
Direct and dam-mediated effects of prenatal dexamethasone on emotionality, cognition and HPA axis in...
12
Direct and dam-mediated effects of prenatal dexamethasone on emotionality, cognition and HPA axis in adult Wistar rats Jonas Hauser, Joram Feldon ⁎, Christopher R. Pryce Behavioural Neurobiology Laboratory, Swiss Federal Institute of Technology Zurich, Schoerenstrasse 16, CH - 8603 Schwerzenbach, Switzerland abstract article info Article history: Received 4 March 2009 Revised 3 July 2009 Accepted 7 July 2009 Available online 15 July 2009 Keywords: Prenatal programming HPA axis Memory Learning Endocrinology Prenatal stress can affect foetal neurodevelopment and result in increased risk of depression in adulthood. It promotes increased maternal hypothalamo–pituitary–adrenal gland (HPA) secretion of glucocorticoid (GC), leading to increased foetal and maternal GC receptor activity. Prenatal GC receptor activity is also increased during prenatal treatment with dexamethasone (DEX), which is commonly prescribed as a prophylactic treatment of preterm delivery associated morbid symptoms. Here, we exposed pregnant Wistar rats to 0.1 mg/kg/d DEX during the last week of pregnancy and performed cross-fostering at birth. In the adult offspring we then studied the effects of prenatal DEX exposure per se and the effects of rearing by a dam exposed to prenatal DEX. Offspring were assessed in the following paradigms testing biobehavioural processes that are altered in depression: progressive ratio schedule of reinforcement (anhedonia), Porsolt forced swim test (behavioural despair), US pre-exposure active avoidance (learned helplessness), Morris water maze (spatial memory) and HPA axis activity (altered HPA function). Responsiveness to a physical stressor in terms of HPA activity was increased in male offspring exposed prenatally to DEX. Despite this increased HPA axis reactivity, we observed no alteration of the assessed behaviours in offspring exposed prenatally to DEX. We observed impairment in spatial memory in offspring reared by DEX exposed dams, independently of prenatal treatment. This study does not support the hypothesis that prenatal DEX exposure leads to depression-like symptoms in rats, despite the observed sex-specific programming effect on HPA axis. It does however emphasise the importance of rearing environment on adult cognitive performances. © 2009 Elsevier Inc. All rights reserved. Introduction Since its first use in 1972 by Liggins and Howie (1972), exposure to synthetic glucocorticoid (GC; such as dexamethasone (DEX)) has become common practice in prenatal medicine to prophylactically impede preterm delivery associated morbid symptoms, such as respira- tory distress syndrome and intra-ventricular haemorrhage. Synthetic GC act through specific binding to the GC receptor (GR), which can modulate the expression of genes containing GC responsive elements in their promoter (De Kloet et al., 1998). This treatment has been shown to increase the survival rate of preterm infants, but there are major concerns about the potentially harmful long-term effects, which could be mediated by prenatal programming of the GR. Prenatal programming is the setting up of adulthood receptor levels, based on the availability of its ligand during early stages of development (Seckl, 2004). In fact, although the American National Institute of Health supports the use of prenatal GC exposure in cases of women at risk of preterm delivery (National Institutes of Health Consensus Development Panel, 1995), it has also emphasised the importance of performing further research into the potentially harmful long-term effects of prenatal exposure to repeated doses of GC on physiology, endocrinology and behaviour (National Institutes of Health Consensus Development Panel, 2001). It is commonly accepted that many diseases have a component of developmental origin (Barker, 2004; Nathanielsz and Padmanabhan, 2006). Among the environmental challenges known to affect foetal development is sustained exposure of the pregnant female to stress. A stressful event results in increased hypothalamo–pituitary–adrenal (HPA) axis activity in the pregnant female, which in turn leads to increased plasma GC (cortisol in primates and corticosterone in rodents (Ganong, 1999)), that is, the natural ligand of GR. Maternal stress has been shown to induce increased maternal and foetal GC levels (Erisman et al., 1990), despite the presence of an enzymatic barrier in the placenta (Benediktsson et al., 1997). The enzymatic barrier that reduces maternal-to-foetal GC exchange is ineffective against synthetic GC. Therefore prenatal stress and prenatal GC exposure both result in GR activation in the foetus. In rat, both prenatal stress and prenatal GC exposure have been associated in adulthood with: increased anxiety (Estanislau and Morato, 2006; Welberg et al., 2001), impaired HPA negative feedback (Weinstock, 2005; Welberg et al., 2001), and impairment in memory (Brabham et al., 2000; Zagron and Weinstock, 2006). Furthermore, prenatal stress has been associated with increased behavioural despair (Smith et al., 2004) and impaired social behaviour (Patin et al., 2005). Such a constellation of phenotypes following prenatal GR activation is of Hormones and Behavior 56 (2009) 364–375 ⁎ Corresponding author. Fax: +41 44 655 72 03. E-mail address: [email protected] (J. Feldon). 0018-506X/$ – see front matter © 2009 Elsevier Inc. All rights reserved. doi:10.1016/j.yhbeh.2009.07.003 Contents lists available at ScienceDirect Hormones and Behavior journal homepage: www.elsevier.com/locate/yhbeh
Transcript of Direct and dam-mediated effects of prenatal dexamethasone on emotionality, cognition and HPA axis in...

Hormones and Behavior 56 (2009) 364–375
Contents lists available at ScienceDirect
Hormones and Behavior
j ourna l homepage: www.e lsev ie r.com/ locate /yhbeh
Direct and dam-mediated effects of prenatal dexamethasone on emotionality,cognition and HPA axis in adult Wistar rats
Jonas Hauser, Joram Feldon ⁎, Christopher R. PryceBehavioural Neurobiology Laboratory, Swiss Federal Institute of Technology Zurich, Schoerenstrasse 16, CH - 8603 Schwerzenbach, Switzerland
⁎ Corresponding author. Fax: +41 44 655 72 03.E-mail address: [email protected] (J. Feldon
0018-506X/$ – see front matter © 2009 Elsevier Inc. Adoi:10.1016/j.yhbeh.2009.07.003
a b s t r a c t
a r t i c l e i n f oArticle history:Received 4 March 2009Revised 3 July 2009Accepted 7 July 2009Available online 15 July 2009
Keywords:Prenatal programmingHPA axisMemoryLearningEndocrinology
Prenatal stress can affect foetal neurodevelopment and result in increased risk of depression in adulthood. Itpromotes increased maternal hypothalamo–pituitary–adrenal gland (HPA) secretion of glucocorticoid (GC),leading to increased foetal and maternal GC receptor activity. Prenatal GC receptor activity is also increasedduring prenatal treatment with dexamethasone (DEX), which is commonly prescribed as a prophylactictreatment of preterm delivery associated morbid symptoms. Here, we exposed pregnant Wistar rats to0.1 mg/kg/d DEX during the last week of pregnancy and performed cross-fostering at birth. In the adultoffspring we then studied the effects of prenatal DEX exposure per se and the effects of rearing by a damexposed to prenatal DEX. Offspring were assessed in the following paradigms testing biobehaviouralprocesses that are altered in depression: progressive ratio schedule of reinforcement (anhedonia), Porsoltforced swim test (behavioural despair), US pre-exposure active avoidance (learned helplessness), Morriswater maze (spatial memory) and HPA axis activity (altered HPA function). Responsiveness to a physicalstressor in terms of HPA activity was increased in male offspring exposed prenatally to DEX. Despite thisincreased HPA axis reactivity, we observed no alteration of the assessed behaviours in offspring exposedprenatally to DEX. We observed impairment in spatial memory in offspring reared by DEX exposed dams,independently of prenatal treatment. This study does not support the hypothesis that prenatal DEX exposureleads to depression-like symptoms in rats, despite the observed sex-specific programming effect on HPA axis.It does however emphasise the importance of rearing environment on adult cognitive performances.
© 2009 Elsevier Inc. All rights reserved.
Introduction
Since its first use in 1972 by Liggins and Howie (1972), exposure tosynthetic glucocorticoid (GC; such as dexamethasone (DEX)) hasbecome common practice in prenatal medicine to prophylacticallyimpede preterm delivery associated morbid symptoms, such as respira-tory distress syndrome and intra-ventricular haemorrhage. Synthetic GCact through specific binding to theGC receptor (GR),which canmodulatethe expression of genes containing GC responsive elements in theirpromoter (De Kloet et al., 1998). This treatment has been shown toincrease the survival rate of preterm infants, but there aremajor concernsabout thepotentially harmful long-termeffects,which couldbemediatedby prenatal programming of the GR. Prenatal programming is the settingup of adulthood receptor levels, based on the availability of its ligandduring early stages of development (Seckl, 2004). In fact, although theAmerican National Institute of Health supports the use of prenatal GCexposure in cases of women at risk of preterm delivery (NationalInstitutes of Health Consensus Development Panel, 1995), it has alsoemphasised the importance of performing further research into thepotentially harmful long-term effects of prenatal exposure to repeated
).
ll rights reserved.
doses of GC on physiology, endocrinology and behaviour (NationalInstitutes of Health Consensus Development Panel, 2001).
It is commonly accepted that many diseases have a component ofdevelopmental origin (Barker, 2004; Nathanielsz and Padmanabhan,2006). Among the environmental challenges known to affect foetaldevelopment is sustained exposure of the pregnant female to stress. Astressful event results in increased hypothalamo–pituitary–adrenal(HPA) axis activity in the pregnant female, which in turn leads toincreased plasma GC (cortisol in primates and corticosterone inrodents (Ganong, 1999)), that is, the natural ligand of GR. Maternalstress has been shown to induce increased maternal and foetal GClevels (Erisman et al., 1990), despite the presence of an enzymaticbarrier in the placenta (Benediktsson et al., 1997). The enzymaticbarrier that reduces maternal-to-foetal GC exchange is ineffectiveagainst synthetic GC. Therefore prenatal stress and prenatal GCexposure both result in GR activation in the foetus. In rat, bothprenatal stress and prenatal GC exposure have been associated inadulthood with: increased anxiety (Estanislau and Morato, 2006;Welberg et al., 2001), impaired HPA negative feedback (Weinstock,2005; Welberg et al., 2001), and impairment in memory (Brabhamet al., 2000; Zagron and Weinstock, 2006). Furthermore, prenatalstress has been associated with increased behavioural despair (Smithet al., 2004) and impaired social behaviour (Patin et al., 2005). Such aconstellation of phenotypes following prenatal GR activation is of

365J. Hauser et al. / Hormones and Behavior 56 (2009) 364–375
potential relevance to the DSM-IV symptomatology of depression,which has two mandatory symptoms: depressed mood and anhedo-nia. Additional symptoms/features include diminished ability to thinkand concentrate and HPA alterations. In fact, Weinstock has proposedprenatal stress in the rat as a developmental model of depressionsymptoms (Weinstock, 2000).
