
Distinct roles of trauma and transfusion in induction of immune modulation after injury
21
TRANSFUSION PRACTICE Distinct roles of trauma and transfusion in induction of immune modulation after injuryRachael P. Jackman, Garth H. Utter, Marcus O. Muench, John W. Heitman, Matthew M. Munz, Robert W. Jackman, Hope H. Biswas, Ryan M. Rivers, Leslie H. Tobler, Michael P. Busch, and Philip J. Norris BACKGROUND: Trauma and transfusion can both alter immunity, and while transfusions are common among traumatically injured patients, few studies have exam- ined their combined effects on immunity. STUDY DESIGN AND METHODS: We tracked the plasma levels of 41 immunomodulatory proteins in 56 trauma patients from time of injury up to 1 year later. In addition, a murine model was developed to distinguish between the effects of transfusion and underlying injury and blood loss. RESULTS: Thirty-one of the proteins had a significant change over time after traumatic injury, with a mixed early response that was predominantly anti- inflammatory followed by a later increase in proteins involved in wound healing and homeostasis. Results from the murine model revealed similar cytokine responses to humans. In mice, trauma and hemorrhage caused early perturbations in a number of the pro- and anti-inflammatory mediators measured, and transfusion blunted early elevations in interleukin (IL)-6, IL-10, matrix metalloproteinase-9, and interferon-g. Transfu- sion caused or exacerbated changes in monocyte chemotactic protein-1, IL-1a, IL-5, IL-15, and soluble E-selectin. Finally, trauma and hemorrhage alone increased CXCL1 and IL-13. CONCLUSIONS: This work provides a detailed charac- terization of the major shift in the immunologic environ- ment in response to trauma and transfusion and clarifies which immune mediators are affected by trauma and hemorrhage and which by transfusion. T raumatic injury represents a major health concern, resulting in approximately 2.7 million hospital admissions each year in the United States. 1,2 Transfusion of allogeneic blood prod- ucts is a common intervention after traumatic injury, with approximately 9% of trauma patients transfused and approximately one-third of these given massive transfu- sions. 1,3 As a result, 9% of red blood cell (RBC) units in the United States are used in the acute support of traumati- cally injured patients. 4 Traumatic injury has been shown to be a major immunologic event, leading to immune dysregulation ABBREVIATIONS: EGF = epidermal growth factor; FGF-2 = fibroblast growth factor-2; GEE = generalized estimating equations; IP-10 = interferon-g–inducible protein 10; ISS = injury severity score; KC = CXCL1; MCP-1 = monocyte chemot- actic protein-1; MDC = macrophage-derived chemokine; MIF = macrophage migration inhibitory factor; MIP = macro- phage inflammatory protein; MMP-9 = matrix metalloproteinase-9; MPO = myeloperoxidase; sE-selectin = soluble E-selectin; sVCAM-1 = soluble vascular cell adhesion molecule-1; tPAI-1 = total plasminogen activator inhibitor-1; UCDMC = University of California Davis Medical Center; VEGF = vascular endothelial growth factor. From the Blood Systems Research Institute, San Francisco, California; the Departments of Surgery and Political Science, University of California, Davis, California; the Departments of Laboratory Medicine and Medicine, University of California, San Francisco, California; and the Division of Epidemiology, School of Public Health, University of California, Berkeley, California. Address reprint requests to: Rachael P. Jackman, Blood Systems Research Institute, 270 Masonic Avenue, San Francisco, CA 94118; e-mail: [email protected]. This work was supported by NIH RO1 HL-083388-01A1. Received for publication October 14, 2011; revision received January 27, 2012, and accepted January 27, 2012. doi: 10.1111/j.1537-2995.2012.03618.x TRANSFUSION 2012;52:2533-2550. Volume 52, December 2012 TRANSFUSION 2533
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Distinct roles of trauma and transfusion in induction of immune modulation after injury

T R A N S F U S I O N P R A C T I C E
Distinct roles of trauma and transfusion in induction of immunemodulation after injury_3618 2533..2550
Rachael P. Jackman, Garth H. Utter, Marcus O. Muench, John W. Heitman, Matthew M. Munz,Robert W. Jackman, Hope H. Biswas, Ryan M. Rivers, Leslie H. Tobler, Michael P. Busch,
and Philip J. Norris
BACKGROUND: Trauma and transfusion can both alterimmunity, and while transfusions are common amongtraumatically injured patients, few studies have exam-ined their combined effects on immunity.STUDY DESIGN AND METHODS: We tracked theplasma levels of 41 immunomodulatory proteins in 56trauma patients from time of injury up to 1 year later. Inaddition, a murine model was developed to distinguishbetween the effects of transfusion and underlying injuryand blood loss.RESULTS: Thirty-one of the proteins had a significantchange over time after traumatic injury, with a mixedearly response that was predominantly anti-inflammatory followed by a later increase in proteinsinvolved in wound healing and homeostasis. Resultsfrom the murine model revealed similar cytokineresponses to humans. In mice, trauma and hemorrhagecaused early perturbations in a number of the pro- andanti-inflammatory mediators measured, and transfusionblunted early elevations in interleukin (IL)-6, IL-10,matrix metalloproteinase-9, and interferon-g. Transfu-sion caused or exacerbated changes in monocytechemotactic protein-1, IL-1a, IL-5, IL-15, and solubleE-selectin. Finally, trauma and hemorrhage aloneincreased CXCL1 and IL-13.CONCLUSIONS: This work provides a detailed charac-terization of the major shift in the immunologic environ-ment in response to trauma and transfusion andclarifies which immune mediators are affected bytrauma and hemorrhage and which by transfusion.
Traumatic injury represents a major healthconcern, resulting in approximately 2.7 millionhospital admissions each year in the UnitedStates.1,2 Transfusion of allogeneic blood prod-
ucts is a common intervention after traumatic injury, withapproximately 9% of trauma patients transfused andapproximately one-third of these given massive transfu-sions.1,3 As a result, 9% of red blood cell (RBC) units in theUnited States are used in the acute support of traumati-cally injured patients.4
Traumatic injury has been shown to be a majorimmunologic event, leading to immune dysregulation
ABBREVIATIONS: EGF = epidermal growth factor; FGF-2 =fibroblast growth factor-2; GEE = generalized estimatingequations; IP-10 = interferon-g–inducible protein 10; ISS =injury severity score; KC = CXCL1; MCP-1 = monocyte chemot-actic protein-1; MDC = macrophage-derived chemokine;MIF = macrophage migration inhibitory factor; MIP = macro-phage inflammatory protein; MMP-9 = matrixmetalloproteinase-9; MPO = myeloperoxidase; sE-selectin =soluble E-selectin; sVCAM-1 = soluble vascular cell adhesionmolecule-1; tPAI-1 = total plasminogen activator inhibitor-1;UCDMC = University of California Davis Medical Center;VEGF = vascular endothelial growth factor.
From the Blood Systems Research Institute, San Francisco,California; the Departments of Surgery and Political Science,University of California, Davis, California; the Departments ofLaboratory Medicine and Medicine, University of California,San Francisco, California; and the Division of Epidemiology,School of Public Health, University of California, Berkeley,California.
Address reprint requests to: Rachael P. Jackman, BloodSystems Research Institute, 270 Masonic Avenue, San Francisco,CA 94118; e-mail: [email protected].
This work was supported by NIH RO1 HL-083388-01A1.Received for publication October 14, 2011; revision
received January 27, 2012, and accepted January 27, 2012.doi: 10.1111/j.1537-2995.2012.03618.xTRANSFUSION 2012;52:2533-2550.
Volume 52, December 2012 TRANSFUSION 2533

that can contribute to further tissue damage as well ascompromise the host’s ability to fight infection. Severeinjury is thought to lead initially to a proinflammatoryresponse or systemic inflammatory response syndromewhich may be important as part of the normal healingprocess, but can cause excessive tissue damage and insome cases contribute to multiple organ failure.5-7 Thisinitial proinflammatory state is thought to be followed bya compensatory anti-inflammatory response syndrome,which while important for controlling inflammation, mayalso lead to an increased susceptibility to infection.5-7 Thismodel has recently been questioned, as gene expressionprofiles of trauma patients suggest that the pro- and anti-inflammatory responses occur simultaneously.8 Humanstudies of peripheral cytokines after trauma haveobserved increases in interleukin (IL)-6, IL-10, IL-1Ra, andIL-8 at various time points after injury, while reports ofincreases in tumor necrosis factor (TNF)-a and IL-4 havebeen inconsistent.9-22 Changes in levels of different anti-body isotype levels have also been reported, withincreased immunoglobulin (Ig)E and decreased IgM.22-24
Human ex vivo cellular assays with peripheral blood fromtrauma patients have found decreased HLA-DR expres-sion and increased IL-6 and IL-10 production by mono-cytes, increased regulatory T-cell activity, and alteredT-cell effector functions.25-32 Mouse models of traumaticinjury using various combinations of femur fractures,hemorrhagic shock, laparotomy, or burn (all under anes-thesia) have found increases in serum IL-6, IL-10, andother cytokines, as well as increased regulatory T-cellactivity, reduced ex vivo dendritic cell activation, andaltered ex vivo T-cell cytokine profiles.33-41 In mouse andrat models of hemorrhage where animals are anesthetizedand either bled a fixed volume or bled to a fixed reducedblood pressure, short-term defects in IL-2 production andT-cell proliferative capacity have been observed.42-44
Sex and age are also contributing factors to theimmune response to trauma. In humans several studieshave found that men have an increased risk of death,sepsis, and multiple organ dysfunction syndrome com-pared to women and that this sex difference is agedependent.45-49 Mouse models of trauma utilizing ovariec-tomized females, castrated males, and administration ofsex hormones show that the sex differences can be over-come by changing the balance of sex hormones, withandrogens suppressing responses to septic challenges andestrogens enhancing these responses.50,51
Transfusion of whole blood or blood components isperhaps the most commonly performed type of allogeneictransplantation and in itself represents a major immuno-logic intervention. In some contexts, transfusion has beenshown to have an immunosuppressive effect. This immu-nosuppression can contribute to positive clinical out-comes such as reduced transplant graft failure, but hasalso been suggested to increase cancer growth and
susceptibility to infection in some patient populationsand animal models.52-57 In spite of the immunologic con-sequences of both trauma and transfusion, and the preva-lence of transfusion among injured patients, very fewstudies have directly examined the combined effect oftraumatic injury and allogeneic transfusion on immunity.Transfusion has been shown to lead to long-term survivalof allogeneic donor cells in 10% to 15% of trauma patients,but this is not observed in surgical patients receivingtransfusions, suggesting that there is some form of uniqueimmunosuppression occurring with traumatic injury.58
Mouse burn models have observed reduced resistance toinfection and altered NK-cell activity after burn and trans-fusion compared with burn alone,59,60 and a murine modelof hemorrhage found that transfusion modulated the exvivo cytokine production of T cells from hemorrhagedmice,61 but all of these studies used anesthetized animals,potentially missing some of the effects of the stressresponse to traumatic injury.
In this study a wide range of immunomodulatoryplasma proteins was evaluated in serial samples collectedfrom transfused and nontransfused trauma patients,enrolled upon arrival in the emergency room and followedfor up to 1 year after injury. A mouse model was developedto investigate the distinct contributions of traumaticblood loss (without anesthesia) and transfusion onimmunity.
MATERIALS AND METHODS
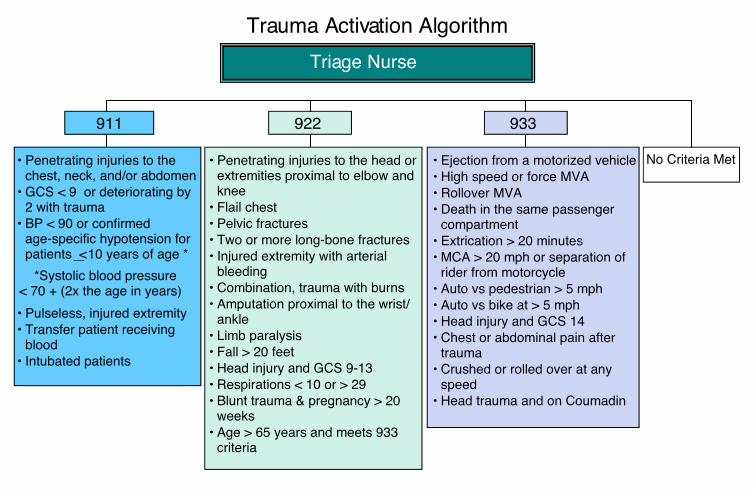
Human subjectsTrauma patients were recruited from the University ofCalifornia Davis Medical Center (UCDMC, Sacramento,CA) as part of a larger study of microchimerism, the per-sistence of donor blood cells in transfusion recipients.From this larger cohort, 56 subjects (39 transfused and 17nontransfused) were selected for cytokine analysis basedon serial sample availability. The University of CaliforniaDavis Institutional Review Board approved the humansubjects portion of our study. Upon arrival to the UCDMCEmergency Department, all injured patients meeting spe-cific institutional triage criteria (Fig. S1, available as sup-porting information in the online version of this paper)associated with a relatively high likelihood of severe injurybetween November 2006 and August 2010 were evaluatedfor enrollment. We excluded subjects less than 12 years ofage, prisoners, patients from whom we were unable toobtain a blood sample before the first transfusion of bloodproducts, and patients who had undergone previoustransplantation (solid organ or hematopoietic transplant).Because the focus of the microchimerism study was onsubjects likely to survive long term after traumatic injury,we also excluded patients who died within 7 days afterinjury. We collected an initial blood sample on all eligiblesubjects and then retained in the study those subjects who
JACKMAN ET AL.
