
Differential expression of heat-shock proteins and spontaneous synthesis of HSP70 during the life...
10
Eur. J. Biochem. 163,211 -220 (1987) 0 FEBS 1987 Differential expression of heat-shock proteins and spontaneous synthesis of HSP70 during the life cycle of Btastocladiella emersonii M. Christina M. BONATO, Aline M. SILVA, Suely L. GOMES, Jost Carlos C. MAIA and Maria Helena JULIANI Departamento de Bioquimica, Instituto de Quimica, Universidade de Siio Paulo (Received July 23/0ctober 27, 1986) - EJB 86 0790 The heat-shock response in Blustocludiellu emrrsonii is dependent on the developmental stage. Cells exposed to elevated temperatures at different stages of the life cycle (sporulation, germination or growth) show a differential synthesis of heat-shock proteins (hsps). Of a total of 22 polypeptides induced, particular subsets of hsps appear in each phase, demonstrating a non-coordinate heat-shock gene expression. In contrast, heat-shock-related proteins (hsp76, hsp70, hsp39a) are spontaneously expressed at a high level during sporulation. By the criteria of two-dimensional gel electrophoresis and partial proteolysis mapping, the 70000-Da protein, whose synthesis is induced spontaneously during sporulation, is indistinguishable from the heat-inducible hsp70. The techniques of in vitro translation, and Northern analysis using a Drosophilu hsp70 probe, demonstrated that enhanced synthesis of hsp70, which occurs during heat-shock treatment and spontaneously during sporulation, is associated with an accumulation of hsp70 mRNA. These observations suggest that hsp70 gene expression is induced during sporulation. The heat-shock response is a typical physiological reaction qf prokaryotic and eukaryotic cells, induced by heat shock and other environmental stresses [I, 21. Certain genes, which are essentially quiescent at normal temperatures, are tran- siently transcribed and a new set of proteins (heat-shock proteins, hsps) is synthesized; frequently there is an ac- companying interruption of normal protein synthesis. Although the pattern of hsps depends on the organism, the cell type, the temperature and the duration of exposure, both the induction mechanism at the transcriptional level and the structure of some of the proteins show a striking degree of conservation. Thus the Drosophilu hsp70 gene hybridizes to genomic sequences from many different organisms [3, 41 and can be induced concomitantly with the heat-shock response in different eukaryotes; heat-shock proteins from a series of eukaryotic organisms are antigenically related [5] and a consensus sequence element was identified in the promotor region of different hsp genes [6, 71. In addition to a tran- scriptional control, selective translation of the heat-shock mRNAs has been demonstrated in some systems [8 - 101. The function of the hsps remains unknown, although the heat-shock response is frequently viewed as a cellular survival function conferring protection against the momentary and subsequent stress to the cell. Besides a role in the response to stress, these proteins may play a role in normal development. This hypothesis is based on the observation that some of the hsps are expressed during oogenesis and pupation in Drosophilu or during the two-cell stage in mouse embryo [II, 121. To understand better the possible role of hsps in differentiation it is important to investigate other develop- mental systems. An ideal model system is that of the aquatic Correspondence to M. H. Juliani, Departamento de Bioquimica, Instituto de Quimica, Universidade de SBo Paulo, Caixa Postal 20780, BR-01498, SZo Paulo, Brazil Abbreviations. SDS, sodium dodecyl sulfate; hsp, heat-shock pro- tein. fungus Blustocludiellu emersonii. This organism undergoes two distinct phases of cell differentiation: germination and sporulation, both of which are rapid and can be induced with a high degree of synchrony. B. emersonii grows as a coenocytic cell. Nutritional starva- tion at any time during its growth induces the sporulation process. This reflects a drastic change in the gene expression program and changes in both translatable RNA species and protein synthesis [13]. This process is synchronous and results in the discharge of highly differentiated cells, the zoospores, after 4 h of starvation. The motile zoospores are quiescent with respect to both protein and RNA synthesis [14]. When placed in an appropriate solution (an inorganic germination solution containing K + as an inducer or a nutrient medium) the zoospores rapidly and synchronously retract their flagella, round up and convert into a germling in a germination process requiring less than one hour. Most of the germination se- quence does not require concomitant transcription, and a great part of the abrupt increase in the rate of protein synthesis is the result of translation of the mRNA transcribed during sporulation and stored in the zoospores [15 - 171. After germination the cells can resume growth if adequate nutritional conditions exist. In the present report we identified the Blustocludiellu heat- shock proteins induced during growth and at different stages of sporulation and germination. A non-coordinate expression of these proteins occurred during the developmental cycle; particular subsets of hsps being heat-induced in each phase. In addition, at physiological temperatures, sporulating cells synthesized high levels of a 70-kDa protein, which is similar, if not identical (by isoelectric focusing, molecular mass and partial proteolytic peptide mapping), to the major heat-shock protein, hsp70, of Blustocludiellu. Furthermore, hybridization experiments using the Drosophilu hsp70 gene and in vitro trans- lation studies confirmed a period of hsp70 gene activity during sporulation.
-
Upload
fio-educacional -
Category
Documents
-
view
1 -
download
0
Transcript of Differential expression of heat-shock proteins and spontaneous synthesis of HSP70 during the life...

Eur. J. Biochem. 163,211 -220 (1987) 0 FEBS 1987
Differential expression of heat-shock proteins and spontaneous synthesis of HSP70 during the life cycle of Btastocladiella emersonii M. Christina M. BONATO, Aline M. SILVA, Suely L. GOMES, Jost Carlos C. MAIA and Maria Helena JULIANI
Departamento de Bioquimica, Instituto de Quimica, Universidade de Siio Paulo
(Received July 23/0ctober 27, 1986) - EJB 86 0790
The heat-shock response in Blustocludiellu emrrsonii is dependent on the developmental stage. Cells exposed to elevated temperatures at different stages of the life cycle (sporulation, germination or growth) show a differential synthesis of heat-shock proteins (hsps). Of a total of 22 polypeptides induced, particular subsets of hsps appear in each phase, demonstrating a non-coordinate heat-shock gene expression. In contrast, heat-shock-related proteins (hsp76, hsp70, hsp39a) are spontaneously expressed at a high level during sporulation. By the criteria of two-dimensional gel electrophoresis and partial proteolysis mapping, the 70000-Da protein, whose synthesis is induced spontaneously during sporulation, is indistinguishable from the heat-inducible hsp70. The techniques of in vitro translation, and Northern analysis using a Drosophilu hsp70 probe, demonstrated that enhanced synthesis of hsp70, which occurs during heat-shock treatment and spontaneously during sporulation, is associated with an accumulation of hsp70 mRNA. These observations suggest that hsp70 gene expression is induced during sporulation.
