
Development of a pyrG Mutant of Aspergillus oryzae Strain S1 as a Host for the Production of...
7
Hindawi Publishing Corporation e Scientific World Journal Volume 2013, Article ID 634317, 7 pages http://dx.doi.org/10.1155/2013/634317 Research Article Development of a pyrG Mutant of Aspergillus oryzae Strain S1 as a Host for the Production of Heterologous Proteins Selina Oh Siew Ling, 1 Reginald Storms, 2,3 Yun Zheng, 2 Mohd Rohaizad Mohd Rodzi, 1 Nor Muhammad Mahadi, 4 Rosli Md Illias, 5 Abdul Munir Abdul Murad, 1 and Farah Diba Abu Bakar 1 1 School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia 2 Centre for Structural and Functional Genomics, Concordia University, 7141 Sherbrooke Street West, Montr´ eal, Qu´ ebec, Canada H4B 1R6 3 Department of Biology, Concordia University, 7141 Sherbrooke Street West, Montr´ eal, Qu´ ebec, Canada H4B 1R6 4 Malaysia Genome Institute, Jalan Bangi, 43000 Kajang, Selangor, Malaysia 5 Department of Bioprocess Engineering, Faculty of Chemical Engineering, Universiti Teknologi Malaysia, 81310 Skudai, Johor, Malaysia Correspondence should be addressed to Farah Diba Abu Bakar; fabyff@ukm.my Received 20 August 2013; Accepted 6 September 2013 Academic Editors: N. F. Abrams and M. D. Vodenicharov Copyright © 2013 Selina Oh Siew Ling et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. e ease with which auxotrophic strains and genes that complement them can be manipulated, as well as the stability of auxotrophic selection systems, are amongst the advantages of using auxotrophic markers to produce heterologous proteins. Most auxotrophic markers in Aspergillus oryzae originate from chemical or physical mutagenesis that may yield undesirable mutations along with the mutation of interest. An auxotrophic A. oryzae strain S1 was generated by deleting the orotidine-5 -monophosphate decarboxylase gene (pyrG) by targeted gene replacement. e uridine requirement of the resulting strain GR6 pyrGΔ0 was complemented by plasmids carrying a pyrG gene from either Aspergillus nidulans or A. oryzae. -Galactosidase expression by strain GR6 pyrGΔ0 transformed with an A. niger plasmid encoding a heterologous -galactosidase was at least 150 times more than that obtained with the untransformed strain. Targeted gene replacement is thus an efficient way of developing auxotrophic mutants in A. oryzae and the auxotrophic strain GR6 pyrGΔ0 facilitated the production of a heterologous protein in this fungus. 1. Introduction In Japan, the filamentous fungus Aspergillus oryzae is exten- sively used in the production of miso (soybean paste), sake (rice wine), and shoyu (soy sauce). Due to its proven safety record in the food industry, A. oryzae has been assigned the status Generally Recognized As Safe (GRAS) by the United States Food and Drug Administration (USFDA) and the World Health Organization (WHO) [1]. A. oryzae is also used for the production of homologous and heterologous proteins and metabolites [2]. e A. oryzae genome consists of eight chromosomes totalling 37 megabases (Mb) predicted to harbour 12,074 protein coding genes [3]. e pyrG gene encodes orotidine-5 -monophosphate (OMP) decarboxylase, an enzyme of the pyrimidine biosyn- thesis pathway that is important for uridine synthesis. e mutagen 5-fluoroorotic acid (5-FOA) inhibits cell prolifer- ation in most organisms by entering the pyrimidine syn- thesis pathway where it is converted into the pyrimidine analogue fluoroorotidine monophosphate. rough the sub- sequent action of ribonucleotide reductase, fluoroorotidine monophosphate is converted into fluorodeoxyuridine, a potent suicidal inhibitor of the enzyme thymidylate syn- thase, which converts deoxyuridine monophosphate into the essential DNA precursor thymidylate [4]. Resistance to 5-FOA can be achieved by blocking the pyrimidine
Transcript of Development of a pyrG Mutant of Aspergillus oryzae Strain S1 as a Host for the Production of...

Hindawi Publishing CorporationThe Scientific World JournalVolume 2013 Article ID 634317 7 pageshttpdxdoiorg1011552013634317
Research ArticleDevelopment of a pyrG Mutant of Aspergillus oryzae Strain S1as a Host for the Production of Heterologous Proteins
Selina Oh Siew Ling1 Reginald Storms23 Yun Zheng2
Mohd Rohaizad Mohd Rodzi1 Nor Muhammad Mahadi4 Rosli Md Illias5
Abdul Munir Abdul Murad1 and Farah Diba Abu Bakar1
1 School of Biosciences and Biotechnology Faculty of Science and Technology Universiti Kebangsaan Malaysia43600 Bangi Selangor Malaysia
2 Centre for Structural and Functional Genomics Concordia University 7141 Sherbrooke Street West MontrealQuebec Canada H4B 1R6
3Department of Biology Concordia University 7141 Sherbrooke Street West Montreal Quebec Canada H4B 1R64Malaysia Genome Institute Jalan Bangi 43000 Kajang Selangor Malaysia5 Department of Bioprocess Engineering Faculty of Chemical Engineering Universiti TeknologiMalaysia 81310 Skudai JohorMalaysia
Correspondence should be addressed to Farah Diba Abu Bakar fabyffukmmy
Received 20 August 2013 Accepted 6 September 2013
Academic Editors N F Abrams and M D Vodenicharov
Copyright copy 2013 Selina Oh Siew Ling et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
The ease with which auxotrophic strains and genes that complement them can bemanipulated as well as the stability of auxotrophicselection systems are amongst the advantages of using auxotrophic markers to produce heterologous proteins Most auxotrophicmarkers inAspergillus oryzae originate from chemical or physical mutagenesis that may yield undesirable mutations along with themutation of interest An auxotrophic A oryzae strain S1 was generated by deleting the orotidine-51015840-monophosphate decarboxylasegene (pyrG) by targeted gene replacement The uridine requirement of the resulting strain GR6 pyrGΔ0 was complemented byplasmids carrying a pyrG gene from either Aspergillus nidulans or A oryzae 120573-Galactosidase expression by strain GR6 pyrGΔ0transformed with an A niger plasmid encoding a heterologous 120573-galactosidase was at least 150 times more than that obtained withthe untransformed strain Targeted gene replacement is thus an efficient way of developing auxotrophic mutants in A oryzae andthe auxotrophic strain GR6 pyrGΔ0 facilitated the production of a heterologous protein in this fungus
1 Introduction
In Japan the filamentous fungus Aspergillus oryzae is exten-sively used in the production of miso (soybean paste) sake(rice wine) and shoyu (soy sauce) Due to its proven safetyrecord in the food industry A oryzae has been assigned thestatus Generally Recognized As Safe (GRAS) by the UnitedStates Food and Drug Administration (USFDA) and theWorld Health Organization (WHO) [1] A oryzae is alsoused for the production of homologous and heterologousproteins and metabolites [2] The A oryzae genome consistsof eight chromosomes totalling 37megabases (Mb) predictedto harbour 12074 protein coding genes [3]
The pyrG gene encodes orotidine-51015840-monophosphate(OMP) decarboxylase an enzyme of the pyrimidine biosyn-thesis pathway that is important for uridine synthesis Themutagen 5-fluoroorotic acid (5-FOA) inhibits cell prolifer-ation in most organisms by entering the pyrimidine syn-thesis pathway where it is converted into the pyrimidineanalogue fluoroorotidine monophosphate Through the sub-sequent action of ribonucleotide reductase fluoroorotidinemonophosphate is converted into fluorodeoxyuridine apotent suicidal inhibitor of the enzyme thymidylate syn-thase which converts deoxyuridine monophosphate intothe essential DNA precursor thymidylate [4] Resistanceto 5-FOA can be achieved by blocking the pyrimidine
2 The Scientific World Journal
Not I 12261
Ssp I 14084 Ssp I 424
Fse I 1488
Nhe I 4929
Mlu I 5444Aat II 5590
Not I 6994
AMA1
AppUC18
14000
12000
10000
8000 6000
4000
2000
ANEp214265bps
GlaTt
LacA
GlaPr
pyrG
(a)
1000
2000
3000
4000
Eco RI 396
Eco RI 2255
pyrG
AppUC19
pUC pyrGAo
4545bps
(b)
Figure 1 Plasmids used for transformation (a) Plasmid ANEp2 containing the pyrG gene of A nidulans and a GlaPr-lacA-GlaTt chimera(b) plasmid pUC pyrGAo containing pyrG (1839 bp) of A oryzae strain S1
pathway through mutational inactivation of orotidine-51015840-monophosphate (OMP) decarboxylase which catalyzes thedecarboxylation of orotidine monophosphate (OMP) toform uridine monophosphate (UMP) In filamentous fungipyrimidine auxotrophs obtained through inactivation of theOMP decarboxylase gene (usually designated pyrG) are aux-otrophic for uridine andor uracil and are resistant to 5-FOA[5 6] The generation of 5-FOA-resistant pyrG mutants hasbeen reported in other fungi including A aculeatus A nigerSclerotinia sclerotiorum and Trichoderma reesei [5 7ndash9]Most pyrGmutants were isolated following either ultraviolet(UV) or chemical mutagenesis Mutations induced in theseways are often unstable (ie they revert to the wild type)andmay includemultiple undesirablemutations In contrastmutants generated by targeted gene replacement are stable asthe targeted gene is removed from the genome It is howevermore difficult to mutate multiple genes this way [10 11]
A BLAST search using the pyrG sequence of A oryzaeS1 against the A oryzae RIB40 genome database at theNational Institute of Technology and Evaluation (NITEhttpwwwbionitegojpdogan) showed that pyrG is presentas a single copy on chromosome 7 (contig SC011 location2219667 2220565)
In this study a gene replacement cassette was constructedconsisting of the native pyrG locus extending 15 to 20 kbbeyond the upstream and downstream termini of the pyrGcoding region but lacking the open reading frame of the geneTransformation of A oryzae strain S1 with the gene replace-ment cassette generated 5-FOA-resistant uracil and uridineauxotrophs Putative pyrGΔ0 mutants were verified by com-paring their growth on selective and nonselective mediaPCR analyses of the pyrG region in the putative mutantsand complementation with plasmids containing wild-type
homologous and heterologous pyrG genes Suitability of thepyrGΔ0mutant as a host for heterologous protein productionwas assessed using strain GR6 harbouring a plasmid inwhich the pyrG and heterologous 120573-galactosidase genes wereintegrated
2 Materials and Methods
21 Fungus Cultivation and Plasmids A oryzae strain S1wasobtained from the Fungal Stock Culture Collection of theMolecular Mycology Laboratory School of Biosciences andBiotechnology Faculty of Science and Technology Uni-versiti Kebangsaan Malaysia The fungus was maintainedon a Potato Dextrose Agar (PDA Difco France) at 30∘Cand subcultured monthly Fungal cultivation was carriedout by inoculating onto a fresh PDA plate Cultures wereincubated for 7 days at 30∘C Plasmid ANEp2 (Figure 1(a))is a shuttle vector capable of autonomous replication inAspergilla [12] In addition to the pUC18 backbone ANEp2carries the A nidulans pyrG gene as a selectable markeran expression cassette (GlaPr-lacA-GlaTt) that expresses anA niger secreted 120573-galactosidase and the AMA1 sequencethat supports autonomous replication in Aspergilli ANEp2was used to functionally test for complementation of uridineauxotrophy and reporter gene expression in anA oryzae pyrGmutant Plasmid pUC pyrGAo containing the pyrG of the Aoryzae strain S1 was also used for complementation of uridineauxotrophy (Figure 1(b)) [13] Escherichia coli strain DH5120572was used for DNA manipulation
22 Media Polypeptone dextrin (PD) medium containing10 (wv) polypeptone 20 (wv) glucose 05 (wv)
The Scientific World Journal 3
Table 1 Primers used in this study
Designation Primer description Primer sequence51015840pyr pyrG 51015840 region sense primer 51015840-GGCGCCACAATCGTTGAGGTA-31015840lowastrevint-XhoI pyrG 51015840 region antisense primer the XhoI site is underlined 51015840-CTCGAGGTTGGCGATGGAGG-31015840lowastfwdint-XhoI pyrG 31015840 region sense primer the XhoI site is underlined 51015840-CTCGAGGTAGTGGTGGATACGTACT-31015840
31015840pyr pyrG 31015840 region antisense primer 51015840-AAGCTTATCAGCTGCATATCTCTAT-31015840
Pyr51015840-out pyrG 51015840 region sense primer outside the pyrG 51015840 genereplacement cassette 51015840-ATTTAGTCATTCCTTTTGGCATC-31015840
Pyr31015840-out pyrG 31015840 region antisense primer outside the pyrG 31015840 genereplacement cassette 51015840-CTAATTTTAATGTCTTCCTTCAG-31015840
lowastrevint-XhoI was used to amplify the targeting region downstream of the pyrG coding regionlowastfwdint-XhoI was used to amplify the targeting region upstream of the pyrG coding region
KH2PO4 01 (wv) NaNO
3 and 005 (wv) MgSO
4sdot7H2O
was used for liquid cultivation of A oryzae [14] Czapek-Dox (CD) medium composed of 02 (wv) NaNO
3 01
(wv) K2HPO4 005 (wv) MgSO
4sdot7H2O 005 (wv)
KCl 0001 (wv) FeSO4and 30 (wv) sucrose at pH 55
(supplemented with 02 (wv) uracil and 20mM uridine)was used for spheroplast cell wall regeneration Spheroplastswere overlaid with CD medium containing 05 (wv) agarand 50 (wv)NaCl ontoCDmediumcontaining 20 (wv)agar and 50 (wv) NaCl [15] Cultivation and selection ofpositive transformants were performed using CD minimalagar media [16] composed of 02 (wv) NaNO
3 01
(wv) KH2PO4 005 (wv) MgSO
4sdot7H2O 005 (wv) KCl
0002 (wv) FeSO4sdot7H2O 20 (wv) glucose 005 (wv)
NaCl and 2 (wv) agar at pH 55 containing 2mgmL 5-FOA 02 (wv) uracil and 20mM uridine
23 Replacement Cassette Construction The cassette used todelete theA oryzae pyrG (Figure 2) was assembled as followsgenomic DNA of A oryzae strain S1 was used as the PCRtemplate to amplify the 16 kb and 20 kb regions upstreamand downstream of the pyrG coding region respectivelyThe upstream region was amplified with the primers 51015840pyrand revint-XhoI while the downstream region was amplifiedusing the primers fwdint-XhoI and 31015840pyr (Table 1) BothPCR products were ligated into the vector pGEM-T Easy(Promega Fitchburg WI USA) Plasmids pGEM-51015840pyr andpGEM-31015840pyr were identified by restriction endonucleasedigestion and verified by sequencing Finally the two regionsflanking the pyrG coding regionwere assembled into the pyrGreplacement cassette by ligating the plasmids pGEM-51015840pyrand pGEM-31015840pyr digested with XhoI and SalI Restrictionendonuclease mapping and sequencing were used to verifythe construction of pGEM-51015840 31015840pyr
24 Spheroplast Preparation and Transformation Sphero-plasts were produced based on the methods of Ushijimaet al [14] and Liew [17] as follows A oryzae conidia (107conidiamL) were cultivated in 50mL PD at 30∘C for 20 hMycelia (3 g wet weight) were pretreated with DTT-lysisbuffer containing 10mM dithiothreitol (DTT) and 10mMTris-HCl for 1 h at 30∘C by shaking at 75 rpm prior to incuba-tion with 70mgmL lysing enzyme (Trichoderma harzianum
Sigma-Aldrich Corp St Louis MO USA) solution contain-ing osmotic buffer (08MMgSO
4in 001M phosphate buffer)
for 4 h at 30∘C (agitationmdash75 rpm) The mixture containingspheroplasts was transferred to a 50mL centrifuge tube(Greiner Bio-One UK) and 06M sorbitol was added slowlyto the mixture The mixture was spun at 4000 rpm 4∘C for30minThe interphase layer between the lysis buffer and sor-bitol was collected and transferred to a new 50mL tube withthe addition of STC buffer (12M sorbitol 001M Tris-HCl001M CaCl
2) Transformation was carried out according to
