
Present-day risk assessment would have predicted the extinction of the passenger pigeon

Journal of Experimental Psychology:Animal Behavior Processes
Copyright © 1978 by the American Psychological Association, Inc.
VOL. 4, No. 1 JANUARY 1978
Discrimination and Emission of Different Key-PeckDurations in the' Pigeon
John M. Ziriax and Alan SilberbergAmerican University
In Experiment 1, food reinforcement depended on pigeons' discriminationsamong three bands of key-peck duration: (a) 0-msec peck (no peck); (b)0-20-msec peck (short pecks) ; and (c) peck durations between 60 and 90,110, or 180 msec, depending on the subject (long pecks). Trials began withthe random selection of a peck-duration band and the presentation of a bluecenter key. The occurrence of a blue-key response within the criterion-bandduration accessed the discrimination phase. This phase consisted of threedifferently colored keys, each color associated with a response-duration band.Only selection of the discrimination-phase color appropriate to the priorblue-key response-duration band was reinforced. All birds accurately dis-criminated prior response bands. In Experiment 2, trials began with a colorindicating the appropriate response band to be emitted, either a short or longpeck. Birds successfully emitted and discriminated both bands, a result in-consistent with Schwartz and Williams's finding that only long pecks canbe reinforced. An interpretation of the peck-duration literature is presented,which views duration as a correlate of response strength.
Recent work, based in large measure on a the response-independent delivery of grain,study by Brown and Jenkins (1968), sug- Successive key light-food trials were sepa-gests that key pecking is not invariably sensi- rated by an intertrial interval (ITI) of 1 min.tive to its consequences. In their study, food- After 20 to 200 trials, pigeons began to peckdeprived pigeons were exposed to the illumi- the lighted key, even though key pecking wasnation of a response key for 8 sec prior to unrelated to food delivery.
Brown and Jenkins noted that while these~ ~ „ . ~ T key pecks were gratuitous, in that reinforce-
This research was supported by National Insti- inHpnwiHwil- nf tVipir nrrnrrmretute of Mental Health Grant MH22881 to The ment was independent ot their occurrence,American University. This report is based on a they nevertheless might have been shaped bythesis submitted by J. Ziriax to The American Uni- an operant process. One need only posit thatversity in partial fulfillment of the requirements the onset Of fae ^ey Ug^t produced anfor a Master's degree. We would like to thank . . , , . c.Scott Parker for helpful comments and suggestions orientation movement toward the key. Sinceand Daniel Najjar for technical assistance. orientation movements would be closely fol-
Requests for reprints should be sent to John lowed by reinforcement, perhaps key pecking
?^^SS^S^^^. *» automatically shaped, as the name given20016. this phenomenon, autoshaping, suggests.
Copyright 1978 by the American Psychological Association, Inc. All rights of reproduction in any form reserved.
1

JOHN M. ZIRIAX AND ALAN SILBERBERG
Despite the plausibility of an operantaccount, subsequent work by Williams andWilliams (1969) showed that any apparentoperant account of the autoshaping phe-nomenon was inadequate. In their study,Brown and Jenkins's autoshaping procedurewas made into an omission paradigm byimposing a negative dependency betweenpecks to the lighted key and reinforcement(also Sheffield, 1965). Consequently, keyillumination was followed by grain presenta-tion only if the pigeon did not peck the key.Despite this negative dependency, pigeonsoften pecked the lighted key, a finding estab-lishing that a stimulus-reinforcer relation issufficient to generate and maintain key peck-ing. As such, key pecks could be viewed asPavlovian conditioned responses, elicited tothe conditioned stimulus (key light) by vir-tue of their contingent association with anunconditioned stimulus (grain; see, however,Woodruff & Williams, 1976).
At the very least, the Williams and Wil-liams (1969) data suggest there may betwo sources of control of the key peck, onebased on response—reinforcer relations, theother based on stimulus-reinforcer relations.This suggestion was made explicit in a studyby Schwartz and Williams (1972). Theyfound topographic differences between keypecks supported by omission and some ope-rant procedures. Operant schedules, such asfixed interval (FI) and fixed ratio (FR),tended to support pecks whose key-contactdurations were two to five times longer (40to 100 msec) than those found in omissionprocedures (20 msec or less). Moreover,Schwartz and Williams found evidence for aprocess difference between each responseband. When food reinforcement was contin-gent on the occurrence of long-duration keypecks, the frequency of these pecks in-creased ; the frequency of short-durationpecks did not increase, however, when rein-forcement was dependent on their occur-rence.
Schwartz (1977b) has recently provided asecond demonstration of the apparent selec-tive sensitivity of long-duration pecks to out-come-contingent control. In this study, pi-geons were exposed to variable-interval (VI)schedules of food reinforcement in which a
key peck occasionally produced a shock tothe wings regardless of peck duration, ordepending on whether it was of short orlong duration. When punishment was de-livered regardless of peck duration or duringthe short-duration response band, no sig-nificant changes occurred in the distributionof key-peck durations. When pecks withinthe long-duration response band werepunished, however, there was a large andreliable drop in the frequency of their emis-sion. Based on this finding, Schwartzargued that only long-duration key peckswere sensitive to the punishment contin-gency. He viewed this finding, in conjunc-tion with the findings of Schwartz and Wil-liams (1972), as supporting the conclusionthat there are two classes of key peck: (a)short-duration pecks, which arise from thepigeon's species-typical feeding pattern andwhich are elicited by food; and (b) long-duration pecks, which are under outcome-contingent control.
The Schwartz (1977b) and Schwartz andWilliams (1972) data are provocative in thatthey invite careful consideration of thetopographic characteristics of the key peck.As recent work has borne out, studying key-peck topographies can lead to new interpre-tations of well-established behavioral phe-nomena. Consider, for example, the phe-nomenon of positive behavioral contrast. AsReynolds (1961) has shown, changing theconditions of multiple-schedule reinforce-ment from, say, multiple VI X—VI X tomultiple VI Jf-Extinction (EXT) often re-sults in a response-rate increase in the un-changed VI component and a rate decreasein the changed component. These ratechanges, which constitute the positive-con-trast effect, have traditionally been attrib-uted to factors such as reduction of responseor reinforcement rate in the changed com-ponent (e.g., Reynolds, 1961; Terrace,1968). Yet, when the topographic char-acteristics of the key peck have been con-sidered—an approach encouraged by theSchwartz and Williams (1972) data—con-trast has appeared to be due to a funda-mentally different process: the addition ofshort-duration, autoshaped key pecks tolong-duration, operant key pecking (Keller,

DISCRIMINATION AND EMISSION OF KEY-PECK DURATION
1974; Schwartz, Hamilton, & Silberberg,1975).
A two-process classification of the keypeck has played a fundamental conceptualrole in the interpretation of many scheduleeffects including autoshaping and omissiontraining, contrast and local contrast (e.g.,Schwartz et al., 1975), performance on dif-ferential reinforcement of low rate (DRL)schedules (e.g., Schwartz, 1977a; Schwartz& Williams, 1971), and matching onmultiple schedules (e.g., Rachlin, 1973).Despite the fact that Schwartz and Wil-liams's (1972) study made this classifkatoryscheme explicit, there have not been manyother published studies using peck durationas a dependent variable (Jenkins & Moore,1973; Schwartz, 1977a, 1977b; Schwartzet al., 1975; Wolin, 1968). The presentstudy addresses this situation directly byinvestigating and comparing the propertiesof short- and long-duration key pecks.
Experiment 1
Two independent accounts immediatelycome to mind for why short-duration pecksseem insensitive to outcome-contingent con-trol in the Schwartz (1977b) and Schwartzand Williams (1972) studies. One possibil-ity is that pigeons might have learned aboutthe reinforcer-prohibiting consequences of akey peck in omission procedures, but per-haps key light—food pairings elicit peckingin the same way that a cinder in the eyeelicits tearing. Thus, the most obviousdemonstration of sensitivity in omission pro-cedures—the withholding of pecking—failsbecause these pecks are controlled by preced-ing stimuli. This, in fact, is a common inter-pretation given to omission effects (seeSchwartz & Gamzu, 1977). A second pos-sibility is that pigeons could control theemission of short-duration pecks, but theyare insensitive to their actual occurrence.For example, Pavlovian conditioned re-sponses, of which short-duration key pecksmay be an analogue, have often been thoughtto offer poorer proprioceptive feedback thanoperants (e.g., Miller & Konorski, 1928/1969; Rescorla & Solomon, 1967). In fact,Sheffield (1965) interpreted his dogs' failure
to inhibit salivation in his omission paradigmas a reflection of their insensitivity to theoccurrence of the conditioned response.
Experiment 1 addresses the second ofthese possible accounts, by determining ifpigeons can discriminate whether a priorkey peck was of short or long duration.The paradigm to be used is a variant of acommon operant procedure, that of condi-tional discriminations (e.g., Gumming &Berryman, 1961). As applied in this study,a conditional-discrimination trial beginswith the illumination of the center key of athree-key chamber with a blue sample stim-ulus. A peck of some experimenter-selectedduration to this key darkens it and illumi-nates all three keys with differently coloredcomparison stimuli. A peck to the correctcomparison key is reinforced by grain pre-sentations, whereas a peck to either of theother two keys is punished by a time-out.Which key is correct on each trial dependson the duration of the peck on the blue keythat accessed the comparison keys. If pi-geons can discriminate among pecks of dif-ferent duration, their accuracy in selectingthe appropriate comparison-stimulus colorshould increase substantially from chancelevels as a function of trials (e.g., Gumming& Berryman, 1961).
By adapting the conditional-discrimina-tion procedure to the study of peck dura-tions, it is possible, at least in principle, todetermine whether pigeons can report theduration of a prior key peck. Three notabledifferences, which distinguish this study'sprocedures from many other conditional-discrimination paradigms, should be empha-sized:
1. In many conditional discriminations,pigeons are required to report an exterocep-tive event, such as the prior key color. Inthe present study, they must report a pro-prioceptive event, the prior peck duration.
2. Conditional discriminations often useonly two comparison stimuli. In the presentstudy, however, three comparison stimuli areused, one comparison stimulus associatedwith each possible sample-stimulus be-havior : short-duration key pecks, long-dura-tion key pecks, and nonpecks (i.e., any re-