The heat-shock response is a typical physiological reaction qf prokaryotic and eukaryotic cells, induced by heat shock and other environmental stresses [I, 21. Certain genes, which are essentially quiescent at normal temperatures, are tran- siently transcribed and a new set of proteins (heat-shock proteins, hsps) is synthesized; frequently there is an ac- companying interruption of normal protein synthesis. Although the pattern of hsps depends on the organism, the cell type, the temperature and the duration of exposure, both the induction mechanism at the transcriptional level and the structure of some of the proteins show a striking degree of conservation. Thus the Drosophilu hsp70 gene hybridizes to genomic sequences from many different organisms [3, 41 and can be induced concomitantly with the heat-shock response in different eukaryotes; heat-shock proteins from a series of eukaryotic organisms are antigenically related [5] and a consensus sequence element was identified in the promotor region of different hsp genes [6, 71. In addition to a tran- scriptional control, selective translation of the heat-shock mRNAs has been demonstrated in some systems [8 - 101.
The function of the hsps remains unknown, although the heat-shock response is frequently viewed as a cellular survival function conferring protection against the momentary and subsequent stress to the cell. Besides a role in the response to stress, these proteins may play a role in normal development. This hypothesis is based on the observation that some of the hsps are expressed during oogenesis and pupation in Drosophilu or during the two-cell stage in mouse embryo [II , 121. To understand better the possible role of hsps in differentiation it is important to investigate other develop- mental systems. An ideal model system is that of the aquatic
Correspondence to M. H . Juliani, Departamento de Bioquimica, Instituto de Quimica, Universidade de SBo Paulo, Caixa Postal 20780, BR-01498, SZo Paulo, Brazil
Abbreviations. SDS, sodium dodecyl sulfate; hsp, heat-shock pro- tein.
fungus Blustocludiellu emersonii. This organism undergoes two distinct phases of cell differentiation: germination and sporulation, both of which are rapid and can be induced with a high degree of synchrony.
B. emersonii grows as a coenocytic cell. Nutritional starva- tion at any time during its growth induces the sporulation process. This reflects a drastic change in the gene expression program and changes in both translatable RNA species and protein synthesis [13]. This process is synchronous and results in the discharge of highly differentiated cells, the zoospores, after 4 h of starvation. The motile zoospores are quiescent with respect to both protein and RNA synthesis [14]. When placed in an appropriate solution (an inorganic germination solution containing K + as an inducer or a nutrient medium) the zoospores rapidly and synchronously retract their flagella, round up and convert into a germling in a germination process requiring less than one hour. Most of the germination se- quence does not require concomitant transcription, and a great part of the abrupt increase in the rate of protein synthesis is the result of translation of the mRNA transcribed during sporulation and stored in the zoospores [15 - 171. After germination the cells can resume growth if adequate nutritional conditions exist.
In the present report we identified the Blustocludiellu heat- shock proteins induced during growth and at different stages of sporulation and germination. A non-coordinate expression of these proteins occurred during the developmental cycle; particular subsets of hsps being heat-induced in each phase. In addition, at physiological temperatures, sporulating cells synthesized high levels of a 70-kDa protein, which is similar, if not identical (by isoelectric focusing, molecular mass and partial proteolytic peptide mapping), to the major heat-shock protein, hsp70, of Blustocludiellu. Furthermore, hybridization experiments using the Drosophilu hsp70 gene and in vitro trans- lation studies confirmed a period of hsp70 gene activity during sporulation.
21 2
MATERIALS AND METHODS
Culture and labeling conditions
Cultures of Blastocladiella emersonii were maintained on plates containing 0.13% peptone, 0.13% yeast extract, 0.3% glucose and 1 YO agar. Zoospores (3 x 10S/ml) were inoculated into DM3 medium [18] followed by growth for 13 h at 19°C. Vegetative cells were then starved by filtering through a Nytex cloth, rinsing and resuspending in sporulation solution (1 mM Tris/maleate buffer, pH 6.8, 1 mM CaC12) at the density of 5 x lo5 cells/ml. Zoospore germination was induced in germination solution (1 mM Tris/maleate buffer, pH 6.8, 1 mM CaC12, 10mM MgC12, 50mM KCl) at 1x106 zoospores/ml. Growth was performed in AL medium (50 mM KCl, 10 mM MgC12, 1 mM CaC12, 0.3% glucose, 40 ng/ml thiamine, 1 mM Tris/maleate, pH 6.8, 0.6 pg/ml FeS04, 0.1 pg/ml CuSO,, 0.2 pg/ml ZnS04, 0.2 pg/ml MnSO,, and the common 18 amino acids present at 0.2 mM each, except for methionine at 5 pM) by inoculation of zoospores at the density of 1 x lo6 cells/ml. Cells were incubated with agitation at 27°C or the heat-shock temperature. Aliquots containing 6 x lo6 zoosporangia, 4 x lo7 germinating cells or 2 x lo7 growing cells were labeled with [3sS]methionine (1 -2 pCi/ ml) in the last 10-20 min incubation at the indicated times and temperature.
