
STUDI KERAGAMAN RAS ISOLAT Pyricularia oryzae Cavara ...
72
STUDI KERAGAMAN RAS ISOLAT Pyricularia oryzae Cavara PENYEBAB PENYAKIT BLAS PADA TANAMAN PADI DARI BEBERAPA KABUPATEN DI SULAWESI SELATAN RIANINGSIH P4100215003 PROGRAM MAGISTER ILMU HAMA DAN PENYAKIT TUMBUHAN FAKULTAS PERTANIAN UNIVERSITAS HASANUDDIN MAKASSAR 2017
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of STUDI KERAGAMAN RAS ISOLAT Pyricularia oryzae Cavara ...

i
STUDI KERAGAMAN RAS ISOLAT Pyricularia oryzae CavaraPENYEBAB PENYAKIT BLAS PADA TANAMAN PADI DARI BEBERAPA
KABUPATEN DI SULAWESI SELATAN
RIANINGSIHP4100215003
PROGRAM MAGISTER ILMU HAMA DAN PENYAKIT TUMBUHANFAKULTAS PERTANIAN
UNIVERSITAS HASANUDDINMAKASSAR
2017

ii
STUDI KERAGAMAN RAS ISOLAT Pyricularia oryzae Cavara PENYEBABPENYAKIT BLAS PADA TANAMAN PADI DARI BEBERAPA KABUPATEN DI
SULAWESI SELATAN
TESISSebagai salah satu syarat untuk mencapai gelar Magister
Program StudiIlmu Hama dan Penyakit Tumbuhan
Disusun dan diajukan oleh :
RIANINGSIHNomor Pokok P4100215003
Kepada
FAKULTAS PERTANIANUNIVERSITAS HASANUDDIN
MAKASSAR2017

iii

iv
PERNYATAAN KEASLIAN TESIS
Yang bertanda tangan di bawah ini:
Nama : Rianingsih
Nomor Pokok : P4100215003
Program Studi : Ilmu Hama dan Penyakit Tumbuhan
Menyatakan dengan sebenarnya bahwa tesis yang saya tulis ini benar-benar
merupakan hasil karya saya sendiri bukan merupakan pengambilalihan tulisan atau
pemikiran orang lain. Apabila dikemudian hari terbukti atau dapat dibuktikan
sebagian atau keseluruhan tesis ini hasil karya orang lain saya bersedia atau
merima sanksi atas perbuatan tersebut
Makassar, 28 Nopember 2017
Yang menyatakan
Rianingsih

v
ABSTRAK
RIANINGSIH. Studi Keragaman Ras Isolat Pyricularia Oryzae Cavara PenyebabPenyakit Blas Pada Tanaman Padi Dari Beberapa Kabupaten Di Sulawesi Selatan.(dibimbing oleh Tutik Kuswinanti dan Andi Nasruddin).
Tujuan penelitian ini adalah untuk memperoleh data tentang penyebaran rasPyricularia oryzae pada beberapa wilayah di Sulawesi Selatan serta analisa genyang terpaut dengan sifat virulensi P. oryzae penyebab penyakit blas.
Penentuan ras penyakit blas menggunakan 1 set varietas diferensial yangterdiri dari 7 varietas yaitu varietas Asahan, Cisokan, IR 64, Krueng Aceh, Cisadane,Cisanggarung dan Kencana Bali. Intensitas serangan blas diamati pada tujuh harisetelah inokulasi menggunakan standar evaluasi IRRI (1996). Tipe bercak skala 5-9digunakan untuk mengidentifikasi tingkat kerentanan inang. Uji genotipe cendawanP.oryzae dilakukan menggunakan marka berbasis gen untuk sifat vilurensi yaitumarkah Erg2 (1.440 bp), Pwl2 (900 bp), dan Cut1 (1.730bp). Pita DNA hasilamplifikasi yang muncul untuk setiap primer dari masing-masing isolat diskordengan nilai 1 (ada) dan 0 (tidak ada).
Hasil isolasi cendawan di temukan total 20 isolat diperoleh dari tigakabupaten yang berbeda di Sulawesi Selatan. Ditemukan 8 isolat dari Kab.Bone, 8isolat di Kab.Maros dan 4 isolat di Kab.Gowa. Berdasarkan morfologi dankenampakkan makroskopis mengacu pada P. oryzae. Inokulasi isolat P. oryzaepada 7 varietas padi diferensial ditemukan 12 ras, Meskipun demikian ras 020 palingdominan ditemukan pada seluruh lokasi penelitian. Menggunakan 3 primer yangmengkode gen virulensi dari 10 isolat P. oryzae, hanya tiga haplotipe yangditemukan yakni C-011, E-010, dan F-110. haplotipe C-011 paling dominan (8isolat), E-010 (1 isolat), dan F-110 (1 isolat).
Kata kunci: Pyricularia oryzae,ras, varietas diferensial, gen virulensi

vi
ABSTRACT
RIANINGSIH. The Studi of the Race Diversity of Pyricularia oryzae Cavara isolate asthe Cause of Blast Disease in Rice Crops in Several Regencies in South Sulawesi(Supervised by Tutik Kuswinanti and Andi Nasruddin)
The research aimed (1) to obtain data about the spreading of the Pyriculariaoryzaerace in several areas in South Sulawesi; and (2) to analyze the genes whichlinked to the virulence nature of P.oryzae causing the blast disease
The Determination of blast disease race used 1 set of differential varietieswhich consisted of 7 varieties of Asahan, Cisokan, IR 64, Krueng Aceh, Cisadane,Cisanggarung and Kencana Bali. The intensity of blast attack was observed onseven days after inoculation using the IRRI evaluation standard (1996). 5-9 scalepatch types were used to identify host vulnerability levels. The P.oryzae fungusgenotype test was perform using gene-based markers for vilurensi properties, i.eErg2 (1,440 bp), Pwl2 (900 bp), and Cut1 (1,730bp) markers. The amplified DNAbands appeared for each primary of each isolate with a value of 1(present) and 0(absent).
The Total of 20 isolates were collected from 3 different regencies of SouthSulawesi: 8 isolates in Bone, 7 isolate in maros, 4 isolate in Gowa. Based on theirmorphological and macroscopic features, the referred to P.oryzae the inoculation ofP.oryzae isolates on 7 differential varieties of rice showed the 12 races were existingbetween P.oryzae isolated tested. However, the 020 races were dominant in allregencies where the samples were collected. By using 3 primers coded for virulencegene of P.oryzae, only three haplotypes were found, only three haplotypes werefound: the haplotypes of C-011, E-010 and F-110. The haplotype of C-011 was foundto be the dominant (8 isolates) while E-010 (1 isolate) and F-110 (1 solate).
Keywords: Pyricularia oryzae, race, differential varieties, virulence genes
\

vii
KATA PENGANTAR
Assalamu Alaikum Warahmatullahi Wabarakatuh.Puji syukur penulis panjatkan kehadirat Allah SWT dan tak lupa kita kirimkan
shalawat dan taslim atas junjungan Nabi Muhammad SAW atas rahmat dan
karunia-Nya penulis dapat menyelesaikan Tesis ini Sebagai syarat untuk dapat
memperoleh gelar Magister Sains.
Selesainya tesis ini tidak terlepas dari doa dari kedua Orang tua tercinta saya
Ayahanda Nandu dan Ibunda Johar, serta Saudara-saudaraku Nining Juliana,
Amd.Keb. dan Haerul Ikram sehingga dapat menyelesaikan penulisan tesis ini.
Pada kesempatan ini pula penulis mengucapkan terima kasih kepada;
1. Ibu Prof. Dr. Tutik Kuswinanti, M.Sc selaku pembimbing 1 dan Bapak Dr.
Ir. Andi Nasruddin, M.Sc selaku pembimbing 2 telah banyak membantu,
memberikan bimbingan, mengarahkan dan memberi saran kepada penulis
dalam menyelesaikan tesis ini.
2. Upacan terima kasih penulis juga disampaikan kepada Bapak Prof. Dr. Ir.
Nur Amin, Dipl.Ing. Agr., Bapak Dr. Ir. Tamrin Abdullah, MS, dan Ibu Dr. Vien
Sartika Dewi yang telah meluangkan waktunya untuk menjadi penguji dan
juga memberikan masukan dalam Seminar proposal, seminar hasil dan saat
ujian tutup.
3. Bapak dan Ibu dosen Program Studi Magister Ilmu Hama dan Penyakit
Tumbuhan, atas ilmu dan perhatian, didikan dan dorongan yang diberikan
kepada penulis sehingga penulis mendapat motivasi dalam menempuh
pendidikan sarjana Strata 2 .
4. Para staf pegawai Departemen Hama dan Penyakit khususnya pak Ardan
dan Pak Kama yang telah membantu penulis dalam menyediakan Alat-alat
penelitian.

