
Die Tierknochen. In: C. Schucany, Die römische Villa von Biberist-Spitalhof/SO (Grabungen 1982,...
75
Transcript of Die Tierknochen. In: C. Schucany, Die römische Villa von Biberist-Spitalhof/SO (Grabungen 1982,...



DIE RÖMISCHE VILLA VON BIBERIST-SPITALHOF/SO (Grabungen 1982, 1983, 1986-1989)
Untersuchungen im Wirtschaftsteil und Überlegungen zum Umland
VON
CATY SCHUCANY
mit Beiträgen von Richard Delage, Eckhard Deschler-Erb, Sabine Deschler-Erb, Cornel Doswald, Gaële Feret, Suzanne Frey-Kupper, Sylvia Fünfschilling,
Adrian Huber, Heide Hüster Plogmann, Stefanie Jacomet, Bruno Kaufmann, Marianne Petrucci-Bavaud, Angela Schlumbaum, Vincent Serneels,
Jorge E. Spangenberg, Richard Sylvestre, Stephan Wyss
digitaler Sonderdruck
Verlag Bernhard Albert Greiner2006

Der Druck des vorliegenden Bandes wurde ermöglicht durch den
Lotteriefonds des Kantons Solothurn
digitaler Sonderdruck
ISBN-10 3-935383-90-8 (Buchpublikation)ISBN-13 978-3-935383-90-5 (Buchpublikation)
1. Auflage 2006
© 2006 by Verlag Bernhard Albert Greiner, 73630 Remshaldenhttp://www.bag-verlag.de
Redaktion Caty Schucany, Bern und Bernhard A. und Claudia Greiner, Remshalden
Digitalisierung & Peter Burry, Repro von Ballmoos, SolothurnFotobearbeitung Jürg Stauffer, Solothurn
Umschlaggestaltung Bernhard A. Greiner, Remshalden & Caty Schucany, Bern
Buch-Layout und Satz Bernhard A. Greiner, Remshalden& Bildbearbeitung
Druck Offset-Druckerei Gunter Dünnbier, 02779 Großschönauhttp://www.druckerei-duennbier.com
Das Werk ist in allen seinen Teilen urheberrechtlich geschützt. Jede Verwertung ist ohne Zustimmung des Verlages unzulässig. Das gilt insbesondere für Vervielfältigungen, Übersetzungen, Mikroverfilmungenund die Einspeicherung und Verarbeitung in elektronischen Systemen.
Titelbild Amt für Verkehr und Tiefbau des Kantons Solothurn, Grabungskampagne 1989
Grafik Caty Schucany, Bern
Ein Titeldatensatz für diese Publikationist bei der Deutschen Nationalbibliothek erhältlich.

BAND I – BEFUND UND AUSWERTUNGZusammenfassung 11Résumé 17Summary 23
I. EINLEITUNG 29I. Einleitung 311. Lage 332. Ältere Nachrichten 353. Die archäologischen Untersuchungen von 1982-1989 37
3.1. Ausgrabung 373.1.1. Sondierung 1982 373.1.2. Flächengrabungen 1982/1983 373.1.3. Flächengrabungen 1986-1989 373.2. Arbeitsmethoden 393.2.1. Grabungstechnik 393.2.2. Fundbergung 393.2.3. Dokumentation 393.3. Auswertung 403.3.1. Vorarbeiten 403.3.2. Auswertungsprojekt 423.3.3. Manuskript 43
4. Erhaltungsbedingungen 454.1. Erhaltung der Befunde 454.2. Erhaltung der Funde 46
5. Topographie 475.1. Geologischer Untergrund 475.2. Römischer Oberboden 475.3. Antikes Geländerelief 47
6. Vorrömische Befunde und Funde 516.1. Steinzeitliche Funde und Befunde 516.1.1. Silices 516.1.2. Steinmeissel 516.1.3. Brandgrube 516.2. Bronzezeitliche Funde und Befunde 536.2.1. Funde aus dem Verwitterungslehm 536.2.2. Mutmassliches Grab 546.2.3. Furt 546.2.4. Holz 556.2.5. Fazit 55
INHALT
V

VI
6.3. Die keltischen Münzen 566.3.1. Befund 566.3.2. Münzen (Suzanne Frey-Kupper) 57
7. Nachrömische Befunde und Funde 617.1. Gräber 617.2. Pfostenbauten und Strassenreste nördlich der römischen Villa (Periode II) 627.3. Gewerbeanlagen 647.4. Mittelalterliches und neuzeitliches Glas (Sylvia Fünfschilling) 667.5. Nachrömische Eisenfunde (Adrian Huber) 707.5.1 Frühmittelalterliche Eisenfunde 707.5.2. Mittelalterliche oder frühneuzeitliche Eisenfunde 72
II. BEFUND UND BAUSTRUKTUREN 738. Einleitung 759. Periode I 77
9.1. Hofmauer 779.2. Gutshofbach 799.3. Aussenniveau 799.4. Werkstatt B-Phase A 80
10. Periode II 8510.1. Anlage 8510.1.1. Hofbegrenzung 8510.1.2. Trennmauer 9310.1.3. Gutshofbach 9510.1.4. Strassen 9510.1.5. Pars urbana 10110.2. Kernzone 10410.2.1. Portikus F 10410.2.2. Gebäude G 11010.2.3. Grabanlage H 11310.2.4. Leichte Bauten 13010.2.5. Aussenniveaus und Gruben 13110.3. östliche Randzone 13210.3.1. Werkstatt B-Phase B 13210.3.2. Werkstatt B-Phase C 13610.3.3. Gebäude A 14110.3.4. Gebäude L 14210.3.5. Holzbau J 14510.3.6. Steinbau J 14910.3.7. Vorplatz westlich des Gebäudes J 16110.3.8. Galerie M 16510.3.9. Gebäude P 16610.3.10. Gebäude O 16910.3.11. Ausserhalb des Gebäudes O 17510.3.12. Gebäude Q 18110.3.13. Leichte Bauten 18510.4. nördliche Randzone 18710.4.1. Gebäude D 18710.4.2. Gebäude C 19710.4.3. Gebäude E 199

VII
11. Periode III 20711.1. Einleitung 20711.2. Innerhalb Gebäude D 20711.3. Gebäude K 20911.4. Gebäude N 211
III. DATIERUNG 21312. Einleitung 215
13. Periode I 21713.1. Gehniveau bei der Nordostecke der Hofmauer 21713.2. Werkstatt B-Phase A 21713.2.1. Bauhorizont 21713.2.2. Benützungsschicht 218
14. Periode II 21914.1. Hofbegrenzung 21914.1.1. Graben 21914.1.2. Hofmauer und Tor 21914.2. Strassen 21914.3. Gebäude A 22014.4. Werkstatt B 22114.4.1 Phase B 22114.4.2. Phase C 22214.5. Gebäude C 22214.6. Gebäude D 22214.7. Gebäude E 22414.8. Portikus F 22514.9. Gebäude G 22614.10. Grabanlage H 22614.10.1. Brandschicht 22614.10.2. Opfergruben 22914.10.3. Zeugen einer späteren Begehung 22914.11. Holzbau J 23014.12. Steinbau J 23114.12.1. Phase 1 23114.12.2. Phase 2 23214.12.3. Phase 3 23214.12.4. Phase 4 23314.13. Westlich des Steinbaus J 23414.13.1. Phase 1 23414.13.2. Phase 2 23514.13.3. Phase 3 23514.14. Gebäude L 23514.15. Galerie M 23514.16. Gebäude O 23514.16.1. Phase 1 23614.16.2. Phase 2 23614.16.3. Phase 3 23714.16.4. Phase 4 23814.17. Westlich und südlich des Gebäudes O 23914.17.1. Phase 1 23914.17.2. Phase 2 24014.17.3. Phase 3 241

VIII
14.18. Gebäude P 24214.19. Gebäude Q 24214.20. Aussenniveau und Gruben 24314.20.1. Kernzone 24314.20.2. Östliche Randzone 244
15. Periode III 24515.1. Gebäude K 24515.2. Gebäude N 246
IV. WEITERFÜHRENDE BETRACHTUNGEN 24716. Fragestellungen 249
17. Siedlungsgeschichte 25117.1. Einleitung 25117.2. Vor der römischen Gutshofanlage 25117.3. Periode I der römischen Gutshofanlage 25317.4. Periode II der römischen Gutshofanlage 25317.4.1. Phase 1 25317.4.2. Phase 2 25517.4.3. Phase 3 25717.4.4. Phase 4 25917.4.5. Phase 5 25917.4.6. Phase 6 26317.4.7. Phase 7 26317.5. Periode III des römischen Gutshofes 26917.6. Nach dem römischen Gutshof 26917.6.1. Dorf und Gräberfelder des 6./7. Jahrhunderts 26917.6.2. Der Spitalhof des 17. Jahrhunderts 270
18. Überlegungen zum Gutsbetrieb 27118.1. Einleitung 27118.1.1. Gesellschaftlicher Hintergrund 27118.1.2. Die antiken Agrarschriftsteller 27218.2. Umland 27318.2.1. Grenzen und Vermessung 27318.2.2. Gliederung der Landschaft 27418.2.3. Organisation des Fundus 27418.2.4. Bewirtschaftung 27818.3. Gutsbetrieb 28218.3.1. Einleitung 28218.3.2. Periode I 28418.3.3. Periode II-Phase 1 28418.3.4. Periode II-Phase 2 28518.3.5. Periode II-Phasen 3-5 28618.3.6. Periode II-Phasen 6 28718.3.7. Periode II-Phase 7 28818.3.8. Periode III 28918.4. Import – Export 28918.4.1. Einleitung 28918.4.2. Auswärtige Güter 28918.4.3. Zum Export produzierte Güter 29018.4.4. Zur Bedeutung des Handwerks im Gutsbetrieb 29118.4.5. Fazit 291

IX
BAND II – FUNDE UND BESONDERE FUNDENSEMBLES
V. FUNDE UND BESONDERE FUNDENSEMBLES 30319. Einleitung 305
20. Die römischen Münzen (Suzanne Frey-Kupper) 30720.1. Einleitung 30720.2. Münzen aus stratigraphischem Zusammenhang 30720.2.1. Voraussetzungen 30720.2.2. Münzen aus Schichten des späten 1. bis mittleren 2. Jahrhunderts 30720.2.3. Münzen aus Schichten des späten 2. bis mittleren 3. Jahrhunderts 30820.3. Das Ende der Gutshofanlagen und das Ende der Münzreihe 31020.4. Verbreitung der Münzen in den Gebäuden 31120.5. Altfunde 31120.5.1. Einzelfunde 31120.5.2. Hort oder Börse 31220.6. Zusammenfassung 312
21. Einige Aspekte der Gefässkeramik 31521.1. Die Keramikkategorien 31521.1.1. Definition 31521.1.2. Entwicklung der Keramikensembles 31621.2. La céramique sigillée moulée de la villa de Biberist-Spitalhof (Richard Delage) 31821.2.1. Introduction 31821.2.2. Approvisionnement du site en céramique sigillée moulée 31921.2.3. Les groupes de production 31921.2.4. Conclusions 32521.3. Amphoren (Stephan Wyss) 35121.3.1. Einleitung und Vorgehen 35121.3.2. Überblick 35121.3.3. Herkunft, Form und Inhalt 35121.3.4. Die importierten Waren 36121.3.5. Die Amphoren aus Grabanlage H 36121.3.6. Zusammenfassung 36321.4. Les graffites sur céramique (Gaële Feret et Richard Sylvestre) 36521.4.1. Introduction 36521.4.2. Des noms et des signes 36521.4.3. Graffites et céramiques 36921.4.4. Graffites et société 37121.4.5. Conclusion 37121.5. Varia 37221.5.1. Gesichtskrug 37221.5.2. Lampe mit Brennstoffrückständen (Jorge E. Spangenberg) 37221.6. Besondere Ensembles 37521.6.1. Das Keramikinventar der Grabanlage H 37521.6.2. Das Keramikinventar des Gebäudes E 38121.6.3. Das Keramikinventar des Holzbaus J 38221.6.4. Das Keramikinventar des Steinbau J-Phase 4 38421.6.5. Das Keramikinventar des Gebäudes O-Phase 4 38521.6.6. Fazit 386

X
22. Die Gläser (Sylvia Fünfschilling) 38722.1. Allgemeine Bemerkungen 38722.2. Glasgefässe 38922.2.1. Formen des 1. Jahrhunderts 38922.2.2. Formen des späteren 1. Jahrhunderts, des 2. Jahrhunderts und jünger 39022.2.3. Glasgefässe unsicherer Zeitstellung 40722.3. Übriges Glas 41122.3.1. Fensterglas 41122.3.2. Perlen 41122.3.3. Varia 41122.4. Glasensembles 41122.4.1. Gebäude A 41122.4.2. Werkstatt B 41122.4.3. Gebäude D 41122.4.4. Gebäude E. 41222.4.5. Grabanlage H 41222.4.6. Holzbau J 41422.4.7. Steinbau J 41422.4.8. Gebäude K 41522.4.9. Gebäude O 41522.4.10. Gebäude P 41522.4.11. Gebäude Q 41522.5. Zusammenfassende Bemerkungen 416
23. Die Funde aus Bronze und Blei (Eck4hard Deschler-Erb) 41723.1. Typologische und chronologische Einordnung 41723.1.1. Hausrat 41723.1.2. Toilettgerät 42723.1.3. Schmuck und Tracht 42923.1.4. Militaria und Transportwesen 44223.1.5. Kleingerät und Fragmente 44523.1.6. Metallverarbeitung 44823.1.7. Funktion unbekannt 44923.1.8. Figürliches 45123.2. Schlussfolgerungen 45223.2.1. Gesamtspektrum 45223.2.2. Verteilung der Funde aus Buntmetall im Villenareal 457
und Überlegungen zur Funktion der betreffenden Häuser
24. Die Funde aus Eisen (Adrian Huber) 45924.1. Einleitung 45924.1.1. Erhaltung 45924.1.2. Sichtung, Auswahl und Dokumentation 45924.2. Verteilung auf die Zeitabschnitte 46024.3. Verteilung auf die Gebäude 46324.4. Typen und Funktion 46424.4.1. Schmuck und Trachtbestandteile 46424.4.2. Toilettgerät 46524.4.3 Stili 46624.4.4. Hausrat und Mobiliar 46824.4.5. Landwirtschaftliches Gerät 48224.4.6. Geräte zur Textil und Lederverarbeitung 48424.4.7. Geräte zur Holzbearbeitung 486

XI
24.4.8. Geräte zur Metallverarbeitung 48924.4.9. Multifunktionale Geräte 49224.4.10. Waffen 49424.4.11. Pferd und Wagen 49824.4.12. Zeugnisse der Eisenverarbeitung und Barren 50224.4.13. Baueisen 50224.4.14. Ketten und Ringe 50424.4.15. Stäbe 50724.4.16. Bänder, Beschläge, Platten und Bleche 50824.4.17. Objekte unbekannter Funktion 51024.5. Zusammenfassende Betrachtungen 51324.5.1. Anteile der Funktionsgruppen 51324.5.2. Räumliche Verteilung der Funktionsgruppen 51324.5.3. Wohn- oder Wirtschaftsbau? - Verhältnis von Hausrat/Mobiliar zu Gerät/Werkzeug 519
25. Les déchets métallurgiques (Vincent Serneels) 52125.1. Introduction 52125.1.1. Répartition spatiale et chronologique 52125.1.2. Représentativité des déchets métallurgiques récoltés 52125.2. Les déchets provenant du bâtiment E 52225.3. Les déchets provenant de l'atelier de forge (atelier B) 52325.3.1. Le forgeage du fer et ses déchets 52425.3.2. Les parois de foyer 52525.3.3. Les scories 52625.3.4. Les scories en forme de calotte 52725.3.5 Les matériaux mis en oeuvre 52925.3.6 Approche quantitative de la première période de fonctionnement de l'atelier B (phase B) 53125.4 Conclusion 532
26. Baumaterialien 53526.1. Einleitung 53526.2. Lehmfachwerk 53526.3. Architekturfragmente aus Stein 53526.4. Verputz 53826.5. Tonsäule 53926.6. Bodenplättchen aus Ton 53926.7. Bestandteile beheizbarer Räume und Heizanlagen 54026.7.1. Tonplatten abgehobener Böden 54026.7.2. Bestandteile hohler Wände 54126.7.3. Terrazzomörtel 542
27. Varia 54327.1. Terrakotten 54327.2. Webgewichte aus Ton 54427.3. Lavezgefässe 54527.3.1. Introduction (Vincent Serneels) 54527.3.2. Repertoire und Datierung 54727.3.3. Description pétrographique des pierre ollaire (Vincent Serneels) 54727.3.4. Quelques réflexions sur les pierres ollaires de Biberist (Vincent Serneels) 54927.3.5. Fazit 54927.4. Die Steingeräte (Cornel Doswald) 55327.4.1. Einleitung 55327.4.2. Mühlsteine 55527.4.3. Mörser 559

XII
27.4.4. Wetz- und Schleifsteine 55927.4.5. Drehpfannen 56027.5. Beinartefakte (Sabine Deschler-Erb) 56027.5.1. Ensemble 56027.5.2. Typologie und Chronologie 56127.5.3. Rohmaterial 57027.5.4. Technologie 57027.5.5. Verteilung im Gutshofareal 57127.5.6. Zusammenfassung 57127.6. Blaue Pigmentkugeln 57227.7. Zusammenfassende Bemerkungen zu den Objekten aus Metall und Bein 573
(Eckhard Deschler-Erb, Sabine Deschler-Erb und Adrian Huber)
28. Samen und Früchte 579(Stefanie Jacomet und Marianne Petrucci-Bavaud, unter Mitarbeit von Marlu Kühn)
28.1. Einleitung 57928.1.1. Forschungsstand 57928.1.2. Ziele 57928.2. Material und Methoden 57928.2.1. Entnahme, Aufbereitung, Datierung und Herkunft der Proben, Erhaltung 57928.2.2. Volumina des untersuchten Materials 58128.2.3. Schlämmtechnik, Auslesen und Bestimmung 58128.2.4. Auswertungsmethoden 58228.2.5. Bemerkung zu den Bestimmungen 58228.3. Übergreifende Betrachtungen 58228.3.1. Einfluss der Schlämm-Methoden auf die Funddichte und das Artenspektrum 58228.3.2. Erhaltung der Reste und deren Einfluss auf die Repräsentanz der Taxa 58328.3.3. Fundspektrum 58328.4. Aussagemöglichkeiten der Archäobotanik zur Interpretation von Schichten und Strukturen 58528.4.1. Einleitung 58528.4.2. Brandschichten inkl. Feuerstellen 58628.4.3. Brandschutt 58728.4.4. Dachschutt 58728.4.5. Gruben- und Grabenfüllungen 58728.4.6. Abfallschichten 58828.4.7. Horizonte und Böden 58828.4.8. Planien und Auffüllungen 58828.4.9. Fazit 58828.5. Die Pflanzenreste aus Gebäuden und Schichten 59028.5.1. Einleitung 59028.5.2. Gebäude D 59028.5.3. Gebäude E 59428.5.4. Grabanlage H 59528.5.5. Holzbau J 59828.5.6. Steinbau J und Umgebung 60528.5.7. Gebäude O und Umgebung 60728.5.8. Gebäude Q 61328.5.9. Werkstatt B 61428.5.10 Pfostenbau 2 61428.5.11 Gruben im Bereich des Tors 61428.6. Weiterführende Betrachtungen 61428.6.1. Produktion und Verarbeitung 61428.6.2. Speisezettel 61528.6.3. Aussagen der Wildpflanzen zum Ackerbau, den Feldern und der Herkunft des Erntegutes 617

XIII
29. Verkohlte Holzreste (Angela Schlumbaum) 62529.1. Einleitung 62529.1.1. Ziel 62529.1.2. Forschungsstand 62529.1.3. Material und Methoden 62529.1.4. Zur Entstehung der Holzkohle (Caty Schucany) 62629.2. Übersicht 62729.2.1. Die Hölzer 62729.2.2. Herkunft der Hölzer 62729.2.3. Vergleich von Holzproben und Schlämmproben 62729.3. Die Holzarten der einzelnen Ensembles 62829.3.1. Werkstatt B-Phase B 62829.3.2. Gebäude D 62829.3.3. Gebäude E 63029.3.4. Grabanlage H 63029.3.5. Holzbau J 63029.3.6. Steinbau J 63129.3.7. Gebäude K 63129.3.8. Gebäude O und Umgebung 63129.3.9. Haus Q 63229.3.10. Bau 2 in der östlichen Randzone 63229.4. Vergleich der Fundschichten 63229.4.1 Abfallschichten 63229.4.2. Gruben 63229.4.3. Horizonte und Böden 63229.4.4. Brandschichten 63229.4.5. Brandschutt 63329.4.6. Dachschutt 63329.4.7. Mauerschutt 63329.5. Zusammenfassung 634
30. Die Tierknochen 635(Sabine Deschler-Erb, mit Beiträgen von Heide Hüster Plogmann, Bruno Kaufmann und Jorge Spangenberg)
30.1. Einleitung 63530.1.1. Häufigkeit, Verteilung und Zusammensetzung des Materials 63530.1.2. Erhaltungszustand 63630.2. Die Tierknochen der einzelnen Gebäude 64030.2.1. Gebäude A 64030.2.2. Werkstatt B 64030.2.3. Gebäude D 64230.2.4. Gebäude E 64530.2.5. Portikus F 64630.2.6. Grabanalage H (Bruno Kaufmann) 64630.2.7. Gebäude J und Umgebung 64730.2.8. Gebäude K 65030.2.9. Gebäude O und Umgebung 65130.2.10. Gebäude P 65230.2.11. Gebäude Q 65230.2.12. Tor und Umgebung 65430.2.13. Rinderkadaver in der Kernzone 65530.3. Die Tierarten im Wandel der Zeit 65630.3.1. Einleitung 65630.3.2. Hausrind 657

XIV
30.3.3. Ovicapriden 65830.3.4. Hausschwein 66030.3.5. Verhältnis der wichtigsten Haustiere zueinander 66030.3.6. Equiden 66130.3.7. Hund 66130.3.8 Hausgeflügel 66130.3.9. Wildtiere 66130.4. Schlussbetrachtungen zur Auswertung der handaufgelesenen Tierknochen 66430.5. Die Tierknochen aus den Schlämmproben (Heide Hüster Plogmann) 66530.5.1. Einleitung 66530.5.2. Spektrum der Tierreste 66630.5.3. Die Ensembles der einzelnen Gebäude 67030.5.4. Schlussbetrachtung 67430.6. Verkohlte Masse aus dem Konservierungsofen in Gebäude Q (Jorge Spangenberg) 67530.6.1. Einleitung 67530.6.2. Material and methods 67530.6.3. Results and discussion 676
31. Anthropologische Untersuchungen (Bruno Kaufmann) 67931.1. Vorbemerkung 67931.2. Der Leichenbrand aus der Grabanlage H 67931.2.1. Einleitung 67931.2.2. Besprechung der einzelnen Kriterien 67931.2.3. Beurteilung der menschlichen Leichenbrände 68131.2.4. Krankhafte Befunde am Skelett ("Pathologika") 68331.2.5. Brandstufen und Brandtemperaturen 68331.2.7. Zusammenfassung der Ergebnisse 68331.3. Die unverbrannten menschlichen Skelettreste aus der Grabanlage H 68531.4. Reste von Säuglignsbestattungen 68531.4.1. Säugling bei Portikus F 68531.4.2. Reste von Säuglingen aus Gebäude E und Umgebung sowie aus Steinbau J 68531.5. Reste erwachsener Menschen 68631.5.1. Portikus F 68631.5.2. Gebäude O-Phase 4 686

XV
BAND III – KATALOGEVI.KATALOGE 693
32. Einleitung zum Befundkatalog 69532.1. Bemerkungen zu Katalog und Schichtansprache 69532.2. Liste der Schichten 695
33. Katalog der vor- und nachrömischen Befunde 70133.1. Vorrömische Befunde 70133.1.1. Steinzeitliche Befunde und Funde 70133.1.2. Bronzezeitliche Befunde und Funde 70133.2. Nachrömische Befunde 70133.2.1. Befunde nördlich der Hofmauer (Periode II) 70133.2.2. Gewerbeanlagen 703
34. Befundkatalog des ersten Gehöfts (Periode I) 70534.1. Hofmauer 70534.2. Gutshofbach 70534.3. Aussenniveau 70534.4. Werkstatt B-Phase A 705
35. Befundkatalog der grossen Villa (Periode II) 70935.1. Anlage 70935.1.1. Begrenzung 70935.1.2. Tor und Umgebung 71035.1.3. Trennmauer 71035.1.4. Gutshofbach 71135.1.5. Strassen 71135.1.6. Pars urbana 71335.2. Gebäude A 71335.3. Werkstatt B 71335.3.1. Phase B 71335.3.2. Phase C 71435.4. Gebäude C 71435.5. Gebäude D 71535.6. Gebäude E 71735.7. Portikus F 71935.8. Gebäude G 71935.9. Grabanlage H 72035.10. Gebäude J und Umgebung 72135.10.1. Holzbau J 72135.10.2. Steinbau J 72235.10.3. Vorplatz westlich von Steinbau J 72635.11. Gebäude L 72635.12. Galerie M 72735.13. Gebäude O und Umgebung 72735.13.1. Gebäude O 72735.13.2. Ausserhalb des Gebäudes O 73035.14. Gebäude P 73235.15. Gebäude Q 73235.16. Leichte Bauten und Aussenniveaus 73435.16.1. Kernzone 73435.16.2. Östliche Randzone 735
36. Befundkatalog des nachfolgenden Gehöfts (Periode III) 73736.1. Innerhalb von Gebäude D 73736.2. Gebäude K 73736.3. Gebäude N 738

XVI
37. Katalog der Münzen 73937.1. Vorbemerkungen 73937.2. Ensemble aus der Ausgrabung 1982-1989 73937.2.1. Kelten, Nordwest- und Westschweiz 73937.3. Einzelfunde aus der Ausgrabung 1982-1989 73937.3.1. Kelten, östliches Mittelgallien 73937.3.2. Rom, Kaiserreich 73937.3.3. Bern, Kanton 74537.4. Katalog der Altfunde 74537.4.1. Im Original noch greifbare Funde 74537.4.2. Im Original nicht mehr greifbar 746
38. Katalog von stratifizierten Funden 74738.1. Vorbemerkung 74738.2. Periode I 74738.2.1. Nordöstlich der Hofmauer Taf. 7 74738.2.2. Werkstatt B-Phase A Taf. 7-10 74738.3. Periode II 74938.3.1. Gruben beim Tor Taf. 10 74938.3.2. Gebäude A Taf. 11 74938.3.3. Werkstatt B-Phase B Taf. 11-17 75038.3.4. Werkstatt B-Phase C Taf. 17 75338.3.5. Gebäude D Taf. 17-22 75338.3.6. Gebäude E Taf. 22-39 75738.3.7. Grabanlage H Taf. 39-53 75838.3.8. Holzbau J Taf. 53-60 78038.3.9. Steinbau J-Phase 1 Taf. 60 78638.3.10. Steinbau J-Phase 2 Taf. 61 78638.3.11. Steinbau J-Phase 3 Taf. 61-62 78638.3.12. Steinbau J-Phase 4 Taf. 62-81 78738.3.13. Gebäude L Taf. 81-83 79938.3.14. Gebäude O-Phase 1 Taf. 83-84 80038.3.15. Gebäude O-Phase 2 Taf. 84-88 80138.3.16. Gebäude O-Phase 3 Taf. 88-96 80438.3.17. Gebäude O-Phase 4 Taf. 96-102 80938.3.18. Ausserhalb des Gebäudes O Taf. 102-108 81338.3.19. Gebäude P Taf. 108-113 81838.3.20. Gebäude Q Taf. 113-119 82038.3.21. Aussenniveau im Villenareal Taf. 119 82538.4. Periode III 82538.4.1. Gebäude K Taf. 120-121 825
39. Katalog der Skelettreste 82739.1. Der Leichenbrand aus der Grabanlage H 82739.2. Unverbrannte menschliche Überreste aus verschiedenen Fundzusammenhängen 83339.2.1. Säugling und weitere Überreste aus Portikus F 83339.2.2. Reste von Säuglingen aus Gebäude E und Umgebung sowie aus Steinbau J 83339.2.3. Reste erwachsener Menschen in Gebäude O-Phase 4 833
40. Anhang (Tabellen – Abkürzungen – Literatur) 83540.1. Tabellen 83540.2. Abkürzungen 94540.3. Literaturverzeichnis 947
TAFELN Taf. 1 – 121



635
30.1. Einleitung
M it über 11’000 untersuchten Tierknochen (ohne dieSchlämmfunde: s. Kap. 30.5) gehört die römische Villa von
Biberist zu den derzeit fundreichsten archäozoologisch unter-suchten Gutshöfen in den nordwestlichen Provinzen2012. Nebender Keramik sind die Tierknochen die häufigste Fundgattung.Obwohl es sich um eine Rettungsgrabung handelt, wurde ihnenwährend und nach der Ausgrabung die gleiche Aufmerksamkeitwie den übrigen Funden entgegengebracht. Die sorgfältige Aus-grabung sowie die Befundauswertung ermöglichen es, die Tier-knochenfunde sowohl in ihrer vertikalen wie horizontalen Ver-teilung detailliert zu analysieren. Mangels Ausgrabung wissen wirzwar nichts über die Verarbeitung und Konsumation tierischerProdukte in der Residenz (pars urbana). Auch lassen die unter-suchten Gebäude des Ökonomiebereichs (pars rustica) kaumSchlüsse auf die übrigen, nicht ausgegrabenen Bereiche zu, denndie Befund- und Fundanalysen zeigen deutlich die Individua-lität eines jeden einzelnen Gebäudes. Trotzdem gewährt die vor-liegende Untersuchung interessante Einblicke in die Wirtschafts-form, Sozialstruktur und geistige Welt einer römischen Villa inden nordwestlichen Provinzen.
30.1.1. Häufigkeit, Verteilung und Zusammensetzung des Materials
Der Menge und der Kosten wegen haben wir nicht alle stra-tifizierten Tierknochen untersuchen können. Insbesondere diez.T. fundreichen Planie- und Auffüllschichten im Umfeld derGebäude J, P, O und Q mussten wir mit wenigen Ausnahmen bei-seite lassen. Untersucht haben wir die Knochen aus allen Gebäu-den, da wir uns Hinweise auf deren Funktion erhofften. Weiterhaben wir Schichten und Ensembles einbezogen, die sich durcheinen überdurchschnittlichen Anteil an Knochen auszeichnen.
Betrachtet man den Fundanfall in den verschiedenen Pha-sen (Abb. 30/1) fällt auf, dass mit Abstand am meisten Knochenaus der letzten Phase 7 der grossen Villa (Periode II) des mitt-leren 3. Jahrhunderts stammen, die die meisten Fundschichtengeliefert hat. Es dürften noch andere Gründe für die relativ gerin-ge Knochenmenge der ersten Phasen vorliegen: Viel Abfall derfrüheren Phasen gelangte gar nicht in die Fundschichten, da erbei Planierungen und Auffüllungen verschleppt wurde oder alsDung auf die Felder kam (Kap. 18.2.4) und zudem einem stär-keren Schwund ausgesetzt war. Speiseabfälle und Tierkadaver imGraben entlang der östlichen Hofmauer (s. Kap. 30.2.12) zeigen,dass man Knochenabfälle auch ausserhalb des Gutshofes ent-sorgte. In der letzten Phase kam es nicht mehr zu grossen bau-lichen Veränderungen. Einiger Abfall dürfte erst kurz vor dem
gewaltsamen Ende der Villa abgelagert worden sein, weshalb esnicht mehr zur Entsorgung ausserhalb des ummauerten Berei-ches kam.
Neben diesen taphonomischen Gründen, nämlich denErhaltungs- und Einlagerungsbedingungen, dürften aber auchwirtschaftliche Gründe vorliegen: Vor allem in den ersten Pha-sen der Villa scheint man die Tierhaltung zu einem grossen Teilzur Produktion sekundärer Güter und zur Nutzung der Arbeits-kraft betrieben zu haben (s. Kap. 30.4.). Bei diesen Wirtschafts-zweigen fallen weniger Tierknochen an als bei einer reinenFleischwirtschaft. Die Anzahl der auf dem Gut lebenden Per-sonen, die Fleisch konsumierten, erreichte erst in der Phase 2,im frühen 2. Jahrhundert, eine gewisse Grösse. Für Schafszuchtund Wollproduktion, wie sie für Biberist wahrscheinlich ge-macht werden können, ist weniger Personal als für einen inten-siven Ackerbau notwendig (s. Kap. 30.3.2; s. auch Kap. 18.3.2;32.3.3.). Für die letzte Phase ist ein Ofen zur Fleischkonservie-rung bezeugt (Kap. 30.2.11), so dass mit einem verstärkten Anfallvon Knochenabfällen zu rechnen ist.
Relativ viele Funde (>80/m3) haben Werkstatt B-Phase A,Gebäude E, Gebäude O-Phasen 3 und 4 und die Schichten west-lich des Steinbaus J geliefert (Abb. 30/1; s. auch Tab. 30/1). Eherwenige Knochen (<20/m3) fanden sich in Gebäude A, PortikusF, Gebäude G, Galerie M und Speicher N, während sie in Gebäu-de C fehlen.
Die beträchtliche Durchmischung, die starke Fragmentie-rung sowie die auf vielen Knochen zu beobachtenden Schnitt-spuren lassen darauf schliessen, dass es sich zum grössten Teilum Speiseabfälle handelt. Es lassen sich demnach vor allem Aus-sagen zur Schlachtrate und zur Fleischkonsumation auf dem Gutmachen. Eine Momentaufnahme eines Schlachtvorganges könn-te der sogenannte „Kuhversturz“ darstellen, ein im grossen Gra-narium D aufgefundener Rinderrumpf (s. Kap.30.2.3), der inZusammenhang mit dem im Gebäude Q eingebauten Konser-vierungsofen stehen könnte (s. Kap. 30.2.11).
Berechnungen zu Herdengrössen und zu den Schwerpunk-ten innerhalb der Viehwirtschaft sind hingegen problematisch.Ist eine Tierart in einem Ensemble sehr häufig vertreten, heisstdies eigentlich nur, dass ihr Fleisch mit Vorliebe hier gegessenwurde. Arbeitstiere oder Tiere, die man hauptsächlich als Lie-feranten sekundärer Produkte wie Wolle oder Milch hielt,können durchaus im Fundmaterial schwächer vertreten sein alsdie reinen Fleischtiere, obwohl sie vielleicht für die Wirtschaft
Kap. 30.1
2012 S. Neftenbach (Deschler-Erb/Schröder Fartash in: Rychener 1999, 412Tab. 10): 25879 Knochen; Dietikon (Fischer in: Ebnöther 1995, 255 Abb. 322):4059 Knochen; Treuchtlingen/Weinbergsdorf (Koch 1993): 2465 Knochen.
30. DIE TIERKNOCHEN
(SABINE DESCHLER-ERB,MIT BEITRÄGEN VON HEIDE HÜSTER PLOGMANN,
BRUNO KAUFMANN UND JORGE SPANGENBERG)
Schucany, Biberist 2006

des Gutes von grosser Bedeutung waren. Für die Bearbeitung derÄcker des Fundus etwa war eine beachtliche Rinderherde nötig(s. Kap. 18.2.4). Weiter ist bei einem ausschliesslich zur Fleisch-produktion gehaltenen Tier wie dem Schwein damit zu rechnen,dass es lebend zum Verkauf weggebracht wurde, seine Knochensomit im Fundmaterial fehlen.
Da auf einem römischen Gutshof mit Selbstversorgung anfleischlichen Produkten und kaum mit einem Zukauf vonSchlachttieren zu rechnen ist, dürfen wir annehmen, dass die inden ersten Phasen des späteren 1. und frühen 2. Jahrhundertszu beobachtende Dominanz von Schaf/Ziegen (Kap. 30.3.1)auch die grosse Bedeutung widerspiegelt, die diesen Tieren zuihrer Lebzeit zukam.
Neben den Speiseabfällen fanden sich Tierknochen in spe-ziellen Fundsituationen: als vergrabene Kadaver bzw. als ritu-
elle Deponierungen (s. Kap. 30.2.12; 30.2.13).Im Vergleich zum Gutshof von Neftenbach,wo zehn Teilskelette verschiedener Nutz- undWildtierarten zum Vorschein kamen, ist inBiberist die Anzahl eher gering2013. Es ist nichtanzunehmen, dass in Biberist weniger Tieredurch Krankheiten verendeten als in Neften-bach. In Biberist scheint man andere Vorstel-lungen bezüglich der Abfallentsorgung gehabtzu haben. Vielleicht war man sich der hygieni-schen Probleme eher bewusst, die sich durchdas Vergraben von Kadavern innerhalb desSiedlungsareales ergeben.
An zwei Stellen, im grossen Granarium Dund bei der Grabanlage H (Kap. 30.2.3; 30.2.6), fanden sich die Überreste von drei Rin-derhornzapfen bzw. eines Bukranions, die imZusammenhang mit symbolisch-religiösenVorstellungen zu sehen sind2014.
30.1.2. Erhaltungszustand
Der Zustand der Tierknochen kann viel-fältige Hinweise auf Ereignisse geben, die zwi-schen der Schlachtung und der Einlagerungstattgefunden haben. Er kann aber auchInformationen zur Schichtgenese und zuVorgängen innerhalb des Sedimentes liefern.Die Erhaltung der Tierknochen kann somitwichtige Informationen zur Befundanalysegeben.
Durchschnittsgewicht Das Durchschnittsgewicht der Tierknochen
schwankt in den einzelnen Ensembles beacht-lich (Abb. 30/2; s. auch Tab. 30/1). Relativ hoheWerte, um 15 g und höher, liegen vor allem fürEnsembles vor, die eher wenig Funde gelieferthaben, etwa in Speicher N aus einer Grube(A116/B075) beim Tor, einer Grube (A034/B155) bei der Trennmauer sowie einer Grube(A095.60/B106.50) in Steinbau J-Phase 4.
Die Knochen der übrigen, meist fundreicheren Schichten wei-sen ein Durchschnittsgewicht von unter 10 g auf. Dabei fällt auf,dass in den Wohngebäuden (E/J/O) in der Regel tiefere Werte(<5 g) erreicht werden als in den Ökonomiegebäuden (Werk-statt B/Vorratsspeicher D) und den Plätzen unter freiem Him-mel (westlich Steinbau J/westlich Gebäude O/Portikus F/bei derHofmauer). Da es sich hauptsächlich um Speiseabfälle handelt,ist anzunehmen, dass sie primär am Ort der Konsumation, alsoin den Wohngebäuden, anfielen. Bei den Knochen aus den Öko-nomiegebäuden und den freien Plätzen dürfte es sich umumgelagerte Funde handeln. Relativ hohe Durchschnittswerte
V. Funde und besondere Fundensembles römischer Zeit – 30. Die Tierknochen
2013 Deschler-Erb/Schröder Fartash in: Rychener 1999, 494.2014 Deschler-Erb 1999.
636
Abb. 30/1. Anteile der Tierknochen je Gebäude (s. Tab. 30/1).
Abb. 30/2. Durchschnittsgewicht (g) der Tierknochen je Gebäude (s. Tab. 30/1).

