Ubiquitous presence of β-glucuronidase (GUS) in plants and its regulation in some model plants
12
Planta (2006) 224: 853–864 DOI 10.1007/s00425-006-0276-2 ORIGINAL ARTICLE Charu Sudan · Shiva Prakash · Prasanna Bhomkar Shalu Jain · Neera Bhalla-Sarin Ubiquitous presence of -glucuronidase (GUS) in plants and its regulation in some model plants Received: 23 February 2006 / Accepted: 25 March 2006 / Published online: 28 April 2006 © Springer-Verlag 2006 Abstract The enzyme -glucuronidase (GUS) is well characterized in animals and microbes. However, this enzyme is not well studied in plants and is widely assumed to be absent in them. In this study we document the ubiq- uitous presence of GUS in the model plants Arabidopsis thaliana, Oryza sativa, Nicotiana tabacum and Zea mays and record its expression pattern. The pH of the assay buVer was found to be critical with pH 4.0 being optimum for detection in all the species. GUS in plants appears to be associated with growth. In general, younger regions of the organs showed more GUS activity than the older and more mature tissues. In Brassica juncea roots stained for GUS, intense blue color could be seen in the trichoblast cells and the growing root hair cells as compared to the non-root hair forming epidermal cells or the fully elon- gated root hairs. Cotton Wbers showed high GUS activity during the initial phase of elongation while the seed coat, from which the Wbers formed, did not stain for GUS activ- ity. The activity in the Wbers disappeared after they were fully elongated. The level of GUS activity increased 2.58 folds in leaf tissues of N. tabacum when cultured in MS medium supplemented with 6-benzylaminopurine, while gibberellic acid enhanced GUS activity 2.9 folds in the inter-nodal regions of rice in 12-h treatment. In addition, elongation of stem, root and root hairs in tobacco seed- lings was strongly inhibited by the speciWc inhibitor of GUS, saccharo-1-4-lactone in a reversible manner. Taken together, these evidences suggest a probable association of plant GUS in cell growth. Keywords Arabidopsis thaliana · -Glucuronidase · Oryza sativa · Nicotiana tabacum · Saccharo-1-4-lactone · Zea mays Abbreviations ABA: Abscisic acid · BAP: 6-Benzylaminopurine · GA 3 : Gibberellic acid · GUS: -Glucuronidase · MUG: 4-Methyl-umbelliferyl- beta-D-glucuronide · NAA: Naphthalene acetic acid · SL: Saccharo-1-4-lactone · X-gluc: 5-Bromo-4-chloro-3-indolyl glucuronide Introduction The enzyme -glucuronidase (GUS) has been widely stud- ied in micro-organisms and animals (reviewed by Fishman 1955). The GUS enzyme from E. coli has been well charac- terized and the GUS gene from the same organism is widely being used as a reporter in plant transformation experiments (JeVerson and Wilson 1991). The enzyme GUS is also known to be present in many bacterial species such as the enterobacterium Shigella, which is closely related to E. coli. Its activity was also observed in non- enterobacterial, anaerobic residents of the digestive tract, such as Bacteroides and Clostridium (Hawkesworth et al. 1971). Other bacterial species with GUS activity are Strep- tococcus, Staphylococcus, Corynebacterium (Levvy and Marsh 1959) and Alcaligenes (JeVerson 1989). In these bacteria, GUS plays a role in carbohydrate and energy metabolism. The GUS activity is very common in virtually all tissues of vertebrates where it is known to play a major role in the stepwise degradation of glycosaminoglucuro- nides and in the release of active hormones from steroid hormone–glucuronide conjugates (Kyle et al. 1992). The enzyme may also participate in the catabolism of polysac- charides containing glucuronic acid (Levvy and Marsh 1959). In the rat preputial gland, GUS is estimated to con- stitute as much as 5% of the total protein (Kyle et al. 1992). Its activity is also known to be present in organisms from invertebrate taxa (Koburger and Miller 1985) such as in snails (Levvy and Marsh 1959), nematodes (Sebasti- ano et al. 1986) and insects (Levvy and Marsh 1959; Lang- ley et al. 1983). GUS activity has also been detected in the houseXy (Musca domestica) and in various locusts (Levvy and Marsh 1959). Charu Sudan and Shiva Prakash, the Wrst two authors, have contributed equally. C. Sudan · S. Prakash · P. Bhomkar · S. Jain · N. Bhalla-Sarin (&) Lab no. 308, School of Life Sciences, JNU, New Delhi 110 067, India E-mail: [email protected] Fax: +91-11-5525397
Transcript of Ubiquitous presence of β-glucuronidase (GUS) in plants and its regulation in some model plants

Planta (2006) 224: 853–864 DOI 10.1007/s00425-006-0276-2
ORIGINAL ARTICLE
Charu Sudan · Shiva Prakash · Prasanna Bhomkar Shalu Jain · Neera Bhalla-Sarin
Ubiquitous presence of �-glucuronidase (GUS) in plants and its regulation in some model plants
Received: 23 February 2006 / Accepted: 25 March 2006 / Published online: 28 April 2006© Springer-Verlag 2006
Abstract The enzyme �-glucuronidase (GUS) is wellcharacterized in animals and microbes. However, thisenzyme is not well studied in plants and is widely assumedto be absent in them. In this study we document the ubiq-uitous presence of GUS in the model plants Arabidopsisthaliana, Oryza sativa, Nicotiana tabacum and Zea maysand record its expression pattern. The pH of the assaybuVer was found to be critical with pH 4.0 being optimumfor detection in all the species. GUS in plants appears tobe associated with growth. In general, younger regions ofthe organs showed more GUS activity than the older andmore mature tissues. In Brassica juncea roots stained forGUS, intense blue color could be seen in the trichoblastcells and the growing root hair cells as compared to thenon-root hair forming epidermal cells or the fully elon-gated root hairs. Cotton Wbers showed high GUS activityduring the initial phase of elongation while the seed coat,from which the Wbers formed, did not stain for GUS activ-ity. The activity in the Wbers disappeared after they werefully elongated. The level of GUS activity increased 2.58folds in leaf tissues of N. tabacum when cultured in MSmedium supplemented with 6-benzylaminopurine, whilegibberellic acid enhanced GUS activity 2.9 folds in theinter-nodal regions of rice in 12-h treatment. In addition,elongation of stem, root and root hairs in tobacco seed-lings was strongly inhibited by the speciWc inhibitor ofGUS, saccharo-1-4-lactone in a reversible manner. Takentogether, these evidences suggest a probable association ofplant GUS in cell growth.