viii
5. Teman -teman Magister IHPT angkatan 2015 yang saya sayangi Ramlah,
Rahma, Fatmawati, Didiana Y. Molebila, Nur Hardina, Jazman dan Zulfikar
yang telah banyak membantu dalam proses penelitian.
6. Tak lupa ucapan terima kasihku kepada Akbar Amri yang telah meluangkan
waktu dalam pengambilan sampel di lapangan.
Penulis menyadari bahwa tulisan ini masih jauh dari kesempurnaan, untuk itu
penulis mengharapkan saran dan kritik dari pembaca yang bersifat membangun
demi kesempurnaan tulisan ini. Akhir kata penulis mengucapkan terima kasih dan
semoga tulisan ini bermanfaat bagi kita semua.
Wassalamu Alaikum Warahmatullahi Wabarakatu
Makassar, Nopember 2017
Penyusun

ix
DAFTAR ISI
Halaman
ABSTRAK .......................................................................................... v
KATA PENGANTAR ........................................................................... vii
DAFTAR GAMBAR ............................................................................. xi
DAFTAR TABEL ................................................................................ xii
I. PENDAHULUAN ........................................................................... 1
1.1 Latar Belakang .............................................................................. 1
1.2 Tujuan............................................................................................ 6
1.3 Kegunaan ..................................................................................... 6
1.4 Rumusan Masalah ....................................................................... 6
1.5. Hipotesis....................................................................................... 7
1.6 Kerangka Pikir Penelitian . ............................................................. 7
II. TINJAUAN PUSTAKA ................................................................... 8
2.1 Penyakit Blas ............................................................................... 8
2.1.1.Sistematika ......................................................................... 9
2.1.2 Biologi Penyakit ................................................................... 9
2.1.3 Gejala Penyakit Blas ........................................................... 10
2.1.4 Keragaman dan penyebaran Ras Pyricularia oryzae Cav .... 12
2.1.5 Epidemiologi Penyakit Blas .................................................. 16
2.1.6.Siklus penyakit Blas ............................................................. 19
2.1.7 Penanda SCAR (Sequence Characterized Amplified
Region Marker) ..................................................................... 20
III. METODE PENELITIAN .................................................................. 22
3.1 Tempat dan Waktu........................................................................ 22
3.2 Bahan dan Alat ............................................................................. 24
3.3 Metode Pelaksanaan .................................................................... 23
3.3.1 Pengambilan sampel ........................................................... 23
3.3.2 Pengamatan Intensitas Serangan dilapangan ..................... 23
3.3.3 Isolasi dan Identifikasi cendawan P. oryzae ........................ 23

x
3.4. Uji Ras iIsolat Cendawan P.oryzae ............................................... 26
3.5 Deteksi Gen virulensi isolat Pyricularia oryzae Cav
dengan Metode PCR .................................................................... 28
3.5.1 Isolasi DNA Genom.............................................................. 28
3.5.2 Amplifikasi PCR ................................................................... 29
3.6. Analis Data ................................................................................... 30
IV. HASIL DAN PEMBAHASAN ......................................................... 31
4.1 Hasil .............................................................................................. 31
4.1.1 Pengamatan Intensitas Serangan Blas Daun ...................... 31
4.1.2 Isolasi dan Karakteristik Morfologi cendawan P. oryzae ....... 31
4.1.3 Uji Ras Isolat- isolat P. oryzae ............................................. 34
4.1.4 Hasil Analisis PCR cendawan P. oryzae .............................. 36
4.2 Pembahasan ................................................................................. 39
V . KESIMPULAN DAN SARAN ........................................................ 47
5.1 Kesimpulan ............................................................................ 47
5.2 Saran ..................................................................................... 47
DAFTAR PUSTAKA............................................................................ 48
LAMPIRAN.......................................................................................... 55

xi
DAFTAR GAMBAR
Teks
Gambar 1.Kerangka Pikir .................................................................... 7
Gambar 2. Gejala blas daun . .............................................................. 11
Gambar 3. Gejala blas pada padi (A) leher malai, (B) kolar. ................ 12
Gambar 4. Morfologi isolat cendawan P.oryzae yang ditemukan ........ 33
Gambar 5. (a dan b) Hifa dan (c ) Klamidospora dari cendawan
P. oryzae ............................................................................ 33
Gambar 6. (A) Pembentukan Konidia, (B) Konidia yang masih
menempel dikonidiofor dengan dua septa, (C ) Konidia
dengan dua septa dengan tiga sel pembesaran 40x
menggunakan mikroskop Adapter...................................... 34
Gambar 7. Gejala serangan penyakit blas pada tanaman padi diferensial
Setelah inokulasi................................................................ 35
Gambar 8. Pola pita hasil amflipikasi DNA genomic cendawan P.oryzae
Dengan gen Pwl1. .............................................................. 38
Gambar 9. Pola pita hasil amflipikasi DNA genomic cendawan P.
Dengan gen Erg2. .............................................................. 38
Gambar 10. Pola pita hasil amflipikasi DNA genomic cendawan
P.oryzae dengan gen Cut1................................................... 38
LAMPIRAN
Gambar lampiran 1. Survey lokasi serangan blas di kab. Bone ........... 56
Gambar lampiran 2. Survey lokasi serangan blas di kab. Maros ......... 56
Gambar lampiran 2. Survey lokasi serangan blas di kab. Gowa ......... 56

xii
DAFTAR TABEL
TEKS
Tabel 1. Skor keparahan Penyakit (IRRI, 1996) .................................. 27
Tabel 2. Contoh pemberian nomor ras P. oryzae asal padi ................ 28
Tabel 3. Intensitas serangan blas daun pada dua lokasi Pengambilan
Sampel ................................................................................... 31
Tabel 4. Jumlah isolat yang ditemukan pada daerah pengambilan
sampel .................................................................................. 32
Tabel 5. Reaksi Ketahanan variietas padi diferensial terhadap 20 isolat
cendawan P. oryzae............................................................... 36
Tabel 6. Pengelompokan haplotip cendawan Pyricularia oryzae
berdasarkan keberadaan tiga gen virulensi............................ 37

1
BAB IPENDAHULUAN
1.1 Latar Belakang
Upaya peningkatan produktivitas padi terus dilakukan mengingat padi
merupakan tanaman pangan yang menjadi makanan pokok di Indonesia. Tetapi
keberadaan serangan penyakit pada tanaman padi merupakan faktor pembatas
produksi. Penyakit penting yang menyerang padi adalah penyakit blas yang
disebabkan oleh Pyricularia oryzae Cavara sinonim (Pyricularia grisea (Cooke)
Sacc) dengan bentuk teleomorph: Magnaporthe grisea). Blas merupakan penyakit
paling merusak pada tanaman padi (Couch dan Kohn, 2002) yang mulai dari fase
vegetativ sampai stadia pembentukan malai atau generatif. Serangan yang berat
terjadi pada stadia generatif, karena dapat menimbulkan puso dan atau
menggagalkan panen (Santika dan Sunaryo, 2008).
Cendawan P.oryzae dapat menyebabkan bercak pada daun padi, buku
(node blast), leher malai (neck blast), bulir padi (grain blast) (Chen 1993;Scardaci et,
al 1997) dan kolar daun (collar blast) (Scardaci et al. 1997). Bercak pada daun padi
memiliki bentuk yang khas yaitu belah ketupat dengan bagian tepi berwarna coklat
dan bagian tengah berwarna putih keabu-abuan (Santoso dan Nasution, 2012).
Bentuk dan ukuran bercak dipengaruhi oleh kondisi lingkungan dan kultivar.
Cendawan ini tidak hanya menyerang padi, tetapi juga dapat menimbulkan
kerusakan pada tanaman serealia lain seperti gandum dan sorgum (Kahmann dan
Basse 1997)

