. 9). In contrast, neuro- tensin undergoes endopeptidase cleavages that disrupted the C-terminal hexapeptide sequence that bore the whole biolog-

305
E 24-1 6
- 0 10 20 30
NT-(l-8) 1
c NTf-H)
NT-(1-11)
1
.
Pro-lle NT-(l-11)
1 f
0 10 20 30 0 10 20 30 TIME (mln)
0 10 20 30
Fig. 8. HPLC analyses of neurotensb(1-11) by astrocytes and purified endopeptidase 24.16 : effect of CPP-Ala-Ala-Tyr-pAB, phosphodiepryl 03 and Pro-Ile. Neurotensin-(1 - 11) (2 nmol, 20 pM) was incubated at 37 "C with an homogenate of 15-day-old cultured astrocytes or purified rat brain E 24.16 in the absence of inhibitor (A) or in the presence of 1 pM CPP-Ala-Ala-Tyr-pAB (B), 0.5 pM phosphodiepryl 03 (C) and 10 mM Pro-Ile (D). Conditions of HPLC analyzes are described in Material and Methods.
ical activity of the peptide. The two endopeptidases ubiqui- tously involved in the two main cleavages were identified as endopeptidase 24.15 and endopeptidase 24.16. It should be noted that a recent study by Meintlein and Dahms clearly demonstrated that endopeptidases 24.15 and 24.16 mainly participated to the inactivation of neurotensin by plated rat cortical astrocytes (Mentlein and Dahms, 1994). In agreement with such a hypothesis, the immunocytochemical localization of endopeptidase 24.15 in rat brain established that this enzyme displayed a widespread distribution within both neurons and glial cells (Healy and Orlowski, 1992). Furthermore we recently established that endopeptidase 24.16 activity (Checler et al., 1986a) and immunoreactivity (Chabry et al., 1990) could be detected in pure cultured neu- rons. Analyses at the light and electronic microscopic levels confirmed the neuronal occurence of endopeptidase 24.16 and also revealed an astrocytic counterpart of this activity (Woulfe et al., 1992).
The involvement of endopeptidases 24.15 and 24.16 in neurotensin degradation raised the question of the relevance of astrocytes and pure cultured neurons as physiological models for better understanding peptide degradation. Several experimental data concerning endopeptidase 24.16 suggested that these cell cultures could indeed be of help to solve the question of the termination of the neurotensin message. Firstly, we previously showed that a colocalization of endo-
peptidase 24.16 and neurotensin receptors existed on a discrete population (less than 10% of total cells) of 4-day- old pure cultured neurons (Chabry et al., 1990). Such topo- graphical parallelism is a strong prerequisit to envisage a physiological role of the enzyme in neurotensin inactivation. Secondly, electron microscopy revealed frequent appositions between neuronal and glial endopeptidase-24.16-positive ele- ments in brain zones corresponding to neurotensin affer- ences. Furthermore, clues of synergistic actions of neuronal and glial forms of endopeptidase 24.16 came from evidence of putative endocytic invaginations that could reflect cell to cell translocations of cytosoluble material (Woulfe et al., 1992). Together, these data brought anatomical supports to the hypothesis that membrane-associated but also cytosoluble endopeptidase 24.16 from both astrocytes and neurons in- deed contribute to neurotensin physiological inactivation. Wether such a conclusion could also be stated for endopep- tidase 24.15 remains to be verified by additional anatomical studies.
One copy each of neurotensin and neuromedin N are lo- cated at the C-terminal of the prepro-neurotensin precursor (Dobner et al., 1987). The organization of the gene revealed that the two peptides are encoded by the fourth exon (Kis- lauskis et al., 1988) A recent study seemed to indicate that the processing of the precursor in the brain led to the con- comittant biosynthesis of the two peptides (De Nadai et al.,

306
LNT-(1-1O)J
E24-ll Neurons NT
peptidase
E 24-16
Fig. 9. Model for neurotensin degradation by neurons and astrocytes. Solid arrows corresponded to cleavages by soluble frac- tions and dashed arrows indicated cleavages by membrane-associ- ated fractions. Minor contributions of peptidase activities are indi- cated by small characters. NT = pGlul-Leu2-Tyr3-Glu4-Asn5- Lys6-Pro7-Arg8-Arg9-Pro1O-Tyrl1-Ile12-Leu13-OH.