Keywords Arabidopsis thaliana · �-Glucuronidase · Oryza sativa · Nicotiana tabacum · Saccharo-1-4-lactone · Zea mays
Abbreviations ABA: Abscisic acid · BAP: 6-Benzylaminopurine · GA3: Gibberellic acid · GUS: �-Glucuronidase · MUG: 4-Methyl-umbelliferyl-beta-D-glucuronide · NAA: Naphthalene acetic acid · SL: Saccharo-1-4-lactone · X-gluc: 5-Bromo-4-chloro-3-indolyl glucuronide
Introduction
The enzyme �-glucuronidase (GUS) has been widely stud-ied in micro-organisms and animals (reviewed by Fishman1955). The GUS enzyme from E. coli has been well charac-terized and the GUS gene from the same organism iswidely being used as a reporter in plant transformationexperiments (JeVerson and Wilson 1991). The enzymeGUS is also known to be present in many bacterial speciessuch as the enterobacterium Shigella, which is closelyrelated to E. coli. Its activity was also observed in non-enterobacterial, anaerobic residents of the digestive tract,such as Bacteroides and Clostridium (Hawkesworth et al.1971). Other bacterial species with GUS activity are Strep-tococcus, Staphylococcus, Corynebacterium (Levvy andMarsh 1959) and Alcaligenes (JeVerson 1989). In thesebacteria, GUS plays a role in carbohydrate and energymetabolism. The GUS activity is very common in virtuallyall tissues of vertebrates where it is known to play a majorrole in the stepwise degradation of glycosaminoglucuro-nides and in the release of active hormones from steroidhormone–glucuronide conjugates (Kyle et al. 1992). Theenzyme may also participate in the catabolism of polysac-charides containing glucuronic acid (Levvy and Marsh1959). In the rat preputial gland, GUS is estimated to con-stitute as much as 5% of the total protein (Kyle et al.1992). Its activity is also known to be present in organismsfrom invertebrate taxa (Koburger and Miller 1985) suchas in snails (Levvy and Marsh 1959), nematodes (Sebasti-ano et al. 1986) and insects (Levvy and Marsh 1959; Lang-ley et al. 1983). GUS activity has also been detected in thehouseXy (Musca domestica) and in various locusts (Levvyand Marsh 1959).
Charu Sudan and Shiva Prakash, the Wrst two authors, havecontributed equally.
C. Sudan · S. Prakash · P. Bhomkar · S. Jain · N. Bhalla-Sarin (&)Lab no. 308, School of Life Sciences, JNU, New Delhi 110 067, IndiaE-mail: [email protected]: +91-11-5525397

854
Biochemical studies of GUS in plants are limited.Schulz and Weissenbock (1987) reported partial puriWca-tion and characterization of a speciWc GUS from rye pri-mary leaves and further studies by Anhalt andWeissenbock (1992) implicated GUS in lignin biosynthe-sis in rye. They localized the presence of GUS to the apo-plastic space. In addition, Scutellaria baicalensis has longbeen known to possess a GUS, called baicalinase (Lewy1954). The GUS enzyme from S. baicalensis has beenpuriWed and characterized (Matsuda et al. 1995; Morim-oto et al. 1995). Muhitch (1998) reported endogenousGUS activity in the pedicel of developing maize kernelswhich had an estimated Mw of ca. 32 kDa, stimulated byassay at 60°C and showed inhibition of activity at highionic strength or in the presence of EDTA. Inspite ofthese reports, GUS in plants is not well studied as com-pared to the GUS from animals or micro-organisms. Thewidespread use of E. coli-derived uidA (GUS) reportergene (JeVerson 1987a) as a scorable marker for genetictransformation of plants (Gilissen et al. 1998) perhapsreXects the general notion that the enzyme GUS is absentin plants. There are, however, reports of detection ofGUS activity in speciWc tissues particularly in immaturetissues of a few speciWc plant species, after prolongedincubation in GUS staining buVer. Plegt and Bino (1989)had shown GUS activity in the tapetum and pollen ofuntransformed plants of potato, tomato and Nicotianatabacum. Hu et al. (1990) surveyed 52 plant species forintrinsic GUS like activity using either the quantitativeXourimetric GUS assay where the tissues were assayedfor 24 h or the histochemical assay which required anovernight incubation in the staining solution. Underthese conditions, they reported that with few exceptions,the GUS activity was detected in certain part(s) of thefruit wall, seed coat, endosperm or the embryos of thetested plants. In addition, a number of publications onplant transformation in diVerent plant species report‘background activities’ of GUS (Bekkaoui et al. 1988;Janssen and Gardner 1989; Lee et al. 1988; Martin et al.1990; Raineri et al. 1990; Wenzler et al. 1989; Woznaikand Owens 1994). In most of these reports on GUS activ-ity in plants, the observation was limited to recording theblue color after prolonged incubation in the histochemi-cal assay buVer. In some instances, the endogenous GUSactivity was explained to be due to the endosymbiontbacteria present in these plants (Wilson et al. 1992). Infact, Tor (1992) showed a positive GUS activity associ-ated with the endogenous microbes in yam. In contrast,Muhitch (1998) tested maize kernels for the presence ofendophytes probably being the causers of the endoge-nous GUS activity and could exclude that. Till date, thegenerally held belief is that there is no GUS enzyme inhigher plants, and the blue stain observed after the GUSassay in the untransformed plant tissues was due to thepresence of endosymbiotic micro-organisms or microbialcontamination during the enzyme assay (Raineri et al.1990; Wilson et al. 1992). Hence, eVorts have been mostlyfocused on the suppression of the background activity(Alwen et al. 1992; Hodal et al. 1992; Kosugi et al. 1990)
and little is known about its biochemistry, physiologyand expression pattern in plants.
While standardizing conditions for transformation ofpigeonpea (Cajanus cajan L.), we carried out co-cultiva-tion of cotyledonary node explants (Shiva Prakash et al.1994) with Agrobacterium. When explants were assayedhistochemically for GUS, an intense blue color devel-oped not only in the transformed explants but also in theuntransformed control explants maintained on non-selection medium. Further investigations revealed that inpigeonpea, GUS has an acidic pH optimum. Whenassayed at acidic pH, high GUS activity was detected notonly in majority of the cell types of pigeonpea but also inmost of the cell types of the model plants viz., Arabidop-sis thaliana, Oryza sativa, N. tabacum, Zea mays and 20other species of plants. In this report we document (a)the ubiquitous presence of GUS in several plants includ-ing the model plants A. thaliana, O. sativa, Z. mays andN. tabacum (b) association of GUS activity with growthat organ and cellular level (c) enhancement of GUSactivity in speciWc plant tissues when treated with 6-ben-zylaminopurine (BAP) or gibberellic acid (GA3) and (d)the strong but reversible inhibition of elongation intobacco seedlings treated with saccharo-1-4-lactone(SL), which is a speciWc inhibitor of GUS.
Materials and methods
Plant material
Seeds of Z. mays cv. Sathi and O. sativa cv. Pusa Basmatiwere obtained from Genetics Division, IARI, New Delhi,India; pigeonpea (C. cajan L. cv. Pusa 851) was obtainedfrom Pulse Division, IARI, New Delhi, India. Wild-typeseed stocks of A. thaliana (ecotype Columbia) wereobtained from A. thaliana Biological Resource Center atOhio State University, USA.
Seeds of Z. mays, O. sativa and N. tabacum were sur-face sterilized with 0.1% HgCl2 followed by severalwashes with sterile double distilled water and placed inculture tubes containing 0.6% agar for germination. Allthe cultures were maintained under continuous whitelight (Xuence density of 50 �mol m¡2 s¡1) at 25 § 2°C forgermination. Ten-day-old seedlings of Z. mays and O.sativa and 17-day-old seedlings of N. tabacum were usedto study GUS activity. Arabidopsis seeds were surfacesterilized with 1% bleach for 5 min and incubated at 4°Cfor 48 h before germination on 0.6% agar. Twenty plantsof each of these model plants were transferred to green-house for GUS expression analysis in the reproductivestages.