2
Kehilangan hasil akibat penyakit blas pada padi dari suatu daerah di dunia
diperkirakan berkisar antara 50-100%. Diperkirakan bahwa tiap tahunnya penyakit
ini menghancurkan padi yang akan cukup dimakan oleh lebih dari 60 juta orang.
Kehilangan hasil secara ekonomi tidak dapat terhitung, tetapi beberapa data
menunjukkan nilainya lebih dari 70 milyar dolar pada beberapa negara di Asia
(Scheuermann et al. 2012) sedangkan di Indonesia luas serangan penyakit blas
menduduki urutan kelima berda sarkan kompilasi data statistik pertanian IV yang
dilaporkan oleh Direktorat Perlindungan Tanaman pangan. Rata-rata luas serangan
penyakit blas dalam kurun waktu 10 tahun terakhir adalah 9.778 Ha/tahun (Soetarto
et al. 2001). Pada tahun 2005, penyakit blas menyebabkan pertanaman varietas
Fatmawati mengalami puso sebanyak 500 ha di Tulang Bawang Lampung. Menurut
Amir dan Kardin (1991), serangan blas leher dapat mencapai 90% dan
menyebabkan kehilangan hasil padi mencapai 50 - 90% pada varietas rentan.
Serangan penyakit tersebut cenderung meningkat setiap tahun. (Deptan 2009).
Balai Besar Peneltian Padi (2015) melaporkan bahwa luas serangan penyakit blas
dapat mencapai luas 1.285 juta ha atau sekitar 12% dari total luas areal pertanaman
padi di Indonesia.
Sulitnya mengendalikan penyakit ini karena P.oryzae mempunyai keragaman
genetik yang tinggi dan sifat perkembangan seluler dan morfologi yang sangat
adaptif pada tanaman padi yang diinfeksi (Koizumi, 2009). Sifat-sifat tersebut
menyebabkan ras-ras cendawan P. oryzae dapat berubah sifat virulensinya dalam
waktu singkat, bergantung pada inang dan pengaruh lingkungan (Utami dkk, 2006).
Berkembangnya penyakit blas pada padi sawah diduga berkaitan dengan teknik budi

3
daya padi, khususnya penggunaan pupuk N dosis tinggi, serta penanaman varietas
yang tidak memiliki ketahanan terhadap penyakit ini. Fenomena semacam ini terjadi
juga di beberapa Negara seperti Jepang, Filipina, Vietnam, dan Korea (Kobayashi
et al. 2007).
Cara pengendalian penyakit blas dapat dilakukan dengan berbagai cara di
antaranya dengan teknik budidaya, penanaman varietas tahan, dan penggunaan
fungisida. Penggunaan varietas tahan merupakan cara yang paling efektif,
ekonomis, dan mudah dilakukan. Namun, penggunaan teknologi ini berhadapan
dengan patogen blas yang memiliki keragaman genetik dan kemampuan
beradaptasi yang tinggi sehingga dengan cepat membentuk ras baru yang dapat
mematahkan ketahanan varietas yang baru diperkenalkan (Santoso, dkk 2007,
Fukuta et al, 2009;Lestari dkk 2011). Penyebab terbentuknya populasi bersifat
dinamis ini antara lain adalah adanya kemampuan dalam melakukan rekombinasi
baik secara seksual maupun aseksual (Zeigler, 1998). Sejumlah varietas unggul
yang ditargetkan untuk mengendalikan penyakit blas di suatu lingkungan hanya
dapat berkembang selama dua sampai tiga musim saja (Amir dkk. 2000).
Penelitian mengenai penyebaran ras di Indonesia sudah banyak dilakukan.
Sebanyak 18 ras yaitu ras 001, 021, 040, 041, 051,061, 071, 073, 100, 101, 121,
201, 203, 241, 301, 333, 341, dan 343 telah teridentifikasi di bebrapa lokasi di
Indonesia seperti Sumatera (Kayu Agung, Lampung), Kalimantan Tengah
(Dadahup), Bali (Tabanan) dan Jawa Barat (Kuningan) hasil monitoring tahun 2007
hingga 2008 (Santosa dan Nasution, 2009). Belum ada data tentang penyebaran
ras di Sulawesi Selatan.

4
Menurut Amir dkk (2000), terdapat empat ras yang ada setiap musim tanam
yaitu ras 001, 003, 033, dan 173, artinya isolat ini memiliki penyebaran yang luas.
Isolat ras 001 mempunyai tingkat patogenesitas yang paling rendah virulensinya,
menyebar luas dan mampu bertahan lama di lapangan (Sari 2008; Utami 2000).
Adapun penentuan ras baru dari patogen blas di Indonesia dilakukan dengan
menggunakan varietas diferensial yaitu Asahan, Cisokan, IR64, Krueng Aceh,
Cisadane, Cisanggarung, dan Kencana Bali (Mogi et al. 1991). Oleh karena itu perlu
dilakukan penelitian tentang ras dari P.oryzae dengan memonitoring keberadaan
dan dominasi ras patogen P.oryzae di Sulawesi Selatan sebagai dasar rekomendasi
penanaman varietas tahan sesuai ras yang ada sangat diperlukan (Koizumi, 2009).
Gen-gen yang terdapat dalam varietas differensial standar internasional yang
terdiri dari galur-galur monogenik yang mengandung 25 gen ketahanan pada
penyakit blas yakni (Pia, Pib, Pii, Pik, Pik-h, Pik- m, Pik-p, Pik-s, Pish, Pit, Pita, pita-
2, Piz, Piz-5(=Pi2), Piz-t, Pi 1, Pi 3, Pi5(t), Pi7, Pi9, Pi11(t), Pi12(t), Pi12(t), Pi19(t)
dan Pi20) telah dikembangkan dan digunakan luas dibeberapa wilayah (Kobayashi
et al, 2007). Di Indonesia digunakan galur monogenic IRBLkh-K3, IRBLz5- CA®, dan
IRBLz-Fu yang masing –masing mengandung gen ketahanan terhadap penyakit blas
Pik-h, Pi2(t), dan Piz yang memilki ketahanan yang luas pada isolat-isolat P.oryzae
(Santoso dan Nasution 2009). Sedangkan galur monogenik IRBL-b-B(Pib) dan
IRBLt-K59 (Pit) menunjukkan reaksi rentan tehadap isolat-isolat Blas di Indonesia
sehingga penggunaannya tidak efektif (Santoso dkk, 2007).

5
Analisis keragaman genetik dari patogen blas juga dilakukan menggunakan
marker MGR 586 dan marker spesifik menggunakan Cut1, Erg2. Hasil penelitian
Reflinur dkk (2005), untuk mengetahui keragaman genetik dari 230 isolat cendawan
P. oryzae dari lima lokasi, yaitu Lampung, Sukabumi, Sumatera Utara, Sumatera
Barat, dan Bogor dengan menggunakan primer spesifik gen virulensi yang berperan
dalam pengenalan inang P.oryzae yaitu Cut1, Erg2, dan Pwl2. Gen Cut1 merupakan
lokus menjadikan gen cutinase berfungsi dalam mendegradasi lapisan kutikula
tanaman (Sweigard et al.1992). Gen Erg2 berperan sebagai penyandi metabolit
sekunderpada cendawan menjadi target antifungal pada sel tanaman (Keon et al.,
1994), sedangkan gen Pwl2 berfungsi sebagai gen avirulen yang bersifat sfesifik
inang (Valent dan Chumley, 1994). menghasilkan delapan haplotip, yaitu haplotip A-
000, B-001, C-011, D-111, E-010, F-110, G-100, dan H-101 dimana Haplotip D-111
paling banyak ditemukan pada setiap lokasi asal isolat P. oryzae.
Penelitian yang serupa dilakukan oleh Lestari dkk,(2014) dengan
menggunakan gen spesifik virulensi yaitu Cut1, Erg2, Pwl2 diperoleh 6 haplotipe dari
16 isolat hal ini menunjukkan bahwa isolat-isolat tersebut memiliki keragaman yang
tinggi. Tingginya keragaman haplotipe dipengaruhi oleh faktor lingkungan seperti
suhu dan kelembaban baik pada lokasi yang sama ataupun pada lokasi yang
berbeda.
Berdasarkan uriaian tersebut perlu dilakukan penelitian mengenai marka
molekuler berbasis PCR menjadi metode yang ampuh untuk mendeteksi variasi
genetik dari penyakit tanaman termasuk cendawan Pyricularia oryzae (Annamalai et
al,. 1995; Lestari dkk, 2014). Variasi ini berhubungan erat dengan variasi genetik

