
Different 5′ leader sequences modulate β-glucuronidase accumulation levels in transgenic...
8
i::ul'h.l'tic(J 85: 209 216. 1995. 209 © 1995 K/I/H'C/" AU/tlemi" Pl/lJlishers. Primetl in the Ne/her/antls. Different 5/ leader sequences modulate ,3-glucuronidase accumulation levels in transgenie Nicotiana tabacum plants Marc De Loosel, Xavier Danthinne 2 , Erik Van Bockstaele l , Marc Van Montagu 2 & Ann Depicker 2 I RijksslC/tion voor Plantenveredeling, Cenlrum voor Landbouwkundig Onderzoek Gent, Burg. Van Gansherghe/aan /09, B-9820 Mere/beke, Belgium; 2 Laboralorium voor Genetica, Universiteil Gent, KL Ledeganckslraat 35, B-9000 Gent, Be/giunl Key \Yards: ,U-glucuronidase, plant, silencing, translational control, 5'-untranslated region, variation of gene expressIon Summary Three random synthetic leaders and three naturally-occurringleaders, the lObaceo mosaic virus (TMV) coat protein, the satellite tobacco necrosis virus (STNV) and the plant chlorophyll a/b-binding protein (Cab22L), were shown 10 modulate the U-glucuronidase reporter protein accumulation levels in transient expression experiments. The same chimcric constructs also confer differential distribution patlems of reporter protein accumuiation in stably- transformed tobacco calli or regenerated transgenic plants. When the highest expression levels with a given leader are compared, the 31-nucleotide random leader stimulates translat ion 20- and 100-fold reiative to the 9- and 4- nucleotide synthetic leaders respectivel y. However, this 3 I-nucleotide random leader is approx. 2 to 3-fold wcaker than the 30-nucleotide STNV leader and even S-fold weaker than both the 79-nuclcotide TMV leader and the 66-nuclcotide Cab22L leader. These results confirm the findings in transient expression experiments and stress the of the S' -untranslated reg ion for the production of heterologous proteins in transgenic plants. Introduction effect on translational enhancement (Gallie & Walbot. J992). The satellitt: tobacco necrosis virus (STNV) Of all the regions of messenger RNA that can be (Danthinne et al., 1991) RNA lacks a conventional cap involved in translational efficiency, the leader sequence structure at the 5' terminus, but neverthe1ess thc viral is probably the most important (Kozak, 1989; Hershey, coat protein is very efficiently translated (Danthinne 1991). Firstly, this region is undoubtedly implicated in et al., 1993). The Cab22L leader also has a stimula- thc process of translational initiation and this is usually tory effect upon the trans-protein accumulation after consiclcred to be the rate-limiting step of translation. transient expression compared with the effect of the Sccondly, the 5' leader can be directly or indircctly cauliAower mosaic virus (CaMV) 3SS 5' leader (Harp- involved in mRNA stability. ster et al., 1988). Plant viral RNA leader sequences, like those of In an in vitro translation system derived from wheat tobacco mosaic virus (TM V), alfalfa mosaic virus germ, thc STNV leader sequence only weakly stimu- (A!MV) and plant potyvirus, have been shown to act lates translation relative to the leaders of TMV and in cis as dlicient enhancers of translation at least in in of Cab22L origin (Danthinne & Van Emmelo, 1990). vitro systems or during transient expression in pro!O- In transient expression the STNV-I, the TMV and the pla:;ts (Gallie et al., 1987a. 1987b; Jobling & Gehrke. Cab22L leaders exhibit a 4-, 12-. and 8-fold enhance- 1987; Sleat ct al., 1987; CalTington & Frecd, 1990, ment respectively, relative to a 31 nucleotide random Nicoiaisen et al., 1992). Motil's in the TMV lead- leader (Danthinne & Van EmmeJo, 1990). er of 77 nucleotides were studicd in detail for their
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Different 5′ leader sequences modulate β-glucuronidase accumulation levels in transgenic...

i::ul'h.l'tic(J 85: 209 216. 1995. 209 © 1995 K/I/H'C/" AU/tlemi" Pl/lJlishers. Primetl in the Ne/her/antls.