1989). Therefore, downstream of the maturation processing, metabolic events could be the way to modulate the lifetime of the two peptides, thereby controlling their neuromodula- tory functions. In this respect, it could be hypothesized that neuromedin N behaves as a short-lived peptide underlying a neurotransmitter function, while neurotensin could be long- term acting with prolonged activity, thereby carrying the neuromodulatory message in the central nervous system.
We wish to acknowledge Drs S. Wilk and M. Orlowski (Mount Sinai School of Medicine, New York) for providing us with Z-pro- prolinal, Bz-Gly-AAF-pAB and CPP-Ala-Ala-Tyr-pAB. We are grateful to Dr V. Dive who synthesized and provided us with phos- phodiepryl 03. We wish to thank Drs P. Crine and G. Boileau (Fa- cult6 de MCdecine de Montreal, Montreal) for giving us the purified endopeptidase 24-11. V. ChCron and F. Aguila are acknowledged for their expert secretarial and artwork assistance. This work was supported by the Centre National de la Recherche Scientifique and the Institut National de la Sante' et de la Recherche Me'dicale.
REFERENCES Almenoff, J., Wilk, S. & Orlowski, M. (1981) Membrane bound
pituitary metalloendopeptidase : apparent identity to enkepha- linase, Biochem. Biophys. Res. Commun. 102, 206-214.
Barelli, H., Vincent, J. P. & Checler, F. (1991) Rat brain endopepti- dase 24-15 : specificity towards opioids, tachykinins and neuro- tensin-related peptides. Comparison with the neurotensin degrad- ing metallopeptidase endopeptidase 24 - 16, Neurochem. Life Sci.
Barelli, H., Dive, V., Yiotakis, A., Vincent, J. P. & Cheder, F. (1992) Potent inhibition of endopeptidase 24.16 and endopeptidase 24.1 5 by the phosphonamide peptide N-(phenylethylphospho- ny1)-Gly-LPro-L-aminohexanoic acid, Biochem. J. 287, 621 - 625.
Barelli, H., Mao, Y. K., Vincent, B., Daniel, E. E., Vincent, J. P. & Cheder, F. (1993) Differential catabolic fate of neuromedin N and neurotensin in the canine intestinal mucosa, Peptides 14,
Chabry, J., Cheder, F., Vincent, J. P. & Mazella, J. (1990) Colocali- zation of neurotensin receptors and of the neurotensin-degrading enzyme endopeptidase 24-16 in primary cultures of neurons, J. Neurosci. 10, 3916-3921.
Checler, F., Mazella, J., Kitabgi, P. & Vincent, J. P. (1986a) High affinity receptor sites and rapid proteolytic inactivation of neuro-
Adv. 10, 115-124.
457-463.
tensin in primary cultured neurons, J. Neurochem. 47, 1742- 1748.
Checler, F., Vincent, J. P. & Kitabgi, P. (1986b) Neuromedin N: high affinity interaction with brain neurotensin receptors and rapid inactivation by brain synaptic peptidases, Eur: J. Pharmacol. 126, 239-244.
Cheder, F., Vincent, J. P. & Kitabgi, P. (1986~) Purification and characterization of a novel neurotensin degrading peptidase from rat brain synaptic membranes, J. Biol. Chem. 261, 11274- 11 281.
Cheder, F., Barelli, H., Kitabgi, P. & Vincent, J. P. (1988) Neuroten- sin metabolism in various tissues from central and peripheral origins. Ubiquitous involvement of a novel neurotensin degrad- ing metalloendopeptidase, Biochimie (Paris) 70, 75 - 82.
