
Avivi, S. and Sudarsono. MEKANISME KETAHANAN TANAMAN TRANSGENIK Nicotiana benthamiana::cp PStV...
15
MEKANISME KETAHANAN TANAMAN TRANSGENIK Nicotiana benthamiana::cp PStV GENERASI T 1 DAN FENOMENA HETEROLOGOUS PROTECTION Sholeh Avivi Staf Pengajar Jurusan Budidaya Pertanian Fakultas Pertanian Universitas Jember Jl. Kalimantan, Jember 68121 e-mail: [email protected] Sudarsono Staf Pengajar Jurusan Budidaya Pertanian Fakultas Pertanian Institut Pertanian Bogor ABSTRACT Avivi, S. and Sudarsono. 2004. Transgenic Nicotiana benthamiana::cp PStV Resistance Mechanism of T 1 Generation and Heterologous Protection Phenomenon. J. Agrikultura 15(3):137-145. The aims of this research were to: (1) investigate the effectiveness of the 4 cp gene constructs (pBINRCP1, pBINRCP2, pBINRCP3, and pBINRCP4) to protect N. benthamiana T 1 against PStV infection; (2) investigate the resistance mechanism of transgenic N. benthamiana, and (3) investigate the heterologous protection phenomenon. To achieve those objectives, transgenic N. benthamiana plants were regenerated, PCR tested, segregation tested, and inoculated with PStV using biological analysis methods. The result showed that all of PStV construct gave the resistancy against PStV inoculation. The transgenic plants contained mostly one or two functional loci of nptII and the resistance phenotype observed in the T 0 generation was inherited by the T 1 progenies. We proposed that the resistant mechanism was RNA-mediated resistance and the heterologous protection pheno- menon was worked in our plant. Key words: cp PStV, resistance mechanism, heterologous protection ABSTRAK Tujuan penelitian ini adalah untuk: (i) mempelajari efektivitas 4 konstruksi gen cp PStV dalam menimbulkan reaksi ketahanan terhadap inokulasi PStV pada generasi T 1 ; (ii) mempelajari mekanisme ketahanan tanaman transgenik terhadap PStV; (iii) mempelajari fenomena heterologous protection. Untuk merealisasikan tujuan tersebut, beberapa tanaman transgenik N. benthamiana diregenerasikan hingga menghasilkan biji T 1 dan dievaluasi ketahanannya terhadap PStV, diuji tingkat segregasinya dan jumlah lokus fungsional gennya. Juga dihipotesakan mekanisme ketahanan tanaman transgenik terhadap PStV dan dipelajari adanya fenomena heterologous protection. Dari hasil pengujian menunjukkan beberapa
Transcript of Avivi, S. and Sudarsono. MEKANISME KETAHANAN TANAMAN TRANSGENIK Nicotiana benthamiana::cp PStV...

MEKANISME KETAHANAN TANAMAN
TRANSGENIK Nicotiana benthamiana::cp PStV GENERASI T1
DAN FENOMENA HETEROLOGOUS PROTECTION
Sholeh Avivi
Staf Pengajar Jurusan Budidaya Pertanian
Fakultas Pertanian Universitas Jember
Jl. Kalimantan, Jember 68121
e-mail: [email protected]
Sudarsono Staf Pengajar Jurusan Budidaya Pertanian
Fakultas Pertanian Institut Pertanian Bogor
ABSTRACT
Avivi, S. and Sudarsono. 2004. Transgenic Nicotiana benthamiana::cp PStV
Resistance Mechanism of T1 Generation and Heterologous Protection
Phenomenon. J. Agrikultura 15(3):137-145.
The aims of this research were to: (1) investigate the effectiveness of the 4 cp
gene constructs (pBINRCP1, pBINRCP2, pBINRCP3, and pBINRCP4) to protect N.
benthamiana T1 against PStV infection; (2) investigate the resistance mechanism of
transgenic N. benthamiana, and (3) investigate the heterologous protection
phenomenon. To achieve those objectives, transgenic N. benthamiana plants were
regenerated, PCR tested, segregation tested, and inoculated with PStV using
biological analysis methods. The result showed that all of PStV construct gave the
resistancy against PStV inoculation. The transgenic plants contained mostly one or
two functional loci of nptII and the resistance phenotype observed in the T0
generation was inherited by the T1 progenies. We proposed that the resistant
mechanism was RNA-mediated resistance and the heterologous protection pheno-
menon was worked in our plant.
Key words: cp PStV, resistance mechanism, heterologous protection
ABSTRAK
Tujuan penelitian ini adalah untuk: (i) mempelajari efektivitas 4 konstruksi
gen cp PStV dalam menimbulkan reaksi ketahanan terhadap inokulasi PStV pada
generasi T1; (ii) mempelajari mekanisme ketahanan tanaman transgenik terhadap
PStV; (iii) mempelajari fenomena heterologous protection. Untuk merealisasikan
tujuan tersebut, beberapa tanaman transgenik N. benthamiana diregenerasikan
hingga menghasilkan biji T1 dan dievaluasi ketahanannya terhadap PStV, diuji
tingkat segregasinya dan jumlah lokus fungsional gennya. Juga dihipotesakan
mekanisme ketahanan tanaman transgenik terhadap PStV dan dipelajari adanya
fenomena heterologous protection. Dari hasil pengujian menunjukkan beberapa

2
tanaman transgenik mengandung 1-2 lokus fungsional dan setiap tipe konstruksi gen
dapat menghasilkan ketahanan terhadap inokulasi PStV hingga turunan T1.
Sedangkan mekanisme ketahanannya diduga disebabkan akibat RNA-mediated
resistance dan tanaman yang mengandung gen cp tipe pBINRCP4 dapat tahan
terhadap infeksi PVY, dengan demikian diduga kuat terdapat fenomena heterologous
protection.