6
yang sangat dipengaruhi oleh sejumlah tekanan seleksi patogen (Chada dan
Gopallehisnan, 2005; Shrinivasachary et al,. 2002) sehingga penggunaan marka
spesifik gen virulensi dapat membantu dalam memberikan informasi tentang
mekanisme potensial dan variasi ras cendawan Pyricularia oryzae (Lounge dan de
Wilt, 1988).
1.2 Tujuan
Tujuan penelitian ini adalah untuk memperoleh data tentang penyebaran ras
Pyricularia oryzae pada beberapa wilayah di Sulawesi Selatan serta analisa gen
yang terpaut dengan sifat virulensi Pyricularia oryzae penyebab penyakit blas.
1.3 Kegunanan Penelitian
Sebagai acuan atau rujukan tentang keberadaan ras-ras P. oryzae yang ada di
Sulawesi Selatan, serta informasi marka spesifik yang terpaut dengan gen
virulensinya. Data yang diperoleh dapat bermanfaat bagi pemulia tanaman dalam
upaya merakit varietas-varietas padi yang tahan terhadap penyakit blas.
1.4 Rumusan Masalah
1. Berapa banyak ras patogen Pyricularia oryzae yang ada di Sulawesi
Selatan?
2. Bagaimana penyebaran ras Pyricularia oryzae yang ada di Sulawesi
selatan?
3. Deteksi gen virulensi dari isolat-isolat Pyricularia oryzae yang ditemukan?

7
1.5 Hipotesis penelitian
1. Terdapat keragaman ras pada isolat-isolat P. oryzae yang dikoleksi
2. Terdapat ras yang dominan pada tiap-tiap lokasi yang diuji
3. Terdapat perbedaan gen virulensi pada masing-masing ras P. oryzae
1.6 Kerangka Pikir
Gambar 1. Kerangka pikir penelitian
Padi (Oryza sativa L
Penyakit blas (Pyricularia oryzae Cavara)
Varietas padi diferensial
Indonesia
Uji ras fisiologi
Karaterisasi gen virulensicendawan menggunakanprimer spesifik
AsahanCisokanIR64,Krueng Aceh,CisadaneCisanggarungKencana Bali
Keragaman Ras
Isolasi Cendawan P. oryzae Karakterisasi morfologi

8
BAB IITINJAUAN PUSTAKA
2.1 Penyakit Blas
Penyakit blas disebabkan oleh cendawan Pyricularia oryzae Cavara.
Termasuk dalam kelompok ascomycetes dan bersifat heretalik (Ziegler, 1998). Di
Alam, Cendawan ini ditemukan hanya dalam bentuk asesksualnya. Sedangkan
bentuk seksual hanya diperoleh dengan pengkulturan di Laboratorium ( Valent,
1990). Secara morfologi cendawan P. oryzae mempunyai konidia berbentuk bulat,
lonjong, tembus cahaya, dan bersekat dua dengan tiga ruangan (Ou, 1985).
Serangan blas leher dapat mencapai 90% dan menyebabkan kehilangan
hasil padi mencapai 50 - 90% pada varietas rentan (Amir dan Kardin, 1991).
Serangan penyakit tersebut cenderung meningkat setiap tahun. (Deptan, 2009).
Balai Besar Peneltian Padi (2015), melaporkan bahwa luas serangan penyakit blas
dapat mencapai luas 1.285 juta ha atau sekitar 12% dari total luas areal pertanaman
padi di Indonesia. Di Sulawesi Selatan, ledakan penyakit blas pernah terjadi di
Kabupaten Sinjai dan Bulukumba pada tahun 1980, lebih dari 900 ha lahan sawah
yang ditanami padi varietas Semeru terinfeksi blast leher berkisar antara 6 sampai
85% di Bulukumba dan 80 ha lebih dari 85% infeksi blast leher, dan sekitar 390 ha
terinfeksi blas daun (Wakman et al., 1980)
Daerah endemik penyakit blas di Indonesia adalah Lampung, Sumatra
Selatan, Jambi, Sumatra Barat, Sulawesi Tengah, Sulawesi Tenggara, dan Jawa
Barat (Sukabumi). Penyakit blas, khususnya blas leher, menjadi tantangan yang
makin serius karena banyak ditemukan pada beberapa varietas padi sawah di Jawa

9
barat (Sukabumi, Kuningan), Lampung (Tulang Bawang, Lampung Tengah), dan
Bali (Tabanan) (Yuliani dan Maryana.,2014).
2.1.1 Sistematika
Menurut CABI (2016), klasifikasi dari cendawan Pyricularia oryzae penyebab
penyakit blas sebagai berikut:
Domain : Eukaryota
Kingdom : Fungi
Phylum : Ascomycota
Subphylum : Pezizomycotina
Class : Sordariomycetes
Subclass : Sordariomycetidae
Family : Magnaporthacea
Genus : Magnaporthe
Species : Magnaporthe oryzae
SinonimDactylaria oryzae ( Cavara) Sawada, (1917)
Pyricualaria grisea Sacc., (1880) (anamorph)
Pyricularia oryzae Cavara, (1891)
2.1.2 Biologi Penyakit
Pyricularia oryzae menghasilkan spora seksual (askospora) dalam struktur
yang disebut asci dan diklasifikasikan kedalam famili Magnaporthaceae. Asci
ditemukan dalam struktur khusus disebut perithecia. Miselium dari M. oryzae ini
bersepta dan spora cendawan ini bersifat haploid. Reproduksi secara seksual atau
teleomorph dari cendawan M.oryzae dapat diproduksi di Laboratorium jika isolat
dari jenis yang berbeda dipasangkan. Cendawan ini bersifat heterotalik dengan
sistem kawin bipolar (dikendalikan oleh dua alel yang berbeda pada lokus tunggal)
dengan gen tambahan mengendalikan siklus seksual (Tebeest, 2007).

10
Tahap aseksual disebut Pyricularia oryzae merupakan bentuk spora yang
paling umum ditemukan, dapat ditemukan pada luka atau gejala dari penyakit ini.
Konidia dari cendawan ini bersel tiga dan diproduksi di apex pada konidiofor.
Sporulasi koloni pada media agar biasanya fleecy dan berwarna agak keabu-abuan
(Tebeest, 2007).
2.1.3 Gejala Penyakit Blas
Penyakit blas menyerang tanaman padi mulai dari persemaian sampai
pengisian bulir padi. Cendawan P. oryzae dapat membentuk bercak pada daun padi,
buku batang, leher malai, cabang malai, bulir padi, dan kolar daun (Chen, 1993;
Scardaci et al., 1997). Bercak pada pelepah daun padi jarang ditemukan. Penyakit
blas menimbulkan dua gejala khas, yaitu blas daun yang menyerang tanaman padi
pada fase vegetatif dan blas leher yang menyerang pada awal pembungaan
(Bonman, 1992). Serangan serius pada fase vegetatif dapat menyebabkan matinya
tanaman padi dan pada fase generatif dapat menyebabkan patahnya leher malai
dan hampanya bulir padi.
Bentuk khas dari bercak blas daun adalah belah ketupat dengan dua
ujungnya kurang lebih runcing. Awalnya bercak berukuran kecil berwarna hijau
gelap, abu-abu sedikit kebiru-biruan. Bercak ini terus membesar pada varietas yang
rentan, khususnya bila dalam keadaan lembab. Bercak yang telah berkembang
terlihat pada bagian tepi berwarna coklat dan bagian tengah berwarna putih keabu-
abuan. Bercak yang telah berkembang penuh mencapai panjang 1–1,5 cm dan lebar
0,3–0,5 cm dengan tepi berwarna coklat (Ou, 1985).