637
zeigen sich in den Bauhorizonten von Gebäude Q und der erstenPhase des Steinbaus J. Es dürfte sich hierbei um Material han-deln, dass im Zusammenhang mit dem Bau der Gebäude voneiner anderer Stelle hierher gelangte.
Dass Funde aus Planien ein höheres Durchschnittsgewichtals solche aus Benutzungsschichten aufweisen, hat bereits amBeispiel der Tierknochen aus Insula 23 in Augusta Raurica unddem Gebäude 25 der Villa von Neftenbach gezeigt werden kön-nen, was auf vergleichbare Ablagerungsvorgänge in Stadtund Villa hindeutet2015. Dort wird die grössere Beanspru-chung im Sediment oder ein systematisches Wegwischen vongrösserem Abfall für die geringeren Durchschnittswerte in denBenutzungsschichten diskutiert. Dabei hat sich eher die Bean-spruchung im Sediment, vor allem durch die Begehung derBewohner, als hauptsächlicher Faktor für ein geringes Durch-
schnittsgewicht herausgestellt. Dies dürfteauch in Biberist zutreffen: Die besagten Pla-nien weisen nicht einen übermässigen Anteilan grossen und somit auch schweren Rin-derknochen auf. In den Planien westlich desSteinbaus J und westlich des Gebäudes O so-wie den Strukturen bei der Hofmauer sinddie kleineren Schweineknochen gut oder so-gar dominant vertreten (Tab. 30/2). Darausist zu schliessen, dass die vorliegenden Pla-nieschichten nicht dadurch entstanden, dassbewusst grösserer Abfall aus den Wohnge-bäuden entfernt wurde, sondern eher zufälligim Zusammenhang mit baulichen Massnah-men.
Oberflächenerhaltung Bei guter Erhaltung ist eine Knochen-
oberfläche glatt und hart. Durch verschiedeneBodeneinflüsse (ph-Wert, Durchlässigkeit desBodens, wechselndes Milieu) wird der Kno-chen brüchig, die Oberfläche oder ganze Teileder Kompakta splittern ab. Dass die Oberflä-chenerhaltung nichts mit anthropogenen Ein-flüssen zu tun hat, lässt sich daran erkennen,dass keine klaren Unterschiede zwischen Gru-ben, Wohnhäusern und Ökonomiegebäudenbestehen.
Die mit Abstand am schlechtesten erhalte-nen Knochen stammen aus dem am nördlich-sten gelegenen Gebäude E (Abb. 30/3), das sichfast zuoberst auf dem Moränerücken über derAareniederung befand. Im kiesigen Materialherrschte wohl ein stark wechselndes Milieu.
Obwohl sich die Biberister Sedimenteschlecht auf die Erhaltung der Tierknochenauswirkten, begünstigten sie hingegen dieErhaltung der Hornscheide des Bukranions beiGrab H2016.
BestimmbarkeitDie Zuweisung eines Tierknochens an eine bestimmte Tier-
art hängt eng mit dem Durchschnittsgewicht, aber auch mit derErhaltung zusammen. In Biberist ist der Anteil der bestimm-baren Knochen starken Schwankungen unterworfen (Abb.30/4; s. auch Tab. 30/1). In Gebäude E, wo das Durchschnitts-gewicht relativ tief ist und die Erhaltung sehr schlecht, ist auchder Bestimmbarkeitsgrad nicht sehr hoch (>20%). In denübrigen Ensembles werden zwar häufiger Werte von über 50%erreicht; allerdings ist festzustellen, dass im Vergleich zur eben-
Kap. 30.1
2015 Deschler-Erb 1991, 369, Abb. 209; Deschler-Erb/Schröder Fartash in: Ryche-ner 1999, 262-264.2016 Kap. 30.2.6; wie Anm. 2015, 101 Abb. 3.
Abb. 30/3. Anteil (n%) schlecht erhaltener Tierknochen je Gebäude (s. Tab. 30/1).
Abb. 30/4. Anteil (n%) der bestimmbaren Tierknochen je Gebäude (s. Tab. 30/1).

falls sehr sorgfältig durchgeführten Grabungbeim Augster Theater, die Werte tiefer liegen2017.Der Zustand der Biberister Knochen ist durch-schnittlich schlechter, als jener in Augst2018.Einige Untersuchungen zur Metrik und zumIndividualalter sind deshalb nicht möglich.
BruchkantenEin frisch gebrochener Knochen weist
eckige, spitz zulaufende Bruchkanten auf.Durch Abrieb im Sediment, Umlagerung derSchichten, längere Lagerung an der Oberflächeoder Wassereinfluss runden sich die Bruch-kanten ab. Ein hoher Verrundungsgrad deutetsomit auf eine bewegte Ablagerungsgeschich-te hin.
Hohe Anteile von verrundeten Bruchkan-ten (>15%) fanden sich in der allerdings fund-armen Grube (A118/B075) beim Tor, sowie inGrube (A110/B050) westlich des Gebäudes Ound in der vor dem Bau der Werkstatt B-Phase C eingebrachten Planie (Tab. 30/1; s.auch Abb. 30/5). Das entsprechende Materialmuss vor seiner definitiven Einlagerung län-gere Zeit herumgelegen sein. Recht hohe An-teile von verrundeten Bruchkanten weisenauch die übrigen Ensembles der Werkstatt Bauf. In den Wohngebäuden E, J und O sowiein Gebäude Q ist der Anteil der verrundetenBruchkanten eher klein. Leicht höhere Wertefinden sich hingegen in der Werkgrube (A049/B228) südöstlich des Gebäudes E. Dies lässtdarauf schliessen, dass die Verrundung weni-ger bei der Beanspruchung in den Benutzungs-schichten der Wohngebäude als vielmehr beider Umlagerung sowie der Lagerung ausser-halb der Gebäude entstanden ist.
Verbissspuren Der Anteil der Knochen mit Verbissspuren
hängt davon ab, wie lange diese frei zugänglichfür Hunde und Schweine herumlagen. Oft
fallen Verbissspuren mit einem hohen Verrundungsgrad zusam-men, was für eine längere, bewegte Ablagerungsgeschichtespricht (Abb. 30/5; s. auch Tab. 30/1). Dies gilt etwa für das vordem Bau der Werkstatt B-Phase C eingebrachte Material. Bei denKnochen der Phase 1 des Steinbaus J korreliert ein hoher Anteilan Verbissspuren mit einem relativ hohen Durchschnittsgewicht,was für eine Planieschicht spricht. Die höchsten Anteile von fast35% finden sich in Grube (A110/B050) westlich des GebäudesO und in der Traufrinne nördlich der Portikus F, die allerdingsnur wenige Knochen geliefert haben.
V. Funde und besondere Fundensembles römischer Zeit – 30. Die Tierknochen
2017 Zu Augst: Furger/Deschler-Erb 1992, 359 Abb. 87.2018 Das mag auch auf die starke Entkalkung des Bodens zurückzuführen sein(s. Kap. 4.2 mit Anm. 55)
638
Abb. 30/6. Anteil (n%) der Tierknochen mit Schnittspuren je Gebäude (s. Tab. 30/1).
Abb. 30/7. Entwicklung des Schnittspurenanteils (n%) bei den Tier-knochen in der grossen Villa (Periode II).
Abb. 30/5. Anteile (n%) der Knochen mit verrundeten Bruchkanten und Verbissspuren (s. Tab. 30/1).

639
Hohe Anteile an Verbissspuren liegen für die Dachschutt-schichten in den Gebäude Q und K vor, nicht aber für die Dach-schuttschichten der Gebäude D, E, O und Steinbau J. Es ist denk-bar, dass bei ersteren die Knochen nur locker bedeckt und daherleichter zugänglich für herumstreunende Tiere waren, was beiGebäude K, einem Fachwerkhaus, auch von der Konstruktionher nahe liegt, während letztere, insbesondere Gebäude D, E undSteinbau J nachweislich durch mächtige Mauerschuttschichtenbesser geschützt waren.
In den meisten Ensembles des Landarbeiterhauses O sind dieVerbissspuren etwa doppelt so häufig wie im Vilicushaus J. Mög-licherweise ist daraus zu schliessen, dass die Hofhunde bei denLandarbeitern lebten oder zumindest von diesen versorgt wur-den.
In den Schichten des Wohnhauses E und des Getreidespei-chers D sind Verbissspuren selten. Bei Gebäude E dürfte dies ander dort zu beobachtenden schlechten Oberflächenerhaltung derKnochen liegen (s. oben), was verunmöglicht, Verbissspuren zuerkennen.
Schnitt- und HackspurenSchnitt- und häufig auch Hackspuren gelangen einerseits
ohne Absicht während des Schlachtvorganges an den Knochen,wenn mit dem Werkzeug zu weit ins Fleisch eingedrungen odereine Sehne zu heftig durchgetrennt wird. Andererseits sind diegrösseren Rinderknochen in der Regel auch stärker zerlegt undweisen mehr Schlachtspuren auf als die Schaf-/Ziegen- undSchweineknochen2019. Die Häufigkeit von Schnitt- und Hack-spuren kann charakteristisch für eine bestimmte Schlacht-technik und sogar für bestimmte Bevölkerungsgruppen sein, wiedie folgenden Ausführungen zeigen.
Tierknochen mit Schnittspuren kommen im ganzen Ville-nareal vor (Abb. 30/6), was nicht bedeutet, dass überall ge-schlachtet und/oder gegessen wurde, sondern eher für eine inter-ne Verlagerung des Abfalls spricht. In Gebäude E, dem mut-masslichen Sitz des Verwalters des grossen Getreidespeichers D,
fanden sich kaum Knochen mit Schlachtspu-ren, was indes nur zu einem gewissen Teil mitder erwähnt schlechten Oberflächenerhaltungin Zusammenhang zu bringen ist; deutlicheSchnittspuren wären auch bei schlechter Er-haltung noch zu erkennen gewesen. In denPhasen des Steinbaus J, des Vilicushauses, lie-gen die Anteile unter einem Prozent (Tab.30/1). Hingegen findet sich in einigen Ensem-bles des Gebäudes O ein Anteil an Schlacht-spuren von um die 4% und höher. Mögli-cherweise liegt hier ein sozialer Unterschiedvor: Vielleicht schlachteten die Landarbeiterselbst, während die Verwalter von einem pro-fessionellen Metzger schlachten liessen. Rela-tiv hohe Anteile (4-12%) werden auch inallen Ensembles der Werkstatt B erreicht. Beidiesen Knochen könnte es sich um einge-brachtes Material handeln, das z.T. aus demLandarbeiterhaus A stammt. Hier sind viel-leicht die Betreiber der Werkstatt zu suchen(s. Kap. 10.3.1; 30.2.2).
Betrachtet man die Entwicklung der Schlachtspurenanteile anden Rinder-, Schaf-/Ziegen- und Schweineknochen, zeigensich deutliche Schwankungen (Abb. 30/7): In den Phasen 1-2 dergrossen Villa (Periode II) liegen sie bei allen drei Nutztierartenam höchsten. In den folgenden Phasen 3-4 sowie 5-6 gehen siedeutlich zurück. In der letzten Phase 7 erhöhen sie sich wiede-rum. Die Veränderungen von Phase 2 zu Phase 3 bzw. vonPhase 6 zu Phase 7 finden in den tief greifenden Umbauten imganzen Villenareal eine Entsprechung, die zumindest für die letz-te Phase mit einem Besitzerwechsel in Verbindung gebracht wer-den kann (s. Kap. 17.4.2-3; 17.4.6-7). Dies bedeutet, dass nebenzeitgleichen sozialen Unterschieden innerhalb des Gutshofesauch zeitliche Veränderungen festgestellt werden können.
BrandspurenBrandspuren entstehen beim Braten des den Knochen um-
gebenden Fleisches oder wenn Knochenabfälle in ein Herdfeu-er oder einen Hausbrand geraten.
Knochen mit Brandspuren haben die meisten grösseren En-sembles des Gutshofinneren geliefert (Abb. 30/8; s. auch Tab.30/1). Hohe Anteile finden sich in den Brandschichten des Holz-baus J, der Gebäude K und O-Phase 2, die alle einem Brand zumOpfer gefallen sind. Die höchsten Anteile überhaupt liegen fürdie Brandschichten des Holzbaus J vor (Abb. 30/8), also für dieverbrannte Benützungsschicht, den darüber gelegenen Brand-schutt und den in Mauerausbruchgruben planierten Brand-schutt, was für einen intensiven Brand spricht.
Wie die Brandspuren der Knochen aus den anderen Ensem-bles zu erklären sind, muss offen bleiben. Nur wenige verbrannteKnochen fanden sich ausserhalb der Hofmauer.
Kap. 30.1
Abb. 30/8. Anteil (n%) der der verbrannten Tierknochen je Gebäude (s. Tab. 30/1).
2019 Wie Anm. 2017, 414.

30.2. Die Tierknochen dereinzelnen Gebäude
Mehrere Gebäude haben keine oder so we-nige Tierknochen geliefert, dass wir auf einestatistische Auswertung vollständig verzichte-ten (Gebäude C/Gebäude G/Gebäude L/Gale-rie M/Speicher N). Bei anderen Gebäuden liessdie Anzahl zwar eine Tierartenanalyse zu.Weiterführende Untersuchungen, etwa zumSkelettteilspektrum oder zur Altersstrukturwerden zum Teil versucht, sind aber aufgrundder geringen Datenbasis als sehr unsicher zubezeichnen.
30.2.1. Gebäude A
Aufgrund seiner Anlage und Ausstattungist das wohl im ganzen 2. Jahrhundert beste-hende Gebäude A als Wohngebäude einerLandarbeiterfamilie zu interpretieren (Kap. 10.3.3). Die nur gerade 52 Tierknochen lassenkeine weitreichenden Interpretationen zu(Tab.30/2-30/7); sie zeigen lediglich die dreiwichtigsten Haustierarten, was mit der gerin-gen statistischen Basis zu erklären ist. Die Ab-fälle aus Gebäude A dürften anderswo entsorgtworden sein. In Frage kommt der später pla-nierte Abfallhaufen bei Werkstatt B-Phase B.
30.2.2. Werkstatt B
Phase A Aufgrund der Installationen – eine Werk-
grube, ein Becken und mehrere Kanäle – wirddie schon für das erste Gehöft (Periode I) um75 n.Chr. errichteten Werkstatt B als Wäsche-rei und/oder Walkerei gedeutet, die bis insfrühe 2. Jahrhundert bestand (Kap. 9.4).Grundsätzlich lassen sich diese Funktions-
deutungen aufgrund der mitgefundenen Tierknochen nichtbeurteilen, da bei den entsprechenden Tätigkeiten kein Tier-knochenabfall direkt anfällt.
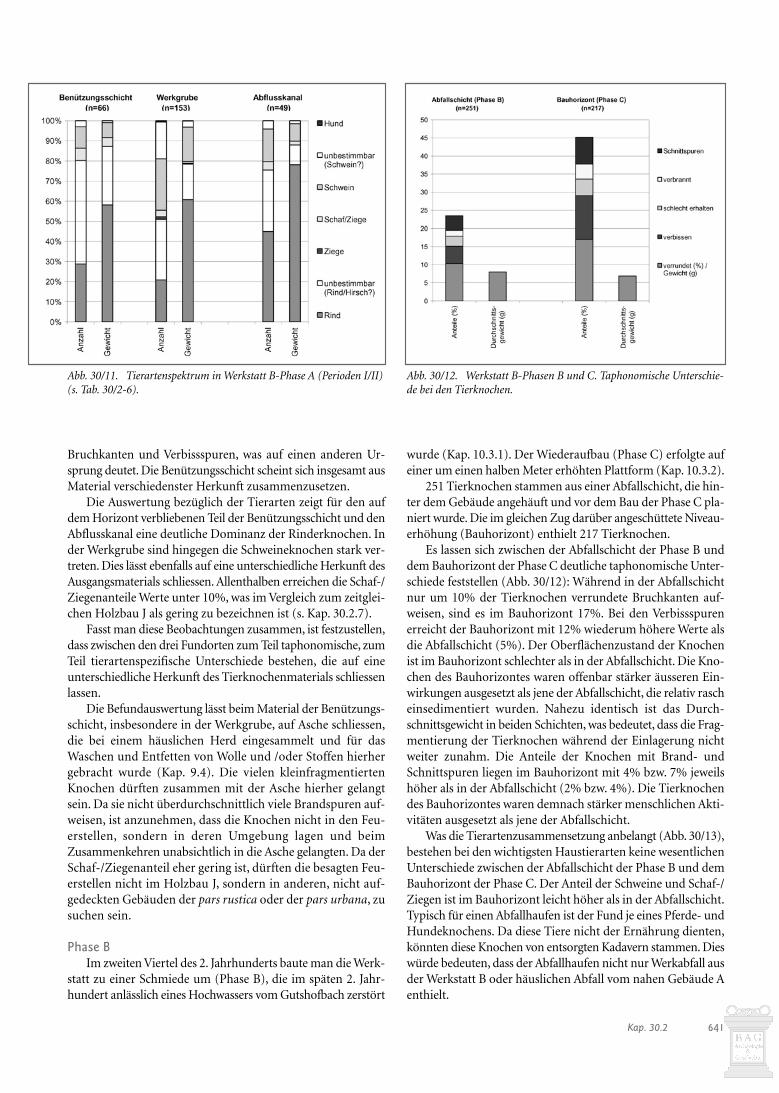
Ingesamt stammen 274 Tierknochen aus der Benützungs-schicht, die beim Bau der folgenden Phase z.T. in die Vertie-fungen einplaniert wurde (Tab. 30/1). Die einzelnen Strukturenhaben unterschiedlich viele Tierknochen geliefert (Abb. 30/9):In der Benützungsschicht und dem Abflusskanal kamen deut-lich weniger Tierknochen zum Vorschein als in der Werkgrube.In Bezug auf das Schichtvolumen ist der Anteil der Knochen inden einzelnen Schichten miteinander vergleichbar.
Die schwarze Benützungsschicht hat den grössten Anteil vonTierknochen mit Brandspuren geliefert (Abb. 30/10), wobeiweniger verbrannte Knochen aus der Werkgrube stammen, garkeine aus dem Abflusskanal. Der Zustand der Knochen aus demAbflusskanal scheint auch sonst anders zu sein als bei jenen desnicht planierten Teils der Benützungsschicht und der Werk-grube. So finden sich hier die höchsten Werte an verrundeten
V. Funde und besondere Fundensembles römischer Zeit – 30. Die Tierknochen640
Abb. 30/9. Werkstatt B-Phase A. Fundanteil (pro m3) und Durchschnittsgewicht (g/n) derverschiedenenTierarten.
Abb. 30/10. Werkstatt B-Phase A. Anteil verbrannter, verrundeter undverbissener Tierknochen.
641
Bruchkanten und Verbissspuren, was auf einen anderen Ur-sprung deutet. Die Benützungsschicht scheint sich insgesamt ausMaterial verschiedenster Herkunft zusammenzusetzen.
Die Auswertung bezüglich der Tierarten zeigt für den aufdem Horizont verbliebenen Teil der Benützungsschicht und denAbflusskanal eine deutliche Dominanz der Rinderknochen. Inder Werkgrube sind hingegen die Schweineknochen stark ver-treten. Dies lässt ebenfalls auf eine unterschiedliche Herkunft desAusgangsmaterials schliessen. Allenthalben erreichen die Schaf-/Ziegenanteile Werte unter 10%, was im Vergleich zum zeitglei-chen Holzbau J als gering zu bezeichnen ist (s. Kap. 30.2.7).
Fasst man diese Beobachtungen zusammen, ist festzustellen,dass zwischen den drei Fundorten zum Teil taphonomische, zumTeil tierartenspezifische Unterschiede bestehen, die auf eineunterschiedliche Herkunft des Tierknochenmaterials schliessenlassen.
Die Befundauswertung lässt beim Material der Benützungs-schicht, insbesondere in der Werkgrube, auf Asche schliessen,die bei einem häuslichen Herd eingesammelt und für dasWaschen und Entfetten von Wolle und /oder Stoffen hierhergebracht wurde (Kap. 9.4). Die vielen kleinfragmentiertenKnochen dürften zusammen mit der Asche hierher gelangtsein. Da sie nicht überdurchschnittlich viele Brandspuren auf-weisen, ist anzunehmen, dass die Knochen nicht in den Feu-erstellen, sondern in deren Umgebung lagen und beimZusammenkehren unabsichtlich in die Asche gelangten. Da derSchaf-/Ziegenanteil eher gering ist, dürften die besagten Feu-erstellen nicht im Holzbau J, sondern in anderen, nicht auf-gedeckten Gebäuden der pars rustica oder der pars urbana, zusuchen sein.
Phase BIm zweiten Viertel des 2. Jahrhunderts baute man die Werk-
statt zu einer Schmiede um (Phase B), die im späten 2. Jahr-hundert anlässlich eines Hochwassers vom Gutshofbach zerstört
wurde (Kap. 10.3.1). Der Wiederaufbau (Phase C) erfolgte aufeiner um einen halben Meter erhöhten Plattform (Kap. 10.3.2).
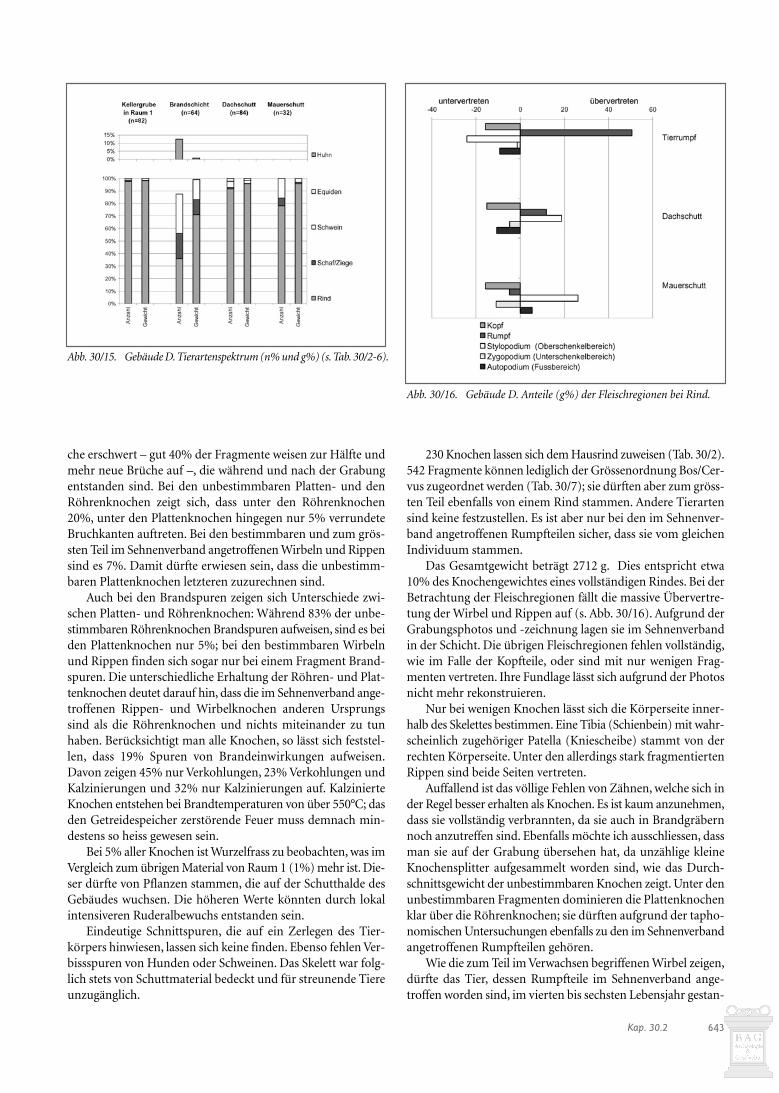
251 Tierknochen stammen aus einer Abfallschicht, die hin-ter dem Gebäude angehäuft und vor dem Bau der Phase C pla-niert wurde. Die im gleichen Zug darüber angeschüttete Niveau-erhöhung (Bauhorizont) enthielt 217 Tierknochen.
Es lassen sich zwischen der Abfallschicht der Phase B unddem Bauhorizont der Phase C deutliche taphonomische Unter-schiede feststellen (Abb. 30/12): Während in der Abfallschichtnur um 10% der Tierknochen verrundete Bruchkanten auf-weisen, sind es im Bauhorizont 17%. Bei den Verbissspurenerreicht der Bauhorizont mit 12% wiederum höhere Werte alsdie Abfallschicht (5%). Der Oberflächenzustand der Knochenist im Bauhorizont schlechter als in der Abfallschicht. Die Kno-chen des Bauhorizontes waren offenbar stärker äusseren Ein-wirkungen ausgesetzt als jene der Abfallschicht, die relativ rascheinsedimentiert wurden. Nahezu identisch ist das Durch-schnittsgewicht in beiden Schichten, was bedeutet, dass die Frag-mentierung der Tierknochen während der Einlagerung nichtweiter zunahm. Die Anteile der Knochen mit Brand- undSchnittspuren liegen im Bauhorizont mit 4% bzw. 7% jeweilshöher als in der Abfallschicht (2% bzw. 4%). Die Tierknochendes Bauhorizontes waren demnach stärker menschlichen Akti-vitäten ausgesetzt als jene der Abfallschicht.
Was die Tierartenzusammensetzung anbelangt (Abb. 30/13),bestehen bei den wichtigsten Haustierarten keine wesentlichenUnterschiede zwischen der Abfallschicht der Phase B und demBauhorizont der Phase C. Der Anteil der Schweine und Schaf-/Ziegen ist im Bauhorizont leicht höher als in der Abfallschicht.Typisch für einen Abfallhaufen ist der Fund je eines Pferde- undHundeknochens. Da diese Tiere nicht der Ernährung dienten,könnten diese Knochen von entsorgten Kadavern stammen. Dieswürde bedeuten, dass der Abfallhaufen nicht nur Werkabfall ausder Werkstatt B oder häuslichen Abfall vom nahen Gebäude Aenthielt.
Kap. 30.2
Abb. 30/11. Tierartenspektrum in Werkstatt B-Phase A (Perioden I/II) (s. Tab. 30/2-6).
Abb. 30/12. Werkstatt B-Phasen B und C. Taphonomische Unterschie-de bei den Tierknochen.

Die Auswertung der Rinderknochen nach Fleischregionenzeigt im Bauhorizont der Phase C ein dem rezenten Vergleichs-skelett sehr ähnliche Verteilung (Abb. 30/14)2020. Daraus lässt sichschliessen, dass bei diesem Material – im Gegensatz zur Abfall-schicht der Phase B – in nächster Umgebung ein oder mehrereRinder geschlachtet wurden und ihre Knochen mehr oder we-niger vollständig in den Boden gelangten. Bei den Schweine-knochen ist in beiden Schichten eine immer wieder zu beob-achtende deutliche Untervertretung der Rumpfteile festzustellen.
Aus archäozoologischer Sicht ist zu vermuten, dass die meis-ten Tierknochen der Abfallschicht der Phase B nicht mit derSchmiede in der Werkstatt B zusammenhängen, sondern vonanderswoher stammen. Sie scheinen besser erhalten zu sein alsdas Material des Bauhorizontes der Phase C (s. Abb. 30/12). DieZusammensetzung deutet auf Material hin, das an verschiede-nen Orten anfiel und hier aufgehäuft wurde. In der vor dem Bauder Phase C über der Abfallschicht eingebrachten Niveauerhö-hung (Bauhorizont C) dürften Schlachtabfälle von Rindern ent-halten sein, die in nächster Nähe angefallen waren.
30.2.3. Gebäude D
Das im zweiten Viertel des 2. Jahrhunderts errichtete, z.T.umgebaute grosse Gebäude ist aufgrund des Grundrisses undder botanischen Reste als Getreidespeicher zu interpretieren(Kap. 10.4.1), der kurz nach der Mitte des 3. Jahrhundert einemverheerenden Feuer zum Opfer fiel. Insgesamt kamen 1468 Tier-knochen zum Vorschein, wovon aber ein grosser Teil auf zweiSpezialbefunde, einen Tierrumpf (772) und eine Kellergrube(108), entfällt (Tab. 30/1).
Tierknochen ohne speziellen Befund588 Tierknochen stammen zum grossen Teil aus der Brand-
schicht in Raum 1 und dem Dach- sowie Mauerschutt. AufVergleiche zwischen den einzelnen Räumen wird daher im Fol-
genden verzichtet. Eine vergleichende Untersuchung der Pha-sen 1 und 2 macht ebenfalls wenig Sinn, da lediglich die beimUmbau erstellten Kiesböden mit Unterlage von Raum 1 (22Tierknochen) und Raum 4 (21 Tierknochen) einer früherenPhase zuzurechnen sind.
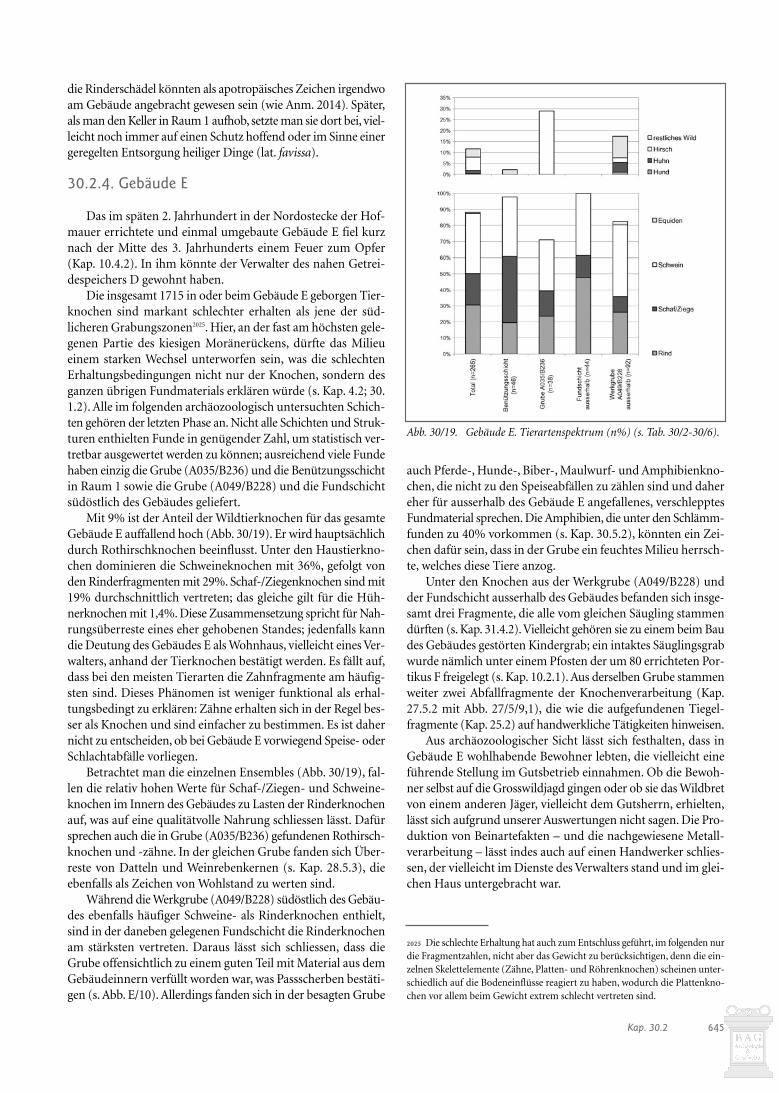
Insgesamt herrschen die Rinderknochen vor (Abb. 30/15).Beim Skelettteilspektrum der Rinderknochen aus dem Dach-und dem Mauerschutt zeigt sich eine auffällige Dominanz desStylopodiums (Abb. 30/16); im Dachschutt sind auch die Rumpf-teile deutlich übervertreten. Bei den Rinderknochen handelt essich somit nicht um primäre Schlachtabfälle, sondern umÜberreste bereits zerlegter Körperteile, die an den in der Süd-westecke des Raumes gefundenen Rumpf erinnern (s. unten).
Tierrumpf (A983/B175) in Raum 1In der Südwestecke von Raum 1 fanden sich, teils in der
Brandschicht, teils im Dachschutt, konzentriert auf rund einenQuadratmeter insgesamt 772 Tierknochenfragmente2021. Bereitswährend der Ausgrabung war ersichtlich, dass einige Knochenin anatomisch richtiger Lage zueinander im Boden lagen (Abb.30/17). Handelt es sich um ein Tier, das unter den zusammen-stürzenden Gebäudemassen ums Leben kam, oder haben wireine Entsorgung von Schlachtabfall oder eines Kadavers vor uns?Die folgende Tierknochenanalyse hat zum Ziel, Pro und Con-tra dieser Deutungen zu überprüfen.
Zu bemerken ist, dass die Knochen nicht in einer Grubegefunden wurden, was für die Entsorgung von Schlachtabfällenoder eines Kadavers spräche, sondern an der Bodenoberfläche,zwischen Brandschicht und Dachschutt. Eine Beurteilung desBruchkantenzustandes wird durch den hohen Anteil neuer Brü-
V. Funde und besondere Fundensembles römischer Zeit – 30. Die Tierknochen
2020 Zu den Fleischregionen und dem Vergleich mit den rezenten Vergleichs-skeletten siehe Rychener 1999, 30, Tab. 244.2021 Brandschicht: Inv.-Nr. 14/91053; 14/9/1054; 14/9/1055; Dachschutt Inv.-Nr. 14/9/978; 14/9/980.
642
Abb. 30/13. Werkstatt B-Phasen B und C. Tierartenspektrum (n%) (s. Tab. 30/2-6).
Abb. 30/14. Werkstatt B-Phasen B und C. Anteile (g%) der Fleisch-regionen bei Rind und Schwein.
643
che erschwert – gut 40% der Fragmente weisen zur Hälfte undmehr neue Brüche auf –, die während und nach der Grabungentstanden sind. Bei den unbestimmbaren Platten- und denRöhrenknochen zeigt sich, dass unter den Röhrenknochen20%, unter den Plattenknochen hingegen nur 5% verrundeteBruchkanten auftreten. Bei den bestimmbaren und zum grös-sten Teil im Sehnenverband angetroffenen Wirbeln und Rippensind es 7%. Damit dürfte erwiesen sein, dass die unbestimm-baren Plattenknochen letzteren zuzurechnen sind.
Auch bei den Brandspuren zeigen sich Unterschiede zwi-schen Platten- und Röhrenknochen: Während 83% der unbe-stimmbaren Röhrenknochen Brandspuren aufweisen, sind es beiden Plattenknochen nur 5%; bei den bestimmbaren Wirbelnund Rippen finden sich sogar nur bei einem Fragment Brand-spuren. Die unterschiedliche Erhaltung der Röhren- und Plat-tenknochen deutet darauf hin, dass die im Sehnenverband ange-troffenen Rippen- und Wirbelknochen anderen Ursprungssind als die Röhrenknochen und nichts miteinander zu tunhaben. Berücksichtigt man alle Knochen, so lässt sich feststel-len, dass 19% Spuren von Brandeinwirkungen aufweisen.Davon zeigen 45% nur Verkohlungen, 23% Verkohlungen undKalzinierungen und 32% nur Kalzinierungen auf. KalzinierteKnochen entstehen bei Brandtemperaturen von über 550°C; dasden Getreidespeicher zerstörende Feuer muss demnach min-destens so heiss gewesen sein.
Bei 5% aller Knochen ist Wurzelfrass zu beobachten, was imVergleich zum übrigen Material von Raum 1 (1%) mehr ist. Die-ser dürfte von Pflanzen stammen, die auf der Schutthalde desGebäudes wuchsen. Die höheren Werte könnten durch lokalintensiveren Ruderalbewuchs entstanden sein.
Eindeutige Schnittspuren, die auf ein Zerlegen des Tier-körpers hinwiesen, lassen sich keine finden. Ebenso fehlen Ver-bissspuren von Hunden oder Schweinen. Das Skelett war folg-lich stets von Schuttmaterial bedeckt und für streunende Tiereunzugänglich.
230 Knochen lassen sich dem Hausrind zuweisen (Tab. 30/2).542 Fragmente können lediglich der Grössenordnung Bos/Cer-vus zugeordnet werden (Tab. 30/7); sie dürften aber zum gröss-ten Teil ebenfalls von einem Rind stammen. Andere Tierartensind keine festzustellen. Es ist aber nur bei den im Sehnenver-band angetroffenen Rumpfteilen sicher, dass sie vom gleichenIndividuum stammen.
Das Gesamtgewicht beträgt 2712 g. Dies entspricht etwa10% des Knochengewichtes eines vollständigen Rindes. Bei derBetrachtung der Fleischregionen fällt die massive Übervertre-tung der Wirbel und Rippen auf (s. Abb. 30/16). Aufgrund derGrabungsphotos und -zeichnung lagen sie im Sehnenverbandin der Schicht. Die übrigen Fleischregionen fehlen vollständig,wie im Falle der Kopfteile, oder sind mit nur wenigen Frag-menten vertreten. Ihre Fundlage lässt sich aufgrund der Photosnicht mehr rekonstruieren.
Nur bei wenigen Knochen lässt sich die Körperseite inner-halb des Skelettes bestimmen. Eine Tibia (Schienbein) mit wahr-scheinlich zugehöriger Patella (Kniescheibe) stammt von derrechten Körperseite. Unter den allerdings stark fragmentiertenRippen sind beide Seiten vertreten.
Auffallend ist das völlige Fehlen von Zähnen, welche sich inder Regel besser erhalten als Knochen. Es ist kaum anzunehmen,dass sie vollständig verbrannten, da sie auch in Brandgräbernnoch anzutreffen sind. Ebenfalls möchte ich ausschliessen, dassman sie auf der Grabung übersehen hat, da unzählige kleineKnochensplitter aufgesammelt worden sind, wie das Durch-schnittsgewicht der unbestimmbaren Knochen zeigt. Unter denunbestimmbaren Fragmenten dominieren die Plattenknochenklar über die Röhrenknochen; sie dürften aufgrund der tapho-nomischen Untersuchungen ebenfalls zu den im Sehnenverbandangetroffenen Rumpfteilen gehören.
Wie die zum Teil im Verwachsen begriffenen Wirbel zeigen,dürfte das Tier, dessen Rumpfteile im Sehnenverband ange-troffen worden sind, im vierten bis sechsten Lebensjahr gestan-
Kap. 30.2
Abb. 30/15. Gebäude D. Tierartenspektrum (n% und g%) (s. Tab. 30/2-6).
Abb. 30/16. Gebäude D. Anteile (g%) der Fleischregionen bei Rind.