2534 TRANSFUSION Volume 52, December 2012

subsequently provided informed consent. We approachedfor consent all transfused subjects and a subset of non-transfused subjects with comparable significant injuries,enrolled at an approximate ratio of one nontransfusedsubject per four transfused subjects over the life of thestudy. Transfused blood products were all leukoreduced.
Modeling traumatic blood loss and transfusionin miceFemale BALB/cJ and C57Bl/6J mice were purchased fromThe Jackson Laboratory (Bar Harbor, ME) and allowed toacclimate for a minimum of 2 weeks before use at age 9weeks and 2 to 3 months, respectively. Mice were main-tained in a specific pathogen-free vivarium under barrierconditions at Blood Systems Research Institute (San Fran-cisco, CA). All research was performed with approval andoversight of the Institutional Animal Care and Use Com-mittee at ISIS Services LLC (San Carlos, CA).
To model traumatic blood loss, BALB/cJ mice werebled 25% to 30% of their total estimated blood volumebased on weight. The volume of blood to be removed wascalculated as
25% in L weight in g 20 ;µ( ) = ( ) ! ( )
30% in L weight in g 24 .µ( ) = ( ) ! ( )
Mice were bled at the submandibular vein cluster using asterile lancet without anesthesia. This method allows forcollection of high blood volumes as well as rapid control ofbleeding once the desired volume is collected.62 Afterbleeding, mice were placed in a recovery area with gentleheat and close monitoring until able to resume groomingand other light activity. C57Bl/6J blood donor mice wereexsanguinated via orbital enucleation under deep anes-thesia. Blood was collected from multiple mice into asingle tube containing CPDA-1 (taken from a 500 mL WBFdouble CPDA-1 blood bag unit, Pall Medical, East Hills,NY) at 14% final volume and gently mixed between collec-tions. Donor blood was centrifuged, and then a portion ofthe plasma fraction was removed to bring to a hematocritof approximately 75% and gently mixed. Blood transfu-sions consisting of 100 mL of fresh RBCs (<6 hr old) and400 mL of sterile 0.9% sodium chloride (Baxter HealthcareCorp., Deerfield, IL) were administered intravenously bytail vein, as were injections of normal saline alone(500 mL). Blood was administered on the same day as col-lection to simplify the model and avoid introducingfurther variation that might be associated with age ofblood products.
Sample collection and processingHuman blood samples were collected into 10 mL plasticspray-coated K2EDTA tubes (Vacutainer, BD, Franklin
Lakes, NJ) at UCDMC and shipped via overnight courierservice (FedEx) to Blood Systems Research Institute. Uponarrival, the plasma fraction was isolated, aliquoted, andstored at -80°C until use. Murine blood samples were col-lected by exsanguination via orbital enucleation underanesthesia (isoflurane) into tubes without anticoagulantadditives. Blood samples were allowed to clot for 20 to 30minutes after collection and then centrifuged at highspeed to isolate serum. Serum was aliquoted and stored at-80°C until use.
Cytokine detectionCytokines were measured using a multiplex analyzer(Luminex 100 platform, Luminex, Austin, TX) and com-puter software (BioManager, Bio-Rad, Hercules, CA) foranalysis. The following multiplexing kits were purchasedfrom Millipore (Billerica, MA): the Milliplex MAP humancytokine and chemokine kit containing IL-1a, IL-1Ra,IL-9, IL-12p40, IL-15, IL-17, epidermal growth factor(EGF), eotaxin, fibroblast growth factor-2 (FGF-2), fracta-lkine, interferon (IFN)-g–inducible protein 10 (IP-10;CXCL10), monocyte chemotactic protein (MCP)-1,MCP-3, macrophage-derived chemokine (MDC), mac-rophage inflammatory protein (MIP)-1a, MIP-1b, sIL-2Ra, TNF-b, and vascular endothelial growth factor(VEGF); the Milliplex MAP high-sensitivity human cytok-ine kit containing IL-1b, IL-2, IL-4, IL-5, IL-6, IL-7, IL-8,IL-10, IL-12p70, IL-13, IFN-g, granulocyte-macrophage–colony-stimulating factor (GM-CSF), and TNF-a; the Mil-liplex MAP human sepsis and apoptosis kit containingsoluble vascular cell adhesion molecule-1 (sVCAM-1),soluble intercellular adhesion molecule 1 (sICAM-1),sFas, sFasL, macrophage migration inhibitory factor(MIF), and total plasminogen activator inhibitor-1 (tPAI-1); the Milliplex MAP human cardiovascular diseasePanel 1 kit containing soluble E-selectin (sE-selectin),matrix metalloproteinase (MMP)-9, and myeloperoxi-dase (MPO); the Milliplex MAP mouse cytokine andchemokine Panel 1 kit containing eotaxin, GM-CSF,IFN-g, IL-10, IL-12 (p40), IL-12 (p70), IL-13, IL-15, IL-17,IL-1a, IL-1b, IL-2, IL-4, IL-5, IL-6, IL-7, IP-10, KC(CXCL1), M-CSF, MCP-1, MIP-1a, MIP-1b, MIP-2, andTNF-a; and the Milliplex MAP mouse cardiovasculardisease Panel 1 containing MMP-9, tPAI-1, sE-selectin,sICAM-1, and sVCAM-1. Samples were run in dupli-cate according to manufacturer instructions. For thehuman studies, four plates were required foreach kit so the same lots were used for all platesfrom the same kit, and additional internal controls wereadded to all plates to confirm minimal plate-to-platevariation. For the murine studies, each experiment wasfit on a single plate for each kit to avoid plate-to-platevariation.
IMMUNE RESPONSE TO TRAUMA AND TRANSFUSION
Volume 52, December 2012 TRANSFUSION 2535

Statistical analysis
Human data
Cytokine and clinical data were loaded into a database(SQL, MySQL Version 5.1.7, http://www.mysql.com/;using SQuirreLSQL, Client Version 2.6.9). Undetectablevalues were assigned a value of zero for the purposes ofanalysis. Data were then imported into a statistical analy-sis package (Stata Special Edition, Version 10.1, StataCorp,College Station, TX). While all follow-up samples wereprocessed within 24 hours from collection, a subset ofindex samples were (out of necessity) collected overweekends and holidays and were therefore delayed in pro-cessing for an extra 24 to 48 hours. To address this, 10additional trauma index samples received within 24 hoursafter collection were subaliquoted with portions pro-cessed immediately and 24, 48, and 72 hours after arrivaland analyzed for cytokine levels.63 For all proteins with asignificant change over time, older index samples weredropped from the analysis. Concentration of all proteinsin pg/mL were log transformed using ln(concentration +1). For reference, untransformed median values withinterquartile ranges are reported for all cytokines on Day 0in Table S1 (available as supporting information in theonline version of this paper). Age, injury severity score(ISS), number of units transfused in the first 48 hours, andtime were converted into categorical variables. General-ized estimating equations (GEE) was used to model thechanges in each cytokine over time. This method allowsfor repeated measures, accounting both for variationbetween individuals and for differences between indi-viduals over time. Exchangeable correlation was used toallow for gaps in sample collection or loss to follow-upover time. For modeling time since trauma, linear, qua-dratic, and cubic functions were evaluated for eachprotein, and the function with the best overall fit waschosen. In addition to time since trauma, the followingindependent variables were included in the model: age(<25, 25-34, 35-44, or >44 years), sex (male or female), ISS(1-9, 10-14, 16-24, 25-43, 45-75), number of units trans-fused in the first 48 hours (expressed as the indicator vari-ables “no transfusion” and “large transfusion” [>4 units]each compared against “modest transfusion” [!4 units]),injury type (blunt or penetrating), and microchimeric (yesor no). The Wald chi-square statistic was used to assessoverall significance with a cutoff of p value of less than0.05. This tests against the hypothesis that none of thevariables have an effect on the protein concentrations.A coefficient for an individual variable was consideredsignificant if the p value was less than 0.05. Graphswere generated in a statistical analysis package (StataSpecial Edition, Version 10.1, StataCorp). Maximum andminimum values over a set range for each time functiongenerated by the model were calculated and a percentageincrease was calculated as
100 e e e 1Maximum Minimum Minimum! "( ) [ ]"( ).
Animal dataConcentrations of each protein analyzed were comparedbetween treatment groups using one-way analysis of vari-ance (ANOVA); each treatment group was then comparedto the untreated control group using Dunnett’s multiple-comparison posttest. A graphing and analysis softwarepackage (Prism, Version 5.0a, GraphPad Software, Inc., LaJolla, CA) was used to generate figures.
RESULTS
Human response to trauma and transfusionTo assess the human immune response to trauma andtransfusion, serial blood samples were collected fromtrauma patients at UCDMC. The first blood sample wascollected upon arrival in the emergency room, withfollow-up samples collected at regular intervals up to1 year after injuries. Of the 56 subjects included in theanalysis, 75% were male, 73% suffered blunt trauma, and70% were transfused. The median age was 30.5 years, themedian ISS was 17, and a median of 4 RBC units weregiven in the first 48 hours after injury to those whoreceived a transfusion (Table 1).
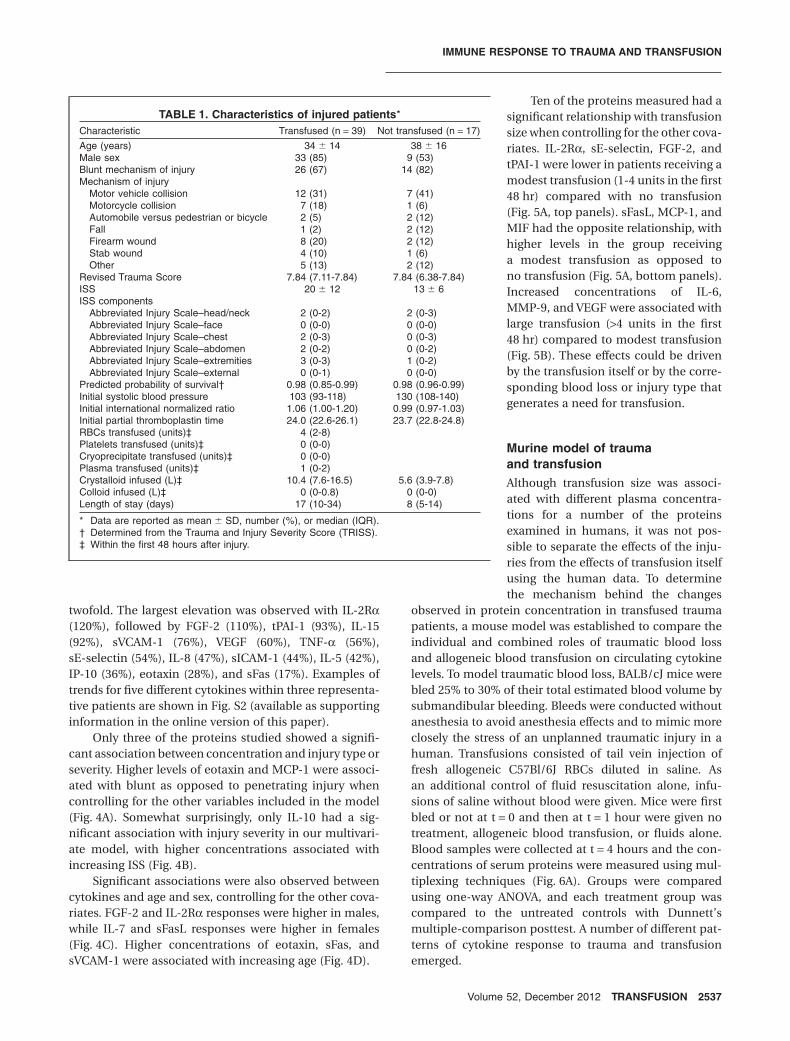
Plasma was isolated from the blood and assayed for41 cytokines, chemokines, and other immunomodulatingproteins using multiplexing techniques. GEE models wereused to examine the change in protein concentrationsover time for each protein, controlling for the includedclinical data. Thirty-one of the proteins had a significantchange over time after trauma, controlling for the othervariables in the model. Of these, eight proteins wereelevated at the time of arrival in the emergency room: IL-6,IL-10, IL-1Ra, MIF, MPO, MCP-1, MMP-9, and sFasL(Fig. 1). The elevations observed for IL-6 and IL-10 werethe highest, with the models predicting a percentageincrease at maximum (over minimum) of 550 and 450%,respectively. IL-1Ra showed the next largest increase at330%, followed by MPO (210%), MMP-9 (200%), sFasL(98%), MIF (83%), and MCP-1 (61%).