Different 5/ leader sequences modulate ,3-glucuronidase accumulation levels in transgenie Nicotiana tabacum plants
Marc De Loosel, Xavier Danthinne2, Erik Van Bockstaele l , Marc Van Montagu 2 &
Ann Depicker2
I RijksslC/tion voor Plantenveredeling, Cenlrum voor Landbouwkundig Onderzoek Gent, Burg. Van
Gansherghe/aan /09, B-9820 Mere/beke, Belgium; 2 Laboralorium voor Genetica, Universiteil Gent, KL Ledeganckslraat 35, B-9000 Gent, Be/giunl
Key \Yards: ,U-glucuronidase, plant, silencing, translational control, 5'-untranslated region, variation of gene expressIon
Summary
Three random synthetic leaders and three naturally-occurringleaders, the lObaceo mosaic virus (TMV) coat protein, the satellite tobacco necrosis virus (STNV) and the plant chlorophyll a/b-binding protein (Cab22L), were shown 10 modulate the U-glucuronidase reporter protein accumulation levels in transient expression experiments. The same chimcric constructs also confer differential distribution patlems of reporter protein accumuiation in stablytransformed tobacco calli or regenerated transgenic plants. When the highest expression levels with a given leader are compared, the 31-nucleotide random leader stimulates trans lat ion 20- and 100-fold reiative to the 9- and 4nucleotide synthetic leaders respectivel y. However, this 3 I-nucleotide random leader is approx. 2 to 3-fold wcaker than the 30-nucleotide STNV leader and even S-fold weaker than both the 79-nuclcotide TMV leader and the 66-nuclcotide Cab22L leader. These results confirm the findings in transient expression experiments and stress the import~lncC of the S' -untranslated reg ion for the production of heterologous proteins in transgenic plants.
Introduction effect on translational enhancement (Gallie & Walbot. J992). The satellitt: tobacco necrosis virus (STNV)
Of all the regions of messenger RNA that can be (Danthinne et al., 1991) RNA lacks a conventional cap involved in translational efficiency, the leader sequence structure at the 5' terminus, but neverthe1ess thc viral is probably the most important (Kozak, 1989; Hershey, coat protein is very efficiently translated (Danthinne 1991). Firstly, this region is undoubtedly implicated in et al., 1993). The Cab22L leader also has a stimulathc process of translational initiation and this is usually tory effect upon the trans-protein accumulation after consiclcred to be the rate-limiting step of translation. transient expression compared with the effect of the Sccondly, the 5' leader can be directly or indircctly cauliAower mosaic virus (CaMV) 3SS 5' leader (Harpinvolved in mRNA stability. ster et al., 1988).
Plant viral RNA leader sequences, like those of In an in vitro translation system derived from wheat tobacco mosaic virus (TM V), alfalfa mosaic virus germ, thc STNV leader sequence only weakly stimu(A!MV) and plant potyvirus, have been shown to act lates translation relative to the leaders of TMV and in cis as dlicient enhancers of translation at least in in of Cab22L origin (Danthinne & Van Emmelo, 1990). vitro systems or during transient expression in pro!O In transient expression the STNV-I, the TMV and the pla:;ts (Gallie et al., 1987a. 1987b; Jobling & Gehrke. Cab22L leaders exhibit a 4-, 12-. and 8-fold enhance1987; Sleat ct al., 1987; CalTington & Frecd, 1990, ment respectively, relative to a 31 nucleotide random Nicoiaisen et al., 1992). Motil's in the TMV lead leader (Danthinne & Van EmmeJo, 1990). er of 77 nucleotides were studicd in detail for their

210
This work analyzes whether the effect of the leader sequence on heterologous protein accumulation seen after transient expression (Danthinne & Van Emmelo, J990) can be extrapolated to that which is found in stably-transfonned plant tissues. The work also aims to determine whether the leader affects the distribution of protein accumulation levels beyond the variation of expression levels due to position and silencing effects. Therefore, the expression of the different chimeric gene constructions was studied first of all by comparing the values for the GUS activities in two series of 30 different calli. Subsequently the expression of the same chimeric constructs was evaluated in regenerated transgenie plants and their progeny.
Materials and methods
Plasmid constructions
The cauliftower mosaic virus promoter (P35S) was mutagenized to create a PstI restriction site (Danthinne & Van Emmelo, 1990) immediately downstream from the three putative transcript ion start sites (Ow et al., 1987; Sanders et al., 1987; Harpster et al., 1988). All leader sequences were fused to the ,8-g1ucuronidase (gus)-coding sequence at the Ncol site positioned at the first codon. Downstream, the gus-coding sequence was fused to the 3' region of the nopaline synthase gene (Depieker et al., J982; Ingelbrecht et al., 1989) at the EcoRI site located 6 nucleotides downstrcam from the stop codon (Jefferson, 1987). The chimeric gus genes were put under the control of the mutagenized 35S promoter in such a way that only three heterologous nucleotides GAC are present at the 5' end of all the transcripts. The chimeric genes were introduced into the XbaI site of the binary T-DNA vector pDEIOOI (Denecke el al., 1992) in both orientations relative to the right border. The resulting plasmids (pXD601 10
pXD612) were given an odd or an even number when the gus caSsette pointed to the inside or to the outside of the T-DNA, respectively (see Figs IA and 1B).