11
Gambar 2. Gejala blas daun
Bercak pada daun yang rentan tidak membentuk tepi yang jelas. Bercak
tersebut dikelilingi oleh warna kuning pucat (halo area), terutama pada lingkungan
yang kondusif seperti keadaan lembab dan ternaungi. Selain itu, perkembangan
bercak juga dipengaruhi oleh kerentanan varietas dan umur bercak itu sendiri.
Bercak tidak akan berkembang dan tetap seperti titik kecil pada varietas yang tahan.
Hal ini disebabkan oleh proses perkembangan konidia dari cendawan P. oryzae
dalam jaringan inangnya terhambat. Pada lingkungan yang kondusif, bercak-bercak
tersebut dapat menyatu dan menyebabkan rusaknya sebagian besar daun. Blas
daun dapat menyebabkan kematian keseluruhan tanaman pada varietas rentan
yang masih muda sampai stadia anakan (Scardaci et al.1997).
Blas leher berupa bercak coklat kehitaman pada pangkal leher yang dapat
mengakibatkan leher malai tidak mampu menopang malai dan patah karena tangkai
malai membusuk. Apabila infeksi patogen P. oryzae terjadi sebelum pengisian bulir
dapat menyebabkan kehampaan bulir tanaman padi. Serangan patogen blas tidak

12
hanya pada bagian daun dan malai, namun bagian batang juga dapat terinfeksi
sehingga batang padi membusuk dan rebah (Ou, 1985). Serangan P. oryzae pada
kolar daun (daerah pertemuan antara helaian daun dan pelepah) menimbulkan
gejala blas kolar berwarna coklat. Blas kolar yang terjadi pada daun bendera atau
pada daun kedua terakhir dapat menyebabkan pengaruh yang nyata pada produksi
padi (Scardaci et al. 1997)
Gambar 3. Gejala blas pada padi (A) leher malai, (B) kolar.
2.1.4 Keragaman dan penyebaran Ras Pyricularia oryzae Cav.
Keberadaan ras-ras dari patagen P.oryzae telah dicatat oleh Sasaki di
Jepang pada tahun 1922. Sejak tahun 1950-an, beberapa negara terutama Jepang,
Amerika, dan Taiwan mulai mempelajari ras-ras dari cendawan patogen tersebut
lebih intensif. Masing-masing negara menggunakan set yang berbeda dari varietas
yang beragam. Banyak ras yang diidentifikasi di setiap negara tersebut. Kemudian
pada tahun 1960-an mulai ada laporan mengenai ras berikutnya dari Philipina,
Indenesia, Korea, dan Kolombia (Bonman, 1992).
A B

13
P. oryzae diketahui mempunyai banyak ras fisiologi yang berbeda-beda sifat
dan virulensinya. Pada tahun 1975, IRRI sudah melaporkan adanya 260 ras fisiologi
(Semangun, 1991). Konidium-konidium yang dihasilkan oleh suatu biakan murni
yang berasal dari konidium tunggal dapat berkembang menjadi banyak ras
(Thurston, 1984). Patogenisitas dipengaruhi oleh perbedaan mekanisme
metabolisme dan senyawa-senyawa kimia yang terdapat pada cendawan P. oryzae
(Otsuka et al., 1963). Para peneliti Jepang dan India membuktikan bahwa P.oryzae
mampu membentuk beberapa macam toksin, antara lain ß- picolinic acid, pyricularin,
pyriculol, dan tenuazonic acid (Ou, 1985).
Perbedaan reaksi suatu varietas terhadap blas disebabkan oleh adanya
perbedaan dan perubahan ras antar lokasi dan adanya perubahan ras yang
dominan di suatu wilayah sebaran. Hingga saat ini telah terdeteksi 64 ras blas,
beberapa diantaranya terdapat di Sitiung (Sumatera Barat). Ras baru ini terutama
ditemukan di Karang Agung (Sumatera Selatan) dan Jawa Barat. Ras-ras blas
tersebut dapat menyerang varietas Lematang, Kapuas, Krueng Aceh, IR64, Cisokan
dan Cisadane. Di Sitiung, ras-ras blas dapat menyerang padi gogo varietas Sentani,
Tondano, Maninjau, Ranau, Arias, Bicol, dan C-22. Sedangkan varietas Semariti dan
Sirendah masih mampu bertahan (Nasution dkk. 1992).
Hasil monitoring terhadap perkembangan populasi patogen blas yang
dilakukan di Lampung dari tahun 2000 hingga 2004 menunjukkan bahwa di wilayah
tersebut setiap musim tanam dapat diidentifikasi 13-17 ras yang berbeda dengan
proporsi yang beragam. Selama lima tahun monitoring tersebut diperoleh total 26 ras
yang berbeda di antaranya terdapat 7 ras yang selalu muncul di setiap tahun yaitu

14
ras 001, 023, 033, 073, 101, 133, dan 173. Hasil monitoring 2007 hingga 2008 dari
beberapa lokasi di Indonesia seperti Sumatera (Kayu Agung, Lampung), Kalimantan
Tengah (Dadahup), Bali (Tabanan) dan Jawa Barat (Kuningan) terindentifikasi
sebanyak 18 ras seperti ras 001, 021, 040, 041, 051, 061, 071, 073, 100, 101, 121,
201, 203, 241, 301, 333, 341, dan 343 (Santoso dan Nasution, 2009).
Sampel daun sakit di Propinsi D.I. Yogyakarta menunjukkan bahwa penyakit
blas yang berkembang adalah ras 033, 133, dan 173. Di Propinsi Jawa Barat yaitu
Kabupaten Sukabumi telah diketahui sebagai daerah endemik penyakit blas. Data
monitoring menunjukkan bahwa tingkat keragaman populasi patogen blas di
Sukabumi cukup tinggi. Beberapa ras yang ditemukan di Kab. Sukabumi yaitu Ras
001, 123, 133, 173, dan 243 (Santoso dkk, 2007). Ras pada setiap musim tanam
selalu ada perubahan baik dalam jumlah dan dominasinya. Ras 001, 003, 033, dan
173 merupakan ras yang selalu ada pada setiap musim tanam dan ras 041, 043,
051, 061, 073, 141, 161, 163, dan 173 adalah ras-ras yang dapat menyerang
varietas IR64 (Amir dkk, 2000).
Keragaman ras fisiologis patogen blas merupakan tantangan utama dalam
pengembangan padi gogo di lahan kering marginal. Populasi P. oryzae sangat
beragam dan terdiri dari individu-individu ras yang mempunyai sifat virulensi yang
berbeda (Zeigler et al. 1994). Dominasi ras P. oryzae di suatu wilayah dengan
wilayah lain yang berbeda memungkinkan varietas padi di suatu wilayah tahan tetapi
rentan di wilayah lain. Informasi sebaran ras P. oryzae sangat diperlukan untuk
memprediksi kesesuaian varietas yang akan dilepas (spesifik lokasi). Hal ini sangat
penting untuk regionalisasi varietas sesuai dengan sebaran ras P. oryzae di wilayah

15
tersebut. Dengan diketahuinya sebaran ras P.oryzae yang dominan di suatu wilayah
endemik blas maka pengendalian penyakit blas akan lebih efektif dengan
menggunakan varietas tahan yang disesuaikan dengan ras P. oryzae di wilayah
tersebut.
Pyricularia oryzae penyebab blas padi memiliki keragaman ras fisiologi tinggi.
Jumlah ras fisiologi meningkat di beberapa negara (Ou 1980). Banyak varietas baru
yang resisten terhadap penyakit blas hanya bertahan selama beberapa tahun saja,
menjadi rentan terhadap strain Pyricularia dengan virulen baru. Sejumlah ras berada
di lapangan sebagai hasil perubahan terus menerus dalam patogenisitas dan jumlah
konidium yang sangat banyak (Ou, 1980;Namai, 2011). Sebaliknya frekuensi ras
fisiologi cendawan blas pada padi relatif stabil di Korea dan Filipina. Resistensi padi
secara kuantitatif terhadap penyakit blas bukan akibat ketidakstabilan cendawan
patogen yang ekstrim (Bonman et al. 1987).
Karthikeyan et al (2013), melaporkan bahwa dari 600 sampel daun padi
yang terinfeksi blas daun dan blas leher, dalam karakterisasi virulensinya terhadap
padi, 198 ras P. grisea dilakukan dengan 9 NILs (Near-isogenic rice lines). Dalam
analisis yang dilaksankan pada tahun 2001, diidentifikasi 15 patotipe baru di antara
49 strains Kerala. Dalam tahun 2002, 14 patotipe diidentifikasi dari 26 strain Tamil
Nadu dan 9 patotipe dari 22 strains Karnataka. Hal serupa dilakukan oleh Neto et al.,
(2010), terhadap 479 sampel isolate P. grisea yang diperoleh dari lahan sawah
irigasi Araguaia River Valley, Tocantins, Brazil. Sejalan dengan itu, Nasution dkk.
(2012), melaporkan bahwa hasil identifikasi ras 175 isolat P. grisea yang diperoleh
dari padi sawah di daerah Subang, Indramayu, Kuningan, Bogor, Sukabumi, dan