Leaf, stem and root samples of Abelmoschus esculen-tus, Amaranthus paniculatus, Brassica juncea, Cocosnucifera, Eleusine coracana, Glycine max, Gossypiumhirsutum, Hibiscus rosa chinensis, Helianthus annus,Lathyrus sativus, Litchi chinensis, Petunia hybrida, Pisumsativum, Secale cereale, Solanum tuberosum, Triticum

855
aestivum, Tagetes erecta, Vigna radiata, Vigna mungoand Vigna unguiculata were collected from the localbotanical garden, dipped in 0.1% Tween 20 for 10 minand washed three times with sterile water.
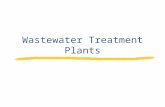
From Wve 40-day-old tobacco plants grown in vitro,the fourth inter-nodes (measuring 2.5–4 cm countedfrom the base of the plants) were harvested. Each of theinter-nodes was cut into three equal segments and GUSactivity was measured, respectively, in each segment(see Fig. 6).
From 12-week-old rice seedlings, 5–6 cm long stemsegments containing the uppermost inter-node wereexcised. Each of the inter-nodes was cut into three equalsegments. GUS activity was measured in each of the seg-ments (see Fig. 6).
Quantitative assay
Enzyme extraction and determination of protein content
Plant tissue was ground with extraction buVer 0.1 M cit-rate phosphate buVer, 1 mM EDTA, 5.6 mM �-mercapto-ethanol, adjusted to pH 4.0 at the ratio 1:5 of fresh weight(g) to buVer volume (ml). The homogenate was centri-fuged at 600 g for 5 min at 4°C. The supernatant was re-centrifuged at 20,000 g for 20 min. All extraction proce-dures were performed on ice. Assays were carried out withthis Wnal supernatant (tissue extract). Protein concentra-tion was determined by using the Bradford’s dye bindingassay (Bradford 1976) with BSA as standard.
Flourimetric assay of GUS activity
An aliquot of the supernatant containing a Wxed amountof total protein in the range of 5–50 �g dissolved in100 �l extraction buVer was added to 100 �l of 2 mM 4-methyl-umbelliferyl-beta-D-glucuronide (MUG) madein the extraction buVer. The mixture was vortexed brieXyand incubated at 37°C for 60 min. The reaction wasstopped by the addition of 900 �l 0.2 M Na2CO3 andXuorescence was measured at 455 nm using a ShimadzuRF-540 spectroXuorimeter set at an excitation wave-length of 365 nm.
Determination of pH optima
The eVect of pH on GUS activity was determined usingcitrate-phosphate and sodium phosphate buVers in thepH range of 3.0–6.0 and 7.0–8.0, respectively.
Histochemical GUS assay
Plant materials were kept in 2 ml Eppendorf tubes andcompletely immersed in GUS-staining solution (0.1 MNaH2PO4 pH 4.0, 0.5 mM K3[Fe(CN)6], 0.5 mMK4[Fe(CN)6]·3H2O, 1 mM EDTA, 0.1% Triton X-100,1 mg/ml 5-bromo-4-chloro-3-indolyl glucuronide (X-gluc), 0.02% sodium azide). Only one seedling/tissuesection was kept in each tube. Paint brush was used for
handling the tissue samples. The samples were vacuuminWltrated for 5 min (ca. 200 mbar) to facilitate penetra-tion of the staining solution. Sodium azide was includedin the buVer to prevent microbial growth. X-gluc stainingof the tissues was conducted at 37°C for 6 h. The stainedsamples were immersed in 70% ethanol to removechlorophyll. The ethanol solution was changed thrice at1 h interval. GUS expression was examined understereomicroscope.
Hormonal induction
Calli of Z. mays were generated as reported previously(Gordon-Kamm et al. 1990). Tissue samples viz., youngas well as mature leaves or mature roots from 40-day-oldN. tabacum plants, young or mature leaves as well asmature roots or calli of Z. mays were cultured on MSbasal medium or MS medium supplemented with BAP(0.44–44.4 �M) or naphthalene acetic acid (NAA; 0.54–53.7 �M) or GA3 (50–500 �M) or abscisic acid (ABA)(0.1–5.0 �M) or ethrel (2.7–276.8 �M). ABA and ethrel,when used, were Wlter sterilized before adding to theautoclaved medium. After 7 days of culture, tissue sam-ples were assayed for GUS activity.
Gibberellic acid treatment of rice inter-nodes was per-formed as described by Cho and Kende (1997). BrieXy,5–6 cm long stem segments containing the uppermostinter-node were excised from 12-week-old plants. Fifteento twenty sections were incubated in distilled water or inGA3 (50–500 �M) or BAP (0.44–44.4 �M) or NAA(0.54–53.7 �M) or ABA (0.1–5.0 �M) solution such thatthe liquid remained below the basal node for 12 h at25 § 2°C under continuous light (56 �mol m¡2 s¡1).These inter-nodal segments were analyzed both quanti-tatively and qualitatively for GUS activity.
SL treatment
Surface sterilized tobacco seeds (ca. 40–50) were soakedin 100 �l of sterile water in a 2.0 ml Eppendorf tubes andincubated in light for 24 h. Subsequently, seeds weretransferred to 2.0 ml Eppendorf tubes (two seeds pertube) containing 50 �l of MES buVer (100 mM, pH 5.2)with SL (10 mM) and incubated in dark for 10 days. Themedium in the Eppendorf tubes was replaced with freshmedium every 2 days. Seeds similarly germinated in MESbuVer without SL were used as control. To test the eVectof removal of SL, 12 seedlings treated with SL (for10 days) were transferred to 2.0 ml Eppendorf tubes (2seedlings per tube) containing 100 �l MES buVer(100 mM, pH 5.2) after washing with the same buVer andobserved for 7 days for recording their growth. Thelength of three longest root hairs of Wve diVerent plantsfrom each of the treatments was measured.
Statistical analysis
Each experiment was performed at least twice withminimum Wve replicates. The information on the num-

856
ber of replicates and the number of times each experi-ment was repeated is included in the correspondinglegends of the Wgures. Data presented were subjected toF test to detect diVerences between treatments andTukey’s multiple comparison test was used to separatetreatment means.
Results
pH optima of GUS from C. cajan L. is in acidic range
�-glucuronidase activity of proteins extracted frompigeonpea leaves assayed at diVerent pH values is shownin Fig. 1. A high activity of GUS was observed betweenpH 3.0 and 5.0, with the maximum activity at pH 4.0. Incontrast, E. coli-derived GUS had a diVerent activityproWle with the optimum at pH 7.0 (JeVerson 1987b).Hence for all further experiments including histochemi-cal assays, the pH of the assay buVer as well as the pH ofthe extraction buVer was kept at 4.0.
