
agreement for synchronous operation of captive power plants ...

Diet and phylogeny shape the gut microbiota ofAntarctic seals: a comparison of wild andcaptive animals
Tiffanie M. Nelson,1,2* Tracey L. Rogers,1,2
Alejandro R. Carlini4 and Mark V. Brown1,3
1Evolution and Ecology Research Centre, 2School ofBiological, Earth and Environmental Sciences, and3School of Biotechnology and Biomolecular Sciences,University of New South Wales, Kensington, New SouthWales, Australia.4Departmento Ciencias Biológicas, Instituto AntárticoArgentina, Buenos Aires, Argentina.
Summary
The gut microbiota of mammals underpins the meta-bolic capacity and health of the host. Our understand-ing of what influences the composition of thiscommunity has been limited primarily to evidencefrom captive and terrestrial mammals. Therefore, thegut microbiota of southern elephant seals, Miroungaleonina, and leopard seals, Hydrurga leptonyx, inhab-iting Antarctica were compared with captive leopardseals. Each seal exhibited a gut microbiota dominatedby four phyla: Firmicutes (41.5 � 4.0%), Fusobacteria(25.6 � 3.9%), Proteobacteria (17.0 � 3.2%) andBacteroidetes (14.1 � 1.7%). Species, age, sex andcaptivity were strong drivers of the composition ofthe gut microbiota, which can be attributed to differ-ences in diet, gut length and physiology and socialinteractions. Differences in particular prey itemsconsumed by seal species could contribute to theobserved differences in the gut microbiota. Thelonger gut of the southern elephant seal provides ahabitat reduced in available oxygen and more suitableto members of the phyla Bacteroidetes comparedwith other hosts. Among wild seals, 16 ‘core’ bacterialcommunity members were present in the gut of atleast 50% of individuals. As identified between south-ern elephant seal mother–pup pairs, ‘core’ membersare passed on via vertical transmission from a young
age and persist through to adulthood. Our study sug-gests that these hosts have co-evolved with their gutmicrobiota and core members may provide somebenefit to the host, such as developing the immunesystem. Further evidence of their strong evolutionaryhistory is provided with the presence of 18 shared‘core’ members in the gut microbiota of related sealsliving in the Arctic. The influence of diet and otherfactors, particularly in captivity, influences the com-position of the community considerably. This studysuggests that the gut microbiota has co-evolved withwild mammals as is evident in the shared presence of‘core’ members.
Introduction
Bacteria first colonize the gut of mammals during passagethrough the birth canal. As the mammal ages, the gut isexposed to microbes through the consumption of dietaryitems, social interactions and stochastically from thesurrounding environment (Palmer et al., 2007). The gutmicrobiota performs numerous beneficial functions for thehost, such as harvesting energy, regulating immunity andaiding cellular maturation (Savage, 1977; Hooper andGordon, 2001; Turnbaugh et al., 2006).
Colonization of the gut by bacteria is thought to bedeeply rooted in the evolution of the mammal (Ley et al.,2008; Lombardo, 2008). Recent investigations into the gutmicrobiota of adult mammals have identified numerousfactors influencing the unique and variable communitypresent in each individual (e.g. Ley et al., 2006; Rawlset al., 2006; Castillo et al., 2007; Dethlefsen et al., 2008;Benson et al., 2010; Biagi et al., 2010). These include:diet composition and type, host genotype, gut morphologyand physiology, social interactions, exercise, health, hostmass and antibiotic use (Dethlefsen et al., 2007; Leyet al., 2008; Turnbaugh et al., 2009). This knowledge hasbeen derived primarily from studies on humans orlaboratory-reared, agricultural and captive animals (e.g.Hayashi et al., 2002; Bernbom et al., 2006; Castillo et al.,2007; Thompson et al., 2008; Kovacs et al., 2011).Natural systems, however, are inherently more variableand are likely to result in different influences impactingcolonization of the gut or host responses. Captive
Received 25 June, 2012; revised 27 September, 2012; accepted 8October, 2012. *For correspondence. E-mail [email protected];Tel. (+61) (08) 8920 9281, (+61) (0) 425 335 143; Fax (+61) (08)8920 9222.
bs_bs_banner
Environmental Microbiology (2012) doi:10.1111/1462-2920.12022
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd

mammals, for instance, may be fed unnatural diets,inhabit unnatural environments, and have limited ability toexhibit social behaviours. Additionally many of thesemammals have been influenced by routine administrationof antibiotics, which have a profound impact on the gutmicrobiota (Dethlefsen et al., 2008; Dethlefsen andRelman, 2011). Therefore, the current knowledge offactors influencing the gut microbiota may not adequatelyrepresent mammals inhabiting natural or wild systems.Establishing such an understanding in wild mammals,living under natural conditions, has the potential toprovide further insights into the ecological and evolu-tionary relationship between mammals and their gutmicrobiota.
The constraints of an underwater lifestyle have causedmarine mammals to undergo considerable physiological,anatomical and ecological adaptations during their evolu-tion. Many adaptations maximize underwater foragingtimes (Rosen et al., 2007) and this may have implicationsfor the composition of the gut microbiota. For instance, thegastro-intestinal tract of marine mammals is compara-tively longer than terrestrial mammals (Mårtensson et al.,1998). They also have unusual metabolic adaptability; insome cases, they possess elevated metabolic rates dueto their large size yet are able to reduce this considerablyduring foraging dives (Hurley and Costa, 2001; Williamset al., 2001). Marine mammals are also one of the mostunderstudied group of mammals living in natural systems,and although few studies have looked at the gut micro-biota of wild mammals in general (e.g. Nelson et al.,2003; Uenishi et al., 2007), even fewer have investigatedmarine mammals (i.e. Tsukinowa et al., 2008; Glad et al.,2010).
Few comparisons have been made of the gut micro-biota of mammals inhabiting wild and captive environ-ments. Ley and colleagues (2008) in their study onnumerous terrestrial mammals, concluded that bacterialcomposition in the gut was generally more similar amongconspecific hosts than among hosts of different species,regardless of whether they were from a captive or wildenvironment. A few studies have observed considerabledifferences between wild and captive individuals, butamong mammals, these have been limited to primatespecies (Uenishi et al., 2007; Villers et al., 2008; Naka-mura et al., 2011), while several other studies have inves-tigated such questions in birds (Scupham et al., 2008;Xenoulis et al., 2010; Wienemann et al., 2011) and fish(Dhanasiri et al., 2011).
The aim of the present study was to investigate factorsinfluencing the gut microbiota of wild marine mammalsand also compare this with captive equivalents. Faecalsamples were collected from two species of wild marinemammals, the southern elephant seal and the leopardseal, inhabiting Antarctica. These were compared with
faecal samples from leopard seals inhabiting TarongaZoo, Sydney, NSW, Australia. Next-generation sequenc-ing technologies, utilizing tagged PCR amplification andsequencing of hyper-variable regions of the 16S rRNAgene, were applied to DNA extracted from faecal materialto understand bacterial diversity present in the gut. Thetwo species were chosen because they are closelyrelated, which may limit the differences between factorssuch as evolutionary history, diet, gut morphology andenvironment, which have been shown to influence the gutmicrobiota of mammals (Ley et al., 2008). These speciesare two of 15 extant species (Davis et al., 2004; Higdonet al., 2007) from the phylogenetic family, Phocidae. Bothspecies inhabit the Antarctic and Southern Ocean Region,allowing us to control for or minimize the influence ofstochastic exposure to the gut from the environment.These species also both forage for and consume a car-nivorous diet and possess a simple gut morphology, i.e. asingle-chambered stomach, a reduced cecum and longsmall intestine (Lavigne et al., 2007). To investigate theinfluence of captivity on the gut microbiota, leopard sealshoused in Taronga Zoo (Sydney, Australia) were alsosampled, to allow for comparison with the wild individuals.Southern elephant seals were sampled at three distinctage-classes to identify the effect of age-dependantfactors, including lactation, on the composition of the gutmicrobiota. To investigate the broader influences affectingthe gut microbiota of these hosts, their gut microbiota wascompared with data from a previous study, which identi-fied the gut microbiota present in three species of sealinhabiting the Arctic, also from the family Phocidae (Gladet al., 2010).
Results
16S rRNA gene pyrosequencing of the sealgut microbiota
A total of 251 648 sequences from the hyper-variableV1–V3 region of the 16S rRNA gene were obtained. Thisresulted in a total of 6159 operational taxonomic units(OTUs) with � 97% sequence similarity from a total of 19bacterial phyla. Each individual’s community averaged343 � 20 OTUs dominated by (98.2 � 0.5% of total abun-dance) representatives from four phyla: Firmicutes(41.5 � 4.0%), Fusobacteria (25.6 � 3.9%), Proteobacte-ria (17.0 � 3.2%), and Bacteroidetes (14.1 � 1.7%).