16
Cianjur, diperoleh 22 kelompok ras. Diantara ras yang diidentifikasi, ras 101 adalah
ras yang paling dominan sebesar 24,6 % disusul ras 001 sebesar 18,3 % , ras 041
sebesar 17,7% dan ras 033 sebesar 11,4%. , ras 001(12,5%) dan ras 073 sebesar
10,7%. Ras P. grisea yang paling dominan di Kabupaten Subang adalah ras 053
(18%), di Karawang ras 003 (25%) dan di Kabupaten Indramayu ras 003 (21%).
Komposisi dan sebaran ras jamur P. grisea di Kabupaten Subang, Karawang, dan
Indramayu berbeda-beda, di Kabupaten Subang ditemukan 16 kelompok ras, di
Karawang 11 kelompok, dan di Indramayu ada 12 kelompok ras P. grisea.
Sudir dkk (2013), melaporkan hasil pemantauan ras blas di enam kabupaten
di Jawa Tengah pada musim tanam 2013 diperoleh 122 isolat jamur P. grisea yang
terdiri atas 23 ras. Ras 013 dan 153 yang paling dominan yaitu masing-masing
sebesar 9,84%, dan ras 001, 113 dan 151 masing-masing sebesar 7,38%.
Komposisi dan sebaran ras jamur P. grisea berbeda antar lokasi.
2.1.5 Epidemiologi Penyakit Blas
Cendawan Pyricularia oryzae termasuk ke dalam kelompok Ascomycetes
dan bersifat heterotalik. Cendawan ini ditemukan di alam dalam bentuk aseksualnya
saja sedangkan bentuk seksualnya, yaitu Magnaporthe grisea (Herbert) Barr, hanya
dihasilkan dengan pengkulturan di laboratorium. Secara morfologi, cendawan P.
grisea mempunyai konidia berbentuk bulat, lonjong, tembus cahaya, dan bersekat
dua (3 ruangan) (Ou, 1985). Konidia cendawan P. grisea akan berkecambah pada
kondisi optimum dengan cara membentuk buluh-buluh perkecambahan yang
selanjutnya menjadi appresoria (Bourett dan Howard, 1990). Appresoria akan

17
menembus kutikula daun dengan bantuan melanin yang ada pada dinding
appresoria. Proses penetrasi appresoria pada kondisi optimum berlangsung selama
8–10 jam (Chumley dan Valent, 1990).
Infeksi blas meningkat pada kombinasi suhu air rendah 17 °C dan suhu
udara sedang 32 °C (Tasugi dan Yoshida, 1959). Perkembangan penyakit blas juga
dipengaruhi oleh faktor lingkungan yaitu kesuburan tanah dan kelembaban udara.
Pada wilayah dengan kelembaban udara > 90% tanaman mengalami masa
berembun yang panjang (dew period). Hal ini mendukung perkembangan blas.
Hujan lebat yang terjadi dalam waktu singkat tidak membantu perkembangan
penyakit blas. Sebaliknya, hujan rintik-rintik tetapi lama merupakan kondisi yang
menguntungkan bagi blas untuk berkembang dan menginfeksi tanaman (Amir dan
Nasution, 1993). Winarti (1992), melaporkan bahwa intensitas serangan blas leher
pada tanaman padi dipengaruhi oleh curah hujan, suhu, kelembaban, lama
penyinaran matahari, dan intensitas radiasi matahari.
Kelembaban udara dan kelembaban tanah memengaruhi patogenisitas dan
pertumbuhan cendawan. Serangan penyakit blas lebih berat pada lahan kering
daripada lahan sawah, namun tergantung pada varietas padi yang ditanam.
Kelembaban udara mempengaruhi perkembangan bercak. Peran kelembaban udara
baik iklim makro maupun mikro serta pembentukan embun sangat menentukan
perkembangan penyakit blas. Di pesemaian, infeksi di bagian tengah lebih berat
dibandingkan bagian pinggir. Faktor naungan memiliki pengaruh terhadap
perkembangan bercak. Pesemaian dalam rumah kaca, akan lebih rentan apabila
kondisi lingkungan sedikit teduh dan ternaungi. Patogen blas berkembangbiak cepat

18
pada tanaman padi yang berjarak tanam rapat. Pada jarak tanam yang rapat
memiliki kelembaban udara mikro yang tinggi. Kecepatan pertumbuhan cendawan P.
oryzae juga akan semakin tinggi jika pemupukan urea dilakukan secara berlebihan.
Penyebaran penyakit blas sangat luas dan bersifat destrukif jika kondisi
lingkungan menguntungkan (Scardaci et al, 1997). Namun epidemi penyakit hanya
berkembang apabila terjadi kombinasi dan perkembangan yang baik dari banyak
faktor lingkungan seperti: kelembaban, suhu dan angin yang bertepatan dengan
tingkat kerentanan tanaman dengan produksi, penyebaran, inokulasi, penetrasi,
infeksi dan reproduksi patogen (Agrios, 1996). Menurut Ou (1979), penyebaran
spora blas dapat terjadi melalui angin, benih, sisa gabah, jerami tanaman sakit, sisa
tanaman padi dilapangan dan tanaman inang lainnya terutama dari golongan
graminae/rerumputan. Pada daerah tropik, sumber inokulum selalu ada sepanjang
tahun, karena adanya spora di udara dan tanaman inang selain padi (Santoso dan
Nasution, 2009). Cendawan P. oryzae dapat menjadi patogen pada beberapa
tanaman penting lainnya seperti gandum, sorghum dan serealia lainnya (Kahmann
dan Basse 1997), serta lebih dari 40 spesies gulma rumput-rumputan dan gulma
lainnya (Ou, 1985).
Patogen P. oryzae memiliki kemampuan untuk membentuk ras baru dengan
cepat menyebabkan pengendalian penyakit ini sulit dilakukan oleh petani. Virulensi
ras mudah bergeser ketingkat yang lebih tinggi dari ras semula sehingga dengan
cepat dapat mematahkan ketahanan varietas. Beberapa varietas unggul padi gogo
yang semula tahan terhadap penyakit blas setelah 2-3 musim ditanam di lapangan

19
berubah menjadi rentan. Selain itu, P. oryzae mempunyai banyak ras dan
virulensinya berbeda berdasarkan tempat dan musim tanam
2.1.6 Siklus penyakit Blas yang disebabkan oleh Pyricularia oryzae
Siklus penyakit dari penyakit blas meliputi tiga fase, yaitu infeksi, kolonisasi
dan sporulasi (Leung dan Shi, 1994). Infeksi dimulai dengan pembentukan konidia
bersepta dua yang dilepaskan oleh konidiofor, konidia berpindah ke permukaan
daun yang tidak terinfeksi melalui percikan air atau dengan bantuan angin. Konidia
mempunyai perekat atau getah sehingga dapat melengket pada daun (Hamer et
al.,1988). Selanjutnya appresoria akan menembus kutikula daun dengan bantuan
melanin yang ada pada dinding appresoria. Kondisi optimum untuk penetrasi
appresoria berlangsung 8-10 jam (Chumley dan Valent,1990). Pada tanaman yang
rentan dengan kondisi yang lembab konidia akan muncul selama 3-4 hari. Konidia
akan mudah menyebar dan menjadi inokulum untuk infeksi selanjutnya.
Besar kecilnya bercak menentukan banyaknya spora yang dihasilkan . bercak
yang kecil mampu menghasilkan 50-300 konidia selama 6-7 hari sedangkan bercak
yang besar menghasilkan konidia 2.500 – 6.000 konidia selama 14 hari di
Laboraturium (Hashioka,1965). Pembentukan spora mencapai puncaknya dalam
waktu 3-8 hari setelah timbul gejala awal pada daun dan 10-12 hari setelah
timbulnya gejala pada malai. Spora yang dihasilkan oleh bercak daun pada lima
daun dari atas dapat menginfeksi leher malai pada saat berbunga awal. umumnya
spora dilepaskan pada dini hari antara pukul 02.00-06.00. Air hujan sangat berperan
terhadap pelepasan spora (Kato et al., 1970), pada daerah tropis spora akan