JOHN M. ZIRIAX AND ALAN SILBERBERG
sponse but key pecking). The purpose of thenonpeck class is to minimize the possibilityof pigeons' comparison-key choices beinggoverned by either "long versus not-long" or"short versus not-short" discriminations.
3. In many conditional-discriminationprocedures, the accuracy of comparison-stimulus choices is controlled by two out-comes—whether a reinforcer or a time-outresults. In the present study, however, com-parison-stimulus selection is often guidedexteroceptively as well. Early in training,the appropriate comparison-stimulus coloris briefly presented on all keys following asample-stimulus peck. As accuracy improves,exteroceptive-stimulus support is graduallyeliminated. This "titration" procedure hasbeen shown to optimize prospects for master-ing conditional discriminations (e.g., seeSidman & Rosenberger, 1967).
Method
Subjects
Four adult white Carneaux pigeons (B6, B7,B8, and B9), maintained at 80% of their free-feeding weights, served as subjects. They had ex-tensive experience with conditional-discriminationprocedures prior to their use in Experiment 1 (seePreliminary training in Procedure section).
Table 1Subjects' Sample Response Class, Short-and Long-Peck Response Durations, andCorrect Comparison Stimulus
Comparison
Birdno.
6
7
8
9
Sampleresponse
type
NonpeckShortLongNonpeckShortLongNonpeckShortLongNonpeckShortLong
Responseduration(in msec)
0-2060-180
0-2060-90
0-2060-110
0-2060-110
stimulus
Training
RedGreenWhiteWhiteRedGreenRedWhiteGreenWhiteGreenRed
Reversal
WhiteRedGreenRedGreenWhiteWhiteGreenRedRedWhiteGreen
Apparatus
Each of two identical chambers, measuring 27.5 X32.5 X 29.0 cm, served as the experimental space.With the exception of the stainless steel responsepanel, all walls were composed of galvanized steel.The distances from the wire mesh floor to the bot-tom of the S.S X 5 cm hopper aperture and thehouse-light were 5.5 and 26.2 cm, respectively.Three 2.54-cm-diameter Gerbrands-type normallyclosed response keys, spaced 6.5 cm apart centerto center, were located 21 cm above the chamberfloor. Each key required a force of .IS N tooperate. The keys were transilluminated with dif-ferent hues by Industrial Electronics Engineersmultistimulus projectors. A PDP-8/e minicomputercontrolled all experimental events and data collec-tion.
Procedure
Main experimental paradigm. In the final ex-perimental procedure, all subjects were exposed toa three-key conditional-discrimination procedurein which there were three response classes to bediscriminated: (a) short-duration key pecks (key-peck durations between 0 and 20 msec) ; (b) long-duration key pecks (key-peck durations longer than60 msec for all birds, and shorter than 180 msecfor B6, 90 msec for B7, and 110 msec for B8 andB9) ; and (c) nonpecks, defined as the nonoccur-rence of a key peck for a period specified by a vari-able-time (VT) schedule. The upper limit on thelong-duration key pecks and the value of the VTschedule for nonpecking were set for each birdso that the three types 'of sample response in thepresence of the blue key were roughly equiprobable,based on data from a pilot experiment (see Pre-liminary training below).
One of three key colors (red, white, and green)served as the comparison stimulus for each type ofsample-stimulus behavior. The sample response, theduration in milliseconds of the short- and long-duration response bands, and the correct compari-son-stimulus color for all subjects throughout theexperiment are presented in Table 1. The value ofthe VT schedule specifying the duration of thenonpeck response class was 4 sec for B6 and B8,and 6.7 sec for B7 and B9.
The first trial of each session began with the com-puter's selection of the sample-response band, eachband being equiprobable, and the illumination ofthe center key with blue light (see Figure 1). Ablue-key response was either within the responseband selected (Ra,mpu in Figure 1) or not (Rotner).Blue-key responses not within the sample-responseband resulted in a 2-sec time-out. During the time-out, the chamber was completely darkened; anytime-out key pecks restarted the time-out. Afterthe time-out, the trial was restarted with thesample-response band as before. When the responsewas a member of the sample-response band (RB»mp iein Figure 1), a ,70-sec time-out resulted. Key pecks

DISCRIMINATION AND EMISSION OF KEY-PECK DURATION
during this time-out reintroduced the 2-sec time-out (R,ny in Figure 1). These two time-outs iso-lated the same response in time from other keypecks. When an appropriate, temporally isolatedsample response occurred, the .70-sec time-out wasfollowed by the illumination of all three keys withthe color of the correct comparison stimulus forthat particular sample-response band (Faded Sig-nal in Figure 1). The duration of this signal was1.1 sec on the first trial of each session. Signalduration on subsequent trials varied according tothe titration schedule described below. Responses onany key during the signal had no scheduled conse-quences. After the signal terminated, each of thethree comparison stimuli was randomly assignedto a response key. A peck on the key displayingthe correct comparison stimulus (Rcorrect in Figure1) resulted in a correct trial. A peck on eitherof the other two keys resulted in an error trial(Rerror in Figure 1).
Correct trials resulted in access to mixed grainfor 3 sec for Sessions 1-19 for B6, 1-40 for B7,1-38 for B8, and 1-36 for B9. Subsequently, the3-sec grain presentation was available on an FR 2schedule with unreinforced correct trials resultingin a .20-sec hopper flash. The rationale for intro-ducing hopper flashes is that brief-stimulus changesfollowing the completion of a component of aschedule often facilitate stimulus control (e.g.,see Kelleher, 1966, pp. 200-206). All error trialsresulted in a 10-sec time-out, and the FR 2 counterwas reset to zero. Thus, when the FR 2 schedulewas in effect, access to grain resulted only follow-ing two consecutive correct trials.
To facilitate acquisition of the conditional dis-crimination, a titration procedure was used (seeSidman & Rosenberger, 1967). Following each cor-rect sample response, the correct comparison stim-ulus to be selected was presented on all three keys(see Figure 1). If the correct sample stimuluswas then chosen, the duration of this titrated signalwas reduced on subsequent trials. The titrated sig-nal was reduced in six steps: 1.1 to .9, .9 to .7, ,7to .5, .5 to .3, .3 to .1, and .1 to 0 sec. If twotrials in a row were terminated by pecks on otherthan the correct comparison-stimulus key (Re rro»in Figure 1) and the duration of the signal wasless than 1.1 sec, the duration of the signal onthe next trial was increased by one step by thetitration schedule.
Daily sessions ended after 60 reinforcements or30 min without a correct trial. After 90 sessions(Training in Table 1), response-band comparison-stimulus pairings were changed so that all re-sponse bands were assorted with different compari-son stimuli. As can be seen in Table 1, both ofthe two possible transformations of pairings wereused across subjects in making this assortment.Subjects were exposed to this reversal conditionuntil their performances were stable, based onexperimenter judgment.
Preliminary training. The experimental proceduredescribed above was actually the last of three re-
© © ©
© © ©
Figure 1. Outline of procedure for Experiment 1.(B, R, W, and G in circles = blue, red, white, and
green key lights; R = response; T.O. = time-out.See text for details.)
lated procedures developed to study peck duration.The first procedure incorporated neither the time-outs isolating sample-stimulus behaviors nor thetitration schedule described above. The second ap-proximation to the final procedure introduced thetitration schedule. Each of these preliminary pro-cedures lasted 25 sessions. After readjusting sample-stimulus band durations so that their relative fre-quencies were nearly equal and altering response-band comparison-stimulus pairings, we introducedall subjects to the procedure described above inMain experimental paradigm.
Results
Each panel of Figure 2 presents the per-cent of no-peck (stars), short-peck (closedcircles), and long-peck (open circles) com-parison-stimulus selections during trialswithout a titration signal as a function of

6 JOHN M. ZIRIAX AND ALAN SILBERBERG
five-session blocks. Comparison-stimulus pie response. Sample-response types arechoices are separated into panels on the basis labeled in each left-hand panel. For example,of the response type (no peck, short- or the top left-hand panel presents the percentlong-duration peck) that served as the sam- of each of the three types of comparison-
60
20
60
20V)UJO
I 60O
O 20
H-
UJ 60
£2060
20
60
20
,B6 LONG
TRAININGB6 NO PECK
REVERSAL
-*••* .,*,..•*••*•••• **. .*••*•.*••*••...•'
B6 SHORT A 0_o'°'°Xo-°-° I/ 0-0X o~0-o'
r^O^'^r
B7 NO PECK .„-*"*••*..,..*..*..„"" *
B7 SHORT
•B7 LONG
10 30 50 70 90 10 30 50SESSIONS
Figure 2. Percent of no-peck -(stars), short-peck (closed circles), and long-peck (open circles)comparison-stimulus choices as a function of five-session blocks during training and reversalconditions for Birds B6 and B7. (Sample-response types are labeled in each left-hand panel.)