Conjup,ation and plant cell transformations
All the T-DNA vectors were mobilized from Escherichia coli MC 1061 (Casadaban & Cohen, 1980) to the Ap,robacrerium strain C58C I, containing plasrnid pGV2260 (Deblaere et al., 1987). Transconjugants harbouring the correct T-DNA constructs werc co-cultivated with Nicotiana rabacul11 SR I protoplasts
(Depieker et al., 1985) or with leaf discs (De Block et al., 1987). Kanamycin-resistant calli were grown on B5 medium (Gamborg et al., 1968) supplemented with honnones and 50 lig/mi kanamycin. Each T-DNA COnstruct was transformed twice. From both independentIy performed co-cultivations, 30 calli wcre analyzccl for GUS activity. The GUS-negative calli were no\ considered. The calli are indicated by CXD followed by the number of the T-DNA plasmid construct and a number to designate the individual clone. Kanamycinresistant shoots were cut from leaf discs and grown in glass containers. The independently obtained plant lines are indicated by LXD followed by the number of the corresponding T-DNA construct and a number indicating each individual shoot. For each construcl (except XD603), between 4 and 20 axenically-grown transgenics were analyzed at the 6-1eaf stage approximately 3 months after their isolation from the leaf disc. Subsequently, 2 to 5 of the highest expressing transgenics were transferred to soil, grown in the greenhouse and allowed to set seed. About 50 secds of the RI generation were sown on Murashige and Skooi' medium containing 50 pg/ml kanamycin. The r lil between kanamycin-resist<mt and kanamycin-sensitive seedlings was detennined. FIOm lines with a single TDNA locus, eight to twelve kanamycin-resistant Rl plants were grown in vitro up to the 6-leaf stage anel analyzed for their GUS content.
GUS assays
Fig.!.The enzyme activities (Jefferson, 1987) were detel P35S-1 mined by means of continuous kinetic reactions per indic31
Pnos.formed spectrophotometrically (340 ATTe, SLT or the noiEMS, Labsystems Inc.) or ftuorimetrically (FluB. NUl
oroscan, Labsystems Inc.) in a computer-directed the Val
,G-gluc
(1993). The detection limit (values two-fold abov· background) was 0.1 U GUS/mg protein. The pr<
microtitre plate reader as described by Breyne et al
mant:tein concentrations were determined by the method ol casse'Bradford (1976) using the kit supplied by Bio-Rad. positi the si nurnb
Results
DisfnT-DNA construcrs
ealli .'
The basic structure of the set of constructed T-DNA Distri
vectors is presented in Fig. J. All T-DNAs carry the genic
same neomycin phosphotransferase hybrid gene allo\.\!els. F
ing the select ion for kanamycin-resistant transfor

:Iock 'n on with Conjentyzed
not lwed nd a Icin'own )Iant nber nber truct 'own 'oxiiisc. 'anseen-f the :oog ratio itive Ie T
RI and
~ter-
perI or Flucted t al. love prodof
'NA the
owfor-
LB RBA. 211 odd number
3' oes np til P nos 3'nas gus P 355
LB RB
even mumber
3' Des np ril P nos P 355 gus 3' nos
B. Plasmid constructionsLeaders
pXD601 pXD6024nt random leader
(i.ce.uei-
pXD603 pXD6049nt random leader
G .. CUCGACC ... uCi -
30nt STNV leader pXD605 pXD606
ei ...cu ........G... c ... GG ........ Cuuu .. CUG ACU -cc ~
pXD607 pXD60B31 nt random leader
GACGU ...CCCGGGG ....ucc:JC'.l ...G....GucO..ccc~
79nt TMV leader pXD609 pXD610I G"'C G"U cc Gu"'uuuu IJ AC ..... c ....uIJ"C CAC" ..CA ..... .a.c ...... c .. ..C..... "C'" AC"U IJ AC ......VVV "'CU ...uu Cu"'O ......UIJ ...cc~
66nt Cab22L leader pXD611 pXD612
I co ",cu CG "'Geve.u IJ IJ =veu "uu ..euve ..Gc .....u .....c...... .toG ....CUcuuvucuCUUCuv ..uu "',. "'cc "'UG
Fig. I. Description of the T·DNA consUlJcts. A. Schematic represenlalion of the T-DNA construcLs used. The veclors containing the cassene P35S-leader-gus-3' nos oriented to the insiJe or the outside of the T-DNA are indicated with odd and even numbers respeclively. The star symbol Ifldicatcs the position where the various leaders, described in Fig. IB. were cloned. Abbrevialions: P35S. caulillower mosaic virus promoter; Pnos. nopaline synthase promoter; SliS. O-glucuronidase gene; nptll. neomycin phosphouansferase ll-coding sequenee; 3' nns. 3' region of !he nopaline synthase gene; 3'ocs. 3' region of the octopinc synthase gene; RB. T-DNA right border region; LB. T-DNA lef! border region. Il. Nucleotide sequence of the leaders tesled for uanslational enhancement and the nomenclature of the vee tors earrying the T-DNAs with 'he various leader fusions. The tirst residues are derivcd from the uanscription start of the CaMV 35S promoter. The initialion codon of the ./-glucuronidasecoding sequenee is underlined.