Influence of host species and captivity on thegut microbiota
Identification of the gut microbiota was performed andcompared between southern elephant seal adultsand sub-adults (n = 18), wild leopard seals (n = 12) and
2 T. M. Nelson, T. L. Rogers, A. R. Carlini and M. V. Brown
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

captive leopard seals (n = 2). Community compositionof the gut microbiota displayed significant separationbetween host species groups [PERMANOVA (Permuta-tional multivariate analysis of variance): F = 4.6477,PPERM = 0.0001; Table 1]. Pair-wise tests revealed eachhost species group was significantly different from theother two (Table 1). Abundances of major phyla differedconsiderably between hosts species groups (Fig. 1). Thephylum Proteobacteria contributed more to the gut micro-biota of wild leopard seals (31.1 � 6.9%) than to captiveleopard seals (3.6 � 1.8%) or southern elephant seals
(14.7 � 4.2%), although these trends were not significant(Student’s t-test: P = 0.0395 and 0.0408 respectively;Fig. 1). The phylum Bacteroidetes contributed signifi-cantly more to the gut microbiota of southern elephantseals (20.7 � 2.9%) than wild leopard seals (8.3 � 2.6%)or captive leopard seals (2.2 � 1.0%) (Student’s t-test:P = 0.0059 and 0.0080 respectively; Fig. 1). Microbialrichness of the gut microbiota was significantly higher forsouthern elephant seals compared with wild leopard seals(Student’s t-test: P � 0.0001; Fig. 2). However, captiveleopard seals possessed a gut microbiota with richness
Table 1. Effect of species, location, age-class and sex on the gut microbiota of hosts by a PERMANOVA.
Data Source of variation d.f. SS F P
Southern elephant seals, wild leopard seals, captive leopard seals Species and location 2 30241 4.6477 0.0001***Pair-wise comparison t P
Southern elephant seals, wild leopard seals 2.3479 0.0001***Southern elephant seals, captive leopard seals 1.983 0.0003***Wild leopard seals, captive leopard seals 1.9878 0.0004***
Southern elephant seals Age 2 15870 2.8356 0.0001***Pair-wise comparison t P
Adult, sub-adult 1.2799 0.0398Sub-adult, pup 1.8357 0.001**Adult, pup 1.9902 0.0001***
Southern elephant seal mothers and pups Age 1 10930 4.7383 0.0019Southern elephant seals Sex 1 7151.8 2.3829 0.0033**Wild leopard seals Sex 1 4567.4 1.3732 0.1209Captive leopard seals Individual 1 2751.4 1.079 0.4199
PERMANOVA of gut microbial abundance data to generate a permutated F statistic (F) and permutated P-value (P) with calculated degrees offreedom (d.f.) and sums of squares (SS) noted. P-values given in italics were obtained using Monte Carlo samples from the asymptotic permutationdistribution. Pair-wise a posteriori tests between host species groups and age-classes were conducted using the t-statistic (t). Significance level:***P � 0.001,**P = 0.01, *P = 0.025.
Fig. 1. Average relative abundance of major phyla in the gut microbiota of seal host species groups. Average relative abundance of OTUs foreach major phyla in the gut microbiota of each host species group. Southern elephant seals were separated to reflect similarity in gutmicrobiota. Error bars represent SE. Student’s paired t-test were conducted between groups of individuals for each phylum. Significance level:***P � 0.001, **P = 0.01, *P = 0.025.
The gut microbiota of wild and captive Antarctic seals 3
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

significantly higher than southern elephant seals, as wellas wild leopard seals (Student’s t-test: P = 0.0196 and0.0001 respectively; Fig. 2).
The contribution of OTUs to the average dissimilaritybetween host species groups identified by SIMPER (simi-larity percentages procedure) analysis and, therefore,characteristic of the gut microbial community structurediffered considerably between host species groups(Fig. 3). In southern elephant seals the characteristicOTUs that were relatively more abundant than in wildleopard seals or captive leopard seals were from thegenus Ilyobacter from the phylum Fusobacteria, and thegenus Subdoligranulum from the phylum Firmicutes(Fig. 3A). Characteristic OTUs present in high abundancein wild leopard seals included representatives from thegenus Oceanospirillales from the phylum Proteobacteriaand from the family Clostridiales XI from the phylum Fir-micutes (Fig. 3B). In captive leopard seals the character-istic OTUs were from the genera Subdoligranulum andSporobacter from the phylum Firmicutes, and the generaFusobacterium and Cetobacterium from the phylum Fuso-bacteria (Fig. 3C).
Influence of age-class on the gut microbiota of southernelephant seals
The gut microbiota of southern elephant seals differedsignificantly at each of the three age-classes (F = 2.8356,PPERM = 0.0001). The strongest separation as revealedin pair-wise tests occurred between pups and adults(t = 1.9902, PPERM = 0.0001) and between pups and sub-adults (t = 1.8357, PPERM = 0.001) (Table 1). The gutmicrobiota of adults and sub-adults was not significantlydifferent (t = 1.2799, PPERM = 0.0398; Table 1), so theywere grouped together in all future analyses. Significant
differences were noted between age-classes using PER-MDISP (permutational analysis of multivariate disper-sions) which tests the homogeneity of multivariatedispersions within groups (PERMDISP: F = 16.73,PPERM = 0.0008). However, PERMANOVA results wereaccepted as a reflection of true differences in the meansas distinct groups were observed in the non-metric multi-dimensional scaling (nMDS) ordination plot (Fig. S1) andsignificant differences using analysis of similarity(ANOSIM) (Table S1), which is more robust to heterog-enous dispersions, confirmed the PERMANOVA results.
The gut microbiota of adults and sub-adults possesseda significantly greater abundance of the phylum Firmi-cutes (43.3 � 5.4%) compared with pups (18.0 � 2.7%;Fig. 1). Instead, pups possessed a significantly greaterabundance of the phylum Fusobacteria (62.0 � 4.4%) intheir gut microbiota compared with adults and sub-adults(20.3 � 3.9%; Fig. 1). These OTUs included representa-tives from the genera Ilyobacter, Fusobacterium and Psy-chrilyobacter. Southern elephant seal pups possessed agut microbiota significantly reduced in OTU richness asidentified using the non-parametric estimator of speciesrichness, Chao 1 (Chao, 1984), when compared withsouthern elephant seal adults and sub-adults (Fig. 2). Inaddition, there was a non-significant trend (i.e. P � 0.025)towards an increased OTU richness in the gut microbiotaof southern elephant seals with age-class.
The influence of sex and free-living/environmentalsources on the gut microbiota
There was a significant difference in the gut microbiota ofmale and female southern elephant seals (F = 7151.8,PPERM = 0.0033; Table 1). This difference was notobserved between male and female wild leopard seals
Fig. 2. Microbial richness of the gutmicrobiota of seal host species groups.Microbial richness of the gut microbiota asmeasured using Chao 1 mean with error barsrepresenting SE. Southern elephant sealswere separated to reflect similarity in gutmicrobiota. Student’s paired t-test conductedbetween seal groups. Significance level:***P � 0.001, **P = 0.01, *P = 0.025.
4 T. M. Nelson, T. L. Rogers, A. R. Carlini and M. V. Brown
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology
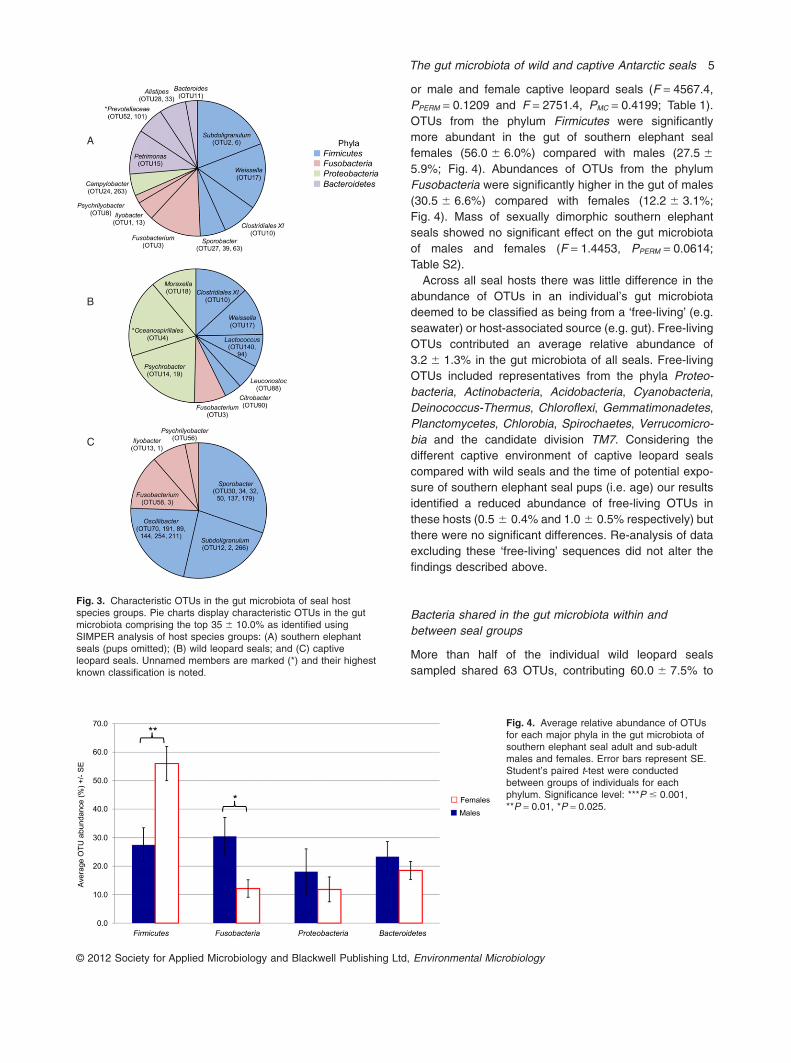
or male and female captive leopard seals (F = 4567.4,PPERM = 0.1209 and F = 2751.4, PMC = 0.4199; Table 1).OTUs from the phylum Firmicutes were significantlymore abundant in the gut of southern elephant sealfemales (56.0 � 6.0%) compared with males (27.5 �
5.9%; Fig. 4). Abundances of OTUs from the phylumFusobacteria were significantly higher in the gut of males(30.5 � 6.6%) compared with females (12.2 � 3.1%;Fig. 4). Mass of sexually dimorphic southern elephantseals showed no significant effect on the gut microbiotaof males and females (F = 1.4453, PPERM = 0.0614;Table S2).