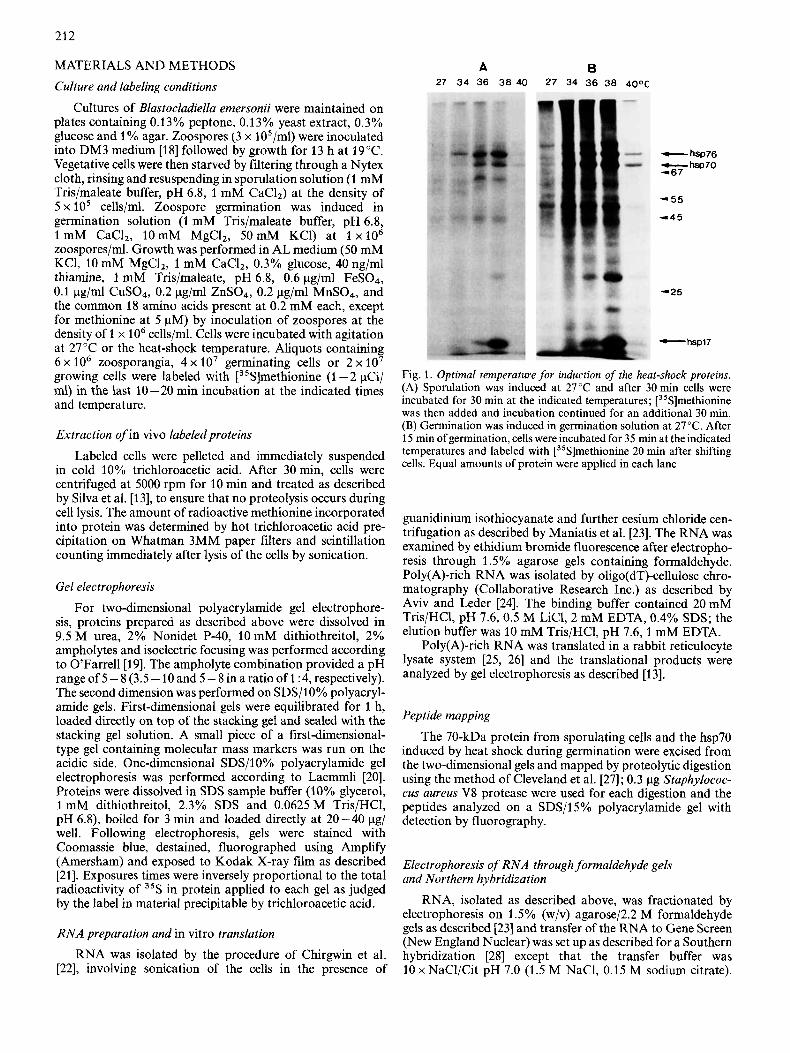
A B 27 3 4 36 38 40 27 34 36 38 40oC
Extraction of in vivo labeled proteins
Labeled cells were pelleted and immediately suspended in cold 10% trichloroacetic acid. After 30 min, cells were centrifuged at 5000 rpm for 10 min and treated as described by Silva et al. [13], to ensure that no proteolysis occurs during cell lysis. The amount of radioactive methionine incorporated into protein was determined by hot trichloroacetic acid pre- cipitation on Whatman 3MM paper filters and scintillation counting immediately after lysis of the cells by sonication.
Gel electrophoresis
For two-dimensional polyacrylamide gel electrophore- sis, proteins prepared as described above were dissolved in 9.5 M urea, 2% Nonidet P-40, 10 mM dithiothreitol, 2% ampholytes and isoelectric focusing was performed according to O’Farrell[19]. The ampholyte combination provided a pH range of 5 - 8 (3.5 - 10 and 5 - 8 in a ratio of 1 : 4, respectively). The second dimension was performed on SDS/lO% polyacryl- amide gels. First-dimensional gels were equilibrated for 1 h, loaded directly on top of the stacking gel and sealed with the stacking gel solution. A small piece of a first-dimensional- type gel containing molecular mass markers was run on the acidic side. One-dimensional SDS/IO% polyacrylamide gel electrophoresis was performed according to Laemmli [20]. Proteins were dissolved in SDS sample buffer (1070 glycerol, 1 mM dithiothreitol, 2.3% SDS and 0.0625 M Tris/HCI, pH 6 4 , boiled for 3 min and loaded directly at 20-40 pg/ well. Following electrophoresis, gels were stained with Coomassie blue, destained, fluorographed using Amplify (Amersham) and exposed to Kodak X-ray film as described [21]. Exposures times were inversely proportional to the total radioactivity of 3sS in protein applied to each gel as judged by the label in material precipitable by trichloroacetic acid.
RNA preparation and in vitro translation
RNA was isolated by the procedure of Chirgwin et al. [22], involving sonication of the cells in the presence of
- hsp76 --hsp70 -67
- 55
-45
-25
C- hsp17
Fig. 1. Optimal temperature for induction of the heat-shock proteins. (A) Sporulation was induced at 27°C and after 30 min cells were incubated for 30 min at the indicated temperatures; [35S]methionine was then added and incubation continued for an additional 30 min. (B) Germination was induced in germination solution at 27°C. After 15 min of germination, cells were incubated for 35 min at the indicated temperatures and labeled with [35S]methionine 20 min after shifting cells. Equal amounts of protein were applied in each lane
guanidinium isothiocyanate and further cesium chloride cen- trifugation as described by Maniatis et al. [23]. The RNA was examined by ethidium bromide fluorescence after electropho- resis through 1.5% agarose gels containing formaldehyde. Poly(A)-rich RNA was isolated by oligo(dT)-cellulose chro- matography (Collaborative Research Inc.) as described by Aviv and Leder [24]. The binding buffer contained 20 mM Tris/HCl, pH 7.6,0.5 M LiC1, 2 mM EDTA, 0.4% SDS; the elution buffer was 10 mM Tris/HCl, pH 7.6, 1 mM EDTA.
Poly(A)-rich RNA was translated in a rabbit reticulocyte lysate system [25, 261 and the translational products were analyzed by gel electrophoresis as described [13].
Peptide mapping
The 70-kDa protein from sporulating cells and the hsp70 induced by heat shock during germination were excised from the two-dimensional gels and mapped by proteolytic digestion using the method of Cleveland et al. [27]; 0.3 pg Staphylococ- cus aureus V8 protease were used for each digestion and the peptides analyzed on a SDS/15Yo polyacrylamide gel with detection by fluorography.
Electrophoresis of RNA through formaldehyde gels and Northern hybridization
RNA, isolated as described above, was fractionated by electrophoresis on 1.5% (w/v) agarose/2.2 M formaldehyde gels as described [23] and transfer of the RNA to Gene Screen (New England Nuclear) was set up as described for a Southern hybridization [28] except that the transfer buffer was 10 x NaCl/Cit pH 7.0 (1.5 M NaCl, 0.15 M sodium citrate).