mams. The different chimeric P35S-leader-gus-3'nos cassettes were all insened into one vector at the same position in two orientations (Fig. I A). Figure 18 shows lhe six different leaders analyzed in this work and the number of the constructs in which they OCCUr.
Dislribulion ofCUS aClivilies in dijferenllransgenic calLi series
Distributions of the GUS activities of the CXD transgenie ealli were obtained from GUS accumulation lev'1:'. figurc 2A shows the GUS amounts in 16 calli
of all transgenic series plotted on a Jogarithmic scale. To emphasize the large differences in absolute GUS amounts among the different series as weil as within one series, the same results were pJotted on a Jinear scale (Fig. 28). Each series contained calli with low, intennediate and high GUS accumulation levels. The same range of variation with the same absolute amounts for the lowest and highest expressing calli was confirmed in a second experiment in which a funher 30 calli were analyzed. Thc main ditlerence bet ween the results of the two consecutive experiments was the number of calli with low and high GUS activ

212
U GUS/mg protein
A. 10,000,
I I
1,000 I I
100
0.1
1000 U GUS/mg protein
B. 10
8
6
4
2
0 c)'
C:) C)'l,.
C:) c:::,"IQ dJ<
IQ
'~I C)0
IQ c:::,re,
IQ c)/\..
IQ
ii c:::,'ó
IQ C)~
IQ ,c:::,
re, ,,'re, ,,'l,.
re, 1-<:) 1-<:) 1-<:) 1-<:) 1-<:)1-<:) 1-<:) 1-<:) 1-<:) 1-<:) 1-<:) 1-<:)
G G G G G G G G G G G G I I I I I I I I I I I I 4 nt RL 9 nt RL 30 nt 31 nt RL 79 nt 66 nt
STNV L TMV L Cab22L L
CXD calli Fig. 2. DislIihulion of GUS aClivilics in lransgenic calli series. GUS aClivilies in lIansgenic calli series were plolIed on a. logarithmic (Al anti a linear (B) scaIe. The G US aClivilics delermined in (ach callus numher of lhe different scrics arc ft'prcsc/lted by bars. Within each scrics. t.hc numbers of calli arc arranged accorJing 10 their increasing GUS conlent.
ity. A comparison shows lhat lhe distributions of GUS 1l1e 4-nllcleotide random leader (RL) yielded thc accumuJation levels in series containing an identical lowest expression but was significantly above bad chil1leric f!,/IS gene but in opposite orientations. were ground. Higher GUS levels \\'ere fOllnd in calli witl: similar. the 9-nucleolide RL transcript (series CXD60] ;(IHI
However. the GUS activity dislributions of series CXD604) and still higher levels were found in tran~
containing J;US COnSlrlJCIS wilh different leaders dis genic calli with a 31-nllcleolide RL in the RUS triU1scrip\ play a pronounced difference (Figs 2A and 28). (series CXD607 and CXD608). The highest aCClll1lll
lati pat I
ST]
nllc and and
Infl pril
For wel ma in t
gro GU wel 3A p]al ry/ GU pro teir the nllc for the mll sho trar
cu
Onl lint owi RI ber plal DN seg sen 3: I mal ana of tI mu and
the in t

nel h~
Je
kth Id spt J-
lat ion levels and more or less comparabie distribution pattems were found in thescries with the 30-nuc\eotide STNV leader (series CX 605 and CXD606), the 77nucleotide TMV leader (series CXD609 and CXD61 0), and the 66-nucleotide Cab22L leader (series CXD611 and CXD612).