In the current study, the overall aimwas to describe the long-termeffects of prenatal GC exposure on behavioural and endocrineresponses to challenge tests in rats. To achieve this we exposedpregnant Wistar rats to DEX (0.1 mg/kg/d per os) during the lastweek of pregnancy: this is the time-regimen that has been used mostfrequently in rat studies and we wanted to be consistent with theseprior studies (e.g. Brabham et al., 2000; Welberg et al., 2001). In theclinic, if there is a risk of preterm delivery then, at foetal age 24–32 weeks, one course of 2–4 doses at 12–24 -h intervals isrecommended, although repeated dosing for as long as the foetusremains in utero are also common (National Institutes of HealthConsensus Development Panel, 1995, 2001). As there are reports ofinteractions between prenatal treatment and maternal behaviour(Brabham et al., 2000; Hauser et al., 2006), we performed cross-fostering at birth, to enable separation of any direct prenatal effectfrom any rearing dam-mediated effect of the treatment. The adultmale and female offspringwere tested in terms of prenatal and rearingdam treatment effects on behaviour in open field for locomotoractivity, elevated plus maze test of anxiety, progressive ratio scheduleof appetitive motivation/anhedonia, Porsolt forced swim test ofcoping, US pre-exposure test of escape and avoidance coping, Morriswater maze test of spatial learning/memory using both working andreference memory paradigms; and effects on HPA activity in basal andrestraint-stress states.
Materials and methods
Subjects, prenatal treatment and cross-fostering
Wistar rats (Harlan, Borchen, Germany) used for breedingwere leftundisturbed for two weeks after arrival, to allow for habituation. Thebreeding and experimental animals were housed in social groups of 2–4 animals in Macrolon cage type IV (195 mm×380 mm×600 mm),under a reversed light–dark cycle (light ON: 2000, OFF: 0800) in adedicated animal facility with controlled temperature (21±1 °C) andhumidity (55±5%). The animals had access to food (Kliba 3430,Klibamühlen, Kaiseraugst, CH) and tapwater ad libitum, except duringthe test of progressive ratio schedule of reinforcement (see below). Allthe experimentswere performed in accordancewith the regulations ofthe Swiss Federal Veterinary Office.
Breeding females were handled daily for one week. The followingweek, their vaginal impedancewas measured daily (EC40 Estrus CycleMonitor; FST; Foster City, USA) until a peak impedance N8000 Ω wasobserved, indicating that the female was in pro-oestrus. On that day,taken as embryonic day 0 (E0), the female was placed with a male at1500. At 0800 the following day the male was removed to its socialgroup, and the female was transferred to a Macrolon cage type II(140 mm×180 mm×345 mm).
Pregnant females were assigned randomly to one of the treatmentgroups. The treatment was performed as described in Hauser et al.(2006). Briefly, between E 15 and birth, pregnant females wereadministered either 0.1 mg/kg/d DEX (D-1756, Sigma, Buchs,Switzerland) or vehicle (0.01% ethanol) via the drinking water. Thedata reported here were obtained from the offspring of two cohortsthat underwent the same treatment and cross-fostering procedures:Cohort 1 was composed of 8 VEH and 10 DEX dams and Cohort 2 wascomposed of 12 VEH and 14 DEX dams. From E21, dams were checkedtwice daily for birth, at 0900 and 1800. The day that pups were firstobserved was taken as postnatal day 1 (PND 1). On PND 1, the waterbottles were replaced by new ones containing tap water, and pups
were sexed, weighed and toe-clipped for identification (DEX versusVEH). Culling was performed to yield litters of 4 female and 4 malepups. On the same day, cross-fostering was carried out to yield littersof 2 VEH males, 2 DEX males, 2 VEH females and 2 DEX females, withall pups per litter unrelated to each other and to the dam towhich theywere allocated. This procedure produced four treatment groups interms of the combination of prenatal drug exposure of the offspringand pregnancy drug exposure of the dam: VEHO–VEHD (i.e. offspringexposed to VEH in utero and reared by a VEH-treated dam); VEHO–
DEXD; DEXO–VEHD and DEXO–DEXD (N=16males and 16 females pertreatment per cohort).
All offspringwereweaned on PND 21 and put in social groups of 2–4 animals from different dams but of the same treatment and sex.Offspring body weight was measured weekly, during the cagechanging procedure, until weaning, and also at PND 50. Beginning atage PND 90, subjects in Cohort 1 were tested sequentially in theprogressive ratio schedule of reinforcement (PND 100), Porsolt forcedswim test (PND 175) and US pre-exposure/active avoidance test (PND190). Subjects in Cohort 2 were bred following the same procedure,and then tested sequentially in the elevated plus maze (PND 90), openfield (PND 92), the HPA axis response to restraint stress (PND 100),and the water maze using protocols for assessment of both referenceand working memory (PND 300).
Behavioural tests
Elevated plus mazeThe elevated plus maze consisted of a central 10(L)×10(W) cm
square platform connected to four arms 10 (W)×45 (L) cm, with 2opposing arms closed by walls 40 cm high and the 2 other opposingarms open, with just an edge of 2 mm. The maze was elevated at 1 mon a central support. The testing room was illuminated by low light(15 lx in the open arms,10 lx in the centre and 5 lx in the closed arms).A video camera was fixed above the plus maze to allow videorecording as well as tracking motion for analysis using the Ethovi-sion© system (Noldus Information Technology, Wageningen, Nether-lands), which computed time spent in each arm and total distancemoved. Each test lasted 5 min , starting with the rat placed on thecentral platform facing an open arm. The Ethovision system calculatedthe total distance moved and the time spent in each arm. From thevideo tapes, a trained experimenter scored entries in the open andclosed arms and the total number of each of the following 4behaviours: a) stretch attend posture: the rat stretches forward andthen retracts to its original position, with the back of its body stayingin the same position; b) grooming: the rat grooms its body with handsor mouth; c) rearing: the rat lifts its front paws from the floor, exceptwhen performing grooming; d) head dipping: the animal looksdownwards over the edge of the open arm. Total arm entries anddistance moved were used as an index of locomotion; the ratio ofopen-arm entries to total arm entries and the ratio of time spent in theopen arms to total time spent in all arms were used as indices ofanxiety. The maze was cleaned with ethanol 50% between each trialand allowed to dry. Males were tested first, then females on asubsequent day.
Open fieldThe open field arena consisted of a grey plastic square with walls
76(L)×76(W)×49(H). The testing roomwas illuminated by low light(15 lx). A video camera was fixed above the arenas to allow videorecording as well as tracking motion for analysis using the Ethovi-sion© system (Noldus Information Technology, Wageningen, Nether-lands). Each trial lasted 1 h, starting with the animal placed in thecentre of the arena. The Ethovision system calculated the distancemoved, which was taken as an index of locomotor activity. Subjectswere tested 4 at a time in four different open fields, counterbalancingfor treatment. The open fields were cleaned with ethanol 50% and

366 J. Hauser et al. / Hormones and Behavior 56 (2009) 364–375
allowed to dry between each trial. Males were tested first, thenfemales on a subsequent day.
Progressive ratio schedule of appetitive motivationAnimals were tested in four test chambers (E10-10TC, Coulbourn
Instruments, Allentown, USA) enclosed within sound-attenuatingboxes (E10-20) and fitted with a removable response lever and aliquid dipper (E14-05). The internal dimensions of each chamber were30(L)×25(W)×21.5(H) cmwith a grid floor. During the experimentalsessions, each chamber was illuminated by a diffuse dim light source(house light, 4 lx), mounted 19 cm above the grid floor in the centre ofthe left side wall. The removable lever was 3 cm above the grid flooron the right side wall. The reward magazine was placed in the centreof the right side wall, 2 cm above the grid and 3 cm from the responselever. When a rat had performed the necessary number of leverpresses, the liquid dipper transferred a drop of sucrose solution(0.01 ml, 7%) from a reservoir into the reward magazine. Each nosepoke in the reward magazine was counted by photocell detectors. Apersonal computer (Compaq), together with universal environmentinterfaces (E91-12), controlled the sessions and recorded thebehavioural data.
All animals received an initial 1-h exposure to a 7% sucrose solutionin their home cage. Within the next three days, subjects were placedprogressively on water restriction of 23 h, before lever-press trainingbegan. On each fixed ratio (FR) training day, animals received waterfor 1 h in the home cage at least one hour after the training session.Following presentation of 50 free reinforcements (each one sucrosedrop), the animals were trained to press a lever for sucrosereinforcement, according to a FR-1 schedule. On each trial, the leverwas presented for 15 s followed by an inter-trial interval of 5 s, duringwhich the lever was retracted. The lever was retracted if more than120 s passed after performance of a correct lever-press responsewithout consumption of the sucrose drop. FR-1 training wascompleted when an animal earned and consumed at least 40 sucrosereinforcements during a 20-min training session. After all animalsreached this criterion, they were gradually introduced to an FR-5schedule. FR training was completed for all animals within 7 days.
After all animals completed FR-5 training, they were tested on aprogressive ratio (PR) schedule. Water was freely available in thehome cage except for 2 h of water deprivation directly before each PRtest session. Under the PR schedule, the number of lever pressesrequired in order to receive sucrose reinforcementwas incremented inthe followingmanner: 1, 2, 3, 4, 5, 6, 7, 8,10,12,14,16,18, 20, 22, 24, 28,32, 36, 40, 44, 48, 52, 56, 64, 72, 80, 88, 96, 104, 112, 120, 136, 152. Thebreak point ratio is the final response ratio completed, e.g. an animalwhich reaches the 20 lever presses ratio but then only presses 12times at ratio 22 will have a break point ratio of 20. Each PR sessionlasted 30 min, and on each trial the lever was presented for 300 smaximum. Subjects were tested over 10 days. On day 11, a 7% sucrosesolution free consumption test was run, where the subjects had accessfor 10 min to a bottle of sucrose 7% solution, and the volumeconsumed was calculated.
Aversive US pre-exposure test using the two way active avoidanceprocedure
Behavioural testing was conducted in eight identical shuttle boxes(Coulbourn Instruments, Allentown, PA; model E10-16TC), each set ina sound- and light-attenuating ventilated shell (model E10-20). Theshuttle box chambers measured 35(L)×17(W)×21.5(H) cm, with agrid floor divided into two identical compartments by an aluminiumhurdle (17(W)×4(H) cm). The hurdlewas thin (1 mm) to prevent theanimals from balancing on it to avoid the footshocks, which weresupplied to the grid floor by a constant direct current source (modelE13-14) and a scanner (model E13-13) set at 0.5 mA intensity. Duringthe experimental sessions each chamber was illuminated by a diffuselight source (house light, 8 lx), mounted 19 cm above the grid floor in
the centre of the side walls. The conditioned stimulus (CS) was a 10-stone of 85 dB, produced by a tone module (model E12-02) placedbehind the shuttle box on the floor of the shell. Background noise wasprovided by a ventilation fan affixed to each isolation cubicle.