21 3
Table 1. Blastocladiella heat-shock proteins identified by two-dimensional gel electrophoresis
Ih 6h 4 h t I I
zoospore round cell qermlinq syncytiurn sporanqiurn \Tp 15-20min 40-60min
9 - O - q - hSP PH Germination Growth Sporulation
early late
kDa 84 82a 82b 76 70 60 50 45a 45b 4% 39a 39b 39c 39d 30a 30b 30c 30d 25 24 17a 17b
6.5-7.0 5.0-5.5 5.5 - 6.0 5.0-5.6" 5.2-5.7 5.5-6.5 6.3-6.8 6.1 -6.6 6.5-7.0 6.7-7.2 5.2-5.7 7.0-7.5 7.2-7.7 7.5-8.0 5.0-5.5 7.0 - 7.5 7.5-8.0 > 8.0 5.3 - 5.8 7.5-8.0 5.8-6.3 6.7-8.2
+ + + + + + + - - -
+ -
- -
+ + + + + + + +
+ + + + + + + + + + + + +
-
-
- -
-
+ + + -
a During sporulation hsp76 is at a pH posisition more acidic than hsp70, but during germination it is more basic. 2 - 5 polypeptides together in this molecular mass range.
A 3 x 103-base BamHI restriction fragment of the Drosophila hsp70 gene, inserted into pBR322 (plasmid B8; a gift of Dr E. Craig), was nick-translated [23] and used as a probe (specific activity > lo7 cpm/pg). The filter was first prehybridized with 6 x NaCl/Cit, 5 x Denhardt's solution (1 mg/ml bovine serum albumin, 1 mg/ml Ficoll and 1 mg/ml polyvinylpyrollidone), 0.5% SDS, 100 pg/ml calf thymus DNA, 0.01 M EDTA at 60°C for 2 h. This buffer was then replaced with another portion of the same buffer containing the denatured 32P- labeled probe DNA and incubation was continued for at least another 16 h at 60°C. The filter was washed with four changes of 4 x NaCl/Cit, 0.1 YO SDS at 60 "C for a total of 2 h. The filter was allowed to dry at room temperature and then exposed to X-ray film using a Cronex intensifying screen.
Immunoprecipitation assay Immunoprecipitation of cell extracts was carried out as
described [29] with modifications. Labeled cells were lysed by sonication in wash buffer (0.05 M Tris/HCl, pH 8.3, 0.45 M NaCl, 0.5% Triton X-100) containing 0.5 mM puromycin, 50 pg/ml antipain and 1 mM phenylmethylsulfonyl fluoride. The antiserum against Escherichia coli dnaK protein was a gift of Dr C. P. Georgopoulos.
RESULTS
Heat-shock proteins are strongly induced at 38 "C in sporangium and germling cells without a decline in the overall protein synthesis
The fluorogram in Fig. 1 shows the electrophoretic dis- tribution of total zoosporangium and germling cell proteins labeled with [35S]methionine after heat shock at various temperatures. In both the sporulation and germination pro- cesses a new set of proteins (heat-shock proteins, hsps) in the 70 - 84-kDa and 45 - 60-kDa range is induced when cultures are shifted from 27 "C to 34 - 38 "C. Methionine incorporation in hsps gradually increases with temperature up to 38"C, followed by an abrupt decline in both hsp and normal proteins at 40°C. Apparently at 40°C the efficiencies of normal and heat-shock protein expression are affected to the same extent. Small hsps (17-30 kDa) have a much narrower induction range with maximal incorporation at 38 "C and very low incor- poration immediately above or below this temperature. The maximum level of hsps synthesis at 38 "C is not accompanied by a shut-down in the overall protein synthesis. In Blasto- cladiella at 38 "C, translation of normal messages is apparently concomitant with increased heat-shock mRNA translation. Three prominent proteins (hsp76, hsp70 and hspl7) are ob-
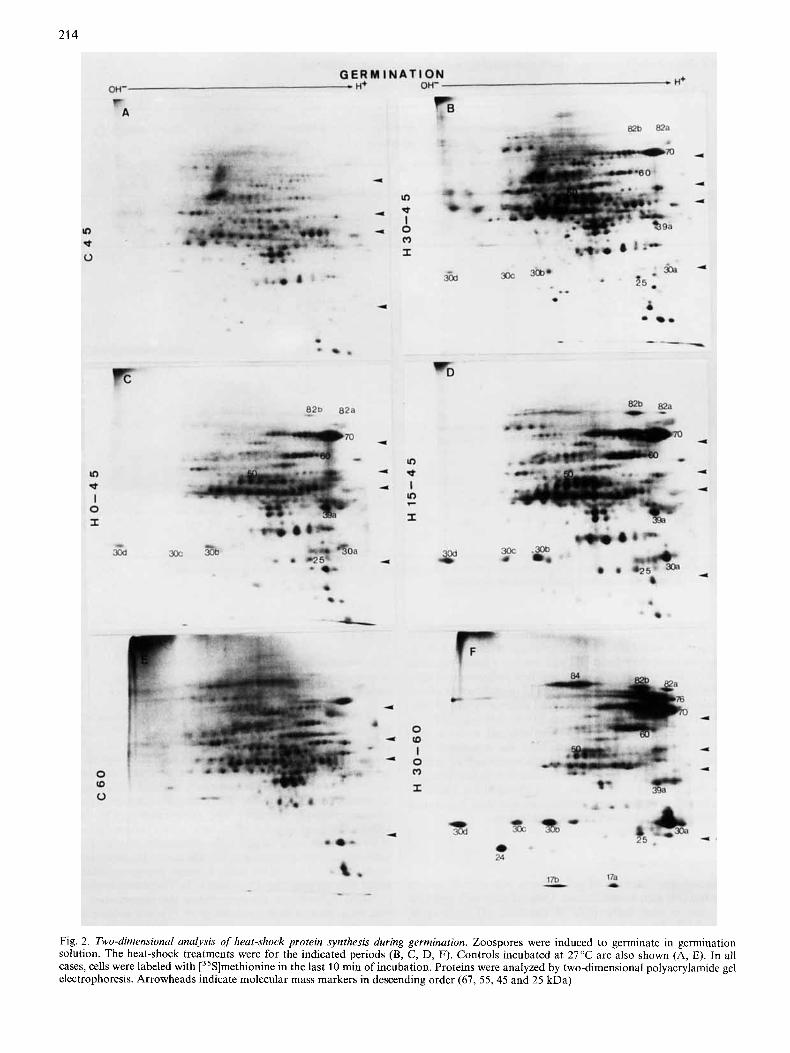
214
Fig. 2. Two-dimensional analysis of heat-shock protein synthesis during germination. Zoospores were induced to germinate in germination solution. The heat-shock treatments were for the indicated periods (B, C, D, F). Controis incubated at 27°C are also shown (A, E). In all cases, cells were labeled with [35S]methionine in the last 10 min of incubation. Proteins were analyzed by two-dimensional polyacrylamide gel electrophoresis. Arrowheads indicate molecular mass markers in descending order (67, 55,45 and 25 kDa)

215
Fig. 3. Two-dimensional gel electrophoresis of proteins synthesized during heat shock in growing cells. Zoospores were induced to germinate in AL medium at 27°C. After 2 h of growth, aliquots of growing cells were shifted to 38°C and heat-shocked for 30 min (B). Control cells were kept at 27°C (A). Cells were labeled in the last 10 rnin of incubation
served in the fluorogram of both sporulating and germinating cells. Several other proteins are clearly induced, but at lower levels. By microscopic examination at different times we ob- served that the germination, including germ tube formation, was not affected by the hyperthermia up to 38°C. However, at 40°C the zoospores encysted, but germ tube formation was inhibited. During sporulation, a shift to higher temperature (38 "C or 40 "C) caused a certain delay in the process.