den haben. Da die Wirbelepiphysen noch nicht ganz mit demWirbelkörper verwachsen sind, haben wir auf ein Vermessen derWirbel verzichtet. Nach der optischen Einschätzung dürfte essich um ein relativ grosses Individuum gehandelt haben, wie siein römischer Zeit immer wieder vorkommen.
Zusammenfassend ist festzustellen, dass alle unter den Schutt-massen des Gebäudes D begrabenen Tierknochen von Rindernstammen. Die Rumpfknochen fanden sich zumindest zum Teil imanatomischen Verband und müssen vom gleichen Individuumstammen. Allerdings ist mit der angetroffenen Skeletteilverteilungund -häufigkeit kein vollständiges Individuum zu rekonstruieren.Würde es sich um ein verunglücktes Tier handeln, müsste ein Teildes Skelettes vollständig verbrannt sein, so dass nur noch Ascheübriggeblieben wäre. Für diese Theorie spricht das gehäufte Vor-kommen von Brandspuren an den Röhrenknochenfragmenten.Diese stammen von den Extremitäten, die im Material extremuntervertreten sind. In diesem Falle wäre das vom Feuer über-raschte Tier auf die Seite gefallen. Die Extremitätenknochen, diezum grossen Teil nur mit Haut überzogen sind, wären rasch derHitze ausgesetzt gewesen und verbrannt. Das gleiche Schicksalwäre dem Kopf und den bodenabgewandten Rumpfteilen wider-fahren. Die Rippen und Wirbel, die direkt auf dem Boden lagen,wären durch die sie bedeckenden Körpermassen geschützt gewe-sen und so besser erhalten geblieben. Ähnliche Beobachtungenhaben sich bei einem experimentellen Versuch einer Bustum-Bestattung feststellen lassen, bei der man zwei Schweine ver-brannte2022: Auch hier blieben die dem Boden aufliegendenRumpfteile vollständiger erhalten als die anderen.
Bei dem eben vorgestellen Erklärungsmodell bietet allerdingsdie Tatsache, dass die Stylopodiumknochen (Scapula, Humerus,Pelvis, Femur), die beim Rind mit viel Muskelfleisch umgebensind, so gut wie fehlen, Schwierigkeiten. Gerade die dem Bodenzugewandte Scapula (Schulterblatt) wäre von allen Knochen deram besten geschützte gewesen. Dieses Indiz und die taphono-mischen Resultate sprechen somit gegen die Theorie eines ver-unfallten Tieres. Aufgrund aller Fakten scheint es eher, dass hiernoch am Knochen anhaftende Rindfleischteile bis zum endgül-tigen Konsum zwischengelagert waren, zu dem es indes nichtmehr kam. Entweder waren diese Teile unbehandelt und zum fri-schen Verbrauch bestimmt, der sich in einem kalten Winterdurchaus über mehrere Wochen hinziehen konnte. Heutzutagelässt man Kuhfleisch bis zu vier Wochen abhängen, wodurch eszarter, mürber und aromatischer wird2023. Oder das Fleisch wurde
hier nur zwischengelagert, um dann anderswo, vielleicht im Ofendes Gebäudes Q, weiter verarbeitet zu werden (Kap. 30.2.11).
Da bei den übrigen Rinderknochen aus der Brandschichtvon Raum 1 sowie dem Dach- und Mauerschutt eine Dominanzdes Stylopodiums und der Rumpfteile festzustellen ist (s. Abb.30/16), könnte der hier zufällig erhaltene Rumpf keinen Ein-zelfall darstellen, sondern bedeuten, dass noch weiter Rinder-teile im Gebäude D gelagert wurden.
Kellergrube (A979/B181) in Raum 1Aus der beim Umbau an der Wende zum 3. Jahrhundert ver-
füllten Kellergrube der Phase a stammen insgesamt 108 Tier-knochen, von denen 80 sich als Rind bestimmen lassen(s. Abb. 30/15; Tab. 30/2-30/7)2024. Jeweils ein Knochen stammtvom Schwein und von Schaf/Ziege. 26 Knochen lassen sich derGrössengruppe „Bos/Cervus“ zuordnen; sie dürften wahr-scheinlich ebenfalls vom Hausrind stammen.
Bei den Rinderknochen handelt es sich hauptsächlich umSchädelteile des Hinterhauptes, die mindestens von drei Indivi-duen stammen müssen. In allen drei Fällen handelt es sich um dierechte Schädelseite.Von zwei Schädeln ist der rechte Hornzapfenso gut erhalten, dass das Geschlecht als männlich bestimmt wer-den kann. Einer der Hornzapfen weist etwa in der Mitte ein ca.2 cm grosses, künstliches Loch auf (Abb. 30/18).Vielleicht war dasHorn ursprünglich mit einem Nagel an einer Wand aufgehängtgewesen. Vom dritten Schädel liessen sich zahlreiche Fragmentezusammensetzen. Es scheint, dass dieser Schädel schon in römi-scher Zeit bei der Sagittalnaht, der Mittellinie des Schädels,getrennt worden war und nur die rechte Hälfte in die Kellergru-be gelangte. Die Zahl drei, die speziellen Skeletteile, die jeweils glei-che Seite, das Aufhängeloch und schliesslich die Versenkung ineiner Grube lassen an einen kultischen Zusammenhang denken;
V. Funde und besondere Fundensembles römischer Zeit – 30. Die Tierknochen
2022 Tierknochenauswertung: Manuskript Sabine Schröder-Fartash, siehe auchVeszeli 2000, 174-175.2023 Stump 1986, 125.2024 Inv.-Nr. 14/9/1066; 14/9/1125.
644
Abb. 30/18. Gebäude D. Rinderschädel aus der Kellergrube(A979/B181) in Raum 1. Foto J. Stauffer, Solothurn.
Abb. 30/17. Gebäude D. Reste eines Rinderrumpfs (A983/B175) inder Brandschicht in Raum 1. Aufnahme gegen Süden.
645
die Rinderschädel könnten als apotropäisches Zeichen irgendwoam Gebäude angebracht gewesen sein (wie Anm. 2014). Später,als man den Keller in Raum 1 aufhob, setzte man sie dort bei, viel-leicht noch immer auf einen Schutz hoffend oder im Sinne einergeregelten Entsorgung heiliger Dinge (lat. favissa).
30.2.4. Gebäude E
Das im späten 2. Jahrhundert in der Nordostecke der Hof-mauer errichtete und einmal umgebaute Gebäude E fiel kurznach der Mitte des 3. Jahrhunderts einem Feuer zum Opfer(Kap. 10.4.2). In ihm könnte der Verwalter des nahen Getrei-despeichers D gewohnt haben.
Die insgesamt 1715 in oder beim Gebäude E geborgen Tier-knochen sind markant schlechter erhalten als jene der süd-licheren Grabungszonen2025. Hier, an der fast am höchsten gele-genen Partie des kiesigen Moränerückens, dürfte das Milieueinem starken Wechsel unterworfen sein, was die schlechtenErhaltungsbedingungen nicht nur der Knochen, sondern desganzen übrigen Fundmaterials erklären würde (s. Kap. 4.2; 30.1.2). Alle im folgenden archäozoologisch untersuchten Schich-ten gehören der letzten Phase an. Nicht alle Schichten und Struk-turen enthielten Funde in genügender Zahl, um statistisch ver-tretbar ausgewertet werden zu können; ausreichend viele Fundehaben einzig die Grube (A035/B236) und die Benützungsschichtin Raum 1 sowie die Grube (A049/B228) und die Fundschichtsüdöstlich des Gebäudes geliefert.
Mit 9% ist der Anteil der Wildtierknochen für das gesamteGebäude E auffallend hoch (Abb. 30/19). Er wird hauptsächlichdurch Rothirschknochen beeinflusst. Unter den Haustierkno-chen dominieren die Schweineknochen mit 36%, gefolgt vonden Rinderfragmenten mit 29%. Schaf-/Ziegenknochen sind mit19% durchschnittlich vertreten; das gleiche gilt für die Hüh-nerknochen mit 1,4%. Diese Zusammensetzung spricht für Nah-rungsüberreste eines eher gehobenen Standes; jedenfalls kanndie Deutung des Gebäudes E als Wohnhaus, vielleicht eines Ver-walters, anhand der Tierknochen bestätigt werden. Es fällt auf,dass bei den meisten Tierarten die Zahnfragmente am häufig-sten sind. Dieses Phänomen ist weniger funktional als erhal-tungsbedingt zu erklären: Zähne erhalten sich in der Regel bes-ser als Knochen und sind einfacher zu bestimmen. Es ist dahernicht zu entscheiden, ob bei Gebäude E vorwiegend Speise- oderSchlachtabfälle vorliegen.
Betrachtet man die einzelnen Ensembles (Abb. 30/19), fal-len die relativ hohen Werte für Schaf-/Ziegen- und Schweine-knochen im Innern des Gebäudes zu Lasten der Rinderknochenauf, was auf eine qualitätvolle Nahrung schliessen lässt. Dafürsprechen auch die in Grube (A035/B236) gefundenen Rothirsch-knochen und -zähne. In der gleichen Grube fanden sich Über-reste von Datteln und Weinrebenkernen (s. Kap. 28.5.3), dieebenfalls als Zeichen von Wohlstand zu werten sind.
Während die Werkgrube (A049/B228) südöstlich des Gebäu-des ebenfalls häufiger Schweine- als Rinderknochen enthielt,sind in der daneben gelegenen Fundschicht die Rinderknochenam stärksten vertreten. Daraus lässt sich schliessen, dass dieGrube offensichtlich zu einem guten Teil mit Material aus demGebäudeinnern verfüllt worden war, was Passscherben bestäti-gen (s. Abb. E/10). Allerdings fanden sich in der besagten Grube
auch Pferde-, Hunde-, Biber-, Maulwurf- und Amphibienkno-chen, die nicht zu den Speiseabfällen zu zählen sind und dahereher für ausserhalb des Gebäude E angefallenes, verschlepptesFundmaterial sprechen. Die Amphibien, die unter den Schlämm-funden zu 40% vorkommen (s. Kap. 30.5.2), könnten ein Zei-chen dafür sein, dass in der Grube ein feuchtes Milieu herrsch-te, welches diese Tiere anzog.
Unter den Knochen aus der Werkgrube (A049/B228) undder Fundschicht ausserhalb des Gebäudes befanden sich insge-samt drei Fragmente, die alle vom gleichen Säugling stammendürften (s. Kap. 31.4.2).Vielleicht gehören sie zu einem beim Baudes Gebäudes gestörten Kindergrab; ein intaktes Säuglingsgrabwurde nämlich unter einem Pfosten der um 80 errichteten Por-tikus F freigelegt (s. Kap. 10.2.1). Aus derselben Grube stammenweiter zwei Abfallfragmente der Knochenverarbeitung (Kap.27.5.2 mit Abb. 27/5/9,1), die wie die aufgefundenen Tiegel-fragmente (Kap. 25.2) auf handwerkliche Tätigkeiten hinweisen.
Aus archäozoologischer Sicht lässt sich festhalten, dass inGebäude E wohlhabende Bewohner lebten, die vielleicht eineführende Stellung im Gutsbetrieb einnahmen. Ob die Bewoh-ner selbst auf die Grosswildjagd gingen oder ob sie das Wildbretvon einem anderen Jäger, vielleicht dem Gutsherrn, erhielten,lässt sich aufgrund unserer Auswertungen nicht sagen. Die Pro-duktion von Beinartefakten – und die nachgewiesene Metall-verarbeitung – lässt indes auch auf einen Handwerker schlies-sen, der vielleicht im Dienste des Verwalters stand und im glei-chen Haus untergebracht war.
Kap. 30.2
Abb. 30/19. Gebäude E. Tierartenspektrum (n%) (s. Tab. 30/2-30/6).
2025 Die schlechte Erhaltung hat auch zum Entschluss geführt, im folgenden nurdie Fragmentzahlen, nicht aber das Gewicht zu berücksichtigen, denn die ein-zelnen Skelettelemente (Zähne, Platten- und Röhrenknochen) scheinen unter-schiedlich auf die Bodeneinflüsse reagiert zu haben, wodurch die Plattenkno-chen vor allem beim Gewicht extrem schlecht vertreten sind.

30.2.5. Portikus F
Aus den drei Schichten der um 80 n.Chr. errichteten Porti-kus F, der Steinunterlage, der Bodenoberfläche und dem Mau-erschutt der im 3. Jahrhundert verfallenen nördlichen Trenn-mauer, stammen – was auch für die übrigen Fundgattungenzutrifft – nur wenige Tierknochen (Tab. 30/2-30/7). In allen dreiSchichten sind die Rinder die am stärksten vertretene Tierart.Auf der Bodenoberfläche fanden sich ebenso viele Schaf-/Zie-gen- wie Rinderknochen, was allerdings aufgrund der geringenFundzahlen nicht sehr aussagekräftig ist. Bei den Rinderknochenfallen die gut vertretenen Stylopodiumsteile (Oberschenkel/-arme) auf, was darauf schliessen lässt, dass hier Speiseabfälle inden Boden gelangten.
30.2.6. Grabanalage H
(BRUNO KAUFMANN)Der Leichenbrand der im dritten Viertel des 2. Jahrhundert
vollzogenen Bestattung (Kap. 10.2.3; Kap. 31.2) enthielt in fastallen Fundkomplexen auch Tierknochen, welche gesamthaftrund einen Viertel (24%) des Knochengewichtes ausmachen. Dasie normalerweise wesentlich dichter gebaut sind, aber die glei-che Verbrennungszeit wie die menschlichen Reste hatten, ist ihreBrandstufe im Mittel etwas geringer als die der Menschenkno-chen (Kap. 31.2.5). Gelegentlich waren sie vermutlich auch amRand des Scheiterhaufens gelagert und daher nur angekohlt. InBiberist kamen auch bei den Tieren neben den üblichen ver-brannten Knochen vereinzelt unverbrannte Zähne und Knochenvor (Tab. 30/9).
Unverbrannte Tierknochen11 von den 59 Fundkomplexen mit Tierknochen enthielten
unverbrannte Tierreste. Diese geben – wie die unverbranntenMenschenknochen (s. Kap. 31.3) – viele Fragen auf. Mischungenzwischen unverbrannten und verbrannten Tierknochen sind all-gemein sehr selten. In Biberist sind es die beiden Pferdeknochen(ein Wirbel und ein unterer Eckzahn), von den übrigen Tierenfast nur Kiefer- und Zahnreste sowie Teile des Fussskelettes. EineAusnahme bilden nicht näher bestimmbare Röhren-knochenfragmente (einmal evtl. Mensch?) sowie ein Schweine-humerus und ein Femur von Schwein oder von Schaf/Ziege2026.Da viele dieser Tierknochen metamorph sind, stellt sich die Frage,ob es sich nicht um gekochte Speisebeigaben handeln könnte.Gegen diese Annahme spricht allerdings der Umstand, dass dieunverbrannten Menschenknochen auch metamorph sind.
Die Tierarten.Das Huhn ist durch Wirbel, Rippen, zwei Coracoide, vier
Langknochen und ein Tarsometatarsus nachgewiesen; alleKnochen können vom gleichen Tier stammen.
Der Hund ist durch einen Unterkiefer belegt sowie mögli-cherweise durch ein Hüftbeinfragment und eine Eckzahnwur-zel, doch ist das erhaltene Wurzelstück zu klein, um es eindeu-tig dem Hund zuschreiben zu können2027. Es könnte auch voneinem Fuchs oder einer (männlichen) Wildkatze stammen. Fallses ein Hund war, so hatte er etwa die Grösse eines Spitzes.
Die beiden erhaltenen Pferdeknochen sind unverbrannt.
Dies gilt sowohl für den Wirbel wie auch für den unteren Eck-zahn. Die Zuordnung ist bei beiden Objekten nicht ganz gesi-chert, auch wenn sehr wahrscheinlich. Die erhaltenen Reste deu-ten auf ein kleingewachsenes Tier hin.
Das Rind ist durch zwei Hornzapfen, ein Hirnschädelfrag-ment, sechs Zähne und ein Unterkieferfragment belegt. Nebstzahlreichen Röhrenknochenfragmenten sind je zwei Wirbel undRippen und Fragmente vom distalen Radius, der proximalenTibia, zwei Calcanei, vier Mittelfussknochen und drei Zehen-knochen näher lokalisierbar. Es dürfte sich um Reste eines nachrömischen Verhältnissen etwa mittelgrossen Tieres handeln.Unverbrannte Rinderknochen sind nicht belegt.
Von Schaf und/oder Ziege sind folgende Knochen belegt:Vom Schädel liegen ein Hornzapfen, acht Zähne und ein un-verbrannter Unterkiefer vor; das postkraniale Skelett wirddurch zahlreiche Röhrenknochenfragmente sowie durch einenWirbel, einen proximalen Humerus, drei Fusswurzelknochen(zwei Calcanei, ein Astragalus), zwei Mittelfussknochen und vierZehenknochen vertreten. Es handelt sich um Reste von min-destens drei Tieren: zwei etwa ausgewachsene (belegt durchunverbrannte sowie verbrannte Molaren) und ein Milchlammoder Zicklein, vertreten durch einen unverbrannten Unterkiefer.
Zahlenmässig dürften die Schweineknochen diejenigen vonSchaf oder Ziege leicht übertreffen. Auch bezüglich der Min-destindividuenzahl dürften etwa vier bis fünf Schweine dreiSchaf/Ziegen gegenüberstehen. Alle Tiere wurden vor dem voll-ständigen Auswachsen geschlachtet, doch lassen sich keine Span-ferkel nachweisen. Die Schweine sind mit allen Körperregionenvertreten, aber auch hier hat man den Eindruck, dass vorwie-gend minderwertige Fleischstücke mitgegeben wurden. So lie-gen vom Hirnschädel vier Stücke vor, vom Gesichtsschädel zweiOberkiefer und vier Zähne. Am häufigsten sind die Wirbel-fragmente (14). Auch die Unterarme (drei Radien, sechs Ellen)und Unterschenkel (zwei Tibien, eine Patella) und die Füsse (vierAstragali, zwei Calcanei, zwei weitere Fusswurzel- und siebenZehenknochen) gehören eher zu den minderen Fleischstücken.Die hochwertigen sind durch eine Rippe, vier Humeri, drei Hüft-beine und zwei Oberschenkelbeine belegt; zu bedenken ist aller-dings, dass die besten Stücke keine Knochen haben und sichsomit nicht nachweisen lassen! Ein distaler Femur („Jambon-neau“) ist unverbrannt. Wie die drei Molaren und der Prämo-lar zu interpretieren sind, ist nicht klar. Die postmortalenKnochenveränderungen an einem Humerus und an anderenRöhrenknochen können am ehesten damit erklärt werden, dassgekochte Fleischstücke mitgegeben wurden2028. Wenn wir dieunverbrannten Schweineknochen miteinbeziehen, dürfen wirmit einer Fleischbeigabe von fünf bis sieben Schweinen rechnen.
Nicht zu den Haustieren gehören die Knochenreste einesSingvogels, die im unteren Teil der Kremationsgrube lagen, wodie Leichen beim Einbrechen des Scheiterhaufens zu liegengekommen waren (s. Kap. 10.2.3), und der ebenfalls verbrann-te Knochen eines Amphibiums (Frosch, Kröte?)2029.
V. Funde und besondere Fundensembles römischer Zeit – 30. Die Tierknochen
2026 Inv.-Nr.14/9/1150; 14/9/1496; 14/9/1497; 14/9/1536 (evtl. Mensch?).2027 Inv.-Nr.14/9/1293; 14/9/1381; 14/9/1545.2028 Inv.-Nr.14/9/1497.2029 Inv.-Nr.14/9/1460 (Amphibium); 14/9/1515 (Singvogel).
646

647
Einige vor allem sehr kleine Knochensplitter konnten kei-ner Tierart zugewiesen werden. Bei der Mehrzahl handelt es sichum Fragmente, die ihrer Grösse nach zu Rind, Schwein oderSchaf/Ziege gehören können. Anhand der wenigen erhaltenenGelenkanteile – Wirbel und Röhrenknochen mit durchwegsnoch offener Epiphysenfuge – handelt es sich um nicht ausge-wachsene Tiere, weshalb eine Zuordnung anhand der Knochen-grösse nicht in Frage kommt.
FazitAnhand der Knochen können ein Huhn, ein Hund von
Spitzgrösse, ein kleines Pferd und ein mittelgrosses Rind nach-gewiesen werden. Hinzu kommen zwei ältere Schafe oder Zie-gen und ein Jungtier (Lamm oder Zicklein) sowie fünf bis sie-ben Schweine. Wenige Knochen belegen ein Amphibium(Frosch oder Kröte) und einen Singvogel. Trotz dieser grossenListe weisen die erhaltenen Knochen vorwiegend auf minder-wertige Fleischstücke hin; am häufigesten kommen Knochen desSchädels und der Unterschenkel sowie der Füsse vor. Diese zahl-reichen tierischen Beigaben sprechen für die Bedeutung der inder Grabanlage H bestatteten Personen.
Es liegen keine anderen Tierarten vor als im übrigen Mate-rial der Villa. Bei den unverbrannten Knochen könnte es sichauch um Abfall handeln, der im Innenhof schon herum lag undzufällig in die Kremationsgrube gelangt war.
Grube (A004/B135) (Sabine Deschler-Erb) Nahe der Grabumfriedung fand sich in einer Grube (A004/
B135) ein Rinderschädel (Bukranion) (s. Abb. 10/H18), auf des-sen knöchernen Hornzapfen z.T. noch die Hornscheide lag. Eshandelt sich dabei um einen der seltenen mir bekannten Fällevon Hornerhaltung in römischer Zeit. Ursprünglich war derSchädel wahrscheinlich beim Grab als apotropäisches Zeichenaufgehängt und wurde bei der Aufhebung des Grabes im zwei-ten Viertel des 3. Jahrhunderts hier vergraben (wie Anm. 2014;s. auch Kap. 10.2.3).
30.2.7. Gebäude J und Umgebung
Vom Gebäude J, das aufgrund seiner Nähe zum Tor als Vili-cushaus gedeutet wird (Kap. 10.3.5-6), liegen am meisten Kno-chen vor, wobei der Anteil pro m3 durchaus nicht aus dem Rah-men fällt (s. Abb. 30/1). Westlich des Steinbaus fanden sich soviele Knochen, dass spezielle Schlachtabfälle zu vermuten wa-ren und eine archäozoologische Untersuchung lohnend erschien.
Holzbau JNach Fragmentzahlen sind in dem Ende des 1. Jahrhunderts
abgebrannten Holzbau J (Kap. 10.3.5) die Schaf-/Ziegenknochenabsolut dominant (Abb. 30/20). Diese Dominanz scheint sich bisin Steinbauphase 2 weiterzuziehen (s. Abb. 30/25), allerdings istdie statistische Basis für diese Phase relativ gering. Nur drei Kno-chen können als Schaf identifiziert werden. Da unter den übri-gen Knochen keine einzige Ziege bestimmt werden kann, ist nichtauszuschliessen, dass die nicht näher als Schaf oder Ziegebestimmbaren Knochen von Schafen stammen. Die zweitwich-tigste Tierart ist das Schwein, dessen Knochen indes nur um die20% der Bestimmbaren ausmachen. Schlecht vertreten sind dieRinderknochen, die in allen Schichten unter 20% liegen.
Weiter fanden sich relativ viel Hühnerknochen. Zu erwäh-nen sind auch zwei Hasenknochen aus der Brandschicht(Tab. 30/4). All diese Tiere sprechen für eine qualitätvolle Flei-schnahrung. Der hohe Anteil an Hundeknochen dürfte hinge-gen nichts mit der Ernährung zu tun haben, sondern könnte miteinem verlagerten Skelett in Zusammenhang stehen. In denSchlämmfunden fanden sich Reste von Aesche, Schleie undBrachse sowie mehreren Singvögeln, was ebenfalls für eine qua-litätvolle Nahrung spricht (Kap. 30.5.2).
Das Nutztierspektrum des Holzbaus J ist als ausserge-wöhnlich zu bezeichnen. Kaum eine Fundstelle in den westlichenProvinzen hat einen annähernd so hohen Anteil an Schaf-/Zie-genknochen geliefert2030. Eine Ausnahme bildet ein Ensemble ausdem frühkaiserzeitlichen Militärlager der Kaiseraugster Unter-stadt2031. Allerdings erreichen dort die Schaf-/Ziegenknochenunter den bestimmbaren Tierknochen nur knapp 40%, gefolgtvon Schweine- und Rinderknochen. Die Tierartenzusammen-setzung hat dazu geführt, die Herkunft der hier Fleisch konsu-mierenden Truppen im mediterranen Bereich zu vermuten. Ineiner jüngeren Untersuchung hat sich dann aufgezeigen lassen,dass der Anteil der kleinen Wiederkäuer in den Anfangsphasenrömischer Siedlungen immer relativ hoch liegt, um dann im Ver-laufe der Zeit allmählich abzunehmen2032. Dies dürfte mit dernoch grösseren Bedeutung der Subsistenzwirtschaft und einemnicht vollständig entwickelten Wirtschaftsnetz in den Anfängender Gutshofwirtschaft zu erklären sein.
Der hohe Anteil alt-adulter Schafe/Ziegen in Holzbau J deu-tet daraufhin (Abb. 30/21), dass in Biberist diese Tiere in ersterLinie für die Produktion von Sekundärprodukten wie Wolle oderMilch gehalten wurden. Dass ihr Fleisch dann schliesslichgegessen wurde, spricht nicht unbedingt für einen gehobenen
Kap. 30.2
2030 Peters 1998, 429, Abb. 77-83.2031 Deschler-Erb in Deschler-Erb u.a. 1991.2032 Deschler-Erb 1998.
Abb. 30/20. Holzbau J. Tierartenspektrum (n%) (s. Tab. 30/2-30/6).

Ernährungsstil, sondern eher für bäuerlichen Pragmatismus. Imerwähnten Kastell der Kaiseraugster Unterstadt scheinen dieSchafe/Ziegen durchschnittlich denn auch jünger gewesen zusein als in Biberist2033. In der Holzbauphase von Gebäude J kannsomit aufgrund der archäozoologischen Auswertungen aufeine mit der Viehzucht der Villa eng verbundene Bewohnerschaftgerechnet werden, die neben etwas Hühner- und Hasenfleischnicht übermässig qualitätvolles Fleisch konsumierte. Es istsomit weniger mit einer Veteranenfamilie zu rechnen, die ur-sprünglich aus dem Mittelmeerraum stammte und deshalbgerne Schaf/Ziegenfleisch konsumierte, sondern eher mit einemauf die sekundäre Produktion von Wolle und/oder Milch spe-zialisierten bäuerlichen Betrieb.
Die Tierknochen stammen aus drei Schichten, die in undüber der mutmasslichen Küche zum Vorschein kamen: Brand-schicht, Brandschutt und der beim Bau der Phase 2 des Stein-baus J in Mauerausbruchgruben verlagerte Teil des Brand-schuttes. Unterschiede zwischen den Schichten ergeben sich imAnteil der verbrannten Tierknochen: Während in der eigent-
lichen Brandschicht nur 5% Brandspuren aufweisen, sind es inder Brandschuttschicht 44% und in den Mauerausbruchgrubensogar 74% (Abb. 30/23). Dies belegt zum einen, dass die Brand-spuren nicht bei der Zubereitung der Fleischmahlzeiten, sondernbeim Brand des Gebäudes entstanden waren. Dabei müssen dieKnochen aus den Schuttschichten dem Feuer stärker ausgesetztgewesen sein als jene der Brandschicht, die vielleicht schon seiteiniger Zeit in der dabei verbrannten Benützungsschicht ein-gelagert waren. Auch anhand des Tierartenspektrums lassen sichgewisse Unterschiede erkennen: In der eigentlichen Brandschichtmachen die kleinen Wiederkäuer lediglich 54% der bestimm-baren Tierknochen aus, im Brandschutt sind es 67% und in demin die Mauerausbruchgruben verlagerten Brandschutt 63%(s. Abb. 30/20). Im Brandschutt ist das viel Fleisch tragende Sty-lopodium deutlich besser vertreten als in der Brandschicht(Abb. 30/24); diese Knochen könnten von Fleischstücken stam-men, die beim Brand noch in der Küche auf Gestellen lagen oderim Obergeschoss, wo ausser Gerste (Kap. 28.5.5) noch andereVorräte lagerten, wie die in den Brandschuttschichten haupt-sächlich auftretenden Dolien- und Amphorenscherben zeigen.Die höheren Schaf-/Ziegenanteile in den Schuttschichten gehenzu Lasten der hier geringeren Anteile an Rindern. Diese Beob-achtungen finden gewisse Entsprechungen bei der Untersuchungder Keramik: Während 70% des Tafelgeschirrs, insbesondere dievielen Servierschüsseln, der Fundlage nach mit den einstür-zenden Lehmwänden hinuntergefallen waren, müssen dieKochtöpfe nahe am Boden gestanden sein (s. Abb. 10/J6). Inrömischer Zeit scheint vorwiegend Fleisch von kleinen Tierennoch am Knochen anhaftend serviert worden zu sein, währenddie Knochen von Rindern hauptsächlich in der Küche blieben,nachdem sie ihren Dienst z.B. als Suppenmarkknochen erfüllthatten, was sich in der Brandschicht durch die höheren Rin-deranteile manifestiert.
V. Funde und besondere Fundensembles römischer Zeit – 30. Die Tierknochen
2033 Wie Anm. 2032, 127 Abb. 82.
648
Abb. 30/21. Holzbau J. Alterstruktur (n%) der Schaf/Ziegenreste (n=21).
Abb. 30/23. Holzbau J. Anteil (n%) verbrannter Tierknochen (Tab. 30/1).
Abb. 30/22. Steinbau J. Altersstruktur (n%) der Schaf/Ziegen im Ver-gleich mit Holzbau J.
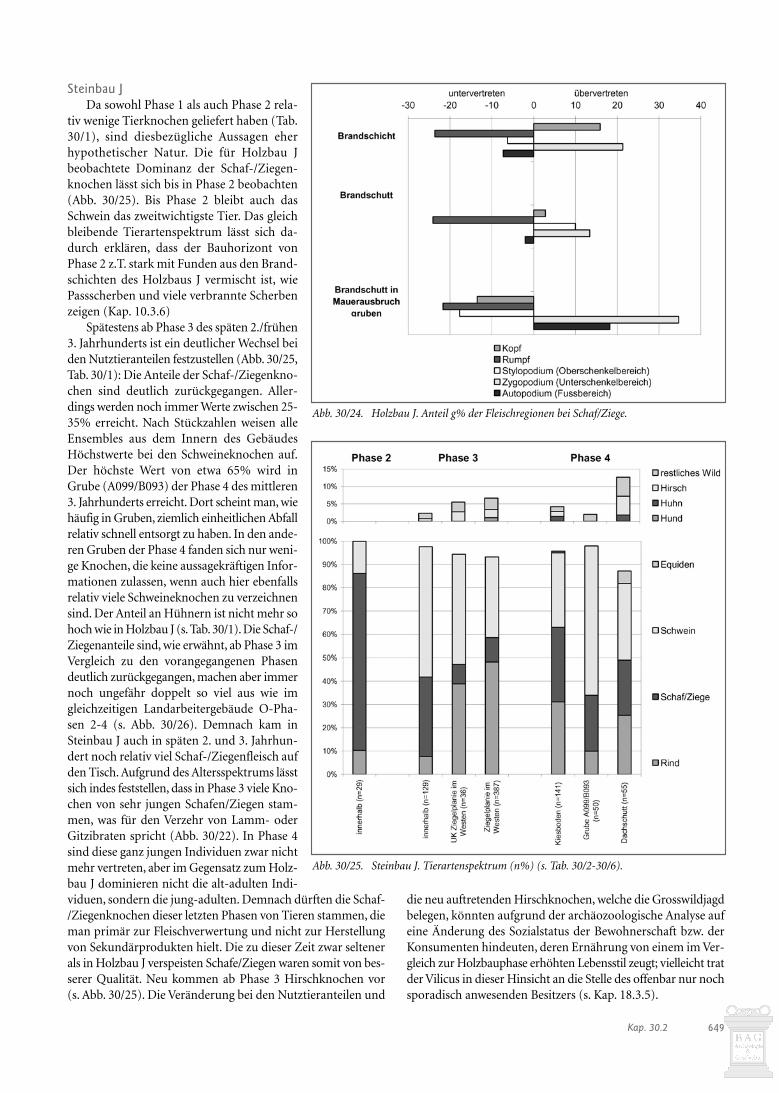
649
Steinbau JDa sowohl Phase 1 als auch Phase 2 rela-
tiv wenige Tierknochen geliefert haben (Tab.30/1), sind diesbezügliche Aussagen eherhypothetischer Natur. Die für Holzbau Jbeobachtete Dominanz der Schaf-/Ziegen-knochen lässt sich bis in Phase 2 beobachten(Abb. 30/25). Bis Phase 2 bleibt auch dasSchwein das zweitwichtigste Tier. Das gleichbleibende Tierartenspektrum lässt sich da-durch erklären, dass der Bauhorizont vonPhase 2 z.T. stark mit Funden aus den Brand-schichten des Holzbaus J vermischt ist, wiePassscherben und viele verbrannte Scherbenzeigen (Kap. 10.3.6)
Spätestens ab Phase 3 des späten 2./frühen3. Jahrhunderts ist ein deutlicher Wechsel beiden Nutztieranteilen festzustellen (Abb. 30/25,Tab. 30/1): Die Anteile der Schaf-/Ziegenkno-chen sind deutlich zurückgegangen. Aller-dings werden noch immer Werte zwischen 25-35% erreicht. Nach Stückzahlen weisen alleEnsembles aus dem Innern des GebäudesHöchstwerte bei den Schweineknochen auf.Der höchste Wert von etwa 65% wird inGrube (A099/B093) der Phase 4 des mittleren3. Jahrhunderts erreicht. Dort scheint man, wiehäufig in Gruben, ziemlich einheitlichen Abfallrelativ schnell entsorgt zu haben. In den ande-ren Gruben der Phase 4 fanden sich nur weni-ge Knochen, die keine aussagekräftigen Infor-mationen zulassen, wenn auch hier ebenfallsrelativ viele Schweineknochen zu verzeichnensind. Der Anteil an Hühnern ist nicht mehr sohoch wie in Holzbau J (s. Tab. 30/1). Die Schaf-/Ziegenanteile sind, wie erwähnt, ab Phase 3 imVergleich zu den vorangegangenen Phasendeutlich zurückgegangen, machen aber immernoch ungefähr doppelt so viel aus wie imgleichzeitigen Landarbeitergebäude O-Pha-sen 2-4 (s. Abb. 30/26). Demnach kam inSteinbau J auch in späten 2. und 3. Jahrhun-dert noch relativ viel Schaf-/Ziegenfleisch aufden Tisch. Aufgrund des Altersspektrums lässtsich indes feststellen, dass in Phase 3 viele Kno-chen von sehr jungen Schafen/Ziegen stam-men, was für den Verzehr von Lamm- oderGitzibraten spricht (Abb. 30/22). In Phase 4sind diese ganz jungen Individuen zwar nichtmehr vertreten, aber im Gegensatz zum Holz-bau J dominieren nicht die alt-adulten Indi-viduen, sondern die jung-adulten. Demnach dürften die Schaf-/Ziegenknochen dieser letzten Phasen von Tieren stammen, dieman primär zur Fleischverwertung und nicht zur Herstellungvon Sekundärprodukten hielt. Die zu dieser Zeit zwar seltenerals in Holzbau J verspeisten Schafe/Ziegen waren somit von bes-serer Qualität. Neu kommen ab Phase 3 Hirschknochen vor(s. Abb. 30/25). Die Veränderung bei den Nutztieranteilen und
die neu auftretenden Hirschknochen, welche die Grosswildjagdbelegen, könnten aufgrund der archäozoologische Analyse aufeine Änderung des Sozialstatus der Bewohnerschaft bzw. derKonsumenten hindeuten, deren Ernährung von einem im Ver-gleich zur Holzbauphase erhöhten Lebensstil zeugt; vielleicht tratder Vilicus in dieser Hinsicht an die Stelle des offenbar nur nochsporadisch anwesenden Besitzers (s. Kap. 18.3.5).
Kap. 30.2
Abb. 30/24. Holzbau J. Anteil g% der Fleischregionen bei Schaf/Ziege.
Abb. 30/25. Steinbau J. Tierartenspektrum (n%) (s. Tab. 30/2-30/6).
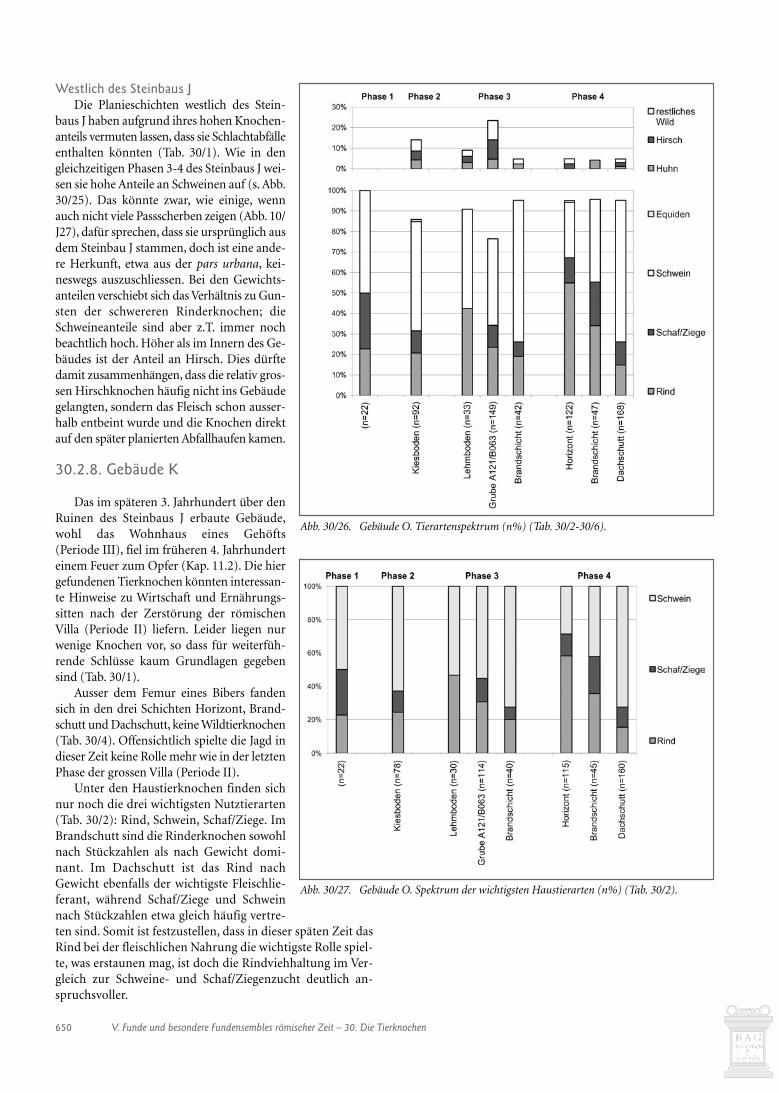
Westlich des Steinbaus JDie Planieschichten westlich des Stein-
baus J haben aufgrund ihres hohen Knochen-anteils vermuten lassen, dass sie Schlachtabfälleenthalten könnten (Tab. 30/1). Wie in dengleichzeitigen Phasen 3-4 des Steinbaus J wei-sen sie hohe Anteile an Schweinen auf (s. Abb.30/25). Das könnte zwar, wie einige, wennauch nicht viele Passscherben zeigen (Abb. 10/J27), dafür sprechen, dass sie ursprünglich ausdem Steinbau J stammen, doch ist eine ande-re Herkunft, etwa aus der pars urbana, kei-neswegs auszuschliessen. Bei den Gewichts-anteilen verschiebt sich das Verhältnis zu Gun-sten der schwereren Rinderknochen; dieSchweineanteile sind aber z.T. immer nochbeachtlich hoch. Höher als im Innern des Ge-bäudes ist der Anteil an Hirsch. Dies dürftedamit zusammenhängen, dass die relativ gros-sen Hirschknochen häufig nicht ins Gebäudegelangten, sondern das Fleisch schon ausser-halb entbeint wurde und die Knochen direktauf den später planierten Abfallhaufen kamen.
30.2.8. Gebäude K
Das im späteren 3. Jahrhundert über denRuinen des Steinbaus J erbaute Gebäude,wohl das Wohnhaus eines Gehöfts(Periode III), fiel im früheren 4. Jahrhunderteinem Feuer zum Opfer (Kap. 11.2). Die hiergefundenen Tierknochen könnten interessan-te Hinweise zu Wirtschaft und Ernährungs-sitten nach der Zerstörung der römischenVilla (Periode II) liefern. Leider liegen nurwenige Knochen vor, so dass für weiterfüh-rende Schlüsse kaum Grundlagen gegebensind (Tab. 30/1).
Ausser dem Femur eines Bibers fandensich in den drei Schichten Horizont, Brand-schutt und Dachschutt, keine Wildtierknochen(Tab. 30/4). Offensichtlich spielte die Jagd indieser Zeit keine Rolle mehr wie in der letztenPhase der grossen Villa (Periode II).
Unter den Haustierknochen finden sichnur noch die drei wichtigsten Nutztierarten(Tab. 30/2): Rind, Schwein, Schaf/Ziege. ImBrandschutt sind die Rinderknochen sowohlnach Stückzahlen als nach Gewicht domi-nant. Im Dachschutt ist das Rind nachGewicht ebenfalls der wichtigste Fleischlie-ferant, während Schaf/Ziege und Schweinnach Stückzahlen etwa gleich häufig vertre-ten sind. Somit ist festzustellen, dass in dieser späten Zeit dasRind bei der fleischlichen Nahrung die wichtigste Rolle spiel-te, was erstaunen mag, ist doch die Rindviehhaltung im Ver-gleich zur Schweine- und Schaf/Ziegenzucht deutlich an-spruchsvoller.
V. Funde und besondere Fundensembles römischer Zeit – 30. Die Tierknochen650
Abb. 30/26. Gebäude O. Tierartenspektrum (n%) (Tab. 30/2-30/6).
Abb. 30/27. Gebäude O. Spektrum der wichtigsten Haustierarten (n%) (Tab. 30/2).