Storms et al [12] andWernars et al [18] using 16 times 106 viablespheroplasts in STC mixed with 10120583g purified PCR product(36 kb amplified DNA fragment consisting of the 51015840pyr and31015840pyr regions of the pyrG gene replacement cassette) and12M sorbitol as the osmotic stabilizer Subsequently 05mLaliquots containing 8 times 105 transformed spheroplasts weremixed into 05 CD overlay agar poured onto 2 CD agarplates supplemented with 02 uracil and 20mM uridineand incubated at 30∘C for 4ndash6 days
25 Screening and Analysis of Transformants 5-FOA resis-tant colonies were tested for auxotrophy by patching onCD medium and CD medium with uracil and uridineVerification that the 5-FOA-resistant auxotrophs arose viadeletion of the pyrG coding region was done by PCRanalysis of total DNA using primers Pyr51015840-out and Pyr31015840-out (Table 1) designed to be complementary to the knowngenomic (obtained from the A oryzae RIB40 NITE genomedatabase) sequence just beyond the upstream and down-stream boundaries of the replacement cassette The pyrGΔ0mutants were also tested by complementation with plasmidspUC pyrGAo and ANEp2 harbouring A oryzae and A nigerpyrG respectively
26 120573-Galactosidase Expression Levels of 120573-Galactosidaseactivity expressed by A oryzae strain GR6 pyrGΔ0 and GR6pyrGΔ0 transformants harbouring ANEp2 were determinedusing filtered medium from 7-day old static cultures Staticcultures were generated by inoculating 10 times 106 conid-iospores into Petri dishes containing 25mL of MMJ mediumas described by Kafer [19] using 15 maltose as the carbonsource (along with the necessary supplements) [20] at 30∘C
4 The Scientific World Journal
Digest with SalI and XhoI
Ligate
5998400pyr
pyrG upstream
XhoI 1639 fwdint XhoI
XhoI 62
pyrG downstream
SalI 2083
revint XhoI
pGEM-5998400pyr4608bp
pGEM-3998400pyr5008bp
3998400pyr
XhoI
5998400pyr
6585bppGEM-5998400 3998400pyr
SalI 1683
3998400pyr
pGEM-Treg EasypGEM-Treg Easy
pGEM-Treg Easy
Figure 2 Schematic diagram outlining assembly of the pyrG gene replacement cassette
120573-Galactosidase assays were performed by adding culture fil-trates to a reaction-buffer containing 50mM sodium acetate(pH 41) and 4mgmL of orthonitrophenyl-120573-D-galactoside(ONPG) and incubating for 10min at 37∘C Assays werestopped by adding 1 volume of 1M sodium carbonate andthe absorbance was read at 415 nm 120573-Galactosidase activityis expressed in units (U) per mL of culture where one unit isthe amount of enzyme needed to hydrolyze 1 120583moL of ONPGto orthonitrophenol per min at 37∘C Western blot analyseswere carried out using 20 120583L samples of concentrated culturefiltrate using ultra-filtration spin columns with a molecularweight cut-off of 10 kDa (ldquoVivaspinrdquo Sartorius GottingenGermany) and fractionated by 10 SDS-PAGE Fractionatedproteins were transferred to nitrocellulose membranes andvisualized using a rabbit anti-lac A antiserum an anti-rabbitIgG labelled with horseradish peroxidase (Promega) and achemiluminescent detection kit (Thermo Fisher ScientificWaltham MA USA) Rabbit serum containing polyclonalantibodies directed against A niger 120573-galactosidase wasgenerated as follows A niger 120573-galactosidase was expressedand purified by ion exchange chromatography Purified120573-galactosidase (200120583g with complete Freundrsquos Adjuvantfor immunization) was introduced by subcutaneous injec-tions into New Zealand white rabbits The initial injectionwas followed by two booster injections (100120583g purified120573-galactosidase with Incomplete Freundrsquos Adjuvant) For
serum preparation the collected blood was stored at 4∘Covernight and centrifuged for 30min at 4000 rpm Thecollected supernatant was stored at minus20∘C Enzyme-linkedimmunosorbent assay (ELISA) was used for the evaluationof antibody positive titer in the rabbit serum
3 Results
31 Deletion of the A oryzae Strain S1 pyrG The 36kb pyrGΔ0 gene replacement cassette was amplified usingprimers 51015840pyr and 31015840pyr and plasmid pGEM-51015840 31015840pyr as thetemplate (Figure 4(a)) Generation of a pyrGΔ0 derivativeof A oryzae was performed by transforming strain S1 withthe 36 kb pyrGΔ0 cassette Initial attempts to regeneratetransformed spheroplasts were performed on CD platessupplemented with uracil uridine and 5-FOAThis strategywhich selects for pyrGΔ0 gene replacements immediatelyafter transformation of spheroplasts with the PCR productis similar to the approach used to isolate pyrG mutantsof Trichoderma reesei (irradiated spores are platedout onminimal medium containing uridine and 5-FOAmdashLonget al [5]) However colonies derived from the experimentsdescribed here with pyrG failed to yield FOA resistantcolonies Perhaps 5-FOA-resistant pyrGΔ0 mutants werenot obtained because multiple nuclei were present in thespheroplasts and therefore a single pyrGΔ0 locus could
The Scientific World Journal 5
(a) Wild type (b) GR6 mutant
(c) CD plates (minimal medium)
(d) CD (uracil uridineand 5-FOA) plates
Figure 3 Media plates showing growth or no growth of the Aoryzae S1 wild type (column (a)) and GR6 pyrG mutant (column(b)) Column (a) ability of the wild type strain S1 (row (c)) andColumn (b) the inability of the GR6 pyrGmutant strain (row (c)) togrow on minimal medium CD plates Column (a) inability of thewild type strain S1 (row (d)) and Column (b) ability of the GR6pyrGmutant strain (row (d)) to grow onminimalmediumCDplatessupplemented with uracil uridine and 5-FOA
not prevent production of the suicide inhibitor FdUMPTo address this possibility the transformation protocol wasmodified by allowing nuclei in which the pyrG gene wasdeleted to segregate from wild-type nuclei by plating thetransformed spheroplasts (about 8 times 105 per plate) ontononselective regeneration plates containing uraciluridineOnce a lawn of conidia was produced the conidia werecollected diluted and plated at 8 times 106 spores per plateon CD selective medium with 5-FOA uracil and uridineTen randomly selected colonies were chosen from over 100colonies that formed and purified to homogeneity by tworounds of plating on CD uracil and uridine plates Conidiaprepared using the purified colonies were harvested andplated onto CD plates without uracil and uridine (106 conidiaper plate) Only one of the putative mutants designated GR6(Gene Replacement mutant number 6) did not grow and wastherefore assumed to be completely stable (Figure 3)
32 Verifying that GR6 Harbours a pyrGΔ0 Locus PCRamplifications with primers Pyr51015840-out and Pyr31015840-out usingthe genomic DNA of strain S1 and strain GR6 as the tem-plates showed that a 09 kb deletion was introduced into thepyrG region of strain GR6 (Figure 4(b)) which is consistentwith generation of the desired pyrGΔ0 locus by homologousrecombination
Then the pyrG auxotroph GR6 was transformed withplasmids pUC pyrGAo and ANEp2 These plasmids containthe wild-type pyrG from A oryzae (in pUC19) and Anidulans respectively Both plasmids were introduced intoand restored uracil and uridine prototrophy of GR6 withfrequencies of three transformants per 120583g of plasmid DNAfor pUC pyrGAo and 50 transformants per 120583g of plasmidDNA for ANEp2 Furthermore the frequency ratio of pyrGprototrophs obtained when transformation was performedwith the pUC pyrGAo versus the AMA plasmid was about01 This is in accordance with the range of prototrophy fre-quency ratios obtained with integrating versus autonomously
1kb 1
35kb
30kb sim36kb
(a)
50kb40kb35kb
sim45kb
sim36kb
1kb 1 2
(b)
Figure 4 Gel electrophoretic profile of the (a) (lane 1) genereplacement cassette consisting of the 51015840 and 31015840pyrG DNA inserts(without the coding region) with the size of sim36 kb and (b) genomicPCR product of the A oryzae S1 wild type strain (lane 1) and itsmutant derivative pyrGΔ0 strain GR6 (lane 2) Lane M 1 kb DNAladder (Fermentas Canada)
replicating plasmids previously reported with other fungalspecies [12 21]
33 Protein Expression Total secreted proteins in culturefiltrates of the untransformed strain GR6 pyrGΔ0 and GR6pyrGΔ0 transformed with ANEp2 were analyzed by SDS-PAGE (Figure 5(a)) and Western blotting (Figure 5(b))SDS-PAGE showed a very prominent 110 kDa proteinwhich is about the size expected of the A niger 120573-galactosidase (Swiss-Prot accession number P29853) (Unit-ProtKB httpwwwuniprotorguniprotP29853) [22] in theculture filtrate of ANEp2 GR6 pyrGΔ0 which was absentin the untransformed strain GR6 pyrGΔ0 Western blotanalyses confirmed that the major band was the A niger 120573-galactosidase As expected from the above culture filtratesof the ANEp2 GR6 pyrGΔ0 transformants possessed highlevels of 120573-galactosidase activity (135UmL) as compared tothe untransformed strain GR6 pyrGΔ0 which expressed 120573-galactosidase at 080UmL
4 Discussion
Replacement of a gene requires homologous recombinationof the introduced DNA with the genome gene replacementis achieved by a precise double crossover event [23] We firstsought these events by selecting for FOA-resistant clonesFOA inhibits the growth of the fungus but permits multipli-cation of cells deficient in OMP decarboxylase and becauseof this are auxotrophic for uridine Initially we selected forpyrGΔ0 gene replacements immediately after transfectionusing a similar plating procedure as has been used toisolate pyrG mutants of T reesei A aculeatus and Penicil-lium nalgiovense from irradiated spores [5 7 21] In ourhands this technique failed to yield FOA resistant colonies
6 The Scientific World Journal
M 1 2+ve
175kDa
80kDa
58kDa
46kDa
30kDa(a)
1 2+ve
110kDa
(b)
Figure 5 Analysis of GR6 culture filtrates Panel (a) lane M SDS-PAGE profile of prestained protein marker (New England Biolabs USA)lane +ve purified A niger 120573-galactosidase isolated from a culture filtrate of A niger transformed with the GlaPr-lacA-GlaTt expressioncassette lane 1 culture filtrate prepared using A oryzae strain GR6 pyrGΔ0 and lane 2 culture filtrate prepared using GR6 pyrGΔ0transformed with ANEp2 Panel (b) Western blot performed by probing the gel shown in Panel (a) with polyclonal rabbit antibodies directedagainst A niger 120573-galactosidase
This was probably because multiple nuclei were present ineach spheroplast and therefore a single pyrGΔ0 locus couldnot prevent production of the suicide inhibitor FdUMPTo test this the transformation protocol was modified toallow cells containing putative pyrG deletions to segregatefrom wild-type ones by plating the transformed spheroplastsonto nonselective regeneration plates containing uracil anduridine Then the conidia from the nonselective plates wereplated-out on CD selectivemedium containing 5-FOA uraciland uridine This led to isolation of the mutant strain GR6pyrGΔ0 PCR based tests confirmed that the pyrG region ofstrain GR6 was deleted
It was possible to complement the uridine requirementof GR6 pyrGΔ0 by introducing the plasmids pUC pyrGAoand ANEp2 carrying A oryzae or A nidulans pyrG respec-tively This suggests that the AMA1 region of ANEp2 whichsupports autonomous plasmid replication in other fungi alsosupports autonomous replication in A oryzae Furthermorethe transformation frequency of ANEp2 was about 10-foldgreater than that obtained with the integrative plasmidpUC pyrGAo Transformation frequency ratios obtainedwith pUC pyrGAo as compared to plasmids harbouringAMA1 were in accordance with those obtained with inte-grating versus autonomously replicating plasmids [12] AlsoFierro et al [21] transformed P nalgiovense with integrativeversus autoreplicative plasmids and found 60-fold higherfrequencies for AMA1-based autonomously replicating plas-mids compared with integrative plasmids Autonomouslyreplicating plasmids carrying the AMA1 region from Anidulans transformed P nalgiovense very efficiently and theseplasmids also showed mitotic stability with low degrees ofreorganization120573-Galactosidase expression showed that the A oryzae
strain GR6 pyrGΔ0 was able to efficiently produce heterolo-gous secreted proteins using the A niger transcriptional and
translational controls introduced via ANEp2The strain GR6pyrGΔ0 transformed with ANEp2 containing lacA encodingA niger 120573-galactosidase was more than 150 times higher thanthat obtained with the untransformed strain The findingof this study is promising and this paves the way for theapplication of A oryzae strain GR6 pyrGΔ0 in heterologousprotein production given that this strain is amenable togenetic manipulation and that A oryzae is widely used inbiotechnology and the food industry
5 Conclusion
Mutant strain GR6 a pyrGΔ0 derivative of the A oryzaestrain S1 was constructed using targeted gene replacementThe pyrG auxotrophs were verified by assessing their abilityto grow on selective and nonselective media and PCRanalysis Plasmids pUC pyrGAo and ANEp2 harbouring thepyrG genes from A oryzae and A nidulans respectivelycomplemented the uracil and uridine requirement of strainGR6 Transformants of the A oryzae GR6 mutant carryingthe heterologous expression vector ANEp2 were able tofunctionally express a secreted 120573-galactosidase at high levels
Acknowledgments
This work was supported by the Ministry of Science Tech-nology and Innovation (MOSTI) Malaysia under the GrantUKM-MGI-NBD0014-2007 Natural Sciences and Engineer-ing Research Council of Canada Grant STPGP 336896 anda scholarship from the Canadian Commonwealth Exchangeprogram (Asia-Pacific) The authors would also like to thankProfessorWilliam J Broughton for the helpful discussion andsuggestions for improving the paper
The Scientific World Journal 7
References
[1] T Kobayashi K Abe K Asai et al ldquoGenomics of Aspergillusoryzaerdquo Bioscience Biotechnology and Biochemistry vol 71 no3 pp 646ndash670 2007
[2] O P Ward W M Qin J Dhanjoon J Ye and A Singh ldquoPhys-iology and biotechnology of Aspergillusrdquo Advances in AppliedMicrobiology vol 58 pp 1ndash75 2005
[3] M Machida K Asai M Sano et al ldquoGenome sequencing andanalysis of Aspergillus oryzaerdquo Nature vol 438 no 7071 pp1157ndash1161 2005
[4] J D Boeke F LaCroute andG R Fink ldquoA positive selection formutants lacking orotidine-51015840-phosphate decarboxylase activityin yeast 5-fluoro-orotic acid resistancerdquoMolecular and GeneralGenetics vol 197 no 2 pp 345ndash346 1984
[5] H Long T-H Wang and Y-K Zhang ldquoIsolation of Tricho-derma reesei pyrG negative Mutant by UV Mutagenesis and ItsApplication in Transformationrdquo Chemical Research in ChineseUniversities vol 24 no 5 pp 565ndash569 2008
[6] MHeerikhuisen CVan denHondel andP J Punt ldquoAspergillussojaerdquo in Production of Recombinant ProteinsmdashNovel Microbialand Eukaryotic Expression Systems pp 191ndash214 Wiley-VCHWeinheim Germany 2005
[7] S Kanamasa K Yamaoka T Kawaguchi J-I Sumitani and MAra ldquoTransformation of Aspergillus aculeatus using the drugresistance gene of Aspergillus oryzae and the pyrG gene ofAspergillus nidulansrdquo Bioscience Biotechnology and Biochem-istry vol 67 no 12 pp 2661ndash2663 2003
[8] R V Miller E J Ford N J Zidack and D C Sands ldquoApyrimidine auxotroph of Sclerotinia sclerotiorum for use inbiological weed controlrdquo Journal of General Microbiology vol135 pp 2085ndash2091 1989
[9] W Van Hartingsveldt I E Mattern and C M J Van ZeijlldquoDevelopment of a homologous transformation system forAspergillus niger based on the pyrG generdquoMolecular andGeneralGenetics vol 206 no 1 pp 71ndash75 1987
[10] D Mueller and U Stahl ldquoRNA a powerful tool for recombina-tion and regulated expression of genesrdquo in Progress in Botanypp 31ndash45 Springer Berlin Germany 2005
[11] J Aranda M Poza B G Pardo et al ldquoA rapid and simplemethod for constructing stable mutants of Acinetobacter bau-manniirdquo BMCMicrobiology vol 10 article 279 2010