Kata kunci: cp PStV, mekanisme resistensi, proteksi heterologous
PENDAHULUAN
Penyakit belang pada kacang tanah yang disebabkan oleh PStV (peanut stripe
virus) merupakan penyakit yang dominan dan tersebar di pusat-pusat produksi
kacang tanah di Indonesia. Tingkat serangan PStV di lapang dapat mencapai 100%
dan daerah sebaran penyakit ini diketahui sangat luas (Saleh & Baliadi 1992).
Serangan PStV dapat menurunkan produksi hingga 30-60% (Indonesia), 23% (RRC),
dan 67% (Filipina). Di Indonesia, serangan penyakit PStV dapat menurunkan
produksi antara 30-60%, sementara di RRC 23% dan di Filipina 67% (Natural et al.
1998; Saleh & Baliadi 1992; Sudarsono et al. 1997; Xu et al. 1990). Oleh karena itu
pencegahan terhadap PStV pada tanaman kacang tanah sangat perlu dilakukan.
Terdapat tiga pendekatan yang dapat dilakukan untuk mencegah kehilangan
hasil akibat serangan virus, yaitu (1) penghilangan sumber inokulum virus dengan
cara menggunakan benih atau bibit bebas virus dan penghilangan tanaman yang
terinfeksi di lapang, (2) pencegahan penyebaran virus, dan (3) penggunaan varietas
tanaman yang tahan terhadap virus. Pendekatan ketiga ini mempunyai beberapa
keuntungan dibandingkan dua pendekatan yang lain karena biaya penerapannya lebih
ekonomis, tidak mempunyai dampak ekologi yang negatif seperti halnya penggunaan
pestisida, dan dapat dengan efektif mengendalikan virus (Hull, 1990).
Untuk memperoleh varietas kacang tanah yang tahan terhadap PStV, teknik
rekayasa genetika dengan cara mengintroduksikan gen ketahanan ke dalam tanaman
merupakan salah satu alternatif yang layak dipilih. Gen-gen ketahanan terhadap virus
yang dapat menghasilkan tanaman yang resisten terhadap virus antara lain adalah gen
penyandi satelit RNA, antisense RNA, dan gen coat protein (selubung protein) dari
virus yang bersangkutan (gen cp) serta gen replikase virus (Hull, 1990; Hammond,
1998). Menurut beberapa peneliti terdapat hubungan yang positif antara ketahanan
yang didapat dari gen cp dengan tingkat kesamaan sekuen asam amino antara

3
transgen CP dan CP virus yang menginfeksi (Beachy, 1990; Lindbo, et al. 1993;
Kumagai et al. 1995).
Dengan alasan tersebut gen cp sampai saat ini lebih disukai oleh para peneliti
dibandingkan sekuen satelit RNA dan antisens RNA. Dalam penelitian ini juga
digunakan gen cp yang berasal dari virus PStV isolat Malang untuk mendapatkan
tanaman transgenik yang tahan PStV. Gen cp tersebut dikonstruksi pada 4 plasmid
yaitu: pBINRCP1, pBINRCP2, pBINRCP3, dan pBINRCP4 (Tabel 1).
Tabel 1. Sifat dan pola ekspresi empat tipe gen cp PStV yang diuji keefektifannya
dan tujuan pengujian yang dilakukan untuk mendapatkan tanaman
transgenik tahan PStV dan mempelajari mekanisme ketahanannya.
Tipe gen cp
PStV
Ekspresi gen cp
Tujuan pengujian Akumulasi
mRNA
Akumulasi
coat protein
pBINRCP1 + +
Menguji apakah ketahanan terjadi akibat
akumulasi CP
pBINRCP2 + + Jika ketahanan terjadi akibat akumulasi
CP, tipe ini mencegah penularan oleh
kutu daun
pBINRCP3 + -
Menguji apakah ketahanan terjadi akibat
akumulasi mRNA. Tipe ini merupakan
alternatif yang lebih baik bagi konsumen
pBINRCP4 + +
(lebih kecil
dari CP
normal)
Menguji apakah ketahanan terjadi akibat
akumulasi CP yang terkonservasi untuk
semua potyvirus. Berpotensi untuk
memproteksi terhadap berbagai anggota
potyvirus yang berbeda hanya dengan
satu tipe gen cp.
Selain itu, beberapa peneliti (Anandalakshmi et al. 1998; Angel &
Baulcombe 1997; Frank et al. 1999) mengemukakan hipotesis bahwa gen coat
protein tertentu kemungkinan dapat dipakai untuk melindungi tanaman transgenik
dari beberapa jenis virus yang berbeda (heterologous protection), asal susunan asam
nukleat dari gen cp yang digunakan mempunyai tingkat kesamaan yang tinggi .
Tanaman kacang tanah dapat diinfeksi oleh berbagai virus yang tergolong ke
dalam genus Potyvirus (a.l. PStV, PVY, PmoV). Sebagian besar gen cp dari virus
yang merupakan anggota Potyvirus mempunyai tingkat kesamaan sekuensi asam
nukleat yang tinggi di bagian tengahnya (hingga 70%). Penggunaan gen cp bagian ini

4
diduga akan dapat dipakai untuk melindungi tanaman transgenik dari berbagai virus
tersebut.
Dalam penelitian ini, digunakan tipe gen pBINRCP4 yang merupakan bagian
gen cp yang tingkat kesamaannya sangat tinggi untuk berbagai Potyvirus. Apabila
dengan menggunakan pBINRCP4 ini ditemukan tanaman transgenik yang tahan
PStV maka kemungkinan juga akan tahan terhadap Potyvirus yang lain. Jika hal ini
benar maka akan diperoleh bukti yang mendukung hipotesis tentang adanya
heterologous protection. Dengan demikian tipe gen pBINRCP4 akan dapat dipakai
untuk melindungi tanaman transgenik dari serangan berbagai potyvirus. Dalam
percobaan ini, selain PStV, potyvirus lain yang digunakan adalah PVY.