651
30.2.9. Gebäude O und Umgebung
InnenraumDas im zweiten Viertel des 2. Jahrhunderts errichtete,
mehrmals umgebaute Landarbeiterhaus O fiel kurz nach derMitte des 3. Jahrhunderts einem Feuer zum Opfer (Kap. 10.3.10). Während die Phasen 1-2 mit gut 30 Tierknochen pro m3
im Rahmen liegen, stellen die folgenden Phasen 3-4 mit überhundert die fundreichsten Ensembles der gesamten Villa dar(Tab. 30/1).
Mit Ausnahme des Horizontes der Phase 4 dominieren inallen Phasen die Schweineknochen nach Fragmentzahlen mitWerten zwischen 42% und 72% unter den drei wichtigstenNutztierarten (Abb. 30/26). Aufgrund der Gewichtsanteile sinddie Schweineknochen ebenfalls durchwegs gut vertreten, auchwenn die schwereren Rinderknochen eine deutlich grössereBedeutung haben. Die Schaf-/Ziegenknochen weisen in Phase 1einen Anteil von 27% auf. Allerdings ist dieser Wert angesichtsder geringen Fundmenge vermutlich nicht sehr aussagekräftig.In den folgenden Phasen liegt der Anteil der Schaf-/Ziegen-knochen hingegen meist deutlich unter 20%. Aufgrund desGrundrisses wird für Phase 2 mit einem Wechsel von zwei zueiner Familie gerechnet. Aufgrund der Tierknochen zeichnet sichindes kein Bruch ab. Die Nutztieranteile scheinen sich konti-nuierlich zu entwickeln, eine Änderung der Ernährungssittenzeichnet sich nicht ab.
Die zur Phase 3 des frühen mittleren 3. Jahrhunderts gehö-rige Grube (A121/B063) barg insgesamt 292 Tierknochen,wovon allerdings die Hälfte unbestimmbar bleibt (Tab. 30/1).Unter den bestimmbaren Tierknochen sind 46% dem Haus-schwein zuzuweisen (Abb. 30/26). Auch nach Gewicht sind dieSchweine mit 32% die wichtigste Tierart (Abb. 30/27). Bei derBetrachtung der Fleischregionen (Abb. 30/28) fällt neben derüblichen Untervertretung der Rumpfteile die Übervertretung desStylopodiums und des Zygopodiums auf, was auf Speiseabfäl-le hindeutet.
Mit den relativ geringen Schaf-/Ziegenanteilen ab Phase 2unterscheidet sich das Landarbeiterhaus O deutlich von dengleichzeitigen Phasen 3-4 des Vilicushauses J (s. Abb. 30/26 mitAbb. 30/25): Dort erreichen die Schaf-/Ziegenknochen Wertevon um die 30%. Daraus lassen sich unterschiedliche Essittenvermuten.
Weitere Unterschiede zwischen diesen beiden Gebäuden sindbei den Anteilen der Hühnerknochen festzustellen: Währenddiese in Steinbau J nur in Phase 4 mit Werten unter 4% vor-kommen, sind sie in Gebäude O in allen Phasen stetig vertre-ten und erreichen Werte bis über 5%. Auch die unter die Gross-gruppe Geflügel eingeordneten Enten, Tauben und unbe-stimmbaren Haus-/Wildvogelknochen kommen im Gegensatzzu Steinbau J mit mehreren Vertretern in allen Phasen von Ge-bäude O vor (Tab. 30/3 und 30/6). Lag die Betreuung des Geflü-gelhofes in den Händen der Landarbeiter von Gebäude O? Vordem Bau des Gebäudes, in der Phase 1 der grossen Villa (Peri-ode II), scheint diese Aufgabe noch in den Händen der Bewoh-ner des Holzbaus J gelegen zu haben, da hier der Hühneranteilrelativ hoch ist (S. Abb. 30/20). In Gebäude O fanden sich nebenden zahlreichen Vogelknochen auch Eierschalenfragmente(Kap. 30.5.2). Diese könnten von Brucheiern stammen, die bei
der Eiproduktion immer wieder anfallen. Sie sind nicht langehaltbar und können nicht auf dem Markt verkauft werden,daher werden sie gewöhnlich direkt auf dem Bauernhof kon-sumiert. Wie wichtig Vogeleier für die gehobene römischeErnährung – vor allem als Vorspeise – auch in unseren Regio-nen war, lässt sich z.B. an den an Metalllöffeln anhaftenden Eier-schalen feststellen2034. Es dürfte sich bei der Eierproduktion umein einträgliches Geschäft gehandelt haben.
Legehennen werden gewöhnlich im fortgeschrittenen Altergeschlachtet und taugen höchstens noch als Suppenhühner.Somit mag es nicht erstaunen, dass in Biberist – im Gegensatzzur Koloniestadt Augusta Raurica – die Landarbeiterfamilienvon Gebäude O relativ viel Geflügelfleisch konsumierten2035.
Bei den Hirschknochenanteilen liegen in den Gebäuden Jund O vergleichbare Häufigkeiten vor. Höchste Werte findensich in Grube (A121/B063) des Gebäudes O-Phase 3 (Tab. 30/4)und im Dachschutt des Steinbaus J-Phase 4 (s. Abb. 30/2), diebeide ins zweite Drittel des 3. Jahrhunderts datieren. Auch in dergleichzeitigen Grube (A035/B236) des Gebäudes E finden sichrelativ hohe Hirschanteile (s. Abb. 30/19). Offensichtlich wurdein allen Wohngebäuden Hirschfleisch konsumiert und derAbfall hauptsächlich in Gruben entsorgt. Da sich darunter keineKnochenteile finden, die geräuchert werden (Scapula, Mandi-bula), dürfte es sich nicht um konserviertes Fleisch gehandelthaben. Die Beobachtung, dass zwar zwischen den einzelnenWohngebäuden des mittleren 3. Jahrhunderts Unterschiede inBezug auf die Nutztieranteile bestehen, Hirschfleisch aberüberall in gleichem Masse konsumiert wurde, lässt daraufschliessen, dass entweder alle Bewohner auf die Jagd gingen oderdass sie das Wildfleisch von einem oder mehreren Jäger bezo-gen (s. Kap. 30.2.7).
Westlich des Gebäudes Aus den Schichten westlich des Landarbeiterhauses O wer-
den nur die Knochen des Horizontes und Brandschicht sowieder Grube (A110/B050), die alle der Phase 1 des zweiten Vier-tels des 2. Jahrhunderts zuzuordnen sind, in die Untersuchungeinbezogen. Sie sind Teil des Vorplatzes der Phase 1 des Gebäu-
Kap. 30.2
2034 Riha/Stern 1982, 10-11; zur Bedeutung der Eier: Weeber 1995, 9-11; s. auchdas Preisedikt des Diokletian von 301 n.Chr.: Vier Eier kosteten gleichviel wieein halbes Pfund Rindfleisch (Weeber 1995, 285).2035 Zu Augst: Schibler/Furger 1988, 156-177.
Abb. 30/28. Gebäude O-Phase 3. Anteil (g%) der Fleischregionen beiSchwein in der Grube (A121/B063).

des O, die relativ wenig Tierknochen geliefert hat (Tab. 30/1),so dass ein Vergleich nur hypothetische Aussagen zulässt. Trotz-dem ist festzustellen, dass sowohl im Horizont westlich desGebäudes als auch im Innern die Schweineknochen nach Frag-mentzahlen am stärksten vertreten sind (Tab. 30/2). Ein Zu-sammenhang zwischen diesen Funden ist somit möglich. In derGrube (A110/B050) kamen gleich viele Schweine- wie Rinder-fragmente zum Vorschein; ein Bezug zum Gebäudeinnern istsomit ebenfalls nicht auszuschliessen.
Westlich des Gebäudes fanden sich indes auch Tierarten, dieim Innern des Gebäudes fehlen (Tab. 30/3-30/4): Hund, Huhn,Gans, Wildschwein und Hirsch. Diese Funde könnten darauf-hin deuten, dass zwar ein grosser Teil der Knochen ursprüng-lich in Gebäude O angefallen war, dass aber vor dem Haus eineDurchmischung mit anderen Abfällen stattfand.
30.2.10. Gebäude P
Das zu Beginn des 2. Jahrhunderts errichtete Gebäudewurde beim Bau des Landarbeiterhauses O im früheren 2. Jahr-hundert abgebrochen (Kap. 10.3.9). Das meiste Fundmaterialstammt aus der Abbruchschicht (Tab. 30/1). Da diese aber nur38 bestimmbare Tierknochen geliefert hat, sind nur hypothe-tische Aussagen möglich. Möglicherweise hängt der geringeFundanfall mit der Interpretation dieses Gebäudes als Wachhauszusammen, wo vielleicht weniger Essabfälle zu erwarten sind alsin einem Wohnhaus.
Trotz der geringen Fundmenge fällt auf, dass die Schaf-/Zie-genknochen in der Abbruchschicht eine untergeordnete Rollespielen (Tab. 30/2), im Gegensatz zu den Rinder- und Schwei-neknochen.
30.2.11. Gebäude Q
Das erst in der letzten Siedlungsphase 7 der grossen Villa(Periode II), im zweiten Viertel des 3. Jahrhunderts, vermutlichnach einem Besitzerwechsel errichtete Gebäude fiel kurz nachder Jahrhundertmitte einem Feuer zum Opfer (Kap. 10.3.12).In der Nordwestecke war ein Konservierungsofen eingebaut, des-sen Funktion nur im Zusammenspiel der Auswertung derBaustrukturen, der archäozoologischen Untersuchung sowie derAnalyse einer amorphen Masse aus den Schlämmrückständen(Kap. 30.6) gedeutet werden kann.
Die meisten Tierknochen (n=257) stammen aus dem Dach-schutt und sind trotz guter Erhaltung nur zu 52% bestimmbar(Tab. 30/1). Die übrigen Schichten – Grube (A125/B44) undEinfüllung der Ofenkanäle – haben im einzelnen zwar relativwenig bestimmbares Material geliefert, ergeben aber dennochinteressante Informationen für die Interpretation des Gebäu-des.
Im Fundmaterial des Dachschuttes lässt sich eine sehr auf-fällige Zusammensetzung der Tierarten feststellen (Abb. 30/29):Nach Gewicht sind die Hirschknochen mit 56% dominant, nachStückzahlen werden sie von den Schweinen leicht übertroffen.Rind und Schaf/Ziege spielen eine untergeordnete Rolle. DieseZusammensetzung ist innerhalb der Villa von Biberist ausser-gewöhnlich und spricht für eine spezielle Funktion des Gebäu-des.
Ungewöhnlich ist auch die Skeletteilzusammensetzung derSchweineknochen (Abb. 30/31): Kopf-, Zygopodium (Unterarm-/Unterschenkelbereich) und Autopodium (Fussbereich) nähernsich den Anteilen beim Vergleichsskelett. Die Rumpfteile sind,wie gewohnt, deutlich untervertreten, hingegen lässt sich beimfleischreichen Stylopodium (Oberarm-/Oberschenkelbereich)eine krasse Übervertretung feststellen. Diese wird hauptsächlichdurch die Humeri verursacht, während die Schulterblätterdeutlich seltener sind. Wahrscheinlich liegen hier Abfälle vor, diesich bei der Vorbereitung von Schweineschultern zum Konser-vieren ergeben: Das Schweinefleisch wird bereits in grobe Stü-cke zerlegt in die entsprechende Anlage gebracht. Während dasSchulterblatt ganz im umgebenden Muskelfleisch belassenwird und mit in den Ofen kommt, entfernt man den Humerus,der als Abfall liegen bleibt. Das Schulterblatt fällt erst am Kon-sumationsort als Abfall an. Ob eventuell auch andere Teile, wiez.B. Backenmuskel, Hinterschinken oder Speckseiten verarbei-tet wurden, muss aufgrund der unspezifischen Verteilung desSkeletteilspektrums offen bleiben.
Bei den Hirschknochen des Dachschuttes ist hingegen – wieim gesamten Material der Phase 7 der grossen Villa (Periode II)zu beobachten (Abb. 30/30) – in erster Linie das Autopodiumdominant. Diese Übervertretung ist damit zu erklären, dass dieHirsche bereits auf dem Jagdplatz grob zerlegt wurden, und die
V. Funde und besondere Fundensembles römischer Zeit – 30. Die Tierknochen652
Abb. 30/29. Gebäude Q. Tierartenspektrum (n%) aus dem Dachschutt(Tab. 30/2; 30/4).
Abb. 30/30. Fleischregionen bei Rothirsch (Periode II-Phase 7).

653
grossen Fleischstücke meistens ohne Knochen und ins Fell ein-gewickelt, in dem noch die Fussknochen steckten, in den Guts-hof gelangten (s. Kap. 30.3.9). Der hohe Anteil an Hirschknochendeutet auf eine grosse Bedeutung des Hirschfleisches in Gebäu-de Q hin; wahrscheinlich wurde es hier ebenfalls konserviert.
In der zum Gebäude Q gehörenden Grube (A125/B44) sindSchweine- und Hirschknochen indes von geringer Bedeutung,dafür spielen die Rinderknochen die Hauptrolle (Tab. 30/2),wobei es sich hauptsächlich um Rumpf- und Stylopodiums-teile handelt. Möglicherweise sind diese Knochen ebenfalls inZusammenhang mit der Haltbarmachung von Fleisch zu sehen.Sie erinnern an den Rinderrumpf, der sich im Gebäude D be-fand, als dieses wohl zur selben Zeit niederbrannte wie Gebäu-de Q (Kap. 17.4.7). Möglicherweise war es üblich, grössereFleischstücke nach dem Schlachten im kühlen, dunkeln Erdge-schoss des Horreums einige Tage abhangen zu lassen (s. Kap.30.2.3), bevor man es ins Gebäude Q transportierte, wo es fürdie Konservierung entbeint wurde. Dürfen wir die grosse Gru-be in Gebäude Q in Zusammenhang mit der Schlachtung oderder Konservierung deuten? Vielleicht war dort ein Bottich ein-gelassen, um das Blut der geschlachteten Tiere aufzunehmen oderum die Fleischstücke in einer Salzlake zu pökeln. Es fiel auch eingrosser rechteckiger Stein auf (s. Kap. 10.3.12), der mit seinen Ril-len an die Schlachtbänke von Augusta Raurica erinnert2036.Demnach könnte es sich bei Gebäude Q um ein Schlachthausund eine Produktionsstätte von Fleischkonserven handeln.
Die Idee, Frischfleisch zu konservieren und für Zeiten undOrte, in denen es nicht vorhanden war, verfügbar zu machen,dürfte uralt sein. Für die vorindustriellen Epochen kommen dieKühllagerung in Eis und Schnee, das Einsalzen, Räuchern undTrocknen in Frage, wobei die drei letzteren Methoden den Vor-teil hatten, die Produkte über weite Distanzen verhandeln zukönnen. In Italien scheint am häufigsten Schweinefleisch ver-arbeitet worden zu sein; daneben wurde aber auch Schaf-, Zie-gen, Rind- und Hirschfleisch konserviert2037. Auch über die ver-schiedenen Verarbeitungsmethoden geben diverse schriftlicheQuellen Auskunft, wobei in der Regel das Fleisch einige Zeit inSalz eingelegt und dann mehrere Tage in den Rauch gehängtwurde2038. Seit dem 1. Jahrhundert v.Chr. ist der Export vonSchinken und sonstigen Räucherwaren aus verschiedenen Re-gionen Galliens nach Italien belegt2039.
In Augusta Raurica liessen sich aufgrundvon Durchlochungen an Schulterblättern, diebeim Durchstossen mit einem Haken ent-standen sind, und Konzentrationen von Schul-terblättern und Unterkiefern Speiseabfällevon Räucherwaren nachweisen2040. Diese Abfäl-le finden sich aber nicht direkt im Bereich derÖfen, da die Knochen erst bei der Konsumie-rung des Rauchfleisches anfielen. Schulter-blätter mit Durchlochungen fanden sich auchin Rottweil und in Xanten, wo sich ebenfallsauffallend viele Rinderrippen fanden, die aufeine durchschnittliche Länge von 166 mmzugehauen sind und auf der Innenseite paral-lele Schnittspuren aufweisen2041. Im Zusam-menhang mit den durchlochten Schulterblät-tern werden sie als die Überreste von
zusammenhängenden Brustkorbteilen gedeutet, die man kon-serviert hat. Eine Ansammlung vergleichbarer Rinderrippen, dieim Durchschnitt etwas kürzer sind als jene in Xanten, kam ineiner Kanalverfüllung der Augster Frauenthermen zum Vor-schein, wo sie als Überreste einer Taberne gedeutet werden, inder Fleischeintöpfe gekocht wurden2042. Aufgrund des XantenerBefundes dürfen wir vermuten, dass die Rippen der AugsterKanalverfüllung ebenfalls von konserviertem Brustfleisch stam-men. Auch die Biberister Rinderrumpfteile in Gebäude D undder Grube (A125/B44) in Gebäude Q dürften ähnlich zu deu-ten sein.
Bei der Anlage in Gebäude Q muss es sich aufgrund der Kon-struktion um einen Ofen handeln, der die Hitze gleichmässigverteilte und nur langsam an das Gut abgab. Botanische Reste,die auf einen Darrofen schliessen lassen, kamen in den Schlämm-proben der Ofenkanäle keine zum Vorschein (Kap. 28.5.8). Hin-gegen liegen formlose Brocken vor, die Fett von Schafen oderSchweinen enthalten (Kap. 30.6). Der Ofen in Gebäude Q könn-te demnach zum Konservieren von Hirsch-, Schweine- undRindfleisch gedient haben. Es bleibt zu fragen, ob in BiberistSelbstversorgung vorliegt oder, wie in Neftenbach, eine Über-schussproduktion2043. Die Grösse des Ofens spricht wohl für letz-teres.
Kap. 30.2
2036 Schwarz 1995, 40-42.2037 André 1998, 123.2038 Zitate wie Anm. 2037 mit Anm. 112; s. auch Kap. 18.3.1 mit Anm. 654.2039 Wie Anm. 2037; s. auch Kap. 18.3.1 mit Anm. 657.2040 Schmid 1969, 106; zur Konzentrationen von Schulterblättern und Unter-kiefern: Schibler/Schmid 1989, 25-26.; Furger/Deschler-Erb 1992, 392-393.2041 Kokabi 1982, 44 (Rottweil); Berke 1995, 356-360; 361 (Xanten).2042 Deschler-Erb 1991a, 146-151.2043 Wie Anm. 2013, 455-457.
Abb. 30/31. Gebäude Q. Anteil (g%) der verschiedenen Skelettteile bei Schwein.

30.2.12. Tor und Umgebung
Graben Fast 1100 Knochen fanden sich im Graben entlang der Hof-
mauer, der vor deren Bau zu Beginn des 2. Jahrhunderts dasGutshofareal begrenzte und dann als gedeckter Drainagegrabendiente (Kap. 10.1.1): 597 Knochen gehören zum Bauhorizont deswohl im zweiten Viertel des 2. Jahrhunderts umgebauten Tores,500 Knochen stammen aus der oberen Einfüllung des Grabens,in die die Abdeckung später einstürzte. Trotz der grossen Fund-menge lassen sich wegen der geringen Bestimmbarkeit (Tab.30/1) nur beschränkt Aussagen zur Zusammensetzung machen.Die geringe Bestimmbarkeit hängt in erster Linie mit dem gerin-gen Durchschnittsgewicht zusammen; die Keramikscherben fal-len ebenfalls durch ihre Kleinteiligkeit auf (s. Kap. 10.1.1). ImGraben wurden die gleichen Tierarten wie im Innern der Vil-lenanlage gefunden. Auffällig sind hingegen die hohen Anteile(10%) von Equiden- und Hundeknochen (Tab. 30/3). In denSchichten im Innern der Villenanlage sind sie jeweils nur mitwenigen Knochenfragmenten vertreten, kommen aber ausserhalbder Gebäude häufiger vor als innerhalb. Dies spricht dafür, dassbeide Tierarten kaum gegessen wurden und es sich bei den Ein-zelknochen um verstreute Kadaverteile handelt. Der erhöhteAnteil im Graben ausserhalb des umfriedeten Areals dürftedadurch entstanden sein, dass störender Abfall, vielleicht sogarTeilskelette, dort entsorgt wurden. Der relativ geringe Anteil anVerbissspuren (Tab. 30/1) ist damit zu erklären, dass der Grabenzugedeckt war (Kap. 10.1.1), so dass streunende Hoftiere oderWildtiere aus der Umgebung nicht an die Knochen herankamen.
Hundekadaver Neben der Zufahrtstrasse zum Tor fanden sich Überreste von
mindestens zwei Hundeskeletten. Das Skelett des ersten Hun-des (A119/B082.30) ist zu grossen Teilen erhalten. Im Sehnen-verband und mit unverrundeten Bruchkanten lag es in situ(Abb. 30/32). Es handelt sich um einen mittelgrossen Rüden(Kap. 30.3.7), der zwischen eineinhalb und zwei Jahre altwurde, wie sich anhand der Methode von M.-P. Horard-Herbin,die den Abkauungsgrad des ersten Unterkiefermolares berück-sichtigt, ermitteln lässt2044. Er war somit älter als die zur Fleisch-
produktion genutzten spätlatènezeitlichen Tiere aus Levrouz,aber jünger als die meisten römerzeitlichen Tiere der gleichenFundstelle2045. Er scheint frühzeitig durch eine Krankheit, einenUnfall, bei einem Überfall oder sogar durch Gift ums Lebengekommen zu sein. Da er nahe beim Hoftor wohl kurz nach sei-nem Tod verscharrt worden war, könnte es sich um den Hof-hund handeln, der zu Lebzeiten, vielleicht an einer Kette ange-bunden, das gegen die Strasse Salodurum-Brenodurum/Solo-thurn-Bern gerichtete Hoftor bewachte. Hatte er auch noch nachseinem Tod seine Aufgabe als Wachhund zu erfüllen?
Dem zweiten Hund lassen sich lediglich neun Knochenzuweisen (Tab. 30/3). Das Skelett dürfte verlagert sein. Da keineZähne vorhanden sind, lässt sich sein Alter nicht genauer alsadult bestimmen.
Gruben Die drei bei der Verwüstung der grossen Villa (Periode II-
Phase 7) kurz nach der Mitte des 3. Jahrhunderts noch offenenGruben im Bereich des Tores haben nur wenig Tierknochengeliefert (Tab. 30/1), die wahrscheinlich eher zufällig in diesegelangt sind. In der Grube (A118/B072) an der Innenseite desHofmauer, nach Ausweis der Schlämmfunde eine Latrine (Kap.30.5.3), fällt der hohe Hühnerknochenanteil auf (Tab. 30/3), wasvermuten lässt, dass es sich um Speiseabfälle handelt, die im 5 msüdlich gelegenen Gebäude O angefallen waren, wo ebenfallsrelativ viele Vogelknochen festgestellt werden können (Kap.30.2.9). In der Grube (A116/B075) (Tab. 30/2) innerhalb dersüdlichen Torwange fanden sich hingegen eher Rinder- undSchweineknochen. Der Anteil verbrannter Knochen ist mit 14%relativ hoch (Tab. 30/1). Ein Viertel des Materials weist ver-rundete Bruchkanten auf, auch der Anteil der Knochen mit Ver-bissspuren ist relativ hoch. Die in die Gruben gelangten Kno-chen dürften vorher schon lange herumgelegen sein. Die Grube(A118/B075) ausserhalb der südlichen Torwange hat zu wenigKnochen geliefert, als dass sie ausgewertet werden könnte.
V. Funde und besondere Fundensembles römischer Zeit – 30. Die Tierknochen
2044 Horard-Herbin 2000, 117 fig. 2. Der erste Unterkiefermolar des Hoftor-hundes entspricht dem Abkauungsgrad ‚stage Da‘.2045 Wie Anm. 2044, 120 fig. 5.
654
Abb. 30/32. Hundeskelett (A119/B082.30) vor dem Tor. Aufnahmegeben Süden.
Abb. 30/33. Grube (A002/B152) mit Kadaver eines Rindes südlich dernördlichen Trennmauer. Augnahme gegen Westen.

655
30.2.13. Rinderkadaver in der Kernzone
Grube (A002/B153)Unmittelbar südlich der Portikus F fand sich eine längliche,
nordsüdlich gerichtete Grube (A002/B153), die das nahezu voll-ständige Skelett eines Rindes enthielt (Abb. 30/33). Bereits beider Grabung wurden die Knochen als „teilweise stark brüchigund halb zerfallen„ beschrieben. Nach der zeichnerischen undfotografischen Aufnahme des Skelettes wurden die nur grobgereinigten Knochen in grosse Abfallsäcke oder Alufolie einge-packt, mehr oder weniger nach anatomischen Einheiten geord-net. Bei der zehn Jahre später stattfindenden archäozoologischenBearbeitung haben wir leider feststellen müssen, dass die an denKnochen haftende Erde steinhart ausgetrocknet ist und kaummehr entfernt werden kann. Die Knochen sind noch brüchigerals bei der Grabung beobachtet: Nahezu alle Plattenknochensind bis zur Unkenntlichkeit zerfallen. Der Schädel und dieUnterkiefer werden nur durch die eingetrocknete Erde zusam-mengehalten und präsentieren sich als unförmige Erdklumpen.Die Röhrenknochen sind vergleichsweise gut erhalten. Allerdingssplittert die Oberfläche an vielen Stellen auf. Um einigermassenden Status quo zu erhalten und eine minimale osteologischeBearbeitung zu ermöglichen, habe ich die Röhrenknochenund den Kopf mit einem härtenden Präparat (Osteo-Fix) be-handelt. Zu Beginn der osteologischen Bearbeitung hat uns derextrem schlechte Zustand dieser Kadaverknochen an eine spe-zielle Behandlung, etwa das Zudecken mit ungelöschtem Kalk,denken lassen, was aber nicht überprüfbar ist. Auf die üblicheEinzelaufnahme der Knochenfragmente muss aus besagtenGründen verzichtet werden. Desgleichen kann das Knochen-gewicht nicht berücksichtigt werden, da die anhaftende Erde dasFundgewicht massiv erhöht. Wegen des schlechten Zustandesdes Skelettes muss für die Beantwortung vieler Fragen auf dieZeichnung und deren Beschrieb zurückgegriffen werden, dieimmerhin als überdurchschnittlich ausführlich bezeichnet wer-den darf2046. Die einzelnen Skeletteile befanden sich mehr oderweniger in anatomisch richtiger Lage zueinander. Dies bedeu-tet, dass das Tier noch im Sehnenverband in die Grube gelegtwurde. Das Rind lag auf seiner rechten Seite mit dem Kopf nachSüden; die Beine waren somit nach Osten gerichtet. Die Zeich-
nerin beobachtete eine „unnatürliche Stellung der Beine“. DieKnochen des linken Hinterbeines bildeten an einer Stelle einenrechten Winkel. Von den anderen Beinen seien nicht mehr alleKnochenpartien erhalten. Wahrscheinlich seien der Kuh die Fes-seln gewaltsam gebrochen worden. Auf den Fotos lassen sich beiden Metapodienmitten allerdings gut die sehr hellen neuenBruchkanten ausmachen, die daraufhindeuten, dass die unna-türliche Beinstellung erst durch eine Verschiebung während derGrabung zustandekam. Allerdings fehlen den Beinen so gut wiealle Zehen, beim rechten Hinterteil auch der distale Teil desMetatarsus (Abb. 30/35). Dies könnte ein Hinweis darauf sein,dass dem Rind die Haut abgezogen worden war, in der dieZehenknochen noch steckten; dies ist wiederholt bei Überres-ten von Gerbereien beobachtet worden2047. In Zusammenhangmit dem Abziehen der Haut könnte auch der Zustand des Kop-fes zu sehen sein: Während Ober- und Unterkiefer annäherndvollständig vorzuliegen scheinen, fehlen wesentliche Teile derSchädeldecke, insbesondere die Hornzapfen, wie auch dasHinterhaupt. Weiter muss der Kopf vom Rumpf abgetrennt wor-den sein, denn man hatte ihn zwar an der anatomisch richtigenStelle deponiert, aber um 180° gedreht, sodass die Schädeldeckegegen den Körper hin zu liegen kam. Schnittspuren lassen sichaufgrund des schlechten Zustandes nicht mehr beobachten. Obdie Abtrennung des Kopfes in Zusammenhang mit der Ent-häutung zu sehen ist, muss offen bleiben.
Ausser der Haut und eventuell dem Horn wurde kein Teildes Rindes weiterverwendet. Denn das Erdmaterial zwischenund um die Knochen unterschied sich durch seine dunklereFarbe und einen grösseren organischen Anteil vom übrigen Sedi-ment. Diese Zone entspricht ungefähr dem Fleischvolumen desRindes. Daher ist damit zu rechnen, dass das Rind eines natür-lichen Todes verstorben ist. Altersschwäche als Todesursachedürfte weitgehend auszuschliessen sein, denn aufgrund des deut-lichen, aber nicht übermässig starken Abkauungsgrad des drit-ten Molaren und der nicht verwachsenen Beckensymphyse istdas Sterbealter auf etwa sechs bis sieben Jahre zu schätzen. Obdas aufgrund der Pubisbildung als weiblich zu bestimmende Tierim Zusammenhang mit einer Geburt starb, muss offen bleiben.Ein Fötus wurde nicht angetroffen. Als Todesursache kommt amehesten eine schwere, möglicherweise ansteckende Krankheit inFrage, die die Gutshofbewohner dazu bewog, zwar die Haut,nicht aber das Fleisch des Tieres zu nutzen.
Da der Kadaver mangels Funde archäologisch nicht zu datie-ren ist, haben wir den linken Radius einer Radiokarbondatie-rung unterzogen (s. Kap. 14.20 Anm. 148). Die Analyse ergab einAlter von 381 n.Chr. ± 82, womit der Kadaver zum Gehöft desspäten 3. Jahrhunderts (Periode III) gehören dürfte oder allen-falls zur letzten Phase der Villa (Periode II-Phase 7), die ins mitt-lere 3. Jahrhundert datiert. Zu letzterer gehört das gut 10 m naheGebäude G, das als Stall für Reit- oder Zugtiere (Pflugrinder!)zu interpretieren ist (s. Kap. 17.4.7; 17.5; s. auch Kap. 10.2.2).
Kap. 30.2
2046 Zeichnung und Beschrieb führte Christine Stierli-Blapp durch.2047 Schmid 1969, 105-106.
Abb. 30/34. Grube (A017/B150) mit Resten eines Rinderkadavers.Aufnahme gegen Westen.

geln, kommt die Bedeutung seltenerer und oft-mals kleinerer Tiere wie Geflügel nur bei denKnochenzahlen zum Ausdruck (z. B. Abb.30/15 und 30/19). Trotzdem ergeben sich beider Auswertung sowohl nach Fragmentzahlenwie nach Gewichtswerten im grossen undganzen vergleichbare Tendenzen.
Die Nutzung einer Tierart lässt sich an-hand der Schlachtalterraten, des Skeletteil-spektrums und des Geschlechterverhältnissesfeststellen. Hohe Anteile jugendlicher Tieresprechen in der Regel für die ausschliesslicheNutzung als Fleischtiere, erhöhte Anteile vonalten Individuen hingegen für die Nutzung inerster Linie als Arbeitskraft (Zug-, Last- oderReittiere) oder die Gewinnung von sekundä-ren Produkten (Milch, Wolle) (s. Abb. 30/21;30/22). Zur Weiterzucht sind für einen selbst-versorgenden Gutsbetrieb nur wenige Mut-tertiere notwendig. Die Zucht von Jungtierenzum Verkauf im nächstgelegenen Marktort,etwa im nahen Vicus Salodurum, dürfte sichebenfalls in einer Dominanz adulter Tiere im
Tierknochenmaterial niederschlagen. Trotzdem dürfen wirnicht vergessen, dass Tierhaltung, die zu einem einzigen Zweckbetrieben wird, eine Erfindung unserer Zeit und unserer west-lichen Welt ist. Während des längsten Teils der Menschheitsge-schichte – und heute noch in vielen Ländern – wurde und wirdjedes noch so alte Tier schliesslich geschlachtet und sein Fleischgegessen, es sei denn, es handelte sich um kranke Tiere, wie z.B.der Rinderkadaver in einer Grube in der Kernzone (Kap. 30.2.13)oder Tiere, deren Fleisch tabu war, wie z.B. der Wachhund beimHoftor (Kap. 30.2.12).
Markante Abweichungen der Skeletteilhäufigkeiten im Ver-gleich zu einem vollständigen Skelett (s. Abb. 30/14; 30/16; 30/24;30/31; 30/30) lassen auf eine bestimmte Nutzung des Tieres nachseinem Tod schliessen. So spricht die Häufung von Schädel- undFussteilen in einem Ensemble für Gerberei und/oder Hornma-nufaktur. Neben diesen Spezialfällen, die hauptsächlich instädtischen Siedlungen mit ihren Gewerbequartierena nzutref-fen sind, ist es auf den Gutshöfen eher die Regel, dass mit denKnochenabfällen Schlacht- und Speiseabfälle unterschieden wer-den können, je nachdem wie gross der Anteil der viel Fleisch tra-genden Skeletteile ist.
Auch das Geschlechterverhältnis kann Informationen zurhauptsächlichen Nutzung einer Tierart liefern.So sprechen z.B.vieleältere Sauen für eine intensive Nachzucht an Ort, viele subadulteEber hingegen für einen häufigen Verzehr von Schweinefleisch.
Im Falle von Biberist ist zu betonen, dass für die meistenTierarten und Fragestellungen höchstens Phase 7 der Villa (Peri-ode II) eine genügend grosse Datenbasis für sichere Aussagengeliefert hat. Gerade für die Frage nach dem Geschlechterver-hältnis liegen meistens zu wenige Daten vor. Aufgrund deshohen Fragmentierungsgrades und der schlechten Erhaltungliessen sich auch nur wenige Masse nehmen. Es ist folglich nurbei wenigen Tierarten eine metrische Auswertung möglich,deren Aussagemöglichkeiten überdies angesichts der geringenstatistischen Basis hypothetisch bleibt.
V. Funde und besondere Fundensembles römischer Zeit – 30. Die Tierknochen656
Abb. 30/35. Grube (A002/B153). Vorhandene Knochen des Rinderkadavers.
Grube A 017/B150Reste eines möglichen weiteren Kadavers kamen in einer
rechteckigen Grube (A017/B150) 4 m südlich des mutmass-lichen Stalls G zum Vorschein (Abb. 30/34), die später von Bach-läufen stark beeinträchtigt wurde (Kap. 10.2.5; s. auch Abb. 4/1).Datierende Funde fehlen.
Die Tierknochen sind in einem extrem schlechten Zustand.Es scheint sich um die Überreste eines Rindes zu handeln. EinFemur und ein Humerus lassen sich identifizieren, allerdings nurdie Diaphyse (Röhre), die Gelenkenden fehlen. Weitere Frag-menten könnten vom relativ dickwandigen Hinterhaupt stam-men. Ein anderes Fragment dürfte einem Wirbel zuzuordnen sein.Alles in allem handelt es sich um Fragmente mit relativ dickerKompakta, was ihre Erhaltung sicher begünstigt hat. Dies bedeu-tet, dass andere Knochenteile, die sich schlechter erhalten (z.B.Plattenknochen), durchaus ursprünglich vorhanden gewesen seinkönnen. Andererseits ist zu bemerken, dass Zähne, die sich in derRegel auch im schlechten Milieu relativ gut erhalten, fehlen.
30.3. Die Tierarten im Wandel der Zeit
30.3.1. Einleitung
In diesem Kapitel werden die Tierarten bezüglich ihresAnteils, des Skeletteilspektrums, der Schlachtalterrate, des Ge-schlechterverhältnisses und, wenn möglich, ihrer osteometri-schen Resultate während den einzelnen Siedlungsphasen derVilla (Periode II) diskutiert. Das Vorgängergehöft (Periode I)und der nachfolgende Hof (Periode III) müssen mangels aus-sagekräftiger Ensembles beiseite gelassen werden (Tab. 30/1). Daverschiedene Ensembles über mehrere Siedlungsphasen reichen(s. Abb. 17/1), haben wir z.T. zwei Phasen zusammengefasst,Phasen 1/2, 3/4 und 5/6, die grössere und damit aussagekräfti-gere Ensembles ergeben.
Während die Gewichtsanteile die Bedeutung einer Tierartfür die Ernährung bzw. Nahrungsmittelproduktion widerspie-