Subjects from each treatment group and each sex (N=12–16)were split into shock pre-exposed (PE, N=6–8 per treatment andsex) and non-pre-exposed (NPE, N=6–8 per treatment and sex)subgroups. The test procedure comprised three stages over threeconsecutive days: two daily sessions of pre-exposure to both the shockand the apparatus (PE group) or to the apparatus only (NPE group)and a conditioning session on the third day. On each of days 1 and 2,the PE rats received 25 presentations of shock of 2 s and 0.5 mA, witha variable inter-stimulus interval of 50±30 s. Each NPE animal wasplaced in the shuttle box, with the house light on, for an equivalentperiod of time. A general evaluation of each animal's activity level wasobtained by recording the total number of crossings during all NPEand PE sessions. On the third day, each animal was placed into ashuttle box and received 100 conditioning trials on a variable intervalschedule of 50±40 s. Each trial began with a 10-s tone followed by a2-s 0.5 mA shock, the tone remaining on with the shock. If the ratcrossed the barrier to the opposite compartment during the tone, thestimulus was terminated and no shock was delivered (avoidanceresponse). A crossing response during the shock terminated the toneand the shock (escape response). If the rat failed to cross during theentire tone–shock trial, the tone and the shock terminated after 12 s(escape failure). As a measure of activity, the total number of inter-trial crossings was also recorded.
Forced swim testWe used the forced swim test as described in the original paper of
Porsolt et al. (1977) with somemodifications. Briefly, a pretest sessionof 15 min was followed 24 h later by a test session of 5 min. For boththe pretest and the test sessions, conducted under low illumination(12 lx), the rats were placed in a plastic cylindrical tank (44 cm highby 32 cm in diameter) filled with tap water at 22 °C, to a depth of30 cm, so that when the head of the rat was above thewater surface itshind limbs and tail did not make contact with the tank floor. A videocamera was placed above the tank, and connected to a video recorderto record each trial for subsequent scoring. From the video tapes,specific behaviours were scored by an observer, blind to animaltreatment, using continuous scoring to determine the exact durationsand entering data into a handheld computer (Workabout, PSION,London, UK) running the Observer Mobile Support Package software(Noldus Information Technology, Wageningen, NL). Behaviouralcategories were exclusive and defined as follows: floating: no bodymovements other than those required to stay afloat; swimming:horizontal movement through the water; and struggling: strongvertical movement. The water was changed after every second sessionduring 15-min pre-testing and after every sixth session during 5-mintesting and faeces were removed after each session. Following eitherpretest or test session, the rats were dried with a towel and kept warmon a heating pad for 30 min in their home cage.
Water mazeThe water maze consisted of a circular fibreglass tank painted
black, with a diameter of 2 m, a height of 60 cm and raised 40 cmabove the floor. It was positioned in the middle of a well-lit testingroom, with several black and white spatial cues on the walls. Everyevening it was filled with a mixture of hot and cold tap water to a levelof 30 cm, and thewater temperaturewasmaintained at 21±1 °C untilthe following testing day. A rough circular platform (diameter: 11 cm)was submerged 2 cm below the water surface. A visual cue (circularwhite disk) could be placed 11 cm above the platform. Four points,equally spaced along the circumference, were arbitrarily assigned asnorth (N), east (E), south (S) and west (W); defining 4 quadrants (NE,SE, NWand SW) of equal size. Five platform locations were set,1 in the

367J. Hauser et al. / Hormones and Behavior 56 (2009) 364–375
middle of each quadrant 50 cm from centre of the pool and one in thecentre of the pool. A video camera was placed above the tank, andconnected to a video recorder and the Ethovision system (NoldusInformation Technology, Wageningen, Netherlands), allowing track-ing of the swim path and automatic calculation of time to reach theplatform, distance moved to reach the platform and swimmingvelocity.
An initial visual training test day was used to familiarize the animalwith the water maze and allow between-treatment comparison ofpossible differences in visual discrimination and motor control. Thevisual test was composed of 4 trials, with the platform equipped withthe visual cue. The platform position was changed for each trial in thefollowing sequence: SE, NE, SW and NW. The starting position of therat was always in the quadrant opposite to that in which the platformwas placed. The trial started by placing the rat gently in the waterfacing the tank wall, and finished when the animal climbed on theplatform and stayed on it for at least 2 s. Escape latency, distancemoved and velocity were directly measured and computed by theEthovision software. If the animal failed to find the platform within90 s, the experimenter guided it to the platform and an escape latencyof 90 s was scored. The rat was left on the platform for 15 s and thenplaced in a dry warm cage for 30 s before beginning the next trial. Atthe end of the last trial, the animal was dried and put in its home cagewhich was temporarily warmed. Test trials began on the next day.
Reference memory protocol. The animals were tested for 6 con-secutive days, with 6 trials per day, with the submerged platform in afixed position (SW) and without the visual cue, using a proceduresimilar to that used for visual training. The starting positions werefixed as N, SE, NW, S,Wand E. On day 7, each animal was given a probetest, by placing it in the maze at position N but without the platform.The time spent and distances moved in each quadrant were computedby the Ethovision software.
Working memory protocol. The animals were tested for 5 consecu-tive days, with 6 trials per day, using a submerged platform theposition of which was changed daily, without the visual cue and usinga procedure similar to that used for visual training. The platformpositions for days 1–5, respectively, were fixed as SE, NE, SW, NE andCentre. The starting position of the rat was dependent on the platformposition, and was as follows: for platform in quadrant NW: between Sand SE, NE, S, E, SW and between E and SE; for platform in quadrantNE: between W and SW, SE, W, S, NW and between S and SW; forplatform in quadrant SW: between E and NE, NW, E, N, SE, between Nand NE; for platform in quadrant SE: between N and NW, SW, N,W, NEand betweenWand NW; and for platform in the centre position: N, E,S, W, NW, SE. Latency and distance moved to find the platform werecomputed by the Ethovision software. The development of perfor-mance across the first two trials was taken as an index of workingmemory.
Basal and challenged HPA axis activity
Blood sampling. For basal sampling, the animals were removed fromtheir home cage and sampledwithin 1min, in an adjacent room underred light. Blood samples were collected from the dorsal tail vein, witheach animal restrained in a towel and a small cut made 0.5 cm fromthe tail tip. By gently stroking the tail, venous blood (0.2 ml) wascollected in pre-chilled ethylenediaminetetraacetic acid (EDTA)-coated capillary tubes (Microvette®, Sarstedt AG, Sevelen, CH) andstored at 4 °C for maximum 1 h. Blood was then centrifuged at 1200 gfor 20min at 4 °C, and plasmawas collected and stored at−80 °C untilRIA analyses. Basal morning (onset of active phase) and afternoon(end of active phase) blood samples were collected between 0700 and0800 (immediately after light Off) and between 1800 and 1900(before light On), respectively, and one week apart. One week later,
the same subjects were placed in restraint stress followed by serialblood sampling, conducted between 1300 and 1800. Each subject wastaken to an adjacent room, a basal blood sample was taken (time=−20 min) and immediately thereafter it was placed in a noveltransparent-plastic restraint tube (8 cm diameter) of adjustablelength, for 20 min. Immediately after restraint, a second blood samplewas taken (time=0 min) and the animal was returned to the homecage. At times 20, 40, 80 and 120 min after the end of restraint,subjects were taken to the adjacent room, bled from the tail (allsamples from the same incision) and returned to the home cage in thecolony room.
Radioimmunoassay for plasma ACTH and corticosterone. The plasmasamples were analysed for immunoreactive ACTH and corticosterone.Plasma ACTH titres were determined in a single undiluted 25 μlsample aliquot using a commercial RIA kit for determination of ACTHin human EDTA plasma (DiaSorin, Stillwater, MN, USA; modified asdescribed in Pryce et al. (2003)). Fifty microlitres of 125I tracer and50 μl of antiserum were added to 25 μl aliquot of the five standards(concentrations ranging from 4 to 120 pg/ml), the two qualitycontrols and the samples. Standards and controls were measured intriplicate. Following 20-h incubation at 4 °C, 250 μl of precipitatingcomplex was added to the tubes, whichwere then vortexed, incubated20min at room temperature and centrifuged at 1500 g for 20min. Thesupernatant was removed and samples were measured in a gammascintillation counter (Minaxi, Packard Instruments, Downers Grove,IL), 3-min count per tube. Assay sensitivity was 0.2 pg/tube at 95%binding. Intra-assay precision was 8.98% (N=10) and inter-assayprecision was 5.09% (N=5).
Plasma immunoreactive corticosterone titres were determinedwith an in-house 3 H RIA validated for rat EDTA plasma (Pryce et al.,2002). Plasma samples were diluted to bring them into the range ofthe standard curve: 1:40 for basal and 1:400 for restraint samples.Plasma samples (250 μl, in duplicate) and corticosterone standards(Sigma, C2505, 12.5–250 pg/250 μl per tube, in duplicate) wereheated in a water bath 90 °C for 5 min for binding proteindenaturation. A rabbit anti-corticosterone serum (07-120016, ICNBiomedicals, Costa Mesa, CA) was incubated with [1,2,6,7-3H]corticosterone (TRK 406, Amersham, Switzerland, ZH) and eitherstandard or sample; separation was achieved with dextran-coatedcharcoal. Assay sensitivity was 2.5 pg/tube/250 μl at 95% binding.Intra-assay precision was 4.63% (n=5) and inter-assay precision was10.89% (n=5).
Statistics
Data are presented as mean±standard error of the mean for allmeasures. The analyses were performed using SPSS 13 software andthe graphics using Sigmaplot 10 software, running onWindows XP. Tominimize the litter effect, a maximum of two littermates from anylitter was used in any test (Lehmann and Feldon, 2000). Forbehavioural and endocrine tests in adulthood, the effects of prenataltreatment (DEX, VEH) and rearing dam treatment (DEX dam, VEHdam) were determined by ANOVA with between-subject factors ofprenatal treatment, rearing dam treatment and sex, and repeatedmeasures factor that depended on the experiment: postnatal age forinfancy body weight; 5-min bins for open field; days and 1-min binsfor forced swim test; daily sessions for progressive ratio schedulereinforcement; in US pre-exposure/active avoidance, where pre-exposure was an additional between-subject factor, days for pre-exposure days and 10-trial blocks for the conditioning day; days andtrials for water maze acquisition and quadrant for probe test; and timepoint relative to end of restraint for hormone titres. The birth weightANOVA was performed on litter average so as to remove litter effect.The body weight at birth and in adulthood and the performance in thewater maze probe test data were analysed with univariate ANOVA. In

368 J. Hauser et al. / Hormones and Behavior 56 (2009) 364–375
the reference memory probe test, the degrees of freedom for thewithin subject factor of percentage time spent in each quadrant wasreduced by 1 (final df=3): because the sum of this dependentmeasure had to be 100% of total time, the degrees of freedom of therelevant interaction and error terms were adjusted accordingly (Lenget al., 2005). For water maze latencies and for hormone titres, anatural logarithmic transformationwas required to obtain the normaldistribution assumed by ANOVA. A p-value lower than 0.05 wasconsidered as significant.