Heat-shock proteins are differentially expressed during germination, sporulation or growth
A more detailed analysis of the heat-shock proteins, in- duced following a shift to 38"C, was performed using two- dimensional gel electrophoresis. The heat-shock response was examined during growth and at various stages of sporulation and germination. Within the limitations of resolution offered by isoelectric focusing gel electrophoresis and size separation on polyacrylamide/SDS gels, at least 22 polypeptides could be identified in the overall life cycle. Table 1 provides a cata- logue of differential expression of heat-shock proteins related to the developmental stage. Though some polypeptides occur in all developmental stages, others are characteristic of one specific phase of the life cycle. hsp70 is the major heat-shock protein in Blastocladiella. hsp82a, hsp82b, hsp76, hsp70, hsp60, hsp25 and hspl7b are common to germination, sporulation and growth. Some hsps are not induced during the germination process (45a, 45b, 45c, 39b, 39c, 39d), whereas hsp24 is typical of germination and hsp39b, hsp39c and hsp39d are typical of sporulation.
Hsps during zoospore germination. Zoospores germinating at 27 "C in inorganic germination solution were shifted to 38 "C after 15 min or 30 min of incubation at the physiological temperature and exposure to heat shock was, respectively, for 30 rnin and 15 min, i.e. cells were collected 45 rnin after
induction of germination. At this time (45 min) the morpho- logical transition from zoospore to germling was complete. At least 11 polypeptides are induced in both cases: hsp82a, hsp82b, hsp70, hsp60, hsp50, hsp39a, hsp30a, hsp30b, hsp30c, hsp30d, hsp25 (Fig. 2B, D). When zoospores are induced to germinate directly at 38°C and collected after 45 min, the same set of hsps is also found (Fig. 2C). In all cases [35S]methionine was present during the last 10 min (35- 45min) of incubation at the heat-shock temperature. No decrease in the total amount of methionine incorporation into protein, determined by hot trichloroacetic acid precipitation on filter-paper and scintillation counting, was observed (data not shown); incorporation at 38 "C typically being greater than at 27°C. In all these situations, germination, including germ tube formation, occurred at 38°C even when the heat shock was initiated at time zero of germination. When heat- shocked (30 - 60 min) germinating cells were labeled at late germination (50 - 60 min) a new additional set of heat-shock proteins is also induced: hsp84, hsp76, hsp24, hspl7a and hspl7b (Fig. 2F and Table 1). These proteins are not induced prior to 45 min of germination, irrespective of the time and duration of heat treatment. They appear only at late germina- tion in cells heat-shocked early during germination, suggesting a developmental control superimposed upon the expression of these proteins.
hsps during growth. Growing cells were shifted from 27 "C to 38°C at 120 min of growth, heat-shocked for 30 rnin or 60 rnin and pulse-labeled with [3sS]methionine in the last 10 rnin of incubation at 38°C. Fourteen heat-shocked polypeptides (82a, 82b, 76, 70, 60, 50,45a, 45b, 45c, 39a, 30b, 30d, 25 and 17b) were detected in the fluorograms, after either 30 min or 60 min of heat treatment, as listed in Table 1, though the heat response was more evident on short heat treatment (30 min) (Fig. 3). hsp70 is the major heat-shock protein and, hsp82b is vigorously induced; several heat shock

216
Fig. 4. Patterns of protein synthesis in normal and heat-shocked sporulating cells. Starvation-induced sporulation was at 27 “C. During the indicated periods aliquots of sporulating cells were heat-shocked at 38°C (B, D, F). Controls incubated at 27°C are also shown (A, C, E). Cells were labeled with [35S]methionine in the last 20 min of incubation. Fluorograms of two-dimensional gels are shown

21 7
A B 1 2 3 1 2 3 4 5 6 7
25 S
1 8 s
Fig. 5. Comparison between the peptide maps of hsp70 from heat- shocked germinating cells and the corresponding protein from 2-h sporulating cells at normal temperature. Starvation-induced sporu- lating cells at 27°C were pulse-labeled with [35S]methionine at 90- 120 rnin (A). During germination, cells were maintained at 38°C from 15 rnin to 45 rnin and labeled with [35S]methionine in the last 30 rnin of incubation (B). Spots corresponding to hsp70 were excised from two-dimensional gels and then mapped on a 15% polyacrylamide gel using 0.3 pg V8 protease as described by Cleveland et al. [27]. Molec- ular mass markers are indicated to the right of the fluorogram of the gel
proteins, such as hsp76, hsp30b, hsp30d or hspl7b, are faintly labeled when compared with the induction obtained in the other developmental stages. Only a slight decrease in the syn- thesis of normal proteins is observed under heat shock. During growth at physiological temperature, two proteins with molecular masses and pH ranges similar to hsp82b and hsp70 are synthesized at basal levels.
hsps during sporulation. Cells were heat-shocked at 38 "C for 30 rnin or 60 rnin at various stages during starvation- induced sporulation. Pulse-labeling with [3sS]methionine was during the last 20 min of incubation. At least 16 polypeptides are induced (84, 82a, 82b, 76, 70, 60, 45a, 45b, 45c, 39b, 39c, 39d, 30a, 25, 17a, 17b) in the first 30min of heat shock (Fig. 4B, D and F). If the cells are exposed to high temperatures for longer periods (60 rnin), the synthesis of several heat-shock proteins decreases (hsp84, hsp39b, hsp39c, hsp39d) and a concomitant increase in the methionine incor- poration at normal proteins is observed (not shown), in- dicating that the changes in the protein pattern induced by heat shock are transient.