BAHAN DAN METODE
Penelitian dilakukan di Lab. Biologi Molekuler Tanaman dan di kebun
percobaan, Jurusan Budidaya Pertanian, Fakultas Pertanian, IPB. Penelitian mulai
dilakukan sejak bulan September 1997 sampai dengan bulan Februari 2000. Metode
regenerasi tanaman transgenik model dilakukan dengan cara standar seperti yang
telah dilakukan oleh Avivi (2000). Isolat PStV yang digunakan diisolasi dari kacang
tanah kultivar Kelinci yang terserang PStV di kebun percobaan Sawah Baru,
Darmaga, IPB.
Uji Segregasi dan Percobaan Inokulasi Tanaman T1. Biji T1 dikecambahkan
dalam media MSO yang mengandung kanamycin (50 mg.L-1
). Segregasi antara
kecambah yang tahan (KanR) dan yang rentan kanamycin (Kan
r) dicatat untuk
menentuan segregasi gen nptII yang ada pada tanaman transgeniknya. Segregasi gen
nptII diharapkan sama dengan segregasi gen cp PStV, mengingat kedua gen ini
berada pada alur konstruksi yang sama. Oleh karenanya, adanya gen nptII dapat
dipakai sebagai prediktor adanya gen cp PStV.
Kecambah yang KanR selanjutnya dipindahkan (transplanting) ke media
tanah untuk pengujian ketahanan terhadap infeksi PStV. Tanaman transgenik T1
diinokulasi dengan PStV. Inokulasi pertama dilakukan pada 4 minggu setelah tanam
(MST) dan inokulasi kedua dilakukan pada 5 MST. Pengamatan terhadap tunas
transgenik yang telah diinokulasi dilakukan antara 7-14 hari setelah diinokulasi.
Gejala serangan PStV pada tanaman N. benthamiana berupa mosaik ringan sistemik

5
pada daun. Hasil pengujian kemudian dipisahkan ke dalam kelompok tanaman yang
rentan, penyembuhan (recovery) dan tahan terhadap infeksi PStV. Hasil pengujian
ini kemudian digunakan sebagai indikasi dapat tidaknya sifat tahan PStV tersebut
diturunkan secara seksual.
Uji Integrasi Gen cp dengan PCR. Deteksi gen cp dalam genom tanaman
dilakukan dengan PCR seperti yang dilakukan oleh Thomson & Dietzgen (1995),
tetapi menggunakan primer dengan sekuen sebagai berikut: PST1: 5’-GCATGCCCT
CGCCATTGCAA-3’ dan PST2: 5’-GCACACACTTCTTGGCATGG-3’. Produk
yang dihasilkan oleh primer PST1 dan PST2 berukuran 234 bp.
Uji mekanisme Heterologous Protection terhadap Potyvirus selain PStV. Hal ini
dilakukan untuk menguji adanya fenomena heterologous protection dengan melihat
apakah tanaman transgenik yang membawa tipe gen pBINRCP4 (gen cp yang tingkat
kesamaan sekuensi asam nukleatnya sangat tinggi dengan gen cp potyvirus lainnya)
akan dapat melindungi tanaman model dari infeksi PStV dan dari potyvirus lainnya.
Jika hal ini benar maka tipe gen pBINRCP4 dapat dipakai untuk mendapatkan
tanaman yang tahan terhadap infeksi berbagai anggota dari group potyvirus.
Pengujian dilakukan dengan cara sebagai berikut: contoh tanaman transgenik yang
membawa tipe gen pBINRCP4 diinokulasi dengan PStV dan PVY secara mekanik.
Pada tanaman transgenik yang diinokulasi diamati ada tidaknya gejala serangan
PStV dan PVY sebagaimana dalam pengujian dengan virus PStV. Hasil pengujian
akan menghasilkan kelompok tanaman yang rentan, penyembuhan dan tahan
terhadap infeksi PStV dan PVY.
HASIL DAN PEMBAHASAN
Dengan sistem transformasi dan regenerasi yang digunakan dalam percobaan
ini diperoleh tanaman transgenik yang tumbuh dan berkembang hingga
menghasilkan biji dengan tingkat keberhasilan sekitar 50% (data tidak ditunjukkan).