Results
Body weight
Prenatal DEX exposure led to a reduction in weight at birth, duringinfancy and in adulthood (Table 1). Furthermore, rearing by a DEX-treated dam also resulted in a reduction of body weight in infancy andadulthood (Table 1). The sex difference, namely reduced body weightof females compared to males, increased throughout infancy and wasclearly visible in adulthood (Table 1). These interpretations weresupported by 2×2×2 (prenatal treatment×rearing dam treatment×sex) ANOVA of body weight at birth, which yielded a main effect ofprenatal treatment (F(1,32)=83.64; pb0.001), there were no maineffects of rearing dam treatment or sex (pb0.1). In infancy, a2×2×2×3 (prenatal treatment×rearing dam treatment×sex×age)split-plot ANOVA of body weight yielded a significant main effect ofage (F(2,228)=4826.43; pb0.001), of prenatal treatment (F(1,114)=141.23; pb0.001) of rearing dam treatment (F(1,114)=4.37, pb0.05)and of sex (F(1,114)=9.74, pb0.005). Furthermore there was asignificant prenatal treatment×age interaction (F(2,228)= 32.8;pb0.001) and a significant sex×age interaction (F(2,228)=4.89;pb0.01). Further restricted ANOVAs were performed at each age andyielded a significant main effect of sex at all ages (F(1,114)=5.84;pb0.05; F(1,114)=7.17; pb0.01and F(1,114)=9.47, pb0.005; forrespectively PND 7, 14 and 21) and a significant effect of rearingdam treatment only at PND 7 (F(1,114)=10.66, pb0.005) and PND 14(F(1,114)=4.70, pb0.05). Adult body weight was analysed with a2×2×2 (prenatal treatment×rearing dam treatment×sex) ANOVA ofbody weight at PND 55, which yielded a significant main effect ofprenatal treatment (F(1,113)=92.58, pb0.001), a significant maineffect of sex (F(1,113)=756.22; pb0.001) and an almost significantmain effect of rearing dam treatment (F(1,113)=3.61; p=0.06).
Open field and elevated plus maze
The animals were subjected to a test of anxiety using the elevatedplus maze. There was no impact of either prenatal treatment orrearing dam treatment on the two anxiety indices used or on anybehaviour scored in this test (open-arm entry ratio: prenatal: DEX:15.9±3.5, VEH: 11.4±3.6; rearing dam: DEX: 11.7±3.6, VEH: 15.7±3.5 and open-arm time ratio: prenatal: DEX: 5.3±1.4, VEH: 3.7±1.4;rearing dam: DEX:3.6±1.4, VEH: 5.4±1.4). Female rats showedincreased open-arm time ratio, open-arm entry ratio and head dipping
Table 1Body weight.
Variable PND1 PND
Prenatal treatment VEH 6.2±0.1 17.5DEX 4.3±0.1a 12.1
Rearing dam treatment VEH n.a. 15.5DEX n.a. 14.0
Sex Female 5.1±0.1 14.2Male 5.4±0.1 15.3
Body weight (mean±SEM) from birth (PND 1) until adulthood (PND55).a Indicates a significant difference in pair-wise comparisons on the specified variable (prb Indicates a trend to a significant difference in pair-wise comparisons on rearing dam tr
relative to males (open-arm entry ratio: F: 18.0±3.7, M: 9.6±3.4; open-arm time ratio: F: 6.1±1.4, M: 2.9±1.2 and head dipping:F:5.75±0.82, M: 2.68±0.51). These interpretations were supported by2×2×2 (prenatal treatment×rearing dam treatment×sex) ANOVAsonopen-arm entry ratio, open-arm time ratio, head dipping, stretchattended postures, grooming and rearing. Therewas amain effect of sexfor open-arm entry ratio (F(1;31)=12.87; pb0.005), open-arm timeratio (F(1;31)=14.96; pb0.005) and head dipping (F(1,31)=11.94;pb0.005), there were no other significantmain effect of sex, prenatal orrearing dam treatment (all p-valuesN0.1). To account for the existingsex difference in time spent in the open arm and the potential confoundthis could be regarding head dipping behaviour, which can only occur inthe open arm, we ran an ANCOVA of the number of head dippingperformed using the time spent in the open arm as a co-variate.Interestingly, in this analysis the sex main effect failed to achievesignificance (F(1,30)=1.31), suggesting that the increasedheaddippingwas due to the increased time spent in the open arm. This finding wasfurther supported by the significant correlation between head dippingand open-arm time (r=0.72, pb0.001).
The rats were tested in the open field to assess spontaneouslocomotor activity and spatial exploration. There was a clearhabituation effect of locomotor activity as evidenced by the declineof the distance moved across the successive 5-min bins over thecourse of the 1-h test period, which was observed in all subjectsindependently of the sex, the prenatal treatment or the rearing damtreatment. Distance moved during the 1-h test period was affectedneither by prenatal treatment nor by rearing dam treatment (distancemoved in meters; prenatal: DEX: 93.3±4.4, VEH: 97.1±3.8; rearingdam: DEX: 98.0±4.2, VEH: 92.2±4.0). Females showed an increasedtotal distance moved compared to males (distance moved in meters:F: 109.1±7.27, M: 66.96±5.34). These interpretations were sup-ported by a 2×2×2×12 (prenatal treatment×rearing dam treat-ment×sex×5-min bins) split-plot ANOVA on distance moved. Therewas a significant main effect of sex (F(1;31)=20.54; pb0.001) and of5-min bins (F(11;341)=43.66; pb0.001). There were no significantmain effects or interactions of either prenatal treatment or rearingdam treatment (pN0.1).
Progressive ratio schedule of reinforcement
The motivation of the subjects for a gustatory reward was assessedin the progressive ratio schedule of reinforcement. All subjectsexhibited a stable performance across the 10 sessions. Female ratsexhibited an increased performance compared to males, starting onthe second session (Table 2). Neither prenatal treatment nor rearingdam treatment affected the performance of the animals in this test.These interpretations were supported by 2×2×2×10 (prenataltreatment×rearing dam treatment×sex×sessions) ANOVAs onnumber of lever presses, number of rewards obtained and breakpoint ratio. There was a significant main effect of sessions (F(9,495)=4.99; pb0.001, F(9,495)=7.54; pb0.001 and F(9,495)=5.10;pb0.001 respectively). There was a significant sex×sessions inter-action in lever presses (F(9,495)=2.59; pb0.05) and in break point
7 PND14 PND21 PND55
±0.3 34.6±0.5 54.9±0.7 222.2±2.0±0.3a 27.1±0.5a 43.9±0.8a 193.9±2.1a
±0.3 31.6±0.4 50.0±0.6 210.8±2.1±0.4a 30.1±0.6a 48.8±0.9 205.3±2.1b
±0.3 29.9±0.5 47.8±0.7 167.7±2.0±0.3a 31.8±0.5a 51.0±0.8a 248.4±2.1a
enatal treatment, rearing dam treatment or sex, with a p-valueb0.05).eatment (p-value=0.06).
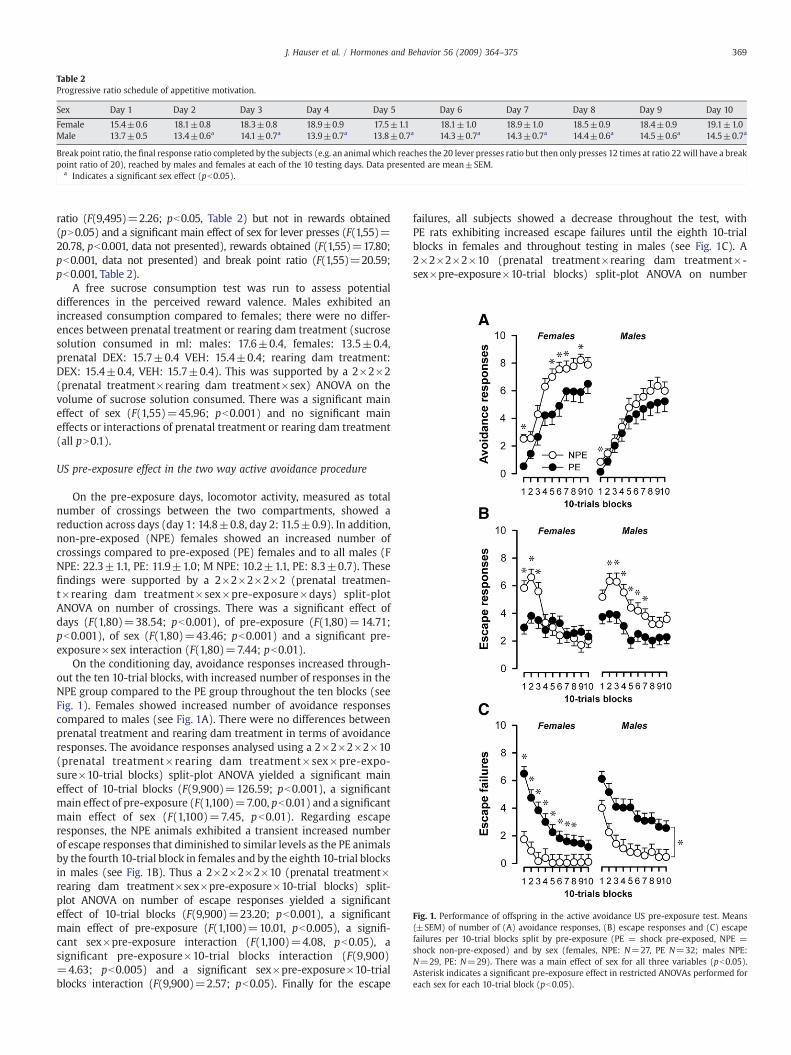
Table 2Progressive ratio schedule of appetitive motivation.
Sex Day 1 Day 2 Day 3 Day 4 Day 5 Day 6 Day 7 Day 8 Day 9 Day 10
Female 15.4±0.6 18.1±0.8 18.3±0.8 18.9±0.9 17.5±1.1 18.1±1.0 18.9±1.0 18.5±0.9 18.4±0.9 19.1±1.0Male 13.7±0.5 13.4±0.6a 14.1±0.7a 13.9±0.7a 13.8±0.7a 14.3±0.7a 14.3±0.7a 14.4±0.6a 14.5±0.6a 14.5±0.7a
Break point ratio, the final response ratio completed by the subjects (e.g. an animal which reaches the 20 lever presses ratio but then only presses 12 times at ratio 22will have a breakpoint ratio of 20), reached by males and females at each of the 10 testing days. Data presented are mean±SEM.
a Indicates a significant sex effect (pb0.05).
Fig. 1. Performance of offspring in the active avoidance US pre-exposure test. Means(±SEM) of number of (A) avoidance responses, (B) escape responses and (C) escapefailures per 10-trial blocks split by pre-exposure (PE = shock pre-exposed, NPE =shock non-pre-exposed) and by sex (females, NPE: N=27, PE N=32; males NPE:N=29, PE: N=29). There was a main effect of sex for all three variables (pb0.05).Asterisk indicates a significant pre-exposure effect in restricted ANOVAs performed foreach sex for each 10-trial block (pb0.05).
369J. Hauser et al. / Hormones and Behavior 56 (2009) 364–375
ratio (F(9,495)=2.26; pb0.05, Table 2) but not in rewards obtained(pN0.05) and a significant main effect of sex for lever presses (F(1,55)=20.78, pb0.001, data not presented), rewards obtained (F(1,55)=17.80;pb0.001, data not presented) and break point ratio (F(1,55)=20.59;pb0.001, Table 2).