20
dilepaskan siang hari setelah turun hujan. Kecepatan angin dan posisi daun
merupakan faktor penentu banyaknya spora yang tertangkap. Bila bercak hanya
berupa titik sebsar ujung jarum dan tidak berkembang lagi, maka varietas yang
diterserang tersebut sangat tahan. Perbedaan bentuk, warna, dan ukuran dari
bercak digunakan untuk membedakan ketahanan varietas.
2.1.6. Penanda SCAR (Sequence Characterized Amplified Region Marker)
Tiga pasang primer SCAR yaitu Cut1, Pwl2, dan Erg2 telah digunakan. Di
Indonesia untuk menunjukkan keragaman genetik Pyricularia patogen padi dari
beberapa daerah endemik blas (Reflinur dkk. 2005). Enzim kutinase Pyricularia padi
yang disandikan oleh gen cut1 tidak mempengaruhi patogenisitas dan kecepatan
konidiasi (Sweigard et al. 1992), tetapi gen cut1 yang diperlukan untuk menginfeksi
tanaman (Skamnioti dan Gurr 2007). Sebanyak delapan gen diperkirakan
menyandikan enzim kutinase Pyricularia dari padi (Dean et al. 2005). Pwl2
merupakan salah satu gen avirulen Pyricularia 70-15 dari padi (Dean et al. 2005).
Kehadiran gen Pwl2 pada Pyricularia dari padi mencegah cendawan ini
menginfeksi inang alternatifnya, yaitu Eragrostis curvula, dan tetap patogen terhadap
padi dan barley. Keragaman fenotipe Cut1 da n Pwl2 mungkin disebabkan oleh
mutasi, yaitu dengan keberadaan aktivitas semacam elemen transposon. Mutasi dan
rekombinasi merupakan sumber utama bagi cendawan patogen tumbuhan dalam
menghasilkan variasi genetik, seperti halnya pada kebanyakan organisme (Burdon &
Silk 1997). Begitu juga penanda Erg2, hanya satu isolat yang tidak menunjukkan
hasil amplifikasinya. Sampai saat ini belum diperoleh informasi penyebab mutasi dari

21
gen Erg2. Gen yang berhubungan dengan sifat virulensi pada P.oryzae secara
genetik telah dikaji oleh beberapa peneliti (Chao dan Ellingboe, 1997; Valent et al.,
2001). Selain itu, beberapa gen resisten terhadap P.oryzae, gen yang mengontrol
kompatibilitas dengan varietas tertentu, dan gen-gen yang mengontrol
perkembangan blas selama infeksi termasuk gen yang mempengaruhi pembentukan
apresorium dan fungsi penetrasi apresorium juga telah dilaporkan (Chao dan
Ellingboe, 1997; Kang dan Lee, 2000; Lau dan Ellingboe, 1993). Famili gen Pwl
selain Pwl1 (Kang dan Lee, 2000) dan beberapa gen spesifik yang berperan dalam
pengenalan inang P.oryzae, yaitu Cut1, Erg2, dan Pwl2. Gen Cut1 merupakan lokus
yang menjadikan gen cutinase berfungsi sebagai pendegradasi lapisan kutikula
tanaman (Sweigard et al., 1992). Gen Erg2 berperan sebagai penyandi metabolit
sekunder pada cendawan yang menjadi target antifungal pada sel tanaman (Keon et
al., 1994), sedangkan Pwl2 dikenal sebagai gen avirulen yang bersifat spesifik inang
(Valent dan Chumley, 1994). Ketiga gen tersebut banyak digunakan sebagai marka
DNA yang dikenal sebagai sequence characterized amplified region (SCAR)
(Soubabere et al., 2001), termasuk penggunaannya dalam studi keragaman genetik
cendawan P.oryzae (Reflinur et al., 2005).

22
BAB IIIMETODE PENELITIAN
3.1 Tempat dan waktu penelitian
Penelitian ini dilakukan di Laboratorium Penyakit Tumbuhan Fakultas
Pertanian, Universitas Hasanuddin, Uji PCR dilakukan di Laboratorium Penelitian,
Rumah Sakit Pendidikan Universitas Hasanuddin dan pengambilan sampel akan
dilakukan pada padi sawah ditiga Kabupaten yang mewakili dua zona iklim di
Sulawesi Selatan yakni Kab Bone, Kab Maros dan Kab. Gowa. Pelaksanaanya
mulai pada bulan Maret 2017- Oktober 2017.
3.2 Bahan dan Alat
Bahan-bahan yang digunakan dalam penelitian ini yaitu agar-agar, kentang,
gula pasir, rolled oats, aquadesh, NaOCl, spiritus, alkohol, plastik, karet gelang,
kertas saring, alumunium foil, wrapping plastic, dan Chloramphenicol.
Alat yang digunakan adalah autoclave, oven, laminar air flow, inkubator,
shacker, neraca o haus, timbangan analitik, hot plate, erlenmeyer, cawan petri, gelas
ukur, korkbohrer, jarum preparat, kaca preparat, deck glass, haemocytometer, alat
semprot, spatula, gelas pelastik, penyaring, pipet mikro, pembakar bunzen,
mikroskop dan kamera

23
3.3 Metde Pelaksanaan
3.3.1 Pengamatan Intensitas Serangan di Lapangan.
Pengamatan intensitas serangan penyakit blas dilakukan pada padi sawah..
Pengamatan dilakukan berupa blas daun (leaf blast). Metode pengamatan dilakukan
secara diagonal pada setiap lahan ditentukan 5 plot (30 tanaman sampel dalam
satu plot) untuk intensitas penyakit. Intensitas penyakit (IP dalam %). Skala yang
digunakan untuk menilai kategori kerusakan adalah:skala IRRI (1996) (Tabel 1).
3.3.2 Pengambilan sampel
Pengambilan sampel yang bergejala dilakukan di Kab. Bone, Kab. Maros dan
Kab. Gowa dengan menggunakan metode purpose sampling atau pengambilan
sampel yang bergejala penyakit blas. Pengambilan sampel dilakukan pada umur
tanaman berada pada fase vegetatif (daun) dan generatif yaitu leher dan kolar.
Tamanan yang bergejala diambil kemudian dimasukkan pada amplop sampel dan
diberikan label identitas berupa jenis varietas, daerah/lokasi, dan tanggal
pengambilan untuk keperluan penyimpanan. Selanjutnya dibawa ke laboraturium
untuk proses isolasi dan identifikasi.
3.3.3 Isolasi dan identifikasi Cendawan Pyricularia oryzae
a. Pembuatan media1. Pembuatan media Potato Dextrose Agar (PDA)
Media PDA terdiri dari (200 g potongan kentang, 20 g sukrosa, agar 17 g dan
1000 ml aquadest). Pertama kentang di potong- potong kecil benbentuk lalu direbus
dengan aquadest sampai mendidih hingga keluar ekstrak dari kentang. Kemudian

24
ekstrak kentang kemudian disaring dan dimasukkan kedalam Erlenmeyer yang telah
berisi 20 g sukrosa dan agar 17 g . Setelah itu, menambahkan aquadest hingga
mencapai volume 1000 ml pada Erlenmeyer sebagai pengganti volume air yang
hilang selama pemanasan. Larutan tersebut dimasak kembali menggunakan hot
plate sambil diaduk-aduk agar semua bahan larut biarkan sampai mendidih. Setelah
mendidih lalu tutup Erlenmeyer menggunakan aluminium foil dan dieratkan kembali
menggunakan plastic wrapping. Selanjutnya media disterilisasi menggunakan
autoclave pada suhu 121o C. sebelum digunakan tambahkan 250 mg-1
Chloramphenicol untuk meminimal kontaminan bakteri
2. Pembuatan Media Oat meal Agar
Media Oat meal Agar terdiri ( 40 g rolled oat, 5 g sukrosa 16 g agar dan 1000
ml aquadest). Pertama Merebus rolled oat kedalam 1000 ml aquades selama 15
menit sambil diaduk- aduk sampai mendidih kemudian disaring. menambahkan 16 g
agar-agar dan 5 g sukrosa lalu panaskan kembali diatas hotplate untuk melarutkan.
Setelah mendidih tutup dengan kapas yang dilapisi aluminium foil dan autoclave dua
kali selama 40 menit pada 15 psi. Sebelum oatmeal agar siap digunakan.
3. Pembuatan media cair PDB
Media PDB digunakan untuk kultur miselia terdiri dari (200 g potongan
kentang, 20 g sukrosa, dan 1000 ml aquadest). Kentang di potong- potong kecil lalu
direbus dengan aquadest sampai mendidih hingga keluar ekstrak dari kentang.
Kemudian ekstrak kentang disaring dan dimasukkan kedalam Erlenmeyer yang telah
berisi 20 g sukrosa. Setelah itu, ditambahkan aquadest hingga mencapai volume