Segregasi Gen nptII Tanaman T1 dan Jumlah Lokus Gen. Tanaman transgenik
T0 yang telah diuji tahan terhadap inokulasi PStV ditumbuhkan hingga diperoleh biji
T1. Dari klon yang menghasilkan biji T1 ini dipilih beberapa klon yang mewakili
masing-masing tipe gen untuk diuji segregasi gen nptII-nya. dan jumlah lokus
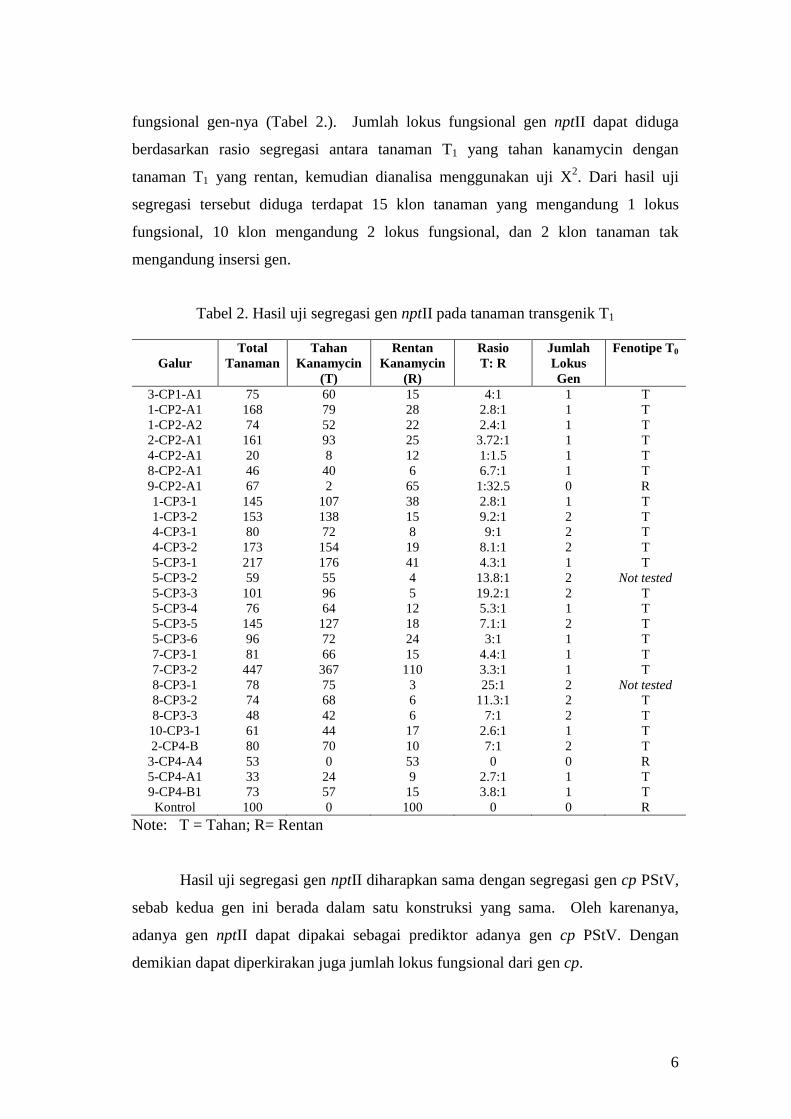
6
fungsional gen-nya (Tabel 2.). Jumlah lokus fungsional gen nptII dapat diduga
berdasarkan rasio segregasi antara tanaman T1 yang tahan kanamycin dengan
tanaman T1 yang rentan, kemudian dianalisa menggunakan uji X2. Dari hasil uji
segregasi tersebut diduga terdapat 15 klon tanaman yang mengandung 1 lokus
fungsional, 10 klon mengandung 2 lokus fungsional, dan 2 klon tanaman tak
mengandung insersi gen.
Tabel 2. Hasil uji segregasi gen nptII pada tanaman transgenik T1
Galur
Total
Tanaman
Tahan
Kanamycin
(T)
Rentan
Kanamycin
(R)
Rasio
T: R
Jumlah
Lokus
Gen
Fenotipe T0
3-CP1-A1 75 60 15 4:1 1 T
1-CP2-A1 168 79 28 2.8:1 1 T
1-CP2-A2 74 52 22 2.4:1 1 T
2-CP2-A1 161 93 25 3.72:1 1 T
4-CP2-A1 20 8 12 1:1.5 1 T
8-CP2-A1 46 40 6 6.7:1 1 T
9-CP2-A1 67 2 65 1:32.5 0 R
1-CP3-1 145 107 38 2.8:1 1 T
1-CP3-2 153 138 15 9.2:1 2 T
4-CP3-1 80 72 8 9:1 2 T
4-CP3-2 173 154 19 8.1:1 2 T
5-CP3-1 217 176 41 4.3:1 1 T
5-CP3-2 59 55 4 13.8:1 2 Not tested
5-CP3-3 101 96 5 19.2:1 2 T
5-CP3-4 76 64 12 5.3:1 1 T
5-CP3-5 145 127 18 7.1:1 2 T
5-CP3-6 96 72 24 3:1 1 T
7-CP3-1 81 66 15 4.4:1 1 T
7-CP3-2 447 367 110 3.3:1 1 T
8-CP3-1 78 75 3 25:1 2 Not tested
8-CP3-2 74 68 6 11.3:1 2 T
8-CP3-3 48 42 6 7:1 2 T
10-CP3-1 61 44 17 2.6:1 1 T
2-CP4-B 80 70 10 7:1 2 T
3-CP4-A4 53 0 53 0 0 R
5-CP4-A1 33 24 9 2.7:1 1 T
9-CP4-B1 73 57 15 3.8:1 1 T
Kontrol 100 0 100 0 0 R
Note: T = Tahan; R= Rentan
Hasil uji segregasi gen nptII diharapkan sama dengan segregasi gen cp PStV,
sebab kedua gen ini berada dalam satu konstruksi yang sama. Oleh karenanya,
adanya gen nptII dapat dipakai sebagai prediktor adanya gen cp PStV. Dengan
demikian dapat diperkirakan juga jumlah lokus fungsional dari gen cp.