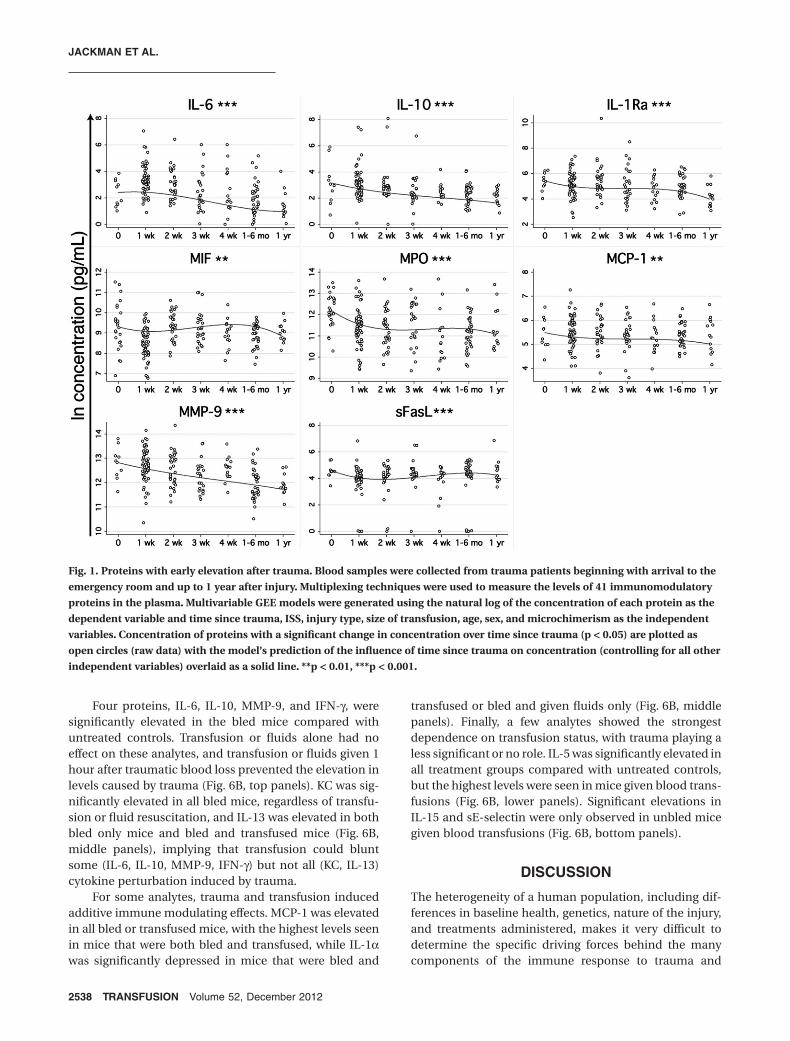
Nine of the proteins with a significant change overtime since trauma showed a depression at the time ofarrival in the emergency room (Fig. 2). The largestdecrease based on the percentage change of the predictedmaximum value over minimum was fractalkine (490%),followed by IL-1a (460%), EGF (210%), IL-7 (190%), IL-9(130%), IL-17 (120%), TNF-b (110%), MIP-1a (90%), andMDC (52%).
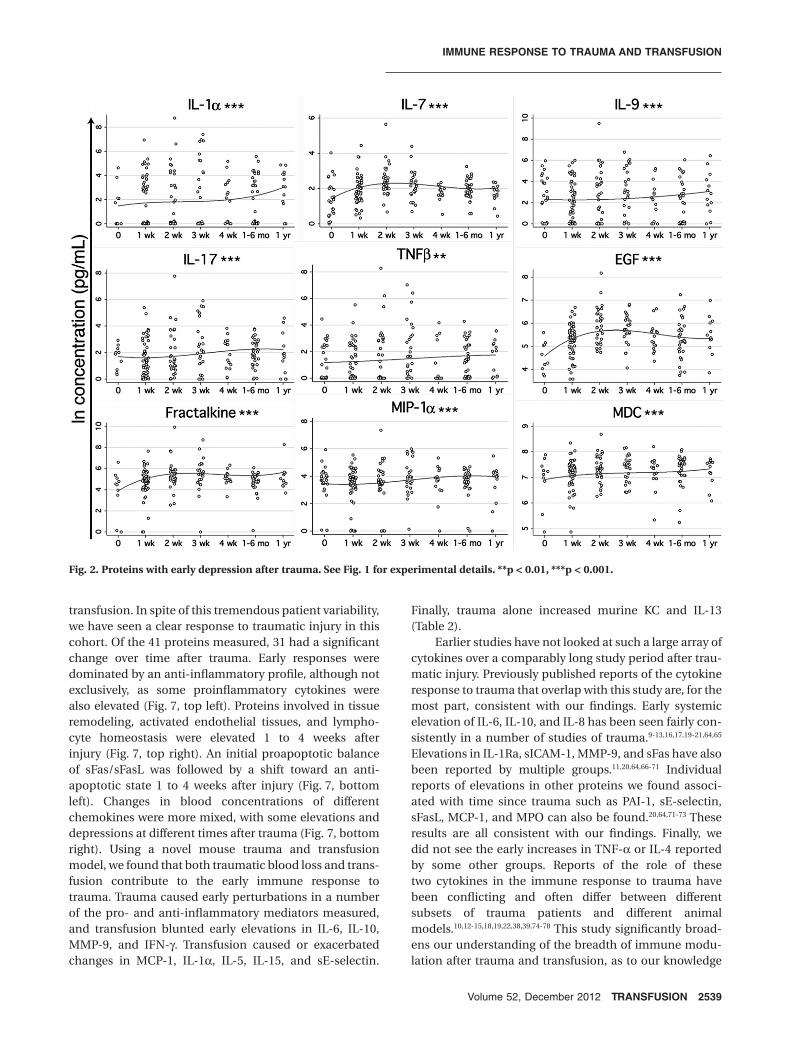
The remaining 14 proteins with a significant changeover time since trauma had a delayed elevation pattern,peaking between 1 and 4 weeks after trauma (Fig. 3). Therange of these elevations was not as wide as what wasobserved for the early responses; most of the proteins withthis late elevation pattern had an increase less than
JACKMAN ET AL.
2536 TRANSFUSION Volume 52, December 2012
twofold. The largest elevation was observed with IL-2Ra(120%), followed by FGF-2 (110%), tPAI-1 (93%), IL-15(92%), sVCAM-1 (76%), VEGF (60%), TNF-a (56%),sE-selectin (54%), IL-8 (47%), sICAM-1 (44%), IL-5 (42%),IP-10 (36%), eotaxin (28%), and sFas (17%). Examples oftrends for five different cytokines within three representa-tive patients are shown in Fig. S2 (available as supportinginformation in the online version of this paper).
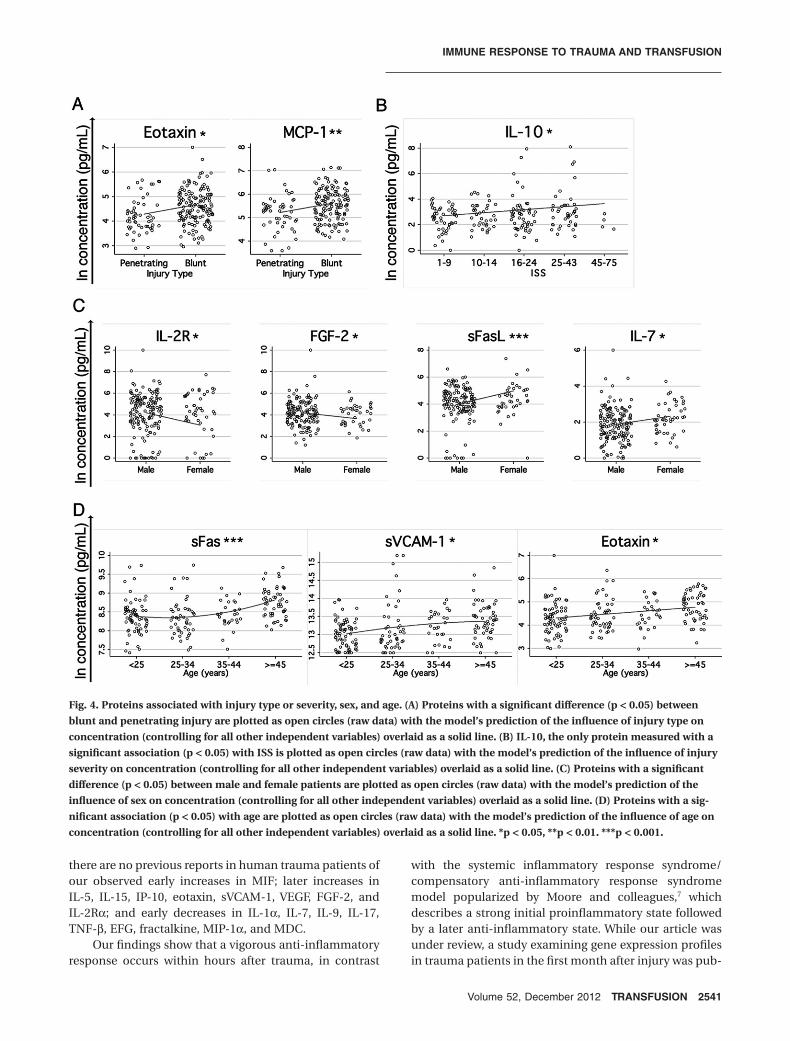
Only three of the proteins studied showed a signifi-cant association between concentration and injury type orseverity. Higher levels of eotaxin and MCP-1 were associ-ated with blunt as opposed to penetrating injury whencontrolling for the other variables included in the model(Fig. 4A). Somewhat surprisingly, only IL-10 had a sig-nificant association with injury severity in our multivari-ate model, with higher concentrations associated withincreasing ISS (Fig. 4B).
Significant associations were also observed betweencytokines and age and sex, controlling for the other cova-riates. FGF-2 and IL-2Ra responses were higher in males,while IL-7 and sFasL responses were higher in females(Fig. 4C). Higher concentrations of eotaxin, sFas, andsVCAM-1 were associated with increasing age (Fig. 4D).