657
30.3.2. Hausrind
In Bezug auf die Gewichtsanteile ist dasHausrind in allen Phasen der Periode II mitAnteilen zwischen 60-70% die wichtigste Tier-art (Abb. 30/36). Dies lässt sich immer wiederin römischem Zusammenhang feststellen, dadieses Tier mit Abstand das grösste der dreiwichtigsten gehaltenen Haustierarten – Haus-rind, Schaf/Ziege, Hausschwein – ist. Als ein-zige Veränderung im Laufe der Zeit ist ledig-lich ein Rückgang um 10% von den Phasen 5/6zu Phase 7 zu nennen.
Nach Fragmentanteilen ist das Rind injeder Phase hingegen nur das zweitwichtigsteTier. Zwischen den Phasen 1/2 und 3/4 ist einleichter Anstieg der Hausrindknochen um8% zu konstatieren. In den Phasen 3/4, 5/6und 7 schwankt der Rinderanteil nur nochzwischen 36% und 38%.
NutzungIn allen Phasen ist der Anteil der adulten
Rinder klar dominant (Abb. 30/37). Somitdürfte die primäre Nutzung als Arbeitstier imVordergrund gestanden haben. (s. auch Kap.18.2.4). Ob auch die Milch genutzt wurde, lässtsich aufgrund der vorliegenden Daten, vorallem mangels geschlechtsbestimmter Kno-chen, nicht beurteilen. Aufgrund schriftlicherQuellen ist bekannt, dass in der römischenKultur Kuhmilch weniger oft Verwendungfand als Ziegenmilch2048. Ob dies allerdingsauch in den nordwestlichen Provinzen der Fallwar, hat die Forschung bis heute nicht klärenkönnen.
In Phase 7 fällt im Vergleich zu den vor-angegangenen Phasen eine Zunahme des Jung-tieranteils auf. Fast ein Viertel der altersbe-stimmten Rinderknochen entfällt nun auf dieGruppe der infantilen-juvenilen Tiere, d.h. derKälber bis zu einem Alter von etwa 6 Monaten.Der Anteil der juvenilen-subadulten Tierebeträgt immerhin noch 10%, und auch neo-nate-infantile Individuen sind belegt. Dieslässt darauf schliessen, dass in dieser letztenPhase der Villa (Periode II) das Rind als Flei-schlieferant an Bedeutung zugenommen hat.
Diese Veränderung in Phase 7 lässt sichauch beim Skeletteilspektrum feststellen (Abb. 30/38): Währenddie Rumpfteile, wie meistens in archäologischem Zusammen-hang zu beobachten, in den Phasen 1 bis 6 gegenüber einemrezenten Vergleichsskelett deutlich untervertreten sind, verbes-sert sich ihr Anteil in der letzten Phase um mehr als 10%. Dieals Schlachtabfälle zu interpretierenden Kopfteile und dasZygopodium sind nun im Gegensatz zu den vorangegangenenPhasen untervertreten. Das relativ gut belegte Autopodium mussnicht auf Schlachtabfall weisen, sondern könnte auch als Koch-
abfall gedeutet werden, z.B. von Rinderfüssen als Beilage in Ein-töpfen. Halbfabrikate und Abfallstücke belegen überdies einelokale Herstellung von Beinartefakten (s. Kap. 27.5). Die stär-kere Bedeutung des Rindes als Fleischlieferant in Phase 7könnte mit dem neu eingerichteten Schlachthaus mit Konser-vierungsofen in Gebäude Q in Zusammenhang stehen.
Kap. 30.3
2048 André 1998, 130-131; s. auch Kap. 18.3.1 mit Anm. 661.663-664.
Abb. 30/36. Entwicklung des Tierartenspektrums (n% und g%) in der grossen Villa (Periode II).
Abb. 30/37. Entwicklung der Alterstruktur (n%) bei Rind in der grossen Villa (Periode II).
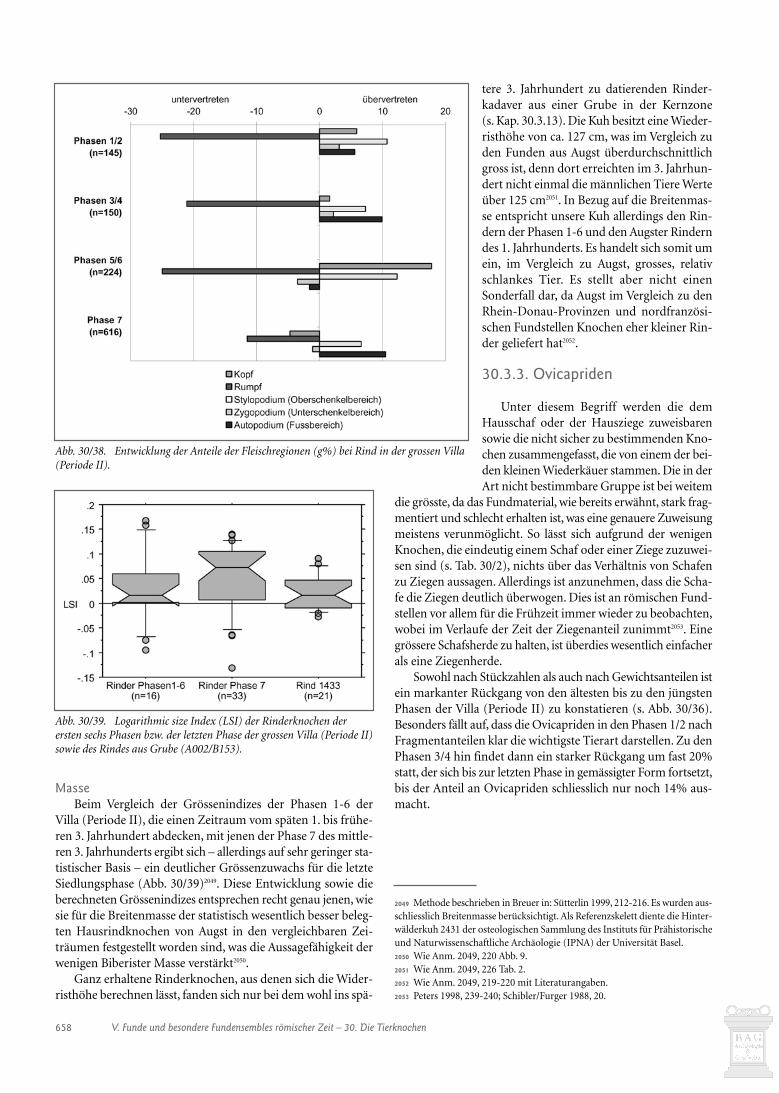
MasseBeim Vergleich der Grössenindizes der Phasen 1-6 der
Villa (Periode II), die einen Zeitraum vom späten 1. bis frühe-ren 3. Jahrhundert abdecken, mit jenen der Phase 7 des mittle-ren 3. Jahrhunderts ergibt sich – allerdings auf sehr geringer sta-tistischer Basis – ein deutlicher Grössenzuwachs für die letzteSiedlungsphase (Abb. 30/39)2049. Diese Entwicklung sowie dieberechneten Grössenindizes entsprechen recht genau jenen, wiesie für die Breitenmasse der statistisch wesentlich besser beleg-ten Hausrindknochen von Augst in den vergleichbaren Zei-träumen festgestellt worden sind, was die Aussagefähigkeit derwenigen Biberister Masse verstärkt2050.
Ganz erhaltene Rinderknochen, aus denen sich die Wider-risthöhe berechnen lässt, fanden sich nur bei dem wohl ins spä-
tere 3. Jahrhundert zu datierenden Rinder-kadaver aus einer Grube in der Kernzone(s. Kap. 30.3.13). Die Kuh besitzt eine Wieder-risthöhe von ca. 127 cm, was im Vergleich zuden Funden aus Augst überdurchschnittlichgross ist, denn dort erreichten im 3. Jahrhun-dert nicht einmal die männlichen Tiere Werteüber 125 cm2051. In Bezug auf die Breitenmas-se entspricht unsere Kuh allerdings den Rin-dern der Phasen 1-6 und den Augster Rinderndes 1. Jahrhunderts. Es handelt sich somit umein, im Vergleich zu Augst, grosses, relativschlankes Tier. Es stellt aber nicht einenSonderfall dar, da Augst im Vergleich zu denRhein-Donau-Provinzen und nordfranzösi-schen Fundstellen Knochen eher kleiner Rin-der geliefert hat2052.
30.3.3. Ovicapriden
Unter diesem Begriff werden die demHausschaf oder der Hausziege zuweisbarensowie die nicht sicher zu bestimmenden Kno-chen zusammengefasst, die von einem der bei-den kleinen Wiederkäuer stammen. Die in derArt nicht bestimmbare Gruppe ist bei weitem
die grösste, da das Fundmaterial, wie bereits erwähnt, stark frag-mentiert und schlecht erhalten ist, was eine genauere Zuweisungmeistens verunmöglicht. So lässt sich aufgrund der wenigenKnochen, die eindeutig einem Schaf oder einer Ziege zuzuwei-sen sind (s. Tab. 30/2), nichts über das Verhältnis von Schafenzu Ziegen aussagen. Allerdings ist anzunehmen, dass die Scha-fe die Ziegen deutlich überwogen. Dies ist an römischen Fund-stellen vor allem für die Frühzeit immer wieder zu beobachten,wobei im Verlaufe der Zeit der Ziegenanteil zunimmt2053. Einegrössere Schafsherde zu halten, ist überdies wesentlich einfacherals eine Ziegenherde.
Sowohl nach Stückzahlen als auch nach Gewichtsanteilen istein markanter Rückgang von den ältesten bis zu den jüngstenPhasen der Villa (Periode II) zu konstatieren (s. Abb. 30/36).Besonders fällt auf, dass die Ovicapriden in den Phasen 1/2 nachFragmentanteilen klar die wichtigste Tierart darstellen. Zu denPhasen 3/4 hin findet dann ein starker Rückgang um fast 20%statt, der sich bis zur letzten Phase in gemässigter Form fortsetzt,bis der Anteil an Ovicapriden schliesslich nur noch 14% aus-macht.
V. Funde und besondere Fundensembles römischer Zeit – 30. Die Tierknochen
2049 Methode beschrieben in Breuer in: Sütterlin 1999, 212-216. Es wurden aus-schliesslich Breitenmasse berücksichtigt. Als Referenzskelett diente die Hinter-wälderkuh 2431 der osteologischen Sammlung des Instituts für Prähistorischeund Naturwissenschaftliche Archäologie (IPNA) der Universität Basel.2050 Wie Anm. 2049, 220 Abb. 9.2051 Wie Anm. 2049, 226 Tab. 2.2052 Wie Anm. 2049, 219-220 mit Literaturangaben.2053 Peters 1998, 239-240; Schibler/Furger 1988, 20.
658
Abb. 30/38. Entwicklung der Anteile der Fleischregionen (g%) bei Rind in der grossen Villa (Periode II).
Abb. 30/39. Logarithmic size Index (LSI) der Rinderknochen derersten sechs Phasen bzw. der letzten Phase der grossen Villa (Periode II)sowie des Rindes aus Grube (A002/B153).

659
NutzungDa für die Phasen 3/4 nur wenige Alters-
bestimmungen vorliegen, lässt sich nicht mitSicherheit feststellen ob der beim Tierarten-spektrum festgestellte abrupte Wechsel zwi-schen den Phasen 1/2 und den folgendenPhasen auch gleichzeitig beim Altersspek-trum stattfand (Abb. 30/40). Vergleicht mandie Altersstruktur in den Phasen 1/2 mit jenerin den Phasen 5/6, so ist ein Wechsel vonhauptsächlich jung- und alt-adulten Indivi-duen zu einer Dominanz der infantil-juveni-len festzustellen. Dies lässt auf eine veränder-te Nutzung schliessen: Zuerst stand die Nut-zung der sekundären Produkte im Vorder-grund. In Frage kommen sowohl Milch wieWolle. In Italien scheint hauptsächlich Zie-genmilch zur Herstellung von Käse verwendetworden zu sein, zum Teil direkt auch auf denHöfen selbst2054. Welche Produktion in Biberistdie wichtigere war, lässt sich aufgrund derkaum unterscheidbaren Schaf/Ziegenknochennicht sagen; die in dieser Zeit noch betriebe-ne Wasch- und Walkanlage in der Werkstatt B-Phase A (Kap. 9.4) dürfte indes für eine bedeu-tende Wollproduktion sprechen. Spätestens abden Phasen 5/6 gewann die Fleischnutzungstark an Bedeutung. In Phase 7 nehmen dieAnteile der jung-und hauptsächlich der alt-adulten Tiere wieder zu, wobei die infantil-juvenilen einen Anteil von 20% ausmachen.Dies deutet auf eine vermehrte Nutzung derTiere auch als Milch- und Wolllieferanten hin.
Die Untersuchung der Fleischregionenlässt nur schwache Unterschiede zwischenden einzelnen Phasen erkennen (Abb. 30/41):Während in den Phasen 1 bis 6 das Zygopo-dium jeweils über 20% über der Normalver-tretung liegt, was eher für Schlachtabfällespricht, ist sein Anteil in der letzten Pha-se zugunsten des Stylopodiums zurückgegan-gen, was für vermehrte Speiseabfälle spricht.
MasseGerade für die Frage nach der Nutzung
sekundärer Produkte wie Wolle wären metri-sche Untersuchungen zu den Ovicapridensehr hilfreich. Leider können bei den sehrfragmentierten und nur in Einzelfällen art-bestimmten Knochen nur wenige Masse ab-genommen werden, anhand derer sich keinerelevanten Aussagen machen lassen.
Anhand zweier Schafsmetacarpen aus Phase 7 lassen sichWiderristhöhen von 54,9 cm bzw. 54,8 cm errechnen2055. Im Ver-gleich zu den gleichzeitigen Augster Werten sind diese Masse alssehr gering zu bezeichnen2056. Sie dürften ein weiteres Zeichendafür sein, dass die kleinen Wiederkäuer in der Spätzeit kaumeine Rolle für die Villenwirtschaft spielten.
Kap. 30.3
2054 André 1998, 130-131; s. auch Kap. 18.3.1 mit Anm. 664.2055 Von den Driesch/Boessneck 1974, 339 Tab.6.2056 Breuer u.a. 2001, 166 Abb. 5.
Abb. 30/40. Entwicklung der Alterstruktur bei Schaf/Ziege in der grossen Villa (Periode II).
Abb. 30/41. Entwicklung der Anteile der Fleischregionen (g%) bei Schaf/Ziege in der gros-sen Villa (Periode II).
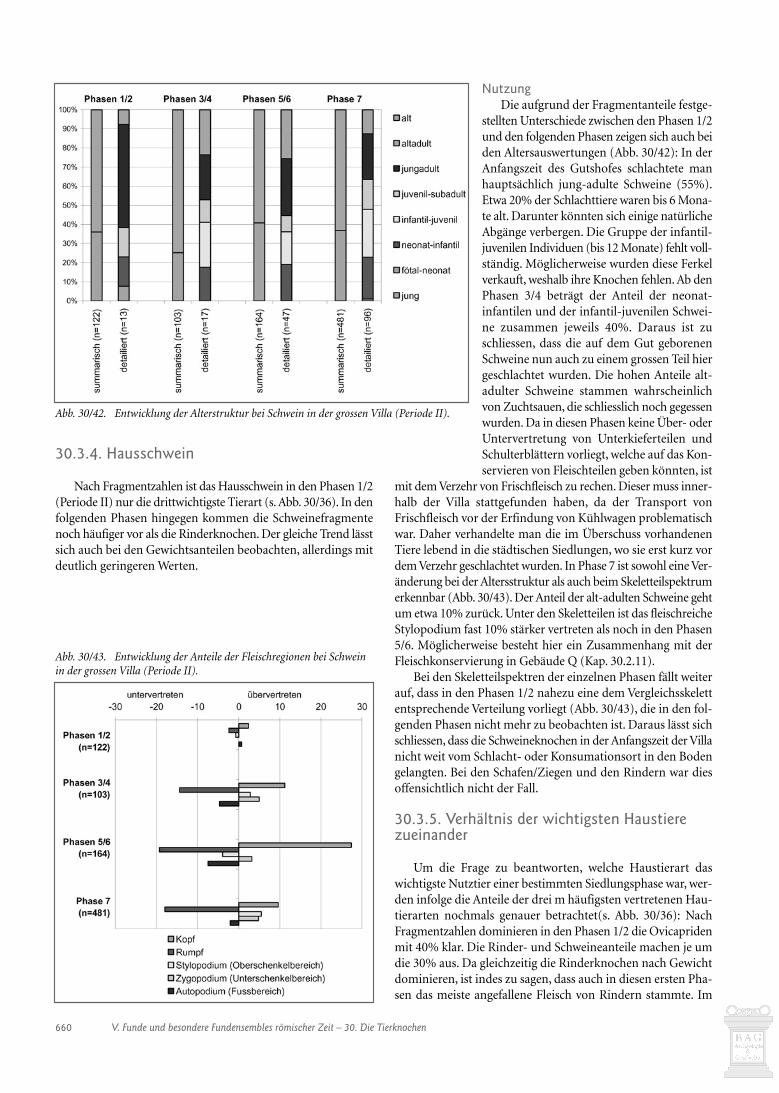
NutzungDie aufgrund der Fragmentanteile festge-
stellten Unterschiede zwischen den Phasen 1/2und den folgenden Phasen zeigen sich auch beiden Altersauswertungen (Abb. 30/42): In derAnfangszeit des Gutshofes schlachtete manhauptsächlich jung-adulte Schweine (55%).Etwa 20% der Schlachttiere waren bis 6 Mona-te alt. Darunter könnten sich einige natürlicheAbgänge verbergen. Die Gruppe der infantil-juvenilen Individuen (bis 12 Monate) fehlt voll-ständig. Möglicherweise wurden diese Ferkelverkauft, weshalb ihre Knochen fehlen. Ab denPhasen 3/4 beträgt der Anteil der neonat-infantilen und der infantil-juvenilen Schwei-ne zusammen jeweils 40%. Daraus ist zuschliessen, dass die auf dem Gut geborenenSchweine nun auch zu einem grossen Teil hiergeschlachtet wurden. Die hohen Anteile alt-adulter Schweine stammen wahrscheinlichvon Zuchtsauen, die schliesslich noch gegessenwurden. Da in diesen Phasen keine Über- oderUntervertretung von Unterkieferteilen undSchulterblättern vorliegt, welche auf das Kon-servieren von Fleischteilen geben könnten, ist
mit dem Verzehr von Frischfleisch zu rechen. Dieser muss inner-halb der Villa stattgefunden haben, da der Transport vonFrischfleisch vor der Erfindung von Kühlwagen problematischwar. Daher verhandelte man die im Überschuss vorhandenenTiere lebend in die städtischen Siedlungen, wo sie erst kurz vordem Verzehr geschlachtet wurden. In Phase 7 ist sowohl eine Ver-änderung bei der Altersstruktur als auch beim Skeletteilspektrumerkennbar (Abb. 30/43). Der Anteil der alt-adulten Schweine gehtum etwa 10% zurück. Unter den Skeletteilen ist das fleischreicheStylopodium fast 10% stärker vertreten als noch in den Phasen5/6. Möglicherweise besteht hier ein Zusammenhang mit derFleischkonservierung in Gebäude Q (Kap. 30.2.11).
Bei den Skeletteilspektren der einzelnen Phasen fällt weiterauf, dass in den Phasen 1/2 nahezu eine dem Vergleichsskelettentsprechende Verteilung vorliegt (Abb. 30/43), die in den fol-genden Phasen nicht mehr zu beobachten ist. Daraus lässt sichschliessen, dass die Schweineknochen in der Anfangszeit der Villanicht weit vom Schlacht- oder Konsumationsort in den Bodengelangten. Bei den Schafen/Ziegen und den Rindern war diesoffensichtlich nicht der Fall.
30.3.5. Verhältnis der wichtigsten Haustierezueinander
Um die Frage zu beantworten, welche Haustierart daswichtigste Nutztier einer bestimmten Siedlungsphase war, wer-den infolge die Anteile der drei m häufigsten vertretenen Hau-tierarten nochmals genauer betrachtet(s. Abb. 30/36): NachFragmentzahlen dominieren in den Phasen 1/2 die Ovicapridenmit 40% klar. Die Rinder- und Schweineanteile machen je umdie 30% aus. Da gleichzeitig die Rinderknochen nach Gewichtdominieren, ist indes zu sagen, dass auch in diesen ersten Pha-sen das meiste angefallene Fleisch von Rindern stammte. Im
V. Funde und besondere Fundensembles römischer Zeit – 30. Die Tierknochen660
Abb. 30/42. Entwicklung der Alterstruktur bei Schwein in der grossen Villa (Periode II).
30.3.4. Hausschwein
Nach Fragmentzahlen ist das Hausschwein in den Phasen 1/2(Periode II) nur die drittwichtigste Tierart (s. Abb. 30/36). In denfolgenden Phasen hingegen kommen die Schweinefragmentenoch häufiger vor als die Rinderknochen. Der gleiche Trend lässtsich auch bei den Gewichtsanteilen beobachten, allerdings mitdeutlich geringeren Werten.
Abb. 30/43. Entwicklung der Anteile der Fleischregionen bei Schweinin der grossen Villa (Periode II).

661
Viehbestand dürften hingegen die Ovicapriden und unter die-sen die Schafe überwogen haben.
In den Phasen 3/4 sind die Verhältnisse völlig verändert: DerAnteil der Rinder- und Schweineknochen liegt bei jeweils 40%,während die Ovicapridenknochen nur noch 20% ausmachen.In Bezug auf das Gewicht ist der Anteil der Rinderknochen zwargleich geblieben, hingegen haben die Schweine auf Kosten derOvicapriden an Bedeutung gewonnen. Der Konsum an Rind-fleisch ist demnach gleich geblieben; er dürfte durch die unver-änderte Nutzung der Rinder als Arbeitstiere bedingt sein. Hin-gegen haben die Schafe/Ziegen zu Gunsten der Schweine starkan Bedeutung verloren. Die Wollproduktion wurde weitgehendaufgegeben, hingegen spielte die Fleischproduktion eine grös-sere Rolle. In den Phasen 5/6 und in Phase 7 liegen ähnliche Ver-hältnisse wie in den Phasen 3/4 vor.
30.3.6. Equiden
Der Erhaltungszustand der Equidenknochen lässt in denmeisten Fällen keine Zuweisung an Pferd, Maultier/Mauleselund Esel zu. Einzig aus der vor dem Bau der Werkstatt B-Phase C(Periode II) eingebrachten Auffüllung stammen ein Femur-fragment und ein Calcaneus, die einem Esel zu gewiesen wer-den können. Eselknochen kommen nördlich der Alpen inrömischem Zusammenhang nur selten vor. Aus einer Schicht des2./3. Jahrhunderts in den Vigier-Häusern in Solothurn (Son-dierung 1981) stammt ebenfalls ein Eselknochen2057. Denkbar istein Zusammenhang zwischen den Eselfunden und dem Bibe-rister Granarium D: Vielleicht wurden Esel beim Transport vonGetreidesäcken oder dem Dreschen des Getreides eingesetzt.
Der Anteil der Equidenknochen ist in allen Phasen geringund liegt nach Fragmentanzahlen deutlich unter 1% (s. Abb.30/36). Es dürfte sich hauptsächlich um Transport- und Reit-tiere handeln, die in der Villa ihr Ende gefunden haben. Wennsie überhaupt gegessen wurden, dürften sie für die Ernährungvon keiner Bedeutung gewesen sein. Ihre Knochen fanden sichdenn auch meistens ausserhalb der Gebäude.
30.3.7. Hund
Auch die Hundeknochen sind selten (s. Abb. 30/36). Wäh-rend sie in den Phasen 1/2 der Villa (Periode II) noch 1.5 % derFragmente ausmachen, liegen ihre Werte in den folgenden Pha-sen unter 1%. In diese Berechnungen nicht miteingeschlossensind die Überreste zweier Hunde, die beim Hoftor zum Vor-schein kamen und zeitlich nicht näher eingeordnet werden kön-nen (Kap. 30.2.12).
MasseDas in der Nähe des Hoftores gefundene (Kap. 30.2.12), zu
grossen Teilen erhaltene Skelett stammt von einem Rüden, dereine Widerristhöhe von 54,4-58 cm aufwies. Nach der Einteilungnach J. Peters gehört er somit zu den mittelgrossen römischenHunden. Der aufgrund des Radius berechnete Schlankheitsin-dex spricht für ein normalwüchsiges Tier. Im Durchschnittwaren Hunde von Gutshöfen und Vici entsprechend ihrerFunktion als Wach- oder Herdenhunde grösser als jene aus Städ-ten, wo vermehrt mit Schosshunden zu rechnen ist2058.
30.3.8 Hausgeflügel
Unter den Vogelknochen sind die Hühnerknochen amhäufigsten vertreten (s. Tab. 30/3). Allerdings machen auch sienie über 2% der bestimmbaren Tierknochen aus. Einzig aus denPhasen 1/2 stammt ein Gänseknochen, der unter den Haustie-ren aufgeführt wird, da Gänse in städtischem Zusammenhang(z.B. Augst) häufig zusammen mit den Haushühnern vorkom-men2059. Das Hausgeflügel war somit von untergeordneter Be-deutung für die Ernährung der Villenbewohner. Nicht auszu-schliessen ist hingegen eine Produktion von Eiern und Federn.
Schwierig ist in römischem Zusammenhang die Zuweisungvon Entenknochen an die domestizierte oder die Wildform. Ausschriftlichen und bildlichen Quellen hauptsächlich Italienswissen wir, dass Enten als Haustiere gehalten wurden2060. Ob diesallerdings in den Nordwestprovinzen auch der Fall war, istschwer zu sagen. Anatomisch-morphologisch ist jedenfallskaum eine Zuweisung möglich. Ein Hinweis auf die Wildformkönnte ihr ausschliessliches Auftreten in den Phasen 5/6 und inPhase 7 sein, in denen ein erhöhter Anteil an Wildtieren fest-zustellen ist2061.
Ähnliche Probleme liegen bei den Tauben vor. Allerdingskann hier argumentiert werden, dass in unserem Gebiet dieWildform fehlt. Trotzdem ist nicht mit Sicherheit zu sagen, obdomestizierte oder wilde Tauben eingeführt wurden, weshalb sieauf den Tierartenlisten ebenfalls unter der Grossgruppe Haus-/Wildvögel aufgeführt sind.
30.3.9. Wildtiere
Der Anteil der Wildtiere unter den bestimmbaren Tier-knochen steigt kontinuierlich von der ersten bis zur letztenPhase der grossen Villa (Periode II) (s. Abb. 30/36). In den Pha-sen 1/2 ist der Anteil mit 0,7% nach Fragmenten bzw. 1,4% nachGewicht noch unbedeutend. Auch in den Phasen 3/4 erreicht ernur 1% nach Fragmenten, allerdings schon 3,1% nach Gewicht.In den Phasen 5/6 wird dann auch beim Fragmentanteil 3,3%erreicht, während sich der Gewichtsanteil leicht steigert. Einbeträchtliche Zunahme ist in der letzten Phase 7 zu verzeichnen:Die Wildtiere erreichen nun einen Fragmentanteil von fast 8%und einen Gewichtsanteil von sogar 13,4%.
Unter den wenigen Wildtierknochen der Phasen 1/2 und 3/4spielt keine Tierart eine besondere Rolle (s. Tab. 30/4): In denPhasen 1/2 sind Hirsch- und Hasenknochen gleich häufig. Inden Phasen 3/4 stehen zwei Wildschweinknochen je einemHirsch- und einem Rehknochen gegenüber. Die Zunahme derWildtiere in den Phasen 5/6, insbesondere aber Phase 7, wirdallein durch höhere Anteile des Rothirsches verursacht, währenddie Anteile der Reh-, Wildschwein- und Hasenknochen sichkaum verändern. Nur für die letzte Phase 7 liegt genügend Mate-rial für eine Alters- und Skeletteilanalyse des Hirsches vor. Die
Kap. 30.3
2057 Unpubliziertes Manuskript Guido Breuer.2058 Peters 1997, 518-519.2059 Zum gemeinsamen Vorkommen: wie Anm. 2017, 376-377.2060 Toynbee 1983, 260-262.2061 Der Unsicherheit wegen wurden die Enten auf den Tabellen der Tierartender unbestimmbaren Grossgruppe zugeordnet.

meisten Knochen des Rothirsches stammen von ausgewachse-nen Tieren (Abb. 30/44). Geweihfunde sind kein Beleg für dieJagd auf männliche Tiere, da es sich auch um im Wald aufgele-sene Abwurfstangen handeln könnte, Knochenfunde hingegenschon. Beim Skeletteilspektrum fällt die deutliche Übervertre-tung des Autopodiums auf (Abb. 30/46). Möglicherweisegelangte nicht das ganze Tier in den Gutshof, sondern nur dasentbeinte Fleisch und das Fell, in dem noch die Fussknochensteckten. Da sich bei einem Jagdausflug in weiter entfernteGebiete die gleichen, beim Schweinefleisch diskutierten Frisch-halteprobleme ergeben, ist anzunehmen, dass die Jagdgebietenicht allzu weit weg lagen, etwa im nahen Oberwald, und in derkälteren Jahreszeit gejagt wurde, in der auf den Feldern weni-ger Arbeit anfiel.
Der Anteil des Rothirsches ist vor allem in Phase 7 so hoch,dass eine Analyse der Grössenindizes gewagt werden kann. AlsVergleich bieten sich die ungleich besser vertretenen Hirsch-knochen im Gutshof von Neftenbach an. Metrische Untersu-chungen haben gezeigt, dass die Hirsche dort im Vergleich zuanderen römischen Fundstellen sehr gross gewachsen waren undeher noch den neolithischen Hirschen entsprechen, weswegenauf ein gutes Biotop und auf eine erfolgreiche Hirschpopulationgeschlossen worden ist2062. Der Vergleich der Grössenindizes derHirsche beider Villen zeigt nun, dass die Biberister Hirschewesentlich kleiner waren (Abb. 30/45). Auch der Vergleich derEinzelmasse bei Astragli und Calcanei zeigt im Vergleich zu Nef-tenbach auffallend geringe Masse2063. Dies lässt vermuten, dassdas Umland der Biberister Villa ein schlechteres Biotop für Hir-sche bot, da es wahrscheinlich seit der Eisenzeit agrarisch stär-ker genutzt wurde (s. aber Kap. 18.2.2). Das Gebiet um Nef-tenbach hingegen bot Hirschen ideale Lebensgrundlagen, wassich dann die Bewohner dieses Gutshofes für die Errichtungeines auf die Hirschfleischproduktion spezialisierten Betriebeszunutze machten2064.
Hohe Anteile an Hasen gelten allgemein als Indikatoren fürdie Öffnung einer Landschaft. Aus den vorliegenden Resultatenkönnte man somit schliessen, dass die Naturlandschaft der Bibe-rister Villa im Verlaufe der 200 Jahre ihrer Dauer kaum ihr Aus-sehen verändert hätte. Lag dies daran, dass – wie bereits erwähnt– das schweizerische Mittelland schon seit der Eisenzeit dichtbesiedelt und agrarisch genutzt wurde und in römischer Zeitkeine zusätzliche Öffnung der Landschaft mehr stattfand2065?Allerdings hat gerade in römischem Zusammenhang der öko-logische Aussagewert von Wildtieren nur bedingt Gültigkeit, dadie Häufigkeiten der in eine Siedlung gelangten Wildtierknochenin erster Linie vom menschlichen Zutun abhängt2066.
In Phase 7 ist zusätzlich eine Vergrösserung der Diversität anWildtieren festzustellen (s. Tab. 4). Neu vertreten ist die Wild-katze mit einem Phalangen aus der Einfüllung eines Ofenkan-als in Gebäude Q, dessen Hohlraum erst nach der Zerstörungaufgefüllt wurde. Die Wildkatze ist ein sehr scheues Wildtier, dassonnige, warme und trockene Standorte bevorzugt2067. Nachweiseauf dem Gebiet der heutigen Schweiz in römischer Zeit wurdennur in juranahen Siedlungen gefunden2068. Mit dem Fund ausBiberist liegt nun ein weiteres Beispiel vor. Vielleicht unternahmman von der Villa aus Jagdausflüge in den nahen Jura. Allerdingsist anzunehmen, dass die Wildkatze nicht ihres Fleisches, son-dern ihres Pelzes wegen erlegt wurde. Da in Biberist eine Pha-
V. Funde und besondere Fundensembles römischer Zeit – 30. Die Tierknochen
2062 Wie Anm. 2013, 454-455.2063 Wie Anm. 2013, 455 Abb.785.2064 Deschler-Erb 2001, 51f.2065 S. die hohe Stetigkeit, mit der Hasenknochen an eisenzeitlichen Fundstel-len auftreten (Schibler u.a. 1999, 133 fig. 61).2066 Deschler-Erb 2001, 47.2067 Hausser 1995, 413-415.2068 Basel-Münsterhügel (Deschler-Erb u.a. 1998, 157); Zusammenstellung(Olive/Deschler-Erb 1999, 37).
662
Abb. 30/44. Alterstruktur bei Hirsch in der letzten Phase der grossenVilla (Periode II-Phase 7).
Abb. 30/45. LSI (Logarithmic Size Index) der Hirschknochen von Biberist-Spitalhof und Neftenbach.

663
lanx gefunden wurde, ist es auch möglich, dass nur der vom Kör-per abgezogene Pelz bis in den Gutshof gelangte.
Zum erstenmal in Phase 7 ist der Biber belegt (s. Tab. 4). EinSchneidezahn stammt aus einer Grube südöstlich des Gebäu-des E und ein Femur aus dem Brandschutt des Gebäudes K desnachfolgenden Gehöfts (Periode III) des späten 3. Jahrhunderts.Die nähere Umgebung bot mit der Aare und ihren Nebenflüs-sen und -bächen einen guten Lebensraum für diesen in Aue-wäldern lebenden Nager. Biber könnten sowohl des Pelzes alsauch ihres Fleisches wegen die Villenbewohner interessierthaben. Schliesslich sei darauf hingewiesen, dass das heutige DorfBiberist seinen Namen dieser Tierart verdankt2069.
Bei den meisten anderen in Phase 7 neu auftretenden, über-dies kleineren Tierarten ist anzunehmen, dass sie ohne mensch-liches Zutun hierher kamen und ihr Ende fanden. Die Reste eini-ger Tiere dürften erst später in die lange an der Erdoberflächegelegenen Ruinen gekommen sein. Es handelt sich um Überrestevon Igeln, kleinen Nagern, Amphibien und Reptilien (s.Tab. 30/5). Eine Muschelschale musste unbestimmt bleiben. DieHausratte aus der Verfüllung der Ofenkanäle in Gebäude Q(Kap. 30.2.11) dürfte ebenfalls zufällig hier verendet sein. Einweiterer Rattenknochen fand sich in den Schlämmfunden desGebäudes O (Kap. 30.5.3). Während die Amphibien aus dernäheren Umgebung der Villa stammen, stellt die Hausratte einenunfreiwilligen menschlichen Import dar. Als Kulturfolger brei-teten sich Ratten über die gesamte antike Welt aus. Ihr Ver-
breitungsgebiet, das sich hauptsächlich den grossen Handels-wegen entlang zieht, deckt sich mit einiger zeitlicher Verzöge-rung mit der Expansion des Römischen Reiches, so dass sich anihrer Verbreitung mehr oder weniger der Stand der Romani-sierung ablesen lässt2070. In Biberist könnte der Rattenfund imZusammenhang mit dem Granarium D zu sehen sein. Mögli-cherweise gelangte das Tier mit Getreidelieferungen, die auf derAare verschifft wurden, in die Villa (s. auch Kap. 28.6.3). Da derFund weit weg von den neuzeitlichen Höfen (Kap. 17.6.2) auffreiem Feld zu Tage kam, dürfte die Ratte wohl schon kurz nachdem Ende der Gutshofanlagen im verfallenen Ofenkanal veren-det sein.
Zusammenfassend lässt sich festhalten, dass die deutlicheZunahme der Wildtieranteile in den Phasen 5/6 und vor allemin Phase 7 hauptsächlich mit der Intensivierung der Hirschjagdzusammenhängt. Aufgrund der Auflassung des Grabes H wirdfür Phase 7 mit einem neuen Besitzer gerechnet (Kap. 17.4.7),der vielleicht ein begeisterter Jäger war und seinem Hobby häu-figer nachging. Mit dem Fund einer Wildkatze sind Beziehun-gen zum Jura belegt2071. Die Hirsche dürfte man hingegen in dernäheren Umgebung der Villa, südlich gelegenen im Oberwald,gejagt haben. Die grössere Diversität an Wildtieren in Phase 7im Vergleich zu den vorangegangenen Phasen der grossenVilla (Periode II) ist mehrheitlich auf die Funde kleinererTierarten zurückzuführen, die unter natürlichen Umständen inden Boden gelangten.
Kap. 30.3
2069 Grossenbacher Künzler 1995, 71-72.2070 Sorge 1995; Audoin-Rouzeau/Vigne 1997.2071 Vergleiche dazu die Ausführungen wie Anm. 2066, 51.
Abb. 30/46. Anteile der Fleischregionen (g%) bei Hirsch in der letzten Phase der grossenVilla (Periode II-Phase 7) (n=87).