A free sucrose consumption test was run to assess potentialdifferences in the perceived reward valence. Males exhibited anincreased consumption compared to females; there were no differ-ences between prenatal treatment or rearing dam treatment (sucrosesolution consumed in ml: males: 17.6±0.4, females: 13.5±0.4,prenatal DEX: 15.7±0.4 VEH: 15.4±0.4; rearing dam treatment:DEX: 15.4±0.4, VEH: 15.7±0.4). This was supported by a 2×2×2(prenatal treatment×rearing dam treatment×sex) ANOVA on thevolume of sucrose solution consumed. There was a significant maineffect of sex (F(1,55)=45.96; pb0.001) and no significant maineffects or interactions of prenatal treatment or rearing dam treatment(all pN0.1).
US pre-exposure effect in the two way active avoidance procedure
On the pre-exposure days, locomotor activity, measured as totalnumber of crossings between the two compartments, showed areduction across days (day 1: 14.8±0.8, day 2: 11.5±0.9). In addition,non-pre-exposed (NPE) females showed an increased number ofcrossings compared to pre-exposed (PE) females and to all males (FNPE: 22.3±1.1, PE: 11.9±1.0; M NPE: 10.2±1.1, PE: 8.3±0.7). Thesefindings were supported by a 2×2×2×2×2 (prenatal treatmen-t×rearing dam treatment×sex×pre-exposure×days) split-plotANOVA on number of crossings. There was a significant effect ofdays (F(1,80)=38.54; pb0.001), of pre-exposure (F(1,80)=14.71;pb0.001), of sex (F(1,80)=43.46; pb0.001) and a significant pre-exposure×sex interaction (F(1,80)=7.44; pb0.01).
On the conditioning day, avoidance responses increased through-out the ten 10-trial blocks, with increased number of responses in theNPE group compared to the PE group throughout the ten blocks (seeFig. 1). Females showed increased number of avoidance responsescompared to males (see Fig. 1A). There were no differences betweenprenatal treatment and rearing dam treatment in terms of avoidanceresponses. The avoidance responses analysed using a 2×2×2×2×10(prenatal treatment× rearing dam treatment× sex×pre-expo-sure×10-trial blocks) split-plot ANOVA yielded a significant maineffect of 10-trial blocks (F(9,900)=126.59; pb0.001), a significantmain effect of pre-exposure (F(1,100)=7.00, pb0.01) and a significantmain effect of sex (F(1,100)=7.45, pb0.01). Regarding escaperesponses, the NPE animals exhibited a transient increased numberof escape responses that diminished to similar levels as the PE animalsby the fourth 10-trial block in females and by the eighth 10-trial blocksin males (see Fig. 1B). Thus a 2×2×2×2×10 (prenatal treatment×rearing dam treatment×sex×pre-exposure×10-trial blocks) split-plot ANOVA on number of escape responses yielded a significanteffect of 10-trial blocks (F(9,900)=23.20; pb0.001), a significantmain effect of pre-exposure (F(1,100)=10.01, pb0.005), a signifi-cant sex×pre-exposure interaction (F(1,100)=4.08, pb0.05), asignificant pre-exposure×10-trial blocks interaction (F(9,900)=4.63; pb0.005) and a significant sex×pre-exposure×10-trialblocks interaction (F(9,900)=2.57; pb0.05). Finally for the escape
failures, all subjects showed a decrease throughout the test, withPE rats exhibiting increased escape failures until the eighth 10-trialblocks in females and throughout testing in males (see Fig. 1C). A2×2×2×2×10 (prenatal treatment×rearing dam treatment×-sex×pre-exposure×10-trial blocks) split-plot ANOVA on number

370 J. Hauser et al. / Hormones and Behavior 56 (2009) 364–375
of escape failures yielded a significant effect of 10-trial blocks (F(9,900)=57.62; pb0.001), a significant main effect of pre-exposure(F(1,100)= 24.45, pb0.001), a significant pre-exposure×10-trialblocks interaction (F(9,900)=5.60; pb0.005) and a significantsex×pre-exposure×10-trial blocks interaction (F(9,900)=4.45;pb0.005).
Forced swim test
In both pretest and test days, subjects exhibited an initial highstruggling, low floating, low swimming pattern, which rapidlyreversed. After the initial struggling period, females reared by aDEX-treated dam showed an increase of floating time independent ofthe day (Fig. 2). Thus a 2×2×2×15 (prenatal treatment×rearing damtreatment×sex×1-min bins) split-plot ANOVA on floating timefor pretest day yielded a significant main effect of 1-min bins(F(14,560)=35.79; pb0.001), a significant main effect of rearingdam treatment (F(1,40)=5.70, pb0.05) and a significant rearing damtreatment×sex interaction (F(1,40)=12.66; pb0.005). There wereno significant effects of prenatal treatment (pN0.1). Swimming andstruggling scores yielded the inverted, and therefore similar, patternto floating (data not presented). The test day performance wasanalysed using a 2×2×2×5 (prenatal treatment×rearing damtreatment×sex×1-min bins) split-plot ANOVA on floating time,which yielded a significant main effect of 1-min bins (F(4,160)=49.74; pb0.001) and a significant rearing dam treatment×sexinteraction (F(1,40)=4.32; pb0.05). Swimming and struggling scoresyielded the inverted, and therefore similar, pattern tofloating (data notpresented).
Water maze
The visual andmotor functions of the animals were assessed in thevisual cued version of the water maze. In this task, all animals showedlearning, measured as the decrease in latency to reach platform (datanot presented). In addition, females exhibited an overall reducedlatency to reach platform (data not presented). These interpretationswere supported by a 2×2×2×4 (prenatal treatment×rearing damtreatment×sex×trials) split-plot ANOVA on latency to reach plat-form, which yielded a significant main effect of trials (F(3,168)=30.18; pb0.001) and a main effect of sex (F(1,56)=5.43; pb0.05).There were no significant main effects or interactions of prenataltreatment or rearing dam treatment. Swimming distance presented asimilar pattern (data not presented).
Fig. 2. Floating time in the Porsolt forced swim test. Means (±SEM) of floating timesplit by rearing dam treatment for females and males during pretest and test days(females N=24; males N=24). There was a significant sex×rearing treatmentinteraction on both pretest and test days (pb0.05). Asterisk indicates a female specificsignificant rearing dam treatment effect (pb0.05).
Reference memoryReference memory performance was measured as the reduction of
latency to reach platform observed across testing days. Interestingly,this learning was reduced in rats reared by DEX-treated dams (Fig.3A). These interpretations were supported by a 2×2×2×6×6(prenatal treatment×rearing dam treatment×sex×trials×days)split-plot ANOVA on latency to reach platform, which yielded asignificant main effect of days (F(5; 280)=61.66; pb0.001) and asignificant main effect of rearing dam treatment (F(1,56)=4.75;pb0.05). Therewere no significantmain or interaction effects of eitherprenatal treatment or sex (pN0.1). Swimming distance presented asimilar pattern (data not presented). For swim velocity there was noeffect of sex, prenatal treatment or rearing dam treatment (pN0.1).
In the probe test, all subjects showed preference for the quadrantthat contained the platform during the acquisition (Fig. 3B). The2×2×2×4 (prenatal treatment×rearing dam treatment×sex×qua-drant) split-plot ANOVA of time spent in each quadrant yielded asignificant main effect of quadrant (F(2,168)=38.57; pb0.001). Therewere no significant effects of prenatal or rearing dam treatment onthese acquisition or retention (pN0.1). Swimming distance presenteda similar pattern (data not presented). For swim velocity there was noeffect of sex, prenatal treatment or rearing dam treatment (pN0.1).
Working memoryWorking memory performance was measured as the reduction of
latency to reach the hidden platform from trial 1 to trial 2. All subjectsexhibited improvement of performance from trial 1 to trial 2, with anattenuated improvement in performance in animals reared by DEX-treated dams (Fig. 4). Additionally, males exhibited reduced latency toreach the platform compared to females (M: 30.4±1.5 and F: 35.5±1.7). Latency to reach the platform on trial 1 and 2 was analysed usinga 2×2×5×2 (prenatal treatment×rearing dam treatment×sex×days×trials) split-plot ANOVA. There was a significant main effect oftrials (F(1,56)=38.87; pb0.001), sex (F(1,56)=4.32; pb0.05) and asignificant rearing dam treatment×trials interaction F(1,56)=38.87;pb0.001). Swimming distance presented a similar pattern (data notpresented). For swim velocity there was no effect of sex, prenataltreatment or rearing dam treatment (pN0.1).
Basal and challenged HPA axis activity
Basal ACTH and corticosteroneACTH titres measured under basal conditions exhibited the
expected increased titres in the morning compared to afternoonsamples in the females, but not in the males (Fig. 5). Neither prenataltreatment nor rearing dam treatment affected basal plasma ACTHtitres. These interpretations of the results were supported by a2×2×2×2 (prenatal treatment×rearing dam treatment×sex×timeof the day) split-plot ANOVA on plasma ACTH titres. There was asignificant main effect of time of the day (F(1,56)=13.55; pb0.005)and a sex×time of the day interaction (F(1,56)=7.89; pb0.01). Thiswas further supported by a posteriori sex-split ANOVA analyses,yielding a significant effect of time in females (F(1,28)=21.02;pb0.001) but not in males (pN0.1). There were no significant mainor interaction effects of prenatal or rearing dam treatment on basalACTH plasma titres (pN0.1).
Corticosterone titres measured under basal condition exhibited forboth males and females the expected increased titres in the morningcompared to the afternoon samples (Fig. 5). Furthermore, ratsexposed prenatally to DEX exhibited a reduced afternoon sampleplasma corticosterone titre. Finally, females exhibited the expectedincreased plasma corticosterone titres compared to males. Theseinterpretations of the results were supported by a 2×2×2×2(prenatal treatment×rearing dam treatment×sex×time of the day)split-plot ANOVA on plasma corticosterone titre. There was asignificant main effect of time of the day (F(1,56)=162.84;

Fig. 3. Performance of 300-day-old offspring in the Morris water maze test using a reference memory paradigm. (A) Means (±SEM) of ln-transformed latency to reach the platformacross the 6 testing days split by prenatal and by rearing dam treatment (N=16 per treatment). There was a significant main effect of rearing dam treatment (pb0.05). (B) Meanpercentage (±SEM) of time spent in each quadrant during the 1-min probe test (target quadrant was SW) split by treatment (N=16 per treatment). The dashed line represents thechance level at 25%. Asterisks indicate a preference for target quadrant (pb0.05).
371J. Hauser et al. / Hormones and Behavior 56 (2009) 364–375
pb0.001), of sex (F(1,56)=11.09; pb0.001) and a significant prenataltreatment×time of the day interaction (F(1,56)=4.84; pb0.05). Thiswas further confirmed by a posteriori post-hoc analyses of morningand afternoon titres separately, which yielded a significant main effectof prenatal treatment in afternoon samples only (F(1,56)=4.32;pb0.05). There were no significant main or interaction effects ofrearing dam treatment on basal corticosterone plasma titres (pN0.1).