High level of GUS activity in all the model plants and other species tested
Quantitative determination of GUS using Xourimetricassay showed high level of GUS activity in diVerent tis-sues of the model plants A. thaliana, O. sativa, Z. maysand N. tabacum when the assay mixture and extractionbuVer were set at pH 4.0 (Table 1). All the other plantspecies tested (A. esculentus, A. paniculatus, B. juncea,C. nucifera, E. coracana, G. max, G. hirsutum, H. rosachinensis, H. annus, L. sativus, L. chinensis, P. hybrida,P. sativum, S. cereale, S. tuberosum, T. aestivum, T.erecta, V. radiata, V. mungo and V. unguiculata) alsoshowed high GUS activity in root, leaf and stem sam-ples (data not shown).
Spatial distribution of GUS in model plants
In Arabidopsis, GUS activity was localized in the vascu-lature of leaf and stem (Fig. 2a–d). Developing pods andXorets stained dark blue (Fig. 2b). Cross sections of stemrevealed higher activity in the cells of the vasculature likexylem and phloem and moderate activity in the collen-chymatous cells as compared to the cells of the pith(Fig. 2d).
In the leaves of O. sativa, the GUS activity wasdetected in most of the cell types viz., mesophyll, xylem,phloem, sclerenchymatous cells of leaf (Fig. 3a, b) and toa lesser extent in epidermis. In the root, most of the GUSactivity was associated with the central vascular cylinder(Fig. 3c). GUS staining was observed predominantly atthe basal region of the inter-nodal stem segments(Fig. 3d). Cross sections of stem revealed higher activityin vascular bundle as compared to the surrounding par-enchymatous cells (Fig. 3e, f). Spikelets and anthersincluding the Wlaments also stained blue showing thepresence of GUS in the reproductive tissues as well(Fig. 3g).
Very high GUS expression was observed in Z. mays(Table 1, Fig. 4). The bundle sheath cells, xylem, phloem,sclerenchyma and mesophyll cells of leaf stained blue(Fig. 4a). All the cells of corn root viz., root hair, epider-mis, endodermis and stele also stained dark blue(Fig. 4b). Staining of the inter-nodal sections showedactivity associated with the phloem region of the vascu-lature (Fig. 4c, d). The glumes and pedicel of the malespikelets ca. 4–5 days prior to anthesis stained blue whilethe anthers did not show GUS activity at the same stage(Fig. 4e). However, the anthers showed GUS staining onthe day of anthesis (Fig. 4f). In the female spikelets ofcorn, the glumes, the basal pedicel as well as nucellusstained dark blue (Fig. 4g).
In 17-day-old seedlings of tobacco, GUS expressionwas observed in the growing root and shoot tips as wellas in the leaf vasculature (Fig. 5). In the roots of 40-day-old tobacco seedlings, GUS activity was seen mostly inthe sclerenchymatous cells (Fig. 5b). GUS activity wasalso detectable in all the Xoral parts (Fig. 5c).
In general, GUS activity could be detected histo-chemically in all the tissues (vegetative as well as repro-ductive) viz., root, stem, leaves, Xowers and fruits ofmodel plant species Arabidopsis, O. sativa, Z. mays and
Fig. 1 EVect of pH on the activity of �-glucuronidase (GUS) proteinextract from pigeonpea leaves. Each data point is the mean of sixreplicates. The experiment was repeated thrice with similar results.Data shown are from a single representative experiment
0.0
5.0
.0
15
10
.0
20.0
25.0
30.0
35.0
3 4 5 6 7 8pH
nmol
es o
f M
U /
min
/ mg
prot
ein
Table 1 Quantitative measurement of �-glucuronidase activity(nmol MU/min/mg protein) in diVerent plant species
Each data point is the mean of Wve replicates. The experiment wasrepeated twice with similar results. Data shown are from a singlerepresentative experiment
Species Leaf Stem Root
Oryza sativa 2.41 § 0.53 2.92 § 0.69 1.35 § 0.27Nicotiana tabacum 1.17 § 0.28 0.63 § 0.17 0.62 § 0.27Zea mays 1.69 § 0.57 1.14 § 0.36 3.85 § 0.91Arabidopsis thaliana 1.12 § 0.26 1.55 § 0.39 0.21 § 0.11

857
N. tabacum (Figs. 2, 3, 4, 5). Moreover, in all these plantspecies, thick-walled cells like the cells of the vascula-ture (including xylem, phloem and sclerenchyma) aswell as collenchymatous cells of Arabidopsis showeddarker staining.
Developmental regulation of GUS expression
The developmental stage of the tissue was found to becritical for the detection of GUS activity in any particu-lar plant organ. In the roots of 17-day-old seedlings of
Fig. 2 Histochemical localiza-tion of GUS activity in diVerent tissues of Arabidopsis thaliana. a Eight days after germination (bar = 2.7 mm), b Xorets (bar = 1.1 mm), c cross section of leaf (bar = 53 �m), d cross section of stem (bar = 71 �m). cc Collenchyma; vb vascular bundle; if inter-fascicular sclerenchyma
Fig. 3 Histochemical localization of GUS activity in diVerent tis-sues of rice (Oryza sativa). a Cross section of leaf whorl(bar = 322 �m), b cross section of leaf (bar = 112 �m), c cross sec-tion of root (bar = 212 �m), d segment of rice with Wrst and second
inter-node (bar = 5 mm). in1 1st inter-node; n node; in2 2nd inter-node. e Inter-node cross section (bar = 392 �m), f close-up view ofa portion of e (bar = 98 �m), g spikelet without the glumes on theday of anthesis (bar = 2 mm)

858
N. tabacum, the root tip zone showed darker stain forGUS activity as compared to the diVerentiated zone(Fig. 5a). Similarly the shoot apex stained darker forGUS as compared to the basal diVerentiated zone. Therice inter-nodes when stained for GUS also showeddarker staining at the basal region (Fig. 3d) which cor-responds to the intercalary meristem and elongationzone (this structure has been described previously byCho and Kende 1998).
Quantitative measurement of GUS activity in diVer-ent segments of tobacco inter-nodes revealed higheractivity in the upper segments as compared to the basal
segments (Fig. 6a). In contrast, in rice higher activity wasseen in the basal segments of the inter-nodes as com-pared to the upper region (Fig. 6b).
Staining of B. juncea roots for GUS activity revealeda striped pattern of darkly stained trichoblast cellsarranged in Wles (Fig. 7a). Staining was also intense inthe elongating root hairs (Fig. 7b). In contrast, GUSstaining intensity was reduced in the mature root hairs(Fig. 7c).