Across all seal hosts there was little difference in theabundance of OTUs in an individual’s gut microbiotadeemed to be classified as being from a ‘free-living’ (e.g.seawater) or host-associated source (e.g. gut). Free-livingOTUs contributed an average relative abundance of3.2 � 1.3% in the gut microbiota of all seals. Free-livingOTUs included representatives from the phyla Proteo-bacteria, Actinobacteria, Acidobacteria, Cyanobacteria,Deinococcus-Thermus, Chloroflexi, Gemmatimonadetes,Planctomycetes, Chlorobia, Spirochaetes, Verrucomicro-bia and the candidate division TM7. Considering thedifferent captive environment of captive leopard sealscompared with wild seals and the time of potential expo-sure of southern elephant seal pups (i.e. age) our resultsidentified a reduced abundance of free-living OTUs inthese hosts (0.5 � 0.4% and 1.0 � 0.5% respectively) butthere were no significant differences. Re-analysis of dataexcluding these ‘free-living’ sequences did not alter thefindings described above.
Bacteria shared in the gut microbiota within andbetween seal groups
More than half of the individual wild leopard sealssampled shared 63 OTUs, contributing 60.0 � 7.5% to
A
B
C
Fig. 3. Characteristic OTUs in the gut microbiota of seal hostspecies groups. Pie charts display characteristic OTUs in the gutmicrobiota comprising the top 35 � 10.0% as identified usingSIMPER analysis of host species groups: (A) southern elephantseals (pups omitted); (B) wild leopard seals; and (C) captiveleopard seals. Unnamed members are marked (*) and their highestknown classification is noted.
Fig. 4. Average relative abundance of OTUsfor each major phyla in the gut microbiota ofsouthern elephant seal adult and sub-adultmales and females. Error bars represent SE.Student’s paired t-test were conductedbetween groups of individuals for eachphylum. Significance level: ***P � 0.001,**P = 0.01, *P = 0.025.
The gut microbiota of wild and captive Antarctic seals 5
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

total abundance of the gut microbiota (Table S3). Most (atleast 50%) southern elephant seals shared 95 OTUs con-tributing 56.4 � 3.6% to total abundance (Table S4).Captive leopard seals shared 49 OTUs at each of thesampling times, contributing an average relative abun-dance of 40.5 � 11.5% per individual (Table S5). Thegreatest abundance of shared OTUs was observedamong southern elephant seal pups with 143 OTUs con-tributing 88.7 � 3.2% to total abundance. The gut micro-biota of pups was significantly different to their mothers(F = 4.1868, PMC = 0.0032; Table 1); yet, they shared20 OTUs that contributed 53.3 � 11.5% to total abun-dance in pups (Table 2). The greatest contributor to abun-dance in the pup gut microbiota was OTU1 from thegenus Ilyobacter in the phylum Fusobacteria with anaverage presence of 30.9 � 4.3% per pup (Table 2). Thethree other highest contributing OTUs in pups werealso from the phylum Fusobacteria from the genera:
Fusobacterium (OTU3), Psychrilyobacter (OTU8) andIlyobacter (OTU13).
Three OTUs belonging to the genera Ilyobacter,Fusobacterium and Psychrilyobacter from the phylumFusobacteria were present in at least 50% of individualswithin each host species group (Table 3). These OTUscontributed an average relative abundance of 9.7 � 2.1%per individual (Fig. 5). At least 50% of individual southernelephant seals and wild leopard seals shared a further10 OTUs, contributing an average relative abundance of21.0 � 2.4% per individual (Table 4; Fig. 5) and includingthe top three most abundant OTUs shared in the gutmicrobiota of at least 50% of southern elephant seal pups(OTU1, OTU3 and OTU8; Table 3). Captive leopard sealsand southern elephant seals shared 21 OTUs, contribut-ing 22.2 � 3.7% to total abundance compared with threeOTUs contributing 4.3 � 2.0% between captive and wildleopard seals (Fig. 5 and Table 4).
Table 2. Shared OTUs in the gut microbiota of mother–pup pairs.
OTU Phylum Genus
Average relative abundance/individual � SE (%)
Mothers Pups
OTU1*∧ Fusobacteria Ilyobacter 2.93 � 1.36 30.91 � 4.32OTU3*∧ Fusobacteria Fusobacterium 5.56 � 4.02 13.00 � 3.85OTU8*∧ Fusobacteria Psychrilyobacter 2.13 � 1.44 6.25 � 0.44OTU13*∧ Fusobacteria Ilyobacter 0.53 � 0.27 6.08 � 0.85OTU11*∧ Bacteroidetes Bacteroides 0.98 � 0.93 4.20 � 2.87OTU2*∧ Firmicutes Subdoligranulum 2.60 � 1.21 1.70 � 0.74OTU27*∧ Firmicutes Sporobacter 3.65 � 1.86 0.16 � 0.07OTU31*∧ Proteobacteria Sutterella 1.60 � 1.53 1.94 � 0.48OTU75*∧ Firmicutes Unnamed member from the family Peptostreptococcaceae 0.16 � 0.07 1.51 � 0.23OTU48*∧ Actinobacteria Collinsella 0.44 � 0.20 1.10 � 0.70OTU287*∧ Firmicutes Unnamed member from the family Peptostreptococcaceae 0.02 � 0.00 0.67 � 0.08OTU81* Proteobacteria Escherichia/Shigella 0.09 � 0.05 1.06 � 0.44OTU151*∧ Firmicutes Anaerococcus 0.69 � 0.40 0.21 � 0.09OTU24*∧ Proteobacteria Campylobacter 0.12 � 0.06 0.49 � 0.26OTU101*∧ Bacteroidetes Unnamed member from the family Prevotellaceae 0.86 � 0.27 0.38 � 0.15OTU110 Firmicutes Sarcina 0.52 � 0.44 0.34 � 0.16OTU388*∧ Fusobacteria Fusobacterium 0.09 � 0.06 0.46 � 0.08OTU17*∧ Firmicutes Weissella 0.32 � 0.12 0.20 � 0.06OTU447*∧ Fusobacteria Ilyobacter 0.06 � 0.02 0.32 � 0.11OTU33*∧ Bacteroidetes Alistipes 0.07 � 0.01 0.16 � 0.11
Average relative OTU abundance in the gut microbiota of mothers or pups � SE. OTUs shared between at least 50% of the individuals: (*)southern elephant seals and wild leopard seals; and (∧) southern elephant seals (mothers and pups omitted) are marked.
Table 3. Shared OTUs in the gut microbiota of host species groups.
OTU Phylum Genus
Average relative abundance/individual � SE (%)
Southern elephant seals(pups omitted)
Wild leopardseals
Captive leopardseals
OTU3 Fusobacteria Fusobacterium 4.4 � 1.8 4.4 � 1.8 3.5 � 2.2OTU1 Fusobacteria Ilyobacter 8.2 � 2.4 8.2 � 2.4 0.5 � 0.2OTU8 Fusobacteria Psychrilyobacter 1.9 � 0.6 1.9 � 0.6 0.9 � 0.5
Average relative abundance of three OTUs in the phylum Fusobacteria that occurred in at least 50% of the individuals in each host species group.
6 T. M. Nelson, T. L. Rogers, A. R. Carlini and M. V. Brown
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

Discussion
A unique gut microbiota shaped byspecies-dependant differences
The significant differences observed in the gut microbiotaof southern elephant and leopard seals suggests thatcolonization of the gut is shaped by stochastic exposuresduring an individual’s lifetime. Similarity of the gut micro-biota between con-specific individuals may be accountedfor by similar diets, behaviours and physiology resulting insimilar exposures of bacteria to the gut.