7
Tabel 3. Klon-klon transgenik T1 yang diuji sifat ketahanannya terhadap PStV Galur #Tanaman Gejala
sistematik
Lesio lokal pada C. amaranticolor Sifat
ketahanan 8 MST 10 MST 12 MST
3-CP1-A1 1 - + - - P
2 - + + - P
3 - - - - T
4 - + + + R
6 + + - - P
7 + - - - T
8-CP2-A1 1 - + + - P
2 - + - - P
4 - - - - T
5 + - - - T
7 + - - - T
8 - - - - T
9-CP2-A1 1 + + + + R
1-CP3-2 1 + + - - P
4 - + + + R
5 + + - - P
6 - + - - P
9 + + + + R
10 - - - - T
5-CP3-6 1 + + + + R
4 - + + + R
5 - - - - T
7 + - - - T
7-CP3-1 1 - + + + R
2 - + - - P
3 + + - - P
4 - + - - P
7 - + - - P
8 - - - - T
9 - - - - T
10 + + + + R
11 + + - - P
12 + + - - P
13 - + + + R
8-CP3-3 1 - - - - T
5-CP4-A1 1 + - - - T
3 - - - - T
4 - + - - P
5 - - - - T
6 + + + + R
9-CP4-B1 1 - - - - T
2 + - - - T
5 + + + + R
6 + + + - P
8 + + - - P
Kontrol
Transformed
1-5 + + + + R
Non Transformed 1-7 + + + + R
Keterangan: T = Tahan; P = Penyembuhan; R = Rentan; + = ada gejala lesiolokal;
- = tak bergejala
Pengujian Ketahanan Tanaman Transgenik T1 Terhadap PStV. Untuk
mengetahui apakah sifat ketahanan terhadap PStV diturunkan secara seksual atau
tidak, tanaman T0 yang tahan PStV, keturunan T1-nya ditanam dan diuji dengan cara
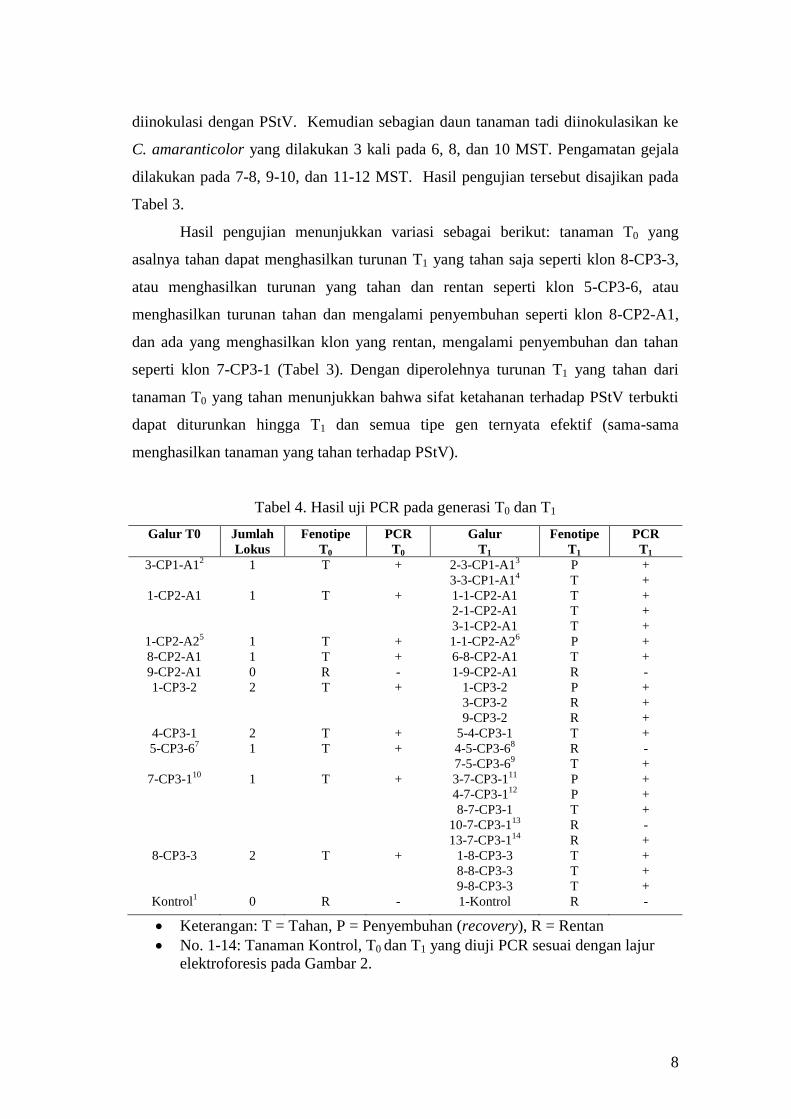
8
diinokulasi dengan PStV. Kemudian sebagian daun tanaman tadi diinokulasikan ke
C. amaranticolor yang dilakukan 3 kali pada 6, 8, dan 10 MST. Pengamatan gejala
dilakukan pada 7-8, 9-10, dan 11-12 MST. Hasil pengujian tersebut disajikan pada
Tabel 3.
Hasil pengujian menunjukkan variasi sebagai berikut: tanaman T0 yang
asalnya tahan dapat menghasilkan turunan T1 yang tahan saja seperti klon 8-CP3-3,
atau menghasilkan turunan yang tahan dan rentan seperti klon 5-CP3-6, atau
menghasilkan turunan tahan dan mengalami penyembuhan seperti klon 8-CP2-A1,
dan ada yang menghasilkan klon yang rentan, mengalami penyembuhan dan tahan
seperti klon 7-CP3-1 (Tabel 3). Dengan diperolehnya turunan T1 yang tahan dari
tanaman T0 yang tahan menunjukkan bahwa sifat ketahanan terhadap PStV terbukti
dapat diturunkan hingga T1 dan semua tipe gen ternyata efektif (sama-sama
menghasilkan tanaman yang tahan terhadap PStV).
Tabel 4. Hasil uji PCR pada generasi T0 dan T1
Galur T0 Jumlah
Lokus
Fenotipe
T0
PCR
T0
Galur
T1
Fenotipe
T1
PCR
T1
3-CP1-A12 1 T + 2-3-CP1-A1
3 P +
3-3-CP1-A14
T +
1-CP2-A1
1
T
+
1-1-CP2-A1 T +
2-1-CP2-A1 T +
3-1-CP2-A1 T +
1-CP2-A25 1 T + 1-1-CP2-A2
6 P +
8-CP2-A1 1 T + 6-8-CP2-A1 T +
9-CP2-A1 0 R - 1-9-CP2-A1 R -
1-CP3-2
2
T
+
1-CP3-2 P +
3-CP3-2 R +
9-CP3-2 R +
4-CP3-1 2 T + 5-4-CP3-1 T +
5-CP3-67
1 T + 4-5-CP3-68 R -
7-5-CP3-69 T +
7-CP3-110
1 T + 3-7-CP3-111
P +
4-7-CP3-112
P +
8-7-CP3-1 T +
10-7-CP3-113
R -
13-7-CP3-114
R +
8-CP3-3
2 T + 1-8-CP3-3 T +
8-8-CP3-3 T +
9-8-CP3-3 T +
Kontrol1 0 R - 1-Kontrol R -
Keterangan: T = Tahan, P = Penyembuhan (recovery), R = Rentan
No. 1-14: Tanaman Kontrol, T0 dan T1 yang diuji PCR sesuai dengan lajur
elektroforesis pada Gambar 2.