30.4. Schlussbetrachtungen zur Auswertung der handaufgelesenen Tierknochen
Die archäozoologischen Untersuchungen haben folgendeResultate erbracht: In der Anfangszeit des grossen Gutsbetrie-bes, Phasen 1-2 der Villa (Periode II) des letzten Viertel des1. Jahrhunderts und ersten Viertel des 2. Jahrhunderts, war dieHaltung von kleinen Wiederkäuern, wohl mehrheitlich Schafenzur Wollproduktion, von grösserer wirtschaftlicher Bedeu-tung. Es besteht wohl ein direkter Zusammenhang zum Betriebder Waschanlage und Walkerei in Gebäude B-Phase A. Obwohlerst in fortgeschrittenem Alter geschlachtet, kamen die Tiere imSitz des Vilicus, Holzbau J, auf den Tisch. Wahrscheinlich warder Betrieb zu dieser Zeit auf eine Überschussproduktion vonWolle und vielleicht Milch bzw. Käse ausgerichtet. Die Rinderwurden, wie auch in den folgenden Phasen, hauptsächlich alsArbeitstiere eingesetzt.
Ab dem zweiten Viertel des 2. Jahrhundert (Phase 3) sind biszum Ende der grossen Villa (Periode II), kurz nach der Mitte des3. Jahrhunderts, die Schweine die am häufigsten vertretene Tier-art; die reine Fleischproduktion ist wichtiger geworden als dieHerstellung sekundärer Produkte. Die Rinder liegen in Bezug aufdie Fragmentanzahl nur an zweiter Stelle. Der Anteil der Scha-fe/Ziegen geht sukzessive zurück, wobei nun die Fleischpro-duktion an erster Stelle steht. In der letzten Phase 7 der Villa(Periode II), im mittleren 3. Jahrhundert, gibt es bei den Haus-tieranteilen keine wesentlichen Veränderungen. Die Bedeutungder Rinder für die Ernährung scheint indes leicht zurückzuge-hen.
Die Grössenindizes der Biberister Rinder entsprechen den-jenigen von Augst und nehmen im gleichen Masse im Verlaufder Zeit zu. Daraus lässt sich schliessen, dass die Viehzucht inrömischer Zeit regional vernetzt gewesen sein dürfte. Zu denanderen Nutztieren lassen sich leider keine metrischen Unter-suchungen durchführen.
Der Wildtieranteil, der durch die Rothirschknochen klardominiert wird, nimmt von der ersten zur letzten Phase der Villa(Periode II) kontinuierlich zu, wobei zwischen Phasen 5/6 undPhase 7 ein markanter Anstieg festzustellen ist, der mit dem pos-tulierten Besitzerwechsel übereinstimmt. Die Hirsche in derUmgebung von Biberist waren durchschnittlich deutlich klei-ner als diejenigen von Neftenbach, was auf unterschiedliche Bio-tope in den beiden Regionen schliessen lässt.
Zwischen den einzelnen Gebäuden liessen sich zum Teildeutliche Unterschiede in der Zusammensetzung der Tierartenfeststellen. Diese sind durch die spezifischen Funktionen zuerklären, die den einzelnen Gebäuden zukam: Die Tierknochender als Waschhaus/Walkerei gedeuteten Werkstatt B-Phase Akönnen direkt keine Hinweise zur Funktion des Gebäudes lie-fern; nur die Dominanz der Schaf/Ziegenknochen im gleich-zeitigen Holzbau J und während der gesamten Phasen 1/2 deu-tet auf Wollverarbeitung in diesem Gebäude hin. Da im Wasch-haus/Walkerei die Rinder- und Schweineknochen wichtig sind,ist anzunehmen, dass die hier gefundenen Knochen nicht ausdem Holzbau J stammen, sondern aus einem oder mehrerenanderen Häusern hierher gebracht worden sind. Auch beimMaterial der Phasen B und C ist anzunehmen, dass die Tier-
knochen nicht in der Werkstatt B, in dieser Zeit eine Schmiede,angefallen, sondern sekundär hier abgelagert worden sind,vielleicht z.T. aus dem nahen Landarbeiterhaus A.
Im Getreidespeicher D fanden sich zwei Spezialbefunde: Ineiner Kellergrube im Hauptraum 1 waren drei rechte Schädel-hälften mit Rinderhornzapfen versenkt, die wahrscheinlich inkultischem Zusammenhang zu sehen sind. Unter den Schutt-massen des kurz nach der Mitte des 3. Jahrhunderts abgebrann-ten Gebäudes fanden sich hauptsächlich Rumpfteile eines Rin-des, die zum Teil im anatomischen Verband angetroffen wurden.Es dürfte sich um die Überreste von grösseren Fleischteilen han-deln, die im Erdgeschoss des Getreidespeichers aufgehängt undwährend kurzer Zeit, bis zur Konsumation oder zur Konser-vierung in der Anlage des Gebäudes Q zwischengelagert waren,was durch den Gebäudebrand verhindert wurde.
Das zwar sehr umfangreiche Material aus Gebäude E istextrem schlecht erhalten, unterstützt aber die Deutung des Ge-bäudes als Wohnhaus. Die Zusammensetzung spricht für Spei-seabfälle wohlhabender Bewohner. Der Hirschanteil ist auffal-lend hoch. Spezifische Abfälle deuten auf die Herstellung vonKnochenartefakten für den Eigenverbrauch hin.
In einer Grube bei der nördlichen Trennmauer fand sich derKadaver einer sechs bis sieben Jahre alten Kuh, die ins spätere3. Jahrhundert, in die letzte Phase der Villa (Periode II) oder indie Zeit des nachfolgenden Gehöfts (Periode III), datiert. Aus-ser dem Hinterhaupt und den Fussknochen ist das Skelett voll-ständig. Auch zeichnete sich das einstige Fleischvolumen imSediment ab, so dass anzunehmen ist, dass das wohl an einerKrankheit gestorbene Tier nur enthäutet und dann im Bodenversenkt worden war. Mögliche Reste eines weiteren vergrabe-nen Rindes fanden sich nur knapp 15 m weiter östlich.
Das Vilicushaus J ist das fundreichste Gebäude der Villa.Unter den Tierknochen des Holzbaus J sind die Schafe/Ziegenaussergewöhnlich häufig. Der hohe Anteil alt-adulter Tierespricht nicht für eine ausgesuchte Küche, sondern zeigt, dass diefür sekundäre Produkte, wohl Wolle, gehaltenen Tiere anschlies-send noch gegessen wurden. Spätestens ab der Phase 3 des Stein-baus J ändert sich das Tierartenspektrum grundlegend: Wäh-rend nun die Schweineknochen dominieren, sind die Anteile derSchaf-/Ziegenknochen deutlich zurückgegangen; der hohe Pro-zentsatz von Jungtieren spricht nun für eine reine Fleischver-wertung. Der Anteil der Schafe/Ziegen ist aber vergleichsweisenoch immer höher als im gleichzeitigen Landarbeiterhaus O,wodurch die sozialen Unterschiede zwischen den Bewohnern derbeiden Gebäude auch archäozoologisch belegt werden können.Die Schnittspurenanalyse zeigt ebenfalls Unterschiede zwi-schen dem Vilicushaus J und dem Landarbeiterhaus O auf.
Im Gebäude P, wohl einem Wachhaus, das der Siedlungs-phase 2 angehört, fanden sich im Gegensatz zum Holzbau J derPhase 1 auffallend wenige Schaf-/Ziegenknochen. Dies belegterneut, dass schon in der Frühphase der Villa mit verschiedenenPersonengruppen zu rechnen ist.
Im Landarbeiterhaus O ist das Schwein die dominante Tier-art. Daneben spielen Geflügelknochen, hauptsächlich Huhn, einewichtigere Rolle. Möglicherweise wurde die Eierproduktiondurch die dort lebenden Bewohner betreut.
Der Ofen des erst in Phase 7 errichteten Gebäudes Q dürf-te aufgrund der archäozoologischen Resultate zur Fleischkon-
V. Funde und besondere Fundensembles römischer Zeit – 30. Die Tierknochen664

665
servierung gedient haben. Die Hirschanteile liegen bei 30% undsind als extrem hoch zu bezeichnen. Bei den Schweineknochenkommen jene Skeletteile vor, die bei der Vorbereitung der Fleisch-teile zum Konservieren anfallen. In einer Grube neben der Anla-ge fanden sich hauptsächlich Rinderrumpfteile, die an diezwischengelagerten Teile in Gebäude D erinnern.
Beim Hoftor fand sich das Skelett eines mittelgrossen Rü-den, der zwischen eineinhalb und zwei Jahre alt wurde. Erscheint frühzeitig ums Leben gekommen sein. Sein Vergrabungs-ort könnte symbolische Bedeutung haben.
Vom Gehöft des späten 3. Jahrhunderts fanden sich nur we-nige Tierknochen, die nur hypothetische Schlüsse zulassen. Eswurde offensichtlich keine Grosswildjagd mehr betrieben. In die-ser Zeit lebte vermutlich nur noch eine Bauernfamilie, die Selbst-versorgung betrieb und nur noch die im nördlichen Teil des ein-stigen Fundus gelegenen Felder bewirtschaftete.
30.5. Die Tierknochen aus denSchlämmproben
(HEIDE HÜSTER PLOGMANN)
30.5.1. Einleitung
Auf dem Gebiet der heutigen Schweiz ist hinsichtlich derUntersuchung von Kleintierresten bislang nur ein römischerGutshof repräsentativ erfasst, nämlich der Gutshof Neften-bach2072. Aber auch andere Siedlungseinheiten sind keineswegshinreichend aufgenommen, so dass das vorliegende Material inmehr als einer Hinsicht hilft, Lücken zu schliessen. Die Fundeumfassen einen Zeitraum vom letzten Viertel des 1. Jahrhundertn.Chr. bis zum dritten Viertel des 3. Jahrhunderts n.Chr., dasMaterial ist zudem feinchronologisch auftrennbar. Damit dür-fen Antworten auf folgende Fragen erhofft werden: WelchenStellenwert hatten einheimische Wirbeltiere und hier besondersdie fast ausschliesslich in Schlämmproben erfassten Fischeund Singvögel in der Ernährung der Bewohner der Villa?Wurde die gesamte Fauna der Umgebung genutzt oder wurdeselektiv gejagt bzw. ausgelesen? Sind Nahrungsmittelimporte ausdem Süden nachweisbar? Und schliesslich: Sind auf dem Arealräumliche Unterschiede in der Ernährung und/oder chronolo-gische Entwicklungen der Esskultur festzustellen?
Material und MethodeDie Untersuchung der Villa von Biberist schloss als eine der
ersten Grabungen in der Schweiz eine systematische Beprobungerfolgversprechender Befunde mit ein (Kap. 3.2.2; 28.2.1).Demzufolge standen schliesslich 252 Proben aus 130 Struktu-ren bzw. Schichten zur Verfügung. Die vorliegende Untersu-chung basiert auf 128 Proben. 34 Proben aus der Grabanlage Hsind zusammen mit den menschlichen Resten bearbeitet wor-den (Kap. 31), weitere Proben sind fundleer bzw. enthalten aus-schliesslich hochfragmentierte, und damit nicht mehr bestimm-bare Reste grösserer Säugetiere; sie alle sind nicht in die vorlie-genden Analysen einbezogen worden.
Von den 128 Proben hat 77 Sabine Deschler-Erb 1996 bear-beitet, die verbleibenden 51 Proben hat Heide Hüster Plogmann2001 analysiert, die für die Auswertung und die Erstellung desManuskriptes verantwortlich zeichnet.
Die vorliegende Untersuchung basiert insgesamt auf 12’594Skelettresten (Tab. 30/12). Bei einem Volumen von mindestenszehn Litern für die „grobgeschlämmten Proben“ (G) bzw. zweiLitern für die sog. Reserveproben (R) je Probe ergibt sich einedurchschnittliche Funddichte von etwa einem Knochenfragmentpro Liter Erdreich2073. Der Anteil bestimmbarer Reste liegt beiknapp 40% (n=4786), so dass die Dichten von Resten einzelnerTierarten pro Liter weit unter einem Fragment liegen. Aus die-sem Grunde wird auf die Berechnung der einzelnen Fund-dichten verzichtet. Die Fundarmut an bestimmbaren zoologi-schen Resten verunmöglicht eine chronologische Betrachtung.In drei der vier definierten Zeithorizonte sind lediglich 65-161Fragmente einer Tierart zuzuordnen. Unter ihnen befinden sichzu 50-75% Reste von Mäusen, sodass Aussagen zu wirtschaft-lichen und/oder kulturellen Entwicklungen in der Ernährungder Bewohner des Gutshauses spekulativ bleiben.
17 der geborgenen Skelettelemente zeigen, dass sie einenDarmtrakt passiert haben. Bei ihnen handelt es sich zumeist umFischwirbel aus der Grube A118/B072 (n=14) zwischen dem Torund dem Landarbeiterhauses O (Kap. 10.1.1). Die Wirbel sind,verursacht durch Verdauungssäfte und Darmperistaltik, intypischer Weise lateral und cranio-caudal gestaucht.
Unter den unbestimmbaren Fragmenten dürfte der grössteTeil nach der Knochenstruktur zu urteilen von mittelgrossen bisgrossen Säugetieren stammen, also vermutlich von Nutzhaus-tieren und vereinzelt möglicherweise von grösserem jagdbarenWild (s. Kap. 30.3). Das Durchschnittsgewicht dieser Fragmenteliegt unter 0,1g, was die Grösse der Fragmente vor Augen füh-ren mag.
Die Bestimmung der Reste erfolgte anhand der Vergleichs-sammlung der Archäozoologischen Abteilung des IPNA der Uni-versität Basel. Ein Teil der Fragmente kann zwar der Gruppe„Fisch“ oder „Vogel“ zugeordnet werden, lässt aber eine nähe-re Bestimmung nicht zu. Bei ihnen handelt es sich meist um sehrkleine Röhrenknochen- oder Wirbelfragmente (Vögel) oderaber, bei den Fischen, um fragmentierte, wenig differenzierteHartteile, wie Flossenträger, Flossenstrahlen, Rippen, Kiemen-deckelstrahlen, Elemente des Branchialskelettes oder Schuppen.Für die Fische wurde nach der Bestimmung versucht, dieLänge der Tiere abzuschätzen. Zum einen wurden ganz erhal-tene Skelettelemente in 5 cm-Klassen eingestuft, fragmentierteElemente wurden dagegen nur sehr grob eingeteilt. Die Rubri-ken „sehr klein“ und „klein“ umfassen Fische, die bis etwa 15 cmgross waren, in der Rubrik „mittel“ finden sich Tiere von etwa20-30 cm Länge, unter „gross“ sind schliesslich Individuen mit35-50 cm Körperlänge anzusiedeln. Alle Schätzungen dieneneiner groben Orientierung und können keine exakte Rekon-struktion der Fischgrössen darstellen2074.
Kap. 30.5
2072 Rychener 1999.2073 Zur Definition der verschiedenen Proben s. Kap. 28.2.1.2074 Casteel 1976.
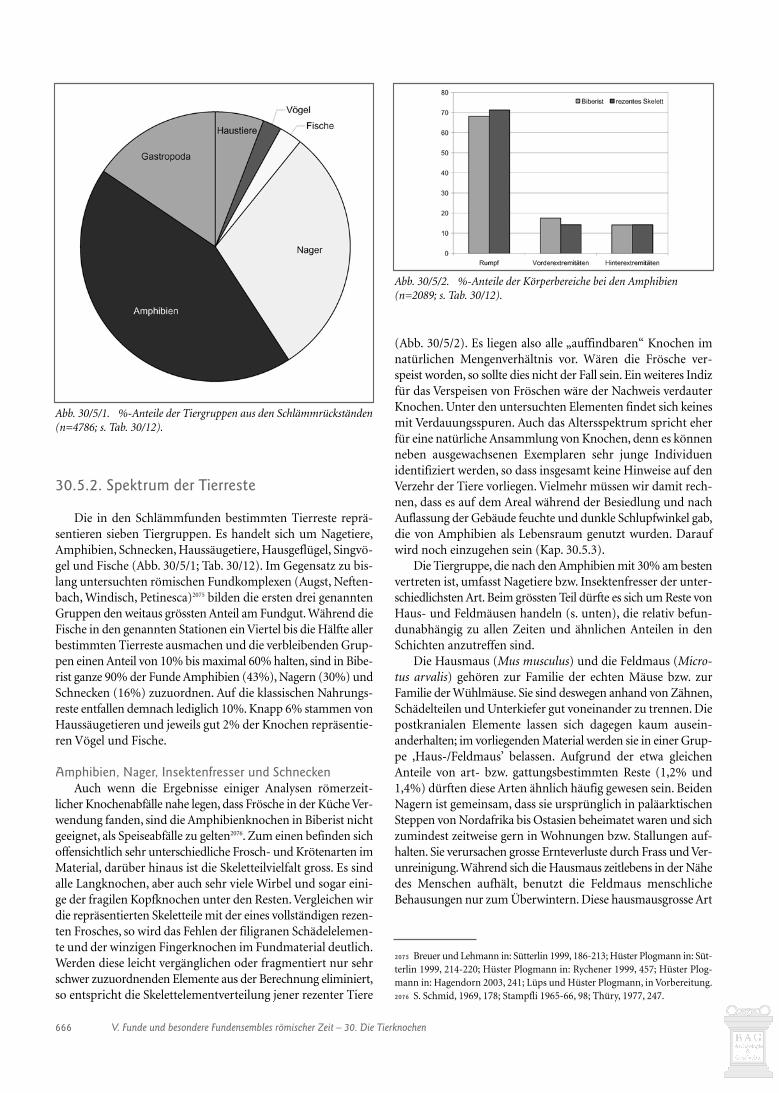
30.5.2. Spektrum der Tierreste
Die in den Schlämmfunden bestimmten Tierreste reprä-sentieren sieben Tiergruppen. Es handelt sich um Nagetiere,Amphibien, Schnecken, Haussäugetiere, Hausgeflügel, Singvö-gel und Fische (Abb. 30/5/1; Tab. 30/12). Im Gegensatz zu bis-lang untersuchten römischen Fundkomplexen (Augst, Neften-bach, Windisch, Petinesca)2075 bilden die ersten drei genanntenGruppen den weitaus grössten Anteil am Fundgut. Während dieFische in den genannten Stationen ein Viertel bis die Hälfte allerbestimmten Tierreste ausmachen und die verbleibenden Grup-pen einen Anteil von 10% bis maximal 60% halten, sind in Bibe-rist ganze 90% der Funde Amphibien (43%), Nagern (30%) undSchnecken (16%) zuzuordnen. Auf die klassischen Nahrungs-reste entfallen demnach lediglich 10%. Knapp 6% stammen vonHaussäugetieren und jeweils gut 2% der Knochen repräsentie-ren Vögel und Fische.
Amphibien, Nager, Insektenfresser und SchneckenAuch wenn die Ergebnisse einiger Analysen römerzeit-
licher Knochenabfälle nahe legen, dass Frösche in der Küche Ver-wendung fanden, sind die Amphibienknochen in Biberist nichtgeeignet, als Speiseabfälle zu gelten2076. Zum einen befinden sichoffensichtlich sehr unterschiedliche Frosch- und Krötenarten imMaterial, darüber hinaus ist die Skeletteilvielfalt gross. Es sindalle Langknochen, aber auch sehr viele Wirbel und sogar eini-ge der fragilen Kopfknochen unter den Resten. Vergleichen wirdie repräsentierten Skeletteile mit der eines vollständigen rezen-ten Frosches, so wird das Fehlen der filigranen Schädelelemen-te und der winzigen Fingerknochen im Fundmaterial deutlich.Werden diese leicht vergänglichen oder fragmentiert nur sehrschwer zuzuordnenden Elemente aus der Berechnung eliminiert,so entspricht die Skelettelementverteilung jener rezenter Tiere
(Abb. 30/5/2). Es liegen also alle „auffindbaren“ Knochen imnatürlichen Mengenverhältnis vor. Wären die Frösche ver-speist worden, so sollte dies nicht der Fall sein. Ein weiteres Indizfür das Verspeisen von Fröschen wäre der Nachweis verdauterKnochen. Unter den untersuchten Elementen findet sich keinesmit Verdauungsspuren. Auch das Altersspektrum spricht eherfür eine natürliche Ansammlung von Knochen, denn es könnenneben ausgewachsenen Exemplaren sehr junge Individuenidentifiziert werden, so dass insgesamt keine Hinweise auf denVerzehr der Tiere vorliegen. Vielmehr müssen wir damit rech-nen, dass es auf dem Areal während der Besiedlung und nachAuflassung der Gebäude feuchte und dunkle Schlupfwinkel gab,die von Amphibien als Lebensraum genutzt wurden. Daraufwird noch einzugehen sein (Kap. 30.5.3).
Die Tiergruppe, die nach den Amphibien mit 30% am bestenvertreten ist, umfasst Nagetiere bzw. Insektenfresser der unter-schiedlichsten Art. Beim grössten Teil dürfte es sich um Reste vonHaus- und Feldmäusen handeln (s. unten), die relativ befun-dunabhängig zu allen Zeiten und ähnlichen Anteilen in denSchichten anzutreffen sind.
Die Hausmaus (Mus musculus) und die Feldmaus (Micro-tus arvalis) gehören zur Familie der echten Mäuse bzw. zurFamilie der Wühlmäuse. Sie sind deswegen anhand von Zähnen,Schädelteilen und Unterkiefer gut voneinander zu trennen. Diepostkranialen Elemente lassen sich dagegen kaum ausein-anderhalten; im vorliegenden Material werden sie in einer Grup-pe ‚Haus-/Feldmaus’ belassen. Aufgrund der etwa gleichenAnteile von art- bzw. gattungsbestimmten Reste (1,2% und1,4%) dürften diese Arten ähnlich häufig gewesen sein. BeidenNagern ist gemeinsam, dass sie ursprünglich in paläarktischenSteppen von Nordafrika bis Ostasien beheimatet waren und sichzumindest zeitweise gern in Wohnungen bzw. Stallungen auf-halten. Sie verursachen grosse Ernteverluste durch Frass und Ver-unreinigung. Während sich die Hausmaus zeitlebens in der Nähedes Menschen aufhält, benutzt die Feldmaus menschlicheBehausungen nur zum Überwintern. Diese hausmausgrosse Art
V. Funde und besondere Fundensembles römischer Zeit – 30. Die Tierknochen
2075 Breuer und Lehmann in: Sütterlin 1999, 186-213; Hüster Plogmann in: Süt-terlin 1999, 214-220; Hüster Plogmann in: Rychener 1999, 457; Hüster Plog-mann in: Hagendorn 2003, 241; Lüps und Hüster Plogmann, in Vorbereitung.2076 S. Schmid, 1969, 178; Stampfli 1965-66, 98; Thüry, 1977, 247.
666
Abb. 30/5/1. %-Anteile der Tiergruppen aus den Schlämmrückständen(n=4786; s. Tab. 30/12).
Abb. 30/5/2. %-Anteile der Körperbereiche bei den Amphibien(n=2089; s. Tab. 30/12).

667
lebt im Sommer vorwiegend auf Wiesen und Weiden. Der Bo-den muss jedoch gut entwässert sein, um den aus der Steppestammenden Kulturfolger zu beherbergen. Von der etwas grös-seren, ebenfalls zu den Wühlmäusen gehörenden Schermaus(Arvicola terrestris) konnten 24 Skelettelemente geborgen wer-den. Primärbiotope der Art sind dicht bewachsene Ufer ste-hender oder langsam fliessender Gewässer, wie sie im Bereichder Villa, aber auch in der unmittelbaren Umgebung der Villaanzutreffen waren (s. Kap. 18.2.2).
Die Hausratte gehört, wie die Hausmaus, zur Familie derEchten Mäuse (Muridae). Sie war ursprünglich auf dem indi-schen Subkontinent beheimatet. Begünstigt durch den überse-eischen Handel und Warenverkehr erreichte sie wohl noch in vor-christlicher Zeit Ostafrika und die Mittelmeerländer2077. SichereNachweise liegen für unseren Raum inzwischen für die römischeZeit vor (Neftenbach, Augst)2078. Auch in Biberist ist die Hausrattedurch drei Reste im Gebäude O in den Phasen 1 sowie in denPhasen 3 und 4 nachgewiesen. Darauf wird noch einzugehen sein(Kap. 30.5.3). In unseren Breiten hält sich die wärmeliebende Artvorzugsweise in Wohngebäuden und Stallungen auf.
Neun Reste stammen von nicht näher zuzuordnendenWaldmäusen (Apodemus). Es dürfte es sich entweder um dieGelbhalsmaus (A. flavicollis) oder die Gemeine Waldmaus (A.sylvaticus) handeln. Beide Arten bewohnen Laub- und Misch-wälder, Waldränder, sie kommen aber auch in Gärten vor undsuchen im Winter oft einzeln stehende Gebäude oder Stallun-gen auf. Die Reste aus Biberist sind über alle Phasen und Gebäu-de verteilt.
Insektenfresser sind im Schlämmaterial durch zwei Artenvertreten, den Maulwurf (Talpa europaeus) und die Wald- bzw.Schabrackenspitzmaus (Sorex araneus/coronatus). 14 Reste deu-ten auf ein Mitglied der Familie der Spitzmäuse (Soricidae) hin,allerdings ist auch hier die Artzuweisung unsicher. Es kann sichsowohl um die Wald- oder die Schabrackenspitzmaus (Sorex ara-neus/coronatus) handeln. Wald- und Schabrackenspitzmäusesind besonders im Alpenvorland schwer voneinander zu tren-nen und in der Regel nur durch präzise Masse am Unterkiefer,biochemische Analysen oder ein Studium der Chromosomen zuunterschieden2079. Die 66-88 mm (Kopf-Rumpf-Länge) kleinenTiere konkurrenzieren zudem die gleichen Lebensräume. Wich-tig ist eine gute, den Boden bedeckende Pflanzenschicht. Nahebei Häusern werden sie in der Regel von den Hausspitzmäusenverdrängt. In Biberist stammen 10 der Spitzmausreste aus denKanälen des Gebäude Q, zwei weitere aus dem mutmasslichenGetreidespeicher D und schliesslich zwei aus der viertenPhase des Steinbaus J.
Der Maulwurf (Talpa europaeus) ist schliesslich durch 17gleichmässig über Gebäude und Zeitphasen verteilte Reste be-legt. Die 124-142 mm grossen Tiere sind überall dort vertreten,wo der Boden fruchtbar und tief genug ist, um Gänge zu gra-ben. Ursprünglich war der Maulwurf in Laubwäldern heimisch,bevölkerte aber sehr schnell Ackerland, Weiden und Gärten.
Aus Gebäude E und Q stammen schliesslich zwei Reste vonnicht näher zuzuordnenden Schläfern (Gliridae). Die Oberkie-fer- resp. Zahnfragmente deuten von der Grösse her entwederauf Siebenschläfer (Glis glis) oder Gartenschläfer (Eliomysquercinus). Beide Arten können in Gärten und sogar in Gebäu-den angetroffen werden.
16 % der tierischen Rest in Biberist machen Schalenfrag-mente von Gastropoden, also Schnecken, aus. Sie wurden nichtsystematisch bestimmt, sondern nur grob gesichtet. Insgesamthandelt es sich meistens um Reste grösserer Exemplare aus derFamilie der Helicidae. Am häufigsten dürfte unter ihnen diegebänderte Gartenschnecke (Cepea spec.) zu finden sein. Siezeichnet sich durch eine grosse Formen- und Farbenvariationaus, genauso variabel sind die Habitate, in denen sie zu findensind. Sie leben sowohl in Wäldern, Hecken, Grasland wie auchin Gärten. Des Weiteren fanden sich wenige Exemplare vonWeinbergschnecken (Helix pomatia). Diese Tiere belegen ähn-liche Lebensräume wie die Gartenschnecke, lieben jedoch be-sonders kalkhaltige Substrate. Die Schneckenreste fanden sichvorrangig in den Gebäuden D und Q (Tab. 30/5/X1).
SpeiseresteNur 10% der Tierreste aus den Schlämmproben weisen auf
Speisereste hin, die in bislang untersuchten römischen Komple-xen zwischen 70% und 90% der Tierreste aus Schlämmprobenausmachen. Einschränkend muss für das vorliegende Materialerwähnt werden, dass sich unter diesen Knochen auch drei Restevon Hunden und weitere fünf von nicht näher zu identifizie-renden Carnivoren befinden, die nicht als Speisereste eingestuftwerden dürfen.
Säuger und VögelDrei Hasenreste repräsentieren unter den Schlämmfunden
die einzigen verspeisten Wildtiere. Bei ihnen handelt es sich umdrei Oberkieferzähne aus dem Gebäude E. Hasen gelten als be-liebte Speise in römischer Zeit und sind in Biberist im hand-aufgelesenen Fundmaterial wenn auch nicht häufig, so dochregelmässig über das gesamte Areal nachgewiesen (s. Kap. 30.3.9).
Bei den Haussäugetieren liegt der Schwerpunkt der Arten-nachweise aus Schlämmproben notwendigerweise bei den klei-neren Arten, doch finden sich auch Reste grösserer Tiere. Amhäufigsten sind mit 208 Fragmenten (4%) die Schweine. DieWiederkäuer Schaf/Ziege und Rind sind durch 29 bzw. 32 Kno-chenfragmente repräsentiert (<1%). Wenn diese Relationen kei-nesfalls auf den tatsächlichen Verzehr schliessen lassen, so zei-gen sie doch einen grösseren Anteil an Schweinen gegenüberSchafen und Ziegen an. Die Arten haben eine vergleichbareGrösse und sollten demnach etwa gleich häufig im Schlämm-material vertreten sein. Während in den beiden ersten Phasender grossen Villa bei den handaufgelesenen Tierknochen dieOvicapriden überwiegen, dominieren ab der Phase 3 dieSchweine- und Rinderknochen (Kap. 30.3.5). Bei den imSchlämmmaterial gefunden Skelettelementen handelt es sichmeistens um Kleinknochen und Zahnfragmente (die bei hand-aufgelesenem Material unterrepräsentiert sind) sowie um Ske-lettelemente jüngerer Tiere. So machen auch im vorliegendenMaterial Zahnfragmente zwei Drittel der Haussäugerreste aus.
Kap. 30.5
2077 Becker 1978, 387.2078 Hüster Plogmann in: Rychener 1999, 422; Vogel Müller/Deschler-Erb 1996119.2079 Hausser 1995, 23-31.

Hausgeflügel ist in den Schlämmfunden nur durch Hühner(Gallus gallus) vertreten. Auch von dieser Art finden sich in denSiebrückständen eher Kleinknochen und – darüber hinaus –auch Eierschalenreste. Unter den 22 nachgewiesenen Hühner-resten gibt es allerdings nur vier Eierschalenfragmente, die ausden Gebäuden O und Q stammen. Insgesamt dürfte der Anteilder Hühnerknochen etwa um die nicht näher einzuordnendenVogelknochen zu erhöhen sein. Zumindest handelt es sich beidiesen 68 Resten um grössere Vogelartige. Da im handaufgele-senen Material unter dem Hausgeflügel die Hühner deutlichüberwiegen, ist dieser Schluss zulässig (Kap. 30.3.8).
Skelettelemente von Singvögeln – nach Apicius eine begehr-te Speise der römischen Küche – sind ausserhalb der Schlämm-proben sehr selten nachzuweisen. Im vorliegenden Materialnehmen sie einen Anteil von 0,5% oder 22 Fragmenten ein. Dass19 von ihnen nicht näher zugewiesen werden können, liegt ander starken Fragmentierung. Dies darf als Hinweis auf den Ver-zehr der Tiere gewertet werden. Drei Oberarmfragmente kön-nen Amselartigen und Finken zugeordnet werden. Sie stammenaus den Gebäuden O und E, sowie aus einer der Gruben in derNähe des Tores (s. Kap. 30.5.3).
FischeArtenzusammensetzung: Wie die Vögel sind auch die Fische
mit gut 2% des bestimmten Materials vertreten (Tab. 30/5/X1).Die 132 Reste repräsentieren zumeist die Süsswasserfauna derUmgebung der Villa. 31 Fischknochen lassen nur die Aussage zu,dass es sich um Fischreste handelt. Die zumindest bis zur Fami-lie bestimmbaren Fischknochen (n=101) zeigen eine Reprä-sentanz der Lachsartigen (Salmonidae) von 54%. Ihnen folgendie Karpfenartigen (Cyprinidae) mit 37%, das Egli (Perca fluvi-atilis) ist mit 7% am Fundmaterial beteiligt. Schliesslich fandensich drei Fischknochen, die unter „Sonstige“ subsummiertwerden. Es handelt sich bei ihnen zum einen um einen Cau-dalwirbel einer Trüsche (Lota lota), der aus einer der Grubenbeim Tor geborgen wurde. Dieser einzige Vertreter der Familieder Dorschartigen (Gadidae), der im Süsswasser vorkommt, lebtin den gleichen sauerstoffreichen und nährstoffarmen Gewäs-sern wie die Lachsartigen. Trüschen waren bis zum Beginn desletzten Jahrhunderts wegen ihres schmackhaften und festen Flei-sches und ihrer gesunden Leber (Lebertran) beliebte Speisefi-sche. Ausonius, ein im 4. Jahrhundert an der Mosel lebenderrömischer Schriftsteller, rühmt sie als „rühmliche Schützlinge derMosel“ und schätzt bei ihnen, dass sie „lange haltbar (sind) ohnezu verderben“2080.
Die zweite sich hinter den „Sonstigen“ verbergende Art istein Exot in der gallo-römischen Küche. Es handelt sich um einenCaudalwirbel und ein Schädelfragment der Mittelmeermakre-le (Scomber japonicus). So ungewöhnlich das Auftauchen einerArt aus dem Mittelmeer in der Reihe der vertrauten Arten ausder Region ist, es zeichnet sich immer deutlicher eine Vorliebefür diesen Importfisch im ganzen Imperium Romanum ab; derImport dürfte mit dem Import von Fischsauce und anderenFischkonserven in Zusammenhang stehen (s. Kap. 21.3.4). Ausder Schweiz liegen eine Reihe weiterer Nachweise vor, die frü-hesten Nachweise stammen aus der 2. Holzbauphase von Vin-donissa (1. Jahrzehnt v.Chr.) und aus Dangstetten (Ende1. Jahrhundert v.Chr.)2081. Hinweise auf die Mittelmeermakrele
mehren sich im deutschen Rheingebiet (Mainz), in Belgien, denNiederlanden und Grossbritannien. Die Reste von Biberiststammen aus den Phasen 1 und 3 des Gebäudes O.
Sieben Skelettelemente belegen Fang und Konsum des Egli(Perca fluviatilis) aus der Familie der Barsche (Percidae). Siestammen aus den Gebäuden O (4), E (2) und einer der Grubenbeim Tor (n=1). Das Egli hat damit einen Anteil von 7% an denbestimmten Fischresten (Abb. 30/5/3). Egli bewohnen mitVorliebe klare Gewässer mit hartem Grund. Als tolerante Art mitwenig Ansprüchen können sich Egli aber auch in anderen Ge-wässertypen ansiedeln. Besonders Jungfische schliessen sich gernzu Schwärmen zusammen und ziehen so am Ufer entlang. Bisheute gelten Egli als attraktive Speisefische. Ihr festes, kompak-tes Muskelfleisch war schon im 4. Jahrhundert beliebt, dennAusonius berichtet2082:
„Dich auch, Barsch, du Tafeljuwel, darf ich nicht übergehen;
unter den Flussfischen kommst du allein dem Seefisch an Rang gleich,
du vermagst mit der rötlichen Barbe des Meeres dich zu messen:
denn dein Geschmack ist pikant, und die Teile des kernigen Fleisches
drängen sich Schicht an Schicht zusammen, doch trennen sie Gräten.“
Die Familie der Karpfenartigen (Cyprinidae) bildet nach denLachsartigen den zweitgrössten Anteil am Fundmaterial. DieKarpfenartigen sind heute die artenreichste Familie in unserenGewässern und konnten auch problemlos in vielen eutro-phierten Teichen und langsam fliessenden Bächen und Flüssenin unmittelbarer Nähe der Villa erbeutet werden (s. auch Kap.
V. Funde und besondere Fundensembles römischer Zeit – 30. Die Tierknochen
2080 Ausonius, Mosella 103.2081 Hüster Plogmann in: Hagendorn 2003, 422; zu Dangstetten mündliche Mit-teilung von M. Uerpmann.2082 Ausonius, Mosella 115-119.
668
Abb. 30/5/3. %-Anteile der Arten bzw. Familien bei den Fischen(n=101; s. Tab. 30/12).