Comparison of the pattern of proteins synthesized at various stages of sporulation at physiological temperatures
Fig. 6. Hybridization of a Drosophila hsp7O gene probe to RNA isolated from control and heat-shocked cells. RNA was resolved by electrophoresis on agarose gels, transferred to Gene Screen filter and then hybridized to the 32P-labeled nick-translated probe (plasmid B8 containing the Drosophila hsp70 gene). (A) Poly(A)-rich RNA isolated from 1-h (lane l), 2-h (lane 2) and 3-h (lane 3) sporulating cells at 27°C. (B) Total RNA isolated from sporulating cells heat-shocked at 38 "C from 30 min to 60 min (lane 3), 60 min to 90 rnin (lane 4), 90 min to 120 rnin (lane 5) , 90 rnin to 150 rnin (lane 6) or 150 rnin to 180 rnin (lane 7) and control cells 30 min (lane 1) or 180 min (lane 2) after induction of sporulation. In this particular autoradiogram the lower intensity of hybridization observed in lanes 5 and 6 is due to smaller amounts of RNA applied to these lanes
with that induced by heat treatments indicates that three of the proteins (39 kDa, 70 kDa and 76 kDa), synthesized at control temperatures, are similar in molecular mass and pH range to the heat-induced proteins hsp39a, hsp70 and hsp76 respectively (Fig. 4A, C and E), and may thus be related. The 39-kDa protein is constitutively expressed throughout sporulation at physiological temperatures, whereas the 70- kDa and 76-kDa proteins are induced only by 120min of sporulation. Interestingly heat shock treatment does not appreciably increase the level of expression of these two pro- teins at that time (Fig. 4C). Previously we have demonstrated that a 70-kDa protein (denoted protein 50) with electro- phoretic migration properties similar to hsp70 exhibits maximal synthesis at 120 min of sporulation [13].
hsp70 is expressed during sporulation
In order to confirm the identity between the 70-kDa pro- tein induced during sporulation at normal temperature and hsp70, the major heat-shock protein of Blastocladiella, we compared the proteolytic digests resulting from the action of V8 protease on both hsp70 induced by heat shock and the 70- kDa protein induced during sporulation (120-min sporulating cells). Spots corresponding to these proteins were excised from two-dimensional gels and subjected to limited proteolysis ac- cording to Cleveland et al. [27]. As shown in Fig. 5, the patterns of protease V8 peptides from heat-shocked cells and sporulating cells are virtually identical.
In order to determine whether hsp70 mRNA is expressed during sporulation at normal temperatures, the technique of RNA blot analysis (Northern hybridization) was employed. Using the Drosophila hsp70 gene as a probe, we detected, under low-stringency hybridization conditions, a 2600- nucleotide transcript in Northern blots with either total RNA (Fig. 6B) or poly(A)-rich RNA (Fig. 6A). In agreement with the data on the synthesis of the 70-kDa protein during sporulation, maximum accumulation of the 2600-nucleotide transcript occurs by 120 rnin of sporulation at normal temper-

21 8
Fig. 7. Immunoprecipitation of the 70-kDa protein from normal and heat-shocked cells. Labeled extracts were immunoprecipitated with antiserum against E. coli dnaK protein and SDS/polyacrylamide gel electrophoretograms of the precipitated proteins were analyzed by fluorography. Lanes: 1,120-min sporulating cells; 2, sporulating cells heat-shocked from 30 rnin to 60 min; 3, E. coli K12 heat-shocked at 42°C for 12min; 4, E. coli K12 control cells. Labeling with [35S]methionine was done in the last 20 min for Blastocladiella and 7 rnin for E. coli. The arrowhead on the right indicates hsp70
ature (Fig. 6A); hybridization was not detected using either RNA from the earliest times of sporulation or zoospores (not shown). The synthesis of hsp70, induced by heat shock at various stages of sporulation, is also due to an accumulation of hsp70 mRNA, as shown in Fig. 6B.
Additional evidence in support of the identity between the 70-kDa protein and hsp70 of Blustocludiellu is provided by immunological cross-reaction of both proteins with antiserum against dnaK protein from E. coli, the prokaryotic analogue of hsp70 [4]. A protein doublet in the 70-kDa region of the gel is specifically immunoprecipitated from Blustocludiellu cell extracts labeled with [35S]methionine by anti-dnaK antibody (Fig. 7). No attempt was made to quantify the increased synthesis of this protein during sporulation, utilizing the immunoprecipitation assay, because of occasional proteolysis during the incubation with the antibody.
Regulation of heat-shock gene expression is at the transcriptional level
Regulation of gene expression during sporulation is mainly at the transcriptional level [13], whereas during germination mRNAs stored in the zoospore are translated in early germination [15, 161 (Silva et al., unpublished results). Consequently translational activation of pre-existing hsp mRNA could be occurring during zoospore germination. In order to determine whether the regulation of the response to heat shock is different in these two differentiation phases, cells at both stages were submitted to treatment with actino- mycin D during heat shock. Treatment of zoospores with actinomycin D during germination ( 5 pg/ml) inhibited the induction of hsps (not shown) suggesting that control of ex- pression of these proteins is at the transcriptional level. At
Fig. 8. Cell-j?ee translation products of RNA isolated from control and heat-shocked cells. Poly(A)-rich RNA, isolated from each sample, was translated in a rabbit reticulocyte lysate system. The. ["S]methionine proteins were analyzed by two-dimensional gel electrophoresis and fluorography. Sporulating cells, heat-shocked at 38°C from 30 rnin to 60 rnin (A) or 90 rnin to 120 rnin (B) after induction of sporulation; control sporulating cells 120 rnin after induction of sporulation at 27°C (C); zoospores (D). Arrows indicate heat-shock proteins

219
any time during sporulation, hsps synthesis was also inhibited by adding actinomycin D ( 5 pg/ml) just before the shift to 38 "C (not shown).