Infiuence oflhe leader on GUS accumu/alion in primary Iransgenie planls
For each of the T-DNA constructs transgenie plants were obtained and analyzed for their GUS expression in a leafofan in vilro-grown plant (Figs3A and 38). As in the transgenie calli series, the primary axenicallygrown transformants also showed a high variation in GUS units per mg of tot al protein. The obtaincd reslJlts were plotted on a logarithmic and a linear scale in Figs 3A and 38 respectively. As the number of transgenie plants analyzed was limited, it is not possible to carry out a statistical analysis of the data. The highest GUS values per leader construct are 6150 U GUS/rng protein for the Cab22L leader, 3536 U GUS/mg protein for the TMV leader, 1167 U GUS/rng protein for the STNV leader, 309 U GUS/mg protein for the 31nucleotide random leader (RL), 36 U GUS/mg protein for the 9-nucleotide RL and 20 U GUS/mg protein for the 4-nucleotide RL. Thus a comparison of GUS accumulation in leaf material of primary transgenie plants shows the same hierarchy of leader effects as in the transgenie calli series.
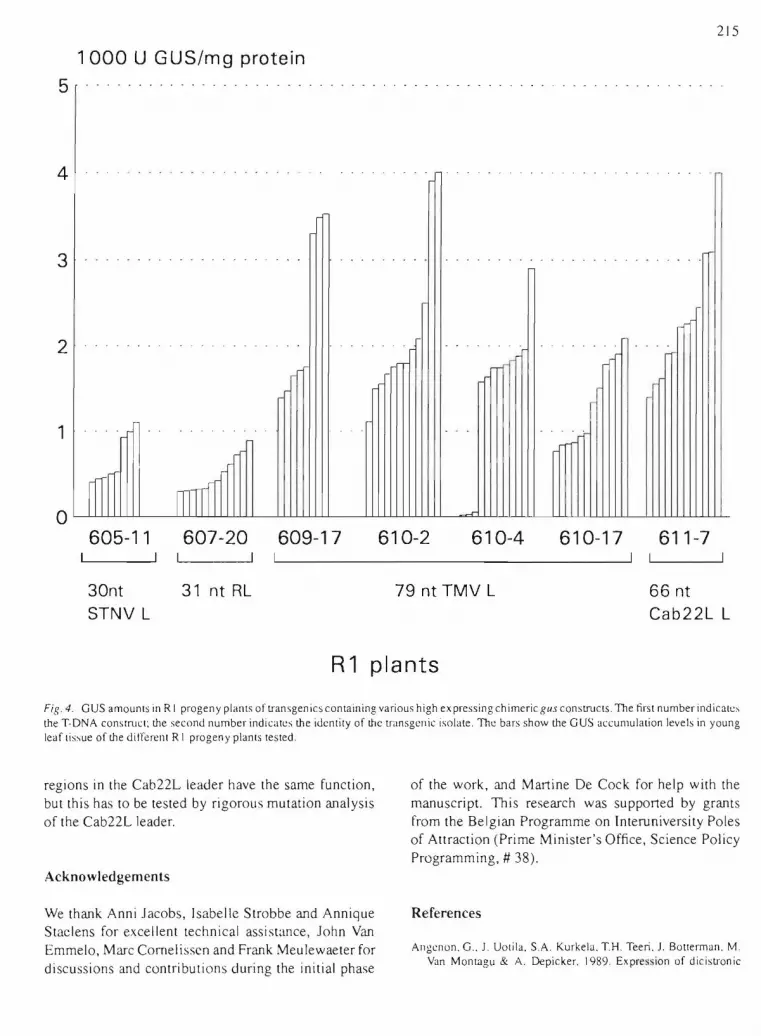
GUS expression in progeny planls
One or more of the highest-expressing, transgenie lines per T-DNA construct, indicated by a dark shadowing in Fig. 38, were allowed to f10wer and the RI seeds were collected. Subsequently the number of kanamycin-resistant versus kanamycin-sensitive plants was determined to estimate the number of TDNA loci. Most of the RI progenies show a 3 to I segregation of kanamycin-resistant versus kanamycinsensitive seedlings. Of the transgenics with c\ear 3: I segregation ratias for the T-DNA-linked selection marker, 8 or J 2 R I in vilro-grown progeny plants were analyzed for GUS accumulation levels (Fig. 4). Most of the progeny shows two distinct classes of GUS accumulation probably corresponding to the heterozygotes and the homozygotes for the T-DNA locus.