For these hosts, diet is considered a pathway to colo-nization. Southern elephant and leopard seals are preda-tors consuming a carnivorous diet and while they overlapin the consumption of some prey, such as fish, they pri-marily forage for different prey items. Southern elephantseals forage for squid and fish (Slip, 1995; Bradshawet al., 2003); whereas, leopard seals consume a variety ofprey such as seals, krill, penguins and fish (Rogers andBryden, 1995; Walker et al., 1998; Hall-Aspland andRogers, 2004). Fusobacteria, which are common to bothsouthern elephant and leopard seals are also found inassociation with several species of fish including noto-thenioid fish inhabiting Antarctica (Ramirez and Dixon,2003; Navarrete et al., 2009; Ward et al., 2009), suggest-ing this as one possible route of colonization to themammal gut. Leopard seals, but not southern elephantseals, possessed higher abundances of OTUs from thephylum Proteobacteria and specifically the genus Psy-chrobacteria. These bacterial species have been found inassociation with Antarctic krill, Euphausia superba, Adeliépenguins, Pygoscelis adeliae and gentoo penguins,P. papua (Denner et al., 2001; Banks et al., 2009), whichare common prey of leopard seals but not southernelephant seals.
Following colonization, physicochemical qualities ofthe gut habitat will strongly influence the ability of colo-nizing bacteria to subsist. The gut of the southernelephant seal is approximately 15 times longer than thatof the leopard seal (Bryden, 1971). This was originallysuggested as an adaptation to the frequent and deepdiving performed by southern elephant seals, wherebyblood flow was redistributed away from the gut to favoursensitive tissues, such as the heart (Krockenberger andBryden, 1994). A follow-up study found no significant
Fig. 5. Shared OTUs within and between host species groups.Venn diagram displays the number of shared OTUs in the gutmicrobiota of at least 50% of the individuals in a host speciesgroup.
Table 4. Shared OTUs in the gut microbiota of southern elephant seals and wild leopard seals.
OTU Phylum Genus
Average relative abundance/individual � SE (%)
Southern elephantseals (pups omitted)
Wild leopardseals
OTU17† Firmicutes Firmicutes 0.2 � 0.1 5.6 � 2.2OTU10† Firmicutes Unnamed member from the family Clostridiales XI 1.1 � 0.1 7.0 � 3.1OTU3† Fusobacteria Fusobacterium 4.4 � 1.8 3.3 � 2.1OTU8 Fusobacteria Psychrilyobacter 1.9 � 0.6 0.5 � 0.4OTU1 Fusobacteria Ilyobacter 8.2 � 2.4 0.2 � 0.1OTU15 Bacteroidetes Petrimonas 4.5 � 2.1 0.8 � 0.4OTU454 Firmicutes Roseburia 0.1 � < 0.1 0.1 � < 0.1OTU151 Firmicutes Anaerococcus 0.3 � 0.1 0.3 � 0.3OTU88 Firmicutes Leuconostoc < 0.1 � < 0.1 1.5 � 0.6OTU167 Firmicutes Anaerococcus 0.1 � 0.1 0.1 � 0.1OTU65 Proteobacteria Unnamed member of the family Pseudomonadaceae 0.5 � 0.3 0.1 � < 0.1OTU999 Firmicutes Unnamed member from the family Clostridiales XI < 0.1 � < 0.1 0.1 � 0.1OTU145 Bacteroidetes Porphyromonas 0.2 � 0.1 0.2 � 0.1
Average relative abundance of OTUs shared in more than half of the individual southern elephant seals and wild leopard seals � SE. OTUsoccurring in the gut microbiota of more than three-quarters of individuals are marked (†).
The gut microbiota of wild and captive Antarctic seals 7
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

correlation between intestinal lengths relative to divingability and concluded that gut length is instead corre-lated with body length (Mårtensson et al., 1998).Regardless, the longer gut of the southern elephant sealis a habitat which is reduced in available oxygen andthe significantly greater abundance of OTUs from thephylum Bacteroidetes, which are known to be obligateanaerobes, would be better suited to this habitat. Con-trastingly, facultative anaerobes, such as OTUs from thephylum Proteobacteria and particularly the Gammapro-teobacteria, are better suited to a habitat with availableoxygen and are present in greater abundance in the gutof leopard seals. Oxygen availability as controlled by gutlength in humans has been previously proposed as afactor contributing to alterations in the gut microbiota(Zhang et al., 2009).
A conserved ‘core’ gut microbiota of phocid seals
Bacteria in the gut of mammals perform numerous func-tions that are beneficial to the host including: immuneregulation, tissue maturation, intestinal development, andthe extraction of energy, nutrients and vitamins fromdietary items (Hooper and Gordon, 2001; Bäckhed et al.,2005). This tight symbiotic relationship suggests bacteriain the gut are evolutionarily linked to their mammalian host(Ley et al., 2008; Ochman et al., 2010; Yildirim et al.,2010). Southern elephant seals and leopard seals shared16 OTUs in their gut microbiota, which is suggested to betheir ‘core’ gut microbiota which has co-evolved with thesemammalian hosts. Three of these ‘core’ OTUs, all from thephylum Fusobacteria, were shared with captive leopardseals, indicating the maintenance of the ‘core’ underaltered conditions.
Bacterial sequences from the gut microbiota ofAntarctic seals in this study were compared with apublished study of seals inhabiting the Arctic (Glad et al.,2010) including the: harbour, Phoca vitulina (n = 1), grey,Halichoerus grypus (n = 1), and hooded, Cystophoracristata (n = 9) seal. Sequence taxonomy of the 319 fulllength 16S rRNA gene sequences from Arctic sealswere assigned using the Ribosomal Database Project(RDP) v.10 Classifier tool (Wang et al., 2007). Acrossthe five species of Antarctic and Arctic seals, allwithin the family Phocidae, 60 OTUs from 18 generawere shared. These included representatives from thegenera Bacteroides, Faecalibacterium, Fusobacterium,Oscillibacter, Sporobacter and Paraprevotella. InGenBank, all of the 60 OTUs possessed greater orequal similarity across the sequence length to thoseobserved in the gut microbiota of seals (98.8 � 0.1%)than any other source (95.8 � 0.2%). As dietary differ-ences exist due to geographic separation of these sealhost species, the number and similarity of shared OTUs
in the gut microbiota of individuals highlights that theseOTUs may be seal-specific, having co-evolved with theirseal hosts.
Nineteen core OTUs were identified as being trans-ferred from southern elephant seal mothers to their pups,including three OTUs shared among all host speciesgroups from the genera Ilyobacter and Psychrilyobacter.The gut microbiota of pups performs different functionscompared with that required by older hosts. In youngmammals, the gut microbiota is required to develop theimmune system, mature the gut and break down the milkin order to provide energy (Palmer et al., 2007; Vael andDesager, 2009; Wall et al., 2009). The transfer of theseOTUs to pups under three weeks of age, and theirapparent persistence through to adulthood, suggeststhey may provide a function which is not related directlyto the diet, as adults and pups consume very differentdiets. Instead, they may serve the purpose of immuneregulation as they are recognized by the innate immunesystem as harmless, which may be an indication of along evolutionary association (Rook and Brunet, 2005).Rather than produce an aggressive immune response,which could result in harm to the new pup, these bacteriamay actually prime the immune system in pups and later,in adults, to regulate other aspects of the immunesystem (Rook et al., 2004). If the role of core OTUs islinked to immune function of the hosts, the maintenanceof the OTUs is ensured through vertical (mother-to-infant) transmission.
Captivity exerts a strong influence on the gut microbiota
The gut microbiota of captive and wild leopard seals isdissimilar as a result of their different exposures andliving environment. Conspecific hosts have been shownto display dissimilarities when sampled from captive andwild environments (e.g. Uenishi et al., 2007; Scuphamet al., 2008; Villers et al., 2008; Xenoulis et al., 2010;Nakamura et al., 2011), although this has rarely beendemonstrated in mammals, and this is the first study toidentify this difference in marine seals. Dietary items candrive differences in the gut microbiota through the intro-duction of OTUs associated with prey. The significantdifference in the gut microbiota of wild and captiveleopard seals may be influenced in part by the distinctOTUs associated with their particular prey items. In cap-tivity, leopard seals are fed a diet of fish caught offSydney, Australia, while wild leopard seals feed on avariety of dietary items including krill, penguins, and fish(Walker et al., 1998; Hall-Aspland and Rogers, 2004;Casaux et al., 2009). Changes in the gut microbiota ofthe Atlantic cod, Gadus morhua, were observed after sixweeks in response to artificial feeding of the fish, i.e. achange in diet, when moved from a wild to a captive
8 T. M. Nelson, T. L. Rogers, A. R. Carlini and M. V. Brown
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

environment (Dhanasiri et al., 2011). In captivity, con-stant social interaction, co-habitation, interaction withhuman keepers, the general public, and other mammals(when kept in holding pens), provides increased oppor-tunities for transmission of OTUs from host-associatedsources capable of colonizing the seals and may con-tribute to the increased OTU richness in the gut micro-biota of the captive leopard seals.
Metabolic rate, body condition, hormonal productionand other functions are known to change in animals whenplaced in a captive environment (Mortensen and Tindall,1981; Rangel-Negrín et al., 2009; Fanson et al., 2010).The external environment for these hosts differs consid-erably, which may influence other aspects of metabolismand thermoregulation, thereby influencing the functionalcapacity of the gut (Hooper et al., 2002). For instance,temperatures experienced by captive leopard seals inSydney range from 5 to 41°C (averaging 14 to 41°C insummer) (Bureau of Meteorology, 2011); whereas wildleopard seals in Antarctica experience temperaturesranging from -16 to 3°C (averaging -1 to 3°C in summer)(Ferron et al., 2004). Additionally, antibiotics administeredroutinely to captive mammals may cause a change, whichcan be long-term, in their gut microbiota as has beenpreviously shown in humans (Dethlefsen and Relman,2011). Other studies investigating the gut bacteria ofcaptive mammals have suggested that the microbial com-munities in these animals serve as an indicator for wildspecies. Although greater replication in the number ofcaptive leopard seals is preferred to reduce the variabilityamong samples, this study provides an appropriate com-parison between wild and captive hosts. It gives a clearindication that the gut microbiota of captive mammalsdoes not provide an accurate indication of the gut micro-biota of wild equivalents, except for a subset of coreOTUs.