DISCRIMINATION AND EMISSION OF KEY-PECK DURATION 7
stimulus selections when the sample response the right side of the figure are from the re-was from the no-peck category. The curve versal condition. Figure 3 is identical tocomposed of stars identifies correct sample- Figure 2 except that its data are from thestimulus selections; the other two curves other two subjects, Birds B8 and B9.constitute errors. Thus, data presented in Generally speaking, the percent of correctthis format permit identification not only of comparison-stimulus selections increasedchanges in the likelihood of a correct com- across sessions for all subjects and condi-parison-stimulus selection but also of the two tions, asymptoting above chance performancetypes of errors that can obtain. Panels from (33%). In all cases, the asymptotic likeli-
coLU
100
60
20
60
20
O20
zLU 60OOC.LUOL 20
60
20
60
20
TRAINING
B8 NO PECK *••*'
REVERSAL
B8 SHORT
-*• ?B8 LONG
B9 ,NO PECK'•* »..*..*.•*. ..*••*.
•*..*•' •*' •*..*••«• •.
B9 SHORTo-o-°
*••*.•*••*•,..„,*••*• "*••;-%..*.•*••*••*••*••* '*••*••*..*.-*•.»..*•' '•*..».•*
B9 LONG
S,' *•*.* *. *.*. .*.. '*••*•.'«' •*•*.*.*•. •«••*•*•*
°-°-° ' %r°-°-o'°^0.0X " N°
I 0' '•*"*••*..*..*......... ._.,..,
10 30 50 70 90 10 30 50 70SESSIONS
Figure 3. Percent of no-peck (stars), short-peck (closed circles), and long-peck (open circles)comparison-stimulus choices as a function of five-session blocks during training and reversalconditions for Birds B8 and B9. (Sample-response types are labeled in each left-hand panel.)

JOHN M. ZIRIAX AND ALAN SILBERBERG
Table 2Percent-Correct Ranges for Last 10 Sessions for Each Sample-Response Class and Number ofLast Session at or Below Chance Level for Any Response Class
Bird no. &condition
Sample-response class
NonpeckShort
key pecksLong
key pecks
No. of lastsession at
chancelevel
B6
TrainingReversal
B7TrainingReversal
B8
TrainingReversal
B9TrainingReversal
69-9067-90
53-67S8-75
76-9859-95
50-7967-88
50-8654-86
56-7140-71
42-7260-84
51-6353-66
67-9662-88
61-8336-59
49-7558-79
47-6743-69
473
6423
6812
4456
hood of a correct comparison-stimulus selec-tion exceeded that of making either of thetwo possible types of errors. When errorsdid occur, subjects were more likely to con-fuse short with long pecks and vice versathan to confuse either of these response typeswith the no-peck category.
Table 2 presents for each subject the low-est and highest percent-correct scores dur-ing the last 10 sessions of the training andreversal conditions for each of the threesample-stimulus behaviors. In the last col-umn of the table is the number of the lastsession during the training or reversalconditions in which accuracy in reportingany of the three sample-stimulus behaviorsdropped to, or below, chance levels (33%).The results from this table, in conjunctionwith those from Figures 2 and 3, show thatpigeons can report with often high levelsof accuracy whether a prior response hasbeen a nonpeck, a short-duration peck, or along-duration peck.
Discussion
The purpose of Experiment 1 was to testwhether there are differences in the pro-prioceptive characteristics of short- and long-
duration pecks. Toward this end, pigeonswere required to report the duration of aprior key peck. As the curves in Figures 2and 3 show, they did admirably well in thistask, reliably reporting their prior behaviorwhether it was a peck of 0-sec duration (i.e.,a nonpeck), a short-duration peck, or along-duration peck. These findings suggestthat each of these behaviors can be uniquelycoded in terms of its duration or duration-linked topography. Hence, it appears thatthe difficulty Schwartz (1977b) andSchwartz and Williams (1972) had in re-inforcing and punishing short-duration peckswas due not to pigeons' insensitivity to theoccurrence of these pecks but, more likely,to pigeons' inability to control their emission.
Behavior as stimulus. The temporal dis-criminations found in Experiment 1 might beconsidered a psychophysical tour de force,because pigeons reliably distinguished be-tween pecks whose durations differed notby seconds—the kinds of temporal discrimi-nations previously demonstrated (e.g.,Rilling, 1967; Stubbs, 1968)—but by tensof milliseconds. Nevertheless, it is unlikelythat pigeons in the present study were dis-criminating time per se in reporting theirsample-stimulus behaviors. Far more likely,