9
Seperti yang telah dikemukakan bahwa jika rasio lokus fungsional dari gen
nptII dapat diketahui, maka akan dapat diduga rasio lokus fungsional dari gen cp
PStV. Pendugaan lokus fungsional gen cp PStV dilakukan dengan melihat nilai rasio
tanaman T1 yang tahan plus tanaman T1 yang mengalami penyembuhan dengan
tanaman T1 yang rentan terhadap PStV, kemudian nilai tersebut dibandingkan
dengan jumlah lokus fungsional gen nptII.
Hasil Uji Integrasi dengan Teknik PCR. Hasil PCR terhadap daun tanaman
transgenik dari sebagian populasi T0 dan T1 menggunakan primer PST1 dan PST2
disajikan pada Tabel 4. Dari hasil PCR tersebut diperoleh beberapa klon tanaman
yang positif mengandung gen cp tetapi tidak memiliki sifat ketahanan terhadap PStV
seperti klon 3-CP3-2 dan 9-CP3-2. Fenomena demikian menarik untuk didiskusikan.
Terdapat beberapa kemungkinan yang dapat menjelaskan hal tersebut misalnya (1)
terjadi mutasi pada satu basa pada titik awal transripsi, sehingga enzim polimerase
tak dapat mengawali proses transkripsi, akibatnya mRNA tidak dapat terbentuk, (2)
terjadi mutasi pada faktor sigma sehingga faktor sigma ini tidak mampu membawa
enzim polimerase mengenali promotor, akibatnya mRNA tidak dapat terbentuk tapi
tidak sempurna, (3) mRNA transgen mengalami proses silencing, dan masih banyak
lagi kemungkinan yang lain.
Gambar 2. Hasil uji elektroforesis pada tanaman transgenik T1
234 bp
(-)
Co
ntr
ol
3-C
P1-
A1
3-C
P1-
A1-
R1
3-C
P1-
A1-
R1
1-C
P2-
A1
1-C
P2-
A1-
R1
5-C
P3-
6
5-C
P3-
6-R
1
5-C
P3-
6-R
1
7-C
P3-
1
7-C
P3-
1-R
1
7-C
P3-
1 -R
1
7-C
P3-
1 -R
1
7-C
P3-
1 -R
1
1 K
b L
add
er
Pla
smid
pC
P1

10
Mekanisme Ketahanan Terhadap PStV (Suatu Hipotesa). Dari keempat tipe
konstruksi yang digunakan apabila ditinjau dari proses ekspresi gennya mempunyai
dua perbedaan pokok yaitu pBINRCP1, pBINRCP3, dan pBINRCP4 dapat
membentuk mRNA dan coat protein. Sedangkan pBINRCP2 tidak membentuk coat
protein, hanya dapat membentuk mRNA saja, disebabkan adanya stop codon pada
awal ORF-nya (Tabel 1).
Dengan menggunakan empat konstruksi gen tersebut dapat kita buat hipotesis
tentang konsep mekanisme ketahanan terhadap PStV. Hipotesisnya dapat dibuat
sebagai berikut:
1) Jika sifat ketahanan hanya dimiliki oleh tanaman transgenik yang memiliki
konstruksi gen yang mampu mengekspresikan coat protein maka dapat diduga
bahwa ketahanan terhadap PStV memerlukan adanya coat protein.
2) Jika sifat ketahanan ternyata dapat diperoleh baik pada tanaman yang diduga
memiliki konstruksi gen yang mampu mengekspresikan coat protein maupun
tanaman yang memiliki konstruksi gen yang mampu mengekspresikan mRNA
saja, maka dapat diduga bahwa ketahanan terhadap PStV tidak memerlukan
adanya coat protein namun cukup hanya dengan mRNA saja.
Berdasarkan hasil penelitian ini hipotesis kedua yang diterima, sebab dari
sejumlah tanaman transgenik yang mengandung keempat tipe gen cp, ternyata
diperoleh bahwa ketahanan terhadap infeksi virus dapat terjadi pada semua tipe
tanaman transgenik. Baik tanaman transgenik tersebut mampu mengekspresikan CP
(tipe konstruksi pBINRCP1, pBINRCP3, dan pBINRCP4) maupun hanya sampai
tingkat mRNA saja (tipe konstruksi pBINRCP2). Bila asumsi-asumsi di atas benar,
berarti dapat diambil suatu kesimpulan bahwa untuk memperoleh ketahanan terhadap
inokulasi PStV, tanaman tidak perlu sampai mengekspresikan CP namun cukup
sampai tingkat mRNA saja.
Sejauh ini terdapat dua hipotesis yang berbeda dalam usaha untuk
menjelaskan mekanisme terhadap virus pada tanaman transgenik yang membawa gen
cp. Kedua hipotesis tersebut sama-sama didukung oleh data kuantitatif hasil
rangkaian percobaan dan juga didukung oleh berbagai penelitian yang dilakukan oleh
banyak peneliti (Angel & Baulcombe 1997; Ruiz et al. 1998; Anandalakshmi et al.
1998; Frank et al. 1999).

11
Hipotesis pertama menyatakan bahwa ketahanan terhadap virus pada tanaman
terjadi akibat adanya akumulasi coat protein (coat protein-mediated resistance) pada
sel tanaman transgeniknya sebelum tanaman terinfeksi oleh virus (Anandalakshmi et
al. 1998; Brigneti et al. 1998; Kasschau & Carrington 1998). Akumulasi protein pada
sel tanaman transgenik diduga menghambat proses replikasi virus yang menyerang
sehingga tanaman yang terinfeksi tidak mengalami kerusakan seberat tanaman
normal (non-transgenik).