We can derive from the RI population analysis that the same hierarchy of expression levels is maintained in the progeny of the primary transgcnics: narnely the
213
9-nucleotide RL < 31-nucleotide RL < STNV leader < TMV and Cab22L leaders. Although the TMV-gu.\ and Cab22L-gus constructs gave rise to similar GUS distributions, it may not be a coincidence that the highest accumulation levels were always found in transgenie material with the Cab22L leader sequence.in the transcript.
Discussion
The results on GUS accumulation levels in different transgenie lines with a particular T-DNA construct illustrate the commonly-observed variation in transgene expression. Clearly, every series contains a number of transgenie calli that express the P35S-gus transgene to a very low extent in comparison with a number of calli with high (normal) expression levels. However, the resulting low accumulation levels were always approximately 0.0 1ofthe highest accumulation level in the respcctive series. The variation in gene expression might be the result of position effects such as f1anking enhancing or silencing DNA sequences, the surrounding chromatin structure or the methylation status of the insert locus. However, other processes not understood such as gene silencing and co-suppression may contribute to the observed variation in gene expression in primary transgenic populations (Hobbs et al., 1990, 1993; Ingelbrecht et al., 1994).
Nevertheless, the results demonstrate the importanee of the leader sequence upon heterologous gene expression. Very often chimeric leaders contain linkers and adaptors which can lower substantially the flnal recombinant protein accumulation (Jones et al., 1985).
The 5'-untrarJslated leader might directly influence the accu mulat ion of the gene product as it is undoubtedIy involved in the initiationoftranslation. This isexemplifled by the pOOI' translation of second cistrons in dicistronic transcript ion units (Angenon et al., 1989). However, the 5' -untranslated leader is also involved in mRNA accumulation levels (Harpster et al., 1988). The leaders may increase the mRNA stability because enhanced translation proteets the transcript from more rapid degradation (Vancanneyt et al., 1990). The analysis and correlation of GUS accumulation levels with transcript accumulation levels wil! allow us to c1arify events.
We conclude from the experiments that both the 5'-untranslated region of the Cab22L gene (Harpster et al., 1988) and the 5' leader of the TMV viral coat

214
Ä. U GUS/mg prolein
10,000
1,000
1000 U GUS/mg prolein
B. 7
4
5
6
3
2
4 nl RL 9 nl RL 30nl 31 nl RL 79 nl 66 nl STNV L TMVL L Cab22 L L
Primary Iransgenie planls
Fig. 3. Distribution of GUS aClivitics in young Icaf matcri,,1 of prilll"ry rransgcnic pianIs conIaining mc various IcmJcr-gu5 conslrUCls. plotll',on a logarithmic scale (Fig 3A) and a Iinear scak (Fig. 38). The GUS activities detcmlincd are reprcsenled by bars plollcd on me ordinale. 1111' are grouped per T-DNA construct. Wimin each group. mc GüS arnounts are arranged according 10 incrcasing alllounis. The blacK bars indiellt' me transgenic plants of which me RI progeny was rurther analy7.cd in detail: LXD6ü4-4. LXD6üS-II. LXD6ü7-20, LXD609-17. LXD610 ,', LXD61O-2.and LXD6II-7.
protein (Gall ie et al., 1987a) enhéUlce the expression éUld Cab22L leaders resulting in a Jack of seconda.rv of chimeric gus genes in transgenic pJants. Both lead structure is nolable. This may allow easy scanning 1'01'
ers show similarities In length and nucleotide compo the first AUG in a favourable context (Kozak, 1989) sition. They are bet ween 50 and 70 nucleotides long, Functional analysis of the TMVleader idenlified 1111
which corresponds to the length of the majority of plant poJy(CAA) region as the primary element responsl5' leaders (Joshi, 1987; Hershey, 1991). The under bie 1'01' enhancement in vivu (Gallie & Walbot, 1992) representation of guanosine nuclcotides in the TMV Therefore, it is possible that the analogous CA-rich
Fig. !he !caf
reg but 01'1
Ac:
We Sta Em disl
215
1000 U GUS/mg protein
5
4
3
:ed ICY
ale -4,
ry or I),
le
;1
~ ),
:h
2
1
605-11 607-20 609-17 610-2 610-4 610-17 611-7
30nt 31 nt RL 79 nt TMV L 66 nt STNV L Cab22L L
R1 plants
Fig. 4. GUS amOUnIS in R I progeny plants of transgen ics containing various high ex pressing chimeric gus constrllCIS. The nrs! number indicalc, the T-DNA construct; the second number indicatcs the idenlily of me transgcnic isolale. Thc bars show the GUS accumulation levels in young leaf tissue of me dil'l'crellt RI progeny plants tested.