[12] R Storms Y Zheng H Li S Sillaots AMartinez-Perez and ATsang ldquoPlasmid vectors for protein production gene expressionandmolecular manipulations inAspergillus nigerrdquo Plasmid vol53 no 3 pp 191ndash204 2005
[13] S S L Oh J M Leong A M A Murad N M Mahadi andF D Abu Bakar ldquoCloning and analysis of pyrG gene encod-ing orotidine 5-monophosphate decarboxylase of Aspergillusoryzae strain Slrdquo Sains Malaysiana vol 40 no 4 pp 331ndash3372011
[14] S Ushijima T Nakadai and K Uchida ldquoInterspecific electrofu-sion of protoplasts between Aspergillus oryzae and Aspergillussojaerdquo Agricultural and Biological Chemistry vol 55 pp 129ndash136 1991
[15] Y Iimura K Gomi H Uzu and S Hara ldquoTransformationof Aspergillus oryzae through plasmid-mediated complementa-tion of themethionine-auxotrophicmutationrdquoAgricultural andBiological Chemistry vol 51 pp 323ndash328 1987
[16] J J Feng T Takahashi M Machida and Y Koyama ldquoIdenti-fication of a basic helix-loop-helix-type transcription regulator
gene in Aspergillus oryzae by systematically deleting large chro-mosomal segmentsrdquo Applied and Environmental Microbiologyvol 75 no 18 pp 5943ndash5951 2009
[17] Y R Liew Protoplasting and cell wall regeneration of the filamen-tous fungi Aspergillus oryzae strain S1 [BSc thesis] UniversitiKebangsaan Malaysia Malaysia 2008
[18] K Wernars T Goosen B M Wennekes K Swart CA Vanden Hondel and HW Van den Broek ldquoCotransformationof Aspergillus nidulans a tool for replacing fungal genesrdquoMolecular and General Genetics vol 209 no 1 pp 71ndash77 1987
[19] E Kafer ldquoMeiotic andmitotic recombination in Aspergillus andits chromosomal aberrationsrdquo Advances in Genetics vol 19 pp33ndash131 1977
[20] ldquoLiquid media plates and overlayers for A nigerrdquo 2012 httpcloneconcordiacaaspergillusprotocolsnigmediahtml
[21] F Fierro F Laich R O Garcıa-Rico and J F Martın ldquoHighefficiency transformation of Penicillium nalgiovense with inte-grative and autonomously replicating plasmidsrdquo InternationalJournal of Food Microbiology vol 90 no 2 pp 237ndash248 2004
[22] ldquoUnitProtKBrdquo 2012 httpwwwuniprotorguniprotP29853[23] V L Prabha and N S Punekar ldquoGenetic transformation in
Aspergilli tools of the traderdquo Indian Journal of Biochemistry andBiophysics vol 41 no 5 pp 205ndash215 2004

2 The Scientific World Journal
Not I 12261
Ssp I 14084 Ssp I 424
Fse I 1488
Nhe I 4929
Mlu I 5444Aat II 5590
Not I 6994
AMA1
AppUC18
14000
12000
10000
8000 6000
4000
2000
ANEp214265bps
GlaTt
LacA
GlaPr
pyrG
(a)
1000
2000
3000
4000
Eco RI 396
Eco RI 2255
pyrG
AppUC19
pUC pyrGAo
4545bps
(b)
Figure 1 Plasmids used for transformation (a) Plasmid ANEp2 containing the pyrG gene of A nidulans and a GlaPr-lacA-GlaTt chimera(b) plasmid pUC pyrGAo containing pyrG (1839 bp) of A oryzae strain S1
pathway through mutational inactivation of orotidine-51015840-monophosphate (OMP) decarboxylase which catalyzes thedecarboxylation of orotidine monophosphate (OMP) toform uridine monophosphate (UMP) In filamentous fungipyrimidine auxotrophs obtained through inactivation of theOMP decarboxylase gene (usually designated pyrG) are aux-otrophic for uridine andor uracil and are resistant to 5-FOA[5 6] The generation of 5-FOA-resistant pyrG mutants hasbeen reported in other fungi including A aculeatus A nigerSclerotinia sclerotiorum and Trichoderma reesei [5 7ndash9]Most pyrGmutants were isolated following either ultraviolet(UV) or chemical mutagenesis Mutations induced in theseways are often unstable (ie they revert to the wild type)andmay includemultiple undesirablemutations In contrastmutants generated by targeted gene replacement are stable asthe targeted gene is removed from the genome It is howevermore difficult to mutate multiple genes this way [10 11]
A BLAST search using the pyrG sequence of A oryzaeS1 against the A oryzae RIB40 genome database at theNational Institute of Technology and Evaluation (NITEhttpwwwbionitegojpdogan) showed that pyrG is presentas a single copy on chromosome 7 (contig SC011 location2219667 2220565)
In this study a gene replacement cassette was constructedconsisting of the native pyrG locus extending 15 to 20 kbbeyond the upstream and downstream termini of the pyrGcoding region but lacking the open reading frame of the geneTransformation of A oryzae strain S1 with the gene replace-ment cassette generated 5-FOA-resistant uracil and uridineauxotrophs Putative pyrGΔ0 mutants were verified by com-paring their growth on selective and nonselective mediaPCR analyses of the pyrG region in the putative mutantsand complementation with plasmids containing wild-type
homologous and heterologous pyrG genes Suitability of thepyrGΔ0mutant as a host for heterologous protein productionwas assessed using strain GR6 harbouring a plasmid inwhich the pyrG and heterologous 120573-galactosidase genes wereintegrated
2 Materials and Methods
21 Fungus Cultivation and Plasmids A oryzae strain S1wasobtained from the Fungal Stock Culture Collection of theMolecular Mycology Laboratory School of Biosciences andBiotechnology Faculty of Science and Technology Uni-versiti Kebangsaan Malaysia The fungus was maintainedon a Potato Dextrose Agar (PDA Difco France) at 30∘Cand subcultured monthly Fungal cultivation was carriedout by inoculating onto a fresh PDA plate Cultures wereincubated for 7 days at 30∘C Plasmid ANEp2 (Figure 1(a))is a shuttle vector capable of autonomous replication inAspergilla [12] In addition to the pUC18 backbone ANEp2carries the A nidulans pyrG gene as a selectable markeran expression cassette (GlaPr-lacA-GlaTt) that expresses anA niger secreted 120573-galactosidase and the AMA1 sequencethat supports autonomous replication in Aspergilli ANEp2was used to functionally test for complementation of uridineauxotrophy and reporter gene expression in anA oryzae pyrGmutant Plasmid pUC pyrGAo containing the pyrG of the Aoryzae strain S1 was also used for complementation of uridineauxotrophy (Figure 1(b)) [13] Escherichia coli strain DH5120572was used for DNA manipulation
22 Media Polypeptone dextrin (PD) medium containing10 (wv) polypeptone 20 (wv) glucose 05 (wv)
The Scientific World Journal 3
Table 1 Primers used in this study
Designation Primer description Primer sequence51015840pyr pyrG 51015840 region sense primer 51015840-GGCGCCACAATCGTTGAGGTA-31015840lowastrevint-XhoI pyrG 51015840 region antisense primer the XhoI site is underlined 51015840-CTCGAGGTTGGCGATGGAGG-31015840lowastfwdint-XhoI pyrG 31015840 region sense primer the XhoI site is underlined 51015840-CTCGAGGTAGTGGTGGATACGTACT-31015840
31015840pyr pyrG 31015840 region antisense primer 51015840-AAGCTTATCAGCTGCATATCTCTAT-31015840
Pyr51015840-out pyrG 51015840 region sense primer outside the pyrG 51015840 genereplacement cassette 51015840-ATTTAGTCATTCCTTTTGGCATC-31015840
Pyr31015840-out pyrG 31015840 region antisense primer outside the pyrG 31015840 genereplacement cassette 51015840-CTAATTTTAATGTCTTCCTTCAG-31015840
lowastrevint-XhoI was used to amplify the targeting region downstream of the pyrG coding regionlowastfwdint-XhoI was used to amplify the targeting region upstream of the pyrG coding region
KH2PO4 01 (wv) NaNO
3 and 005 (wv) MgSO
4sdot7H2O
was used for liquid cultivation of A oryzae [14] Czapek-Dox (CD) medium composed of 02 (wv) NaNO
3 01
(wv) K2HPO4 005 (wv) MgSO
4sdot7H2O 005 (wv)
KCl 0001 (wv) FeSO4and 30 (wv) sucrose at pH 55
(supplemented with 02 (wv) uracil and 20mM uridine)was used for spheroplast cell wall regeneration Spheroplastswere overlaid with CD medium containing 05 (wv) agarand 50 (wv)NaCl ontoCDmediumcontaining 20 (wv)agar and 50 (wv) NaCl [15] Cultivation and selection ofpositive transformants were performed using CD minimalagar media [16] composed of 02 (wv) NaNO
3 01
(wv) KH2PO4 005 (wv) MgSO
4sdot7H2O 005 (wv) KCl
0002 (wv) FeSO4sdot7H2O 20 (wv) glucose 005 (wv)
NaCl and 2 (wv) agar at pH 55 containing 2mgmL 5-FOA 02 (wv) uracil and 20mM uridine
23 Replacement Cassette Construction The cassette used todelete theA oryzae pyrG (Figure 2) was assembled as followsgenomic DNA of A oryzae strain S1 was used as the PCRtemplate to amplify the 16 kb and 20 kb regions upstreamand downstream of the pyrG coding region respectivelyThe upstream region was amplified with the primers 51015840pyrand revint-XhoI while the downstream region was amplifiedusing the primers fwdint-XhoI and 31015840pyr (Table 1) BothPCR products were ligated into the vector pGEM-T Easy(Promega Fitchburg WI USA) Plasmids pGEM-51015840pyr andpGEM-31015840pyr were identified by restriction endonucleasedigestion and verified by sequencing Finally the two regionsflanking the pyrG coding regionwere assembled into the pyrGreplacement cassette by ligating the plasmids pGEM-51015840pyrand pGEM-31015840pyr digested with XhoI and SalI Restrictionendonuclease mapping and sequencing were used to verifythe construction of pGEM-51015840 31015840pyr
24 Spheroplast Preparation and Transformation Sphero-plasts were produced based on the methods of Ushijimaet al [14] and Liew [17] as follows A oryzae conidia (107conidiamL) were cultivated in 50mL PD at 30∘C for 20 hMycelia (3 g wet weight) were pretreated with DTT-lysisbuffer containing 10mM dithiothreitol (DTT) and 10mMTris-HCl for 1 h at 30∘C by shaking at 75 rpm prior to incuba-tion with 70mgmL lysing enzyme (Trichoderma harzianum
Sigma-Aldrich Corp St Louis MO USA) solution contain-ing osmotic buffer (08MMgSO
4in 001M phosphate buffer)
for 4 h at 30∘C (agitationmdash75 rpm) The mixture containingspheroplasts was transferred to a 50mL centrifuge tube(Greiner Bio-One UK) and 06M sorbitol was added slowlyto the mixture The mixture was spun at 4000 rpm 4∘C for30minThe interphase layer between the lysis buffer and sor-bitol was collected and transferred to a new 50mL tube withthe addition of STC buffer (12M sorbitol 001M Tris-HCl001M CaCl
2) Transformation was carried out according to
Storms et al [12] andWernars et al [18] using 16 times 106 viablespheroplasts in STC mixed with 10120583g purified PCR product(36 kb amplified DNA fragment consisting of the 51015840pyr and31015840pyr regions of the pyrG gene replacement cassette) and12M sorbitol as the osmotic stabilizer Subsequently 05mLaliquots containing 8 times 105 transformed spheroplasts weremixed into 05 CD overlay agar poured onto 2 CD agarplates supplemented with 02 uracil and 20mM uridineand incubated at 30∘C for 4ndash6 days
25 Screening and Analysis of Transformants 5-FOA resis-tant colonies were tested for auxotrophy by patching onCD medium and CD medium with uracil and uridineVerification that the 5-FOA-resistant auxotrophs arose viadeletion of the pyrG coding region was done by PCRanalysis of total DNA using primers Pyr51015840-out and Pyr31015840-out (Table 1) designed to be complementary to the knowngenomic (obtained from the A oryzae RIB40 NITE genomedatabase) sequence just beyond the upstream and down-stream boundaries of the replacement cassette The pyrGΔ0mutants were also tested by complementation with plasmidspUC pyrGAo and ANEp2 harbouring A oryzae and A nigerpyrG respectively
26 120573-Galactosidase Expression Levels of 120573-Galactosidaseactivity expressed by A oryzae strain GR6 pyrGΔ0 and GR6pyrGΔ0 transformants harbouring ANEp2 were determinedusing filtered medium from 7-day old static cultures Staticcultures were generated by inoculating 10 times 106 conid-iospores into Petri dishes containing 25mL of MMJ mediumas described by Kafer [19] using 15 maltose as the carbonsource (along with the necessary supplements) [20] at 30∘C
4 The Scientific World Journal
Digest with SalI and XhoI
Ligate
5998400pyr
pyrG upstream
XhoI 1639 fwdint XhoI
XhoI 62
pyrG downstream
SalI 2083
revint XhoI
pGEM-5998400pyr4608bp
pGEM-3998400pyr5008bp
3998400pyr
XhoI
5998400pyr
6585bppGEM-5998400 3998400pyr
SalI 1683
3998400pyr
pGEM-Treg EasypGEM-Treg Easy
pGEM-Treg Easy
Figure 2 Schematic diagram outlining assembly of the pyrG gene replacement cassette
120573-Galactosidase assays were performed by adding culture fil-trates to a reaction-buffer containing 50mM sodium acetate(pH 41) and 4mgmL of orthonitrophenyl-120573-D-galactoside(ONPG) and incubating for 10min at 37∘C Assays werestopped by adding 1 volume of 1M sodium carbonate andthe absorbance was read at 415 nm 120573-Galactosidase activityis expressed in units (U) per mL of culture where one unit isthe amount of enzyme needed to hydrolyze 1 120583moL of ONPGto orthonitrophenol per min at 37∘C Western blot analyseswere carried out using 20 120583L samples of concentrated culturefiltrate using ultra-filtration spin columns with a molecularweight cut-off of 10 kDa (ldquoVivaspinrdquo Sartorius GottingenGermany) and fractionated by 10 SDS-PAGE Fractionatedproteins were transferred to nitrocellulose membranes andvisualized using a rabbit anti-lac A antiserum an anti-rabbitIgG labelled with horseradish peroxidase (Promega) and achemiluminescent detection kit (Thermo Fisher ScientificWaltham MA USA) Rabbit serum containing polyclonalantibodies directed against A niger 120573-galactosidase wasgenerated as follows A niger 120573-galactosidase was expressedand purified by ion exchange chromatography Purified120573-galactosidase (200120583g with complete Freundrsquos Adjuvantfor immunization) was introduced by subcutaneous injec-tions into New Zealand white rabbits The initial injectionwas followed by two booster injections (100120583g purified120573-galactosidase with Incomplete Freundrsquos Adjuvant) For
serum preparation the collected blood was stored at 4∘Covernight and centrifuged for 30min at 4000 rpm Thecollected supernatant was stored at minus20∘C Enzyme-linkedimmunosorbent assay (ELISA) was used for the evaluationof antibody positive titer in the rabbit serum
3 Results
31 Deletion of the A oryzae Strain S1 pyrG The 36kb pyrGΔ0 gene replacement cassette was amplified usingprimers 51015840pyr and 31015840pyr and plasmid pGEM-51015840 31015840pyr as thetemplate (Figure 4(a)) Generation of a pyrGΔ0 derivativeof A oryzae was performed by transforming strain S1 withthe 36 kb pyrGΔ0 cassette Initial attempts to regeneratetransformed spheroplasts were performed on CD platessupplemented with uracil uridine and 5-FOAThis strategywhich selects for pyrGΔ0 gene replacements immediatelyafter transformation of spheroplasts with the PCR productis similar to the approach used to isolate pyrG mutantsof Trichoderma reesei (irradiated spores are platedout onminimal medium containing uridine and 5-FOAmdashLonget al [5]) However colonies derived from the experimentsdescribed here with pyrG failed to yield FOA resistantcolonies Perhaps 5-FOA-resistant pyrGΔ0 mutants werenot obtained because multiple nuclei were present in thespheroplasts and therefore a single pyrGΔ0 locus could
The Scientific World Journal 5
(a) Wild type (b) GR6 mutant
(c) CD plates (minimal medium)
(d) CD (uracil uridineand 5-FOA) plates
Figure 3 Media plates showing growth or no growth of the Aoryzae S1 wild type (column (a)) and GR6 pyrG mutant (column(b)) Column (a) ability of the wild type strain S1 (row (c)) andColumn (b) the inability of the GR6 pyrGmutant strain (row (c)) togrow on minimal medium CD plates Column (a) inability of thewild type strain S1 (row (d)) and Column (b) ability of the GR6pyrGmutant strain (row (d)) to grow onminimalmediumCDplatessupplemented with uracil uridine and 5-FOA