Social interactions increase the transmission of OTUsbetween hosts
Southern elephant seals possessed a richer gut micro-biota and a higher number of shared OTUs among indi-viduals compared with wild leopard seals. This is a resultof the social nature of southern elephant seals, comparedwith wild leopard seals. Southern elephant seals comeashore to give birth, breed and moult, congregating incolonies of 10 s to 100 s of individuals (McCann, 1980;Carlini et al., 2002). The close proximity to large numbersof conspecifics has been identified as an added benefit ofsocial behaviour in animals, providing increased transmis-sion opportunities (Lombardo, 2008). Wild leopard seals,by comparison, spend relatively little time with conspecif-ics and are solitary except for brief periods when matingand in mother–pup pairs post-partum (Siniff, 1991; Hiruki
et al., 1999; Southwell et al., 2003). The greater numberof shared OTUs among southern elephant seals alsosuggests ongoing transfer of bacteria. This pattern hasbeen observed in social bee species, where greater simi-larity was found in social Apis and Bombus species com-pared with the solitary Agapostemon species, indicatingfacilitated transmission of bacteria between individuals(Martinson et al., 2011).
Negligible contributions to the gut microbiota fromfree-living/environmental sources
The proportion of gut microbiota from a free-living or envi-ronmental source (e.g. seawater) was estimated to benegligible (3.2 � 1.3%) across all seal hosts. Currentglobal surveys suggest that dominant members of the gutmicrobiota, such as those from the phyla Firmicutes andBacteroidetes, do not survive outside of their host (Leyet al., 2006). Among the identified free-living OTUs, theyincluded an OTU belonging to the group SAR11, which isglobally distributed in the seawater (López-García et al.,2001; Prabagaran et al., 2007) and an OTU which was99% identical to P. marinus, a bacterium never beforeidentified in the Antarctic marine environment (Baldwinet al., 2005).
Members of the phylum Fusobacteria, including thosegenera observed in the core gut microbiota, Ilyobacterand Psychyrilyobacter, have been identified in habitatssuch as anoxic sediments and anoxic enrichments ofmarine material (Staley and Whitman, 2005). Therefore,exposure from the environment into the gut of these hostsmay have been underestimated. However, the persistentand high abundances of these OTUs in the gut acrossmultiple species and age-classes suggests they are main-tained in the gut habitat through other pathways. In par-ticular, their shared presence between mother–pup pairsis evidence for their vertical transmission.
Sexual dimorphism drives dietary influences on thegut microbiota
Sex differences were only identified in the gut microbiotabetween southern elephant seals, which are sexuallydimorphic. Fully grown males weigh ~ 3000 kg, whereasfemales are commonly ~ 600 kg (Bryden, 1971). Physi-ological and metabolic differences exist between southernelephant seal males and females, resulting in their differ-ent growth strategies and size allowing them to differ intheir capacity to dive deeper, and access distinct preyitems (Hindell, 1991; Lewis et al., 2006). The different gutmicrobiota observed between the sexes of southernelephant seals suggests even fine-scale prey shifts, withinspecies, may cause changes in the gut microbiota.
The gut microbiota of wild and captive Antarctic seals 9
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

Conclusions
This study identified a distinct gut microbiota of southernelephant seals, wild leopard seals and captive leopardseals. Diet type, prey-associated bacteria, gut morphol-ogy, gut physiology and social behaviours have primaryroles in determining community composition within thegut. Free-living bacterial sources are thought to have anegligible role in shaping the community in these hosts. A‘core’ seal gut microbiota was present in these seals,established via mother-to-pup transmission and remain-ing throughout the life of the host. These ‘core’ OTUs mayhave an essential role in the host. Species-specific gutmicrobiota have been previously identified in wild homin-ids, where gut microbial composition is known to mirrorhost phylogeny (Ochman et al., 2010). While the Arcticseal dataset in the present study was not sufficientlyreplicated to perform a similar analysis, the significantdifferences evident between the southern elephant andleopard seal gut microbiota suggest that seal phylogenysimilarly may be reflected by the gut microbiota. CoreOTUs are suggested to have co-evolved with the hostsand perform a significant functional role such as immunesystem regulation or gut development. Experimentalstudies that include the alteration of diets and/or habitatswould provide insight into the relative contribution of dietand evolutionary history in assembling the gut microbiotaof these wild hosts.
Experimental procedures
Faecal sample collection from Antarctic seals
Faecal samples were collected from southern elephant andleopard seal populations in the western Antarctic from Sep-tember 2008 to March 2009. Southern elephant seals weresedated using the protocol described by Carlini and col-leagues (2009) on 25 de Mayo (King George Island), SouthShetland Islands (62°14′S, 58°39′W). Leopard seals weresedated using the protocol described by Higgins and col-leagues (2002) on ice floes off the coastline, Danco Coast,western Antarctic Peninsula (64°09′S, 60°57′W). Seals weresampled within a study area of approximately 5 km in eachgeographic region.
Faecal samples were collected from sedated seals byrectal swabbing using sterile swabs (Oxoid Australia Pty, SA,Australia) that were frozen and then stored at -20°C. Whilethe gut microbiota of mammals is known to vary along thelength of the digestive system and in faeces (Marteau et al.,2001; Mentula et al., 2005), faecal material is indicative of themicrobiota present in the lower intestinal tract (Eckburg et al.,2005; Mentula et al., 2005). Similarity of the lower colon tothe faeces was confirmed for leopard seal hosts in a separatestudy (data not shown, T.M. Nelson, unpublished).
At the time of sample collection sex was noted for eachseal. Additionally, age-class and mass were noted for south-ern elephant seals only. Three distinct age-classes of south-ern elephant seals were estimated based on knowledge of
the morphology and behaviour of an individual, whichchanges as the seal ages. Host age-class of southernelephant seals was classified as follows: adult female (> 4years), adult male (> 6 years), sub-adult female (2–4 years),sub-adult male (2–6 years), and pup (< 3 weeks) (Table S6).Leopard seal adults only were sampled. Mass for southernelephant seals was measured using the method described inCarlini and colleagues (1997). Briefly, this involved suspend-ing sedated seals in a net stretcher from a load cell hangingscale attached to an aluminium tripod. The mass of non-weighed individuals was estimated using their standardlength (nose to tail) and axillary girth with model six and nineas described in Bell and Hindell (1997).
Faecal samples were collected opportunistically from twocaptive leopard seals, one male sub-adult and one femalesub-adult, housed at Taronga Zoo, Sydney, NSW, Australia.
DNA extraction
Total genomic DNA was extracted from 40 faecal samples,including 24 southern elephant seals (four adult male, sevenadult female, four sub-adult male, three sub-adult female,four pup male and two pup female) 12 leopard seals (six adultmale and six adult female) and two captive leopard seals(one male and one female at two times) (Table S6) usingQIAamp Stool DNA Mini Kit (Qiagen Pty, Vic., Australia) fol-lowing the manufacturers protocol.
Community analysis via 16S rRNAgene pyrosequencing
Pyrosequencing of the16S rRNA gene was carried out at theResearch and Testing Laboratory (Lubbock, TX, USA).Primers 27F 5′-AGAGTTTGATCCTGGCTCAG-3′ and 519R5′-GTATTACCGCGGCTGCTG-3′ were used to PCR amplifya ~ 500 base pair (bp) product spanning the hyper-variableregions V1–V3 (Lane et al., 1985). PCR reactions werecarried out individually for each sample using ~ 10 ngof template DNA. Tagged primer pairs and PCR reactionconditions followed the protocol of Dowd and colleagues(2008).
Sequencing was carried out on a Roche 454 titaniumsequencer. Initial quality control measures, used to ensuresequence fidelity, included: the removal of short sequences(< 100 bp), removal of any sequence not perfectly matchingthe 5′ primer, and removal of any sequence containing anunresolved nucleotide.
Sequence data were analysed using the Mothur v.1.13.0suite of programs (Schloss et al., 2009). Potential chimeraswere removed using the Chimera.slayer tool with the minsnpparameter set to 100. A 2% pre-clustering step was used toremove potential errors in sequence data (Huse et al., 2010).Sequences were aligned to the SILVA database (Pruesseet al., 2007) and those that did not align in the appropriatezone were removed. Alignments were trimmed so that allsequences covered the entire alignment length. The finaldataset comprised 251 648 sequences. Clustering was per-formed at a sequence similarity cut-off of 97%. Representa-tives of resultant OTUs were taxonomically identified usingthe SILVA taxonomy tool.