Fig. 4. Performance of 300-day-old offspring in the Morris water maze test using aworking memory paradigm. Means (±SEM) of ln-transformed latency to reachplatform for trials 1 and 2 split by treatment (N=16 per treatment). There was asignificant rearing dam treatment×trials interaction (pb0.05).
ACTH and corticosterone after restraintAll subjects responded to the stressor with a marked increase in
plasma titres of both ACTH and corticosterone. These interpretationswere supported by a 2×2×2×6 (prenatal treatment×rearing damtreatment×sex×time) split-plot ANOVA on plasma ACTH andcorticosterone titre. For both, there was a significant main effect oftime (ACTH: F(5,270)=141.94; pb0.001; corticosterone: F(5,275)=112.63; pb0.001, Fig. 6).
For ACTH, males exposed prenatally to DEX exhibited an increasedelevation in ACTH plasma titres at all time points after the restraintstress (Fig. 6). Females exhibited increased response to the stressor interms of plasma ACTH titres compared to males but reached similartitres after 2 h of recovery. These interpretations were supported by a2×2×2×6 (prenatal treatment×rearing dam treatment×sex×time)split-plot ANOVA on plasma ACTH titre (ln-transformation). Therewas a significant prenatal treatment×sex interaction (F(1,54)=4.12;pb0.05), a significant main effect of prenatal treatment (F(1,54)=6.57; pb0.05), a significant main effect of sex (F(1,54)=19.06;pb0.001) and a significant sex×time interaction (F(5,270)=4.96;pb0.005). There were no significant main or interaction effects ofrearing dam treatment (pN0.1).
For corticosterone, we observed a similar effect as for ACTH, withmales exposed prenatally to DEX exhibiting an increased stressresponse. Again, females exhibited increased plasma corticosteronetitres compared to males. These observations were supported by a2×2×2×6 (prenatal treatment×rearing dam treatment×sex×time)split-plot ANOVA on plasma corticosterone titre (ln-transformation).There was a significant main effect of prenatal treatment (F(1,55)=3.67; p=0.061) and a significant main effect of sex (F(1,55)=19.48;pb0.001), however the prenatal treatment×sex interaction failed toreach significance (F(1,55)=2.13; p=0.15). Given the main effect ofsex, we performed additional sex-split analyses, in which there was asignificant effect of prenatal treatment in males (F(1,28)=8.09;pb0.01; Fig. 6) but not in females (pN0.1), supporting the interpreta-tion of a male-specific effect of prenatal treatment. There were no
significant main or interaction effects of rearing dam treatment(pN0.1).
Discussion
In the present study, we exposed pregnant Wistar rats to DEXduring the last week of pregnancy and assessed their adult offspring inbehavioural tests of anxiety, stress coping and spatial learning andmemory, and in terms of HPA axis basal activity and stressor reactivity.Based on the reports of altered maternal behaviour following prenatalDEX exposure (Brabham et al., 2000; Hauser et al., 2006), weperformed cross-fostering at birth such that direct effects of DEX onoffspring development could be separated from maternally mediatedeffects. Indeed, exposure to inadequate maternal care is predictive ofnumerous behavioural and endocrine alterations relevant to psychia-tric diseases, and the animal evidence provides some supportiveevidence that this association is causal (Pryce et al., 2005). Offspringexposed to DEX prenatally exhibited reduced body weight from birth

Fig. 5. Morning and afternoon basal plasma ACTH and corticosterone titres (ln-transformed; means±SEM) for each sex split by prenatal treatment (N=16 perprenatal treatment). For ACTH, there was a significant sex×time of day interaction anda significant sex effect (pb0.05). For corticosterone, there was a significant prenataltreatment×time of day interaction and a significant time of day main effect (pb0.05).Asterisks represent significant difference between prenatal treatments in the afternoonsamples (pb0.05).
Fig. 6.Male and female offspring ACTH (top) and corticosterone (bottom) plasma titres(ln-transformed; mean±SEM) at time −20, 0, 20, 40, 80, 120 min relative totermination of a 20-min restraint stressor (indicated by the black horizontal bar), splitby prenatal treatment, rearing dam treatment and by sex (N=8 per group). The insetsdepict the same data split only by prenatal treatment and by sex (N=16 per prenataltreatment). For ACTH, there was a significant prenatal treatment×sex interaction and asignificant prenatal treatment main effect (pb0.05). For corticosterone, there was atrend to a main effect of prenatal treatment (p=0.061).
372 J. Hauser et al. / Hormones and Behavior 56 (2009) 364–375
to adulthood, and in males specifically, an increased reactivity of theHPA axis to a restraint stress. Interestingly, on a background ofdecreased body weight and male-specific increase of HPA axisresponsiveness, we did not observe any effects of prenatal DEXexposure on distance moved in the open field, relative time spent inthe open arms in the elevated plus maze, effect of US pre-exposure(USPE) on two way escape and avoidance behaviour, floatingbehaviour in the Porsolt forced swim test (FST), break point ratio inthe progressive ratio schedule of reinforcement (PRSR), or spatialworking and reference memory in the Morris water maze. Regardingthe indirect effect that could have been mediated by the alteredmaternal behaviour, adult offspring reared by DEX exposed damsexhibited reduced body weight from infancy onwards, impairment inworking and reference spatial memory in the Morris water maze, andfemales specifically exhibited an increased floating time in the PorsoltFST.
Direct effects of prenatal exposure to DEX
In this studywe replicated our and others' previous observations ofreduced body weight throughout infancy and in adulthood followingprenatal DEX exposure (Brabham et al., 2000; Hauser et al., 2006).Although the mediating mechanism is not yet understood, it has beenhypothesised to involve the inhibitory action of GCs on DNA synthesisin dividing cells (Sanfacon et al., 1977). However, there are also reportsof an absence of effect on adult body weight following prenatal DEXexposure (Kreider et al., 2005; Oliveira et al., 2006), albeit thesestudies used relatively short treatment (2 days). The study by Oliveiraet al. (2006) included a group with short duration and high dosage ofDEX, which exhibited a significant bodyweight decrease in adulthood.These findings suggest that the effect of prenatal DEX on adulthood
body weight is dose-duration dependant. In a parallel study in ourlaboratory using primates, we reached a similar conclusion regardingthe effect of prenatal DEX exposure on physical growth (Hauser et al.,2006).
Prenatal DEX exposure led to a male-specific increased stressresponse in terms of HPA endocrine activity, replicating a findingreported consistently in the literature (Levitt et al., 1996; Muneoka etal., 1997; Welberg et al., 2001). The effect was independent of theprenatal treatment of the rearing dam, as can only be demonstrated byusing cross-fostering. The effect of prenatal DEX exposure was notabsolutely consistent for ACTH and corticosterone response profiles:the relatively large increase in ACTH plasma titres in prenatal DEXmales was maintained across all time points after restraint, whilst therelatively large increase in corticosterone plasma titres was restrictedto the beginning of the recovery period (post-stressor 20 and 40min).However, based on the cross-fostering design, Brabham et al. (2000)report an increased HPA endocrine response to restraint stress at 0and 30 min after the end of the restraint stress in DEXO–DEXD animalsbut a decreased response at 30 min after restraint stress in DEXO–
VEHD animals. This dependency of the effect of prenatal treatment onrearing dam exposure was not observed in our study, using a similarexperimental design. This absence of interaction between the twoaforementioned treatment groups was supported by pair-wisecomparisons for the plasma ACTH and CORT levels, which failed toyield any significant effects (pN0.1). Thus, whereas in both studiesthe DEXO–DEXD treatment yielded HPA hyper-responsiveness torestraint, in our study DEXO–VEHD treatment also yielded hyper-

373J. Hauser et al. / Hormones and Behavior 56 (2009) 364–375
responsiveness, i.e. the treatment of the dam was irrelevant toprenatal DEX offspring phenotype, whereas in the Brabham et al.study being reared by dams that received VEH actually reversed theeffect of prenatal DEX exposure on the offspring's HPA responsiveness.To our knowledge, the only difference between the two studies is theuse of different rat strains: Brabham et al. studied Sprague–Dawleyrats and we studied Wistar rats. Sprague–Dawley rats have beenreported to show increased endocrine HPA activity following animmobilisation stress episode relative to Wistar rats (Belda et al.,2004). This suggests that the difference between the studies mightoriginate from strain specificity in terms of response to the prenatalDEX exposure direct effects on foetus and/or alteration of maternalbehaviour observed in dams exposed to DEX during pregnancy. Thatthe effects we observed herein are male-specific is consistent withsimilar effects observed in rats subjected to prenatal stress (Bowmanet al., 2004; McCormick et al., 1995; Weinstock et al., 1992), and inguinea-pigs following prenatal DEX exposure (Liu et al., 2001). Thissexual dichotomy might result from differential sensitivity to DEXduring prenatal development. The mechanism mediating theincreased stimulated HPA activity following prenatal DEX exposurehas been suggested to be prenatal programming of GR receptorexpression, which has been reported to be decreased in thehippocampus of offspring exposed prenatally to DEX (Welberg et al.,2001), thus decreasing the negative feedback and consequentlyincreasing the HPA response to a stressor. Brabham et al. (2000)report a decrease in hippocampal GR mRNA in the DEXO–DEXD
subjects, but an increase in the DEXO–VEHD subjects. This paralleledwith a HPA endocrine response to restraint stress (see above).
On this background of clear physiological effects of the treatment,namely reduced body weight and, in males only, increased HPA axisreactivity to a restraint stress, we observed an absence of effect ofprenatal DEX exposure on both locomotor activity in the open fieldand anxiety, as assessed in terms of central open field activity andopen-arm elevated plus maze activity. The lack of impact of prenatalDEX on these two tasks is in contrast to the reports of such treatmentleading to both reduced locomotor activity in the open field andincreased anxiety in the elevated plus maze (Oliveira et al., 2006;Welberg et al., 2001). These divergences might stem from theexperimental differences between the studies. One major differenceis that we conducted tests during the dark phase of the light/darkcycle and under dim light, whereas in the other studies tests wereconducted during the light phase and under bright light. Testingduring the light phase and under high illumination have both beenshown to decrease exploration and to increase anxiogenic behaviourin the elevated plus maze in non-manipulated rats (Bertoglio andCarobrez, 2002; Garcia et al., 2005). Therefore we hypothesise thatanxiogenic effects of prenatal DEX are visible under stressfulbackground conditions only, in this case under bright light and/orduring the light phase, due to prenatal DEX inducing hyper-sensitivityto stress. We also have to acknowledge that repeated testing may havecontributed to some of our effects, but would point out that oursubjects were naivewhen tested in the elevated plusmaze and still weobserved a discrepancy with the findings of Welberg et al. (2001).