not prevent production of the suicide inhibitor FdUMPTo address this possibility the transformation protocol wasmodified by allowing nuclei in which the pyrG gene wasdeleted to segregate from wild-type nuclei by plating thetransformed spheroplasts (about 8 times 105 per plate) ontononselective regeneration plates containing uraciluridineOnce a lawn of conidia was produced the conidia werecollected diluted and plated at 8 times 106 spores per plateon CD selective medium with 5-FOA uracil and uridineTen randomly selected colonies were chosen from over 100colonies that formed and purified to homogeneity by tworounds of plating on CD uracil and uridine plates Conidiaprepared using the purified colonies were harvested andplated onto CD plates without uracil and uridine (106 conidiaper plate) Only one of the putative mutants designated GR6(Gene Replacement mutant number 6) did not grow and wastherefore assumed to be completely stable (Figure 3)
32 Verifying that GR6 Harbours a pyrGΔ0 Locus PCRamplifications with primers Pyr51015840-out and Pyr31015840-out usingthe genomic DNA of strain S1 and strain GR6 as the tem-plates showed that a 09 kb deletion was introduced into thepyrG region of strain GR6 (Figure 4(b)) which is consistentwith generation of the desired pyrGΔ0 locus by homologousrecombination
Then the pyrG auxotroph GR6 was transformed withplasmids pUC pyrGAo and ANEp2 These plasmids containthe wild-type pyrG from A oryzae (in pUC19) and Anidulans respectively Both plasmids were introduced intoand restored uracil and uridine prototrophy of GR6 withfrequencies of three transformants per 120583g of plasmid DNAfor pUC pyrGAo and 50 transformants per 120583g of plasmidDNA for ANEp2 Furthermore the frequency ratio of pyrGprototrophs obtained when transformation was performedwith the pUC pyrGAo versus the AMA plasmid was about01 This is in accordance with the range of prototrophy fre-quency ratios obtained with integrating versus autonomously
1kb 1
35kb
30kb sim36kb
(a)
50kb40kb35kb
sim45kb
sim36kb
1kb 1 2
(b)
Figure 4 Gel electrophoretic profile of the (a) (lane 1) genereplacement cassette consisting of the 51015840 and 31015840pyrG DNA inserts(without the coding region) with the size of sim36 kb and (b) genomicPCR product of the A oryzae S1 wild type strain (lane 1) and itsmutant derivative pyrGΔ0 strain GR6 (lane 2) Lane M 1 kb DNAladder (Fermentas Canada)
replicating plasmids previously reported with other fungalspecies [12 21]
33 Protein Expression Total secreted proteins in culturefiltrates of the untransformed strain GR6 pyrGΔ0 and GR6pyrGΔ0 transformed with ANEp2 were analyzed by SDS-PAGE (Figure 5(a)) and Western blotting (Figure 5(b))SDS-PAGE showed a very prominent 110 kDa proteinwhich is about the size expected of the A niger 120573-galactosidase (Swiss-Prot accession number P29853) (Unit-ProtKB httpwwwuniprotorguniprotP29853) [22] in theculture filtrate of ANEp2 GR6 pyrGΔ0 which was absentin the untransformed strain GR6 pyrGΔ0 Western blotanalyses confirmed that the major band was the A niger 120573-galactosidase As expected from the above culture filtratesof the ANEp2 GR6 pyrGΔ0 transformants possessed highlevels of 120573-galactosidase activity (135UmL) as compared tothe untransformed strain GR6 pyrGΔ0 which expressed 120573-galactosidase at 080UmL
4 Discussion
Replacement of a gene requires homologous recombinationof the introduced DNA with the genome gene replacementis achieved by a precise double crossover event [23] We firstsought these events by selecting for FOA-resistant clonesFOA inhibits the growth of the fungus but permits multipli-cation of cells deficient in OMP decarboxylase and becauseof this are auxotrophic for uridine Initially we selected forpyrGΔ0 gene replacements immediately after transfectionusing a similar plating procedure as has been used toisolate pyrG mutants of T reesei A aculeatus and Penicil-lium nalgiovense from irradiated spores [5 7 21] In ourhands this technique failed to yield FOA resistant colonies
6 The Scientific World Journal
M 1 2+ve
175kDa
80kDa
58kDa
46kDa
30kDa(a)
1 2+ve
110kDa
(b)
Figure 5 Analysis of GR6 culture filtrates Panel (a) lane M SDS-PAGE profile of prestained protein marker (New England Biolabs USA)lane +ve purified A niger 120573-galactosidase isolated from a culture filtrate of A niger transformed with the GlaPr-lacA-GlaTt expressioncassette lane 1 culture filtrate prepared using A oryzae strain GR6 pyrGΔ0 and lane 2 culture filtrate prepared using GR6 pyrGΔ0transformed with ANEp2 Panel (b) Western blot performed by probing the gel shown in Panel (a) with polyclonal rabbit antibodies directedagainst A niger 120573-galactosidase
This was probably because multiple nuclei were present ineach spheroplast and therefore a single pyrGΔ0 locus couldnot prevent production of the suicide inhibitor FdUMPTo test this the transformation protocol was modified toallow cells containing putative pyrG deletions to segregatefrom wild-type ones by plating the transformed spheroplastsonto nonselective regeneration plates containing uracil anduridine Then the conidia from the nonselective plates wereplated-out on CD selectivemedium containing 5-FOA uraciland uridine This led to isolation of the mutant strain GR6pyrGΔ0 PCR based tests confirmed that the pyrG region ofstrain GR6 was deleted
It was possible to complement the uridine requirementof GR6 pyrGΔ0 by introducing the plasmids pUC pyrGAoand ANEp2 carrying A oryzae or A nidulans pyrG respec-tively This suggests that the AMA1 region of ANEp2 whichsupports autonomous plasmid replication in other fungi alsosupports autonomous replication in A oryzae Furthermorethe transformation frequency of ANEp2 was about 10-foldgreater than that obtained with the integrative plasmidpUC pyrGAo Transformation frequency ratios obtainedwith pUC pyrGAo as compared to plasmids harbouringAMA1 were in accordance with those obtained with inte-grating versus autonomously replicating plasmids [12] AlsoFierro et al [21] transformed P nalgiovense with integrativeversus autoreplicative plasmids and found 60-fold higherfrequencies for AMA1-based autonomously replicating plas-mids compared with integrative plasmids Autonomouslyreplicating plasmids carrying the AMA1 region from Anidulans transformed P nalgiovense very efficiently and theseplasmids also showed mitotic stability with low degrees ofreorganization120573-Galactosidase expression showed that the A oryzae
strain GR6 pyrGΔ0 was able to efficiently produce heterolo-gous secreted proteins using the A niger transcriptional and
translational controls introduced via ANEp2The strain GR6pyrGΔ0 transformed with ANEp2 containing lacA encodingA niger 120573-galactosidase was more than 150 times higher thanthat obtained with the untransformed strain The findingof this study is promising and this paves the way for theapplication of A oryzae strain GR6 pyrGΔ0 in heterologousprotein production given that this strain is amenable togenetic manipulation and that A oryzae is widely used inbiotechnology and the food industry
5 Conclusion
Mutant strain GR6 a pyrGΔ0 derivative of the A oryzaestrain S1 was constructed using targeted gene replacementThe pyrG auxotrophs were verified by assessing their abilityto grow on selective and nonselective media and PCRanalysis Plasmids pUC pyrGAo and ANEp2 harbouring thepyrG genes from A oryzae and A nidulans respectivelycomplemented the uracil and uridine requirement of strainGR6 Transformants of the A oryzae GR6 mutant carryingthe heterologous expression vector ANEp2 were able tofunctionally express a secreted 120573-galactosidase at high levels
Acknowledgments
This work was supported by the Ministry of Science Tech-nology and Innovation (MOSTI) Malaysia under the GrantUKM-MGI-NBD0014-2007 Natural Sciences and Engineer-ing Research Council of Canada Grant STPGP 336896 anda scholarship from the Canadian Commonwealth Exchangeprogram (Asia-Pacific) The authors would also like to thankProfessorWilliam J Broughton for the helpful discussion andsuggestions for improving the paper
The Scientific World Journal 7
References
[1] T Kobayashi K Abe K Asai et al ldquoGenomics of Aspergillusoryzaerdquo Bioscience Biotechnology and Biochemistry vol 71 no3 pp 646ndash670 2007
[2] O P Ward W M Qin J Dhanjoon J Ye and A Singh ldquoPhys-iology and biotechnology of Aspergillusrdquo Advances in AppliedMicrobiology vol 58 pp 1ndash75 2005
[3] M Machida K Asai M Sano et al ldquoGenome sequencing andanalysis of Aspergillus oryzaerdquo Nature vol 438 no 7071 pp1157ndash1161 2005
[4] J D Boeke F LaCroute andG R Fink ldquoA positive selection formutants lacking orotidine-51015840-phosphate decarboxylase activityin yeast 5-fluoro-orotic acid resistancerdquoMolecular and GeneralGenetics vol 197 no 2 pp 345ndash346 1984
[5] H Long T-H Wang and Y-K Zhang ldquoIsolation of Tricho-derma reesei pyrG negative Mutant by UV Mutagenesis and ItsApplication in Transformationrdquo Chemical Research in ChineseUniversities vol 24 no 5 pp 565ndash569 2008
[6] MHeerikhuisen CVan denHondel andP J Punt ldquoAspergillussojaerdquo in Production of Recombinant ProteinsmdashNovel Microbialand Eukaryotic Expression Systems pp 191ndash214 Wiley-VCHWeinheim Germany 2005
[7] S Kanamasa K Yamaoka T Kawaguchi J-I Sumitani and MAra ldquoTransformation of Aspergillus aculeatus using the drugresistance gene of Aspergillus oryzae and the pyrG gene ofAspergillus nidulansrdquo Bioscience Biotechnology and Biochem-istry vol 67 no 12 pp 2661ndash2663 2003
[8] R V Miller E J Ford N J Zidack and D C Sands ldquoApyrimidine auxotroph of Sclerotinia sclerotiorum for use inbiological weed controlrdquo Journal of General Microbiology vol135 pp 2085ndash2091 1989
[9] W Van Hartingsveldt I E Mattern and C M J Van ZeijlldquoDevelopment of a homologous transformation system forAspergillus niger based on the pyrG generdquoMolecular andGeneralGenetics vol 206 no 1 pp 71ndash75 1987
[10] D Mueller and U Stahl ldquoRNA a powerful tool for recombina-tion and regulated expression of genesrdquo in Progress in Botanypp 31ndash45 Springer Berlin Germany 2005
[11] J Aranda M Poza B G Pardo et al ldquoA rapid and simplemethod for constructing stable mutants of Acinetobacter bau-manniirdquo BMCMicrobiology vol 10 article 279 2010
[12] R Storms Y Zheng H Li S Sillaots AMartinez-Perez and ATsang ldquoPlasmid vectors for protein production gene expressionandmolecular manipulations inAspergillus nigerrdquo Plasmid vol53 no 3 pp 191ndash204 2005
[13] S S L Oh J M Leong A M A Murad N M Mahadi andF D Abu Bakar ldquoCloning and analysis of pyrG gene encod-ing orotidine 5-monophosphate decarboxylase of Aspergillusoryzae strain Slrdquo Sains Malaysiana vol 40 no 4 pp 331ndash3372011
[14] S Ushijima T Nakadai and K Uchida ldquoInterspecific electrofu-sion of protoplasts between Aspergillus oryzae and Aspergillussojaerdquo Agricultural and Biological Chemistry vol 55 pp 129ndash136 1991
[15] Y Iimura K Gomi H Uzu and S Hara ldquoTransformationof Aspergillus oryzae through plasmid-mediated complementa-tion of themethionine-auxotrophicmutationrdquoAgricultural andBiological Chemistry vol 51 pp 323ndash328 1987
[16] J J Feng T Takahashi M Machida and Y Koyama ldquoIdenti-fication of a basic helix-loop-helix-type transcription regulator
gene in Aspergillus oryzae by systematically deleting large chro-mosomal segmentsrdquo Applied and Environmental Microbiologyvol 75 no 18 pp 5943ndash5951 2009
[17] Y R Liew Protoplasting and cell wall regeneration of the filamen-tous fungi Aspergillus oryzae strain S1 [BSc thesis] UniversitiKebangsaan Malaysia Malaysia 2008
[18] K Wernars T Goosen B M Wennekes K Swart CA Vanden Hondel and HW Van den Broek ldquoCotransformationof Aspergillus nidulans a tool for replacing fungal genesrdquoMolecular and General Genetics vol 209 no 1 pp 71ndash77 1987
[19] E Kafer ldquoMeiotic andmitotic recombination in Aspergillus andits chromosomal aberrationsrdquo Advances in Genetics vol 19 pp33ndash131 1977
[20] ldquoLiquid media plates and overlayers for A nigerrdquo 2012 httpcloneconcordiacaaspergillusprotocolsnigmediahtml
[21] F Fierro F Laich R O Garcıa-Rico and J F Martın ldquoHighefficiency transformation of Penicillium nalgiovense with inte-grative and autonomously replicating plasmidsrdquo InternationalJournal of Food Microbiology vol 90 no 2 pp 237ndash248 2004
[22] ldquoUnitProtKBrdquo 2012 httpwwwuniprotorguniprotP29853[23] V L Prabha and N S Punekar ldquoGenetic transformation in
Aspergilli tools of the traderdquo Indian Journal of Biochemistry andBiophysics vol 41 no 5 pp 205ndash215 2004

The Scientific World Journal 3
Table 1 Primers used in this study
Designation Primer description Primer sequence51015840pyr pyrG 51015840 region sense primer 51015840-GGCGCCACAATCGTTGAGGTA-31015840lowastrevint-XhoI pyrG 51015840 region antisense primer the XhoI site is underlined 51015840-CTCGAGGTTGGCGATGGAGG-31015840lowastfwdint-XhoI pyrG 31015840 region sense primer the XhoI site is underlined 51015840-CTCGAGGTAGTGGTGGATACGTACT-31015840
31015840pyr pyrG 31015840 region antisense primer 51015840-AAGCTTATCAGCTGCATATCTCTAT-31015840
Pyr51015840-out pyrG 51015840 region sense primer outside the pyrG 51015840 genereplacement cassette 51015840-ATTTAGTCATTCCTTTTGGCATC-31015840
Pyr31015840-out pyrG 31015840 region antisense primer outside the pyrG 31015840 genereplacement cassette 51015840-CTAATTTTAATGTCTTCCTTCAG-31015840
lowastrevint-XhoI was used to amplify the targeting region downstream of the pyrG coding regionlowastfwdint-XhoI was used to amplify the targeting region upstream of the pyrG coding region
KH2PO4 01 (wv) NaNO
3 and 005 (wv) MgSO
4sdot7H2O
was used for liquid cultivation of A oryzae [14] Czapek-Dox (CD) medium composed of 02 (wv) NaNO
3 01
(wv) K2HPO4 005 (wv) MgSO
4sdot7H2O 005 (wv)
KCl 0001 (wv) FeSO4and 30 (wv) sucrose at pH 55
(supplemented with 02 (wv) uracil and 20mM uridine)was used for spheroplast cell wall regeneration Spheroplastswere overlaid with CD medium containing 05 (wv) agarand 50 (wv)NaCl ontoCDmediumcontaining 20 (wv)agar and 50 (wv) NaCl [15] Cultivation and selection ofpositive transformants were performed using CD minimalagar media [16] composed of 02 (wv) NaNO
3 01
(wv) KH2PO4 005 (wv) MgSO
4sdot7H2O 005 (wv) KCl
0002 (wv) FeSO4sdot7H2O 20 (wv) glucose 005 (wv)
NaCl and 2 (wv) agar at pH 55 containing 2mgmL 5-FOA 02 (wv) uracil and 20mM uridine
23 Replacement Cassette Construction The cassette used todelete theA oryzae pyrG (Figure 2) was assembled as followsgenomic DNA of A oryzae strain S1 was used as the PCRtemplate to amplify the 16 kb and 20 kb regions upstreamand downstream of the pyrG coding region respectivelyThe upstream region was amplified with the primers 51015840pyrand revint-XhoI while the downstream region was amplifiedusing the primers fwdint-XhoI and 31015840pyr (Table 1) BothPCR products were ligated into the vector pGEM-T Easy(Promega Fitchburg WI USA) Plasmids pGEM-51015840pyr andpGEM-31015840pyr were identified by restriction endonucleasedigestion and verified by sequencing Finally the two regionsflanking the pyrG coding regionwere assembled into the pyrGreplacement cassette by ligating the plasmids pGEM-51015840pyrand pGEM-31015840pyr digested with XhoI and SalI Restrictionendonuclease mapping and sequencing were used to verifythe construction of pGEM-51015840 31015840pyr