10 T. M. Nelson, T. L. Rogers, A. R. Carlini and M. V. Brown
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

Data analyses
Fourth-root-transformed abundance data from faecalsamples were used to generate a resemblance matrix usingthe Bray–Curtis similarity algorithm (Bray and Curtis, 1957).Similarities between sample groups were visualized usingnMDS (Guttman, 1968). Permutational multivariate analysisof variance (PERMANOVA) (Anderson, 2001) was carriedout to test the null hypothesis of no difference in composi-tion of gut microbiota. Each test was done using 9999permutations under Type III sum of squares (SS) and areduced model to generate a permutated F statistic (F) andP-value (P). For some terms in the analysis, there werenot enough permutable units to get a reasonable test bypermutation, so a P-value was obtained using a MonteCarlo random sample from the asymptomatic permutationdistribution (Anderson and Robinson, 2003). In the case of asignificant interaction term and where the number of factorswas greater than three, pair-wise a posteriori tests for allcombinations of factors were conducted using the t-statistic.Homogeneous dispersion of samples within each factorwas assessed by a test of multivariate dispersion (PERM-DISP) (Anderson, 2004). In the case of a positive result,indicating heterogeneous dispersion, significant PER-MANOVA values were discussed. Results were consideredsignificant where P-value � 0.025. The contribution of OTUsto the average dissimilarity between groups was calculatedusing a SIMPER. SIMPER analysis identifies which OTUsaccount for observed differences in microbiota between indi-viduals or groups of individuals. All statistical tests were per-formed using the software PRIMER-E v6 (Clarke andGorley, 2006).
Operational taxonomic units defined as ‘shared’ within agroup, e.g. southern elephant seals, were identified whenpresent in at least 50% of the individuals within that group.The average relative abundance of those OTUs was calcu-lated to give an indication of OTU per individual � standarderror (SE). The non-parametric estimator of species richness,Chao 1 mean (Chao, 1984), was calculated using the onlinesoftware program EstimateS (Colwell, 1997). Significant dif-ferences in microbial richness and abundance of phylabetween groups were tested using a Student’s t-test in Excel2010 (Microsoft Pty).
To obtain contextual information regarding the provenanceof any similar 16S rRNA gene sequences to our dataset,sequences were compared with the National Centre forBiotechnology Information non-redundant database (http://www.ncbi.nlm.nih.gov/) using the Basic Local AlignmentSearch Tool (BLAST) (Altschul et al., 1997) in GenBank(Benson et al., 2011). The source of origin was examined(either from the GenBank entry or the associated paper) ofeach high scoring pair and classified as likely being from afree-living/environmental (e.g. seawater) or host-associatedsource (e.g. gut).
Acknowledgements
The contributors are grateful for the field assistance providedby Instituto Antártico Argentino, members of the AustralianMarine Mammal Research Centre and Taronga Zoo staff.Funding for the field support was provided by the Secretaríade Ciencia y Tecnología and the Dirección Nacional del
Antártico (PICTO No. 36054), the Australian ResearchCouncil, Winifred Scott Foundation, and the Evolution andEcology Research Centre. We dedicate this publication to ourfriend, colleague and contributor, Alejandro Carlini, whopassed away in December 2010.
References
Altschul, S.F., Madden, T.L., Schäffer, A.A., Zhang, J., Zhang,Z., Miller, W., and Lipman, D.J. (1997) Gapped BLAST andPSI-BLAST: a new generation of protein database searchprograms. Nucleic Acids Res 25: 3389–3402.
Anderson, M.J. (2001) A new method for non-parametricmultivariate analysis of variance. Austral Ecol 26: 32–46.
Anderson, M.J. (2004) PERMDISP: A FORTRAN ComputerProgram for Permutational Analysis of Multivariate Disper-sions (for Any Two-Factor ANOVA Design) Using Permu-tation Tests. Auckland, New Zealand: Department ofStatistics, University of Auckland.
Anderson, M.J., and Robinson, J. (2003) Generalized discri-minant analysis based on distances. Aust New Zeal J Stat45: 301–318.
Bäckhed, F., Ley, R.E., Sonnenburg, J.L., Peterson, D.A.,and Gordon, J.I. (2005) Host-bacterial mutualism in thehuman intestine. Science 307: 1915–1919.
Baldwin, A.J., Moss, J.A., Pakulski, J.D., Catala, P., Joux, F.,and Jeffrey, W.H. (2005) Microbial diversity in a PacificOcean transect from the Arctic to Antarctic circles. AquatMicrob Ecol 41: 91–102.
Banks, J.C., Cary, S.C., and Hogg, I.D. (2009) The phyloge-ography of Adélie penguin faecal flora. Environ Microbiol11: 577–588.
Bell, C., and Hindell, M.A. (1997) Estimation of body massin the southern elephant seal, Mirounga leonina, byphotogrammetry and morphometrics. Mar Mammal Sci13: 669–682.
Benson, A.K., Kelly, S.A., Legge, R., Ma, F., Low, S.J., Kim,J., et al. (2010) Individuality in gut microbiota compositionis a complex polygenic trait shaped by multiple environ-mental and host genetic factors. Proc Natl Acad Sci USA107: 18933–18938.
Benson, D.A., Karsch-Mizrachi, I., Lipman, D.J., Ostell, J.,and Sayers, E.W. (2011) GenBank. Nucleic Acids Res 39:D32–D37.
Bernbom, N., Nørrung, B., Saadbye, P., Mølbak, L.,Vogensen, F.K., and Licht, T.R. (2006) Comparison ofmethods and animal models commonly used for investiga-tion of fecal microbiota: effects of time, host and gender.J Microbiol Methods 66: 87–95.
Biagi, E., Nylund, L., Candela, M., Ostan, R., Bucci, L., Pini,E., et al. (2010) Through ageing, and beyond: gut micro-biota and inflammatory status in seniors and centenarians.PLoS ONE 5: e10667.
Bradshaw, C.J.A., Hindell, M.A., Best, N.J., Phillips, K.L.,Wilson, G., and Nichols, P.D. (2003) You are whatyou eat: describing the foraging ecology of southernelephant seals (Mirounga leonina) using blubber fattyacids. P Roy Soc Lond B Bio 270: 1283–1292.
Bray, J.R., and Curtis, J.T. (1957) An ordination of the uplandforest communities of Southern Wisconsin. Ecol Monogr27: 325–349.
The gut microbiota of wild and captive Antarctic seals 11
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

Bryden, M.M. (1971) Size and growth of viscera in the south-ern elephant seal, Mirounga leonina. Aust J Zool 19: 103–120.
Bureau of Meteorology (2011) Climate information. [WWWdocument]. URL http://www.bom.gov.au/climate/.
Carlini, A.R., Daneri, G.A., Marquez, M.E.I., Soave, G.E., andPoljak, S. (1997) Mass transfer from mothers to pups andmass recovery during the post-breeding foraging in south-ern elephant seals (Mirounga leonina) at King GeorgeIsland. Polar Biol 18: 305–310.
Carlini, A.R., Poljak, S., Daneri, G.A., Márquez, M.E.I., andPlötz, J. (2002) Dynamics of male dominance of southernelephant seals (Mirounga leonina) during the breedingseason at King George Island. Pol Polar Res 23: 153–159.
Carlini, A.R., Negrete, J., Daneri, G.A., Rogers, T.L.,Márquez, M.E.I., Ciaglia, M., and Mennucci, J.A. (2009)Immobilization of adult male southern elephant seals (Mir-ounga leonina) during the breeding and molting periodsusing a tiletamine/zolazepam mixture and ketamine. PolarBiol 32: 915–921.
Casaux, R., Baroni, A., Ramón, A., Carlini, A., Bertolin, M.,and DiPrinzio, C.Y. (2009) Diet of the leopard seal (Hydru-rga leptonyx) at the Danco Coast, Antarctic Peninsula.Polar Biol 32: 307–310.
Castillo, M., Martín-Orúe, S.M., Anguita, M., Pérez, J.F., andGasa, J. (2007) Adaptation of gut microbiota to corn physi-cal structure and different types of dietary fibre. Livest Sci109: 149–152.
Chao, A. (1984) Non-parametric estimation of the number ofclasses in a population. Scandinavian J Stat 11: 783–791.
Clarke, K.R., and Gorley, R.N. (2006) PRIMER Version 6:User Manual/Tutorial. Plymouth, UK: PRIMER-E.
Colwell, R. (1997) EstimateS Version 8.0 Statistical Estima-tion of Species Richness and Shared Species fromSamples. Storrs, CT, USA: Department of Ecology andEvolutionary Biology, University of Connecticut.
Davis, C.S., Delisle, I., Stirling, I., Siniff, D.B., and Strobeck,C. (2004) A phylogeny of the extant Phocidae inferred fromcomplete mitochondrial DNA coding regions. Mol Phylo-genet Evol 33: 363–377.
Denner, E.B.M., Mark, B., Busse, H.-J., Turkiewicz, M., andLubitz, W. (2001) Psychrobacter proteolyticus sp. nov.,a psychrotrophic, halotolerant bacterium isolated fromthe Antarctic krill Euphausia superba Dana, excreting acold-adapted metalloprotease. Syst Appl Microbiol 24:44–53.