DISCRIMINATION AND EMISSION OF KEY-PECK DURATION
pigeons discriminated among response topog-raphies, each of which had a characteristicduration. This perspective is suggested bySmith's (1974) finding that different pecktopographies can produce different interre-sponse times (IRTs) ; quite possibly, peckdurations can be similarly determined. Oneneed only assume that during each trial, apigeon chooses among several key-directedmovements within its repertoire, each ofwhich has a characteristic duration. In termsof, .say, the report of a prior peck's duration,the pigeon need only recall the topographyk had selected, because duration is a timeconstant of the selected topography.
Experiment 2
Experiment 1 showed pigeons are cap-able of identifying the temporal duration ofa key contact. Experiment 2 addresses acomplementary issue: Can the emission ofthe behaviors they were so adept at dis-criminating in Experiment 1 be controlledby operant contingencies? To answer thisquestion, the procedure of Experiment 1 wasaltered in two ways. First, the learningtask was simplified by eliminating the non-peck response class. Thus, on certain trialsthe sample stimulus was blue, and only twosample-stimulus behaviors—short- or long-duration key pecks—could produce the com-parison stimuli. Second, during other trials,the sample-stimulus behavior required wasspecified exteroceptively. On those occasions,a trial might begin with a red or green keycolor, indicating that a comparison-stimuluspresentation depends on the emission of, say,a short- or long-duration key peck. The issueof interest is whether the frequencies ofshort-duration pecks when the key is red andlong-duration pecks when the key is greenexceed those respective frequencies when thesample-stimulus behavior is left unspecified(i.e., blue key).
A second issue considered in Experiment2 is whether the discriminations found inExperiment 1 were due not to differencesamong pecks of different duration but tosome other correlated cue such as the numberof blue-key pecks needed to produce the com-parison stimuli. For example, pigeons might
have emitted high frequencies of long-dura-tion pecks only to find that they failed toaccess the discrimination phase of the pro-cedure quickly because a short-duration peckwas the required sample response. On suchoccasions, pigeons might have learned thatdelays in accessing the discrimination phaseof the procedure meant that the short-pecksample response was correct. If pigeonswere discriminating, say, the number of blue-key pecks needed to produce the comparisonstimuli, this discrimination should be evi-denced in the blue-key peck-duration dis-tributions when short- versus long-durationpecks serve as the sample-stimulus behavior.For this reason, blue-key peck durations inExperiment 2 were recorded in two distribu-tions, depending on whether a short or longpeck would effect comparison-stimuluspresentation.
Method
Subjects. The subjects were the same as inExperiment 1.
Apparatus. The apparatus was the same as inExperiment 1.
Procedure. The procedure of Experiment 1 asoutlined in Figure 1, underwent two significantchanges. First, the nonpeck response class andthe VT schedule regulating it were eliminated.Thus, there were only two response classes to bediscriminated, short- and long-duration pecks.Since there were only two appropriate sample-stimulus behaviors, only the two side keys wereilluminated during comparison-stimulus presenta-tions, red and green serving as the comparisonstimuli. The second change from Experiment 1was that trials no longer always began with theillumination of the center key with blue light.Rather, blue-, red- and green-key trials wereequiprobable. As in Experiment 1, blue-key trialsbegan with the computer-controlled selection ofthe class-appropriate sample behavior for thattrial, either a short- or long-duration peck. Sincethe appropriate blue-key sample behavior variedfrom trial to trial, subjects could insure presenta-tion of the comparison stimuli only by emittingboth long and short pecks. Red- and green-keytrials, however, did specify a particular responseclass. Thus, a subject might be required to makea short-duration peck in a red-key trial and along-duration peck in a green-key trial in orderto effect comparison-stimulus presentation.
Short- and long-duration response bands werecued by green and red keys for Birds B6 and B9and red and green keys for B7 and B8. The cor-rect comparison stimulus matched the sample-
10 JOHN M. ZIRIAX AND ALAN SILBERBERG
stimulus color for all red- and green-key trials.Bird B6 was exposed to this procedure for 76sessions, B7 for 71 sessions, B8 for 77 sessions,and B9 for 74 sessions. All data analyses werebased on each subject's last IS sessions. All otherfeatures of the procedure, including the use of thetitration schedule and the FR 2 schedule, werethe same as in Experiment 1.
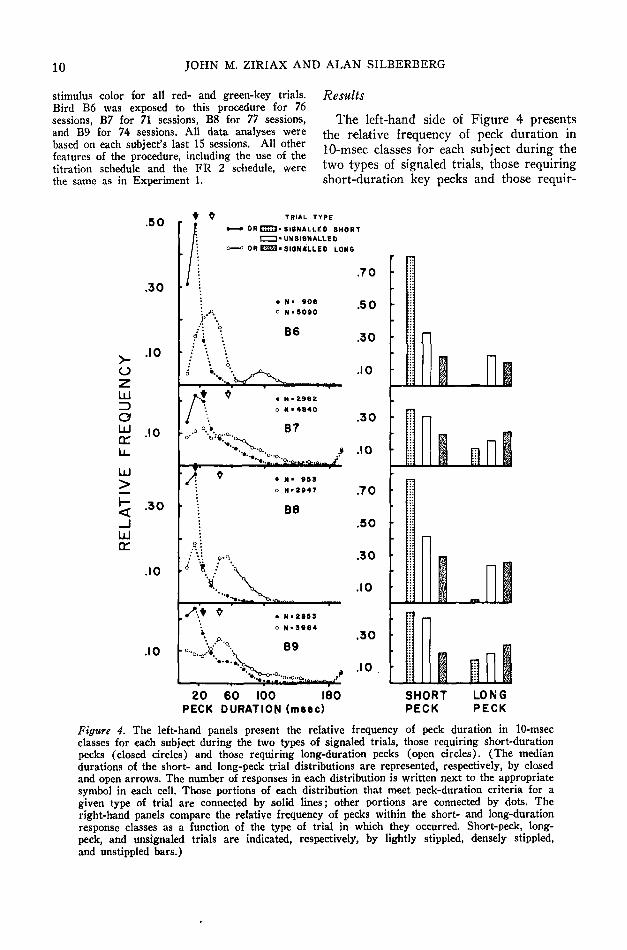
Results
The left-hand side of Figure 4 presentsthe relative frequency of peck duration in10-msec classes for each subject during thetwo types of signaled trials, those requiringshort-duration key pecks and those requir-
.50
.30
O2;UJ
0̂w .10oru_UJ>
S •"_lUJor
.10
.10
T V TRIAL TYPE
? •— • OR CHI- SIGNALLED SHORT/ ITZI'UNSIGNALLEDI o—c ORBS- SIGNALLED LONG
/. j. N- 906 gg
f\ o N-5090t :
l \ \ B6 '30
/ V^-^^-s^ -'0
/
S * N-29B2
\ o N-4840.30
. .0 °,». ..0. B7b'V-̂ ;:,.,..& / -io
f ^/i • N- 98S
: o N-2947 .70
' 1 B8! .50•
2:/'•I ».q .30
•' *./ \a \ ._• "v • ' 0
,*•* ^ • N -2883
\ p. ON ' "84 -30-•o.,..V' '"••• B9
°"° """̂ ->..o..<,,._ / ^ •
.
.
,
1
i Hi_
S pifi!
n
__
m _
I _|| j
i
i ifll20 60 100 180 SHORT LONG
PECK DURATION (msec) PECK PECK
Figure 4. The left-hand panels present the relative frequency of peck duration in 10-msecclasses for each subject during the two types of signaled trials, those requiring short-durationpecks (closed circles) and those requiring long-duration pecks (open circles). (The mediandurations of the short- and long-peck trial distributions are represented, respectively, by closedand open arrows. The number of responses in each distribution is written next to the appropriatesymbol in each cell. Those portions of each distribution that meet peck-duration criteria for agiven type of trial are connected by solid lines; other portions are connected by dots. Theright-hand panels compare the relative frequency of pecks within the short- and long-durationresponse classes as a function of the type of trial in which they occurred. Short-peck, long-peck, and unsignaled trials are indicated, respectively, by lightly stippled, densely stippled,and unstippled bars.)

DISCRIMINATION AND EMISSION OF KEY-PECK DURATION 11
ing long-duration pecks. Three features ofthese data are noteworthy: (a) The modalpeck duration during short-peck trials fellwithin the short-duration response class forall subjects. In fact, for two of the sub-jects (B6 and B8) the majority of pecksduring short-peck trials were in the short-duration class, (b) Long-peck trial distribu-tions tended to be bimodal, with one mode inthe short-peck range and the second mode 10or 20 msec before the beginning of the long-duration response class, (c) As the differ-ence between medians of each trial-typedistribution suggests, more long-durationpecks occurred during long-peck trials thanduring short-peck trials, and more short-duration pecks occurred during short-peck
trials than during long-peck trials for allsubjects.
The right-hand side of Figure 4 comparesthe relative frequency of pecks within theshort- and long-duration response classes asa function of the type of trial in which theyoccurred. These measures were calculatedby dividing the response frequency withina response class during a particular type oftrial by the response total during that trialtype. In all cases, the relative frequency ofshort pecks was highest during short-pecktrials, lowest during long-peck trials, andintermediate during unsignaled trials. Simi-larly, the relative frequency of long-dura-tion pecks was always higher in long-pecktrials than in short-peck trials, and, with
.20• *• \j
>-O
LUID .20
LUCLLL- 0
LU.20•••»
• ^
^I ^
LUa: .20
f\
B6 UNSIGNALLED
. /A UNSIGNALLED
..°-,\ ..o-o REINFORCED
• °' 8'V'% ';.o.
• • "O'̂ Jrf̂ B * * ^̂ ^1f>^̂ *̂
0• './*****.
<f * *
O» *^^*^*"/t_?B"O ^^ "̂O^^4i •, , ^ '̂'̂ '-O-'.*.''*,
•/\B8*.
> C. ^>--.Q..
^^^^»ft • '3 '̂**
.° -̂. B9'.b
• *
9 J~l'''•**»C«$9' *n"fi igi ij"fc_»ifc. .
SHORT •
LONG o
BAND
• n« 916
o n= !889
• n=2657
o n=5798 .«t n.'.*i.-.».-.»l.-.ffi.-.n:
• n = M 5 2
o n«!796
•
• n-4952o n»5300 ,
••
20 60 100 180PECK DURATION (msec)
Figure 5. Relative frequency of sample-stimulus peck durations for each bird in 10-msec classesduring unsignaled short trials (closed circles) and unsignaled long trials (open circles).(Reinforced response-duration bands are signified by solid lines; other portions are connectedby dots. The number of pecks in each distribution is presented within each cell.)

12 JOHN M. ZIRIAX AND ALAN SILBERBERG
Table 3Conditional-Discrimination Performances
Percent correct
Un- Un-Signaled Signaled signaled signaledshort- long- short- long-
Bird peck peck peck peckno. trials trials trials trials
B6B7B8B9
96969785
95949886
94768256
82667550
one exception (B6), long-duration pecksduring unsignaled trials fell in between.
Figure 5 presents the relative frequency ofpeck durations in 10-msec classes for eachsubject during the two types of unsignaledtrials, those requiring short-duration keypecks and those requiring long-durationpecks. The shapes of the two distributionsare largely identical to each other for allof the birds.
Table 3 presents for each subject theaccuracy in selecting the appropriate com-parison stimulus during each type of sample-stimulus presentation. During signaled trials,where conditional discriminations could bebased on both exteroceptive and proprio-ceptive cues, all birds discriminated at levelswell above chance (i.e., 50%). Even duringunsignaled trials, where only the sample-stimulus reponse duration could serve as adiscriminative stimulus, all subjects saveB9 had accuracy levels above chance expec-tations.
Discussion
Experiment 1 showed that pigeons candiscriminate whether the duration of a priorresponse falls within Schwartz and Wil-liams's "reflexive" (0 to 20 msec) or operant(40 or more msec) response bands. Thedata from Table 3 of Experiment 2 cor-roborate these findings in a somewhat dif-ferent procedure: In unsignaled trials, dur-ing which conditional discriminations had tobe based on using peck duration as a dis-criminative event, three of four birds re-
ported the prior peck's duration at levelsof accuracy well above chance. Moreover,Experiment 2 enlarged on the prior study byrecording blue-key peck durations on thebasis of whether the unsignaled trial re-quired a short or long peck to effect com-parison-stimulus presentation (Figure 5).The similarity in this figure between short-peck and long-peck unsignaled duration dis-tributions suggests that no unintended cor-relate of sample-stimulus behavior (e.g.,the number of sample pecks to produce thecomparison stimuli or time since reinforce-ment) served as an alternate cue as to therequired duration of the unsignaled samplepeck.
While this generalized replication of Ex-periment 1 is of some interest, the importantfinding of Experiment 2 was that pigeonscan emit responses whose durations fallwithin the putative reflexive and operantresponse classes. This finding was clearlydemonstrated in Figure 4. The relativefrequencies of short- and long-duration peckswere highest during trials cuing their respec-tive response classes, lowest during trialscuing the alternate response class, and fellin between during those trials where the re-sponse class to be emitted was unspecified.
Despite these results, a two-process classi-fication of the key-peck might be defendedby arguing that increases in the relativefrequency of short-duration pecks duringsignaled short trials were due not to the dif-ferential reinforcement of short pecks butto the differential extinction of pecks oflong duration. In other words, the notionthat only long-duration pecks are sensitiveto operant contingencies is preserved byarguing for an essentially constant popula-tion of short pecks, the relative frequency ofwhich is varied solely by whether long-dura-tion pecks are differentially reinforced.
There are two difficulties with this argu-ment:
1. It is bidirectional. Schwartz and Wil-liams's (1972) finding that only long-dura-tion pecks can be reinforced could be rein-terpreted as a demonstration of the selectivesensitivity of short-duration pecks to dif-ferential extinction.
2. It cannot account for the finding in