24 Spheroplast Preparation and Transformation Sphero-plasts were produced based on the methods of Ushijimaet al [14] and Liew [17] as follows A oryzae conidia (107conidiamL) were cultivated in 50mL PD at 30∘C for 20 hMycelia (3 g wet weight) were pretreated with DTT-lysisbuffer containing 10mM dithiothreitol (DTT) and 10mMTris-HCl for 1 h at 30∘C by shaking at 75 rpm prior to incuba-tion with 70mgmL lysing enzyme (Trichoderma harzianum
Sigma-Aldrich Corp St Louis MO USA) solution contain-ing osmotic buffer (08MMgSO
4in 001M phosphate buffer)
for 4 h at 30∘C (agitationmdash75 rpm) The mixture containingspheroplasts was transferred to a 50mL centrifuge tube(Greiner Bio-One UK) and 06M sorbitol was added slowlyto the mixture The mixture was spun at 4000 rpm 4∘C for30minThe interphase layer between the lysis buffer and sor-bitol was collected and transferred to a new 50mL tube withthe addition of STC buffer (12M sorbitol 001M Tris-HCl001M CaCl
2) Transformation was carried out according to
Storms et al [12] andWernars et al [18] using 16 times 106 viablespheroplasts in STC mixed with 10120583g purified PCR product(36 kb amplified DNA fragment consisting of the 51015840pyr and31015840pyr regions of the pyrG gene replacement cassette) and12M sorbitol as the osmotic stabilizer Subsequently 05mLaliquots containing 8 times 105 transformed spheroplasts weremixed into 05 CD overlay agar poured onto 2 CD agarplates supplemented with 02 uracil and 20mM uridineand incubated at 30∘C for 4ndash6 days
25 Screening and Analysis of Transformants 5-FOA resis-tant colonies were tested for auxotrophy by patching onCD medium and CD medium with uracil and uridineVerification that the 5-FOA-resistant auxotrophs arose viadeletion of the pyrG coding region was done by PCRanalysis of total DNA using primers Pyr51015840-out and Pyr31015840-out (Table 1) designed to be complementary to the knowngenomic (obtained from the A oryzae RIB40 NITE genomedatabase) sequence just beyond the upstream and down-stream boundaries of the replacement cassette The pyrGΔ0mutants were also tested by complementation with plasmidspUC pyrGAo and ANEp2 harbouring A oryzae and A nigerpyrG respectively
26 120573-Galactosidase Expression Levels of 120573-Galactosidaseactivity expressed by A oryzae strain GR6 pyrGΔ0 and GR6pyrGΔ0 transformants harbouring ANEp2 were determinedusing filtered medium from 7-day old static cultures Staticcultures were generated by inoculating 10 times 106 conid-iospores into Petri dishes containing 25mL of MMJ mediumas described by Kafer [19] using 15 maltose as the carbonsource (along with the necessary supplements) [20] at 30∘C
4 The Scientific World Journal
Digest with SalI and XhoI
Ligate
5998400pyr
pyrG upstream
XhoI 1639 fwdint XhoI
XhoI 62
pyrG downstream
SalI 2083
revint XhoI
pGEM-5998400pyr4608bp
pGEM-3998400pyr5008bp
3998400pyr
XhoI
5998400pyr
6585bppGEM-5998400 3998400pyr
SalI 1683
3998400pyr
pGEM-Treg EasypGEM-Treg Easy
pGEM-Treg Easy
Figure 2 Schematic diagram outlining assembly of the pyrG gene replacement cassette
120573-Galactosidase assays were performed by adding culture fil-trates to a reaction-buffer containing 50mM sodium acetate(pH 41) and 4mgmL of orthonitrophenyl-120573-D-galactoside(ONPG) and incubating for 10min at 37∘C Assays werestopped by adding 1 volume of 1M sodium carbonate andthe absorbance was read at 415 nm 120573-Galactosidase activityis expressed in units (U) per mL of culture where one unit isthe amount of enzyme needed to hydrolyze 1 120583moL of ONPGto orthonitrophenol per min at 37∘C Western blot analyseswere carried out using 20 120583L samples of concentrated culturefiltrate using ultra-filtration spin columns with a molecularweight cut-off of 10 kDa (ldquoVivaspinrdquo Sartorius GottingenGermany) and fractionated by 10 SDS-PAGE Fractionatedproteins were transferred to nitrocellulose membranes andvisualized using a rabbit anti-lac A antiserum an anti-rabbitIgG labelled with horseradish peroxidase (Promega) and achemiluminescent detection kit (Thermo Fisher ScientificWaltham MA USA) Rabbit serum containing polyclonalantibodies directed against A niger 120573-galactosidase wasgenerated as follows A niger 120573-galactosidase was expressedand purified by ion exchange chromatography Purified120573-galactosidase (200120583g with complete Freundrsquos Adjuvantfor immunization) was introduced by subcutaneous injec-tions into New Zealand white rabbits The initial injectionwas followed by two booster injections (100120583g purified120573-galactosidase with Incomplete Freundrsquos Adjuvant) For
serum preparation the collected blood was stored at 4∘Covernight and centrifuged for 30min at 4000 rpm Thecollected supernatant was stored at minus20∘C Enzyme-linkedimmunosorbent assay (ELISA) was used for the evaluationof antibody positive titer in the rabbit serum
3 Results
31 Deletion of the A oryzae Strain S1 pyrG The 36kb pyrGΔ0 gene replacement cassette was amplified usingprimers 51015840pyr and 31015840pyr and plasmid pGEM-51015840 31015840pyr as thetemplate (Figure 4(a)) Generation of a pyrGΔ0 derivativeof A oryzae was performed by transforming strain S1 withthe 36 kb pyrGΔ0 cassette Initial attempts to regeneratetransformed spheroplasts were performed on CD platessupplemented with uracil uridine and 5-FOAThis strategywhich selects for pyrGΔ0 gene replacements immediatelyafter transformation of spheroplasts with the PCR productis similar to the approach used to isolate pyrG mutantsof Trichoderma reesei (irradiated spores are platedout onminimal medium containing uridine and 5-FOAmdashLonget al [5]) However colonies derived from the experimentsdescribed here with pyrG failed to yield FOA resistantcolonies Perhaps 5-FOA-resistant pyrGΔ0 mutants werenot obtained because multiple nuclei were present in thespheroplasts and therefore a single pyrGΔ0 locus could
The Scientific World Journal 5
(a) Wild type (b) GR6 mutant
(c) CD plates (minimal medium)
(d) CD (uracil uridineand 5-FOA) plates
Figure 3 Media plates showing growth or no growth of the Aoryzae S1 wild type (column (a)) and GR6 pyrG mutant (column(b)) Column (a) ability of the wild type strain S1 (row (c)) andColumn (b) the inability of the GR6 pyrGmutant strain (row (c)) togrow on minimal medium CD plates Column (a) inability of thewild type strain S1 (row (d)) and Column (b) ability of the GR6pyrGmutant strain (row (d)) to grow onminimalmediumCDplatessupplemented with uracil uridine and 5-FOA
not prevent production of the suicide inhibitor FdUMPTo address this possibility the transformation protocol wasmodified by allowing nuclei in which the pyrG gene wasdeleted to segregate from wild-type nuclei by plating thetransformed spheroplasts (about 8 times 105 per plate) ontononselective regeneration plates containing uraciluridineOnce a lawn of conidia was produced the conidia werecollected diluted and plated at 8 times 106 spores per plateon CD selective medium with 5-FOA uracil and uridineTen randomly selected colonies were chosen from over 100colonies that formed and purified to homogeneity by tworounds of plating on CD uracil and uridine plates Conidiaprepared using the purified colonies were harvested andplated onto CD plates without uracil and uridine (106 conidiaper plate) Only one of the putative mutants designated GR6(Gene Replacement mutant number 6) did not grow and wastherefore assumed to be completely stable (Figure 3)
32 Verifying that GR6 Harbours a pyrGΔ0 Locus PCRamplifications with primers Pyr51015840-out and Pyr31015840-out usingthe genomic DNA of strain S1 and strain GR6 as the tem-plates showed that a 09 kb deletion was introduced into thepyrG region of strain GR6 (Figure 4(b)) which is consistentwith generation of the desired pyrGΔ0 locus by homologousrecombination
Then the pyrG auxotroph GR6 was transformed withplasmids pUC pyrGAo and ANEp2 These plasmids containthe wild-type pyrG from A oryzae (in pUC19) and Anidulans respectively Both plasmids were introduced intoand restored uracil and uridine prototrophy of GR6 withfrequencies of three transformants per 120583g of plasmid DNAfor pUC pyrGAo and 50 transformants per 120583g of plasmidDNA for ANEp2 Furthermore the frequency ratio of pyrGprototrophs obtained when transformation was performedwith the pUC pyrGAo versus the AMA plasmid was about01 This is in accordance with the range of prototrophy fre-quency ratios obtained with integrating versus autonomously
1kb 1
35kb
30kb sim36kb
(a)
50kb40kb35kb
sim45kb
sim36kb
1kb 1 2
(b)
Figure 4 Gel electrophoretic profile of the (a) (lane 1) genereplacement cassette consisting of the 51015840 and 31015840pyrG DNA inserts(without the coding region) with the size of sim36 kb and (b) genomicPCR product of the A oryzae S1 wild type strain (lane 1) and itsmutant derivative pyrGΔ0 strain GR6 (lane 2) Lane M 1 kb DNAladder (Fermentas Canada)
replicating plasmids previously reported with other fungalspecies [12 21]
33 Protein Expression Total secreted proteins in culturefiltrates of the untransformed strain GR6 pyrGΔ0 and GR6pyrGΔ0 transformed with ANEp2 were analyzed by SDS-PAGE (Figure 5(a)) and Western blotting (Figure 5(b))SDS-PAGE showed a very prominent 110 kDa proteinwhich is about the size expected of the A niger 120573-galactosidase (Swiss-Prot accession number P29853) (Unit-ProtKB httpwwwuniprotorguniprotP29853) [22] in theculture filtrate of ANEp2 GR6 pyrGΔ0 which was absentin the untransformed strain GR6 pyrGΔ0 Western blotanalyses confirmed that the major band was the A niger 120573-galactosidase As expected from the above culture filtratesof the ANEp2 GR6 pyrGΔ0 transformants possessed highlevels of 120573-galactosidase activity (135UmL) as compared tothe untransformed strain GR6 pyrGΔ0 which expressed 120573-galactosidase at 080UmL
4 Discussion
Replacement of a gene requires homologous recombinationof the introduced DNA with the genome gene replacementis achieved by a precise double crossover event [23] We firstsought these events by selecting for FOA-resistant clonesFOA inhibits the growth of the fungus but permits multipli-cation of cells deficient in OMP decarboxylase and becauseof this are auxotrophic for uridine Initially we selected forpyrGΔ0 gene replacements immediately after transfectionusing a similar plating procedure as has been used toisolate pyrG mutants of T reesei A aculeatus and Penicil-lium nalgiovense from irradiated spores [5 7 21] In ourhands this technique failed to yield FOA resistant colonies
6 The Scientific World Journal
M 1 2+ve
175kDa
80kDa
58kDa
46kDa
30kDa(a)
1 2+ve
110kDa
(b)
Figure 5 Analysis of GR6 culture filtrates Panel (a) lane M SDS-PAGE profile of prestained protein marker (New England Biolabs USA)lane +ve purified A niger 120573-galactosidase isolated from a culture filtrate of A niger transformed with the GlaPr-lacA-GlaTt expressioncassette lane 1 culture filtrate prepared using A oryzae strain GR6 pyrGΔ0 and lane 2 culture filtrate prepared using GR6 pyrGΔ0transformed with ANEp2 Panel (b) Western blot performed by probing the gel shown in Panel (a) with polyclonal rabbit antibodies directedagainst A niger 120573-galactosidase
This was probably because multiple nuclei were present ineach spheroplast and therefore a single pyrGΔ0 locus couldnot prevent production of the suicide inhibitor FdUMPTo test this the transformation protocol was modified toallow cells containing putative pyrG deletions to segregatefrom wild-type ones by plating the transformed spheroplastsonto nonselective regeneration plates containing uracil anduridine Then the conidia from the nonselective plates wereplated-out on CD selectivemedium containing 5-FOA uraciland uridine This led to isolation of the mutant strain GR6pyrGΔ0 PCR based tests confirmed that the pyrG region ofstrain GR6 was deleted
It was possible to complement the uridine requirementof GR6 pyrGΔ0 by introducing the plasmids pUC pyrGAoand ANEp2 carrying A oryzae or A nidulans pyrG respec-tively This suggests that the AMA1 region of ANEp2 whichsupports autonomous plasmid replication in other fungi alsosupports autonomous replication in A oryzae Furthermorethe transformation frequency of ANEp2 was about 10-foldgreater than that obtained with the integrative plasmidpUC pyrGAo Transformation frequency ratios obtainedwith pUC pyrGAo as compared to plasmids harbouringAMA1 were in accordance with those obtained with inte-grating versus autonomously replicating plasmids [12] AlsoFierro et al [21] transformed P nalgiovense with integrativeversus autoreplicative plasmids and found 60-fold higherfrequencies for AMA1-based autonomously replicating plas-mids compared with integrative plasmids Autonomouslyreplicating plasmids carrying the AMA1 region from Anidulans transformed P nalgiovense very efficiently and theseplasmids also showed mitotic stability with low degrees ofreorganization120573-Galactosidase expression showed that the A oryzae
strain GR6 pyrGΔ0 was able to efficiently produce heterolo-gous secreted proteins using the A niger transcriptional and
translational controls introduced via ANEp2The strain GR6pyrGΔ0 transformed with ANEp2 containing lacA encodingA niger 120573-galactosidase was more than 150 times higher thanthat obtained with the untransformed strain The findingof this study is promising and this paves the way for theapplication of A oryzae strain GR6 pyrGΔ0 in heterologousprotein production given that this strain is amenable togenetic manipulation and that A oryzae is widely used inbiotechnology and the food industry
5 Conclusion
Mutant strain GR6 a pyrGΔ0 derivative of the A oryzaestrain S1 was constructed using targeted gene replacementThe pyrG auxotrophs were verified by assessing their abilityto grow on selective and nonselective media and PCRanalysis Plasmids pUC pyrGAo and ANEp2 harbouring thepyrG genes from A oryzae and A nidulans respectivelycomplemented the uracil and uridine requirement of strainGR6 Transformants of the A oryzae GR6 mutant carryingthe heterologous expression vector ANEp2 were able tofunctionally express a secreted 120573-galactosidase at high levels
Acknowledgments
This work was supported by the Ministry of Science Tech-nology and Innovation (MOSTI) Malaysia under the GrantUKM-MGI-NBD0014-2007 Natural Sciences and Engineer-ing Research Council of Canada Grant STPGP 336896 anda scholarship from the Canadian Commonwealth Exchangeprogram (Asia-Pacific) The authors would also like to thankProfessorWilliam J Broughton for the helpful discussion andsuggestions for improving the paper
The Scientific World Journal 7
References
[1] T Kobayashi K Abe K Asai et al ldquoGenomics of Aspergillusoryzaerdquo Bioscience Biotechnology and Biochemistry vol 71 no3 pp 646ndash670 2007
[2] O P Ward W M Qin J Dhanjoon J Ye and A Singh ldquoPhys-iology and biotechnology of Aspergillusrdquo Advances in AppliedMicrobiology vol 58 pp 1ndash75 2005
[3] M Machida K Asai M Sano et al ldquoGenome sequencing andanalysis of Aspergillus oryzaerdquo Nature vol 438 no 7071 pp1157ndash1161 2005
[4] J D Boeke F LaCroute andG R Fink ldquoA positive selection formutants lacking orotidine-51015840-phosphate decarboxylase activityin yeast 5-fluoro-orotic acid resistancerdquoMolecular and GeneralGenetics vol 197 no 2 pp 345ndash346 1984
[5] H Long T-H Wang and Y-K Zhang ldquoIsolation of Tricho-derma reesei pyrG negative Mutant by UV Mutagenesis and ItsApplication in Transformationrdquo Chemical Research in ChineseUniversities vol 24 no 5 pp 565ndash569 2008