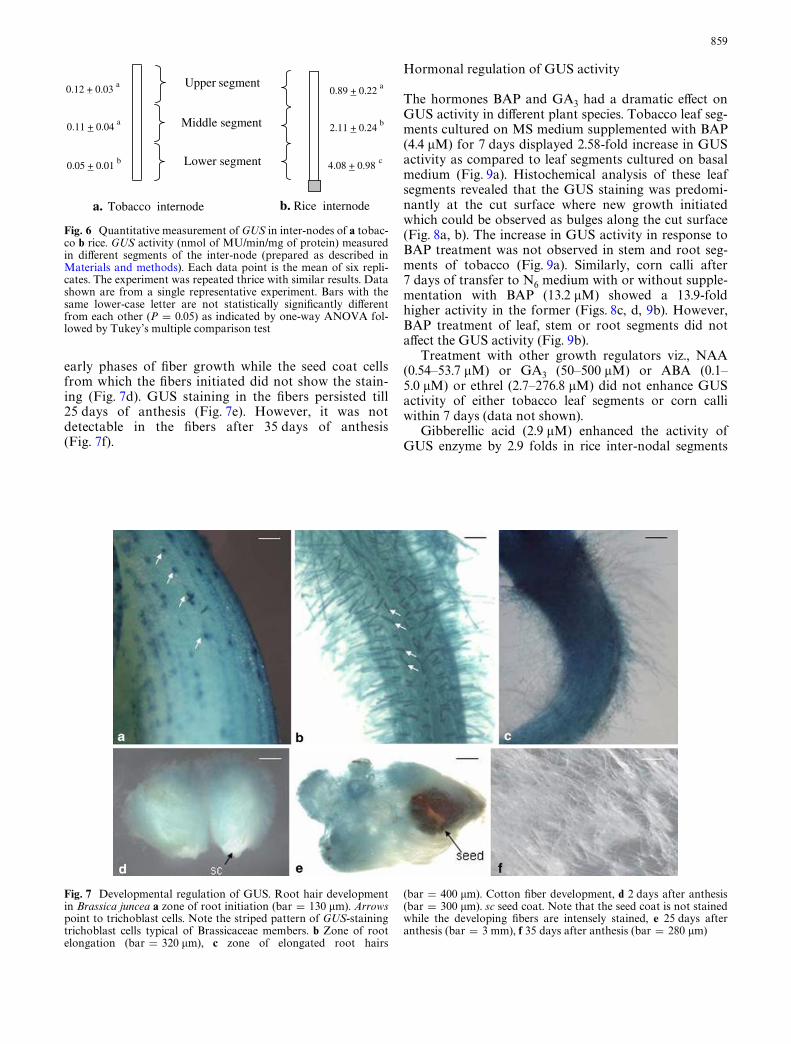
Developing cotton (G. hirsutum) Wbers showed darkblue color within 1 h of incubation in GUS stainingbuVer. The staining intensity of GUS was high in the
Fig. 4 Histochemical localization of GUS activity in diVerent tis-sues of corn (Zea mays). a Cross section of leaf (bar = 112 �m). eEpidermis; m mesophyll; bs bundle-sheath. b Root (bar = 240 �m),c cross section through inter-node (bar = 498 �m), d close up of a
vascular bundle from inter-node (bar = 90 �m). ph Phloem. e Malespikelet ca. 4–5 days prior to anthesis (bar = 1 mm), f an anther onthe day of anthesis (bar = 0.4 mm), g female spikelet (bar = 1 mm)
Fig. 5 Histochemical localization of GUS activity in diVerent tis-sues of tobacco (Nicotiana tabacum). a Seventeen-day-old seedling(bar = 1.7 mm), b cross section of root of 40-day-old seedling
(bar = 570 �m), c Xoral organs on the day of anthesis, i calyx, iigynoecium, iii corolla, iv anther (bar = 1.5 mm)
859
early phases of Wber growth while the seed coat cellsfrom which the Wbers initiated did not show the stain-ing (Fig. 7d). GUS staining in the Wbers persisted till25 days of anthesis (Fig. 7e). However, it was notdetectable in the Wbers after 35 days of anthesis(Fig. 7f).
Hormonal regulation of GUS activity
The hormones BAP and GA3 had a dramatic eVect onGUS activity in diVerent plant species. Tobacco leaf seg-ments cultured on MS medium supplemented with BAP(4.4 �M) for 7 days displayed 2.58-fold increase in GUSactivity as compared to leaf segments cultured on basalmedium (Fig. 9a). Histochemical analysis of these leafsegments revealed that the GUS staining was predomi-nantly at the cut surface where new growth initiatedwhich could be observed as bulges along the cut surface(Fig. 8a, b). The increase in GUS activity in response toBAP treatment was not observed in stem and root seg-ments of tobacco (Fig. 9a). Similarly, corn calli after7 days of transfer to N6 medium with or without supple-mentation with BAP (13.2 �M) showed a 13.9-foldhigher activity in the former (Figs. 8c, d, 9b). However,BAP treatment of leaf, stem or root segments did notaVect the GUS activity (Fig. 9b).
Treatment with other growth regulators viz., NAA(0.54–53.7 �M) or GA3 (50–500 �M) or ABA (0.1–5.0 �M) or ethrel (2.7–276.8 �M) did not enhance GUSactivity of either tobacco leaf segments or corn calliwithin 7 days (data not shown).
Gibberellic acid (2.9 �M) enhanced the activity ofGUS enzyme by 2.9 folds in rice inter-nodal segments
Fig. 6 Quantitative measurement of GUS in inter-nodes of a tobac-co b rice. GUS activity (nmol of MU/min/mg of protein) measuredin diVerent segments of the inter-node (prepared as described inMaterials and methods). Each data point is the mean of six repli-cates. The experiment was repeated thrice with similar results. Datashown are from a single representative experiment. Bars with thesame lower-case letter are not statistically signiWcantly diVerentfrom each other (P = 0.05) as indicated by one-way ANOVA fol-lowed by Tukey’s multiple comparison test
Lower segment
Middle segment
Upper segment 0.12 + 0.03 a
0.11 + 0.04 a
0.05 + 0.01 b 4.08 + 0.98 c
2.11 + 0.24 b
0.89 + 0.22 a
b. Rice internode a. Tobacco internode
Fig. 7 Developmental regulation of GUS. Root hair developmentin Brassica juncea a zone of root initiation (bar = 130 �m). Arrowspoint to trichoblast cells. Note the striped pattern of GUS-stainingtrichoblast cells typical of Brassicaceae members. b Zone of rootelongation (bar = 320 �m), c zone of elongated root hairs
(bar = 400 �m). Cotton Wber development, d 2 days after anthesis(bar = 300 �m). sc seed coat. Note that the seed coat is not stainedwhile the developing Wbers are intensely stained, e 25 days afteranthesis (bar = 3 mm), f 35 days after anthesis (bar = 280 �m)

860
treated for 12 h (Figs. 8e, f, 9c). However, the inter-nodalsegments treated similarly with NAA or ABA did notshow any enhancement of GUS activity (data notshown). Similar treatment of other organs of rice withGA3 did not aVect their GUS activity (Fig. 9c).
EVect of SL
Tobacco seeds, upon germination, showed normal radi-cle emergence within 2–3 days in medium supplementedwith SL as well as in the control medium (without SL).However, diVerences in growth between the two wereobservable from day 4. Elongation of stem and the radi-cle were severely inhibited in all the seedlings grown onthe medium supplemented with SL as compared to those
grown on medium without it (Fig. 10). Root hair forma-tion and elongation (6.27 § 3.53 mm on day 10) was nor-mal in control seedlings, while in SL-treated seedlings,root hair growth was sparse and elongation was severelyinhibited (1.89 § 1.2 mm on day 10). However, growthresumed within 3 days of transfer of the SL-treated seed-lings to control medium (Fig. 10d). The seedlings treatedwith SL when analyzed histochemically showed very lessGUS activity as compared to the untreated controls(Fig. 10c). In contrast, the seedlings which resumedgrowth after the removal of SL treatment showed intensestaining of GUS activity. Moreover, in these seedlings,the staining was observed throughout the stem as com-pared to the control seedlings which stained at the apicalregions only (Fig. 10c, d).