Dethlefsen, L., and Relman, D.A. (2011) Incomplete recoveryand individualized responses of the human distal gutmicrobiota to repeated antibiotic perturbation. Proc NatlAcad Sci USA 108: 4554–4561.
Dethlefsen, L., McFall-Ngai, M., and Relman, D.A. (2007) Anecological and evolutionary perspective on human-microbemutualism and disease. Nature 449: 811–818.
Dethlefsen, L., Huse, S., Sogin, M.L., and Relman, D.A.(2008) The pervasive effects of an antibiotic on the humangut microbiota, as revealed by deep 16S rRNA sequencing.PLoS Biol 6: 2383–2400.
Dhanasiri, A., Brunvold, L., Brinchmann, M., Korsnes, K.,Bergh, Ø., and Kiron, V. (2011) Changes in the intestinalmicrobiota of wild Atlantic cod Gadus morhua L. uponcaptive rearing. Microb Ecol 61: 20–30.
Dowd, S.E., Sun, Y., Wolcott, R.D., Domingo, A., and Carroll,J.A. (2008) Bacterial tag-encoded FLX amplicon pyrose-quencing (bTEFAP) for microbiome studies: bacterialdiversity in the iIleum of newly weaned Salmonella-infectedpigs. Foodborne Pathog Dis 5: 459–472.
Eckburg, P., Bik, E., Bernstein, C., Purdom, E., Dethlefsen,L., and Sargent, M. (2005) Diversity of the human intestinalmicrobial flora. Science 308: 1635–1638.
Fanson, K.V., Wielebnowski, N.C., Shenk, T.M., Jakubas,W.J., Squires, J.R., and Lucas, J.R. (2010) Patterns oftesticular activity in captive and wild Canada lynx (Lynxcanadensis). Gen Comp Endocrinol 169: 210–216.
Ferron, F.A., Simoes, J.C., Aquino, F.E., and Setzer, A.W.(2004) Air temperature time series for King George Island,Antarctica. Pesq Antárt Bras (Braz Antarct Res) 4: 155–169.
Glad, T., Kristiansen, V.F., Nielsen, K.M., Brusetti, L., Wright,A.-D.G., and Sundset, M.A. (2010) Ecological characteri-sation of the colonic microbiota in Arctic and sub-Arcticseals. Microb Ecol 60: 320–330.
Guttman, L. (1968) A general nonmetric technique for findingthe smallest coordinate space for configuration of points.Psychometrika 33: 469–506.
Hall-Aspland, S.A., and Rogers, T.L. (2004) Summer diet ofleopard seals (Hydrurga leptonyx) in Prydz Bay, EasternAntarctica. Polar Biol 27: 729–734.
Hayashi, H., Sakamoto, M., and Benno, Y. (2002) Fecalmicrobial diversity in a strict vegetarian as determined bymolecular analysis and cultivation. Microbiol Immunol 46:819–831.
Higdon, J.W., Bininda-Emonds, O.R.P., Beck, R.M.D., andFerguson, S.H. (2007) Phylogeny and divergence of thepinnipeds (Carnivora: Mammalia) assessed using a multi-gene dataset. BMC Evol Biol 7: 216–235.
Higgins, D.P., Rogers, T.L., Irvine, A.D., and Hall-Aspland,S.A. (2002) Use of midazolam/pethidine and tiletamine/zolazepam combinations for the chemical restraint ofleopard seals (Hydrurga leptonyx). Mar Mamm Sci 18:483–499.
Hindell, M.A. (1991) Some life-history parameters of a declin-ing population of southern elephant seals, Miroungaleonina. J Anim Ecol 60: 119–134.
Hiruki, L.M., Schwartz, M.K., and Boveng, P.L. (1999)Hunting and social behaviour of leopard seals (Hydrurgaleptonyx) at Seal Island, South Shetland Islands, Antarc-tica. J Zool (Lond) 249: 97–109.
Hooper, L.V., and Gordon, J.I. (2001) Commensalhost-bacterial relationships in the gut. Science 292: 1115–1118.
Hooper, L.V., Midtvedt, T., and Gordon, J.I. (2002) Howhost-microbial interactions shape the nutrient environ-ment of the mammalian intestine. Annu Rev Nutr 22: 283–307.
Hurley, J.A., and Costa, D.P. (2001) Standard metabolic rateat the surface and during trained submersions in adultCalifornia sea lions (Zalophus californianus). J Exp Biol204: 3273–3281.
Huse, S.M., Welch, D.M., Morrison, H.G., and Sogin, M.L.(2010) Ironing out the wrinkles in the rare biospherethrough improved OTU clustering. Environ Microbiol 12:1889–1898.
12 T. M. Nelson, T. L. Rogers, A. R. Carlini and M. V. Brown
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

Kovacs, A., Ben-Jacob, N., Tayem, H., Halperin, E., Iraqi, F.,and Gophna, U. (2011) Genotype is a stronger determinantthan sex of the mouse gut microbiota. Microb Ecol 61:423–428.
Krockenberger, M.B., and Bryden, M.M. (1994) Rate ofpassage of digesta through the alimentry tract of southernelephant seals (Mirounga leonina) (Carnivora: Phocidae).J Zool (Lond) 234: 229–237.
Lane, D.J., Pace, B., Olsen, G.J., Stahl, D.A., Sogin, M.L.,and Pace, N.R. (1985) Rapid determination of 16S ribos-omal RNA sequences for phylogenetic analyses. Proc NatlAcad Sci USA 82: 6955–6959.
Lavigne, D.M., Kovacs, K.M., Hiby, L., and Boyd, I.L. (2007)Seals, sea lions, and walruses. In The Encyclopedia ofMammals. Macdonald, D.W. (ed.). Oxford, UK: Oxford Uni-versity Press, pp. 520–573.
Lewis, R., O’Connell, T.C., Lewis, M., Campagna, C., andHoelzel, A.R. (2006) Sex-specific foraging strategies andresource partitioning in the southern elephant seal (Mir-ounga leonina). P Roy Soc B-Biol Sci 273: 2901–2907.
Ley, R., Hamady, M., Lozupone, C., Turnbaugh, P., Ramey,R., Bircher, J., et al. (2008) Evolution of mammals and theirgut microbes. Science 320: 1647–1651.
Ley, R.E., Peterson, D.A., and Gordon, J.I. (2006) Ecologicaland evolutionary forces shaping microbial diversity in thehuman intestine. Cell 124: 837–848.
Lombardo, M.P. (2008) Access to mutualistic endosymbioticmicrobes: an underappreciated benefit of group living.Behav Ecol Sociobiol 62: 479–497.
López-García, P., López-López, A., Moreira, D., andRodri′guez-Valera, F. (2001) Diversity of free-livingprokaryotes from a deep-sea site at the Antarctic PolarFront. FEMS Microbiol Ecol 36: 193–202.
McCann, T.S. (1980) Population structure and social organi-sation of southern elephant seals, Mirounga leonina (L.).Biol J Linn Soc 14: 133–150.
Marteau, P., Pochart, P., Doré, J., Béra-Maillet, C., Bernalier,A., and Corthier, G. (2001) Comparative study of bacterialgroups within the human cecal and fecal microbiota. ApplEnviron Microbiol 67: 4939–4942.
Mårtensson, P.-E., Nordøy, E.S., Messelt, E.B., and Blix, A.S.(1998) Gut length, food transit time and diving habit inphocid seals. Polar Biol 20: 213–217.
Martinson, V.G., Danforth, B.N., Minckley, R.L., Rueppell, O.,Tingek, S., and Moran, N.A. (2011) A simple and distinctivemicrobiota associated with honey bees and bumble bees.Mol Ecol 20: 619–628.
Mentula, S., Harmoinen, J., Heikkila, M., Westermarck, E.,Rautio, M., Huovinen, P., and Kononen, E. (2005)Comparison between cultured small-intestinal and fecalmicrobiotas in beagle dogs. Appl Environ Microbiol 71:4169–4175.
Mortensen, A., and Tindall, A.R. (1981) Caecal decomposi-tion of uric acid in captive and free ranging willow ptarmi-gan (Lagopus lagopus lagopus). Acta Physiol Scand 111:129–133.
Nakamura, N., Amato, K.R., Garber, P., Estrada, A., Mackie,R.I., and Gaskins, H.R. (2011) Analysis of the hydrogeno-trophic microbiota of wild and captive black howlermonkeys (Alouatta pigra) in palenque national park,Mexico. Am J Primatol 73: 909–919.
Navarrete, P., Espejo, R., and Romero, J. (2009) Molecularanalysis of microbiota along the digestive tract of juvenileAtlantic salmon (Salmo salar L.). Microb Ecol 57: 550–561.
Nelson, K.E., Zinder, S.H., Hance, I., Burr, P., Odongo, D.,Wasawo, D., et al. (2003) Phylogenetic analysis of themicrobial populations in the wild herbivore gastrointestinaltract: insights into an unexplored niche. Environ Microbiol5: 1212–1220.
Ochman, H., Worobey, M., Kuo, C.-H., Ndjango, J.-B.N.,Peeters, M., Hahn, B.H., and Hugenholtz, P. (2010) Evo-lutionary relationships of wild hominids recapitulated by gutmicrobial communities. PLoS Biol 8: e1000546.