[6] MHeerikhuisen CVan denHondel andP J Punt ldquoAspergillussojaerdquo in Production of Recombinant ProteinsmdashNovel Microbialand Eukaryotic Expression Systems pp 191ndash214 Wiley-VCHWeinheim Germany 2005
[7] S Kanamasa K Yamaoka T Kawaguchi J-I Sumitani and MAra ldquoTransformation of Aspergillus aculeatus using the drugresistance gene of Aspergillus oryzae and the pyrG gene ofAspergillus nidulansrdquo Bioscience Biotechnology and Biochem-istry vol 67 no 12 pp 2661ndash2663 2003
[8] R V Miller E J Ford N J Zidack and D C Sands ldquoApyrimidine auxotroph of Sclerotinia sclerotiorum for use inbiological weed controlrdquo Journal of General Microbiology vol135 pp 2085ndash2091 1989
[9] W Van Hartingsveldt I E Mattern and C M J Van ZeijlldquoDevelopment of a homologous transformation system forAspergillus niger based on the pyrG generdquoMolecular andGeneralGenetics vol 206 no 1 pp 71ndash75 1987
[10] D Mueller and U Stahl ldquoRNA a powerful tool for recombina-tion and regulated expression of genesrdquo in Progress in Botanypp 31ndash45 Springer Berlin Germany 2005
[11] J Aranda M Poza B G Pardo et al ldquoA rapid and simplemethod for constructing stable mutants of Acinetobacter bau-manniirdquo BMCMicrobiology vol 10 article 279 2010
[12] R Storms Y Zheng H Li S Sillaots AMartinez-Perez and ATsang ldquoPlasmid vectors for protein production gene expressionandmolecular manipulations inAspergillus nigerrdquo Plasmid vol53 no 3 pp 191ndash204 2005
[13] S S L Oh J M Leong A M A Murad N M Mahadi andF D Abu Bakar ldquoCloning and analysis of pyrG gene encod-ing orotidine 5-monophosphate decarboxylase of Aspergillusoryzae strain Slrdquo Sains Malaysiana vol 40 no 4 pp 331ndash3372011
[14] S Ushijima T Nakadai and K Uchida ldquoInterspecific electrofu-sion of protoplasts between Aspergillus oryzae and Aspergillussojaerdquo Agricultural and Biological Chemistry vol 55 pp 129ndash136 1991
[15] Y Iimura K Gomi H Uzu and S Hara ldquoTransformationof Aspergillus oryzae through plasmid-mediated complementa-tion of themethionine-auxotrophicmutationrdquoAgricultural andBiological Chemistry vol 51 pp 323ndash328 1987
[16] J J Feng T Takahashi M Machida and Y Koyama ldquoIdenti-fication of a basic helix-loop-helix-type transcription regulator
gene in Aspergillus oryzae by systematically deleting large chro-mosomal segmentsrdquo Applied and Environmental Microbiologyvol 75 no 18 pp 5943ndash5951 2009
[17] Y R Liew Protoplasting and cell wall regeneration of the filamen-tous fungi Aspergillus oryzae strain S1 [BSc thesis] UniversitiKebangsaan Malaysia Malaysia 2008
[18] K Wernars T Goosen B M Wennekes K Swart CA Vanden Hondel and HW Van den Broek ldquoCotransformationof Aspergillus nidulans a tool for replacing fungal genesrdquoMolecular and General Genetics vol 209 no 1 pp 71ndash77 1987
[19] E Kafer ldquoMeiotic andmitotic recombination in Aspergillus andits chromosomal aberrationsrdquo Advances in Genetics vol 19 pp33ndash131 1977
[20] ldquoLiquid media plates and overlayers for A nigerrdquo 2012 httpcloneconcordiacaaspergillusprotocolsnigmediahtml
[21] F Fierro F Laich R O Garcıa-Rico and J F Martın ldquoHighefficiency transformation of Penicillium nalgiovense with inte-grative and autonomously replicating plasmidsrdquo InternationalJournal of Food Microbiology vol 90 no 2 pp 237ndash248 2004
[22] ldquoUnitProtKBrdquo 2012 httpwwwuniprotorguniprotP29853[23] V L Prabha and N S Punekar ldquoGenetic transformation in
Aspergilli tools of the traderdquo Indian Journal of Biochemistry andBiophysics vol 41 no 5 pp 205ndash215 2004

4 The Scientific World Journal
Digest with SalI and XhoI
Ligate
5998400pyr
pyrG upstream
XhoI 1639 fwdint XhoI
XhoI 62
pyrG downstream
SalI 2083
revint XhoI
pGEM-5998400pyr4608bp
pGEM-3998400pyr5008bp
3998400pyr
XhoI
5998400pyr
6585bppGEM-5998400 3998400pyr
SalI 1683
3998400pyr
pGEM-Treg EasypGEM-Treg Easy
pGEM-Treg Easy
Figure 2 Schematic diagram outlining assembly of the pyrG gene replacement cassette
120573-Galactosidase assays were performed by adding culture fil-trates to a reaction-buffer containing 50mM sodium acetate(pH 41) and 4mgmL of orthonitrophenyl-120573-D-galactoside(ONPG) and incubating for 10min at 37∘C Assays werestopped by adding 1 volume of 1M sodium carbonate andthe absorbance was read at 415 nm 120573-Galactosidase activityis expressed in units (U) per mL of culture where one unit isthe amount of enzyme needed to hydrolyze 1 120583moL of ONPGto orthonitrophenol per min at 37∘C Western blot analyseswere carried out using 20 120583L samples of concentrated culturefiltrate using ultra-filtration spin columns with a molecularweight cut-off of 10 kDa (ldquoVivaspinrdquo Sartorius GottingenGermany) and fractionated by 10 SDS-PAGE Fractionatedproteins were transferred to nitrocellulose membranes andvisualized using a rabbit anti-lac A antiserum an anti-rabbitIgG labelled with horseradish peroxidase (Promega) and achemiluminescent detection kit (Thermo Fisher ScientificWaltham MA USA) Rabbit serum containing polyclonalantibodies directed against A niger 120573-galactosidase wasgenerated as follows A niger 120573-galactosidase was expressedand purified by ion exchange chromatography Purified120573-galactosidase (200120583g with complete Freundrsquos Adjuvantfor immunization) was introduced by subcutaneous injec-tions into New Zealand white rabbits The initial injectionwas followed by two booster injections (100120583g purified120573-galactosidase with Incomplete Freundrsquos Adjuvant) For
serum preparation the collected blood was stored at 4∘Covernight and centrifuged for 30min at 4000 rpm Thecollected supernatant was stored at minus20∘C Enzyme-linkedimmunosorbent assay (ELISA) was used for the evaluationof antibody positive titer in the rabbit serum
3 Results
31 Deletion of the A oryzae Strain S1 pyrG The 36kb pyrGΔ0 gene replacement cassette was amplified usingprimers 51015840pyr and 31015840pyr and plasmid pGEM-51015840 31015840pyr as thetemplate (Figure 4(a)) Generation of a pyrGΔ0 derivativeof A oryzae was performed by transforming strain S1 withthe 36 kb pyrGΔ0 cassette Initial attempts to regeneratetransformed spheroplasts were performed on CD platessupplemented with uracil uridine and 5-FOAThis strategywhich selects for pyrGΔ0 gene replacements immediatelyafter transformation of spheroplasts with the PCR productis similar to the approach used to isolate pyrG mutantsof Trichoderma reesei (irradiated spores are platedout onminimal medium containing uridine and 5-FOAmdashLonget al [5]) However colonies derived from the experimentsdescribed here with pyrG failed to yield FOA resistantcolonies Perhaps 5-FOA-resistant pyrGΔ0 mutants werenot obtained because multiple nuclei were present in thespheroplasts and therefore a single pyrGΔ0 locus could
The Scientific World Journal 5
(a) Wild type (b) GR6 mutant
(c) CD plates (minimal medium)
(d) CD (uracil uridineand 5-FOA) plates
Figure 3 Media plates showing growth or no growth of the Aoryzae S1 wild type (column (a)) and GR6 pyrG mutant (column(b)) Column (a) ability of the wild type strain S1 (row (c)) andColumn (b) the inability of the GR6 pyrGmutant strain (row (c)) togrow on minimal medium CD plates Column (a) inability of thewild type strain S1 (row (d)) and Column (b) ability of the GR6pyrGmutant strain (row (d)) to grow onminimalmediumCDplatessupplemented with uracil uridine and 5-FOA
not prevent production of the suicide inhibitor FdUMPTo address this possibility the transformation protocol wasmodified by allowing nuclei in which the pyrG gene wasdeleted to segregate from wild-type nuclei by plating thetransformed spheroplasts (about 8 times 105 per plate) ontononselective regeneration plates containing uraciluridineOnce a lawn of conidia was produced the conidia werecollected diluted and plated at 8 times 106 spores per plateon CD selective medium with 5-FOA uracil and uridineTen randomly selected colonies were chosen from over 100colonies that formed and purified to homogeneity by tworounds of plating on CD uracil and uridine plates Conidiaprepared using the purified colonies were harvested andplated onto CD plates without uracil and uridine (106 conidiaper plate) Only one of the putative mutants designated GR6(Gene Replacement mutant number 6) did not grow and wastherefore assumed to be completely stable (Figure 3)
32 Verifying that GR6 Harbours a pyrGΔ0 Locus PCRamplifications with primers Pyr51015840-out and Pyr31015840-out usingthe genomic DNA of strain S1 and strain GR6 as the tem-plates showed that a 09 kb deletion was introduced into thepyrG region of strain GR6 (Figure 4(b)) which is consistentwith generation of the desired pyrGΔ0 locus by homologousrecombination
Then the pyrG auxotroph GR6 was transformed withplasmids pUC pyrGAo and ANEp2 These plasmids containthe wild-type pyrG from A oryzae (in pUC19) and Anidulans respectively Both plasmids were introduced intoand restored uracil and uridine prototrophy of GR6 withfrequencies of three transformants per 120583g of plasmid DNAfor pUC pyrGAo and 50 transformants per 120583g of plasmidDNA for ANEp2 Furthermore the frequency ratio of pyrGprototrophs obtained when transformation was performedwith the pUC pyrGAo versus the AMA plasmid was about01 This is in accordance with the range of prototrophy fre-quency ratios obtained with integrating versus autonomously
1kb 1
35kb
30kb sim36kb
(a)
50kb40kb35kb
sim45kb
sim36kb
1kb 1 2
(b)
Figure 4 Gel electrophoretic profile of the (a) (lane 1) genereplacement cassette consisting of the 51015840 and 31015840pyrG DNA inserts(without the coding region) with the size of sim36 kb and (b) genomicPCR product of the A oryzae S1 wild type strain (lane 1) and itsmutant derivative pyrGΔ0 strain GR6 (lane 2) Lane M 1 kb DNAladder (Fermentas Canada)
replicating plasmids previously reported with other fungalspecies [12 21]
33 Protein Expression Total secreted proteins in culturefiltrates of the untransformed strain GR6 pyrGΔ0 and GR6pyrGΔ0 transformed with ANEp2 were analyzed by SDS-PAGE (Figure 5(a)) and Western blotting (Figure 5(b))SDS-PAGE showed a very prominent 110 kDa proteinwhich is about the size expected of the A niger 120573-galactosidase (Swiss-Prot accession number P29853) (Unit-ProtKB httpwwwuniprotorguniprotP29853) [22] in theculture filtrate of ANEp2 GR6 pyrGΔ0 which was absentin the untransformed strain GR6 pyrGΔ0 Western blotanalyses confirmed that the major band was the A niger 120573-galactosidase As expected from the above culture filtratesof the ANEp2 GR6 pyrGΔ0 transformants possessed highlevels of 120573-galactosidase activity (135UmL) as compared tothe untransformed strain GR6 pyrGΔ0 which expressed 120573-galactosidase at 080UmL
4 Discussion
Replacement of a gene requires homologous recombinationof the introduced DNA with the genome gene replacementis achieved by a precise double crossover event [23] We firstsought these events by selecting for FOA-resistant clonesFOA inhibits the growth of the fungus but permits multipli-cation of cells deficient in OMP decarboxylase and becauseof this are auxotrophic for uridine Initially we selected forpyrGΔ0 gene replacements immediately after transfectionusing a similar plating procedure as has been used toisolate pyrG mutants of T reesei A aculeatus and Penicil-lium nalgiovense from irradiated spores [5 7 21] In ourhands this technique failed to yield FOA resistant colonies
6 The Scientific World Journal
M 1 2+ve
175kDa
80kDa
58kDa
46kDa
30kDa(a)
1 2+ve
110kDa
(b)
Figure 5 Analysis of GR6 culture filtrates Panel (a) lane M SDS-PAGE profile of prestained protein marker (New England Biolabs USA)lane +ve purified A niger 120573-galactosidase isolated from a culture filtrate of A niger transformed with the GlaPr-lacA-GlaTt expressioncassette lane 1 culture filtrate prepared using A oryzae strain GR6 pyrGΔ0 and lane 2 culture filtrate prepared using GR6 pyrGΔ0transformed with ANEp2 Panel (b) Western blot performed by probing the gel shown in Panel (a) with polyclonal rabbit antibodies directedagainst A niger 120573-galactosidase
This was probably because multiple nuclei were present ineach spheroplast and therefore a single pyrGΔ0 locus couldnot prevent production of the suicide inhibitor FdUMPTo test this the transformation protocol was modified toallow cells containing putative pyrG deletions to segregatefrom wild-type ones by plating the transformed spheroplastsonto nonselective regeneration plates containing uracil anduridine Then the conidia from the nonselective plates wereplated-out on CD selectivemedium containing 5-FOA uraciland uridine This led to isolation of the mutant strain GR6pyrGΔ0 PCR based tests confirmed that the pyrG region ofstrain GR6 was deleted
It was possible to complement the uridine requirementof GR6 pyrGΔ0 by introducing the plasmids pUC pyrGAoand ANEp2 carrying A oryzae or A nidulans pyrG respec-tively This suggests that the AMA1 region of ANEp2 whichsupports autonomous plasmid replication in other fungi alsosupports autonomous replication in A oryzae Furthermorethe transformation frequency of ANEp2 was about 10-foldgreater than that obtained with the integrative plasmidpUC pyrGAo Transformation frequency ratios obtainedwith pUC pyrGAo as compared to plasmids harbouringAMA1 were in accordance with those obtained with inte-grating versus autonomously replicating plasmids [12] AlsoFierro et al [21] transformed P nalgiovense with integrativeversus autoreplicative plasmids and found 60-fold higherfrequencies for AMA1-based autonomously replicating plas-mids compared with integrative plasmids Autonomouslyreplicating plasmids carrying the AMA1 region from Anidulans transformed P nalgiovense very efficiently and theseplasmids also showed mitotic stability with low degrees ofreorganization120573-Galactosidase expression showed that the A oryzae
strain GR6 pyrGΔ0 was able to efficiently produce heterolo-gous secreted proteins using the A niger transcriptional and
translational controls introduced via ANEp2The strain GR6pyrGΔ0 transformed with ANEp2 containing lacA encodingA niger 120573-galactosidase was more than 150 times higher thanthat obtained with the untransformed strain The findingof this study is promising and this paves the way for theapplication of A oryzae strain GR6 pyrGΔ0 in heterologousprotein production given that this strain is amenable togenetic manipulation and that A oryzae is widely used inbiotechnology and the food industry
5 Conclusion
Mutant strain GR6 a pyrGΔ0 derivative of the A oryzaestrain S1 was constructed using targeted gene replacementThe pyrG auxotrophs were verified by assessing their abilityto grow on selective and nonselective media and PCRanalysis Plasmids pUC pyrGAo and ANEp2 harbouring thepyrG genes from A oryzae and A nidulans respectivelycomplemented the uracil and uridine requirement of strainGR6 Transformants of the A oryzae GR6 mutant carryingthe heterologous expression vector ANEp2 were able tofunctionally express a secreted 120573-galactosidase at high levels
Acknowledgments
This work was supported by the Ministry of Science Tech-nology and Innovation (MOSTI) Malaysia under the GrantUKM-MGI-NBD0014-2007 Natural Sciences and Engineer-ing Research Council of Canada Grant STPGP 336896 anda scholarship from the Canadian Commonwealth Exchangeprogram (Asia-Pacific) The authors would also like to thankProfessorWilliam J Broughton for the helpful discussion andsuggestions for improving the paper