Dalam hal ini, gen yang diintroduksikan harus dapat mengekspresikan coat
protein di dalam sel tanaman dan tanaman transgeniknya harus mengakumulasikan
sejumlah tertentu dari coat proteinnya agar menjadi tahan virus. Kelompok hipotesis
ini didukung oleh hasil penelitian yang menggunakan virus PMTV (potato mop-top
virus) dan berbagai virus yang tergolong dalam genus Tobamovirus, seperti TMV
(tobacco mosaic virus) dan ToMV (Tomato mosaic virus).
Sebaliknya hipotesis yang kedua menyatakan bahwa ketahanan terhadap
virus pada tanaman transgenik tidak memerlukan adanya akumulasi coat protein
tetapi hanya memerlukan adanya akumulasi mRNA dari gen cp (RNA-mediated
resistance). Adanya transkripsi mRNA dari gen cp dalam jumlah yang sangat tinggi
diduga mengaktifkan mekanisme degradasi mRNA melalui proses degradasi RNA
(RNA turn-over) yang disebut sebagai fenomena gene silencing (Angel &
Baulcombe, 1997; Al-Kaff et al. 1998; Atkinson, et al. 1998; Kjemtrup et al. 1998;
Ruiz et al. 1998). Akibat dari proses ini ketika virus menginfeksi tanaman, genom
RNA virusnya (yang juga mempunyai sekuensi homolog dengan gen cp) juga akan
didegradasi melalui mekanisme yang sama sehingga proses infeksi menjadi
terhambat dan tanaman tidak mengalami kerusakan.
Dalam hal ini, gen yang diintroduksikan harus dapat mengespresikan mRNA
dari gen cp dalam jumah banyak sehingga mengaktifkan fenomena gene silencing
dan menyebabkan aktifnya mekanisme degradasi RNA yang dihasilan oleh gen cp
dan RNA yang sejenisnya (genom dari virus yang menginfeksi). Hipotesis ini
didukung hasil penelitian yang menggunakan berbagai virus yang tergolong dalam
genus Potyvirus, seperti PVY (potato virus Y), PVX (potato virus X) dan TEV
(tobacco etch virus). Mekanisme yang sama juga dilaporkan terjadi pada
Geminivirus.

12
Jadi dapat diperkirakan bahwa ketahanan tanaman transgenik terhadap PStV
berhubungan erat dengan adanya akumulasi mRNA dalam tanaman transgenik
(RNA-mediated resistance) dan tidak memerlukan adanya akumulasi coat protein.
Namun demikian kesimpulan mengenai mekanisme ketahanan ini masih
perlu dipastikan dan didukung dangan data-data analisis molekuler yang benar-benar
dapat menunjukkan bahwa tanaman transgenik dengan tipe gen yang digunakan di
atas dapat mengekspresikan coat protein ataukah hanya sampai tingkat mRNA saja.
Uji fenomena Heterologous Protection terhadap Potyvirus selain PStV. Hasil uji
masing-masing klon pBINRCP4 yang diperoleh dan sifat ketahanannya setelah
diinokulasi dengan PStV dan PVY serta setelah diinokulasikan ke C. amaranticolor
disajikan pada Tabel 5.
Tabel 5. Respon tanaman N.benthamiana transgenik pBINRCP4 terhadap inokulasi
PStV dan PVY
Inokulasi &
tanaman yg diuji
Jumlah
Galur
Respon fenotipe
Tahan % Penyembuhan % Rentan %
PStV:
Transgenik CP4
34
20
58.8
6
17.7
8
23.5
Kontrol
Transgenik
5
0
0
0
0
5
100
Kontrol non
Transgenik
10
0
0
0
0
10
100
PVY:
Transgenik CP4
15
12
80.0
0
0
3
20.0
Kontrol
Transgenik
5
0
0
0
0
5
100
Kontrol non
Transgenik
5
0
0
0
0
5
100
Berdasarkan data Tabel 5 dari sebanyak 34 tanaman transgenik yang sudah
diuji dengan PStV diperoleh tanaman yang tahan, mengalami penyembuhan dan
rentan PStV, masing-masing sebagai berikut: 20 tanaman transgenik tahan, 6
tanaman transgenik mengalami penyembuhan dan 8 rentan PStV. Sedangkan dari
sebanyak 15 tanaman transgenik yang sudah diuji dengan PVY diperoleh beberapa
tanaman yang tahan dan rentan PVY, masing-masing sebagai berikut: 12 tanaman
transgenik tahan dan 3 rentan PVY.

13
Seperti yang telah dikemukakan di atas bahwa terdapat hipotesis yang
mengatakan bahwa suatu potongan gen coat protein kemungkinan dapat dipakai
untuk melindungi tanaman transgenik dari beberapa jenis virus yang berbeda
(heterologous protection), asal sekuensi asam nukleat dari gen cp yang digunakan
mempunyai tingkat kesamaan yang tinggi. Dalam penelitian ini, digunakan tipe gen
pBINRCP4 yang merupakan bagian gen cp yang tingkat kesamaan nukleotida-nya
sangat tinggi untuk berbagai Potyvirus. Karena terbukti dapat diperoleh juga tanaman
yang tahan terhadap infeksi PVY maka dapat dibuktikan bahwa hipotesis tentang
adanya heterologous protection adalah benar.
KESIMPULAN DAN SARAN
Kesimpulan
Tanaman transgenik yang diuji mengandung 1-2 lokus fungsional dan setiap
tipe konstruksi gen cp cp dapat menghasilkan ketahanan terhadap inokulasi PStV
hingga turunan T1. Sedangkan mekanisme ketahanannya diduga akibat RNA-
mediated resistance. Karena tanaman yang mengandung gen cp tipe pBINRCP4
dapat tahan terhadap infeksi PVY, dengan demikian penelitian ini membuktikan
terjadinya fenomena heterologous protection.