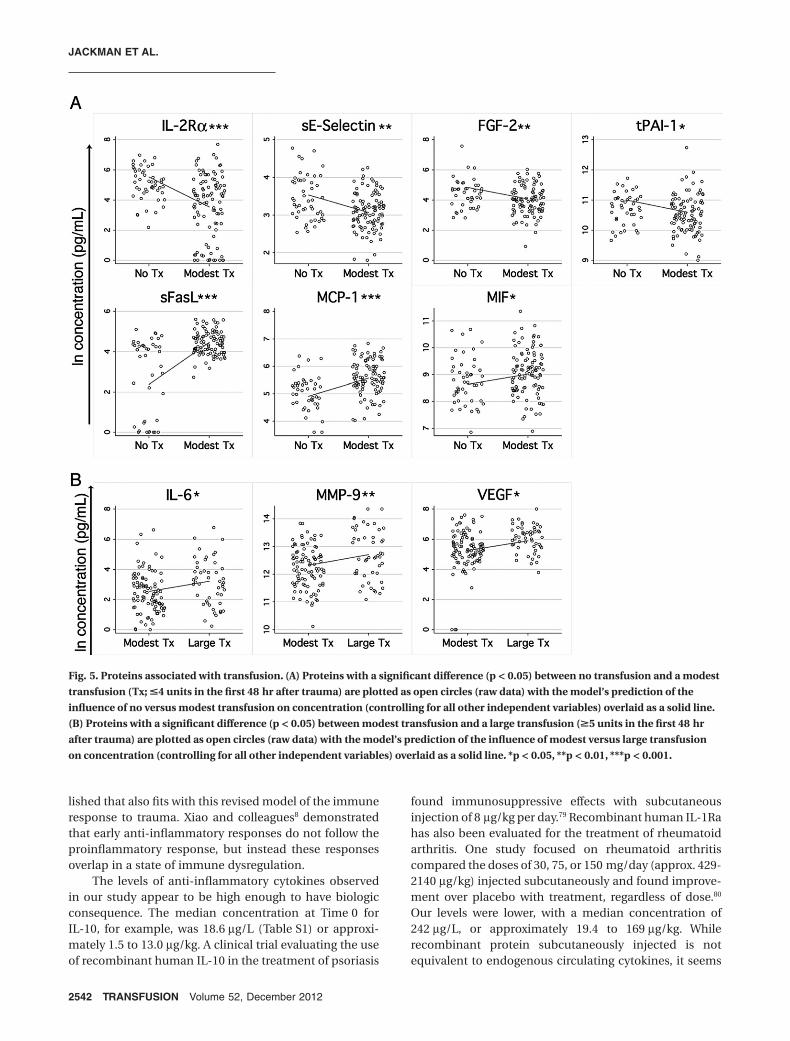
Ten of the proteins measured had asignificant relationship with transfusionsize when controlling for the other cova-riates. IL-2Ra, sE-selectin, FGF-2, andtPAI-1 were lower in patients receiving amodest transfusion (1-4 units in the first48 hr) compared with no transfusion(Fig. 5A, top panels). sFasL, MCP-1, andMIF had the opposite relationship, withhigher levels in the group receivinga modest transfusion as opposed tono transfusion (Fig. 5A, bottom panels).Increased concentrations of IL-6,MMP-9, and VEGF were associated withlarge transfusion (>4 units in the first48 hr) compared to modest transfusion(Fig. 5B). These effects could be drivenby the transfusion itself or by the corre-sponding blood loss or injury type thatgenerates a need for transfusion.
Murine model of traumaand transfusionAlthough transfusion size was associ-ated with different plasma concentra-tions for a number of the proteinsexamined in humans, it was not pos-sible to separate the effects of the inju-ries from the effects of transfusion itselfusing the human data. To determinethe mechanism behind the changes
observed in protein concentration in transfused traumapatients, a mouse model was established to compare theindividual and combined roles of traumatic blood lossand allogeneic blood transfusion on circulating cytokinelevels. To model traumatic blood loss, BALB/cJ mice werebled 25% to 30% of their total estimated blood volume bysubmandibular bleeding. Bleeds were conducted withoutanesthesia to avoid anesthesia effects and to mimic moreclosely the stress of an unplanned traumatic injury in ahuman. Transfusions consisted of tail vein injection offresh allogeneic C57Bl/6J RBCs diluted in saline. Asan additional control of fluid resuscitation alone, infu-sions of saline without blood were given. Mice were firstbled or not at t = 0 and then at t = 1 hour were given notreatment, allogeneic blood transfusion, or fluids alone.Blood samples were collected at t = 4 hours and the con-centrations of serum proteins were measured using mul-tiplexing techniques (Fig. 6A). Groups were comparedusing one-way ANOVA, and each treatment group wascompared to the untreated controls with Dunnett’smultiple-comparison posttest. A number of different pat-terns of cytokine response to trauma and transfusionemerged.
TABLE 1. Characteristics of injured patients*Characteristic Transfused (n = 39) Not transfused (n = 17)Age (years) 34 " 14 38 " 16Male sex 33 (85) 9 (53)Blunt mechanism of injury 26 (67) 14 (82)Mechanism of injury
Motor vehicle collision 12 (31) 7 (41)Motorcycle collision 7 (18) 1 (6)Automobile versus pedestrian or bicycle 2 (5) 2 (12)Fall 1 (2) 2 (12)Firearm wound 8 (20) 2 (12)Stab wound 4 (10) 1 (6)Other 5 (13) 2 (12)
Revised Trauma Score 7.84 (7.11-7.84) 7.84 (6.38-7.84)ISS 20 " 12 13 " 6ISS components
Abbreviated Injury Scale–head/neck 2 (0-2) 2 (0-3)Abbreviated Injury Scale–face 0 (0-0) 0 (0-0)Abbreviated Injury Scale–chest 2 (0-3) 0 (0-3)Abbreviated Injury Scale–abdomen 2 (0-2) 0 (0-2)Abbreviated Injury Scale–extremities 3 (0-3) 1 (0-2)Abbreviated Injury Scale–external 0 (0-1) 0 (0-0)
Predicted probability of survival† 0.98 (0.85-0.99) 0.98 (0.96-0.99)Initial systolic blood pressure 103 (93-118) 130 (108-140)Initial international normalized ratio 1.06 (1.00-1.20) 0.99 (0.97-1.03)Initial partial thromboplastin time 24.0 (22.6-26.1) 23.7 (22.8-24.8)RBCs transfused (units)‡ 4 (2-8)Platelets transfused (units)‡ 0 (0-0)Cryoprecipitate transfused (units)‡ 0 (0-0)Plasma transfused (units)‡ 1 (0-2)Crystalloid infused (L)‡ 10.4 (7.6-16.5) 5.6 (3.9-7.8)Colloid infused (L)‡ 0 (0-0.8) 0 (0-0)Length of stay (days) 17 (10-34) 8 (5-14)
* Data are reported as mean " SD, number (%), or median (IQR).† Determined from the Trauma and Injury Severity Score (TRISS).‡ Within the first 48 hours after injury.
IMMUNE RESPONSE TO TRAUMA AND TRANSFUSION
Volume 52, December 2012 TRANSFUSION 2537
Four proteins, IL-6, IL-10, MMP-9, and IFN-g, weresignificantly elevated in the bled mice compared withuntreated controls. Transfusion or fluids alone had noeffect on these analytes, and transfusion or fluids given 1hour after traumatic blood loss prevented the elevation inlevels caused by trauma (Fig. 6B, top panels). KC was sig-nificantly elevated in all bled mice, regardless of transfu-sion or fluid resuscitation, and IL-13 was elevated in bothbled only mice and bled and transfused mice (Fig. 6B,middle panels), implying that transfusion could bluntsome (IL-6, IL-10, MMP-9, IFN-g) but not all (KC, IL-13)cytokine perturbation induced by trauma.
For some analytes, trauma and transfusion inducedadditive immune modulating effects. MCP-1 was elevatedin all bled or transfused mice, with the highest levels seenin mice that were both bled and transfused, while IL-1awas significantly depressed in mice that were bled and
transfused or bled and given fluids only (Fig. 6B, middlepanels). Finally, a few analytes showed the strongestdependence on transfusion status, with trauma playing aless significant or no role. IL-5 was significantly elevated inall treatment groups compared with untreated controls,but the highest levels were seen in mice given blood trans-fusions (Fig. 6B, lower panels). Significant elevations inIL-15 and sE-selectin were only observed in unbled micegiven blood transfusions (Fig. 6B, bottom panels).
DISCUSSION
The heterogeneity of a human population, including dif-ferences in baseline health, genetics, nature of the injury,and treatments administered, makes it very difficult todetermine the specific driving forces behind the manycomponents of the immune response to trauma and
Fig. 1. Proteins with early elevation after trauma. Blood samples were collected from trauma patients beginning with arrival to theemergency room and up to 1 year after injury. Multiplexing techniques were used to measure the levels of 41 immunomodulatoryproteins in the plasma. Multivariable GEE models were generated using the natural log of the concentration of each protein as thedependent variable and time since trauma, ISS, injury type, size of transfusion, age, sex, and microchimerism as the independentvariables. Concentration of proteins with a significant change in concentration over time since trauma (p < 0.05) are plotted asopen circles (raw data) with the model’s prediction of the influence of time since trauma on concentration (controlling for all otherindependent variables) overlaid as a solid line. **p < 0.01, ***p < 0.001.
JACKMAN ET AL.
2538 TRANSFUSION Volume 52, December 2012
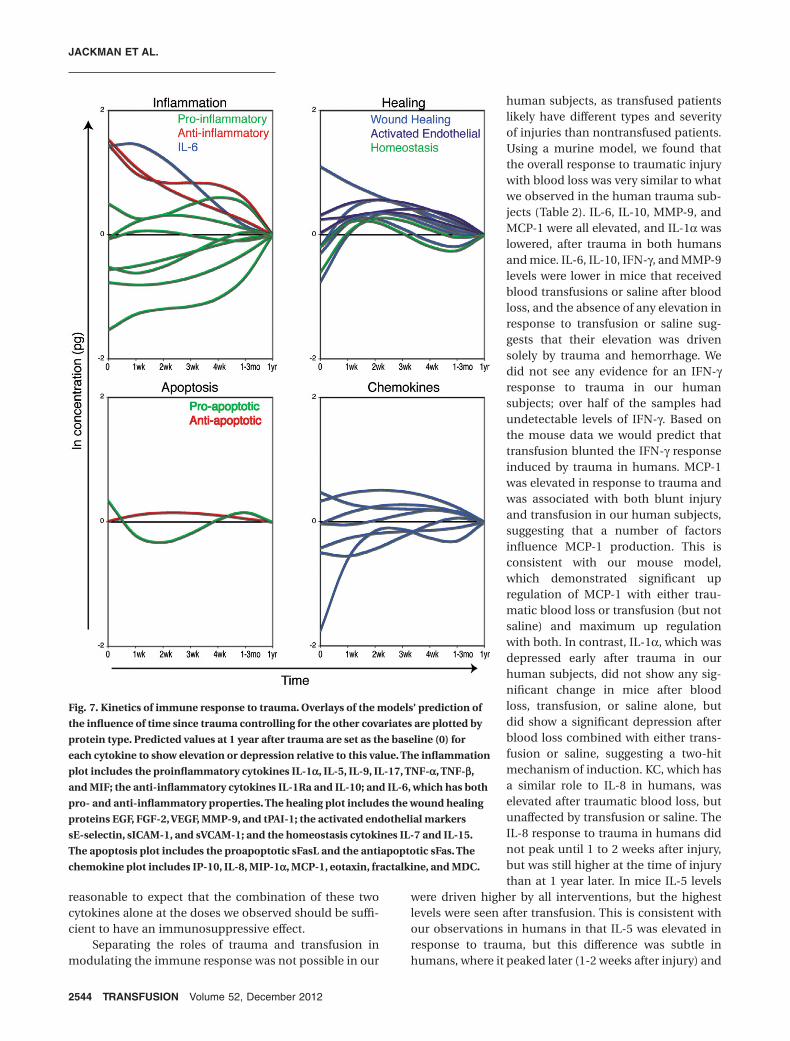
transfusion. In spite of this tremendous patient variability,we have seen a clear response to traumatic injury in thiscohort. Of the 41 proteins measured, 31 had a significantchange over time after trauma. Early responses weredominated by an anti-inflammatory profile, although notexclusively, as some proinflammatory cytokines werealso elevated (Fig. 7, top left). Proteins involved in tissueremodeling, activated endothelial tissues, and lympho-cyte homeostasis were elevated 1 to 4 weeks afterinjury (Fig. 7, top right). An initial proapoptotic balanceof sFas/sFasL was followed by a shift toward an anti-apoptotic state 1 to 4 weeks after injury (Fig. 7, bottomleft). Changes in blood concentrations of differentchemokines were more mixed, with some elevations anddepressions at different times after trauma (Fig. 7, bottomright). Using a novel mouse trauma and transfusionmodel, we found that both traumatic blood loss and trans-fusion contribute to the early immune response totrauma. Trauma caused early perturbations in a numberof the pro- and anti-inflammatory mediators measured,and transfusion blunted early elevations in IL-6, IL-10,MMP-9, and IFN-g. Transfusion caused or exacerbatedchanges in MCP-1, IL-1a, IL-5, IL-15, and sE-selectin.
Finally, trauma alone increased murine KC and IL-13(Table 2).