Fig. 8 Histochemical studies on the eVect of plant hormones on GUS activity. a Tobacco leaf segment cultured on MS basal medium (bar = 1.4 mm), b to-bacco leaf segment cultured on MS basal medium with BAP (4.4 �M) (bar = 1.2 �m) for 7 days. Note that the cut surface of the leaf segments is bulging because of growth and also stained blue as compared to the leaf segment in a. c Corn calli cultured on N6 basal medium (bar = 0.8 mm) for 7 days, d corn calli cultured on N6 medi-um supplemented with BAP (13.2 �M) (bar = 0.75 mm) for 7 days, e rice inter-nodes with-out any hormone treatment (bar = 5 mm), f rice inter-node treated with GA3 (2.9 �M) for 12 h (bar = 4.5 mm). Arrow points to the nodal region of the stem

861
Discussion
The pH is the most critical factor for the detection of plant GUS
The pH of the assay buVer is very critical for detection ofthe GUS activity in plants. At neutral pH, qualitativedetermination of GUS was unsuccessful in most of thetissues tested from diVerent plant species. However,when the assay buVer pH was changed to 4.0, GUS activ-ity could be detected in all the plants and in almost all
the organs both quantitatively and qualitatively(Table 1).
�-glucuronidase in animals (vertebrates and inverte-brates) is also known to have an acidic pH optimum. Themammalian GUS has a pH optimum between 3.5 and 5(Levvy and Marsh 1959; Dutton 1980). In Caenorhabd-itis elegans, the GUS has its optimum activity at pH 5(Sebastiano et al. 1986) and between pH 3 and 5.5 inDrosophila (Langley et al. 1983). It has also beenreported previously that endogenous GUS activity inrye, potato slices, apple seeds and almonds is optimalbetween pH 4 and 5 and in rhubarb at pH 6 (Hodal et al.1992). In contrast, E. coli GUS has an optimum activityat pH of 7.0 (JeVerson 1987b).
Though E. coli GUS gene has been used extensivelyas reporter gene in plant transformation experiments, theexpression of plant gene has largely escaped detectionexcept when reported as background activity. At pH 7.0,plant GUS activity is undetectable by qualitative (histo-chemical) method in most of the plants tested (data notshown). The E. coli-derived GUS has optimum activityat pH 7.0 and hence plant tissues are assayed at neutralpH after transformation. The diVerence in the pHoptima between E. coli-derived GUS reporter and theplant GUS could be the reason why the plant GUS haslargely gone undetected except as a faint background.
Plant GUS is associated with growth
The developmental stage of the tissue/organ is anothercritical factor for the detection of GUS in plants. In gen-eral, young and developing organs showed more GUSactivity than the older and more mature tissues.
In tobacco seedlings, histochemical localization stud-ies revealed GUS expression to be predominant in theroot and shoot apical regions (Fig. 5). Quantitation ofGUS activity in inter-nodal segments of 40-day-oldtobacco plants showed more GUS activity in the upperregion of the segment than in the basal region (Fig. 6a).In contrast higher activity was seen in the basal segmentof rice inter-nodes as compared to the upper segments(Figs. 6b, 3d). This could be explained by the fact that inthe growing inter-nodes of dicots like tobacco, the divid-ing and elongating cells are restricted to the upperregions while the basal region of the same consistsmostly of diVerentiated cells. In contrast, in monocotslike rice, intercalary meristem is present at the basal endsof each of the inter-nodes where cells divide and elon-gate, while the upper segment of the inter-node mostlyconsists of diVerentiated cells. Moreover, the intensity ofGUS staining increased in the region of intercalary meri-stem and the elongating zone of rice inter-nodes whentreated with GA3 (Fig. 8f). This is the region where GA3is known to bring about growth (Cho and Kende 1998).Thus higher level of GUS activity seems to be associatedwith growing regions of the inter-nodes of tobacco andrice.
That the GUS expression is associated with cell elon-gation is illustrated during root hair growth of B. juncea.
Fig. 9 EVect of BAP or GA3 supplementation in the medium onGUS activity of cultured tissue samples. a EVect of BAP on GUSactivity in diVerent tissues of tobacco. b EVect of BAP on GUS activ-ity in diVerent tissues of corn. c EVect of GA3 on GUS activity indiVerent tissues of rice. Each data point is the mean of six replicates.The experiment was repeated thrice with similar results. Data shownare from a single representative experiment. Bars with the same low-er-case letter are not statistically signiWcantly diVerent from eachother (P = 0.05) as indicated by one-way ANOVA followed by Tu-key’s multiple comparison test
0
0.4
0.8
1.2
1.6
2
leaf stem root
nmol
es o
f M
U /
min
/ m
g pr
otei
nnm
oles
of
MU
/ m
in /
mg
prot
ein
nmol
es o
f M
U /
min
/ m
g pr
otei
n
control with BAP
b
a a aa a
control with BAP3.0
0.0
5.0
10.0
15.0
20.0
leaf stem root
control with GA3
0.0
0.5
1.0
1.5
2.0
2.5 c cc
a a
b b
d
leaf stem root calli
c
ba a a a
a
b
c

862
Root hair formation is initiated from specialized elon-gating root epidermis cells called trichoblasts which arearranged in Wles in members of the Brassicaceae givingthem a striped pattern on the root surface (Dolan andCosta 2001). Staining of B. juncea roots for GUS activ-ity revealed a striped pattern of GUS staining tricho-blast cells which were arranged in Wles along thelongitudinal axis of the roots (Fig. 7a). GUS stainingwas also marked in the elongating root hairs while itcould not be detected in the mature root hairs (Fig. 7b).This suggested the association of GUS with cell elonga-tion. Cotton Wber is another interesting system for stud-ies on plant cell elongation. A unique feature ofcottonseed development is that epidermal cells initiateinto Wbers from the outermost layer of the seed coat atanthesis. Each cotton Wber is a single cell and elongatesfrom 10–15 mm up to 2.5–3.0 cm in about 16 days afteranthesis before it switches to secondary cell wall synthe-sis (Basra and Malik 1984; Tiwari and Wilkins 1995).The high expression of GUS in cotton Wbers during itsinitiation and growth (Fig. 7d, e) and its absence in thelater stages (35 days after anthesis) further corroboratesits probable involvement in cell growth.
The darker staining in the thick-walled cells like thevasculature in all the species tested or in the collenchy-matous cells of Arabidopsis was noticeable. Elevatedlevel of expression of cell wall loosening proteins likeexpansins which are involved in cell elongation in thick-walled cells has been previously reported (Cho andKende 1998).
Plant GUS is modulated by plant hormones
The plant hormones BAP and GA3 enhanced GUSactivity in speciWc tissues of Z. mays, N. tabacum and O.sativa. It was observed that 7 days of culture of N. taba-cum leaf segments on MS medium supplemented withBAP resulted in 2.58-fold increase in GUS activity overthe controls grown on MS basal medium. However,treatment with other growth regulators, viz., NAA, GA3,ABA or ethrel (as described in Materials and methods)did not enhance GUS activity in the leaf segments. Also,treatment with BAP for 7 days did not increase GUSactivity in other tissues (stem and root) of tobacco.