The Scientific World Journal 7
References
[1] T Kobayashi K Abe K Asai et al ldquoGenomics of Aspergillusoryzaerdquo Bioscience Biotechnology and Biochemistry vol 71 no3 pp 646ndash670 2007
[2] O P Ward W M Qin J Dhanjoon J Ye and A Singh ldquoPhys-iology and biotechnology of Aspergillusrdquo Advances in AppliedMicrobiology vol 58 pp 1ndash75 2005
[3] M Machida K Asai M Sano et al ldquoGenome sequencing andanalysis of Aspergillus oryzaerdquo Nature vol 438 no 7071 pp1157ndash1161 2005
[4] J D Boeke F LaCroute andG R Fink ldquoA positive selection formutants lacking orotidine-51015840-phosphate decarboxylase activityin yeast 5-fluoro-orotic acid resistancerdquoMolecular and GeneralGenetics vol 197 no 2 pp 345ndash346 1984
[5] H Long T-H Wang and Y-K Zhang ldquoIsolation of Tricho-derma reesei pyrG negative Mutant by UV Mutagenesis and ItsApplication in Transformationrdquo Chemical Research in ChineseUniversities vol 24 no 5 pp 565ndash569 2008
[6] MHeerikhuisen CVan denHondel andP J Punt ldquoAspergillussojaerdquo in Production of Recombinant ProteinsmdashNovel Microbialand Eukaryotic Expression Systems pp 191ndash214 Wiley-VCHWeinheim Germany 2005
[7] S Kanamasa K Yamaoka T Kawaguchi J-I Sumitani and MAra ldquoTransformation of Aspergillus aculeatus using the drugresistance gene of Aspergillus oryzae and the pyrG gene ofAspergillus nidulansrdquo Bioscience Biotechnology and Biochem-istry vol 67 no 12 pp 2661ndash2663 2003
[8] R V Miller E J Ford N J Zidack and D C Sands ldquoApyrimidine auxotroph of Sclerotinia sclerotiorum for use inbiological weed controlrdquo Journal of General Microbiology vol135 pp 2085ndash2091 1989
[9] W Van Hartingsveldt I E Mattern and C M J Van ZeijlldquoDevelopment of a homologous transformation system forAspergillus niger based on the pyrG generdquoMolecular andGeneralGenetics vol 206 no 1 pp 71ndash75 1987
[10] D Mueller and U Stahl ldquoRNA a powerful tool for recombina-tion and regulated expression of genesrdquo in Progress in Botanypp 31ndash45 Springer Berlin Germany 2005
[11] J Aranda M Poza B G Pardo et al ldquoA rapid and simplemethod for constructing stable mutants of Acinetobacter bau-manniirdquo BMCMicrobiology vol 10 article 279 2010
[12] R Storms Y Zheng H Li S Sillaots AMartinez-Perez and ATsang ldquoPlasmid vectors for protein production gene expressionandmolecular manipulations inAspergillus nigerrdquo Plasmid vol53 no 3 pp 191ndash204 2005
[13] S S L Oh J M Leong A M A Murad N M Mahadi andF D Abu Bakar ldquoCloning and analysis of pyrG gene encod-ing orotidine 5-monophosphate decarboxylase of Aspergillusoryzae strain Slrdquo Sains Malaysiana vol 40 no 4 pp 331ndash3372011
[14] S Ushijima T Nakadai and K Uchida ldquoInterspecific electrofu-sion of protoplasts between Aspergillus oryzae and Aspergillussojaerdquo Agricultural and Biological Chemistry vol 55 pp 129ndash136 1991
[15] Y Iimura K Gomi H Uzu and S Hara ldquoTransformationof Aspergillus oryzae through plasmid-mediated complementa-tion of themethionine-auxotrophicmutationrdquoAgricultural andBiological Chemistry vol 51 pp 323ndash328 1987
[16] J J Feng T Takahashi M Machida and Y Koyama ldquoIdenti-fication of a basic helix-loop-helix-type transcription regulator
gene in Aspergillus oryzae by systematically deleting large chro-mosomal segmentsrdquo Applied and Environmental Microbiologyvol 75 no 18 pp 5943ndash5951 2009
[17] Y R Liew Protoplasting and cell wall regeneration of the filamen-tous fungi Aspergillus oryzae strain S1 [BSc thesis] UniversitiKebangsaan Malaysia Malaysia 2008
[18] K Wernars T Goosen B M Wennekes K Swart CA Vanden Hondel and HW Van den Broek ldquoCotransformationof Aspergillus nidulans a tool for replacing fungal genesrdquoMolecular and General Genetics vol 209 no 1 pp 71ndash77 1987
[19] E Kafer ldquoMeiotic andmitotic recombination in Aspergillus andits chromosomal aberrationsrdquo Advances in Genetics vol 19 pp33ndash131 1977
[20] ldquoLiquid media plates and overlayers for A nigerrdquo 2012 httpcloneconcordiacaaspergillusprotocolsnigmediahtml
[21] F Fierro F Laich R O Garcıa-Rico and J F Martın ldquoHighefficiency transformation of Penicillium nalgiovense with inte-grative and autonomously replicating plasmidsrdquo InternationalJournal of Food Microbiology vol 90 no 2 pp 237ndash248 2004
[22] ldquoUnitProtKBrdquo 2012 httpwwwuniprotorguniprotP29853[23] V L Prabha and N S Punekar ldquoGenetic transformation in
Aspergilli tools of the traderdquo Indian Journal of Biochemistry andBiophysics vol 41 no 5 pp 205ndash215 2004

The Scientific World Journal 5
(a) Wild type (b) GR6 mutant
(c) CD plates (minimal medium)
(d) CD (uracil uridineand 5-FOA) plates
Figure 3 Media plates showing growth or no growth of the Aoryzae S1 wild type (column (a)) and GR6 pyrG mutant (column(b)) Column (a) ability of the wild type strain S1 (row (c)) andColumn (b) the inability of the GR6 pyrGmutant strain (row (c)) togrow on minimal medium CD plates Column (a) inability of thewild type strain S1 (row (d)) and Column (b) ability of the GR6pyrGmutant strain (row (d)) to grow onminimalmediumCDplatessupplemented with uracil uridine and 5-FOA
not prevent production of the suicide inhibitor FdUMPTo address this possibility the transformation protocol wasmodified by allowing nuclei in which the pyrG gene wasdeleted to segregate from wild-type nuclei by plating thetransformed spheroplasts (about 8 times 105 per plate) ontononselective regeneration plates containing uraciluridineOnce a lawn of conidia was produced the conidia werecollected diluted and plated at 8 times 106 spores per plateon CD selective medium with 5-FOA uracil and uridineTen randomly selected colonies were chosen from over 100colonies that formed and purified to homogeneity by tworounds of plating on CD uracil and uridine plates Conidiaprepared using the purified colonies were harvested andplated onto CD plates without uracil and uridine (106 conidiaper plate) Only one of the putative mutants designated GR6(Gene Replacement mutant number 6) did not grow and wastherefore assumed to be completely stable (Figure 3)
32 Verifying that GR6 Harbours a pyrGΔ0 Locus PCRamplifications with primers Pyr51015840-out and Pyr31015840-out usingthe genomic DNA of strain S1 and strain GR6 as the tem-plates showed that a 09 kb deletion was introduced into thepyrG region of strain GR6 (Figure 4(b)) which is consistentwith generation of the desired pyrGΔ0 locus by homologousrecombination
Then the pyrG auxotroph GR6 was transformed withplasmids pUC pyrGAo and ANEp2 These plasmids containthe wild-type pyrG from A oryzae (in pUC19) and Anidulans respectively Both plasmids were introduced intoand restored uracil and uridine prototrophy of GR6 withfrequencies of three transformants per 120583g of plasmid DNAfor pUC pyrGAo and 50 transformants per 120583g of plasmidDNA for ANEp2 Furthermore the frequency ratio of pyrGprototrophs obtained when transformation was performedwith the pUC pyrGAo versus the AMA plasmid was about01 This is in accordance with the range of prototrophy fre-quency ratios obtained with integrating versus autonomously
1kb 1
35kb
30kb sim36kb
(a)
50kb40kb35kb
sim45kb
sim36kb
1kb 1 2
(b)
Figure 4 Gel electrophoretic profile of the (a) (lane 1) genereplacement cassette consisting of the 51015840 and 31015840pyrG DNA inserts(without the coding region) with the size of sim36 kb and (b) genomicPCR product of the A oryzae S1 wild type strain (lane 1) and itsmutant derivative pyrGΔ0 strain GR6 (lane 2) Lane M 1 kb DNAladder (Fermentas Canada)
replicating plasmids previously reported with other fungalspecies [12 21]
33 Protein Expression Total secreted proteins in culturefiltrates of the untransformed strain GR6 pyrGΔ0 and GR6pyrGΔ0 transformed with ANEp2 were analyzed by SDS-PAGE (Figure 5(a)) and Western blotting (Figure 5(b))SDS-PAGE showed a very prominent 110 kDa proteinwhich is about the size expected of the A niger 120573-galactosidase (Swiss-Prot accession number P29853) (Unit-ProtKB httpwwwuniprotorguniprotP29853) [22] in theculture filtrate of ANEp2 GR6 pyrGΔ0 which was absentin the untransformed strain GR6 pyrGΔ0 Western blotanalyses confirmed that the major band was the A niger 120573-galactosidase As expected from the above culture filtratesof the ANEp2 GR6 pyrGΔ0 transformants possessed highlevels of 120573-galactosidase activity (135UmL) as compared tothe untransformed strain GR6 pyrGΔ0 which expressed 120573-galactosidase at 080UmL
4 Discussion
Replacement of a gene requires homologous recombinationof the introduced DNA with the genome gene replacementis achieved by a precise double crossover event [23] We firstsought these events by selecting for FOA-resistant clonesFOA inhibits the growth of the fungus but permits multipli-cation of cells deficient in OMP decarboxylase and becauseof this are auxotrophic for uridine Initially we selected forpyrGΔ0 gene replacements immediately after transfectionusing a similar plating procedure as has been used toisolate pyrG mutants of T reesei A aculeatus and Penicil-lium nalgiovense from irradiated spores [5 7 21] In ourhands this technique failed to yield FOA resistant colonies
6 The Scientific World Journal
M 1 2+ve
175kDa
80kDa
58kDa
46kDa
30kDa(a)
1 2+ve
110kDa
(b)
Figure 5 Analysis of GR6 culture filtrates Panel (a) lane M SDS-PAGE profile of prestained protein marker (New England Biolabs USA)lane +ve purified A niger 120573-galactosidase isolated from a culture filtrate of A niger transformed with the GlaPr-lacA-GlaTt expressioncassette lane 1 culture filtrate prepared using A oryzae strain GR6 pyrGΔ0 and lane 2 culture filtrate prepared using GR6 pyrGΔ0transformed with ANEp2 Panel (b) Western blot performed by probing the gel shown in Panel (a) with polyclonal rabbit antibodies directedagainst A niger 120573-galactosidase
This was probably because multiple nuclei were present ineach spheroplast and therefore a single pyrGΔ0 locus couldnot prevent production of the suicide inhibitor FdUMPTo test this the transformation protocol was modified toallow cells containing putative pyrG deletions to segregatefrom wild-type ones by plating the transformed spheroplastsonto nonselective regeneration plates containing uracil anduridine Then the conidia from the nonselective plates wereplated-out on CD selectivemedium containing 5-FOA uraciland uridine This led to isolation of the mutant strain GR6pyrGΔ0 PCR based tests confirmed that the pyrG region ofstrain GR6 was deleted
It was possible to complement the uridine requirementof GR6 pyrGΔ0 by introducing the plasmids pUC pyrGAoand ANEp2 carrying A oryzae or A nidulans pyrG respec-tively This suggests that the AMA1 region of ANEp2 whichsupports autonomous plasmid replication in other fungi alsosupports autonomous replication in A oryzae Furthermorethe transformation frequency of ANEp2 was about 10-foldgreater than that obtained with the integrative plasmidpUC pyrGAo Transformation frequency ratios obtainedwith pUC pyrGAo as compared to plasmids harbouringAMA1 were in accordance with those obtained with inte-grating versus autonomously replicating plasmids [12] AlsoFierro et al [21] transformed P nalgiovense with integrativeversus autoreplicative plasmids and found 60-fold higherfrequencies for AMA1-based autonomously replicating plas-mids compared with integrative plasmids Autonomouslyreplicating plasmids carrying the AMA1 region from Anidulans transformed P nalgiovense very efficiently and theseplasmids also showed mitotic stability with low degrees ofreorganization120573-Galactosidase expression showed that the A oryzae
strain GR6 pyrGΔ0 was able to efficiently produce heterolo-gous secreted proteins using the A niger transcriptional and
translational controls introduced via ANEp2The strain GR6pyrGΔ0 transformed with ANEp2 containing lacA encodingA niger 120573-galactosidase was more than 150 times higher thanthat obtained with the untransformed strain The findingof this study is promising and this paves the way for theapplication of A oryzae strain GR6 pyrGΔ0 in heterologousprotein production given that this strain is amenable togenetic manipulation and that A oryzae is widely used inbiotechnology and the food industry
5 Conclusion
Mutant strain GR6 a pyrGΔ0 derivative of the A oryzaestrain S1 was constructed using targeted gene replacementThe pyrG auxotrophs were verified by assessing their abilityto grow on selective and nonselective media and PCRanalysis Plasmids pUC pyrGAo and ANEp2 harbouring thepyrG genes from A oryzae and A nidulans respectivelycomplemented the uracil and uridine requirement of strainGR6 Transformants of the A oryzae GR6 mutant carryingthe heterologous expression vector ANEp2 were able tofunctionally express a secreted 120573-galactosidase at high levels
Acknowledgments
This work was supported by the Ministry of Science Tech-nology and Innovation (MOSTI) Malaysia under the GrantUKM-MGI-NBD0014-2007 Natural Sciences and Engineer-ing Research Council of Canada Grant STPGP 336896 anda scholarship from the Canadian Commonwealth Exchangeprogram (Asia-Pacific) The authors would also like to thankProfessorWilliam J Broughton for the helpful discussion andsuggestions for improving the paper
The Scientific World Journal 7
References
[1] T Kobayashi K Abe K Asai et al ldquoGenomics of Aspergillusoryzaerdquo Bioscience Biotechnology and Biochemistry vol 71 no3 pp 646ndash670 2007
[2] O P Ward W M Qin J Dhanjoon J Ye and A Singh ldquoPhys-iology and biotechnology of Aspergillusrdquo Advances in AppliedMicrobiology vol 58 pp 1ndash75 2005
[3] M Machida K Asai M Sano et al ldquoGenome sequencing andanalysis of Aspergillus oryzaerdquo Nature vol 438 no 7071 pp1157ndash1161 2005
[4] J D Boeke F LaCroute andG R Fink ldquoA positive selection formutants lacking orotidine-51015840-phosphate decarboxylase activityin yeast 5-fluoro-orotic acid resistancerdquoMolecular and GeneralGenetics vol 197 no 2 pp 345ndash346 1984
[5] H Long T-H Wang and Y-K Zhang ldquoIsolation of Tricho-derma reesei pyrG negative Mutant by UV Mutagenesis and ItsApplication in Transformationrdquo Chemical Research in ChineseUniversities vol 24 no 5 pp 565ndash569 2008
[6] MHeerikhuisen CVan denHondel andP J Punt ldquoAspergillussojaerdquo in Production of Recombinant ProteinsmdashNovel Microbialand Eukaryotic Expression Systems pp 191ndash214 Wiley-VCHWeinheim Germany 2005
[7] S Kanamasa K Yamaoka T Kawaguchi J-I Sumitani and MAra ldquoTransformation of Aspergillus aculeatus using the drugresistance gene of Aspergillus oryzae and the pyrG gene ofAspergillus nidulansrdquo Bioscience Biotechnology and Biochem-istry vol 67 no 12 pp 2661ndash2663 2003
[8] R V Miller E J Ford N J Zidack and D C Sands ldquoApyrimidine auxotroph of Sclerotinia sclerotiorum for use inbiological weed controlrdquo Journal of General Microbiology vol135 pp 2085ndash2091 1989
[9] W Van Hartingsveldt I E Mattern and C M J Van ZeijlldquoDevelopment of a homologous transformation system forAspergillus niger based on the pyrG generdquoMolecular andGeneralGenetics vol 206 no 1 pp 71ndash75 1987
[10] D Mueller and U Stahl ldquoRNA a powerful tool for recombina-tion and regulated expression of genesrdquo in Progress in Botanypp 31ndash45 Springer Berlin Germany 2005
[11] J Aranda M Poza B G Pardo et al ldquoA rapid and simplemethod for constructing stable mutants of Acinetobacter bau-manniirdquo BMCMicrobiology vol 10 article 279 2010