669
18.2.2). Morphologisch sind die Skelettelemente vieler Arten nurschwer zu unterscheiden, insbesondere wenn es sich um Jung-tiere handelt oder um fragmentierte Knochen. Allerdings sindeinige Elemente artspezifisch. Zu ihnen gehören die Schlund-knochen, auf denen – anders als auf den Kieferknochen – art-charakteristische Schlundzähne aufsitzen. Den meisten Kar-pfenfischen ist gemeinsam, dass sie eher in Schwärmen in denbewachsenen Uferregionen zu finden sind. Das gilt ganzbesonders für die Jungfische, die in den flachen, wärmerenRegionen besser gedeihen und zwischen den Pflanzen vorFressfeinden Schutz finden.
Artbestimmt können das Rotauge (Rutilus rutilus), der Brachs-men (Abramis brama), die Rotfeder (Scardinius erythrophthalmus),die Schleie (Tinca tinca), die Blikke (Blicca björka) und die Barbe(Barbus barbus) werden. Gerade die Barbe ist indes kein typischerKarpfenfisch. Sie bevorzugt sauerstoffreiche, klare, grosse Fliess-gewässer mit Sand- oder Kiesgrund, wie es für die nahe Aarezutrifft. Barben leben meist gesellig und sind vorwiegend däm-merungs- und nachtaktiv. Zwischen Mai und Juli ziehen sie in gros-sen Schwärmen zur Fortpflanzung an flachen, kiesigen Stellen imströmenden Wasser flussaufwärts. Gefangen werden die Tiere mitNetzen, Reusen und Angeln. Ihr Fleisch ist zwar grätenreich, abersehr wohlschmeckend, wie auch Ausonius im 4. Jahrhundert in sei-nem Epos auf die Mosel vermerkt2083: „Schmackhafter bist du,kommst du zu Jahren; von allem, was atmet, ward dir allein einAlter gegönnt, dem das Lob nicht versagt ist“.
Das Rotauge ist einer der häufigsten Fische in stehenden undlangsam fliessenden Gewässern. Rotaugen werden heute nichtals Speisefische geschätzt. Im 19. Jahrhundert wurden sie teil-weise als preiswerte Speisefische verkauft, und aus dem Mittel-alter liegen historische Quellen vor, die besagen, dass kleineFische unterschiedlichster Art zu einem stärkenden Trankzubereitet wurden2084.
Einen ähnlichen Lebensraum wie das Rotauge benötigt dieRotfeder; sie bevorzugt zudem Teiche mit weichem Grund, alsoGewässer, die eher als eutrophiert, also nährstoffreich zubezeichnen sind. Als Speisefisch wird sie heute wenig geschätzt,ist aber als Speiserest durchgehend vom Neolithikum bis zumMittelalter nachgewiesen.
Einen weiteren Hinweis auf eutrophe Gewässer mit weichemGrund in der Nähe der Villa erhalten wir im Nachweis derSchleie. In diesen warmen Gewässern hält sie sich vorzugsweisein der Nähe des Schlammgrundes auf und ernährt sich von klei-nen Bodentieren und verrottenden Pflanzenresten. Es ist leichtnachzuvollziehen, dass diese Fische eine „Nachbehandlung“brauchen, will man das zarte und wohlschmeckende Fleischgeniessen. Um dem Fleisch den moderigen Geschmack zu ent-ziehen, werden die Tiere heute noch vor dem Verzehr einige Tagein Frischwasser gehältert.
Auch der Brachsmen und die Blicke bewohnen nährstoff-reiche Seen oder langsam fliessende Gewässer. Beide Arten sindsich in ihrem äusseren Habitus zum Verwechseln ähnlich. AlsSpeisefische sind sie heute wegen ihres grätenreichen Fleischeswenig geschätzt.
Insgesamt weisen also vier der sechs nachgewiesenen Artenaus der Familie der Cyprinidae auf mehr oder weniger eutro-phierte Stillgewässer hin, die, wie die Flurnamen belegen (Kap.18.2.2), zahlreich in der näheren und weiteren Umgebung an-
zutreffen waren, einer Gewässerart, die heute wie in römischerZeit nicht als bevorzugtes Fischereiareal einzustufen ist.
Der grösste Anteil (53%) der nachgewiesenen Fische gehörtzur Familie der Lachsartigen (Salmonidae). Kennzeichnend fürdiese ist ihr Lebensraum in kalten, sauerstoffreichen und klarenGewässern. Im Gegensatz zu den anderen Fischen liegt ihreLaichzeit meist in den Herbst- und Wintermonaten. Das Fleischist ausgesprochen schmackhaft, dementsprechend sind alleSalmonidenarten als Speisefische hoch geschätzt. Die Bestim-mung der einzelnen Arten kann Schwierigkeiten bereiten, denndiese Fischgruppe neigt dazu, lokale Rassen und Formen zu bil-den. Entsprechend hoch ist deswegen in der Regel der Anteil anKnochen, die zwar der Familie, nicht aber einer einzelnen Artzugeordnet werden kann (hier n=13). Eindeutig vertreten sindgrosse Salmoniden. Dabei dürfte es sich entweder um den Lachs(Salmo salar) oder aber die Seerforelle (Salmo trutta trutta) han-deln. Beide Arten sind nahe verwandt und anhand der Mor-phologie der Skelettelemente nur schwer zu trennen. Die ent-sprechenden Brust- und Analflossenstrahlen stammen ausdem Gebäude O und einer der Gruben beim Tor.
Ferner sind die Bachforelle (Salmo trutta f. fario) und dieAesche (Thymallus thymallus) zu nennen. Bei der Bachforellehandelt es sich um eine stationäre Zwergform der Forelle, ihreLänge übersteigt in nährstoffarmen Fliessgewässern eine Längevon 25 cm häufig nicht. Natürlicherweise sind Bachforellen inschnell fliessenden Bächen und Flüssen beheimatet, wie sie inder Umgebung der Villa zu finden waren (Kap. 18.2.2). In Bibe-rist sind Bachforellen mit 24 Resten vertreten.
Auch die Aesche benötigt eine relativ hohe Fliessgeschwin-digkeit des Wohngewässers, im Gegensatz zu den anderen Sal-moniden werden zum Laichen aber bis 24ºC toleriert. Folge-richtig laichen Aeschen als einzige der heimischen Lachsartigennicht im Winter, sondern zwischen März und April. 15 Wirbelund Schuppen weisen den Verzehr in der Villa nach.
Mit den hier vorgestellten Arten dürfte die Fischfauna derUmgebung der Villa gut repräsentiert sein. Dennoch fehlt einewichtige Art: der Hecht (Esox lucius). Da er nach Ausweis dernachgewiesenen Arten, insbesondere der eutrophilen Cyprini-den, sicherlich in den Gewässern des Umlandes präsent war, istdie Abwesenheit dieses Raubfisches als Hinweis auf selektivesFischen zu werten. Dies gilt vor allem, weil die Art auch in ande-ren römerzeitlichen Ensembles fehlt oder aber im Vergleich zuallen anderen Epochen unterrepräsentiert ist. Möglicherweiseist diese Besonderheit auf eine „kulturelle Ächtung“ des Hech-tes zurückzuführen. Ausonius wertet den Hecht als Speisefischim Versepos „Mosella“ folgendermassen2085:
„Stille Gewässer bewohnend und quakenden Fröschen zum Schrecken,
waltet auch, lachhaft benannt mit lateinischem Namen, der Hecht hier,
Lucius, als Herr über Tümpel, Schlamm und Riedgras verdunkeln.
Ihn wird niemand erwählen fürs Mahl an erlesener Tafel;
nur in verräucherter, miefender Kneipe mag man ihn kochen.“
Kap. 30.5
2083 Ausonius, Mosella 91.117.134.2084 19. Jahrhundert: Baumann 1993, 13; Mittelalter: Schibler/Hüster-Plogmann1996, 81.2085 Ausonius, Mosella 120-124.
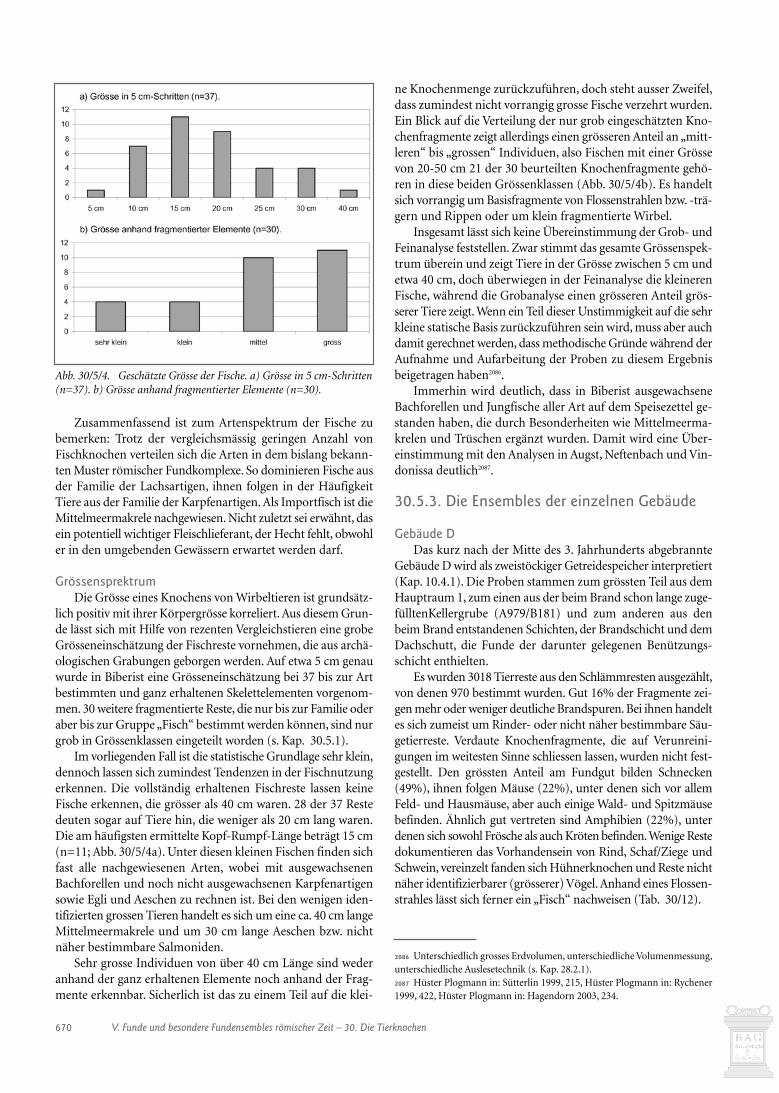
Zusammenfassend ist zum Artenspektrum der Fische zubemerken: Trotz der vergleichsmässig geringen Anzahl vonFischknochen verteilen sich die Arten in dem bislang bekann-ten Muster römischer Fundkomplexe. So dominieren Fische ausder Familie der Lachsartigen, ihnen folgen in der HäufigkeitTiere aus der Familie der Karpfenartigen. Als Importfisch ist dieMittelmeermakrele nachgewiesen. Nicht zuletzt sei erwähnt, dasein potentiell wichtiger Fleischlieferant, der Hecht fehlt, obwohler in den umgebenden Gewässern erwartet werden darf.
GrössensprektrumDie Grösse eines Knochens von Wirbeltieren ist grundsätz-
lich positiv mit ihrer Körpergrösse korreliert. Aus diesem Grun-de lässt sich mit Hilfe von rezenten Vergleichstieren eine grobeGrösseneinschätzung der Fischreste vornehmen, die aus archä-ologischen Grabungen geborgen werden. Auf etwa 5 cm genauwurde in Biberist eine Grösseneinschätzung bei 37 bis zur Artbestimmten und ganz erhaltenen Skelettelementen vorgenom-men. 30 weitere fragmentierte Reste, die nur bis zur Familie oderaber bis zur Gruppe „Fisch“ bestimmt werden können, sind nurgrob in Grössenklassen eingeteilt worden (s. Kap. 30.5.1).
Im vorliegenden Fall ist die statistische Grundlage sehr klein,dennoch lassen sich zumindest Tendenzen in der Fischnutzungerkennen. Die vollständig erhaltenen Fischreste lassen keineFische erkennen, die grösser als 40 cm waren. 28 der 37 Restedeuten sogar auf Tiere hin, die weniger als 20 cm lang waren.Die am häufigsten ermittelte Kopf-Rumpf-Länge beträgt 15 cm(n=11; Abb. 30/5/4a). Unter diesen kleinen Fischen finden sichfast alle nachgewiesenen Arten, wobei mit ausgewachsenenBachforellen und noch nicht ausgewachsenen Karpfenartigensowie Egli und Aeschen zu rechnen ist. Bei den wenigen iden-tifizierten grossen Tieren handelt es sich um eine ca. 40 cm langeMittelmeermakrele und um 30 cm lange Aeschen bzw. nichtnäher bestimmbare Salmoniden.
Sehr grosse Individuen von über 40 cm Länge sind wederanhand der ganz erhaltenen Elemente noch anhand der Frag-mente erkennbar. Sicherlich ist das zu einem Teil auf die klei-
ne Knochenmenge zurückzuführen, doch steht ausser Zweifel,dass zumindest nicht vorrangig grosse Fische verzehrt wurden.Ein Blick auf die Verteilung der nur grob eingeschätzten Kno-chenfragmente zeigt allerdings einen grösseren Anteil an „mitt-leren“ bis „grossen“ Individuen, also Fischen mit einer Grössevon 20-50 cm 21 der 30 beurteilten Knochenfragmente gehö-ren in diese beiden Grössenklassen (Abb. 30/5/4b). Es handeltsich vorrangig um Basisfragmente von Flossenstrahlen bzw. -trä-gern und Rippen oder um klein fragmentierte Wirbel.
Insgesamt lässt sich keine Übereinstimmung der Grob- undFeinanalyse feststellen. Zwar stimmt das gesamte Grössenspek-trum überein und zeigt Tiere in der Grösse zwischen 5 cm undetwa 40 cm, doch überwiegen in der Feinanalyse die kleinerenFische, während die Grobanalyse einen grösseren Anteil grös-serer Tiere zeigt. Wenn ein Teil dieser Unstimmigkeit auf die sehrkleine statische Basis zurückzuführen sein wird, muss aber auchdamit gerechnet werden, dass methodische Gründe während derAufnahme und Aufarbeitung der Proben zu diesem Ergebnisbeigetragen haben2086.
Immerhin wird deutlich, dass in Biberist ausgewachseneBachforellen und Jungfische aller Art auf dem Speisezettel ge-standen haben, die durch Besonderheiten wie Mittelmeerma-krelen und Trüschen ergänzt wurden. Damit wird eine Über-einstimmung mit den Analysen in Augst, Neftenbach und Vin-donissa deutlich2087.
30.5.3. Die Ensembles der einzelnen Gebäude
Gebäude D Das kurz nach der Mitte des 3. Jahrhunderts abgebrannte
Gebäude D wird als zweistöckiger Getreidespeicher interpretiert(Kap. 10.4.1). Die Proben stammen zum grössten Teil aus demHauptraum 1, zum einen aus der beim Brand schon lange zuge-fülltenKellergrube (A979/B181) und zum anderen aus denbeim Brand entstandenen Schichten, der Brandschicht und demDachschutt, die Funde der darunter gelegenen Benützungs-schicht enthielten.
Es wurden 3018 Tierreste aus den Schlämmresten ausgezählt,von denen 970 bestimmt wurden. Gut 16% der Fragmente zei-gen mehr oder weniger deutliche Brandspuren. Bei ihnen handeltes sich zumeist um Rinder- oder nicht näher bestimmbare Säu-getierreste. Verdaute Knochenfragmente, die auf Verunreini-gungen im weitesten Sinne schliessen lassen, wurden nicht fest-gestellt. Den grössten Anteil am Fundgut bilden Schnecken(49%), ihnen folgen Mäuse (22%), unter denen sich vor allemFeld- und Hausmäuse, aber auch einige Wald- und Spitzmäusebefinden. Ähnlich gut vertreten sind Amphibien (22%), unterdenen sich sowohl Frösche als auch Kröten befinden.Wenige Restedokumentieren das Vorhandensein von Rind, Schaf/Ziege undSchwein, vereinzelt fanden sich Hühnerknochen und Reste nichtnäher identifizierbarer (grösserer) Vögel. Anhand eines Flossen-strahles lässt sich ferner ein „Fisch“ nachweisen (Tab. 30/12).
V. Funde und besondere Fundensembles römischer Zeit – 30. Die Tierknochen
2086 Unterschiedlich grosses Erdvolumen, unterschiedliche Volumenmessung,unterschiedliche Auslesetechnik (s. Kap. 28.2.1).2087 Hüster Plogmann in: Sütterlin 1999, 215, Hüster Plogmann in: Rychener1999, 422, Hüster Plogmann in: Hagendorn 2003, 234.
670
Abb. 30/5/4. Geschätzte Grösse der Fische. a) Grösse in 5 cm-Schritten(n=37). b) Grösse anhand fragmentierter Elemente (n=30).

671
Alle untersuchten Proben weisen einen relativ gleichmässi-gen Anteil der schon „obligaten“ Schnecken, Mäuse und Amphi-bien auf. Die Rinderknochen stammen aus dem Dachschutt, woder Rumpf eines Rindes zum Vorschein gekommen ist, und ausder beim Umbau des Gebäudes an der Wende zum 3. Jahrhun-dert verfüllten Kellergrube, wo mehrere Rinderschädelhälftenmitvergraben worden sind (Kap. 30.2.3)2088. Aus dem Dachver-sturz wurden weiter fünf Hühnerreste und der Flossenstrahleines Fisches geborgen, aus der Brandschicht kommen die übri-gen Vogelknochenfragmente, sowie sechs Schweine- und einSchaf- bzw. Ziegenrest2089. Während die Rinderknochen zudem erwähnten Skelett und den Schädelhälften gehören dürf-ten, handelt es sich bei den übrigen Säuger- und Vogelknochenwohl um eher zufällig eingetragene Speisereste. Ihr geringerAnteil am Fundmaterial unterstützt im Zusammenhang mit dergrossen Zahl der verschiedenen nachgewiesenen Mäuse dieInterpretation des Gebäudes D als Getreidespeicher. Möglicher-weise sind die ebenfalls sehr gut vertretenen Schnecken imZusammenhang mit der Lagerung von Frischobst im Speicherzu sehen (s. Kap. 28.5.2).
Gebäude EDas kurz nach der Mitte des 3. Jahrhunderts abgebrannte
Gebäude E wird als Wohntrakt gedeutet, vielleicht als Sitz desIntendanten des Getreidespeichers D (Kap. 10.4.3). Die fünf Pro-ben aus diesem Gebäude stammen hauptsächlich aus einerGrube (A035/B236) innerhalb des Gebäudes und einer Werk-grube (A049/B228) ausserhalb des Gebäudes. Sie enthielten ins-gesamt 1509 tierische Reste, von denen 325 bestimmbar sind.Die Fragmente sind relativ gut erhalten, nur eines zeigt Brand-spuren. Es wurden keine verdauten Knochen identifiziert. DerAnteil der Amphibien im Material liegt bei gut 40%. Wie inGebäude D waren auch hier sowohl Frösche als auch Kröten fest-stellbar; unter den handaufgelesenen Knochen befindet sich einBiber (Kap. 30.2.4). Ausserdem fanden sich 33% Nagetiere, unterdenen hauptsächlich Haus- und Feldmaus vermutet werdendürfen. Allerdings ist auch der Rest einer Schermaus unter denNagerknochen. Bemerkenswert ist der Fund des Zahnfragmenteseines Tieres aus der Familie der Gliridae. Es dürfte sich um denRest eines Siebenschläfers oder Gartenschläfers handeln(Kap. 30.5.2). Der Anteil der Haustiere liegt in Gebäude E bei11%, sie werden durch Schweine (9%) und Schaf/Ziegen (2%)repräsentiert. Die gleichen Arten sind schon bei den han-daufgenommenen Knochen durch einen bedeutenden Anteilaufgefallen (Kap. 30.3.4). Einzelfunde belegen ferner einennicht näher zu identifizierenden kleinen Carnivor und mut-masslich einen Hasen (drei Oberkieferzähne). Vögel sind mitgut 3% im Material vertreten. Im Unterschied zu Gebäude Dfanden sich neben Resten von Hühnern auch solche von Fin-ken. Schliesslich sind die Fische in Gebäude E mit knapp 8%der bestimmbaren Reste beteiligt. Mit knapp 5% sind die Kar-pfenartigen (Cyprinidae) unter den Fischen am besten ver-treten. Unter ihnen war als einzige Art die Barbe (Barbus bar-bus) auszumachen. Die Fische sind in einer Grösse von 20-40 cm verzehrt worden. Analog zu den Cypriniden sind unter1,5% Lachsartigen (Salmonidae) nur Aeschen (Thymallusthymallus) zu nennen. Einzelfunde belegen das Egli (Perca flu-viatilis; Tab. 30/12).
Die beiden Gruben sind durch das Vorhandensein vonAmphibien-, Mäuse-, Schaf/Ziegen- und Schweineknochencharakterisiert. Die Fische, Hühner, Singvögel und der Schläfersind hingegen nur in der Werkgrube (A049/B228) ausserhalbbelegt. Da etwa gleich viele Knochen aus beiden Gruben vor-liegen (n=532 bzw. n=655) ist dieser Unterschied kaum durchdie statistische Grundlage zu erklären. Beide Gruben enthieltensicher Speisereste. Diese deuten aufgrund ihrer Zusammenset-zung auf eine qualitativ gute Ernährung hin. Allerdings ist derrelativ grosse Anteil Karpfenartiger in mittlerer Grösse nicht alsZeichen einer exklusiven Küche zu werten. Dass es sich beiGebäude E um einen Wohntrakt handelt, steht aber auch nachBetrachtung der Kleintierreste ausser Frage.
Holzbau JDer am Ende des 1. Jahrhunderts abgebrannte Holzbau J
dokumentiert eine der frühesten fassbaren Phasen in Biberist(Periode II-Phase 1). Er diente als Sitz des Villenverwalters, des-sen Wohntrakt im Erdgeschoss vermutet wird (Kap. 10.3.5). ImObergeschoss befand sich ein grosser Lagerraum, wo Getreide,sicher Gerste gelagert wurde (s. auch Kap. 28.5.5). Die Probenenthielten 1629 tierische Reste, von denen allerdings nur 161(Tab. 30/12) bestimmbar sind. Verantwortlich für diesen gerin-gen Bestimmungsgrad ist der hohe Anteil calcinierter Knochen,die einem Feuer von mehr als 550°C ausgesetzt gewesen seindürften. Proben aus der Brandschicht sind zu 38% calciniert, dieReste aus dem Brandschutt zeigen einen Anteil von 29% hochverbrannter Knochen. Aus der Grube (A099/B109.50) im nord-westlichen Teil des Hauses wurden lediglich vier Knochensplittergeborgen, die nicht verbrannt sind2090.
Bei der Betrachtung der relativen Anteile der vertretenenTierarten fällt zunächst auf, dass keine Schnecken und kaumAmphibienreste (knapp 4%) anfallen. Die (stark verbrannten)Mäusereste erreichen dafür einen Rekordanteil von 75%! Dergrösste Teil, nämlich 84 der insgesamt 121 Mäusereste stammtaus der Brandschicht. Zwei Proben aus der Küche sind hierbesonders reich an Mäuseknochen (n=54)2091.
Unter den 18 identifizierten Haustierresten dominierenSchweineknochen, drei Fragmente stammen von Schaf oderZiege. Von 12 Vogelknochen stammen fünf von Singvögeln, dierestlichen sieben sind nicht näher zuzuordnen. Vier Knochen-fragmente sind als Fischreste bestimmt worden. Eine Schuppeweist auf die Aesche (Thymallus thymallus), die drei verblei-benden Fragmente stammen von Karpfenartigen. Unter ihnenweisen ein Schlundknochen und ein Wirbel auf jeweils ein 20 cmgrosses Exemplar einer Schleie (Tinca tinca) und eines Brachs-men (Abramis brama) hin. Ein weiteres Wirbelfragment ist nurbis zur Familie der Karpfenartigen zu bestimmen.
Wenn damit insgesamt auch nur wenige Speisereste vorlie-gen, weisen zumindest die Singvögel auf ein mediterran gepräg-tes Essverhalten, das auf ein Mindestmass an Qualitätsbewusst-sein schliessen lässt.
Kap. 30.5
2088 Inv.-Nr.14/9/978; 14/9/1066.2089 Inv.-Nr.14/9/980; 14/9/922.2090 Inv.-Nr.14/9/1899.2091 Inv.-Nr.14/9/2031; 14/9/2032.

Steinbau JAus dem nach dem Brand des Holzbau J an gleicher Stelle
wieder errichteten Steinbau J, der noch immer als Sitz des Vili-cus diente (Kap. 10.3.6), liegen lediglich 80 bestimmbare Tier-reste aus den Schlämmproben vor (Tab. 30/12). Sie zeigen in derZusammensetzung Ähnlichkeiten mit jenen aus dem HolzbauJ: So sind keine Schnecken und nur wenige Amphibienreste aus-zumachen (4%), der Anteil an Mäuseartigen beträgt 68%.Unter den 14 Haustierresten dominieren Schweinknochen, beiden Vögeln ist ein Huhn nachgewiesen. Die Fischreste deutenwiederum auf mehr Karpfenartige als Lachsartige, wobei eineBasis von fünf Skelettelementen keine wertende Aussage zulässt.
Gebäude O und UmgebungDas vierphasige Gebäude O, ein Landarbeiterhaus, wurde im
zweiten Viertel des 2. Jahrhunderts erbaut, während seiner Nut-zung mehrfach umgebaut und fiel kurz nach der Mitte des3. Jahrhunderts einem Brand zum Opfer (Kap. 10.3.10). Insge-samt erbringen die Proben 2205 Knochenfragmente aus denSchlämmproben, von denen 505 bestimmt werden konnten.
Phase 1: Von 683 Fragmente waren 127 artlich zuzuordnen(Tab. 30/12). Der grösste Teil dieser Reste (n=413) stammt auseiner Grube (A110/B050), die in einem um die Mitte des 2. Jahr-hunderts abgebrannten Unterstand westlich des Gebäudes lag,wo offensichtlich Getreide zwischengelagert und vielleichtauch geworfelt wurde (Kap. 28.5.7). Unter den 58 bestimmba-ren Fragmenten finden sich fast ausschliesslich Nagerknochen.Beim grössten Teil dürfte es sich auch hier um Haus- und Feld-mäusen handeln. Wald- und Spitzmäuse sind sporadisch ver-treten. Eine Tibia ist allerdings einer Hausratte (Rattus rattus)zuzuordnen. Neben den Nagerresten finden sich lediglich sie-ben Knochen von Schweinen, Schafen (Ziegen) und Amphibien.27% aller Reste liegen calciniert und damit hochverbrannt vor,was im Einklang mit einem Brand des Unterstandes steht.
Die Zusammensetzung der Tierarten in der Grube (A110/B050) stützt die Hypothese, der Unterstand westlich des Gebäu-des O könnte als Getreidereinigungsplatz gedient haben, und beiden Funden aus der Grube handele es sich um Getreidevorräte,die zum Zeitpunkt des Brandes hier zwischengelagert waren. Diegrosse Zahl der Mäuse (und nicht zuletzt der Ratten) und die nursporadisch auftretenden Speisereste wären so gut zu erklären.
Aus dem Bereich der Feuerstelle innerhalb des Gebäudessind – neben den obligaten Mäusen – vergleichsweise deutlichund variationsreich Speisereste belegt. Unter den Vögeln findensich Hühnerknochen neben Eierschalen und Singvogelknochen.Fische sind durch eine ganze Reihe von Arten belegt. Die Kar-pfenartigen werden durch einen Brachsmen und eine Rotfederrepräsentiert. Erstmals treten in Biberist mehrere Salmonidenauf: Neben zwei nicht näher bestimmten Exemplaren sind zweiBachforellen mittlerer Grösse (20 cm bzw. 25 cm) und eine30 cm grosse Aesche nachgewiesen. Des Weiteren fand sich einSchädelfragment einer Mittelmeermakrele2092. Insgesamt ist mitdiesen Resten auf eine qualitätsvolle und hochwertige Speisen-zubereitung zu schliessen.
Phase 2: Es liegt eine ähnliche Speiserestzusammensetzungvor. Aus dem Feuerstellenbereich und dem Kiesboden innerhalbdes Gebäudes stammen 60 bestimmbare tierische Reste (n=251).Wiederum sind zumeist Mäuse zu identifizieren (n=29). Dane-
ben finden sich sieben Fragmente von Schweineknochen und einSchaf(Ziegen)rest, der Oberarm eines amselartigen Vogels und18 Fischreste. Neben einem nicht näher bestimmbaren Kar-pfenartigen dominieren – wie in Phase 1 – Salmoniden: SechsReste von Bachforellen in einer Grösse zwischen 10 cm und25 cm sowie vier Aeschenreste dürften bei der Zubereitung vonSpeisen in den Küchenboden gelangt sein.
Phase 3: Insgesamt wurden 727 Knochenfragmente ausge-zählt, 236 davon sind bestimmbar. Im Vergleich zu den voran-gehenden Phasen bleibt die Artenzusammensetzung ähnlich,allerdings erfährt die Erhaltung der Knochen eine Verschlech-terung. 28% der Funde aus dem Schlämmmaterial zeigt ober-flächliche Auflösungserscheinungen; die Funde lagen, obwohlüberdeckt von den Schichten der Phase 4, relativ nahe der rezen-ten Oberfläche (s. Abb. 10/O5). Die Tierreste stammen aus einerGrube (A063/B121) sowie aus Brandschichten der Feuerstellenund Kiesböden im Gebäude. Eine unterschiedliche Zusammen-setzung der Reste kann nicht beobachtet werden, weshalb dieFragmente gemeinsam besprochen werden.
Wie auch im Steinbau J sind in allen Phasen des GebäudesO kaum Amphibienreste geborgen worden, dagegen sind Mäuseder unterschiedlichsten Arten überall präsent. Ihr Anteil ambestimmbaren Material beträgt in Phase 3 61% (Tab. 30/12).Ebenfalls omnipräsent, wenn auch in geringer Zahl, sind Über-reste von Hausratten.
Bei den Haustierresten handelt es sich vorrangig um solchevon Hausschweinen (n=28), die Wiederkäuer sind sporadischvertreten. Unter den 9 Vogelresten war eine erste Phalange ausdem Flügel eines Huhnes bestimmbar. 24 Fischreste repräsen-tieren das Egli (n=4), aus der Familie der Karpfenartigen eineBarbe, einen Brachsmen sowie drei nicht näher zu bestimmendeIndividuen. Salmoniden sind ebenfalls mit fünf Fragmenten ver-treten. Zwei Reste sind von Bachforellen, ein Flossenstrahlfrag-ment stammt von einem grossen Salmoniden, bei dem es sichum einen Lachs oder eine Seeforelle handeln könnte. Phase 3 desGebäudes O barg in der Brandschicht einen weiteren Rest einerMittelmeermakrele2093. Es handelt sich um ein Caudalwirbel-fragment eines etwa 40 cm grossen Tieres. Tiere dieser Grössesind mit grosser Wahrscheinlichkeit in Salz konserviert aus demMittelmeerraum in die Provinzen gelangt und waren offenbarregelmässiger Bestandteil eines ausgewogenen Speiseplans.
Phase 4: Aus der letzten Phase, die mit dem Brand des Ge-bäudes O endete, wurden 12 Proben aus der Brandschicht, demDachschutt und dem Horizont des Trockenraums untersucht.Sie enthielten mit 15 bis maximal 80 Fragmenten je Probe nurwenige tierische Reste. Unterschiede zwischen den Proben sindnicht aufgefallen. Von den insgesamt geborgenen 544 Frag-menten sind 82 bestimmbar. Hintergrund für den geringen Be-stimmungsgrad mögen in der Konsistenz der Fragmente liegen:22% der Knochen sind stark verbrannt, 12% eher schlecht erhal-ten. Grundsätzliche Veränderungen im Vergleich zu den vor-angehenden Phasen sind nicht zu erkennen. Allerdings sind nurnoch zwei Fischreste, die je einen Cypriniden und einen Sal-moniden repräsentieren, bestimmt worden.
V. Funde und besondere Fundensembles römischer Zeit – 30. Die Tierknochen
2092 Inv.-Nr.14/9/3179.2093 Inv.-Nr.14/9/2835.
672

673
Zusammenfassend enthält das Gebäude O aus der Sicht derKleintierreste das weitaus interessanteste Inventar. Die Zusam-mensetzung der Vögel- und Fischreste verleitet zu dem Schluss,dass im Wohntrakt während der gesamten Nutzung qualitativhochwertig gekocht wurde (s. auch Kap. 30.2.9). Die Verteilungder Skelettelemente zeigt zu etwa gleichen Teilen Wirbel undeigentliche Zubereitungsreste wie Schuppen und Flossenstrah-len. Wenn wir die Wirbel mit Speiseresten im engeren Sinnegleichsetzen, überwiegen die Speisereste bei weitem. Dennochmögen besonders die leicht vergänglichen Schuppen darauf hin-weisen, dass bei der Fischzubereitung in der Küche Abfälle die-ser Art in grösserer Menge anfielen.
Gebäude QDas nicht vollständig erfasste Gebäude Q wurde zu Beginn
des mittleren Drittel des 3. Jahrhundert an das LandarbeiterhausO angebaut (Kap. 10.3.12). Auffällig ist hier ein zentraler Kon-servierungsofen, der von Gebäude O aus zu befeuern war undin der vermutlich Fleisch zur Vorratshaltung verarbeitet wurde(Kap. 30.2.11). Die meisten Proben stammen aus den zu dieserAnlage gehörenden Kanälen. Ferner wurde eine Grube (A125/B044) in der Mittelachse des Gebäudes beprobt2094. Der Unter-schied in der Tierartenzusammensetzung der Grube und derOfenkanäle ist trotz der geringen Anzahl der Fragmente aus derGrube deutlich. Insgesamt wurden aus der Grube 170, aus denKanälen 2874 Knochenfragmente ausgelesen. Unter den Restender Grube waren 17 Elemente bestimmbar. Fünf von ihnen sindim weitesten Sinne als Speisereste zu bezeichnen. Es handelt sichum vier Fischschuppen (drei nicht weiter bestimmbare Fische,eine Aesche) und einen Unterkieferzahn eines Rindes. Bestimm-bar waren ferner acht Mäuse, drei Amphibien und ein Maulwurf.Während wir es also mit „normalen“ Relationen für die Villa vonBiberist-Spitalhof zu tun haben, zeigen die tierischen Reste ausdem Bereich des Konservierungsofens einen völlig andereCharakter: Der Bestimmungsgrad der Knochenreste aus denKanälen beträgt ungewöhnliche 86(!)%. Dieser erstaunlichhohe Anteil erklärt sich aus der Tatsache, dass 70% der Knochenunfragmentierte Amphibienreste darstellen. Hinzu kommen ca.20% ebenfalls nur wenig zerbrochene Mäusereste und 10%Schnecken. Als Speisereste sind nur Einzelfunde vom Schweinund von Vögeln inklusive einem Schalenfragment eines Hüh-nereies zu werten. Demnach dürfte der grösste Teil der tierischenReste eine Fauna repräsentieren, die sich nach Auflassung desKonservierungsofens bzw. nach dem Brand des Gebäudes Q inden noch hohlen Kanälen des Ofens etabliert hat2095. Zu dieserFauna wäre auch der aus dem handaufgelesenen Knochenma-terial bestimmte Rest einer Hausratte zu zählen (Kap. 30.2.11).In Übereinstimmung mit den botanischen Ergebnissen (Kap. 28.5.8) zeigen auch die Kleintierreste in Gebäude Q kaum Speise-reste und lassen eher auf ein ökonomisch genutztes Gebäudeschliessen.
Gruben am TorIm Bereich des Hoftores wurden innerhalb der Hofmauer
zwei Gruben (A118/B072; A116/B075) beprobt. Die wahr-scheinlich holzverschalten Gruben müssen beim Zerfall der Hof-mauer im 3. Jahrhundert noch offen gewesen sein (Kap. 10.1.1).Die Gruben zeigen vom Kleintierspektrum her jeweils eine völ-
lig unterschiedliche Zusammensetzung. Die direkt an der inne-ren südlichen Seite des Tores gelegene Grube (A116/B075) mageinen Bezug zum Wachraum über dem Tor gehabt haben. Sieentspricht in der Artenzusammensetzung den bislang unter-suchten Proben der Villa: Zwar sind von den 53 geborgenenKnochenresten mit 33 Fragmenten ein grosser Anteil bestimm-bar, doch handelt es sich in den meisten Fällen um Reste vonHaus- oder Feldmäusen (n=25). Weiter finden sich zwei unbe-stimmbare Vogelknochen, zwei Reste von Amphibien, einUnterkieferzahnfragment eines Carnivoren (Hund?) und derFlossenstrahl eines nicht bestimmbaren Fisches sowie derSchlundknochen eines Rotauges. Damit ist der Anteil an Spei-seresten in dieser Grube minimal und widerspiegelt eher dentäglichen Kampf gegen die offenbar überall präsenten Nager aufdem Gut.
Die nur zwei Meter entfernte Grube (A118/B072) lagunmittelbar nördlich des Landarbeiterhauses O und war eben-falls an der Hofmauer situiert. Aus den Schlämmfunden wur-den 378 Fragmente ausgelesen, von denen 45% (n=167)bestimmbar sind. Alle Reste sind gut erhalten, 14 Fischwirbel zei-gen deutliche Spuren von Verdauung. Typischerweise ist derQuerschnitt solcher Wirbel nicht mehr rund, sondern gepresstoval, zudem wirken sie in ihrer Längsachse gestaucht. Demnachist hier mit Fäkalien zu rechnen. Es könnte sich sozusagen umdie nahe beim Tor gut gelegene, zentrale Latrine des Gesindeshandeln, wo der wertvolle menschliche Dung gesammelt wurde(s. Kap. 18.2.4 mit Anm. 640).
Unter den bestimmbaren Resten dominieren nicht, wie inallen anderen Proben, Nager und/oder Amphibien. Sie machenzusammen nur knapp 11% (n=14 bzw. n=4) des Materials aus.Vielmehr sind mit 43% in erster Linie Reste vom Schwein ver-treten. Es handelt sich fast ausschliesslich um Zehenglieder derSchweinsfüsse und um kleine Hand- bzw. Fusswurzelknochen.Weitere 24% stammen von Vögeln. Darunter finden sich achtFragmente von Flügelspitzen von Hühnern. 21 Fragmente vonRöhrenknochen oder Wirbeln sind artlich nicht bestimmbar,aufgrund ihrer Grösse müssen sie jedoch von grösseren Vögelnstammen. Es ist demnach nicht abwegig, hier ebenfalls Reste vonHausgeflügel zu vermuten. Weitere zehn Knochenfragmente sindSingvögeln zuzuordnen. Es liegen Elemente aus nahezu allenKörperbereichen vor. Die Fische machen im Gegensatz zuallen anderen Proben auf dem Gut 21% der bestimmbarenFunde aus. 24 der 32 Fischreste weisen auf Tiere aus der Fami-lie der Lachsartigen. Neben der Bachforelle (n=15) finden sichAeschen (n=2) und grössere Salmoniden (n=7), unter denenLachse oder Seeforellen vermutet werden dürfen. Die beidenWirbel der Aeschen weisen auf Tiere von 15 cm Länge, währenddie Grösse der Bachforellen bei einer Variation von 10-20 cmeine mittlere Länge von 14 cm aufweist. Sechs Fragmentestammen von Karpfenartigen, unter ihnen sind der Brachsmenund die Blikke nachgewiesen. Keines der Fragmente lässt eineGrössenschätzung zu. Weiter fand sich je ein Caudalwirbel desEgli und der Trüsche. Beide Tiere hatten eine Totallänge von
Kap. 30.5
2094 Inv.-Nr.14/9/3298; 14/9/3300.2095 Das Sediment in den Kanälen war ausgesprochen fein und die Funde weit-gehend kleinteilig (Kap. 35.15); es dürfte erst allmählich hineingerieselt sein.

20 cm. Insgesamt überwiegen bei den Fischen Skelettelementeaus dem zentralen Rumpfbereich (20 Wirbel), der Bereich desKopfes (n=6) und des Flossensaumes (n=6) sind sehr viel sel-tener nachgewiesen. Schuppen lassen sich trotz guter Erhaltungnicht nachweisen. Die Übervertretung der Wirbel wird nochdeutlicher, vergegenwärtigt man sich die Zahlenverhältnisse derKnochen in den einzelnen Körperregionen eines Fisches: DasKopfknochen-Wirbelverhältnis beträgt etwa 4:1, die Zahl derFlossenträger und -strahlen variiert stark. Da jeder Flossenstrahleine Vielzahl von „Sollbruchstellen“ hat, die beim Zerfall denAnteil des Flossensaumes sehr schnell in die Höhe treiben kön-nen, mag nachvollziehbar sein, dass die Übervertretung der Wir-bel nicht die natürlichen Anteile der Körperteile widerspiegelt.Hier liegen Speise- und nicht Zubereitungsreste vor. Darüberhinaus legen die verdauten Wirbel den Nachweis der Speiseresteaus Fäkalien nahe. Auch wenn aus heutiger Sicht verwunderlichsein mag, dass Fischwirbel mit verzehrt wurden, belegen Kno-chenreste diese Praxis jedoch bis in die Neuzeit. Auch kleinereSkelettelemente von Geflügel und Haussäugern wurden gern mitverzehrt. In Anbetracht der Tatsache, dass nur ein Bruchteil derverdauten Kleinknochen tatsächlich sichtbare Spuren zeigt,mögen demnach die nachgewiesenen Knochen von Schweine-füssen, Flügelspitzen von Hühnern und Elemente aus dem Ske-lett der Singvögel ebenfalls in die Kategorie Speisereste fallen wiedie beschriebenen Fischwirbel.
Die Artenzusammensetzung in der Grube (A118/B072) ent-spricht mit ihrem überwiegenden Salmonidenanteil den bislangbekannten Verhältnissen in Augst, Neftenbach und Vindonissa(wie Anm. 2087). Innerhalb der Villa von Biberist lässt sich dieVerteilung am besten mit den Resten aus dem nahen GebäudeO parallelisieren. In die gleiche Richtung weist der vergleichs-weise hohe Vogel- und Schweineknochenanteil. Die handauf-genommenen Knochen zeigen ebenfalls Parallelen (Kap. 30.2.12).
30.5.4. Schlussbetrachtung
Die Rolle, die Kleintiere in den römischen Siedlungen, Mili-täranlagen und Gutshöfen spielten, sei es als Schädlinge, „Mit-bewohner“ oder als Nahrung ist bis heute noch relativ unbe-kannt. Indizienschlüsse sind zulässig, nur erfordern sie dasZusammentragen eines grossen Datenmaterials. Trotz grosserFortschritte in der Aufarbeitung und systematischen Bearbei-tung von Schlämmproben und ersten ermutigenden Ergebnis-sen stehen wir noch am Anfang, nämlich in der „Sammelpha-se“. Bis für alle Siedlungs- und Landschaftstypen und für allesozialen Gruppen in den römischen Provinzen eine „Kleintier-charakteristik“ erstellt sein wird, vergehen noch einige Jahre. Eskönnen aber immerhin für die Kleintiergruppe der Fische eini-ge erstaunliche Neuerungen aufgezeigt werden:
Die vor 15 Jahren noch als Sensation gewertete Identifizie-rung von Mittelmeermakrelen in den nördlichen Provinzengehört heute schon fast zum normalen Repertoire einer römi-schen Fundstelle. Zwar finden sich die Tiere noch immer ineinem Umfeld, das zumindest auf einen gehobenen Mittelstandschliessen lässt, doch es wird immer deutlicher, in welch gros-sen Mengen Makrelen aus dem Mittelmeerraum in die Pro-vinzen verhandelt worden sein müssen. Z.T. dürfte dies in den
allerorts zu findenden Fischsaucenamphoren geschehen sein,wenn sie kaum gesiebte Fischsauce (garum) minderer Qualitätoder den gesiebten Rückstand (allec)enthielten2096. Damitschwindet der Hauch von Exklusivität2097.
In den letzten Jahren verstärkt sich der Eindruck von einersehr selektiven Fischerei auch in den Provinzen, etwa im Gebietder heutigen Schweiz: So fehlt der Hecht oder er ist zumindeststark unterrepräsentiert. Die Fische aus der Familie der Lachs-artigen erfahren eine deutliche Zunahme in den meisten römi-schen Fundensembles. Diese Aufwertung geht zu Lasten derTiere aus der Familie der Karpfenartigen. Allerdings gibt es Aus-nahmen: Sie betreffen zum einen die wenigen Cypriniden, diein klaren, sauerstoffreichen Fliessgewässern leben, und zumanderen sehr kleine Individuen. Insgesamt nimmt der Konsumkleiner Fische zu. Auch tauchen neu Arten, die in vorrömischerZeit bedeutungslos waren, sporadisch bis regelmässig im Fund-gut auf. Als Beispiel sei der Aal genannt.
In die meisten dieser aufgezeigten Phänomene reiht sich dasFundmaterial von Biberist ein. Vermutlich würde das noch deut-licher, wenn die Datengrundlage besser wäre. Insgesamt ist dieAusbeute an Fischresten mit 132 Fragmenten so niedrig, dass anmethodische Probleme – welcher Art auch immer – gedachtwerden muss. Möglicherweise sind es aber auch Erhaltungs-probleme, die speziell die Fischreste benachteiligten oder aberdie Bewohner der Villa nutzten das Nahrungsmittel „Fisch“ tat-sächlich nur sehr verhalten. So klein die Datenbasis ist, bei derBetrachtung des Gesamtmaterials zeigt sich: Die Mittelmeer-makrele ist nachgewiesen, während Hechte im Material fehlen,und die Fische aus der Familie der Lachsartigen in den Arten-anteilen dominieren. Als „Exot“ kann die Trüsche betrachtetwerden. Sie ist zwar wohlschmeckend, wird aber relativ seltennachgewiesen. Weiterhin fällt auf, dass die Tiere aus der Fami-lie der Karpfenartigen grösstenteils aus Gewässern stammen, dierelativ stark eutrophiert waren. Arten wie die Schleie, derBrachsmen und die Blikke leben in Gewässern mit schlammi-gem Untergrund und starkem Pflanzenbewuchs. Hier ist an Wei-her oder flache Stillgewässer in unmittelbarer Umgebung derVilla zu denken, wie sie aufgrund der Flurnamen tatsächlich vor-handen gewesen sein müssen (Kap. 18.2.2). Gleichzeitig hat eskleine Fliessgewässer im und um den Gutshof gegeben, etwa derGutshofbach, die Bachforellen und Trüschen beherbergt habenkönnen. Diese Gewässer sind sauerstoffreich, nährstoffarmund kühl. Auch grössere Gewässer dieser Art, wie die nahe Aareoder die etwas weiter entfernte Emme, wurden offenbar befischt.Aeschen und Egli mögen hier gefangen worden sein, währendSeeforellen aus grösseren Seen, etwa dem Bielersee, stammenmögen.
Über zeitliche Entwicklungen in der Konsumation vonFischen kann keine Beurteilung abgegeben werden. Bei der Zu-sammenfassung von Zeithorizonten werden bei der kleinenDatengrundlage eher unterschiedliche Hausinventare gefasst alschronologische Entwicklungen. Innerhalb der Gebäude wäre nurGebäude O von Interesse, hier ist jedoch keine Entwicklung fass-bar.
V. Funde und besondere Fundensembles römischer Zeit – 30. Die Tierknochen
2096 Martin-Kilcher 1994a, 474-476; s. auch Kap. 21.3.4.2097 Ervynck u.a. 2003, 431.
674