The eVect of GA3 on GUS expression in rice inter-nodes is also interesting as it is well known that GA3
Fig. 10 EVect of saccharo-1-4-lactone (SL) on growth of tobacco seedlings. a Ten-day-old control seedlings (untreated with SL). Note the natural elongation of stem and root. Root hair density and their elongation are also normal (bar = 2 mm). b Ten-day-old seedlings treated with SL. Note inhibition of elongation of stem as well as root. In addition root hair density and elongation is adversely aVected (bar = 2 mm). c (i) Untreated 10-day-old control seedling stained for GUS (bar = 2 mm), (ii) 10-day-old seedlings treated with SL stained for GUS (bar = 2 mm), d seedlings 3 days after transfer to MES buVer subsequent to treatment with SL for 10 days, stained for GUS (bar = 4 mm)

863
brings about cell elongation in rice inter-nodes (Cho andKende 1997). GA3 treatment for 12 h increased GUSactivity by 2.9 folds in the rice inter-nodal segments. His-tochemical localization of GUS in rice inter-nodesshowed enhanced activity at the basal meristem regionwhich is known to elongate in response to GA3 treat-ment (Fig. 8f). However, application of GA3 did not haveany appreciable eVect on GUS activity in mature leavesand root of the rice plant. Thus, modulation of GUSactivity by GA3 seems to be associated with growth inrice inter-nodes.
SL inhibits elongation
Saccharo-1-4-lactone, which is a speciWc competitiveinhibitor of GUS, severely retarded stem, root and roothair elongation in tobacco seedlings (Fig. 10). The inhibi-tion of GUS was conWrmed by the near absence of histo-chemical staining in SL-treated plants (Fig. 10c). Theinhibition was reversible as the seedlings transferredback to control medium commenced elongation(Fig. 10d). The fact that the inhibition of GUS stronglyretarded elongation of tobacco seedlings suggested thatplant GUS could be involved in growth. It is noteworthythat the staining for GUS in the seedlings removed fromSL inhibition was observed throughout most part of thestem as compared to the restricted activity at the tips inthe control seedlings which were not treated with SL(Fig. 10c, d). This merits further investigation.
The widespread occurrence of GUS in plants alsoraises the question of its natural substrates within theplant. In the cell walls from diVerent plant species,rhamnogalacturonan fractions of the pectin backbonehave been shown to be composed of galactose, arabinoseand �-D-glucuronic acid (Edashige and Ishii 1997;Aspinall et al. 1967, 1968a, b; An et al. 1994; Cui et al.1996; Renard et al. 1999). The acidic pH optima of plantGUS and the existence of �-D-glucuronic acid in the pec-tin component of cell wall suggest the possibility that thesite of activity of plant GUS may be the cell wall. It needsto be tested whether the �-glucuronic acid residuesknown to be present in the pectin of diVerent plant spe-cies is the endogenous substrate for plant GUS.
Transgenic plants over-expressing E. coli GUS havenot been reported to show any observable phenotypicdiVerence as compared to the non-transgenics (Gilissenet al. 1998). This data have been used as an evidence forthe absence of any substrate for the GUS enzyme inplants. However, the absence of any observable pheno-typic eVect of over-expression of E. coli GUS in trans-genics could be because of the diVerence in the sub-cellular localization of the substrate of endogenousGUS and the site of localization of transferred E. coliGUS which is most often localized in the cytoplasm(Iturriaga et al. 1989). Nonetheless, recently, the expres-sion of E. coli GUS speciWcally in the meristem hasbeen reported to be lethal in pea, alfalfa and Arabidop-sis (Wen et al. 2004). To explain why toxicity of ectopicGUS expression has not been previously reported, it is
argued that achieving an observable phenotype byaltering any given gene product depends on numerousfactors including concentration, timing, intra-cellularlocalization, solubility and tissue speciWcity (Wen et al.2004).
Our results on puriWcation of the enzyme and cloningthe gene for plant GUS will provide further support tothe data presented in this study.
Acknowledgements The authors are grateful to Arabidopsis Bio-logical Resource Center, Ohio, for the gift of A. thaliana seeds; to Dr.P. B. Kirti, Biotechnology centre, IARI, New Delhi for providing themicro-photographic facility; to Prof. S. K. Sopory, Dr. Nirmala andProf. S. C. Maheshwari of International Centre for Genetic Engi-neering and Biotechnology, New Delhi and Prof. A. K. Mattoo,U.S.D.A., U.S.A. for going through the manuscript and their valu-able comments and to Prof. S. C. Sacchar, University of Delhi for hissuggestions. The work is an outcome of a research project funded byDST, Government of India.
References
Alwen A, Moreno RMB, Vicente O, Heberle-Bors E (1992) Plantendogenous activity: how to avoid interference with the use ofE.coli �-glucuronidase as a reporter gene in transgenic plants.Transgenic Res 1:63–70
An J, O’Neill MA, Albersheim P, Darvill AG (1994) Isolation andstructural characterization of �- D-glucosyluronic acid and 4-O-methyl-�-D-glucosyluronic acid containing oligosaccharidesfrom the cell wall pectic polysaccharide, rhamnogalacturonan.Carbohydr Res 252:235–243
Anhalt S, Weissenbock G (1992) Subcellular localization of luteolinglucuronides and related enzymes in rye mesophyll. Planta187:83–88
Aspinall GO, Cottrell IW, Egan SV, Morrison IM, Whyte JNC(1967) Polysaccharides of soybeans. Part IV. Partial hydrolysisof the acid polysaccharide complex from cotyledon meal. J ChemSoc (C) 1071–1078
Aspinall GO, Craig JWT, Whyte JL (1968a) Lemon-peel pectin PartI. Fractionation and partial hydrolysis of water-soluble pectin.Carbohydr Res 7:442 452
Aspinall GO, Gestetner B, Molloy JA, Uddin M (1968b) Pectic poly-saccharides from lucerne (Medicago sativa). Part II. Acidic oligo-saccharides from partial hydrolysis of leaf and stem pectic acids.J Chem Soc (C) 2554–2559
Basra A, Malik CP (1984) Development of the cotton Wber. Int RevCytol 89:65–113
Bekkaoui F, Pilon M, Laine E, Raju DSS, Crosby WL, Dunstan DI(1988) Transient gene expression in electroporated Picea glaucaprotoplasts. Plant Cell Rep 7:481–484
Bradford MM (1976) A rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye binding. Anal Biochem 72:248–254