Earlier studies have not looked at such a large array ofcytokines over a comparably long study period after trau-matic injury. Previously published reports of the cytokineresponse to trauma that overlap with this study are, for themost part, consistent with our findings. Early systemicelevation of IL-6, IL-10, and IL-8 has been seen fairly con-sistently in a number of studies of trauma.9-13,16,17,19-21,64,65
Elevations in IL-1Ra, sICAM-1, MMP-9, and sFas have alsobeen reported by multiple groups.11,20,64,66-71 Individualreports of elevations in other proteins we found associ-ated with time since trauma such as PAI-1, sE-selectin,sFasL, MCP-1, and MPO can also be found.20,64,71-73 Theseresults are all consistent with our findings. Finally, wedid not see the early increases in TNF-a or IL-4 reportedby some other groups. Reports of the role of thesetwo cytokines in the immune response to trauma havebeen conflicting and often differ between differentsubsets of trauma patients and different animalmodels.10,12-15,18,19,22,38,39,74-78 This study significantly broad-ens our understanding of the breadth of immune modu-lation after trauma and transfusion, as to our knowledge
Fig. 2. Proteins with early depression after trauma. See Fig. 1 for experimental details. **p < 0.01, ***p < 0.001.
IMMUNE RESPONSE TO TRAUMA AND TRANSFUSION
Volume 52, December 2012 TRANSFUSION 2539
Fig. 3. Proteins with late elevation after trauma. See Fig. 1 for experimental details. *p < 0.05, **p < 0.01, ***p < 0.001.
JACKMAN ET AL.
2540 TRANSFUSION Volume 52, December 2012
there are no previous reports in human trauma patients ofour observed early increases in MIF; later increases inIL-5, IL-15, IP-10, eotaxin, sVCAM-1, VEGF, FGF-2, andIL-2Ra; and early decreases in IL-1a, IL-7, IL-9, IL-17,TNF-b, EFG, fractalkine, MIP-1a, and MDC.
Our findings show that a vigorous anti-inflammatoryresponse occurs within hours after trauma, in contrast
with the systemic inflammatory response syndrome/compensatory anti-inflammatory response syndromemodel popularized by Moore and colleagues,7 whichdescribes a strong initial proinflammatory state followedby a later anti-inflammatory state. While our article wasunder review, a study examining gene expression profilesin trauma patients in the first month after injury was pub-
Fig. 4. Proteins associated with injury type or severity, sex, and age. (A) Proteins with a significant difference (p < 0.05) betweenblunt and penetrating injury are plotted as open circles (raw data) with the model’s prediction of the influence of injury type onconcentration (controlling for all other independent variables) overlaid as a solid line. (B) IL-10, the only protein measured with asignificant association (p < 0.05) with ISS is plotted as open circles (raw data) with the model’s prediction of the influence of injuryseverity on concentration (controlling for all other independent variables) overlaid as a solid line. (C) Proteins with a significantdifference (p < 0.05) between male and female patients are plotted as open circles (raw data) with the model’s prediction of theinfluence of sex on concentration (controlling for all other independent variables) overlaid as a solid line. (D) Proteins with a sig-nificant association (p < 0.05) with age are plotted as open circles (raw data) with the model’s prediction of the influence of age onconcentration (controlling for all other independent variables) overlaid as a solid line. *p < 0.05, **p < 0.01. ***p < 0.001.
IMMUNE RESPONSE TO TRAUMA AND TRANSFUSION
Volume 52, December 2012 TRANSFUSION 2541
lished that also fits with this revised model of the immuneresponse to trauma. Xiao and colleagues8 demonstratedthat early anti-inflammatory responses do not follow theproinflammatory response, but instead these responsesoverlap in a state of immune dysregulation.
The levels of anti-inflammatory cytokines observedin our study appear to be high enough to have biologicconsequence. The median concentration at Time 0 forIL-10, for example, was 18.6 mg/L (Table S1) or approxi-mately 1.5 to 13.0 mg/kg. A clinical trial evaluating the useof recombinant human IL-10 in the treatment of psoriasis
found immunosuppressive effects with subcutaneousinjection of 8 mg/kg per day.79 Recombinant human IL-1Rahas also been evaluated for the treatment of rheumatoidarthritis. One study focused on rheumatoid arthritiscompared the doses of 30, 75, or 150 mg/day (approx. 429-2140 mg/kg) injected subcutaneously and found improve-ment over placebo with treatment, regardless of dose.80
Our levels were lower, with a median concentration of242 mg/L, or approximately 19.4 to 169 mg/kg. Whilerecombinant protein subcutaneously injected is notequivalent to endogenous circulating cytokines, it seems
Fig. 5. Proteins associated with transfusion. (A) Proteins with a significant difference (p < 0.05) between no transfusion and a modesttransfusion (Tx; !4 units in the first 48 hr after trauma) are plotted as open circles (raw data) with the model’s prediction of theinfluence of no versus modest transfusion on concentration (controlling for all other independent variables) overlaid as a solid line.(B) Proteins with a significant difference (p < 0.05) between modest transfusion and a large transfusion ("5 units in the first 48 hrafter trauma) are plotted as open circles (raw data) with the model’s prediction of the influence of modest versus large transfusionon concentration (controlling for all other independent variables) overlaid as a solid line. *p < 0.05, **p < 0.01, ***p < 0.001.
JACKMAN ET AL.
2542 TRANSFUSION Volume 52, December 2012
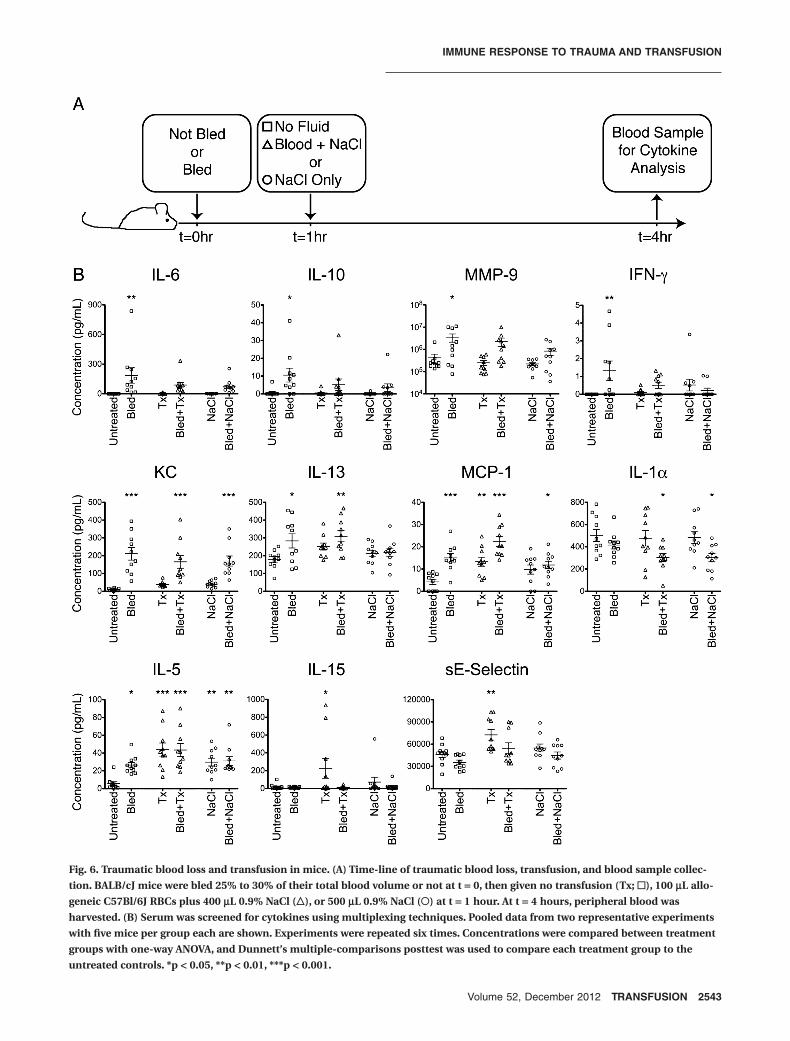
Fig. 6. Traumatic blood loss and transfusion in mice. (A) Time-line of traumatic blood loss, transfusion, and blood sample collec-tion. BALB/cJ mice were bled 25% to 30% of their total blood volume or not at t = 0, then given no transfusion (Tx; !), 100 mL allo-geneic C57Bl/6J RBCs plus 400 mL 0.9% NaCl ("), or 500 mL 0.9% NaCl (#) at t = 1 hour. At t = 4 hours, peripheral blood washarvested. (B) Serum was screened for cytokines using multiplexing techniques. Pooled data from two representative experimentswith five mice per group each are shown. Experiments were repeated six times. Concentrations were compared between treatmentgroups with one-way ANOVA, and Dunnett’s multiple-comparisons posttest was used to compare each treatment group to theuntreated controls. *p < 0.05, **p < 0.01, ***p < 0.001.
IMMUNE RESPONSE TO TRAUMA AND TRANSFUSION
Volume 52, December 2012 TRANSFUSION 2543
reasonable to expect that the combination of these twocytokines alone at the doses we observed should be suffi-cient to have an immunosuppressive effect.
Separating the roles of trauma and transfusion inmodulating the immune response was not possible in our
human subjects, as transfused patientslikely have different types and severityof injuries than nontransfused patients.Using a murine model, we found thatthe overall response to traumatic injurywith blood loss was very similar to whatwe observed in the human trauma sub-jects (Table 2). IL-6, IL-10, MMP-9, andMCP-1 were all elevated, and IL-1a waslowered, after trauma in both humansand mice. IL-6, IL-10, IFN-g, and MMP-9levels were lower in mice that receivedblood transfusions or saline after bloodloss, and the absence of any elevation inresponse to transfusion or saline sug-gests that their elevation was drivensolely by trauma and hemorrhage. Wedid not see any evidence for an IFN-gresponse to trauma in our humansubjects; over half of the samples hadundetectable levels of IFN-g. Based onthe mouse data we would predict thattransfusion blunted the IFN-g responseinduced by trauma in humans. MCP-1was elevated in response to trauma andwas associated with both blunt injuryand transfusion in our human subjects,suggesting that a number of factorsinfluence MCP-1 production. This isconsistent with our mouse model,which demonstrated significant upregulation of MCP-1 with either trau-matic blood loss or transfusion (but notsaline) and maximum up regulationwith both. In contrast, IL-1a, which wasdepressed early after trauma in ourhuman subjects, did not show any sig-nificant change in mice after bloodloss, transfusion, or saline alone, butdid show a significant depression afterblood loss combined with either trans-fusion or saline, suggesting a two-hitmechanism of induction. KC, which hasa similar role to IL-8 in humans, waselevated after traumatic blood loss, butunaffected by transfusion or saline. TheIL-8 response to trauma in humans didnot peak until 1 to 2 weeks after injury,but was still higher at the time of injurythan at 1 year later. In mice IL-5 levels
were driven higher by all interventions, but the highestlevels were seen after transfusion. This is consistent withour observations in humans in that IL-5 was elevated inresponse to trauma, but this difference was subtle inhumans, where it peaked later (1-2 weeks after injury) and
Fig. 7. Kinetics of immune response to trauma. Overlays of the models’ prediction ofthe influence of time since trauma controlling for the other covariates are plotted byprotein type. Predicted values at 1 year after trauma are set as the baseline (0) foreach cytokine to show elevation or depression relative to this value. The inflammationplot includes the proinflammatory cytokines IL-1a, IL-5, IL-9, IL-17, TNF-a, TNF-b,and MIF; the anti-inflammatory cytokines IL-1Ra and IL-10; and IL-6, which has bothpro- and anti-inflammatory properties. The healing plot includes the wound healingproteins EGF, FGF-2, VEGF, MMP-9, and tPAI-1; the activated endothelial markerssE-selectin, sICAM-1, and sVCAM-1; and the homeostasis cytokines IL-7 and IL-15.The apoptosis plot includes the proapoptotic sFasL and the antiapoptotic sFas. Thechemokine plot includes IP-10, IL-8, MIP-1a, MCP-1, eotaxin, fractalkine, and MDC.
JACKMAN ET AL.
2544 TRANSFUSION Volume 52, December 2012

showed no association with transfusion. Similarly, IL-13was elevated in bled or bled and transfused mice, but notin our human cohort. This may be the result of the moredistinct Th2-type responses to trauma in mice comparedwith humans that has been previously reported.39,74-76,78
Finally, IL-15 and sE-selectin were both elevated in themice given transfusion alone, but not the other groups.There was no transfusion only group for the human sub-jects, and the elevations in the proteins seen in humanswere at later time points than we examined in the mice.Furthermore, while our human subjects were given leuko-reduced blood, leukoreduction was not used in the mousemodel, which may have contributed to this observedresponse to transfusion alone in the mice.