675
Zur Einordnung der Gebäude ist zusammenfassend fol-gendes zu bemerken: Die Gebäude D und Q sind fast aus-schliesslich durch Reste von Amphibien, Nager und Schneckenzu charakterisieren. Beide Gebäude enthielten aus Sicht derKleintierknochen so gut wie keine Speisereste; die Interpreta-tion, es handele sich nicht um Wohngebäude, sondern umeinen Getreidespeicher bzw. ein Schlachthaus kann demnachunterstützt werden. Dagegen stellen Gebäude E, Holzbau J,Steinbau J und Gebäude O sicher Wohntrakte dar. In allenHäusern waren – neben erheblichen Anteilen von Nagetieren– Speisereste zu identifizieren. Unter den Haussäugern domi-nieren in allen Gebäuden Reste vom Schwein. Es wurde über-all Geflügel und die eine gute und mediterran geprägte Kücheanzeigenden Singvögel identifiziert. In Haus E, Holzbau J undSteinbau J, alles Häuser, in denen Personen der Villenverwal-tung ihren Sitz hatten, dominieren unter den Fischen Kar-pfenartige aus eutrophen Gewässern in einer mittleren Grös-se. Vom bisherigen Kenntnisstand her war der Verzehr dieserFische in römischer Zeit gesellschaftlich nicht sehr angesehen,wenn auch nicht geächtet, wie das beim Hecht wohl der Fallwar (Kap. 30.5.2).
Im Gegensatz dazu findet sich in Gebäude O, einem Land-arbeiterhaus, eine vergleichsweise grosse Zahl Lachsartiger undMittelmeermakrelen. Eine ähnliche Zusammensetzung derSpeisereste wie im Gebäude O findet sich in der nahen Grube(A118/B072). 4% der Speisereste liegen verdaut vor. Dies unddas identifizierte Skelettteilspektrum weist auf eine Konzen-tration von Fäkalien hin; die Grube dürfte als Latrine gedienthaben. Es ist die einzige Struktur der Villa, wo die grossen Men-gen an Nagetieren fehlen.
30.6. Verkohlte Masse aus dem Konservierungsofen in Gebäude Q
(JORGE SPANGENBERG)
30.6.1. Einleitung
Unter den Schlämmrückständen aus der Einfüllung desKanalsystems des Ofens in dem im früheren 3. Jahrhunderterrichteten Gebäude Q (Kap. 10.3.12) fanden sich mehrere ver-kohlte, amorphe Brocken2098. In der Hoffnung, Hinweise auf dasim Ofen verarbeitete Gut zu erhalten, haben wir sie Jorge E.Spangenberg, Universität Lausanne, zur Analyse übergeben.Diese ergeben, dass es sich um tierisches Fett von Wiederkäu-ern handelt.
30.6.2. Material and methods
A sample from “black carbon” was submitted for lipid cha-racterization at the Laboratories of Stable Isotopes of the Uni-versity of Lausanne (IMG-UNIL). The sample consists ofblack, dull, not very much hard coarse grained solid organic car-bon, and arrived wrapped with paper in a plastic box. Somemegascopically recognizable “not-organic” material was remo-ved by handpicking with dichloromethane-pre-washed forceps.The black carbon was homogenized and finely powderedmanually in an agate mortar.
Sample preparation The sample (619.4 mg) was refluxed with 150 ml of dichlo-
romethane (Fluka, Switzerland, glass distilled) for 4 days, withchange of solvent after the first 2 days.
The solvent (containing the extracted lipids) were combinedand reduced by roto-evaporation, and gently evaporated to dry-ness. The extracts were desulfurized with activated copper pow-der (24 h, room temperature). Separation and methylation ofthe fatty acids from the lipids extract was performed by alkali-ne hydrolysis with aqueous ethanolic potassium hydroxide, andextracted with hexane. The fatty acids were derivatized withmethanolic BF
3. to fatty acid methyl esters (FAMEs). The
FAMES were stored with 0.5 ml hexane in 2 ml vials with PTFE-lined cap at +4°C until gas chromatographic analysis2099.
Gas chromatography/mass spectrometry detection(GC/MSD)
Chemical characterization of the lipids was performedwith a Hewlett-Packard GC HP 6890 coupled to a HP 5973 qua-drupole mass selective detector. The system was equipped witha HP-FFAP fused silica capillary column (50 m × 0.20 mm i.d.)coated with polyethylene glycol-TPA modified as stationaryphase (film thickness 0.33 µm). Helium was used as carrier gas(1 ml/min flow rate), and the injection was made splitless manu-ally at a temperature of 200°C, in order to prevent potential iso-merization of the unsaturated fatty acids. After an initial peri-od of 2 min, the column was heated to 250°C at 5°C/min fol-lowed by an isothermal period of 20 min. The MSD was ope-rated in the electron impact mode at 70 eV, source temperatureof 250°C, emission current of 1 mA and multiple-ion detectionwith a mass range from 50 to 700 amu.
Isotopic analysis of individual fatty acids by GC/C/IRMS The compound specific carbon isotope analyses of the
FAMEs were obtained by the use of a Hewlett-Packard 6890 GCcoupled to a Finnigan MAT Delta S isotope ratio mass spectro-meter by a combustion (C) interface III (GC/C/IRMS) under acontinuous helium flow. The combustion interface is compri-sed of a ceramic furnace with copper oxide and platinum cata-lyst at a temperature of 940°C. A He-flushed Nafion membra-ne prevented water from reaching the ion source of the IRMS.The IRMS ion source pressure was 6×10-6 Bar. The GC was ope-rated with the same type of column and temperature programused for GC/MS analyses. The performance of the GC/C/IRMSsystem, including the GC and combustion furnace, was evalu-ated every 10 analyses by injection of a mixture of FAMEs ofknown δ13C values. The background subtraction and δ13C valueswere calculated using the ISODAT 7.4 software. The reprodu-cibility assessed from four replicate analyses of the samples ran-ged between ±0.1 and ±0.4‰. The accuracy of the GC/C/IRMSanalyses was monitored by coinjection of a FAME laboratorystandard of known isotopic composition. The stable carbon iso-tope ratios are reported in the delta (δ) notation as the per mil (‰)
Kap. 30.6
2098 Inv.-Nr.14/9/2807.2099 Spangenberg u.a. 1998; Spangenberg/Ogrinc 2001; Spangenberg/Dionisi2001.

deviations relative to the Vienna Pee Dee Belemnite limestone(VPDB) standard (δ13C ≡ 0‰): δ13C = [(R
sample- R
standard)/ R
standard] × 103 where R = 13C/12C.
The isotopic shift due to the carbon introduced in the fattyacid methylation was corrected by a mass balance equation:
δ13CFAME
= fFAδ13C
FA+ f
MeOHδ13C
MeOH where δ13C
FAMEδ13C
FA, and
δ13CMeOH
are the carbon isotope compositions of the fatty acidmethyl ester, the fatty acid, and the methanol used for methy-lation of the fatty acid, respectively, and f
FAand f
MeOHare the car-
bon fractions in the fatty acid methyl ester due to the underi-vatized fatty acid and methanol, respectively.
30.6.3. Results and discussion
The gas chromatogram of the fatty acids extracted from the“black carbon” is presented in fig. 30/6/1. The saturated fattyacids (n-alkanoic acids) range from 10 to 18 carbon atoms,maximizing at 12:0 (lauric acid). The main monounsaturatedfatty acids are the palmitoleic (16:1) and oleic acid (18:1). Bran-ched fatty acids elute (almost coelute) with the n-alkanoic acidsof odd carbon number (e.g., 15:0, 17:0, b-14:0, b-16:0). Signi-ficant peaks of phthalates indicate a contamination of the sam-ple by plastic material.
The stable carbon isotopic compositions (δ13C values) of thefatty acids in lipids extracted from archaeological samples hasshown to give valuable information on the vessel use, food con-sumption type, and the origin of dairying remains (e.g.)2100. Theδ13C values of the major fatty acids are given in (fig. 30/6/2).
As pointed out by Dudd/Evershed (1998) the covariation ofthe stable carbon isotopic compositions of the palmitic (16:0)and stearic (18:0) fatty acids are sensitive tools for distinguis-hing the milk, grease, and vegetables based dietary.
The δ13C16:0
vs. δ13C18:0
covariation of the “black carbon” resi-due fall above the field of ruminant fats (fig. 30/6/3). The sub-stantial enrichment in heavy carbon isotope (13C) of the black
carbon compared to the ovine/bovine fats may indicate diffe-rent lipid sources or preferential thermally induced degradationof some fatty acids. This last explanation is further supportedby the high content of short chain alkanoic acid (12:0) originatedfrom oxidative or thermal degradation of longer chains acids(16:1, 18:1, 16:0, 18:0). The isotopically lighter acids may be pre-ferentially degraded, and result in 13C-depleted shorter chainalkanes. This may explain why n-12:0 is up to 1.5 ‰ lighter thanthe palmitic (n-16:0) and stearic (n-18:0) acids.
Concluding remarksFor classification of archeological fats, Evershed u.a. (1999)
use δ13C16:0
and δ13C18:0
values from modern vegetable and animallipids. The carbon isotopic composition of these materials (pri-mary producers and consumers) depends on the δ13C value ofthe atmospheric CO
2fixed into organic compounds by photo-
synthesis. The pre-industrial atmospheric CO2
was isotopical-ly heavier (by ~1.6 ‰ between 1800 and 1980: Freyer 1986) thanin present time. Therefore, assuming that the isotopic fractio-nation in the pre-industrial (e.g. 230 n.Chr.) biogechemical car-bon cycle was determined by the today’s known photosynthe-tic mechanisms and metabolic pathways, we could expect theδ13C
16:0 vs. δ13C
18:0 covariation fields (for plants and consumers, fig.
30/6/3) at that time to be slightly shifted toward more positiveδ13C values.
In summary, the isotopic composition of the main fatty acidsextracted from a “black carbon” residue found at the roman villaof Biberist-Spitalhof exclude a vegetal origin, and point to ther-mally/chemically degraded fats from ruminant (ovine/bovine).
V. Funde und besondere Fundensembles römischer Zeit – 30. Die Tierknochen
2100 Dudd/Evershed 1998; Evershed u.a. 1999.
676
Fig. 30/6/1. GC/MS chromatogram of the fatty acid methyl esters ofthe lipids extracted from a "black carbon" residue. Peaks identification: 1 = dimethyl phthalate; 2 = cyclodecane; 3 = 2,4 dimethylphenol; 4 = lauric adic (12:0) methylester; 5 = 1-tetradecene; 6 = Myristic acid(14:0) methylester; 7 = pentadecanoic acid (15:0) methylester; 8 = ben-zenedicarboxylic acid methylester; 9 = palmitoleic acid (16:1) methyl-ester; 10 = palmitic acid (16:0) methylester; 11 = phthalate; 12 = oleicacid (18:1) methylester; 13 = stearic acid (18:0) methylester; 14 = phthalate.
Fig. 30/6/2. Stable carbon isotope ratios of the fatty acids extractedfrom a "black carbon" residue.

677Kap. 30.6
Fig. 30/6/3. Plot of the d13C values of the major saturated fatty acids(palmitic = 16:0, oleic = 18:1) from the lipids extracted from the blackcarbon. For comparison are plotted the oil lamp residue and soil fillingthe lamp (cf. chap. 21.5.2). The ringed fields encompasses the ranges forpresent-day animal fats (data from Evershed u.a. 1999). The field forpresent-day European C3-vegetable oil lipids is from Spangenberg et al.1998 and Spangenberg/Ogrinc 2001.


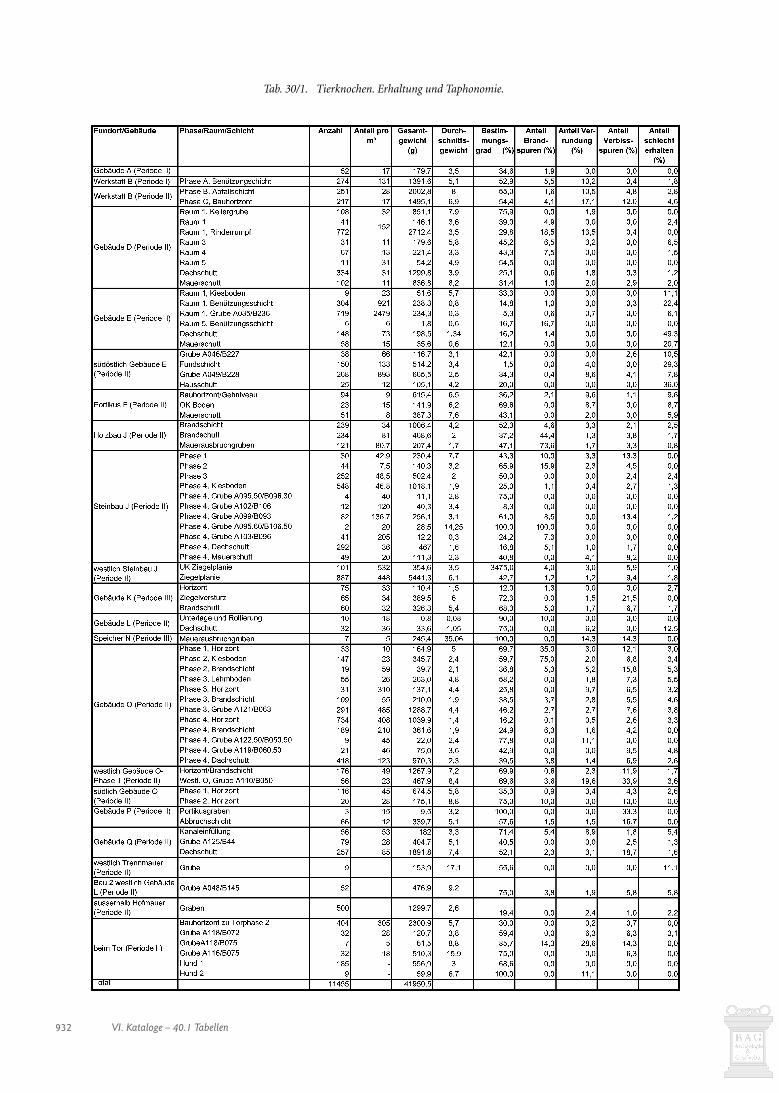
VI. Kataloge – 40.1 Tabellen932
Tab. 30/1. Tierknochen. Erhaltung und Taphonomie.
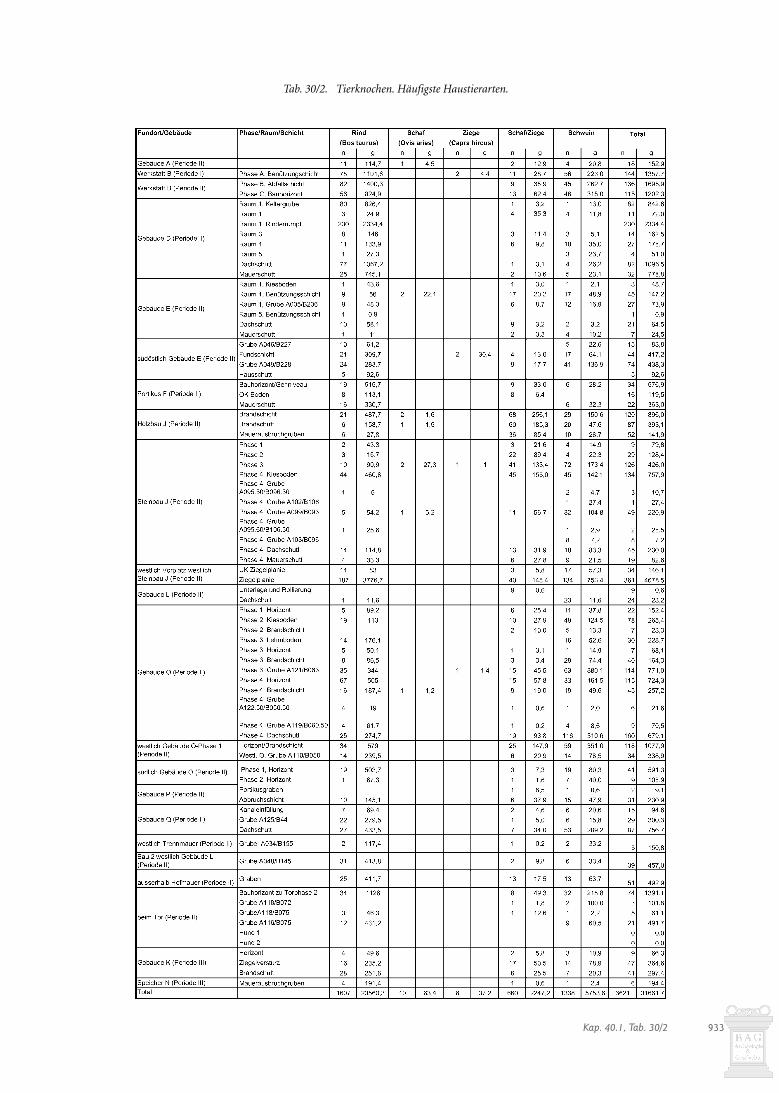
933Kap. 40.1, Tab. 30/2
Tab. 30/2. Tierknochen. Häufigste Haustierarten.
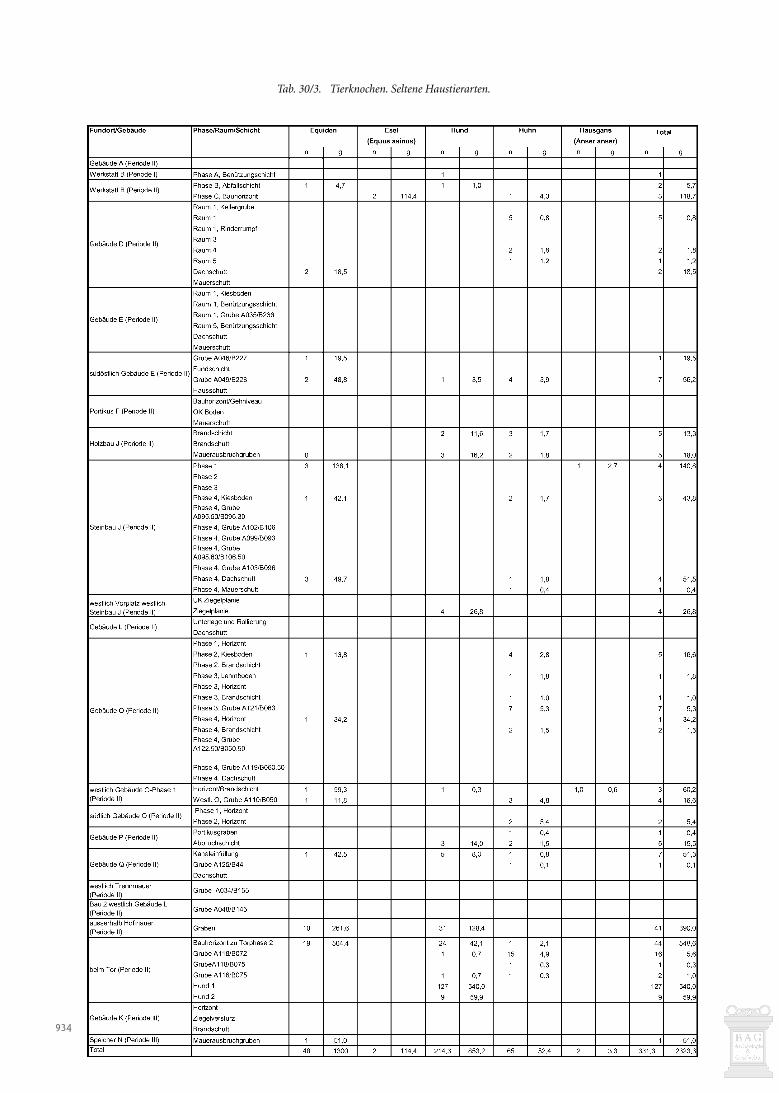
934
Tab. 30/3. Tierknochen. Seltene Haustierarten.
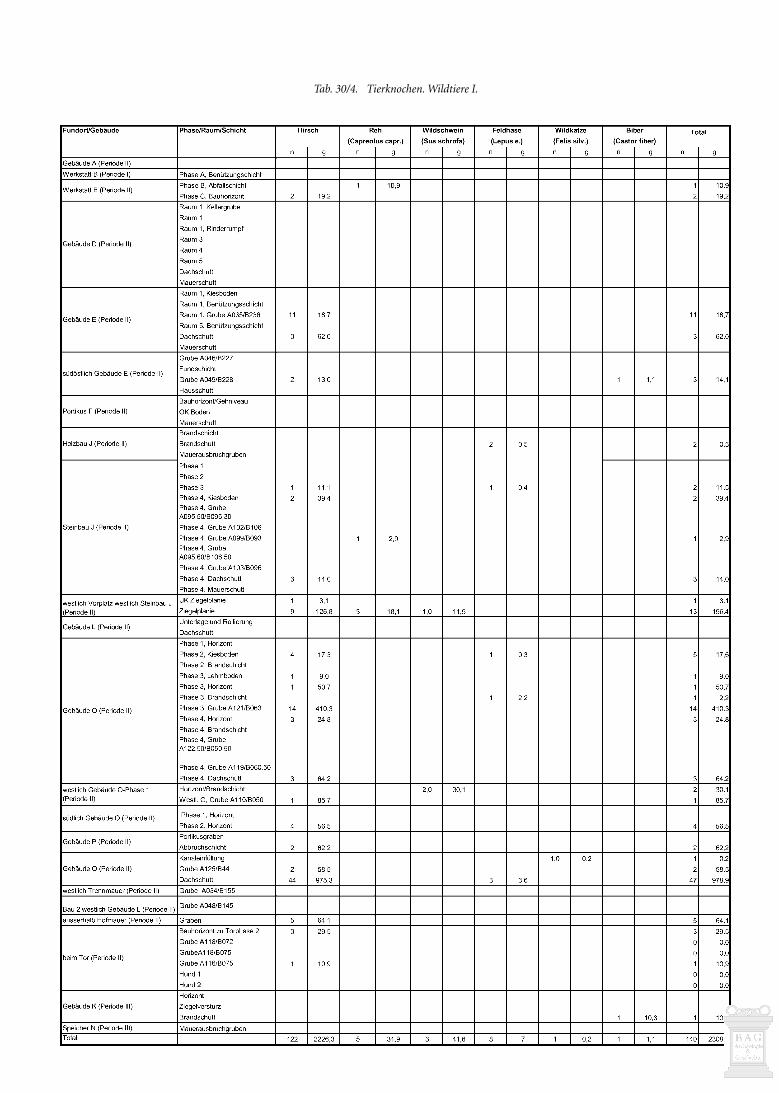
Tab. 30/4. Tierknochen. Wildtiere I.

VI. Kataloge – 40.1 Tabellen936
Tab. 30/5. Tierknochen. Wildtiere II.

937Kap. 40.1, Tab. 30/6
Tab. 30/6. Tierknochen. Grossgruppen.
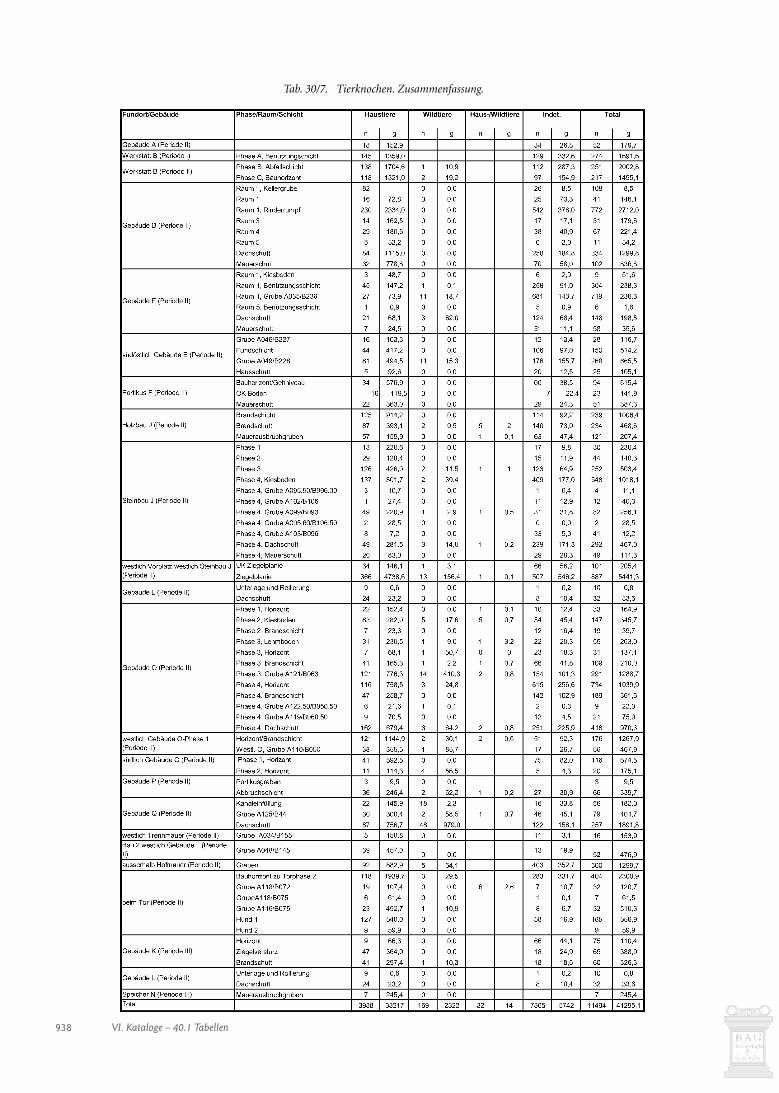
VI. Kataloge – 40.1 Tabellen938
Tab. 30/7. Tierknochen. Zusammenfassung.
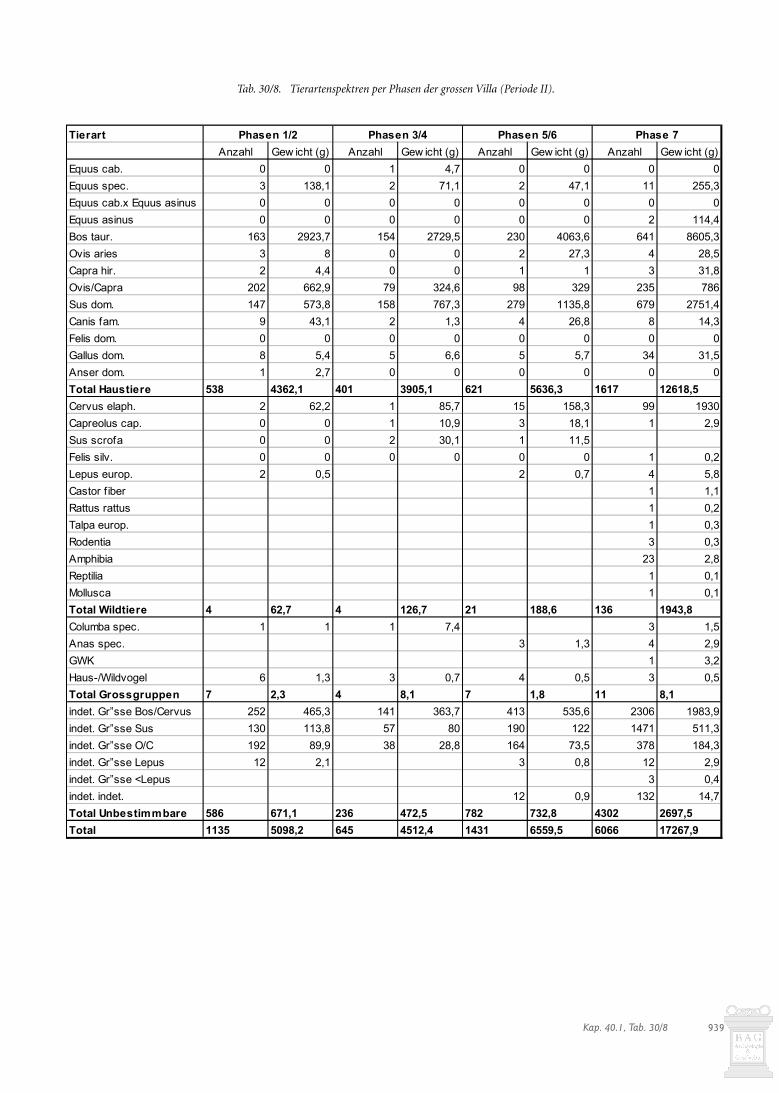
TierartAnzahl Gew icht (g) Anzahl Gew icht (g) Anzahl Gew icht (g) Anzahl Gew icht (g)
Equus cab. 0 0 1 4,7 0 0 0 0Equus spec. 3 138,1 2 71,1 2 47,1 11 255,3Equus cab.x Equus asinus 0 0 0 0 0 0 0 0Equus asinus 0 0 0 0 0 0 2 114,4Bos taur. 163 2923,7 154 2729,5 230 4063,6 641 8605,3Ovis aries 3 8 0 0 2 27,3 4 28,5Capra hir. 2 4,4 0 0 1 1 3 31,8Ovis/Capra 202 662,9 79 324,6 98 329 235 786Sus dom. 147 573,8 158 767,3 279 1135,8 679 2751,4Canis fam. 9 43,1 2 1,3 4 26,8 8 14,3Felis dom. 0 0 0 0 0 0 0 0Gallus dom. 8 5,4 5 6,6 5 5,7 34 31,5Anser dom. 1 2,7 0 0 0 0 0 0Total Haustiere 538 4362,1 401 3905,1 621 5636,3 1617 12618,5Cervus elaph. 2 62,2 1 85,7 15 158,3 99 1930Capreolus cap. 0 0 1 10,9 3 18,1 1 2,9Sus scrofa 0 0 2 30,1 1 11,5Felis silv. 0 0 0 0 0 0 1 0,2Lepus europ. 2 0,5 2 0,7 4 5,8Castor f iber 1 1,1Rattus rattus 1 0,2Talpa europ. 1 0,3Rodentia 3 0,3Amphibia 23 2,8Reptilia 1 0,1Mollusca 1 0,1Total Wildtiere 4 62,7 4 126,7 21 188,6 136 1943,8Columba spec. 1 1 1 7,4 3 1,5Anas spec. 3 1,3 4 2,9GWK 1 3,2Haus-/Wildvogel 6 1,3 3 0,7 4 0,5 3 0,5Total Grossgruppen 7 2,3 4 8,1 7 1,8 11 8,1indet. Gr”sse Bos/Cervus 252 465,3 141 363,7 413 535,6 2306 1983,9indet. Gr”sse Sus 130 113,8 57 80 190 122 1471 511,3indet. Gr”sse O/C 192 89,9 38 28,8 164 73,5 378 184,3indet. Gr”sse Lepus 12 2,1 3 0,8 12 2,9indet. Gr”sse <Lepus 3 0,4indet. indet. 12 0,9 132 14,7Total Unbestimmbare 586 671,1 236 472,5 782 732,8 4302 2697,5Total 1135 5098,2 645 4512,4 1431 6559,5 6066 17267,9
Phasen 1/2 Phasen 3/4 Phasen 5/6 Phase 7
939Kap. 40.1, Tab. 30/8
Tab. 30/8. Tierartenspektren per Phasen der grossen Villa (Periode II).

VI. Kataloge – 40.1 Tabellen940
Tab. 30/9. Tierknochen aus Fundkomplexen der Grabanlage H (unverbrannt kursiv).

941
Tab. 30/10. Masse der Rinder in den Phasen der grossen Villa (Periode II).

Phasen 3/4SD Bp Dp SD Bd Dd GLl GLm Dl Dm GL GB Glpe DLS Ld MBS
Radius 47,86Radius und Ulna 46,6Tibia 49,21Talus 30,63 53,63 50,69 32,63Talus 55,87 52,41 29,52 31,84Calcaneus 118,07Calcaneus 33,39Calcaneus 120,61 32,08Centrotarsale 41,14Centrotarsale 43,81Metacarpus III+IV( 30) 41,94 30,32 25,24 40,57 28,35 265Metacarpus III+IV 42,3 28,42Metatarsus III+IV ( 50) 42,32 35,02 22,49 39,54 306Metatarsus III+IV ( 50) 32,32Phalanx 1 anterior/posterior ( 65) 20,56 17,17 19,53 59,81Phalanx 1 anterior/posterior ( 65) 19,4 13,89 16,47 50,05Phalanx 2 anterior/posterior 16,9Phalanx 1 posterior 18,64Phalanx 3 anterior/posterior 47,08 44,14 12,87
Phase 7CERVUS ELAPHUS [ 70]
VI. Kataloge – 40.1 Tabellen942
Tab. 30/11. Masse der Hirsche in den Phasen der grossen Villa (Periode II).
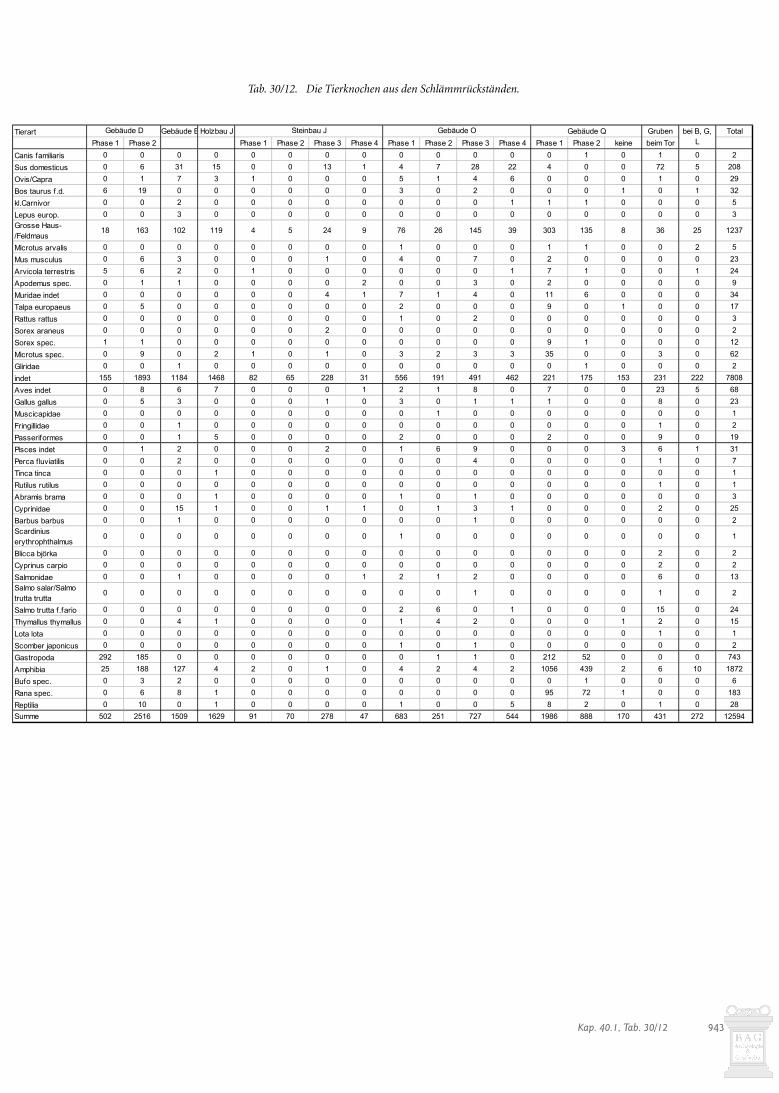
Tierart Gebäude E Holzbau J Gebäude Q Gruben TotalPhase 1 Phase 2 Phase 1 Phase 2 Phase 3 Phase 4 Phase 1 Phase 2 Phase 3 Phase 4 Phase 1 Phase 2 keine beim Tor
Canis familiaris 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 2Sus domesticus 0 6 31 15 0 0 13 1 4 7 28 22 4 0 0 72 5 208Ovis/Capra 0 1 7 3 1 0 0 0 5 1 4 6 0 0 0 1 0 29Bos taurus f.d. 6 19 0 0 0 0 0 0 3 0 2 0 0 0 1 0 1 32kl.Carnivor 0 0 2 0 0 0 0 0 0 0 0 1 1 1 0 0 0 5Lepus europ. 0 0 3 0 0 0 0 0 0 0 0 0 0 0 0 0 0 3Grosse Haus-/Feldmaus
18 163 102 119 4 5 24 9 76 26 145 39 303 135 8 36 25 1237
Microtus arvalis 0 0 0 0 0 0 0 0 1 0 0 0 1 1 0 0 2 5Mus musculus 0 6 3 0 0 0 1 0 4 0 7 0 2 0 0 0 0 23Arvicola terrestris 5 6 2 0 1 0 0 0 0 0 0 1 7 1 0 0 1 24Apodemus spec. 0 1 1 0 0 0 0 2 0 0 3 0 2 0 0 0 0 9Muridae indet 0 0 0 0 0 0 4 1 7 1 4 0 11 6 0 0 0 34Talpa europaeus 0 5 0 0 0 0 0 0 2 0 0 0 9 0 1 0 0 17Rattus rattus 0 0 0 0 0 0 0 0 1 0 2 0 0 0 0 0 0 3Sorex araneus 0 0 0 0 0 0 2 0 0 0 0 0 0 0 0 0 0 2Sorex spec. 1 1 0 0 0 0 0 0 0 0 0 0 9 1 0 0 0 12Microtus spec. 0 9 0 2 1 0 1 0 3 2 3 3 35 0 0 3 0 62Gliridae 0 0 1 0 0 0 0 0 0 0 0 0 0 1 0 0 0 2indet 155 1893 1184 1468 82 65 228 31 556 191 491 462 221 175 153 231 222 7808Aves indet 0 8 6 7 0 0 0 1 2 1 8 0 7 0 0 23 5 68Gallus gallus 0 5 3 0 0 0 1 0 3 0 1 1 1 0 0 8 0 23Muscicapidae 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 1Fringillidae 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 1 0 2Passeriformes 0 0 1 5 0 0 0 0 2 0 0 0 2 0 0 9 0 19Pisces indet 0 1 2 0 0 0 2 0 1 6 9 0 0 0 3 6 1 31Perca f luviatilis 0 0 2 0 0 0 0 0 0 0 4 0 0 0 0 1 0 7Tinca tinca 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1Rutilus rutilus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1Abramis brama 0 0 0 1 0 0 0 0 1 0 1 0 0 0 0 0 0 3Cyprinidae 0 0 15 1 0 0 1 1 0 1 3 1 0 0 0 2 0 25Barbus barbus 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 2Scardinius erythrophthalmus
0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1
Blicca björka 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 2 0 2Cyprinus carpio 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 2 0 2Salmonidae 0 0 1 0 0 0 0 1 2 1 2 0 0 0 0 6 0 13Salmo salar/Salmo trutta trutta
0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 1 0 2
Salmo trutta f.fario 0 0 0 0 0 0 0 0 2 6 0 1 0 0 0 15 0 24Thymallus thymallus 0 0 4 1 0 0 0 0 1 4 2 0 0 0 1 2 0 15Lota lota 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1Scomber japonicus 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 0 0 2Gastropoda 292 185 0 0 0 0 0 0 0 1 1 0 212 52 0 0 0 743Amphibia 25 188 127 4 2 0 1 0 4 2 4 2 1056 439 2 6 10 1872Bufo spec. 0 3 2 0 0 0 0 0 0 0 0 0 0 1 0 0 0 6Rana spec. 0 6 8 1 0 0 0 0 0 0 0 0 95 72 1 0 0 183Reptilia 0 10 0 1 0 0 0 0 1 0 0 5 8 2 0 1 0 28Summe 502 2516 1509 1629 91 70 278 47 683 251 727 544 1986 888 170 431 272 12594
Gebäude D Steinbau J bei B, G, L
Gebäude O
943Kap. 40.1, Tab. 30/12
Tab. 30/12. Die Tierknochen aus den Schlämmrückständen.




