In this study we did not observe effects of prenatal DEX onanhedonia assessed with operant responding for palatable reward ona PR schedule. We observed a similar lack of effect of prenatal DEX inthis test in a nonhuman primate, the common marmoset (Hauser etal., 2008). Stress coping, as assessed using both pre-exposure toinescapable footshock followed by monitoring escape or avoidanceresponses and the forced swim test, was also unaffected by prenatalDEX. These negative findings suggest that the existing evidence forprenatal stress as an animal model of depression (reviewed inWeinstock (2000)) is not exclusively mediated by cortisol-dependentGR activation but requires other components of the stress response.The presence of a significant effect of prenatal exposure to DEXon HPAactivity in the absence of effects of this treatment on emotional
behaviour in a range of assays suggests that the effects of prenatal DEXwere at the level of the HPA axis per se and not at the level of brainregions that regulate both HPA axis function and emotional behaviour,including the amygdala. Furthermore, it is interesting that the alteredHPA stress activity did not feed forward to lead to effects of prenatalDEX treatment in the water maze task: Cognitive behaviour in thelatter has been shown to be influenced by the activity of the HPA axis,actually in an inverted-U shaped relationship, with low stress/corticosterone enhancing water maze performance, and both highstress/corticosterone and experimental elimination of endogenouscorticosterone impairing it (Akirav et al., 2004; Diamond et al., 1992).Thus the male offspring exposed to prenatal DEX would be expectedto exhibit impaired water maze performance, as their HPA stressresponse to the water maze would be expected to be high. Possibly,repeated exposure to the water maze leads to an adaptation of HPAreactivity and a lessening of the prenatal DEX effect on increased HPAreactivity, relative to that observed in subjects exposed to a novelstressor, as was the case in the restraint experiment.
Maternally mediated indirect effects of prenatal DEX exposure
Rearing by a DEX-treated dam resulted in reduced body weightfrom infancy onwards. Interestingly, in a previous study using thesame procedure we reported similar results concomitant withincreased maternal pup-directed behaviour (Hauser et al., 2006).This surprising association has been suggested to result from animpaired ability of the dam to nourish the offspring and/or from theinability of the offspring to digest lipids (Lordache et al., 2004).
Being reared by a DEX-treated dam, regardless of prenataltreatment, led to adult offspring with impairments in both referenceand working spatial memory, as assessed in the water maze test.Specifically, offspring reared by DEX dams showed an increasedlatency to reach the platform in the referencememory paradigm and areduction in the performance improvement between trials 1 and 2 inthe working memory paradigm. Several previous studies havereported that prenatal DEX treatment leads to impairment in learningand memory in the water maze in reference memory (Brabham et al.,2000; Rayburn et al., 1997) or working memory (Emgard et al., 2007).However, only one of these studies (Brabham et al., 2000) also usedthe cross-fostering design that allows separation of direct versus dam-mediated effects. That study reports moderately impaired referencememory, limited to testing days 2 and 3, in offspring that were bothexposed to prenatal DEX and reared by a DEX dam, but a moderatelyimproved reference memory if the subjects exposed to DEX prenatallywere reared by VEH dams. The differences between findings, ofgeneral impairment in offspring reared by DEX dams in the currentstudy versus the interaction of prenatal and rearing dam treatment inthe Brabham et al. study, could originate from ages of the offspring attesting, which were 300 and 70 days, respectively. Indeed, rats exhibitimpaired performance in the water maze with aging (Gallagher andPelleymounter, 1988). One potential mechanism through whichrearing by a DEX-treated dam might impact on the performance inthe water maze is the associated increased levels of maternalbehaviour exhibited by such dams (Hauser et al., 2006). Such apotential causal relationship is supported by existing reports of alteredperformance in water maze in animals that were exposed to early-lifemanipulation impacting on maternal behaviour (Aisa et al., 2007;Lehmann et al., 1999; Pryce et al., 2003).
In both the PR schedule motivation test and the US pre-exposuretest, we did not observe any effect of rearing dam treatment or sex. Inthe FST, females reared by DEX dams demonstrated increasedimmobility. Although the FST is commonly accepted as a screeningmethod for antidepressants (predictive validity), its relevance (facevalidity) as an animal model of depression, either per se or incombination with a preceding manipulation, has been questioned(reviewed in Holmes (2003)). Given that the effect was observed in

374 J. Hauser et al. / Hormones and Behavior 56 (2009) 364–375
both pretest and test, it is possible that this reflects a trait of reducedlocomotor activity in water; however, this is unlikely given that in thevisual cue test performed in the water maze experiment, femalesreared by DEX dams did not demonstrate any behavioural differencesrelative to their vehicle dam counterparts. Another interpretation,according to the original report of Porsolt et al. (1977), is increasedbehavioural despair. Again, this is unlikely as there was no effect ofrearing dam treatment in USPE active avoidance, which is alsoproposed to assess behavioural despair. Therefore, we suggest that theincreased floating observed in females reared by DEX-treated dams isdue to an altered strategy in terms of a relatively early shift from activeto passive coping in an inescapable stressful situation. In the absenceof studies assessing the maternally mediated effects of prenatal DEXexposure or using a cross-fostering design, we have to refer to studiesusing prenatal DEX treatment and assessing performance in thePorsolt FST. Oliveira et al. (2006) also reported a lack of effect ofprenatal DEX exposure on behaviour in the FST, although studyingonly males. Welberg et al. (2001), in contrast, reported decreasedfloating time, in both pretest and test, in male offspring exposedprenatally to DEX. It is noteworthy, however, thatWelberg et al. used awater depth of 25 cm only, and the resultant potential for animals tocontact the tank floor could have produced treatment group-specificconfounding effects, i.e. the DEX-treated subjects might have beenmore likely to contact the tank floor and maintain mobility.
Sex differences
Additional to the aforementioned findings, we observed sexdifferences in several behavioural paradigms. The female ratsexhibited reduced anxiety, increased locomotor activity, increasedmotivation for palatable reward, reduced behavioural despair, andsimilar reference and working memory in the water maze, comparedto male rats. Some of these findings replicate established sexdifferences: reduced female anxiety in the elevated plus maze(Johnston and File, 1991), increased female locomotor activity inopen field (Lehmann et al., 1999) and decreased female helplessnessin the US pre-exposure using the active avoidance (Dalla et al., 2008;Steenbergen et al., 1989). However for other tasks, the presentfindings were not consistent with previous reports: motivation in theprogressive ratio schedule of reinforcement has been reported to besimilar between male and female Wistar rats (van Hest et al., 1988),and immobility in the Porsolt forced swim test has been reported to beincreased in females (Alonso et al., 1991). These discrepancies mightoriginate from protocol differences between the studies, or from theuse of specific strains. Indeed Porsolt et al. (1978) reported straindifferences in the direction of sex differences for both the basal leveland the effect of antidepressant in the forced swim task. For the watermaze, in a recent meta-analysis of the literature, Jonasson (2005)reported improvedmemory in males compared to females, but only in6 out of 14 studies, suggesting that such differences are far fromconsistent. Indeed Jonasson (2005) further reported that the strain isimportant and at least one report on Wistar rats reported similarperformance by males and females in the water maze (Blokland et al.,2006), as found in the present study.
Conclusion
We reproduced the known physiological effects of prenatal DEXexposure, namely reduction of body weight and heightened sensitiv-ity of the HPA axis to stress, although this last effect was male-specificin our study, suggesting interaction between the treatment and sex-specific developmental processes. On this background, we did notobserve evidence of a clear association between prenatal DEXexposure and behavioural despair, as assessed in Porsolt FST or USPEactive avoidance, nor did we observe effects on incentive motivation/anhedonia as assessed in a PR reinforcement schedule. This absence of
effects suggests that other components of the stress response areresponsible for the causal relationship between prenatal stress anddepression-like symptoms. Although both prenatal stress and pre-natal DEXyield an important activation of GR, prenatal stress results ina broader response involving, for example, the autonomic nervoussystem and also an increased activation of mineralocorticoid recep-tors, whereas prenatal DEX, through the GR mediated negativefeedback, results in negligible levels of corticosterone and therebymight well reduce MR activity. Finally, we observed a dam-mediatedeffect of the prenatal DEX treatment as a reduction of bodyweight andas an impairment of both reference and working memory in theMorris water maze, whichwe hypothesise to be due to the exposure toaltered maternal behaviour. In addition to these findings on the directand indirect effect of prenatal DEX exposure, we also observednumerous differences between sexes including some sex-dependenteffects of prenatal DEX exposure, which indicates the importance ofthe use of both sexes in animal studies. These findings suggest thatstudies using prenatal treatment would benefit from the use of cross-fostering, as there might be indirect effects of such treatment. Despitethe absence of depression-like behaviours in offspring exposed toprenatal DEX, the clear effect observed on HPA axis and the indirecteffect on learning and memory would tend to suggest a carefulrethinking of the routine clinical prophylactic use of repeated GCtreatment in pregnancies at risk of preterm delivery.
Acknowledgments
We are extremely grateful to Jeanne Michel and Pascal Guela foranimalmaintenance, Peter Schmid for technical assistance, Else-MariePedersen-Christensen and Corinne Späte for technical assistance,Frank Bootz for veterinary supervision, and Weining Zhang forscientific support. This research was funded by the ETH Zurich.
References
Aisa, B., Tordera, R., Lasheras, B., Del Rio, J., Ramirez, M.J., 2007. Cognitive impairmentassociated to HPA axis hyperactivity after maternal separation in rats. Psychoneur-oendocrinology 32 (3), 256–266.
Akirav, I., Kozenicky, M., Tal, D., Sandi, C., Venero, C., Richter-Levin, G., 2004. A facilitativerole for corticosterone in the acquisition of a spatial task under moderate stress.Learn. Mem. 11 (2), 188–195.
Alonso, S.J., Castellano, M.A., Afonso, D., Rodriguez, M., 1991. Sex differences inbehavioral despair: relationships between behavioral despair and open fieldactivity. Physiol. Behav. 49 (1), 69–72.
Barker, D.J., 2004. The developmental origins of well-being. Philos. Trans. R. Soc. Lond. B.Biol. Sci. 359 (1449), 1359–1366.
Belda, X., Marquez, C., Armario, A., 2004. Long-term effects of a single exposure to stressin adult rats on behavior and hypothalamic–pituitary–adrenal responsiveness:comparison of two outbred rat strains. Behav. Brain Res. 154 (2), 399–408.
Benediktsson, R., Calder, A.A., Edwards, C.R., Seckl, J.R., 1997. Placental 11 beta-hydroxysteroid dehydrogenase: a key regulator of fetal glucocorticoid exposure.Clin. Endocrinol. (Oxf). 46 (2), 161–166.
Bertoglio, L.J., Carobrez, A.P., 2002. Behavioral profile of rats submitted to session 1-session 2 in the elevated plus-maze during diurnal/nocturnal phases and underdifferent illumination conditions. Behav. Brain Res. 132 (2), 135–143.
Blokland, A., Rutten, K., Prickaerts, J., 2006. Analysis of spatial orientation strategies ofmale and femaleWistar rats in aMorris water escape task. Behav. Brain Res.171 (2),216–224.
Bowman, R.E., MacLusky, N.J., Sarmiento, Y., Frankfurt, M., Gordon, M., Luine, V.N., 2004.Sexually dimorphic effects of prenatal stress on cognition, hormonal responses, andcentral neurotransmitters. Endocrinology 145 (8), 3778–3787.