regions in the Cab22L leader have the same function, of the work, and MaI1ine De Cock for help with the but this has to be tested by rigorous mutation analysis manuscript. This research was suppoI1ed by grants of the Cab22L leader. from the Belgian Program me on Interuniversity Poles
of Allraction (Prime Minister's Office, Science Policy Programming, # 38),
Acknowledgements
We thank Anni Jacobs, Isabellc Strobbe and Annique References Staelens for excellent technical assistance, John Van
Angcnon, G .. J. UOlila, S.A. Korkela, T.H. Teen, J. Botterman, M.Emmelo, Marc Comelisscn and Frank Meulewaeter for Van Montagu & A. Depicker. 1989. Expression of dicistronic discussions and contributions during the initial phase

216
transcriptional units in trans&cllic tobacco. Mol. Cell. Biol. 9: 567&- :;6X4.
Bradford. M.M. 1976. A rapid and sensilivc me!hod for Ihe quantiwtion of microgram quanlities of prolein utilizing !he principle of prolcin-dyc binding. Anal. Biochem. 72: 24X-254.
Breyne. P. M. De Loose, A. Dedonder. M, Van Montagu & A. Depicker. 1993. Quantitative kinetic analysis of :J-glueuronidasc aClivities usin~ a computer-directed microtiter plale reader, Plant Mol. Biol. Rep. 11: 21-31,
Carringwn. J ,C & D.D. Freed. 1990, Cap-independemenhnncement of nanslalion by a plant potyvirus 5' nontranslated region, J. Virology 64: 1590-1597.
Casadaban. M.l. & S,N. Cohen. 19RO, Analysis of gene conITol signals by DNA fusion and doning in 1:.'Icherichia col/, J. Mol. Biol. IlR: 179-207,
Dan!hinne. X, & J. Van Emmelo. 1990. Studies on tJlc translational properties of STNV RNA non-coding rq:ions. Med, Fac. Landbouww. Rijksuniv. Gent 55 (Ja): 1037.. 1045.
DanlJlinne, X.. J. Seurinck, M. Van Montagu. C.W.A. Pleij & J. Van Emmelo, 1991, SlTUctural similaritics between !he R~As of two satellites of tobacco necrosis virus, Virology IR5: 605-614.
Dan!hinne. X., J. Seurinck, F. Meu\cwaeter. M. Van Montagu & M, Cornelissen, 1993. The 3' untrun,lated region of !he satellite lobacco necrosis virus RNA stimulates translalion in ,·i!rn. Mol. Cell. Biol. 13: 3340--3349.
De Bloek. M.. J. Botterman. M, Vandewiele, J. Doch, C Thoen. V. Gosselé. R, Movva. C Thompson, M. Van MOnlagu & J, Leemans. 19R7, Engineering herbicide resislatlL'c in plants by expression of a deloxifying enzyme. EMUO J, 6: 2513-2518,
Deblacre, R.. A. Reynaens, H, Höfte, J .-P Hernalstccns. J. Leemans & M, Van Monlagu, 1987, Vcctors for cloning in plant ceiis. In R. Wu & L Grossman (Eds) Recombinant DNA, part 0 (Me!hods in Enzymology, Vol. 153). pp. 277-292, Academic Press. New York.
Denecke. J .. R. De Rycke & J. Botterman. 1992, Plant and mammalian sorting signals for prolein retention in !he endoplasmic reticulum conlain a conservedepitope, EMBO J. 11: 2345-2355.
Depicker, A,. S. Stachel. P Dhaese, P Zambryski & H.M, Goodman. 1982. Nopalinc synulase: transcripl mapping and DNA sequence. J. Mol. Appl. Gene!. I: 561-573,
Depicker. A.. L. Hcnnan. A. Jacobs. J. Scheil & M. Van Mon[;lgu, 1985. Frequencies of simultaneOlls tTanSfOrmalion wi!h dirfcrenl T-DNAs and !heir relevance to !he Alirohaclerium/plant cell inleraction. Mol. Gen. Genet 20 I' 477-484.
Gallic. OK & V. Walbot, 1992. Idcntilication of !he motifs wi!hin !he tobacco mosaic virus 5' -Ieadcr responsible for enhancing translation. Nuc\. Acids Res, 20: 4631-4638.