Cho HT, Kende H (1997) Expression of expansin genes is correlatedwith growth in deepwater rice. Plant Cell 9:1661–1671
Cho HT, Kende H (1998) Tissue localization of expansins in deep-water rice. Plant J 15:805–812
Cui W, Eskin MNA, Biliaderis CG, Marat K (1996) NMR charac-terization of a 4-O-methyl-�-D-glucuronic acid-containingrhamnogalacturonan from yellow mustard (Sinapis alba L.)mucilage. Carbohydr Res 292:173–183
Dolan L, Costa S (2001) Evolution and genetics of root hair stripesin the root epidermis. J Expt Bot 52:413–417
Dutton CJ (1980) Glucuronidation of drugs and other compounds.CRC Press, Boca Raton
Edashige Y, Ishii T (1997) Rhamnogalacturonan I from xylemdiVerentiating zone of Cryptomeria japonica. Carbohydr Res304:357–365

864
Fishman WH (1955) �-D-glucuronidase. Adv Enzymol Relat SubjBiochem 16:361–409
Gilissen LJ, Metz PL, Stiekema WJ, Nap JP (1998) Biosafety of E.coli �-glucuronidase (GUS) in plants. Transgenic Res 7(3):157–163
Gordon-Kamm WJ, Spencer TM, Mangano M L, Adams TR, Da-nies RJ, Start WG, O’Brien JV, Chambers SA, Adams WR, Wil-lets NG, Rice TB, Mackey CJ, Krueger RW, Kauseh AP,Lemaux PG (1990) Transformation of maize cells and regenera-tion of fertile transgenic plants. Plant Cell 2:603–618
Hawkesworth G, Draser BS, Hill MJ (1971) Intestinal bacteria andthe hydrolysis of glycosidic bonds. J Med Microbiol 4:451–459
Hodal L, Bochardt A, Neilson JE, Mattson O, Okkels FT (1992)Detection, expression and speciWc eliminaton of endogenous �-glucuronidase activity in transgenic and non-transgenic plants.Plant Sci 87:115–122
Hu C, Chee PP, Chesney RH, Zhou JH, Miller PD, O’Brien TO(1990) Intrinsic GUS like activities in seed plants. Plant Cell Rep9:1–5
Iturriaga G, JeVerson RA, Bevan MW (1989) Endoplasmic reticu-lum targeting and glycosylation of hybrid proteins in transgenictobacco. Plant Cell 1:381-390
Janssen BJ, Gardner RC (1989) Localized transient expression ofGUS in leaf discs following cocultivation with Agrobacterium.Plant Mol Biol 14:61–72
JeVerson RA (1987a) GUS -fusions: �-glucuronidase as a sensitiveand versatile gene fusion marker in higher plants. EMBO J6:3901–3907
JeVerson RA (1987b) Assaying chimeric genes in plants: The �-glu-curonidase gene fusion system. Plant Mol Biol Rep 5:387–405
JeVerson RA (1989) The GUS reporter gene system. Nature342:837–838
JeVerson RA, Wilson KJ (1991) The GUS gene fusion system. PlantMol Biol Manual B14:1–33
Koburger JA, Miller ML (1985) Evaluation of a Xuorogenic MPNprocedure for determining Escherichia coli in oysters. J FoodProtect 48:244–245
Kosugi S, Ohashi Y, Nakajima K, Arai Y (1990) An improved assayfor � -glucuronidase activity in transformed cells: methanol al-most completely suppresses a putative endogenous �-glucuroni-dase activity. Plant Sci 70:133–140
Kyle JW, Galvin N, Vogler C, Grubb JH (1992) GUS (�-glucuroni-dase) assay in animal tissue. In: Gallagher SR (ed) GUS proto-cols: using the �- glucuronidase gene as a reporter of geneexpression. Academic, San Diego, pp 189–203
Langley SD, Wilson SD, Gros AS, Warner CK, Finnerty V (1983) Agenetic variant of �-glucuronidase in Drosophila melanogaster. JBiol Chem 258:7416–7424
Lee BT, Murdoch K (1988) Isolation, culture and morphogenesisfrom wheat protoplasts and study of expression of DNA con-structs by direct gene transfer. Plant Cell Tiss Org Cult 12:223–226
Levvy GA, Marsh CA (1959) Preparation and properties of �-glucu-ronidase. Adv Carbohydr Chem 14:381–428
Lewy GA (1954) Baicalinase, a plant �-glucuronidase. Biochem J56:462–469
Martin T, Scmidt R, Altman T, Frommer WB (1990) Non-destruc-tive assay system for detection of �-glucuronidase activity inhigher plants. Plant Mol Biol Rep 10:37–46
Matsuda T, Hatano K, Harioka T, Taura F, Tanaka H, Tateishi N,Shan S, Morimoto S, ShoyamaY (1995) Histochemical investiga-tion of �-glucuronidase in culture cells and regenerated plants ofScutellaria baicalensis Georgi. Physiol Plant 19(4):390–394
Morimoto S, Harioka T, Shoyoma Y (1995) PuriWcation and char-acterization of Xavone-speciWc �-glucuronidase from callus cul-tures of Scutellaria baicalensis Georgi. Planta 195:535–540
Muhitch MJ (1998) Characterization of pedicel �-glucuronidaseactivity in developing maize (Zea mays) kernels. Physiol Plant104:423–430
Plegt L, Bino J (1989) �-glucuronidase activity during developmentof male gametophyte from transgenic and non-transgenic plants.Mol Gen Genet 216:321–327
Raineri DM, Bottino P, Gordon MP Nester EW (1990) Agrobacte-rium-mediated transformation of Oryza sativa (Oriza sativa L.).Biotechnology 8:33–38
Renard CMGC, Crépeau M, Thibault J (1999) Glucuronic acid di-rectly linked to galacturonic acid in the rhamnogalacturonanbackbone of beet pectins. Eur J Biochem 266:566–574
Schulz M, Weissenbock G (1987) Partial puriWcation and character-ization of a luteolin-glucuronide-speciWc �-glucuronidase fromrye primary leaves (Secale cereale). Phytochemistry 26:933–937
Sebastiano M, D’Alessio M, Bazzicalupo P (1986) �-glucuronidasemutants of the nematode Caernohabditis elegans. Genetics112:459–468
Shiva Prakash N, Pental D, Bhalla-Sarin N (1994) Regeneration ofpigeonpea (Cajanus cajan L.) through multiple shoot-formation.Plant Cell Rep 13:623–627
Tiwari SC, Wilkins TA (1995) Cotton (Gossypium hirsutum) seedtrichomes expand via diVuse growing mechanism. Can J Bot73:746–757
Tor M (1992) Endophytic bacteria expressing �-glucuronidase causefalse positives in transformtion of Dioscorea species. Plant CellRep 11:452–456
Wen F, Woo H, Hirsch AM, Hawes MC (2004) Lethality of induc-ible, meristem-localized ectopic �-glucuronidase expression inplants. Plant Mol Biol Rep 22:7–14
Wenzler H, Mignery G, May G, Park W (1989) A rapid and eYcienttransformation method for the production of large numbers oftransgenic potato plants. Plant Sci 63:79–85
Wilson KJ, JeVerson RA, Hughes SG (1992) The Escherichia coli gusoperon: Induction and expression of the gus operon in Escheri-chia coli and the occurrence and use of GUS in other bacteria. In:Gallagher S (ed) GUS Protocls: Using the GUS gene as reporterof gene expression. Academic Press Inc, New York, vol 1. pp 7–40
Woznaik CA, Owens LD (1994) Native �-glucuronidase activity insugarbeet (Beta vulgaris). Physiol Plant 90:763–771