DISCRIMINATION AND EMISSION OF KEY-PECK DURATION 13
Figure 4 that long-duration pecks evi-denced less plasticity than short-durationpecks. In particular, the modal duration dur-ing signaled short-peck trials always fellwithin the short-duration response band, yetthe modal duration during signaled long-peck trials never fell within the long-dura-tion response band. Consequently, this inter-pretation must argue that while only longpecks are sensitive to operant contingencies,they are nevertheless less likely than shortpecks to meet the contingencies of reinforce-ment.
General Discussion
The initial question the present study ad-dressed was why short-duration pecks wereselectively insensitive to their consequencesin the Schwartz (1977b) and Schwartz andWilliams (1972) studies. One possibilityadvanced was that the proprioceptive proper-ties of these pecks were somehow less salientthan those of long duration. Even thoughrespondents, to which short-duration pecksmight be related, have often been viewed asfeedback-poor (e.g., Miller & Konorski,1928/1969; Sheffield, 1965), Experiments 1and 2 conclusively showed no class dif-ferences as a function of peck duration inproprioception-based discriminability. Thus,it seems unreasonable to argue that pigeonscan discriminate the occurrence of a peck,except if it is of short duration. Experi-ment 2 assessed the adequacy of a secondexplanation of the selective insensitivity ofshort-duration pecks to outcome-contingentcontrol—that pigeons actually cannot con-trol their emission. By this account, pigeonscan discriminate the occurrence of this "re-flexive" peck, but its evocation is not gov-erned by response-reinforcer relations. Ex-periment 2 showed this explanation also toto be invalid. Pigeons could emit short-dura-tion pecks as readily and possibly morereadily than long-duration pecks. Ratherthan explaining the selective insensitivity ofshort-duration pecks to operant control, theresults of the present study support an un-anticipated conclusion: No meaningful pro-cess distinction can be made between short-and long-duration key pecks.
At first blush, this conclusion may seeminconsistent with most of the data alreadycollected on peck durations. This, in fact,is not the case. Even prior to the presentstudy's results, much of the data on peckdurations defied easy classification in termsof Schwartz and Williams's (1972) two-process definition of the key-peck response.A frequent finding in these data is that thepeck duration obtained seemed inappropriateto the processes generally thought to char-acterize a particular procedure. For example,autoshaping procedures, which are generallythought to tap a respondentlike process (seeBrown & Jenkins, 1968; Gamzu & Williams,1971, 1973), maintain key pecks from thelong-duration, operant response class. Alter-natively, the first few sessions on continuous-reinforcement (CRF) schedules, which pre-sumably control the key peck via an operantprocess, are primarily populated by re-sponses from the short-duration, reflexiveresponse class (see Schwartz & Williams,1972, Figures 6 & 7). Complicating peckclassification still further are Gamzu's(1972; see also Schwartz & Gamzu, 1977,pp. 68-69) and Schwartz's (1977a) findingsthat length of training can substantially in-fluence median peck duration. Early in train-ing, both autoshaping and CRF proceduressupported short-duration pecks; however,late in training, long-duration pecks emergedon both these procedures.
Other peck-duration data not self-evi-dently appropriate to the procedures onwhich they obtained can be seen inSchwartz's (1977a) analysis of performanceson discrete-trials DRL, FI, and FR sched-ules. On discrete-trials DRL schedules,Schwartz found that pecks during the first 2sec after trial onset are primarily from theshort-duration response class, whereas pecksof longer latency are from the long-durationresponse class. A quartile analysis of FIand FR schedules also showed peck-durationchanges within each schedule. Schwartzfound peck durations increased from shortto long as time in the interval passed butdecreased as reinforcement was approachedon FR schedules.
More data not easily related to a two-process account of the key peck come from

14 JOHN M. ZIRIAX AND ALAN SILBERBERG
Schwartz et al. (1975). In one experiment,they recorded peck duration in a two-keymultiple schedule. One key (the autoshapingkey) signaled which multiple-schedule com-ponent was in effect, and a second key(the operant key) provided response-de-pendent reinforcement. During the VI com-ponent of multiple VI-EXT schedules, peck-ing occurred to both keys. Presumably, pecksto the autoshaping key during this differen-tial condition were controlled by the rela-tion between the key light and reinforce-ment, while pecks to the operant key werecontrolled by response-reinforcer relations(also see Keller, 1974). Despite this ap-parent between-key process difference, peckdurations to both keys were from the long-duration, operant response class.
Much of the data where peck durationhas served as a dependent variable is sum-marized above. Generally speaking, thesedurations do not correspond in an obviousway with classifications of procedures suchas "operant" or "respondent." In any case,some procedures, for instance those basedon FI or FR schedules, could not be cata-logued as wholly operant or respondent any-way, because the prominence of short- andlong-duration pecks changes within eachschedule.
Complicating matters further, differentkinds of explanations have been offered toaccommodate each of these findings to a two-process account. For example, the gradualemergence of long-duration pecks on CRFprocedures has been explained in terms ofwhat Schwartz and Gamzu (1977) call the"minimal-unit" hypothesis. In brief, thishypothesis views short-duration key pecks,which naturally occur on Pavlovian pro-cedures, as the biological substrata fromwhich long-duration, operant pecks emerge.As regards CRF schedules, pecking earlyin training is thought to be directly en-hanced by food. Hence, short pecks (i.e.,the minimal units) predominate. With fur-ther training, however, the schedule's oper-ant contingencies gain control, causing long-duration pecks eventually to emerge.
The within-schedule peck-duration changeson FI, FR, and discrete-trial DRL sched-ules are explained by Schwartz (1977a)
in another way—in terms of the presumedtemporal properties of operant and respond-ent conditioning. On FI schedules, pecksearly in the interval presumably have nooperant basis. Hence, what few pecks thereare are of short duration. However, as timein the interval passes and the prospects ofresponse-dependent reinforcement increase,operant factors gain prominence, leading tomore pecks of long duration. On FR sched-ules, all pecks participate in producing re-inforcement. Therefore, long-duration keypecks are prominent throughout the sched-ule. However, those pecks in closest temporalproximity to reinforcement are also in-fluenced by the imminence of food delivery.Consequently, consistent with the principlesof Pavlovian temporal conditioning, the fre-quency of short-duration responding in-creases as the ratio requirement is com-pleted. Finally, on discrete-trials DRLschedules, pecks sensitive to the operantcontingencies are of long latency. Therefore,Schwartz argued that those pecks that con-tinued to occur with short latencies werefrom the reflexive, short-duration responseclass.
Certain unexpected response-duration datafrom Schwartz et al. (1975) and Gamzu(1972) were explained in yet a third way—in terms of adventitious reinforcement.Schwartz et al. theorized that a superstitiousresponse chain had formed between keysbecause changeovers from the autoshapingkey to the operant key occasionally resultedimmediately in reinforcement. To test thisthesis, they imposed a temporal delay be-tween pecking the autoshaping key andoperant-key reinforcement. With this delayin force, they found substantially shorterautoshaping key-peck durations, a conse-quence presumably of eliminating adventi-tiously maintained long-duration operantsfrom that key's duration distributions. Thelengthening of durations on autoshapingtrials in Gamzu's study was also attributedto adventitious reinforcement, based on thefact that autoshaped key pecking is oftenimmediately followed by reinforcement.Hence, long-duration operants, maintainedby a specious response-reinforcer relation-ship, eventually abounded.