Gallic. D.R., D,E. Slea\. J,W Walts. Pc. Turner & TM.A. Wilson. 1'lR7a. 'nlC 5'-leader sequence of tobacco mosaic virus RNA enhances!he cxpression of foreign gene transcripts in vilra and in \'ilO, Nuc\. Acids Res, 15: 3257-3273,
Gallie. OR .. DE Sieat, J.W Walts. PC Turner & T.M.A. Wilson, 19X7b. /\ comparison of eukaryotic viral 5' -leader sequences as cnhanccrs of mRNA expression in \'ivo. Nuc\. Acids Res. 15: Ri\'l.'- X7 1I,
Garnborg. OL. R.A. Miller & K. Ojima, 1968. NUlrient rCljllirements of suspension cultures of soybean rOOI ce lis. Exp. Cc 11 Res. 50: 151-15X
Harpster. M.H.. J,A. Townsend. J.D.G. Jones. J. Bedbrook & P. Dunsmuir. 1988. Relative stTcngths ofule 35S cauliflowcr mosaic virus. I'. 2'. and nopaline syn!hasc prnrnoters in transformed lobacco sugarbeet and oiiseed rape callus tissue. \101. Gen, ('enet. 212: 182-190
Hershey. J.W8. 1991. Translational con trol in rnammalian cells. Ann. Rev. 8iocl1em, 60: 717-755.
Hobbs. SLA, P. Kpodar & CM.O, DeLong, 1990. The effect of T-DNA copy number. position and me!hylation on rcporter gene expression in tobacco transformants. Plant Mo\. Bio\. 15: 851864.
Hobbs. SL.A. TD, Warkenlin & CM,O, Delong. 1993, Transgene copy number can be positively or ncgativcly associalcd wi!h transgene expression, Plant Mol. Biol. 21, 17-26.
lngelbrecht, \.L.W.. L.M.F. Hernlan, R.A, Dekeyser. M,C Van Mon tag u & A,G, Depicker, J 9X9, Different 3' end rcgions strongly influence!he level of gene expression in plant cells, Plant Cell I
671-680. Ingelbrecht. I" H. Van Houdl. A. Oepieker & M, Van Montagu,
1994. POSl-transcriptional silencing of reporter genes in tobacco correlated wi!h DNA me!hylnlion. Proc. Nall. Acad. Sci. USA. 91: 10502- 10506.
Jefferson. R,A,. 1987. Assaying chimeric genes in plants: !he GUS gene fusion system, Plant Mol. Biol. Rep. 5: 387-405,
Jobling, S,A. & L. Gehrke, 1987. Enhanced translation ofchimaeric rncssenger RNAs containing a plant viral untranslated leader sequence, Nature 325: 622-625.
Joncs, J.D.G" P Duosmuir & J. Bedbrook. 1985, High level expression of introdllced chirnaeric genes in regencrated Iransforrned planLs. EMBO J, 4: 241 J -2418,
Joshi. CP" 1987, An inspection of !he domain belween pUlative TATA box and translalion start silc in 79 plant genes. Nucl. Acid, Res. 15: 6643-6653,
Kozak. M., 1989 The scanning model for Iranslation: an update. J. Cell Biol. 108: 229-241.
Nicolaisen, M.. E, Johansen, G.B, Poulsen & B. Borkhardl. 1992. 'nll~ 5' ulllranslalcd reg ion from pea seedborne Illosaic pot)'vil1JS RNA as a translalional enhancer in pea and tobacco protoplasts, FEBS Lelt. 303: 169-172,
Ow, O.W. J,D. Jacobs& S.H. Howell, 1987. Functional rcgionsof !he cauliflower mOsaie virus 35S RNA promotér détctlllined b~
use ofthc lircfly Iliciferase gene as a reporter of promoter activit). Proe Natl. Acad. Sci USA 84 4870-4874.
Sanders. PR .. J,A, Winter, A.R. Barnason, S.G, Rogers & R.T. Fralcy. 1987. Comparison of cauliflower mosaic virus 35S and nopalinc syn!hase promo Iers in transgenic plan". Nucl. Acids Res. 15 1543- 1558,
Sieat. D.E., D,R, Gallie. R.A, Jcfferson, M.W. Bevan, PC Turner & TM .A, Wiison, 1987, Characlerisalion of !he 5' -leader seqllencl' oftobacco rnosaic virus RNA as a general enhanccroftranslalicl! in virrn, Gene 60: 217-225.
Vancallneyt. G.. S, Rosahl & L. Willlllitzer, 1990, Translalabilil)' Dj
a plalll-mRNA strongly inllllcnces its accumulation in transgcnic plants. Nucl. Acids Res. 18: 2917-2921,
s
b SI
bI
tr
re af
Ir
Gei, cr In
br m;
du en cl!
of re~
In)
pi, tra or
a! bel tra me tra tax
bet triU




