Generally, saline infusion had a “rescue” effect similarto that of transfusion (Table 2). This was seen with IL-6,IL-10, MMP-9, and IFN-g, where administration of eitherblood or saline alone after trauma brought concentrationsdown to a level where they were no longer significantlydifferent from untreated controls. Similarly, the reductionin IL-1a seen after trauma and transfusion was also seenwith trauma and saline. For other cytokines, the effectsdriven by transfusion could not be replicated by saline,suggesting that transfusion of fresh allogeneic blood canmodulate even early immune responses (Table 2). Thispattern was seen with MCP-1 and IL-5, where salineeffects were weaker than what was observed with bloodand with IL-15 and sE-selectin, where elevation was onlyseen after blood transfusion alone.
Our murine model differs fromexisting animal models of traumaticinjury in that it involves less tissuedamage than those involving femurfracture or large lacerations, enabling usto ethically bleed the mice withoutanesthesia. The disadvantage of oursystem is that the tissue damage isprobably less than that of our clinicalpatients, although the bleeding processdoes involve a 5-mm-deep stab requir-ing moderate force for delivery. Theadvantage of this model is that it moreclosely mimics the stress of accidentalinjury than those requiring anesthesia,which arguably mimic surgical injury.Our study examined a larger array ofcytokines and involved a different typeof injury than has been previouslyassessed in other murine models oftrauma, making it difficult to fullycompare our results with models usinganesthetized animals. While trends ofelevated IL-6, IL-10, MCP-1, and KCwere similar to what has been observedin models utilizing anesthesia,36,38
further work is required to determine if anesthesia modu-lates the immune response to trauma.
One of the unanticipated findings of the current studywas that IL-1a, IL-7, IL-9, IL-17, TNF-b, EGF, fractalkine,MIP-1a, and MDC were depressed early after trauma inhumans (the early levels were lower than late “baseline”time points; Fig. 2). We believe that this is the first report ofreduced concentrations of circulating cytokines aftertrauma, although there are reports of reduced capacity ofcells from trauma patients or injured mice to producecytokines ex vivo.32,37,75 Since it is not possible to collectsamples before injury in humans, and because variation ishigh between individuals, we determined that samplescollected months after injury were likely to be the closestto baseline for these patients. An alternative, less likelyexplanation to the observed trends is that they are actuallydelayed responses to trauma that persist for up to 1 year. Itis worth noting, however, that of the cytokines measuredboth in mice and in humans, the cytokine with the largestpercentage decrease in our trauma patients, IL-1a, wasalso significantly reduced in our mouse model aftertrauma and transfusion.
In our cohort the severity (measured by ISS) and typeof injury (blunt versus penetrating) did not have a signifi-cant effect on the concentrations of most of the cytokineswe evaluated. A number of studies have linked con-centrations of various cytokines with injury severityincluding IL-10 as we observed here, but also IL-6, IL-8,and IL-4.10,15,17,21,22 One explanation for the lack of signifi-
TABLE 2. Summary of the effects of trauma and transfusion onprotein levels*
ProteinHuman
Mouse: trauma or transfusionTrauma TransfusionIL-6 # #‡ # trauma drives, fluids rescueIL-10 # # trauma drives, fluids rescueMMP-9 # #‡ # trauma drives, fluids rescueINF-g # trauma drives, fluids rescueIL-8/KC #† # trauma drivesIL-13 # trauma drivesMCP-1 # # # both trauma and tx but not NaCl drive, highest
with bothIL-1a $ $ requires both trauma and fluidsIL-5 #† # both trauma and fluids drive, highest levels
with txIL-15 #† # tx drivessE-selectin #† $ # tx drivesTNF-a #†Eotaxin #†IP-10 #†sICAM-1 #†sVCAM-1 #†tPAI-1 #† $IL-7 $IL-17 $MIP-1a $* Includes all proteins measured in both human and mouse where there was a signifi-
cant association with trauma or transfusion (tx).† Delayed change.‡ Greater than 4 units compared with less than or equal to 4 units in first 48 hours.
IMMUNE RESPONSE TO TRAUMA AND TRANSFUSION
Volume 52, December 2012 TRANSFUSION 2545

cant statistical association between severity of injuryand most of the cytokines we evaluated is that our multi-variable model controlled for potential confounders.Alternatively, the difference might be explained by theoverall severity of injury in our cohort compared withothers. For example, the median ISS in our study was 17,and Hoch and colleagues15 saw significant increases inIL-6 and IL-8 with ISS of at least 25. The lower ISS in ourcohort may be partially explained by the selection criteriaused for our study. Our cohort was selected for long-termassessment of microchimerism, so patients who diedin the first 7 days were excluded. Three proteins, IL-6,MMP-9, and VEGF, were seen at higher levels in patientsreceiving large transfusions of 5 or more units in the first48 hours after injury. While our model controlled forthe severity and type of injury to some extent by includingISS and blunt versus penetrating injury, the differencesseen in these patients may still be due more to the type ofinjury that results in a massive transfusion than fromthe transfusion itself. All three of these proteins havebeen associated with ischemia-reperfusion injuriesand/or hypoxia in different clinical and experimentalsettings.20,81-86
We also observed a few differences associated withage and sex in our trauma patient cohort. We sawincreased sFasL in women compared with men andincreasing sFas with age, which is consistent with a studyfrom Kavathia and colleagues.87 Eotaxin and sVCAM-1also increased with age in our cohort, as found in studiesin humans and rats.88-92 Similarly, the higher IL-7 levels weobserved in women are consistent with a study that found40% higher levels of IL-7 in human immunodeficiencyvirus (HIV)-positive women compared with HIV-positivemen.93 Differences in the cytokine response to traumabetween men and women may help to explain thereduced rates of death, sepsis, and organ failure observedamong women compared with men after trauma.45-49
These findings support the observation in mouse modelsthat sex hormones alter the immune response totrauma.50,51
Overall, we have demonstrated that there is a massivechange in the cytokine environment after traumaticinjury and transfusion in humans. Furthermore, we havebeen able to effectively model this change in mice, allow-ing us to distinguish between those factors regulated bytraumatic blood loss, transfusion, or both. This work sug-gests that the immune response to trauma is significantand that transfusion and fluid administration have thepotential to alter the immunologic consequences oftrauma. Given the rates of transfusion among traumapatients, the interplay of the responses to trauma andtransfusion has important implications for immunologi-cally driven clinical complications such as multiple organfailure, sepsis, transfusion-related acute lung injury, andalloimmunization.
ACKNOWLEDGMENTS
We thank Dr Marina E. Fomin for her assistance with animal careand Dr Brian S. Custer for consultation on statistical methods.
CONFLICT OF INTEREST
The authors have no competing interests.
REFERENCES
1. Hess JR, Hiippala S. Optimizing the use of blood productsin trauma care. Crit Care 2005;9 Suppl 5:S10-4.
2. Trunkey DD. History and development of trauma care inthe United States. Clin Orthop Relat Res 2000;(374):36-46.
3. Como JJ, Dutton RP, Scalea TM, Edelman BB, Hess JR.Blood transfusion rates in the care of acute trauma. Trans-fusion 2004;44:809-13.
4. US Department of Health and Human Services. The 2009national blood collection and utilization survey report.2011. Washington, DC: US Department of Health andHuman Services.
5. Giannoudis PV. Current concepts of the inflammatoryresponse after major trauma: an update. Injury 2003;34:397-404.
6. Mannick JA, Rodrick ML, Lederer JA. The immunologicresponse to injury. J Am Coll Surg 2001;193:237-44.
7. Moore FA, Sauaia A, Moore EE, Haenel JB, Burch JM,Lezotte DC. Postinjury multiple organ failure: a bimodalphenomenon. J Trauma 1996;40:501-10; discussion 510-2.
8. Xiao W, Mindrinos MN, Seok J, Cuschieri J, Cuenca AG,Gao H, Hayden DL, Hennessy L, Moore EE, Minei JP,Bankey PE, Johnson JL, Sperry J, Nathens AB, Billiar TR,West MA, Brownstein BH, Mason PH, Baker HV, FinnertyCC, Jeschke MG, Lopez MC, Klein MB, Gamelli RL, Tomp-kins RG et al. A genomic storm in critically injuredhumans. J Exp Med 2011;208:2581-90.
9. Foex BA, Lamb WR, Roberts TE, Brear SG, Macartney I,Hammer M, Brenchley PE. Early cytokine response to mul-tiple injury. Injury 1993;24:373-6.
10. Mimasaka S, Funayama M, Hashiyada M, Nata M, Tsune-nari S. Significance of levels of IL-6 and IL-8 after trauma:a study of 11 cytokines post-mortem using multipleximmunoassay. Injury 2007;38:1047-51.
11. Partrick DA, Moore FA, Moore EE, Biffl WL, Sauaia A,Barnett CC Jr., Jack A. Barney Resident Research Awardwinner. The inflammatory profile of interleukin-6,interleukin-8, and soluble intercellular adhesionmolecule-1 in postinjury multiple organ failure. Am J Surg1996;172:425-9; discussed 429-31.
12. Martin C, Boisson C, Haccoun M, Thomachot L, Mege JL.Patterns of cytokine evolution (tumor necrosis factor-alphaand interleukin-6) after septic shock, hemorrhagic shock,and severe trauma. Crit Care Med 1997;25:1813-9.
13. Endo S, Inada K, Yamada Y, Takakuwa T, Kasai T, Nakae H,
JACKMAN ET AL.
2546 TRANSFUSION Volume 52, December 2012

Yoshida M, Ceska M. Plasma endotoxin and cytokine con-centrations in patients with hemorrhagic shock. Crit CareMed 1994;22:949-55.
14. Ferguson KL, Taheri P, Rodriguez J, Tonapi V, Cardellio A,Dechert R. Tumor necrosis factor activity increases inthe early response to trauma. Acad Emerg Med 1997;4:1035-40.
15. Hoch RC, Rodriguez R, Manning T, Bishop M, Mead P,Shoemaker WC, Abraham E. Effects of accidental traumaon cytokine and endotoxin production. Crit Care Med1993;21:839-45.
16. Biffl WL, Moore EE, Moore FA, Peterson VM. Interleukin-6in the injured patient. Marker of injury or mediator ofinflammation? Ann Surg 1996;224:647-64.
17. Giannoudis PV, Smith RM, Perry SL, Windsor AJ, DicksonRA, Bellamy MC. Immediate IL-10 expression followingmajor orthopaedic trauma: relationship to anti-inflammatory response and subsequent development ofsepsis. Intensive Care Med 2000;26:1076-81.
18. Rabinovici R, John R, Esser KM, Vernick J, Feuerstein G.Serum tumor necrosis factor-alpha profile in traumapatients. J Trauma 1993;35:698-702.
19. Roumen RM, Hendriks T, van der Ven-Jongekrijg J, Nieu-wenhuijzen GA, Sauerwein RW, van der Meer JW, Goris RJ.Cytokine patterns in patients after major vascular surgery,hemorrhagic shock, and severe blunt trauma. Relationwith subsequent adult respiratory distress syndrome andmultiple organ failure. Ann Surg 1993;218:769-76.
20. Seekamp A, Jochum M, Ziegler M, van Griensven M,Martin M, Regel G. Cytokines and adhesion molecules inelective and accidental trauma-related ischemia/reperfusion. J Trauma 1998;44:874-82.
21. Sherry RM, Cue JI, Goddard JK, Parramore JB, DiPiro JT.Interleukin-10 is associated with the development of sepsisin trauma patients. J Trauma 1996;40:613-6; discussion616-7.
22. DiPiro JT, Howdieshell TR, Goddard JK, Callaway DB,Hamilton RG, Mansberger AR Jr. Association ofinterleukin-4 plasma levels with traumatic injury and clini-cal course. Arch Surg 1995;130:1159-62; discussion 1162-3.
23. DiPiro JT, Hamilton RG, Howdieshell TR, Adkinson NF Jr.,Mansberger AR Jr. Total IgE in plasma is elevated aftertraumatic injury and is associated with sepsis syndrome.Ann Surg 1992;215:460-5; discussion 465-6.
24. Bjornson AB, Altemeier WA, Bjornson HS. Host defenseagainst opportunist microorganisms following trauma. II.Changes in complement and immunoglobulins in patientswith abdominal trauma and in septic patients withouttrauma. Ann Surg 1978;188:102-8.
25. Faist E, Kupper TS, Baker CC, Chaudry IH, Dwyer J, BaueAE. Depression of cellular immunity after major injury. Itsassociation with posttraumatic complications and its rever-sal with immunomodulation. Arch Surg 1986;121:1000-5.
26. O’Mahony JB, Palder SB, Wood JJ, McIrvine A, Rodrick ML,Demling RH, Mannick JA. Depression of cellular immunity
after multiple trauma in the absence of sepsis. J Trauma1984;24:869-75.