DISCRIMINATION AND EMISSION OF KEY-PECK DURATION 15
The plausibility of Schwartz and Wil-liams's (1972) two-process account of thekey peck is weakened by the multiplicity ofspecial explanations advanced to maintainit. Moreover, appeal to notions such asminimal units and adventitious reinforce-ment obfuscates a salient aspect of between-schedule peck-duration data: Each schedulefor which durations have been recordedtends to have a characteristic median dura-tion. The differences in median peck dura-tion that different schedules engender presenta most serious challenge to a two-processaccount.
Consider, for example some of the peck-duration data from Schwartz (1977a). Inthis study he shows (a) that median dura-tions on CRF schedules increase as a func-tion of exposure to this schedule from ap-proximately 20 to 35 msec; (b) that peckdurations on FI schedules increase through-out each quartile of the interval from about35 to 45 msec; and (c) that interquartilechanges on FR schedules are in the oppositedirection, shortening from approximately55 to 45 msec. Not only are median dura-tions on each of these schedules substantiallydifferent but also these between-schedulepeck-duration differences do not follow in anobvious way from any operant-reflexiveclassification of minimal-unit interpreta-tion of key pecks. For example, why areCRF-schedule pecks, even after long expo-sure to operant contingencies, no longer induration than the key pecks during the firstquartile of FI, where most pecks are pre-sumably elicited? Why are last-quartile FRkey pecks, where the contribution of reflex-ive factors is presumably greatest, of essen-tially the same duration as in the most"operant" period of the FI (i.e., the lastquartile on FI) ? Finally, why are medianFR-schedule durations, regardless of theirquartile of occurrence, substantially longerthan median peck durations on CRF, alsoan operant schedule of reinforcement?
Based on Schwartz and Williams's (1972)finding of bimodal peck-duration distribu-tions, it might be argued that differingblends of short- and long-duration pecksproduce the differences in median durationfound on particular schedules. However, this
"blending" account is predicated on theexistence of separable response classes; andit is this crucial test that this account fails,for all peck-duration data from Schwartz(1977a) are unimodal. Rather thanestablishing that between-schedule peck-duration differences are due to differingmixes of two types of key peck, Schwartz'sdata suggest that each schedule type hasits own characteristic median peck duration.
The problem that Schwartz's data pre-sent for a two-class characterization of thekey peck is obvious. If peck-duration dif-ferences correspond to differences in process,and if each schedule's duration distributionis unique to that type of schedule, thenthere are more kinds of key pecks than thetwo catalogued by Schwartz and Williams(1972). This point is clearly made in Fig-ure 6. Each panel's x axis may be ignoredfor the moment. The y-axis coordinates ofthe dots in each panel present the medianof the distribution of peck-duration mediansfor all subjects on a given schedule. Thesources and data from which medians andstandard errors were determined are pre-sented in Table 4. In terms of median dura-tions and their standard errors, one couldargue not just for two kinds of key pecksbut for four: one for omission procedures,one for DRL, CRF, and autoshaping pro-cedures, another for FI schedules, andfinally, one for VI and FR schedules.
We believe that even without consideringthe present study's results, this analysis ofbetween-schedule peck-duration differencesrobs the two-process characterization of thekey-peck and the minimal-unit hypothesis oftheir conceptual utility. Still unanswered,however, is why peck duration varies withdifferent types of schedules. No conclusiveanswer can be given to this question at thistime. However, a plausibility argument canbe advanced that is based on the left-to-rightordering of schedules in Figure 6. If thisrank order corresponds to differences in thestrength of behavior each of these sched-ules maintains, then between-schedule peck-duration differences are likely to be aconsequence, at least in part, of schedule dif-ferences in response strength. Of course, thisargument is predicated on a demonstration

16 JOHN M. ZIRIAX AND ALAN SILBERBERG
Table 4Data Sources for Figure 5: Number of Observations, Mean Latencies, Mean Rates, andMedian Durations
Data provided
No. of Mean Mean Medianobser- latency rate duration
Schedule
OM
DRLCRF
POS
FI
VI
FR
Reference
Schwartz & Williams,1972
Schwartz, 1977bBrownstein & Balsam,
1975Schwartz, 1977bSchwartz, 1977bBrandauer, 1959Schwartz & Williams,
1972Schwartz, 1977bGonzalez, 1973Schwartz & Williams,
1972Schwartz, 1977b
Schwartz, Hamilton, &Silberberg, 1975
Schwartz, 1977aSchwartz & Williams,
1972Schwartz, 1977b
Data source
Figure 1
Figures 1, 2, 3, 4Tables 1, 3
Figures 8, 9Figures 1, 2, 3, 4Table 3Figures 1, 2, 3, 4
Figures 1, 2, 3, 4Figure 2Figures 5, 6, 7
Figures 5, 6, 7Table 1Figure 2Table 1Figures 1, 3Figures 5, 6, 7
Figure 5Table 1
vations
4
246
42438
2434
16
8
204
16
(in sec) (resp/sec) (in msec)
18
186.1
5.2 3033
2.540
331.6
34
40 42
46 48
45 5344
65 48
Note. OM = omission training; DRL = differential reinforcement of low rate; CRF = continuous rein-forcement; POS = autoshaping procedure; FI = fixed interval reinforcement schedule; VI = variableinterval reinforcement schedule; FR = fixed ratio reinforcement schedule.
that the reponse strength associated withomission training is less than that asso-ciated with DRL, and that DRL isless than CRF, and so forth. In orderto make this assessment, key-peck durationson each of these schedules were related totwo conventional measures of responsestrength, the reciprocal of response latencyand the rate of response. The x axes ofeach panel of Figure 6 are presented interms of the logarithm of each of these mea-sures. CRF schedules, for which responserates cannot be readily calculated, and trialprocedures are presented in terms of thelatency measure, and other, free-operantschedules are presented in terms of the re-sponse-rate measure. The ^r-axis coordinatesof each dot present the mean performance interms of latencies or rate from the datasources listed in Table 4.
Any conclusions drawn from considerationof Figure 6 must be tempered by the factthat in several cases a data point's x- and3»-axis values are not only not from thesame subject but not even from the samestudies (see Table 4). For example, CRFresponse latencies are from Brandauer's(1959) doctoral dissertation, while CRFpeck durations are from Schwartz (1977a).This amalgam of results was necessary be-cause much of the data on peck durationsis not also reported in terms of latenciesor rates. Therefore, the only way durationscould be related to these other measures wasby between-study comparisons. In addition,the response latencies from Schwartz's(1977a) DRL data have been recalculatedto make them comparable to the omission-latency data presented by Brownstein andBalsam (1975). Comparability was achieved

DISCRIMINATION AND EMISSION OF KEY-PECK DURATION 17
by defining all DRL latencies between 8 and12 sec in the Schwartz study as being of8-sec duration, the longest possible latencyin Brownstein and Balsam's omission trials.Also adjusted were Vl-schedule responserates from Schwartz et al. (1975). Unlikethe response-rate data from FI and FRschedules presented in Figure 6, their ratesincluded hopper times in the rate calculation.As adjusted in the figure, VI rates excludethe hopper cycle. Finally, Schwartz (1977a)reported DRL, FI, and FR peck durationsin terms of their latencies or quartiles ofoccurrence. As presented in Figure 6, thesedata are based on the average durations ofall pecks regardless of latencies or quartilesof occurrence.
Despite the limitations inherent in com-bining data from different sources and recal-culating data from figures, Figure 6 showsa positive relation between peck durationand two correlates of strength: the recipro-cal of latency and the rate of response. Atleast to a small extent, schedule differencesin median peck duration may be a conse-quence of the strength of responding thatthese schedules engender and not a conse-
quence of the degree of their contingent orreflexive control.
The relation attributed between responsestrength and peck duration implies peckdurations are multiply controlled. On theone hand, duration is sensitive to outcomes,as was seen in Experiment 2; on the otherhand, it is a consequence of the strength aschedule engenders. This duality of controlhas been encountered before in the experi-mental analysis of behavior. For example,response rate covaries with the density ofreinforcement a VI schedule provides, eventhough rate changes produce no largechanges in the time rate of reinforcement(e.g., see Catania & Reynolds, 1968). Yeton rate-shaping schedules such as DRL,which selectively reinforce long IRTs, highreinforcement frequencies usually supportlow response rates (e.g., Staddon, 1965).The problem with response rate, as with peckduration, is that it is not simply a dependentvariable but, as schedules such as DRLshow, a dimension of behavior sensitive tothe very contingencies it is intended tomeasure.
One way of reconciling this apparent in-
50
S 40CC3Q
Uju 30
oui
20
DRLCRF
OM
POS
FI
VI
CDFR
-.8 -.6 -.4 -.2 1.6 1.7 1.8LOG (MEAN LATENCY IN SEC"') LOG ( M E A N RE S PONSE S/ MINI
Figure 6. Median peck duration as a function of two other measures, the log of the reciprocalof the mean latency in sec (left panel) and the log of the mean resiponse rate in min (rightpanel). (The rectangles surrounding each data point represent one standard error, plus andminus, of the distribution of median durations or mean latencies and rates from which thedata were drawn. The schedule each data point represents is presented over the data point:DRL = differential reinforcement of low rate; CRF = continuous reinforcement; FI = fixedinterval reinforcement schedule; FR = fixed ratio reinforcement schedule; VI = variable intervalreinforcement schedule; OM and POS, respectively, signify omission training and autoshapingprocedures. Also see Table 4.)