Additional evidence that hsps mRNAs are accumulated during heat shock comes from in vitro translation data using a rabbit reticulocyte cell-free protein-synthesizing system. Examination, by one-dimensional gel electrophoresis, of the spectrum of proteins in the translation products of reactions programmed with poly(A)-rich RNA from either heat- shocked cells or normal cells revealed a marked increase in the relative labeling of proteins having electrophoretic mobilities similar to hsp70, hsp30, hsp25, and hspl7 (not shown). There- fore, the increased labeling of these proteins reflects an increase in the abundance of their respective mRNAs. Two- dimensional gels of these translation products confirm that the mRNAs for hsp70, hsp30a, hsp25, hspl7a and hspl7b are induced during heat shock (Fig. 8A and B). Furthermore, in the one-dimensional pattern of synthesis directed by poly(A)- rich RNA from control cells a band corresponding to hsp70 is clearly visible only at 120 min (not shown) and a cor- responding spot of electrophoretic mobility similar to hsp70 can be seen in the two-dimensional gel (Fig. SC), in agreement with our hybridization data, which show that the mRNA for this protein accumulates at 120 min of sporulation. We have shown that the mRNA for this 70-kDa protein (previously denoted 50) has accumulated at this stage of the sporulation process [13]. Fig. 8 D shows also that these heat-shock mRNAs are not stored in the zoospores, confirming the ex- periments with actinomycin D showing that, even during germination, the regulation of heat-shock gene expression is probably at the transcriptional level.
DISCUSSION
When B l ~ s t o c l ~ d i e l l ~ cells are transferred to 38°C at various stages of the life cycle, the synthesis of at least 22 polypeptides is markedly induced. The most abundant heat- shock protein induced in Blastocladiella, hsp70, is analogous to the Drosophila hsp70 and to E. coli dnaK protein. Immunoprecipitation of a 70-kDa protein, induced by heat shock, with anti-dnaK serum and the detection of its mRNA using a Drosophila hsp70 probe provide strong evidence for the identity of these proteins. Blastocladiella hsps have quite different and individual heat-induction characteristics with respect to the developmental stage of the organism. Some hsps are not induced in certain phases even under heat treatment, whereas the synthesis of others is induced by heat shock only at specific stages of the life cycle. For example, hsp24 is typical of germination, and not expressed at other developmental stages, while hsp39b, hsp39c and hsp39d are only expressed during sporulation. It is not clear why a particular subclass of hsps would be preferentially induced at different develop- mental stages; this selectivity may reflect more general regula- tory phenomena fundamental to the differentiation of the organism. The behavior of these hsps contrasts to seven other hsps (82a, 82b, 76, 70, 60, 25, 17b), which are induced throughout germination, sporulation and growth. Thus, it may be that it is these seven hsps, which are principally in- volved in the acquisition of thermotolerance, which can be induced in this organism (unpublished results).
Regulation of hsps synthesis in B. emersonii occurs not only in response to temperature elevation but actually occurs as part of the normal development of the organism. Three hsps (hsp70, hsp76, hsp39a) are synthesized at physiological
temperatures during specific phases of sporulation; they are not spontaneously induced at other times. Northern hybrid- ization and in vitro translation studies showed that the in- creased synthesis of hsp7O during sporulation reflects the accumulation of hsp70 mRNA at this specific developmental stage. By the criteria of peptide mapping, the hsp70 gene, expressed spontaneously during sporulation at normal temperatures, is that which is also induced by increased tem- perature. It is also important to note that studies using [35S]methionine indicate that the levels of synthesis of hsp70, hsp76, and hsp39a are not significantly increased if, during the time of spontaneous hsp expression, the organism is heat- induced. We conclude that a developmental control is appar- ently superimposed on the heat inducibility of hsps synthesis.
Protein synthesis is essential to the zoospore germination process. The transition from zoospore to germling uses essentially the mRNAs stored in the zoospore [15, 161 (Silva et al., unpublished results). In contrast we have evidence that the control of the expression of heat-shock genes is exerted at the transcriptional level. No heat-shock protein mRNA is present in the zoospore, despite the induction of the hsp70- related protein during sporulation.
Presumably the increased synthesis of hsp70, and possibly other hsps (76 and 39a), during sporulation is not simply a response to stress caused by the starvation conditions. The appearance of hsp70 and hsp76 occurs only 120min after induction of starvation, whereas the pattern of protein syn- thesis starts to change much earlier during sporulation. Therefore, the activation of the hsp7O gene during sporulation is not part of a response to starvation, but rather part of a developmental program. These heat-shock proteins might have important functions during sporulation.
A family of hsp70-related genes was found in Drosophilu and yeast genomes [3, 301. In both cases some members are heat-shock-inducible while others are transcribed under normal growth conditions and not inducible by heat treat- ment. The protein products encoded by these genes are similar but not identical when analyzed by two-dimensional gel or peptide mapping [31,32]. The function of these cognate genes remains obscure.
Despite the lack of information about the function of heat- shock proteins, our data reinforce the idea that heat-shock proteins are not merely associated with protection against hyperthermia, i.e. that they must exert important metabolic and structural functions in the cell as well. As the metabolic and structural necessities of an organism change during the developmental process, the response against the environmen- tal stress must be adapted to the new internal reality, leading to the synthesis of other heat-shock proteins in addition to those which always appear to be necessary for the initial heat- shock response. It has been proposed that the function of the heat-shock proteins is to limit or repair damaged, abnormal or denatured proteins, with hsp70 playing a role in repair of heat-damaged nucleoli [7]. Following heat shock, hsp70 initially binds tightly to some nuclear components and then to nucleoli [33, 341. hsp70 has a high affinity for ATP [35] and the analogous protein from E. coli (the dnaK product) has an ATPase activity [36]. It has been suggested that a system of ubiquitin-linked protein degradation may have a key role in the heat-shock response [37]. A relationship to the dramatic increase in protein turnover, which occurs during sporulation in B l ~ , ~ t o c l ~ d ~ e l l ~ [13], is suggested by the spontaneous induc- tion of some hsps during the sporulation process, with the accumulation of hsp70 mRNA at a specific stage near the midpoint of sporulation. In this context, work in progress

220
directed towards the isolation of the Blustocladiellu hsp70 gene may contribute to a better understanding of the highly conserved heat shock response.