Palmer, C., Bik, E.M., DiGiulio, D.B., Relman, D.A., andBrown, P.O. (2007) Development of the human infant intes-tinal microbiota. PLoS Biol 5: 1556–1573.
Prabagaran, S.R., Manorama, R., Delille, D., and Shivaji, S.(2007) Predominance of Roseobacter, Sulfitobacter, Gla-ciecola and Psychrobacter in seawater collected offUshuaia, Argentina, sub-Antarctica. FEMS Microbiol Ecol59: 342–355.
Pruesse, E., Quast, C., Knittel, K., Fuchs, B.M., Ludwig, W.,Peplies, J., and Glöckner, F.O. (2007) SILVA: a compre-hensive online resource for quality checked and alignedribosomal RNA sequence data compatible with ARB.Nucleic Acids Res 35: 7188–7196.
Ramirez, R.F., and Dixon, B.A. (2003) Enzyme production byobligate intestinal anaerobic bacteria isolated from oscars(Astronotus ocellatus), angelfish (Pterophyllum scalare)and southern flounder (Paralichthys lethostigma). Aquac-ulture 227: 417–426.
Rangel-Negrín, A., Alfaro, J.L., Valdez, R.A., Romano, M.C.,and Serio-Silva, J.C. (2009) Stress in Yucatan spidermonkeys: effects of environmental conditions on fecal cor-ticol levels in wild and captive populations. Anim Conserv12: 496–502.
Rawls, J.F., Mahowald, M.A., Ley, R.E., and Gordon, J.I.(2006) Reciprocal gut microbiota transplants fromzebrafish and mice to germ-free recipients reveal hosthabitat selection. Cell 127: 423–434.
Rogers, T.L., and Bryden, M.M. (1995) Predation of Adéliepenguins (Pygoscelis adeliae) by leopard seals (Hydrurgaleptonyx) in Prydz Bay, Antarctica. Can J Zool 73: 1001–1004.
Rook, G.A.W., and Brunet, L.R. (2005) Microbes, immu-noregulation, and the gut. Gut 54: 317–320.
Rook, G.A.W., Adams, V., Hunt, J., Palmer, R., Martinelli, R.,and Brunet, L.R. (2004) Mycobacteria and other environ-mental organisms as immunomodulators for immuno-regulatory disorders. Springer Semin Immunopathol 25:237–255.
Rosen, D.A.S., Winship, A.J., and Hoopes, L.A. (2007)Thermal and digestive constraints to foraging behaviour inmarine mammals. Philos T Roy Soc B 362: 2151–2168.
Savage, D.C. (1977) Microbial ecology of the gastrointestinaltract. Annu Rev Microbiol 31: 107–133.
Schloss, P.D., Westcott, S.L., Ryabin, T., Hall, J.R., Hart-mann, M., Hollister, E.B., et al. (2009) Introducing Mothur:open-source, platform-independent, community-supportedsoftware for describing and comparing microbial commu-nities. Appl Environ Microbiol 75: 7537–7541.
The gut microbiota of wild and captive Antarctic seals 13
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

Scupham, A., Patton, T., Bent, E., and Bayles, D. (2008)Comparison of the cecal microbiota of domestic and wildturkeys. Microb Ecol 56: 322–331.
Siniff, D.B. (1991) An overview of the ecology of Antarcticseals. Am Zool 31: 143–149.
Slip, D. (1995) The diet of southern elephant seals(Mirounga leonina) from Heard Island. Can J Zool 73:1519–1528.
Southwell, C., Kerry, K., Ensor, P., Woehler, E.J., andRogers, T. (2003) The timing of pupping by pack-ice sealsin East Antarctica. Polar Biol 26: 648–652.
Staley, J.T., and Whitman, W.B. (2005) Phylum XIX. Fuso-bacteria Garrity and Holt 2001, 140. In Bergey’s Manual ofSystematic Bacteriology Volume 4: The Bacteroidetes, Spi-rochaetes, Tenericutes (Mollicutes), Acidobacteria, Fibro-bacteres, Fusobacteria, Dictyoglomi, Gemmatimonadetes,Lentisphaerae, Verrucomicrobia, Chlamydiae, and Planc-tomycetes. Krieg, N.R., Ludwig, W., Whitman, W.B.,Hedlund, B.P., Paster, B.J., Staley, J.T., et al. (eds). NewYork, USA: Springer, pp. 747–774.
Thompson, C.L., Wang, B., and Holmes, A.J. (2008) Theimmediate environment during postnatal development haslong-term impact on gut community structure in pigs. ISMEJ 2: 739–748.
Tsukinowa, E., Karita, S., Asano, S., Wakai, Y., Oka, Y.,Furuta, M., and Goto, M. (2008) Fecal microbiota of adugong (Dugong dugong) in captivity at Toba Aquarium. JGen Appl Microbiol 54: 25–38.
Turnbaugh, P., Ley, R., Mahowald, M., Magrini, V., Mardis, E.,and Gordon, J. (2006) An obesity-associated gut microbi-ome with increased capacity for energy harvest. Nature444: 1027–1031.
Turnbaugh, P.J., Hamady, M., Yatsunenko, T., Cantarel, B.L.,Duncan, A., Ley, R.E., et al. (2009) A core gut microbiomein obese and lean twins. Nature 457: 480–485.
Uenishi, G., Fujita, S., Ohashi, G., Kato, A., Yamauchi, S.,Matsuzawa, T., and Ushida, K. (2007) Molecular analysesof the intestinal microbiota of chimpanzees in the wild andin captivity. Am J Primatol 69: 367–376.
Vael, C., and Desager, K. (2009) The importance of thedevelopment of the intestinal microbiota in infancy. CurrOpin Pediatr 21: 794–800.
Villers, L.M., Jang, S.S., Lent, C.L., Lewin-Koh, S.-C.,and Norosoarinaivo, J.A. (2008) Survey and comparisonof major intestinal flora in captive and wild ring-tailedlemur (Lemur catta) poulations. Am J Primatol 70: 175–184.
Walker, T.R., Boyd, I.L., McCafferty, D.J., Huin, N., Taylor,R.I., and Reid, K. (1998) Seasonal occurrence and diet ofleopard seals (Hydrurga leptonyx) at Bird Island, SouthGeorgia. Antarct Sci 10: 75–81.
Wall, R., Ross, R.P., Ryan, C.A., Hussey, S., Murphy, B.,Fitzgerald, G.F., and Stanton, C. (2009) Role of gut
microbiota in early infant development. Clin Med Pediatr2009: 45–54.
Wang, Q., Garrity, G.M., Tiedje, J.M., and Cole, J.R. (2007)Naive bayesian classifier for rapid assignment of rRNAsequences into the new bacterial taxonomy. Appl EnvironMicrobiol 73: 5261–5267.
Ward, N., Steven, B., Penn, K., Methé, B., and Detrich, W.(2009) Characterization of the intestinal microbiota of twoAntarctic notothenioid fish species. Extremophiles 13:679–685.
Wienemann, T., Schmitt-Wagner, D., Meuser, K., Segel-bacher, G., Schink, B., Brune, A., and Berthold, P. (2011)The bacterial microbiota in the ceca of capercaillie (Tetraourogallus) differs between wild and captive birds. Syst ApplMicrobiol 34: 542–551.
Williams, T.M., Haun, J., Davis, R.W., Fulman, L.A., andKohin, S. (2001) A killer appetite: metabolic consequencesof carnivory in marine mammals. Comp Biochem Physiol AMol Integr Physiol 129: 785–796.
Xenoulis, P.G., Gray, P.L., Brightsmith, D., Palculict, B.,Hoppes, S., Steiner, J.M., et al. (2010) Molecular charac-terization of the cloacal microbiota of wild and captiveparrots. Vet Microbiol 146: 320–325.
Yildirim, S., Yeoman, C.J., Sipos, M., Torralba, M., Wilson,B.A., Goldberg, T.L., et al. (2010) Characterization of thefecal microbiome from non-human wild primates revealsspecies specific microbial communities. PLoS ONE 5:e13963.
Zhang, H., DiBaise, J.K., Zuccolo, A., Kudrna, D., Braidotti,M., Yu, Y., et al. (2009) Human gut microbiota in obesityand after gastric bypass. Proc Natl Acad Sci USA 106:2365–2370.
Supporting information
Additional Supporting Information may be found in the onlineversion of this article:
Fig. S1. Gut microbiota of southern elephant seals of differ-ent age-classes. nMDS plots the relative dissimilarity of anindividuals’ gut microbiota.Table S1. Effect of age-class on the gut microbiota ofsouthern elephant seals by ANOSIM.Table S2. Effect of mass (kg) on the gut microbiota ofsouthern elephant seals by PERMANOVA.Table S3. OTUs shared in the gut microbiota of most wildleopard seals.Table S4. OTUs shared in the gut microbiota of most south-ern elephant seal adults and sub-adults.Table S5. Shared OTUs in the gut microbiota of captiveleopard seals over time.Table S6. Characteristics of seals used for samplecollection.
14 T. M. Nelson, T. L. Rogers, A. R. Carlini and M. V. Brown
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology
Copyright © 2022 FDOKUMEN




