18 JOHN M. ZIRIAX AND ALAN SILBERBERG
compatibility is based on the notion of sched-ule homogeneity (see Herrnstein & Love-land, 1975). On relatively homogeneousschedules such as VI, where the probabilityof reinforcement is largely unrelated to re-sponse rate, rates are free to vary with thestrength supported by that schedule's den-sity of reinforcement. On DRL schedules,however, the conditions of reinforcement arenonhomogeneous—that is, the rate of rein-forcement changes dramatically with changesin response rate. Under these conditions,where microcontingencies based on the IRTcontrol response emission, response rate isnot free to vary. Hence, low rates obtaindespite high time rates of reinforcement.
A similar explanation can be used toreconcile the results of Experiment 2 andthe data presented in Figure 6. In Experi-ment 2, the conditions of reinforcement werenonhomogeneous—that is, the probability ofreinforcement varied dramatically as a func-tion of a peck's duration. These constrainingmicrocontingencies optimized prospects fordemonstrating the sensitivity of the peck'sduration to differential reinforcement. In allof the studies presented in Figure 6, how-ever, peck duration was unconstrained—that is, in no case did the probability ofreinforcement vary with the peck's duration.Under these homogeneous circumstances,peck duration was free to vary with thestrength that the underlying schedule of rein-forcement naturally supported. Conse-quently, the use of peck's duration as a cor-relate of strength may only be appropriateunder conditions where reinforcement is notdirectly contingent on peck duration.
Despite this caveat, positing a relationbetween response strength and peck dura-tion holds the promise of integrating largeportions of the peck-duration literaturewithin a single conceptual framework. Itaccounts not only for the between-scheduledifferences in peck duration shown in Fig-ure 6 but also explains some peck-durationchanges found within a single type of sched-ule. For example, Gamzu (reported inSchwartz & Gamzu, 1977) noted that therate of autoshaped key pecking was in-fluenced by the subject's history of exposureto response-indpendent schedules of rein-
forcement. In particular, he found that expo-sure to nondifferential conditions, in whichthe frequency of reinforcement in the pres-ence of the trial and the ITI are the same,led to low response rates and short peckdurations in subsequent autoshaping pro-cedures. When not preceded by nondifferen-tial conditions, autoshaping rates were higherand median durations were longer. WhileSchwartz and Gamzu interpreted these re-sults in terms of the minimal-unit hypothesisand a two-process classification of the keypeck, the results in Figure 6 suggest analternative explanation: Because the strengthof behavior sustained by low-rate subjectswas less than that sustained by high-ratesubjects, low-rate subjects' key pecks wereof shorter duration.
The results of Schwartz et al. (1975)can also be reinterpreted in terms of thisresponse-strength account. In their two-keymultiple schedule, the introduction of atemporal delay led to susbtantial shorteningof median peck durations on the signal key,a result they attributed to breaking anadventitiously maintained response chain;however, the delay also reduced signal-keypecking rates substantially. A response-strength account suggests that the shortersignal-key peck durations found with thedelay had less to do with adventitious rein-forcement than with the attendant reductionin response rate. Finally, Schwartz and Wil-liams (1972) found that reducing the rein-forcement probability at the end of auto-shaping trials in steps from 1.0 to .5 led toprogressively higher response rates andlonger peck durations. While they offeredno account for this finding, it is obviouslyconsistent with the response-strength inter-pretation of peck durations just advanced.
In these examples, a response-strengthaccount does a good job of explaining peck-duration changes that occur within a singletype of schedule or procedure. This accom-plishment complements the demonstration inFigure 6 that a notion of strength is usefulin relating between-schedule peck-durationdifferences. Nevertheless, two other sets ofdata (Schwartz, 1977a, 1977b) must beaccounted for in terms of strength before

DISCRIMINATION AND EMISSION OF KEY-PECK DURATION 19
such an account can be seriously considered.These data are discussed below.
On a baseline VI schedule of food rein-forcement, Schwartz (1977b) imposed re-sponse-dependent shock for pecks of eithershort or long duration. When shock was de-pendent on the occurrence of long-durationpecks, the relative frequency of those pecksdecreased substantially from baseline levels.When short-duration pecks were punished,however, there was virtually no change intheir relative frequency when compared tobaseline performances. Schwartz interpretedthis selective sensitivity to punishment interms of a reflexive-operant process differ-ence between pecks of short and long dura-tion. While his data seem consistent with thistwo-process account, another interpretationis possible, because in only three of eightsubjects was the frequency of punished long-duration pecks lower than the frequency ofshort-duration pecks when they werepunished. Consequently, the apparent insen-sitivity of short-duration pecks to a punish-ment contingency might have been due to afloor effect. Long-duration pecks, which pre-dominate on VI schedules (see Figure 6),could be reduced to this basal level bypunishment; however, short-duration pecks,which are infrequent on VI schedules, couldnot be lowered substantially because theirrelative frequency was already at the basallevel.
In Schwartz's (1977a) study, peck dura-tions during DRL, FI, and FR scheduleswere analyzed in terms of their latencies orquartiles of occurrence. Schwartz found thatpeck durations increased throughout FIschedules. Since response rate, a presumedcorrelate of strength, also increased with timein the FI, the corresponding interquartile in-creases in median peck duration are entirelyconsistent with a response-strength account.On FR schedules, Schwartz reported theopposite relation. Peck durations decreasedas FR reinforcement was approached. How-ever, this conclusion is at best only weaklysupported by Schwartz's data. In only 14 of20 observations of exposure to FR 40 andFR 80 schedules were peck durations in-versely related to their quartile of occur-rence. Moreover, the average difference in
duration between the first and last quartiles,based on all subjects exposed to FR sched-ules in Figure 5 of the Schwartz study, wasless than 5 msec. In any case, these data donot violate the predictions of a strengthaccount. Response rate, an index of strength,can only be calculated for exposure to FR40 schedules, because response rates arenot presented for performances on FR 80schedules. The mean FR 40 response ratesfor the first and last quartiles, averagedacross observations, were, respectively, 62.5and 66. 6 responses per minute. Interquartilemedian peck-duration changes were similarlyinsubstantial. In only 6 of 12 observationsdid peck durations decrease across quartiles,the main effect reported by Schwartz. More-over, there was essentially no difference be-tween the initial and terminal quartiles'median peck durations when averaged acrossobservations (calculated from Figure 5 ofthe Schwartz study). Insofar as rate (a cor-relate of strength) and duration were essen-tially unchanged across quartiles of occur-rence, these data seem quite compatible withan account based on strength.
As regards discrete-trials DRL schedules,Schwartz (1977a) found that short-latencypecks were predominantly of short duration,while long-latency pecks were primarily fromthe long-duration response class. As wasshown in Figure 6, a strength account pre-dicts an inverse relation between response la-tency and peck duration. Consequently, thisfinding seems, at first blush, incompatiblewith a response-strength interpretation ofpeck duration. Yet, another explanation ofthese results can be offered that is consistentwith a strength account. In the Schwartzstudy, DRL peck-latency distributions werebimodal, one mode being in the vicinity of.5 to 1.0 sec and the second mode beinggreater than 6,0 sec. Based on Reynolds's(1966) demonstration that pigeons are moreskilled at discriminating DRL intervals thanis suggested by their IRT distributions, it isassumed that Schwartz's subjects discrimi-nated that short-latency pecks were notreinforced but emitted them nonetheless.Surely, the strength of these pecks, whichoccur in EXT, is less than that for long-latency pecks, which could result in rein-