Saran
Masih diperlukan data-data moleuler seperti hasil uji ELISA, oligo dt
coloumn, atau run on RNA assay untuk data pendukung mekanisme resistensi
tanaman transgenik terhadap serangan PStV.
UCAPAN TERIMA KASIH
Terimakasih diucapkan kepada lembaga pemberi dana untuk penelitian ini:
SEARCA Thesis grant, Los Banos, Philippine (cq. Sholeh Avivi), Sandwich program
batch II, URGE Project, MOEC, Indonesia (cq. Dr. Sudarsono). Kepada Erna
Rochiyati juga diucapkan terimakasih atas perannya sebagai penyelaras bahasa.

14
DAFTAR PUSTAKA
Avivi, S. 2000. Berbagai tipe konstruksi gen cp PStV yang dapat memproteksi
tanaman Nicotiana benthamiana transgenik terhadap infeksi PStV dan
transformasi gen cp PStV pada kacang tanah. Disertasi. IPB. Bogor. 173p.
Al-Kaff, N.S. S.N. Covey, M.M Kreike, A.M. Page, R. Pinder, and P.J. Dale, 1998.
Transcriptional and post-transcriptional plant gene silencing in response to a
pathogen. Science, 279:2113-2115.
Anandalakshmi, R., G.J. Pruss, X. Ge, R. Marathe T.H. Smith, and V.B. Vance.
1998. A viral suppressor of gene silencing in plant. Proc. Natl. Acad. Sci.
USA. 95:13079-13084.
Angel, S.M. and D.C. Baulcombe. 1997. Consistent gene silencing in transgenic
plants expressing a replicating potato virus X RNA. EMBO J. 16:3675-3684.
Atkinson, R.G., L.R.F. Bieleski, A.P. Gleave, B.J. Jannsen, and B.A.M. Morris.
1998. Post-trnascriptional silencing of chalcone synthase in petunia using a
geminivirus-based episoma vector. Plant J. 15:593-604.
Beachy, RN. 1990. Coat protein Mediated resistance in transgenic plants. p 13-22, In
T.P. Pirone and J.G. shaw (eds). Viral Genes and Plant Pathogenesis.
Springer-Verlag. New York. 215p.
Brigneti, G.,O. Voinnet, L. Wan-Xiang J. Liang-Hui, S.W. Ding, and D.C.
Baulcombe. 1998. Viral pathogenicity determinant are suppresors of
transgene siencing in Nicotiana bethamiana. EMBO J. 17:6739-6746.
Frank G.F., A. Stuart., MacFarlane, and D.C. Baulcombe. 1999. Gene silencing
without DNA: RNA-mediated cross-protection between viruses. The Plant
Cell. 11:1207-1215.
Hammond, J. 1998. Resistance to plant viruses–an overview. p163-171. In A. Hadidi
R.K. Khetarpa, and H. oganezawa (eds.). Plant Virus Disease Control. APS
Press. The American Phytopathological Society. St. Paul, Minnesota.
Hull, R. 1990. Non-conventional resistance to viruses in pants concepts and ris.
P289-303. In J.P. Gustafson (ed.) Gene Manipulation in Plant Improvement
II. Penum Press, New York.
Kasschou, K.D. and J.C. Carrington. 1998. A counter-defensive strategy of plant
viruses: Suppression of post-transcriptional gene silencing. Cell. 95:461-470.
Kjemtrup, S., K.S. Sampson. C.G. Peele. L.V. Nguyen, M.A. Conkling, W.F/
Thompson, and D. Robertson. 1998. Gene silencing from plant DNA carried
by a geminivirus plant J. 14:91-100.

15
Kumagai, M.H., J. Donson, G. Dela-Cioppa, D. Harvey, K. Hanley, and L.K. Grill.
1995. Cytoplasmic inhibition of carotenoid biosynthesis with virus-derived
RNA. Proc. Natl. Acad. Sci. USA. 92:1679-1683.
Lindbo, J. A., L. Silva-Rosales, W.M. Proebsting, and W.G. Dougherty. 1993.
Induction of a highly specific antiviral state in transgenic plants: Implications
for regulation of gene expression and virus resistance. Plant Cell. 5:1749-
1759.
Natural, M. P., F. L. Mangaban, and L. D. Valencia. 1989. Groundnut research in the
Phippines. Second Coordinators Meeting on Peanut Stripe Virus (PStV).
Agrikam 5(2):71-83.
Newton, TR. 1997. Agrobacterium mediated transformation of peanut. Honours
Thesis. The University of Queensland.
Ruiz, M.T., O. Voinnet, and D.C. Baulcombe. 1998. Initiation and maintenance of
virus-induced gene silencing. Plant Cell. 10:937-946.
Saleh, N and Y Baliadi. 1992. Penyakit virus bilur kacang tanah (peanut stripe virus)
dan usaha pengendaliannya. Balai Penelitian Tanaman Pangan. Malang 22p.
Sudarsono, S Tumbelaka, dan S. Ilyas. 1997. Penurunan hasil akibat peanut stripe
virus dan penularan virus lewat benih pada kacang tanah. Hayati:55-58.
Thomson, D. and R.G. Dietzgen. 1995. Detection of DNA and RNA plant viruses by
PCR and RT-PCR using a rapid virus release protocol without tissue
homogenization. J. Virological Methods 54:85-95.
Xu, Z., C. Kunrong Z. Zongyi, C. Jinxiang, and K.J. Middleton. 1990. Research on
peanut stripe virus (PStV) in China. 6p. (Unpublished).




