27. Keane RM, Birmingham W, Shatney CM, Winchurch RA,Munster AM. Prediction of sepsis in the multitraumaticpatient by assays of lymphocyte responsiveness. SurgGynecol Obstet 1983;156:163-7.
28. Livingston DH, Appel SH, Wellhausen SR, Sonnenfeld G,Polk HC Jr. Depressed interferon gamma production andmonocyte HLA-DR expression after severe injury. ArchSurg 1988;123:1309-12.
29. Szabo G, Kodys K, Miller-Graziano CL. Elevated monocyteinterleukin-6 (IL-6) production in immunosuppressedtrauma patients. I. Role of Fc gamma RI cross-linkingstimulation. J Clin Immunol 1991;11:326-35.
30. Faist E, Schinkel C, Zimmer S, Kremer JP, Von Donners-marck GH, Schildberg FW. Inadequate interleukin-2 syn-thesis and interleukin-2 messenger expression followingthermal and mechanical trauma in humans is caused bydefective transmembrane signalling. J Trauma 1993;34:846-53; discussion 853-4.
31. Lyons A, Kelly JL, Rodrick ML, Mannick JA, Lederer JA.Major injury induces increased production ofinterleukin-10 by cells of the immune system with a nega-tive impact on resistance to infection. Ann Surg 1997;226:450-8; discussion 458-60.
32. MacConmara MP, Maung AA, Fujimi S, McKenna AM,Delisle A, Lapchak PH, Rogers S, Lederer JA, Mannick JA.Increased CD4+ CD25+ T regulatory cell activity in traumapatients depresses protective Th1 immunity. Ann Surg2006;244:514-23.
33. Murphy TJ, Ni Choileain N, Zang Y, Mannick JA,Lederer JA. CD4+CD25+ regulatory T cells control innateimmune reactivity after injury. J Immunol 2005;174:2957-63.
34. Kelly JL, Lyons A, Soberg CC, Mannick JA, Lederer JA. Anti-interleukin-10 antibody restores burn-induced defects inT-cell function. Surgery 1997;122:146-52.
35. Ni Choileain N, MacConmara M, Zang Y, Murphy TJ,Mannick JA, Lederer JA. Enhanced regulatory T cell activityis an element of the host response to injury. J Immunol2006;176:225-36.
36. Hsieh CH, Frink M, Hsieh YC, Kan WH, Hsu JT, SchwachaMG, Choudhry MA, Chaudry IH. The role of MIP-1 alphain the development of systemic inflammatory responseand organ injury following trauma hemorrhage. J Immunol2008;181:2806-12.
37. Kawasaki T, Fujimi S, Lederer JA, Hubbard WJ, ChoudhryMA, Schwacha MG, Bland KI, Chaudry IH. Trauma-hemorrhage induces depressed splenic dendritic cell func-tions in mice. J Immunol 2006;177:4514-20.
38. Kobbe P, Vodovotz Y, Kaczorowski DJ, Mollen KP, BilliarTR, Pape HC. Patterns of cytokine release and evolution ofremote organ dysfunction after bilateral femur fracture.Shock 2008;30:43-7.
39. Mack VE, McCarter MD, Naama HA, Calvano SE, Daly JM.
IMMUNE RESPONSE TO TRAUMA AND TRANSFUSION
Volume 52, December 2012 TRANSFUSION 2547

Dominance of T-helper 2-type cytokines after severeinjury. Arch Surg 1996;131:1303-8; discussion 1308-9.
40. Kang SC, Matsutani T, Choudhry MA, Schwacha MG, RueLW, Bland KI, Chaudry IH. Are the immune responses dif-ferent in middle-aged and young mice following bone frac-ture, tissue trauma and hemorrhage? Cytokine 2004;26:223-30.
41. Wichmann MW, Ayala A, Chaudry IH. Severe depression ofhost immune functions following closed-bone fracture,soft-tissue trauma, and hemorrhagic shock. Crit Care Med1998;26:1372-8.
42. Abraham E, Chang YH. Cellular and humoral bases ofhemorrhage-induced depression of lymphocyte function.Crit Care Med 1986;14:81-6.
43. Abraham E, Freitas AA. Hemorrhage produces abnormali-ties in lymphocyte function and lymphokine generation.J Immunol 1989;142:899-906.
44. Stephan RN, Kupper TS, Geha AS, Baue AE, Chaudry IH.Hemorrhage without tissue trauma produces immunosup-pression and enhances susceptibility to sepsis. Arch Surg1987;122:62-8.
45. Morris JA Jr., MacKenzie EJ, Damiano AM, Bass SM. Mor-tality in trauma patients: the interaction between hostfactors and severity. J Trauma 1990;30:1476-82.
46. Oberholzer A, Keel M, Zellweger R, Steckholzer U, TrentzO, Ertel W. Incidence of septic complications and multipleorgan failure in severely injured patients is sex specific.J Trauma 2000;48:932-7.
47. Schroder J, Kahlke V, Staubach KH, Zabel P, Stuber F.Gender differences in human sepsis. Arch Surg 1998;133:1200-5.
48. Wichmann MW, Inthorn D, Andress HJ, Schildberg FW.Incidence and mortality of severe sepsis in surgical inten-sive care patients: the influence of patient gender ondisease process and outcome. Intensive Care Med 2000;26:167-72.
49. Wohltmann CD, Franklin GA, Boaz PW, Luchette FA,Kearney PA, Richardson JD, Spain DA. A multicenter evalu-ation of whether gender dimorphism affects survival aftertrauma. Am J Surg 2001;181:297-300.
50. Choudhry MA, Bland KI, Chaudry IH. Trauma and immuneresponse—effect of gender differences. Injury 2007;38:1382-91.
51. Yokoyama Y, Schwacha MG, Samy TS, Bland KI, ChaudryIH. Gender dimorphism in immune responses followingtrauma and hemorrhage. Immunol Res 2002;26:63-76.
52. Burrows L, Tartter P. Effect of blood transfusions oncolonic malignancy recurrent rate. Lancet 1982;2:662.
53. Okuno K, Ozaki M, Shigeoka H, Nakajima I, Nakamura K,Hirohata T, Jinnai H, Yasutomi M. Effect of packed red celland whole blood transfusion on liver-associated immunefunction. Am J Surg 1994;168:340-4.
54. Opelz G, Sengar DP, Mickey MR, Terasaki PI. Effect ofblood transfusions on subsequent kidney transplants.Transplant Proc 1973;5:253-9.
55. Pirenne J, Kitade H, Kawai M, Koshiba T, Van Damme B,Mathieu C, Waer M. Regulatory cells, TH1/TH2 unbalance,and antibody-induced chronic rejection in operational tol-erance induced by donor-specific blood transfusion. Trans-plantation 2005;79:S25-7.
56. Salvatierra O Jr., Vincenti F, Amend W, Potter D, Iwaki Y,Opelz G, Terasaki P, Duca R, Cochrum K, Hanes D, StoneyRJ, Feduska NJ. Deliberate donor-specific blood transfu-sions prior to living related renal transplantation. A newapproach. Ann Surg 1980;192:543-52.
57. Singal DP, Joseph S. Role of blood transfusions on theinduction of antibodies against recognition sites on T lym-phocytes in renal transplant patients. Hum Immunol 1982;4:93-108.
58. Utter GH, Reed WF, Lee TH, Busch MP. Transfusion-associated microchimerism. Vox Sang 2007;93:188-95.
59. Gianotti L, Pyles T, Alexander JW, Babcock GF, Carey MA.Impact of blood transfusion and burn injury on microbialtranslocation and bacterial survival. Transfusion 1992;32:312-7.
60. Winslow GA, Shelby J, Nelson EW, Saffle JR. Influence ofallogeneic blood transfusion on natural killer cell activityin burn-injured mice. J Burn Care Rehabil 1996;17:117-23.
61. Sullivan DJ, Barton RG, Edelman LS, Shao Y, Nelson EW,Shelby J. Distinct effects of allogeneic blood transfusion onsplenocyte cytokine production after hemorrhagic shock.J Surg Res 1998;75:54-60.
62. Golde WT, Gollobin P, Rodriguez LL. A rapid, simple, andhumane method for submandibular bleeding of miceusing a lancet. Lab Anim (NY) 2005;34:39-43.
63. Jackman RP, Utter GH, Heitman JW, Hirschkorn DF, LawJP, Gefter N, Busch MP, Norris PJ. Effects of blood sampleage at time of separation on measured cytokine concentra-tions in human plasma. Clin Vaccine Immunol 2011;18:318-26.
64. Gardlund B, Sjolin J, Nilsson A, Roll M, Wickerts CJ,Wretlind B. Plasma levels of cytokines in primary septicshock in humans: correlation with disease severity. J InfectDis 1995;172:296-301.
65. Perl M, Gebhard F, Knoferl MW, Bachem M, Gross HJ, KinzlL, Strecker W. The pattern of preformed cytokines in tissuesfrequently affected by blunt trauma. Shock 2003;19:299-304.
66. Law MM, Cryer HG, Abraham E. Elevated levels of solubleICAM-1 correlate with the development of multiple organfailure in severely injured trauma patients. J Trauma 1994;37:100-9; discussion 109-10.
67. Grossetete M, Phelps J, Arko L, Yonas H, Rosenberg GA.Elevation of matrix metalloproteinases 3 and 9 in cere-brospinal fluid and blood in patients with severe traumaticbrain injury. Neurosurgery 2009;65:702-8.
68. Ulrich D, Noah EM, von Heimburg D, Pallua N. TIMP-1,MMP-2, MMP-9, and PIIINP as serum markers for skinfibrosis in patients following severe burn trauma. PlastReconstr Surg 2003;111:1423-31.
69. Crespo AR, Da Rocha AB, Jotz GP, Schneider RF, Grivicich
JACKMAN ET AL.
2548 TRANSFUSION Volume 52, December 2012

I, Pinheiro K, Zanoni C, Regner A. Increased serum sFasand TNFalpha following isolated severe head injury inmales. Brain Inj 2007;21:441-7.
70. Lenzlinger PM, Marx A, Trentz O, Kossmann T, Morganti-Kossmann MC. Prolonged intrathecal release of soluble Fasfollowing severe traumatic brain injury in humans.J Neuroimmunol 2002;122:167-74.
71. Paunel-Gorgulu A, Flohe S, Scholz M, Windolf J, Logters T.Increased serum soluble Fas after major trauma is associ-ated with delayed neutrophil apoptosis and developmentof sepsis. Crit Care 2011;15:R20.
72. Semple BD, Bye N, Rancan M, Ziebell JM, Morganti-Kossmann MC. Role of CCL2 (MCP-1) in traumatic braininjury (TBI): evidence from severe TBI patients and CCL2-/-mice. J Cereb Blood Flow Metab 2010;30:769-82.
73. Pincemail J, Deby-Dupont G, Deby C, Thirion A, Torpier G,Faymonville ME, Damas P, Tomassini M, Lamy M, Fran-chimont P. Fast double antibody radioimmunoassay ofhuman granulocyte myeloperoxidase and its application toplasma. J Immunol Methods 1991;137:181-91.
74. De AK, Kodys KM, Pellegrini J, Yeh B, Furse RK, Bankey P,Miller-Graziano CL. Induction of global anergy rather thaninhibitory Th2 lymphokines mediates posttrauma T cellimmunodepression. Clin Immunol 2000;96:52-66.
75. O’Sullivan ST, Lederer JA, Horgan AF, Chin DH, MannickJA, Rodrick ML. Major injury leads to predominance ofthe T helper-2 lymphocyte phenotype and diminishedinterleukin-12 production associated with decreased resis-tance to infection. Ann Surg 1995;222:482-90; discussion490-2.
76. Puyana JC, Pellegrini JD, De AK, Kodys K, Silva WE, MillerCL. Both T-helper-1- and T-helper-2-type lymphokines aredepressed in posttrauma anergy. J Trauma 1998;44:1037-45; discussion 1045-6.
77. Stylianos S, Wakabayashi G, Gelfand JA, Harris BH.Experimental hemorrhage and blunt trauma do notincrease circulating tumor necrosis factor. J Trauma 1991;31:1063-7.
78. Wick M, Kollig E, Muhr G, Koller M. The potential patternof circulating lymphocytes TH1/TH2 is not altered aftermultiple injuries. Arch Surg 2000;135:1309-14.
79. Asadullah K, Docke WD, Ebeling M, Friedrich M, Belbe G,Audring H, Volk HD, Sterry W. Interleukin 10 treatment ofpsoriasis: clinical results of a phase 2 trial. Arch Dermatol1999;135:187-92.