20 JOHN M. ZIRIAX AND ALAN SILBERBERG
forcement. Consistent with a strength ac-count, the lower response strength associatedwith short-latency pecks results in a shortermedian peck duration than for long-latencypecks, the very results Schwartz obtained.
One finding remains to be evaluated interms of a strength account before this dis-cussion can be considered essentially com-plete: Schwartz and Williams's (1972)original finding that only pecks of longduration are sensitive to outcome-contingentcontrol. In their study, peck-duration dis-tributions were recorded from pigeons ex-posed to VR 5 schedules of reinforcement.Then, depending on the subject, peck dura-tions that fell within the shortest third,quartile, or quintile of the distribution werereinforced. After several sessions of expo-sure to this differential reinforcement-of-duration procedure, pigeons were returned tothe original VR 5 schedule. Based on thepeck-duration distributions from this secondexposure to VR 5 schedules, the fourth long-est quintile of each subject's durations wasnow reinforced. Schwartz and Williamsfound that pigeons had difficulty selectivelyemitting short-duration pecks, it taking anaverage of between 8 and 15 responses toproduce each reinforcement. However, whenlong-duration pecks were reinforced, theproportion of long-duration emissions in-creased substantially, so that every secondor third peck, on average, produced rein-forcement. Based on these data, they con-cluded that only long-duration pecks can bedifferentially reinforced.
Our reinterpretation of these results isbased on the often-reported finding that VRschedules produce high rates of responding(see Ferster & Skinner, 1957), a finding thatsuggests that the response strength en-gendered by these schedules is also high. Ifpeck duration is a correlate of strength,the pigeons exposed to VR schedules in theSchwartz and Williams (1972) study arelikely to have naturally emitted high fre-quencies of long-duration pecks. When theselong-duration pecks were differentially rein-forced, the schedule-dictated increases intheir frequencies readily occurred, becausethese increases were consonant with thedurations these schedules normally support.Therefore, long-duration pecks appeared to
be sensitive to outcome-contingent control.However, when short-duration pecks weredifferentially reinforced, the long-durationpecks engendered by the high-strength VRschedules ran counter to the short-durationpeck contingencies imposed. Hence, it ap-peared that short-duration pecks were in-sensitive to reinforcement. While Schwartzand Williams were right in concluding thatshort-duration pecks were selectively insen-sitive to differential reinforcement, they wereright for the wrong reasons. Selective insen-sitivity was not due to a process differencebetween long- and short-duration pecks butto the difficulty of emitting short-durationpecks on high-rate, high-strength schedules.
The present study's failure to find aprocess difference between pecks of short andlong duration necessitated a reevaluation ofSchwartz and Williams's (1972) two-process account of the key peck and of theliterature that has been interpreted in termsof it. It should be acknowledged that much ofthis reinterpretation, which has been basedon a strength account, is ad hoc. For thisreason, it is still premature to label con-clusively peck duration as an epiphenomenalcorrelate of the response strengths that dif-ferent schedules engender. This limitationnotwithstanding, we know of no alternativeto an account based on strength that holdsgreater promise of integrating the peck-durations literature within a single concep-tual framework.
References
Brandauer, C. M. The effects of uniform prob-abilities of reinforcement upon the response rateof the pigeon (Doctoral dissertation, ColumbiaUniversity, 1958). Dissertation Abstracts Inter-national, 1959, 19, 3371. (University MicrofilmsNo. 59-1478).
Brown, P. L., & Jenkins, H. M. Auto-shaping ofthe pigeon's key-peck. Journal of the Experi-mental Analysis of Behavior, 1968, 11, 1-8.
Brownstein, A. J., & Balsam, P. D. A search forconditioned reinforcement effects in negativeautomaintenance of keypecking. Bulletin of thePsychonomic Society, 1975, 6, 165-168.
Catania, A. C., & Reynolds, G. S. A quantitativeanalysis of the responding maintained by intervalschedules of reinforcement. Journal of the Ex-perimental Analysis of Behavior, 1968, 11, 327-383.
Gumming, W. W., & Berryman, R. Some data onmatching behavior in the pigeon. Journal of the

DISCRIMINATION AND EMISSION OF KEY-PECK DURATION 21
Experimental Analysis a} Behavior, 1961, 4,281-284.
Ferster, C. B., & Skinner, B. F. Schedules of rein-forcement. New York: Appleton-Century-Crofts,19S7.
Gamzu, E. Associative and instrumental factorsunderlying the performance of a complex skeletalresponse (Doctoral dissertation, University ofPennsylvania, 1971). Dissertation Abstracts In-ternational, 1972, 32, 4887. (University Micro-films No. 72-6152).
Gamzu, E., & Williams, D. R. Classical condition-ing of a complex skeletal act. Science, 1971, 171,923-925.
Gamzu, E., & Williams, D. R. Associative factorsunderlying the pigeon's key pecking in auto-shaping procedures. Journal of the ExperimentalAnalysis of Behavior, 1973, 19, 225-232.
Gonzalez, F. A. Effects of partial reinforcement(25%) in an autoshaping procedure. Bulletinof the Psychonomic Society, 1973, 2, 299-301.
Herrnstein, R. J., & Loveland, D. H. Maximizingand matching on concurrent ratio schedules.Journal of the Experimental Analysis of Be-havior, 1975, 24,107-116.
Jenkins, H. M., & Moore, B. R. The form of theauto-shaped response with food or water rein-forcers. Journal of the Experimental Analysis ofBehavior, 1973, 20, 163-181.
Kelleher, R. T. Chaining and conditioned rein-forcements. In W. K. Honig (Ed.), Operantbehavior: Areas of research and application.Englewood Cliffs, N. J.: Prentice-Hall, 1966.
Keller, K. The role of elicited responding in be-havioral contrast. Journal of the ExperimentalAnalysis of Behavior, 1974, 21, 249-257.
Miller, S., & Konorski, J. On a particular formof conditioned reflex. Journal of the Experi-mental Analysis of Behavior, 1969, 12, 187-189.[Originally published in Les comptes rendus desseances de la societe de biologie. (Societe polo-naise de biologie), 1928, 99, 1155-1158.]
Rachlin, H. Contrast and matching. PsychologicalReview, 1973, 80, 217-234.
Rescorla, R. A., & Solomon, R. L. Two-processlearning theory: Relationships between Pavlovianconditioning and instrumental learning. Psycho-logical Review, 1967, 74, 151-182,
Reynolds, G. S. Behavioral contrast. Journal ofthe Experimental Analysis of Behavior, 1961,4, 57-71.
Reynolds, G. S. Discrimination and emission oftemporal intervals by pigeons. Journal of theExperimental Analysis of Behavior, 1966, 9,65-68.
Rilling, M. Number of responses as a stimulus infixed interval and fixed ratio schedules. Jour-nal of Comparative and Physiological Psychol-ogy, 1967, 63,60-65.
Schwartz, B. Studies of operant and reflexive keypecks in the pigeon. Journal of the ExperimentalAnalysis of Behavior, 1977, 27, 301-313. (a)
Schwartz, B. Two types of pigeon key pecking:Suppression of long but not short-duration key
pecks by duration-dependent shock. Journal ofthe Experimental Analysis of Behavior, 1977,27, 393-398. (b)
Schwartz, B., & Gamzu, E. Pavlovian control ofoperant behavior: An analysis of autoshapingand its implications for operant conditioning. InW. K. Honig & J. E. R. Staddon (Eds.),Handbook of operant behavior. Englewood Cliffs,N. J.: Prentice-Hall, 1977.
Schwartz, B., Hamilton, B., & Silberberg, A.Behavioral contrast in the pigeon: A study of theduration of key pecking maintained on multipleschedules of reinforcement. Journal of the Ex-perimental Analysis of Behavior, 1975, 24, 199-206.
Schwartz, B., & Williams, D. R. Discrete-trialsspaced responding in the pigeon: The dependenceof efficient performance on the availability of astimulus for collateral pecking. Journal of theExperimental Analysis of Behavior, 1971, 16,155-160.
Schwartz, B., & Williams, D. R. Two differentkinds of key peck in the pigeon: Some propertiesof responses maintained by negative and posi-tive response-reinforcer contingencies. Journal ofthe Experimental Analysis of Behavior, 1972,18, 201-216.
Sheffield, F. D. Relation between classical condi-tioning and instrumental learning. In W. F. Pro-kasy (Ed.), Classical conditioning. New York:Appleton-Century-Crofts, 1965.
Sidman, M., & Rosenberger, P. B. Several methodsfor teaching serial position sequences to monkeys.Journal of the Experimental Analysis of Be-havior, 1967, 10, 467-478.
Smith, R. F. Topography of the food-reinforcedkey peck and the source of 30-millisecond inter-response times. Journal of the ExperimentalAnalysis of Behavior, 1974, 21, 541-551.
Staddon, J. E. R. Some properties of spaced re-sponding in pigeons. Journal of the ExperimentalAnalysis of Behavior, 1965, 8, 19-27.
Stubbs, D. A. The discrimination of stimulus dura-tion by pigeons. Journal of the ExperimentalAnalysis of Behavior, 1968, 11, 223-238.
Terrace, H. S. Discrimination learning, the peakshift, and behavioral contrast. Journal of theExperimental Analysis of Behavior, 1968, 11,724-741.
Williams, D. R., & Williams, H. Automaintenancein the pigeon: Sustained pecking despite contin-gent nonreinforcement. Journal of the Experi-mental Analysis of Behavior, 1969, 12, 511-520.
Wolin, B. R. Difference in manner of pecking a keybetween pigeons reinforced with food and withwater. In A. C. Catania (Ed.), Contemporaryresearch in operant behavior. Glenview, 111.:Scott, Foresman, 1968.
Woodruff, G., & Williams, D. R. The associativerelation underlying autoshaping in the pigeon.Journal of the Experimental Analysis of Be-havior, 1976, 26, 1-13.
Received May 24, 1977Revision received September 23, 1977 •
Copyright © 2022 FDOKUMEN




