[12] R Storms Y Zheng H Li S Sillaots AMartinez-Perez and ATsang ldquoPlasmid vectors for protein production gene expressionandmolecular manipulations inAspergillus nigerrdquo Plasmid vol53 no 3 pp 191ndash204 2005
[13] S S L Oh J M Leong A M A Murad N M Mahadi andF D Abu Bakar ldquoCloning and analysis of pyrG gene encod-ing orotidine 5-monophosphate decarboxylase of Aspergillusoryzae strain Slrdquo Sains Malaysiana vol 40 no 4 pp 331ndash3372011
[14] S Ushijima T Nakadai and K Uchida ldquoInterspecific electrofu-sion of protoplasts between Aspergillus oryzae and Aspergillussojaerdquo Agricultural and Biological Chemistry vol 55 pp 129ndash136 1991
[15] Y Iimura K Gomi H Uzu and S Hara ldquoTransformationof Aspergillus oryzae through plasmid-mediated complementa-tion of themethionine-auxotrophicmutationrdquoAgricultural andBiological Chemistry vol 51 pp 323ndash328 1987
[16] J J Feng T Takahashi M Machida and Y Koyama ldquoIdenti-fication of a basic helix-loop-helix-type transcription regulator
gene in Aspergillus oryzae by systematically deleting large chro-mosomal segmentsrdquo Applied and Environmental Microbiologyvol 75 no 18 pp 5943ndash5951 2009
[17] Y R Liew Protoplasting and cell wall regeneration of the filamen-tous fungi Aspergillus oryzae strain S1 [BSc thesis] UniversitiKebangsaan Malaysia Malaysia 2008
[18] K Wernars T Goosen B M Wennekes K Swart CA Vanden Hondel and HW Van den Broek ldquoCotransformationof Aspergillus nidulans a tool for replacing fungal genesrdquoMolecular and General Genetics vol 209 no 1 pp 71ndash77 1987
[19] E Kafer ldquoMeiotic andmitotic recombination in Aspergillus andits chromosomal aberrationsrdquo Advances in Genetics vol 19 pp33ndash131 1977
[20] ldquoLiquid media plates and overlayers for A nigerrdquo 2012 httpcloneconcordiacaaspergillusprotocolsnigmediahtml
[21] F Fierro F Laich R O Garcıa-Rico and J F Martın ldquoHighefficiency transformation of Penicillium nalgiovense with inte-grative and autonomously replicating plasmidsrdquo InternationalJournal of Food Microbiology vol 90 no 2 pp 237ndash248 2004
[22] ldquoUnitProtKBrdquo 2012 httpwwwuniprotorguniprotP29853[23] V L Prabha and N S Punekar ldquoGenetic transformation in
Aspergilli tools of the traderdquo Indian Journal of Biochemistry andBiophysics vol 41 no 5 pp 205ndash215 2004
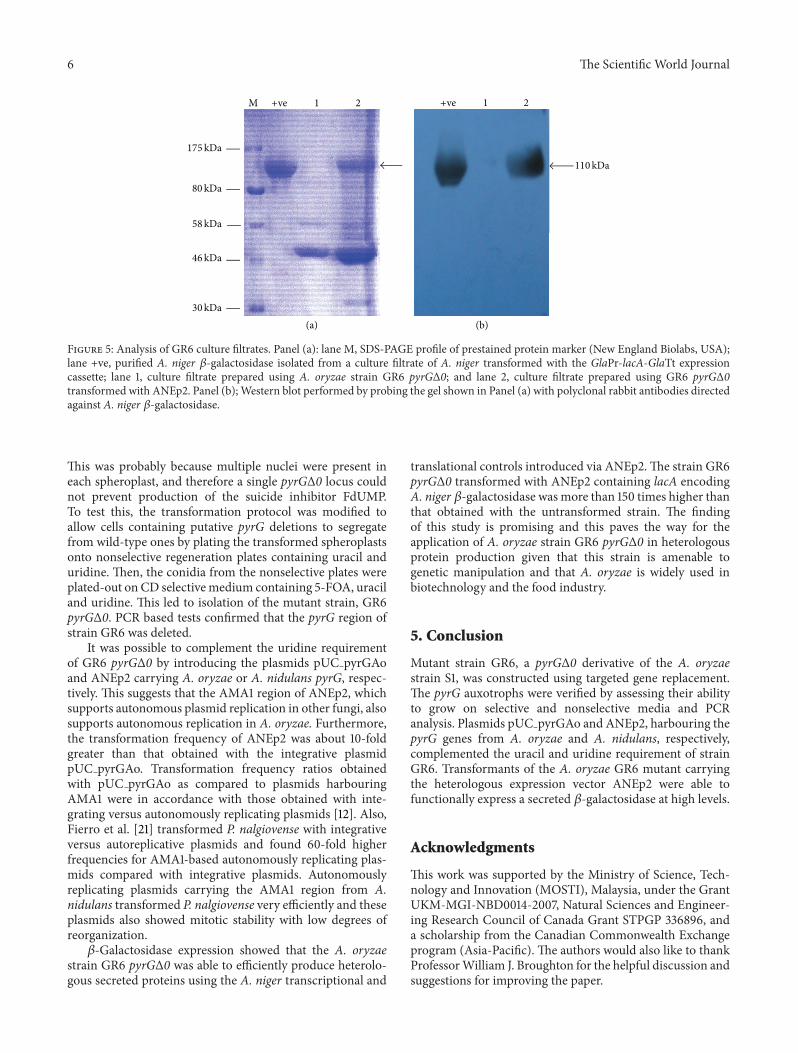
6 The Scientific World Journal
M 1 2+ve
175kDa
80kDa
58kDa
46kDa
30kDa(a)
1 2+ve
110kDa
(b)
Figure 5 Analysis of GR6 culture filtrates Panel (a) lane M SDS-PAGE profile of prestained protein marker (New England Biolabs USA)lane +ve purified A niger 120573-galactosidase isolated from a culture filtrate of A niger transformed with the GlaPr-lacA-GlaTt expressioncassette lane 1 culture filtrate prepared using A oryzae strain GR6 pyrGΔ0 and lane 2 culture filtrate prepared using GR6 pyrGΔ0transformed with ANEp2 Panel (b) Western blot performed by probing the gel shown in Panel (a) with polyclonal rabbit antibodies directedagainst A niger 120573-galactosidase
This was probably because multiple nuclei were present ineach spheroplast and therefore a single pyrGΔ0 locus couldnot prevent production of the suicide inhibitor FdUMPTo test this the transformation protocol was modified toallow cells containing putative pyrG deletions to segregatefrom wild-type ones by plating the transformed spheroplastsonto nonselective regeneration plates containing uracil anduridine Then the conidia from the nonselective plates wereplated-out on CD selectivemedium containing 5-FOA uraciland uridine This led to isolation of the mutant strain GR6pyrGΔ0 PCR based tests confirmed that the pyrG region ofstrain GR6 was deleted
It was possible to complement the uridine requirementof GR6 pyrGΔ0 by introducing the plasmids pUC pyrGAoand ANEp2 carrying A oryzae or A nidulans pyrG respec-tively This suggests that the AMA1 region of ANEp2 whichsupports autonomous plasmid replication in other fungi alsosupports autonomous replication in A oryzae Furthermorethe transformation frequency of ANEp2 was about 10-foldgreater than that obtained with the integrative plasmidpUC pyrGAo Transformation frequency ratios obtainedwith pUC pyrGAo as compared to plasmids harbouringAMA1 were in accordance with those obtained with inte-grating versus autonomously replicating plasmids [12] AlsoFierro et al [21] transformed P nalgiovense with integrativeversus autoreplicative plasmids and found 60-fold higherfrequencies for AMA1-based autonomously replicating plas-mids compared with integrative plasmids Autonomouslyreplicating plasmids carrying the AMA1 region from Anidulans transformed P nalgiovense very efficiently and theseplasmids also showed mitotic stability with low degrees ofreorganization120573-Galactosidase expression showed that the A oryzae
strain GR6 pyrGΔ0 was able to efficiently produce heterolo-gous secreted proteins using the A niger transcriptional and
translational controls introduced via ANEp2The strain GR6pyrGΔ0 transformed with ANEp2 containing lacA encodingA niger 120573-galactosidase was more than 150 times higher thanthat obtained with the untransformed strain The findingof this study is promising and this paves the way for theapplication of A oryzae strain GR6 pyrGΔ0 in heterologousprotein production given that this strain is amenable togenetic manipulation and that A oryzae is widely used inbiotechnology and the food industry
5 Conclusion
Mutant strain GR6 a pyrGΔ0 derivative of the A oryzaestrain S1 was constructed using targeted gene replacementThe pyrG auxotrophs were verified by assessing their abilityto grow on selective and nonselective media and PCRanalysis Plasmids pUC pyrGAo and ANEp2 harbouring thepyrG genes from A oryzae and A nidulans respectivelycomplemented the uracil and uridine requirement of strainGR6 Transformants of the A oryzae GR6 mutant carryingthe heterologous expression vector ANEp2 were able tofunctionally express a secreted 120573-galactosidase at high levels
Acknowledgments
This work was supported by the Ministry of Science Tech-nology and Innovation (MOSTI) Malaysia under the GrantUKM-MGI-NBD0014-2007 Natural Sciences and Engineer-ing Research Council of Canada Grant STPGP 336896 anda scholarship from the Canadian Commonwealth Exchangeprogram (Asia-Pacific) The authors would also like to thankProfessorWilliam J Broughton for the helpful discussion andsuggestions for improving the paper
The Scientific World Journal 7
References
[1] T Kobayashi K Abe K Asai et al ldquoGenomics of Aspergillusoryzaerdquo Bioscience Biotechnology and Biochemistry vol 71 no3 pp 646ndash670 2007
[2] O P Ward W M Qin J Dhanjoon J Ye and A Singh ldquoPhys-iology and biotechnology of Aspergillusrdquo Advances in AppliedMicrobiology vol 58 pp 1ndash75 2005
[3] M Machida K Asai M Sano et al ldquoGenome sequencing andanalysis of Aspergillus oryzaerdquo Nature vol 438 no 7071 pp1157ndash1161 2005
[4] J D Boeke F LaCroute andG R Fink ldquoA positive selection formutants lacking orotidine-51015840-phosphate decarboxylase activityin yeast 5-fluoro-orotic acid resistancerdquoMolecular and GeneralGenetics vol 197 no 2 pp 345ndash346 1984
[5] H Long T-H Wang and Y-K Zhang ldquoIsolation of Tricho-derma reesei pyrG negative Mutant by UV Mutagenesis and ItsApplication in Transformationrdquo Chemical Research in ChineseUniversities vol 24 no 5 pp 565ndash569 2008
[6] MHeerikhuisen CVan denHondel andP J Punt ldquoAspergillussojaerdquo in Production of Recombinant ProteinsmdashNovel Microbialand Eukaryotic Expression Systems pp 191ndash214 Wiley-VCHWeinheim Germany 2005
[7] S Kanamasa K Yamaoka T Kawaguchi J-I Sumitani and MAra ldquoTransformation of Aspergillus aculeatus using the drugresistance gene of Aspergillus oryzae and the pyrG gene ofAspergillus nidulansrdquo Bioscience Biotechnology and Biochem-istry vol 67 no 12 pp 2661ndash2663 2003
[8] R V Miller E J Ford N J Zidack and D C Sands ldquoApyrimidine auxotroph of Sclerotinia sclerotiorum for use inbiological weed controlrdquo Journal of General Microbiology vol135 pp 2085ndash2091 1989
[9] W Van Hartingsveldt I E Mattern and C M J Van ZeijlldquoDevelopment of a homologous transformation system forAspergillus niger based on the pyrG generdquoMolecular andGeneralGenetics vol 206 no 1 pp 71ndash75 1987
[10] D Mueller and U Stahl ldquoRNA a powerful tool for recombina-tion and regulated expression of genesrdquo in Progress in Botanypp 31ndash45 Springer Berlin Germany 2005
[11] J Aranda M Poza B G Pardo et al ldquoA rapid and simplemethod for constructing stable mutants of Acinetobacter bau-manniirdquo BMCMicrobiology vol 10 article 279 2010
[12] R Storms Y Zheng H Li S Sillaots AMartinez-Perez and ATsang ldquoPlasmid vectors for protein production gene expressionandmolecular manipulations inAspergillus nigerrdquo Plasmid vol53 no 3 pp 191ndash204 2005
[13] S S L Oh J M Leong A M A Murad N M Mahadi andF D Abu Bakar ldquoCloning and analysis of pyrG gene encod-ing orotidine 5-monophosphate decarboxylase of Aspergillusoryzae strain Slrdquo Sains Malaysiana vol 40 no 4 pp 331ndash3372011
[14] S Ushijima T Nakadai and K Uchida ldquoInterspecific electrofu-sion of protoplasts between Aspergillus oryzae and Aspergillussojaerdquo Agricultural and Biological Chemistry vol 55 pp 129ndash136 1991
[15] Y Iimura K Gomi H Uzu and S Hara ldquoTransformationof Aspergillus oryzae through plasmid-mediated complementa-tion of themethionine-auxotrophicmutationrdquoAgricultural andBiological Chemistry vol 51 pp 323ndash328 1987
[16] J J Feng T Takahashi M Machida and Y Koyama ldquoIdenti-fication of a basic helix-loop-helix-type transcription regulator
gene in Aspergillus oryzae by systematically deleting large chro-mosomal segmentsrdquo Applied and Environmental Microbiologyvol 75 no 18 pp 5943ndash5951 2009
[17] Y R Liew Protoplasting and cell wall regeneration of the filamen-tous fungi Aspergillus oryzae strain S1 [BSc thesis] UniversitiKebangsaan Malaysia Malaysia 2008
[18] K Wernars T Goosen B M Wennekes K Swart CA Vanden Hondel and HW Van den Broek ldquoCotransformationof Aspergillus nidulans a tool for replacing fungal genesrdquoMolecular and General Genetics vol 209 no 1 pp 71ndash77 1987
[19] E Kafer ldquoMeiotic andmitotic recombination in Aspergillus andits chromosomal aberrationsrdquo Advances in Genetics vol 19 pp33ndash131 1977
[20] ldquoLiquid media plates and overlayers for A nigerrdquo 2012 httpcloneconcordiacaaspergillusprotocolsnigmediahtml
[21] F Fierro F Laich R O Garcıa-Rico and J F Martın ldquoHighefficiency transformation of Penicillium nalgiovense with inte-grative and autonomously replicating plasmidsrdquo InternationalJournal of Food Microbiology vol 90 no 2 pp 237ndash248 2004
[22] ldquoUnitProtKBrdquo 2012 httpwwwuniprotorguniprotP29853[23] V L Prabha and N S Punekar ldquoGenetic transformation in
Aspergilli tools of the traderdquo Indian Journal of Biochemistry andBiophysics vol 41 no 5 pp 205ndash215 2004

The Scientific World Journal 7
References
[1] T Kobayashi K Abe K Asai et al ldquoGenomics of Aspergillusoryzaerdquo Bioscience Biotechnology and Biochemistry vol 71 no3 pp 646ndash670 2007
[2] O P Ward W M Qin J Dhanjoon J Ye and A Singh ldquoPhys-iology and biotechnology of Aspergillusrdquo Advances in AppliedMicrobiology vol 58 pp 1ndash75 2005
[3] M Machida K Asai M Sano et al ldquoGenome sequencing andanalysis of Aspergillus oryzaerdquo Nature vol 438 no 7071 pp1157ndash1161 2005
[4] J D Boeke F LaCroute andG R Fink ldquoA positive selection formutants lacking orotidine-51015840-phosphate decarboxylase activityin yeast 5-fluoro-orotic acid resistancerdquoMolecular and GeneralGenetics vol 197 no 2 pp 345ndash346 1984
[5] H Long T-H Wang and Y-K Zhang ldquoIsolation of Tricho-derma reesei pyrG negative Mutant by UV Mutagenesis and ItsApplication in Transformationrdquo Chemical Research in ChineseUniversities vol 24 no 5 pp 565ndash569 2008
[6] MHeerikhuisen CVan denHondel andP J Punt ldquoAspergillussojaerdquo in Production of Recombinant ProteinsmdashNovel Microbialand Eukaryotic Expression Systems pp 191ndash214 Wiley-VCHWeinheim Germany 2005
[7] S Kanamasa K Yamaoka T Kawaguchi J-I Sumitani and MAra ldquoTransformation of Aspergillus aculeatus using the drugresistance gene of Aspergillus oryzae and the pyrG gene ofAspergillus nidulansrdquo Bioscience Biotechnology and Biochem-istry vol 67 no 12 pp 2661ndash2663 2003
[8] R V Miller E J Ford N J Zidack and D C Sands ldquoApyrimidine auxotroph of Sclerotinia sclerotiorum for use inbiological weed controlrdquo Journal of General Microbiology vol135 pp 2085ndash2091 1989
[9] W Van Hartingsveldt I E Mattern and C M J Van ZeijlldquoDevelopment of a homologous transformation system forAspergillus niger based on the pyrG generdquoMolecular andGeneralGenetics vol 206 no 1 pp 71ndash75 1987
[10] D Mueller and U Stahl ldquoRNA a powerful tool for recombina-tion and regulated expression of genesrdquo in Progress in Botanypp 31ndash45 Springer Berlin Germany 2005
[11] J Aranda M Poza B G Pardo et al ldquoA rapid and simplemethod for constructing stable mutants of Acinetobacter bau-manniirdquo BMCMicrobiology vol 10 article 279 2010
[12] R Storms Y Zheng H Li S Sillaots AMartinez-Perez and ATsang ldquoPlasmid vectors for protein production gene expressionandmolecular manipulations inAspergillus nigerrdquo Plasmid vol53 no 3 pp 191ndash204 2005
[13] S S L Oh J M Leong A M A Murad N M Mahadi andF D Abu Bakar ldquoCloning and analysis of pyrG gene encod-ing orotidine 5-monophosphate decarboxylase of Aspergillusoryzae strain Slrdquo Sains Malaysiana vol 40 no 4 pp 331ndash3372011
[14] S Ushijima T Nakadai and K Uchida ldquoInterspecific electrofu-sion of protoplasts between Aspergillus oryzae and Aspergillussojaerdquo Agricultural and Biological Chemistry vol 55 pp 129ndash136 1991
[15] Y Iimura K Gomi H Uzu and S Hara ldquoTransformationof Aspergillus oryzae through plasmid-mediated complementa-tion of themethionine-auxotrophicmutationrdquoAgricultural andBiological Chemistry vol 51 pp 323ndash328 1987
[16] J J Feng T Takahashi M Machida and Y Koyama ldquoIdenti-fication of a basic helix-loop-helix-type transcription regulator
gene in Aspergillus oryzae by systematically deleting large chro-mosomal segmentsrdquo Applied and Environmental Microbiologyvol 75 no 18 pp 5943ndash5951 2009
[17] Y R Liew Protoplasting and cell wall regeneration of the filamen-tous fungi Aspergillus oryzae strain S1 [BSc thesis] UniversitiKebangsaan Malaysia Malaysia 2008
[18] K Wernars T Goosen B M Wennekes K Swart CA Vanden Hondel and HW Van den Broek ldquoCotransformationof Aspergillus nidulans a tool for replacing fungal genesrdquoMolecular and General Genetics vol 209 no 1 pp 71ndash77 1987
[19] E Kafer ldquoMeiotic andmitotic recombination in Aspergillus andits chromosomal aberrationsrdquo Advances in Genetics vol 19 pp33ndash131 1977
[20] ldquoLiquid media plates and overlayers for A nigerrdquo 2012 httpcloneconcordiacaaspergillusprotocolsnigmediahtml
[21] F Fierro F Laich R O Garcıa-Rico and J F Martın ldquoHighefficiency transformation of Penicillium nalgiovense with inte-grative and autonomously replicating plasmidsrdquo InternationalJournal of Food Microbiology vol 90 no 2 pp 237ndash248 2004
[22] ldquoUnitProtKBrdquo 2012 httpwwwuniprotorguniprotP29853[23] V L Prabha and N S Punekar ldquoGenetic transformation in
Aspergilli tools of the traderdquo Indian Journal of Biochemistry andBiophysics vol 41 no 5 pp 205ndash215 2004




