Checler, F., Dauch, P., Masuo, Y. & Vincent, J. P. (1991) Neuroten- sin and neuromedin N are differently metabolized in ventral teg- mental area and nucleus accumbens, J. Neurochem. 56, 1320- 1328.
Cheder, F. (1 993) Neuropeptide-degrading peptidases in Methods in neurotransmitter and neuropeptide research (Nagatsu, T., Par- vez, H., Naoi, M. & Parvez, s., eds) chapter 13, (in the press) Elsevier Science Publishers, Amsterdam.
Cuber, J. C., Hemnann, C., Kitabgi, P., Bosshard, A,, Bernard, C., De Nadai, F. & Chayvialle, J. A. (1990) Neuromedin-N is not released with neurotensin from rat ileum, Endocrinology 126, 1584- 1592.
Dauch, P., Vincent, J. P. & Cheder, F. (1991) Specific inhibition of endopeptidase 24.16 by dipeptides, Eur. J. Biochem. 202, 269- 276.
Dauch, P., Masuo, Y., Vincent, J. P. & Cheder, F. (1993) A survey of the cerebral regionalization and ontogeny of eight exo- and endo- peptidases in murines, Peptides 14, 593-599.
De Nadai, F., Cuber, J. C. & Kitabgi, P. (1989) The characterization and regional distribution of neuromedin N-like immunoreactivity in rat brain using a highly sensitive and specific radio- immunoassay. Comparison with the distribution of neurotensin, Brain Res. 500, 193-198.
Dobner, P. R., Barber, D. L., Villa-Komaroff, L. & McKierman, C. (1987) Cloning and sequence analysis of cDNA for the canine neurotensidneuromedin N precursor, Proc. Nut1 Acad. Sci. USA 84, 3516-3520.
Healy, D. & Orlowski, M. (1 992) Immunocytochemical localization of endopeptidase 24-15 in rat brain, Brain Res. 571, 121-128.
Horsthemke, B., Hamprecht, B. & Bauer, K. (1983) Heterogeneous distribution of enkephalin-degrading peptidases between neuro- nal and glial cells, Biochem. Biophys. Res. Commun. 115, 423- 429.
Kalivas, P. W., Richardson-Carlson, R. & Duffy, P. (1986) Neu- romedin N mimics the actions of neurotensin in the ventral teg- mental area but not in the nucleus accumbens, J. Pharmacol. Exp. Ther: 238, 1126-1131.
Kislauskis, E., Bullock, B., Mc Neil, S. & Dobner, P. R. (1988) The rat gene encoding neurotensin and neuromedin N. Structure, tissue-specific expression, and evolution of exon sequences, J . Biol. Chem. 263, 4963-4968.
Lentzen, H., Monden, I., Linke, J. & Palenker, J. (1983) No evidence of enkephalinase A on neuronal cells, Life Sci. 33, 105-108.
Marcel, D., Pollard, H., Verroust, P., Schwartz, J. C. & Beaudet, A. (1 990) Electron microscopic localization of immunoreactive enkephalinase (EC 3.4.24.11) in the neostriatum of the rat, J. Neurosci. 10, 2804-2817.
Mentlein, R. & Dahms, P. (1994) Endopeptidases 24.16 and 24.15 are responsible for the degradation of somatostatin, neurotensin and other neuropeptides by cultivated rat cortical astrocytes, J. Neurochem. 62, 27-36.
Orlowski, M., Michaud, C. & Molineaux, C. J. (1988) Substrate- related potent inhibitors of brain metalloendopeptidase, Bio- chemistry 27, 597-602.
Woulfe, J., Checler, F. & Beaudet, A. (1992) Light and electron microscopic localization of the neutral metalloendopeptidase EC 3.4.24.16, in the mesencephalon of the rat, Eur: J. Neurosci. 4, 1 309 - 1 3 19.




