80. Nuki G, Bresnihan B, Bear MB, McCabe D. Long-term safetyand maintenance of clinical improvement following treat-ment with anakinra (recombinant human interleukin-1receptor antagonist) in patients with rheumatoid arthritis:extension phase of a randomized, double-blind, placebo-controlled trial. Arthritis Rheum 2002;46:2838-46.
81. Caron A, Desrosiers RR, Beliveau R. Ischemia injury altersendothelial cell properties of kidney cortex: stimulation ofMMP-9. Exp Cell Res 2005;310:105-16.
82. de Vries DK, Lindeman JH, Tsikas D, de Heer E, Roos A, de
Fijter JW, Baranski AG, van Pelt J, Schaapherder AF. Earlyrenal ischemia-reperfusion injury in humans is dominatedby IL-6 release from the allograft. Am J Transplant 2009;9:1574-84.
83. Detmar M, Brown LF, Berse B, Jackman RW, Elicker BM,Dvorak HF, Claffey KP. Hypoxia regulates the expression ofvascular permeability factor/vascular endothelial growthfactor (VPF/VEGF) and its receptors in human skin.J Invest Dermatol 1997;108:263-8.
84. Kukielka GL, Youker KA, Hawkins HK, Perrard JL, MichaelLH, Ballantyne CM, Smith CW, Entman ML. Regulation ofICAM-1 and IL-6 in myocardial ischemia: effect of reperfu-sion. Ann N Y Acad Sci 1994;723:258-70.
85. Kuyvenhoven JP, Molenaar IQ, Verspaget HW, VeldmanMG, Palareti G, Legnani C, Moolenburgh SE, Terpstra OT,Lamers CB, van Hoek B, Porte RJ. Plasma MMP-2 andMMP-9 and their inhibitors TIMP-1 and TIMP-2 duringhuman orthotopic liver transplantation. The effect of apro-tinin and the relation to ischemia/reperfusion injury.Thromb Haemost 2004;91:506-13.
86. Proczka RM, Malecki M, Chorostowska-Wynimko J, Polan-ski JA. Vascular-endothelial growth factor (VEGF) inpatients with peripheral ischemia. J Physiol Pharmacol2006;57 Suppl 4:305-11.
87. Kavathia N, Jain A, Walston J, Beamer BA, Fedarko NS.Serum markers of apoptosis decrease with age and cancerstage. Aging (Albany NY) 2009;1:652-63.
88. Shurin GV, Yurkovetsky ZR, Chatta GS, Tourkova IL, ShurinMR, Lokshin AE. Dynamic alteration of soluble serumbiomarkers in healthy aging. Cytokine 2007;39:123-9.
89. Miller SJ, Watson WC, Kerr KA, Labarrere CA, Chen NX,Deeg MA, Unthank JL. Development of progressive aorticvasculopathy in a rat model of aging. Am J Physiol HeartCirc Physiol 2007;293:H2634-43.
90. Richter V, Rassoul F, Purschwitz K, Hentschel B, Reuter W,Kuntze T. Circulating vascular cell adhesion moleculesVCAM-1, ICAM-1, and E-selectin in dependence on aging.Gerontology 2003;49:293-300.
91. Zou Y, Jung KJ, Kim JW, Yu BP, Chung HY. Alteration ofsoluble adhesion molecules during aging and their modu-lation by calorie restriction. FASEB J 2004;18:320-2.
92. Li L, Smith A, Hagen TM, Frei B. Vascular oxidative stressand inflammation increase with age: ameliorating effectsof alpha-lipoic acid supplementation. Ann N Y Acad Sci2010;1203:151-9.
93. Napolitano LA, Burt TD, Bacchetti P, Barron Y, French AL,Kovacs A, Anastos K, Young M, McCune JM, GreenblattRM. Increased circulating interleukin-7 levels in HIV-1-infected women. J Acquir Immune Defic Syndr 2005;40:581-4.
SUPPORTING INFORMATION
Additional Supporting Information may be found in theonline version of this article:
IMMUNE RESPONSE TO TRAUMA AND TRANSFUSION
Volume 52, December 2012 TRANSFUSION 2549

Fig. S1. Triage criteria for inclusion in study. Patients inthe 911 and 922 triage groups were included in the study.Fig. S2. Examples of individual patient trends. Levels offive cytokines for three representative patients (0013,0114, and 0153) are plotted as examples of the observedtrends within individuals. All three patients are transfusedmales with blunt wounds. 0013 is age 34 with an ISS of 32,
0114 is age 43 with an ISS of 14, and 0153 is age 69 with anISS of 14. (A) IL-6 and IL-10 are shown as examples ofcytokines elevated early after trauma. (B) IL-17 is shown asan example of a cytokine that is depressed after trauma.(C) tPAI-1 is shown as an example of a cytokine that has alater elevation after trauma.Table S1. Median concentration of proteins at day 0.
JACKMAN ET AL.
2550 TRANSFUSION Volume 52, December 2012
Triage Nurse
No Criteria Met
Trauma Activation Algorithm
• Penetrating injuries to the
chest, neck, and/or abdomen
• GCS < 9 or deteriorating by
2 with trauma
• BP < 90 or confirmed
age-specific hypotension for
patients < 10 years of age *
*Systolic blood pressure
< 70 + (2x the age in years)
• Pulseless, injured extremity
• Transfer patient receiving
blood
• Intubated patients
911
• Penetrating injuries to the head or
extremities proximal to elbow and
knee
• Flail chest
• Pelvic fractures
• Two or more long-bone fractures
• Injured extremity with arterial
bleeding
• Combination, trauma with burns
• Amputation proximal to the wrist/
ankle
• Limb paralysis
• Fall > 20 feet
• Head injury and GCS 9-13
• Respirations < 10 or > 29
• Blunt trauma & pregnancy > 20
weeks
• Age > 65 years and meets 933
criteria
922
• Ejection from a motorized vehicle
• High speed or force MVA
• Rollover MVA
• Death in the same passenger
compartment
• Extrication > 20 minutes
• MCA > 20 mph or separation of
rider from motorcycle
• Auto vs pedestrian > 5 mph
• Auto vs bike at > 5 mph
• Head injury and GCS 14
• Chest or abdominal pain after
trauma
• Crushed or rolled over at any
speed
• Head trauma and on Coumadin
933
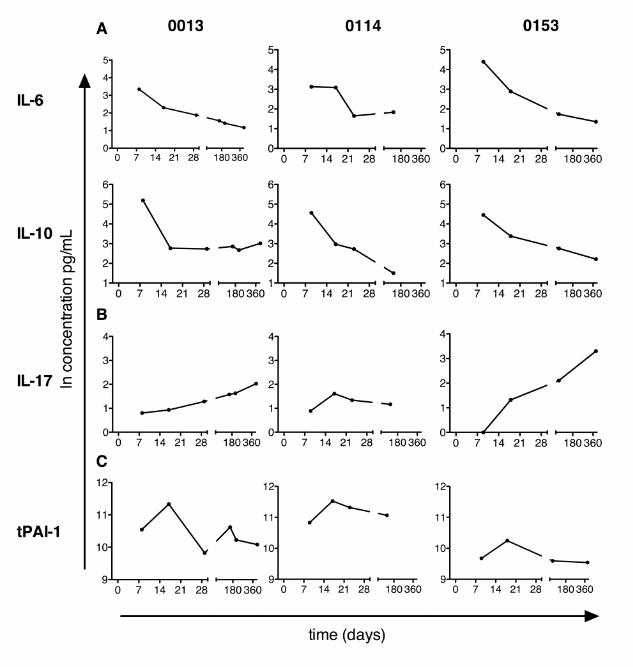
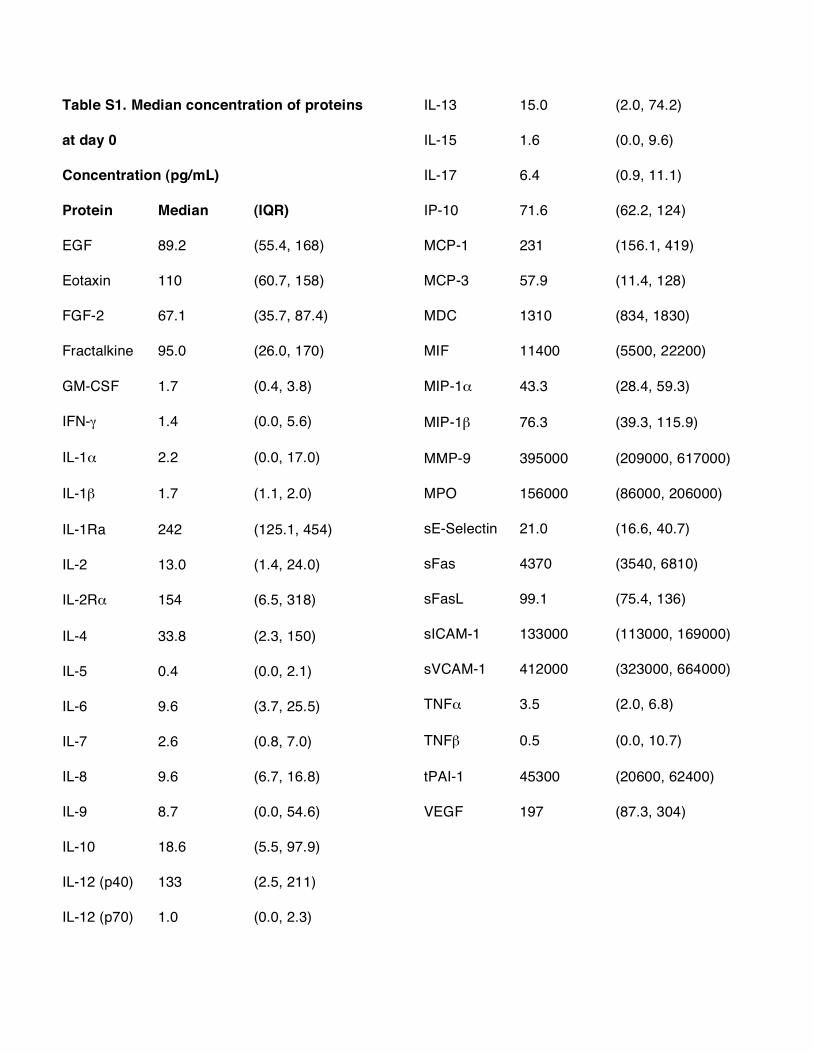
Table S1. Median concentration of proteins
at day 0
Concentration (pg/mL)
Protein Median (IQR)
EGF 89.2 (55.4, 168)
Eotaxin 110 (60.7, 158)
FGF-2 67.1 (35.7, 87.4)
Fractalkine 95.0 (26.0, 170)
GM-CSF 1.7 (0.4, 3.8)
IFN-! 1.4 (0.0, 5.6)
IL-1" 2.2 (0.0, 17.0)
IL-1# 1.7 (1.1, 2.0)
IL-1Ra 242 (125.1, 454)
IL-2 13.0 (1.4, 24.0)
IL-2R" 154 (6.5, 318)
IL-4 33.8 (2.3, 150)
IL-5 0.4 (0.0, 2.1)
IL-6 9.6 (3.7, 25.5)
IL-7 2.6 (0.8, 7.0)
IL-8 9.6 (6.7, 16.8)
IL-9 8.7 (0.0, 54.6)
IL-10 18.6 (5.5, 97.9)
IL-12 (p40) 133 (2.5, 211)
IL-12 (p70) 1.0 (0.0, 2.3)
IL-13 15.0 (2.0, 74.2)
IL-15 1.6 (0.0, 9.6)
IL-17 6.4 (0.9, 11.1)
IP-10 71.6 (62.2, 124)
MCP-1 231 (156.1, 419)
MCP-3 57.9 (11.4, 128)
MDC 1310 (834, 1830)
MIF 11400 (5500, 22200)
MIP-1" 43.3 (28.4, 59.3)
MIP-1# 76.3 (39.3, 115.9)
MMP-9 395000 (209000, 617000)
MPO 156000 (86000, 206000)
sE-Selectin 21.0 (16.6, 40.7)
sFas 4370 (3540, 6810)
sFasL 99.1 (75.4, 136)
sICAM-1 133000 (113000, 169000)
sVCAM-1 412000 (323000, 664000)
TNF" 3.5 (2.0, 6.8)
TNF# 0.5 (0.0, 10.7)
tPAI-1 45300 (20600, 62400)
VEGF 197 (87.3, 304)




