Brabham, T., Phelka, A., Zimmer, C., Nash, A., Lopez, J.F., Vazquez, D.M., 2000. Effects ofprenatal dexamethasone on spatial learning and response to stress is influenced bymaternal factors. Am. J. Physiol. Regul. Integr. Comp. Physiol. 279 (5), 899–909.
Dalla, C., Edgecomb, C., Whetstone, A.S., Shors, T.J., 2008. Females do not expresslearned helplessness like males do. Neuropsychopharmacology 33 (7), 1559–1569.
De Kloet, E.R., Vreugdenhil, E., Oitzl, M.S., Joels, M., 1998. Brain corticosteroid receptorbalance in health and disease. Endocr. Rev. 19 (3), 269–301.
Diamond, D.M., Bennett, M.C., Fleshner, M., Rose, G.M., 1992. Inverted-U relationshipbetween the level of peripheral corticosterone and the magnitude of hippocampalprimed burst potentiation. Hippocampus 2 (4), 421–430.
Emgard, M., Paradisi, M., Pirondi, S., Fernandez, M., Giardino, L., Calza, L., 2007. Prenatalglucocorticoid exposure affects learning and vulnerability of cholinergic neurons.Neurobiol. Aging 28 (1), 112–121.

375J. Hauser et al. / Hormones and Behavior 56 (2009) 364–375
Erisman, S., Carnes, M., Takahashi, L.K., Lent, S.J., 1990. The effects of stress on plasmaACTH and corticosterone in young and aging pregnant rats and their fetuses. Life.Sci. 47 (17), 1527–1533.
Estanislau, C., Morato, S., 2006. Behavior ontogeny in the elevated plus-maze: prenatalstress effects. Int. J. Dev. Neurosci. 24 (4), 255–262.
Gallagher, M., Pelleymounter, M.A.,1988. Spatial learning deficits in old rats: amodel formemory decline in the aged. Neurobiol. Aging 9 (5–6), 549–556.
Ganong, F.W., 1999. Review of Medical Physiology. Appleton and Lange, Stanford.Garcia, A.M., Cardenas, F.P., Morato, S., 2005. Effect of different illumination levels on rat
behavior in the elevated plus-maze. Physiol. Behav. 85 (3), 265–270.Hauser, J., Feldon, J., Pryce, C.R., 2006. Prenatal dexamethasone exposure, postnatal
development, and adulthood prepulse inhibition and latent inhibition in Wistarrats. Behav. Brain Res. 175 (1), 51–61.
Hauser, J., Knapman, A., Zurcher, N.R., Pilloud, S., Maier, C., Diaz-Heijtz, R., Forssberg, H.,Dettling, A., Feldon, J., Pryce, C.R., 2008. Effects of prenatal dexamethasonetreatment on physical growth, pituitary–adrenal hormones, and performance ofmotor, motivational, and cognitive tasks in juvenile and adolescent commonmarmoset monkeys. Endocrinology 149 (12), 6343–6355.
Holmes, P.V., 2003. Rodent models of depression: reexamining validity withoutanthropomorphic inference. Crit. Rev. Neurobiol. 15 (2), 143–174.
Johnston, A.L., File, S.E., 1991. Sex differences in animal tests of anxiety. Physiol. Behav.49 (2), 245–250.
Jonasson, Z., 2005. Meta-analysis of sex differences in rodent models of learning andmemory: a review of behavioral and biological data. Neurosci. Biobehav. Rev. 28 (8),811–825.
Kreider, M.L., Levin, E.D., Seidler, F.J., Slotkin, T.A., 2005. Gestational dexamethasonetreatment elicits sex-dependent alterations in locomotor activity, reward-basedmemory and hippocampal cholinergic function in adolescent and adult rats.Neuropsychopharmacology 9 (30), 1617–1623.
Lehmann, J., Feldon, J., 2000. Long-term biobehavioral effects of maternal separation inthe rat: consistent or confusing? Rev. Neurosci. 11 (4), 383–408.
Lehmann, J., Pryce, C.R., Bettschen, D., Feldon, J., 1999. The maternal separationparadigm and adult emotionality and cognition in male and female Wistar rats.Pharmacol. Biochem. Behav. 64 (4), 705–715.
Leng, A., Jongen-Relo, A.L., Pothuizen, H.H., Feldon, J., 2005. Effects of prenatalmethylazoxymethanol acetate (MAM) treatment in rats on water maze perfor-mance. Behav. Brain. Res. 161 (2), 291–298.
Levitt, N.S., Lindsay, R.S., Holmes, M.C., Seckl, J.R., 1996. Dexamethasone in the last weekof pregnancy attenuates hippocampal glucocorticoid receptor gene expression andelevates blood pressure in the adult offspring in the rat. Neuroendocrinology 64 (6),412–418.
Liggins, G.C., Howie, R.N., 1972. A controlled trial of antepartum glucocorticoidtreatment for prevention of the respiratory distress syndrome in prematureinfants. Pediatrics 50 (4), 515–525.
Liu, L., Li, A., Matthews, S.G., 2001. Maternal glucocorticoid treatment programs HPAregulation in adult offspring: sex-specific effects. Am. J. Physiol. Endocrinol. Metab.280 (5), E729–E739.
Lordache, C., Drozdowski, L., Clandinin, T., Wild, G., Todd, Z., Thomson, A.B., 2004.Dexamethasone plus glucagon-like peptide 2 given to lactating rat dams has a lateeffect on intestinal lipid uptake in the weanling offspring. JPEN. J. Parenter. Enteral.Nutr. 28 (5), 355–363.
McCormick, C.M., Smythe, J.W., Sharma, S., Meaney, M.J., 1995. Sex-specific effects ofprenatal stress on hypothalamic–pituitary–adrenal responses to stress and brainglucocorticoid receptor density in adult rats. Brain Res. Dev. Brain Res. 84 (1), 55–61.
Muneoka, K., Mikuni, M., Ogawa, T., Kitera, K., Kamei, K., Takigawa, M., Takahashi, K.,1997. Prenatal dexamethasone exposure alters brain monoamine metabolism andadrenocortical response in rat offspring. Am. J. Physiol. 273 (5 Pt. 2), 669–675.
Nathanielsz, P.W., Padmanabhan, V., 2006. Developmental origin of health and disease.J. Physiol. 572 (Pt. 1), 3–4.
National Institutes of Health Consensus Development Panel, 1995. Consensus develop-ment panel on the effect of corticosteroids for fetal maturation on perinataloutcomes: effect of corticosteroids for fetal maturation on perinatal outcomes.JAMA 273 (5), 413–418.
National Institutes of Health Consensus Development Panel, 2001. Consensus devel-opment conference statement: antenatal corticosteroids revisited: repeat courses.Obstet. Gynecol. 98 (1), 144–150.
Oliveira, M., Bessa, J.M., Mesquita, A., Tavares, H., Carvalho, A., Silva, R., Pego, J.M.,Cerqueira, J.J., Palha, J.A., Almeida, O.F., Sousa, N., 2006. Induction of a hyperanxiousstate by antenatal dexamethasone: a case for less detrimental natural corticoster-oids. Biol. Psychiatry 59 (9), 844–852.
Patin, V., Lordi, B., Vincent, A., Caston, J., 2005. Effects of prenatal stress on anxiety andsocial interactions in adult rats. Brain Res. Dev. Brain Res. 160 (2), 265–274.
Porsolt, R.D., Le Pichon, M., Jalfre, M., 1977. Depression: a new animal model sensitive toantidepressant treatments. Nature 266 (5604), 730–732.
Porsolt, R.D., Bertin, A., Jalfre, M., 1978. “Behavioural despair” in rats and mice: straindifferences and the effects of imipramine. Eur. J. Pharmacol. 51 (3), 291–294.
Pryce, C.R., Palme, R., Feldon, J., 2002. Development of pituitary–adrenal endocrinefunction in the marmoset monkey: infant hypercortisolism is the norm. J. Clin.Endocrinol. Metab. 87 (2), 691–699.
Pryce, C.R., Bettschen, D., Nanz-Bahr, N.I., Feldon, J., 2003. Comparison of the effects ofearly handling and early deprivation on conditioned stimulus, context, and spatiallearning and memory in adult rats. Behav. Neurosci. 117 (5), 883–893.
Pryce, C.R., Ruedi-Bettschen, D., Dettling, A.C., Weston, A., Russig, H., Ferger, B., Feldon, J.,2005. Long-term effects of early-life environmental manipulations in rodents andprimates: potential animal models in depression research. Neurosci. Biobehav. Rev.29 (4–5), 649–674.
Rayburn, W.F., Christensen, H.D., Gonzalez, C.L., 1997. A placebo-controlled comparisonbetween betamethasone and dexamethasone for fetal maturation: differences inneurobehavioral development of mice offspring. Am. J. Obstet. Gynecol. 176 (4),842–850 discussion 850–1.
Sanfacon, R., Possmayer, F., Harding, P.G., 1977. Dexamethasone treatment of the guineapig fetus: its effects on the incorporation of 3H-thymidine into deoxyribonucleicacid. Am. J. Obstet. Gynecol. 127 (7), 745–752.
Seckl, J.R., 2004. Prenatal glucocorticoids and long-term programming. Eur. J.Endocrinol. 151 (Suppl. 3), 49–62.
Smith, J.W., Seckl, J.R., Evans, A.T., Costall, B., Smythe, J.W., 2004. Gestational stressinduces post-partum depression-like behaviour and alters maternal care in rats.Psychoneuroendocrinology 29 (2), 227–244.
Steenbergen, H.L., Heinsbroek, R.P., Van Haaren, F., Van de Poll, N.E., 1989. Sex-dependent effects of inescapable shock administration on behavior and subsequentescape performance in rats. Physiol. Behav. 45 (4), 781–787.
van Hest, A., van Haaren, F., van de Poll, N.E., 1988. The behavior of male and femaleWistar rats pressing a lever for food is not affected by sex differences in foodmotivation. Behav. Brain Res. 27 (3), 215–221.
Weinstock, M., 2000. Behavioral and neurohormonal sequelae of prenatal stress: asuggested model of depression. Contemporary Issues in Modeling Psychopathol-ogy. Kluwer Academic Publishers, Boston, pp. 45–54.
Weinstock,M., 2005. The potential influence ofmaternal stress hormones ondevelopmentand mental health of the offspring. Brain Behav. Immun. 19 (4), 296–308.
Weinstock, M., Matlina, E., Maor, G.I., Rosen, H., McEwen, B.S., 1992. Prenatal stressselectively alters the reactivity of the hypothalamic–pituitary adrenal system in thefemale rat. Brain Res. 595 (2), 195–200.
Welberg, L.A., Seckl, J.R., Holmes, M.C., 2001. Prenatal glucocorticoid programming ofbrain corticosteroid receptors and corticotrophin-releasing hormone: possibleimplications for behaviour. Neuroscience 104 (1), 71–79.
Zagron, G., Weinstock, M., 2006. Maternal adrenal hormone secretion mediatesbehavioural alterations induced by prenatal stress in male and female rats. Behav.Brain Res. 175 (2), 323–328.




