During the course of this work two papers have appeared suggesting that the synthesis of some heat-shock proteins in Bacillus subtilis (hsp84, hsp3 1 ) and yeast (hsp84, hsp26 and hsp70-related proteins) may also to be under sporulation regu- lation [38, 391. Such observations reinforce our hypothesis that hsps may perform functions of fundamental importance during sporulation.
We thank Dr C. Klein for her reading and helpful comments concerning this manuscript and Dr F. H. Quina for his help with the English. We also thank Dr E. Craig and Dr C. P. Georgopoulos for their generous gift of heat-shock gene clone and dnaK antibody and Drs C. L. S. Santos and R. R. Brentani for generously providing reticulocyte lysates. This work was supported by grants from the Fundaciio de Amparo a Pesquisa do Estado de SZo Paulo (FAPESP 8513412-6), the Conselho Nacional de Desenvolvimento Cientifico e Tecnoldgico (CNPq 40339/84) and the Financiadora de Estudos e Pro-
jetos (FINEP 4384072500). A.M.S. was a recipient of a FAPESP predoctoral fellowship. M.C.M.B. was a Visiting Scientist from Universidade Federal da Paraiba supported by FAPESP and CNPq.
REFERENCES 1.
2.
3.
4.
5.
6. 7. 8. 9.
10.
Schlesinger, M. J., Aliperti, G. & Kelley, P. M. (1982) Trends
Neidhardt, F. C., VanBogelen, R. A. &Vaughn, V. (1984) Annu.
Ingolia, T. D., Slater, M. R. & Craig, E. A. (1982) Mol. Cell Biol.
Bardwell, J. C . A. & Craig, E. A. (1984) Proc. Natl Acad. Sci.
Kelley, P. M. & Schlesinger, M. (1982) Mol. Cell Biol. 2, 267-
Bienz, M. (1985) Trends Biochem. Sci. 10, 157-161. Pelham, H. R. B. (1985) Trends Genet. I , 31-35. Lindquist, S. (1981) Nature (Lond.) 293, 311 -314. Bienz, M. & Gurdon, J. B. (1982) Cell29, 811 -819. Collatz, E., Plesset, J., Foy, J. J. & McLaughlin, C. S. (1985)
Biochem. Sci. 7,222 - 225.
Rev. Genet. 18,295-329.
2,1388-1398.
USA 81,848-852.
274.
Yeast I , 49 - 56.
11. Zimmerman, J. L., Petri, W. & Meselson, M. (1983) Cell 32,
12. Bensaude, O., Babinet, C., Morange, M. & Jacob, F. (1983)
13. Silva, A. M., Maia, J. C. C. & Juliani, M. H. (1986) Cell Differ.
14. Lovett, J. S. (1975) Bacteriol. Rev. 39, 345-404. 15. Soll, D. R. & Sonneborn, D. R. (1971) J. CellSci. 9, 679-699. 16. Leaver, C. J. & Lovett, J. S. (1974) Cell Differ. 3, 165-192. 17. Jaworski, A. J. & Thomson, K. (1980) Dev. Biol. 75, 343-357. 18. Bonato, M. C. M., Silva, A. M., Maia, J. C. C. & Juliani, M. H.
19. O’Farrell, P. H. (1975) J. Biol. Chem. 250,4007-4021. 20. Laemmli, U. K. (1970) Nature (Lond.) 226, 680-685. 21. Laskey, R. A. & Mills, A. D. (1975) Eur. J . Biochem. 56, 335-
341. 22. Chirgwin, J. M., Przybyla, A. E., Mac Donald, R. J. & Rutter,
W. J. (1979) Biochemistry 18, 5294-5299. 23. Maniatis, T., Fritsch, E. F. & Sambrook, J. (1982) Molecular
cloning. A laboratory manual. Cold Spring Harbor Laboratory Press, NY.
24. Aviv, H. & Leder, P. (1972) Proc. Natl Acad. Sci. USA 69,1408 - 1412.
25. Pelham, H. R. B. & Jackson, R. J. (1976) Eur. J . Biochem. 67,
26. Jackson, R. J. & Hunt, T. (1983) Methods Enzymol. 96, 51 -74. 27. Cleveland, D. W., Fisher, S. G., Kirschner, M. W. & Laemmli,
28. Southern, E. M. (1975) J . Mol. Biol. 98, 503-517. 29. Gomes, S. L. & Shapiro, L. (1984) J. Mol. Biol. 178, 551 -568. 30. Ingolia, T. D. & Craig, E. A. (1982) Proc. Nut1 Acad. Sci. USA
31. Wadsworth, S. C. (1982) Mol. Cell Biol. 2,286-292. 32. Craig, E. A. & Jacobsen, K. (1985) Mol. CellBiol. 5,3517-3524. 33. Velazquez, J. M. & Lindquist, S. (1984) Cell36, 655-662. 34. Welch, W. J. & Feramisco, J. R. (1984) J. Biol. Chem. 259,4501 -
35. Welch, W. J. & Feramisco, J. R. (1985) Mol. Cell Biol. 5, 1229-
36. Zylicz, M., LeBowitz, J. H., McMacken, R. & Georgopoulos, C.
37. Munro, S. & Pelham, H. (1985) Nature (Lond.) 317,477-478. 38. Todd, J. A., Hubbard, T. J. P., Travers, A. A. & Ellar, D. J. (1985)
39. Kurtz, S., Rossi, J., Petko, L. & Lindquist, S. (1986) Science (Wash.
1161 - 1170.
Nature (Lond.) 305, 331 -333.
4,263 - 274.
(1984) Eur. J . Biochem. 144, 597-606.
247 - 256.
U. K. (1977) J. Biol. Chem. 252,1102-1106.
79, 525 - 529.
4513.
1237.
(1983) Proc. Natl Acad. Sci. USA 80,6431 -6435.
FEBS Lett. 188,209-214.
DC) 231,1154-1157.




