25
1000 ml. Selanjutnya media diautoclave pada suhu 121oC, sebelum digunakan
ditambahkan 250 mg/L Chloramphenicol untuk menghindari kontaminasi bakteri.
b. Isolasi Cendawan Patogen Pyricularia oryzae
Patogen diisolasi mengikuti prosedur isolasi jaringan standar seperti
dijelaskan dalam Tuite (1969). Jaringan tanaman yang sakit dipotong dari margin
bagian yang terinfeksi dan diiris secara membujur menggunakan gunting steril dan
dicuci dalam air keran selama dua menit di petridish terpisah untuk meminimalkan
kontaminan permukaan. Selanjutnya, sterilisasi permukaan mengikuti metode dari
Aneja (2005) yang telah di modifikasi. Jaringan tanaman yang telah dipotong
kemudian dicelupkan ke dalam etanol 70% selama 1, 5 menit, NaoCl 5% selama 1,5
menit dan dibilas tiga kali dengan aquades steril untuk menghilangkan kontaminan
yang tersisa. Jaringan permukaan yang telah disterilkan kemudian dipindahkan pada
media potato dextrose agar (PDA) dengan menambahkan Chloramphenicol 250
mg/L untuk menekan pertumbuhan bakteri kontaminan. kemudian diinkubasi selama
3-4 hari sampai muncul mesilia dan sporulasi. Untuk membuktikan bentuk spora
cendawan blas,miselium ditumbuhkan kembali pada media oat meal agar (OA)
(30 g oat meal, 20 g agar,dan g sukrosa) diinkubasi selama 7–8 hari pada
suhu ruang.
c. Identifikasi Cendawan Pyricularia oryzae
Identifikasi morfologi cendawan patogen P. oryzae dilakukan berdasarkan
Bonman et al., (1986) dan diamati di bawah mikroskop cahaya dengan berbagai
perbesaran. Cendawan P. oryzae dapat diidentifikasi dengan adanya konidia yang

26
berbentuk mirip gada warna abu-abu dan umumnya memiliki dua septa, namun
kadang-kadang ditemukan juga satu atau tiga septa.
3.4 Uji Ras Isolat Cendawan Pyricularia Oryzae
Penentuan ras penyakit blas sesuai dengan metode dari BB Padi Sukamandi
yang dimodifikasi oleh Prabawa dkk (2014), menggunakan 1 set varietas diferensial
yang terdiri dari 7 varietas yaitu varietas Asahan, Cisokan, IR 64, Krueng Aceh,
Cisadane, Cisanggarung dan Kencana Bali. Metode penentuan ras penyakit blas
dilakukan dengan menanam ketujuh varietas secara berurutan mulai dari varietas
Asahan hingga varietas Kencana Bali pada lokasi penelitian. Tahapan pengujian
diawali dengan persiapan tanah mengikuti metode dari (lestari dkk, 2016). Tanah
dicampur dengan pupuk kandang dengan perbandingan 10 kg : 1 kg.
Persiapan inokulum cendawan P. oryzae mengikuti metode (Listiyowati,
2012) Konidium dipanen sebagai berikut: air steril yang mengandung 0,02% Tween
20 dituang ke permukaan koloni dan permukaan koloni digosok memakai kwas
gambar no. 10. Konidium dikumpulkan dan dibuat suspensi pada konsentrasi 1x106
konidium m/L untuk digunakan sebagai inokulum.
Intensitas serangan blas diamati pada tujuh hari setelah inokulasi
menggunakan standar evaluasi IRRI (1996) yang disajikan pada Tabel 1. Tipe
bercak skala 5-9 digunakan untuk mengidentifikasi tingkat kerentanan inang. Nomor
ras diberikan berdasarkan pola reaksi varietas padi diferensial Indonesia terhadap
hasil inokulasi (Tabel 2). Kemudian dilakukan perhitungan intensitas penyakit
dengan rumus berikut:

27
S= ∑( )( ) %Dimana:
S :keparahan penyakit
n: jumlah daun dengan skor tertentu
v:skor daun yang terserang
N: jumlah daun yang diamati
V: skala skor tertinggi
Tabel 1. Skor Keparahan Penyakit (IRRI,1996)
Skor Kerusakan Daun Klasifikasi0 Tidak ada bercak Sangat tahan
1 Bercak kecil berwarna cokelat sebesar ujung jarum Tahan
2 Bercak nekrotik kecil membulat, abu-abu, sedikit
memanjang, panjang 1-2 mm, tepi cokelat, bercak
banyak ditemukan di bagian bawah daun
Cukup tahan
3 Tipe bercak mirip dengan skor 2, tetapi sejumlah
besar bercak sudah ditemukan pada bagian atas daun
Agak tahan
4 Bercak khas blas (belah ketupat), panjang 3 mm atau
lebih, luas daun terserang kurang dari 2 %
Moderat
5 Bercak khas blas, panjang 3 mm atau lebih, luas daun
terserang 2-10%
Rentan
6 Bercak khas blas, panjang 3 mm atau lebih, panjang 3
mm atau lebih, luas daun terserang 11-25%
Rentan
7 Bercak khas blas, panjang 3 mm atau lebih, luas daun
terserang 26- 50%
Rentan
8 Bercak khas blas, panjang 3 mm atau lebih, luas daun
terserang 51 -75%, beberapa daun mulai mati
Rentan
9 Semua daun mati, luas daun terserang lebih dari 75% Sangat rentan

28
Tabel 2. Contoh pemberian nomor ras Pyricularia asal padi (Mogi et al. 1991)
VarietasDiferensial
Nokode
No Ras
001 003 011 053 153 221
Asahan 200 T T T T T R
Cisokan 100 T T T T R T
IR 64 40 T T T R R T
Krueng Aceh 20 T T T R R R
Cisadane 10 T T R R R T
Cisanggarung 2 T R T R R T
Kencana Bali 1 R R R R R R
R= Rentan, T= Tahan
3.5 Deteksi Gen virulensi isolat Pyricularia oryzae Cav dengan Metode PCR
3.5.1 Isolasi DNA genom
Isolasi DNA genom 10 isolat cendawan P. oryzae dilakukan dari kultur
miselia cendawan. Penyiapan kultur miselia cendawan mengikuti metode Lestari, et
al (2014). Ekstraksi DNA dengan mengikuti prosedur gSYNCTm DNA Extraction Kit
sebanyak 25 mg. Miselia cendawan dipindahkan ke dalam tabung centrifugasi
ukuran 1,5 ml kemudian miselia dicentrifugasi selama 5 menit, ditambahkan 200 μl
Buffer GST dan 20 μl protease K lalu miselia digerus menggunakan Micropestle
kemudian di vortex secara cepat. Selanjutnya diinkubasi semalaman pada suhu 60o
C sampai sampel menjadi lysate. Apabila masih terdapat gumpalan setelah inkubasi,
maka dilakukan centrifugasi selama 2 menit pada 14-16.000x g kemudian
supernatan dipindahkan secara hati-hati pada tabung microcentrifugasi 1,5 ml yang
baru dan tambahkan GSB Buffer kemudian di vortex lalu ditambahkan 200 μl
ethanol absolut pada sampel dan vortex secara sedang selama ± 10 detik sampel

29
kemudian di tuang kedalam GD coloum dalam 2 ml collection tube lalu centrifugasi.
DNA yang tertampung pada GD coloum kemudian ditambahkan 400 μl w1 Buffer
lalu di centrifugasi selama 30 detik hingga ditemukan DNA yang Murni
3.5.2 Amplifikasi PCR
Uji genotipe cendawan P.oryzae dilakukan menggunakan marka berbasis gen
untuk sifat vilurensi yaitu markah Erg2, Pwl2 dan Cut1, yaitu markah yang
mempunyai hubungan virulensi (Soubabere et al. 2001). Identifikasi dilakukan
dengan PCR sepasang primer spesifik untuk gen Erg2 (1.440 bp), Pwl2 (900 bp),
dan Cut1 (1.730bp). Reaksi PCR menggunakan Enzim Go taq® Green Master mix
dimana terdapat 2x green Go Taq® Buffer reaksi (pH 8.5) 400 μl dATP, dGTP, 400
μM dTTP dan 3mM Mgcl2, H2O dan 3 μl template DNA. Urutan basa dari primer-
primer tersebut adalah:
a) Cut1 (F:5’-TATAGCGTTGACCTTGTGGA3’;
R:5’TAAGCATCTCAGACCGAACC-3’)
b) Erg2 (F:5’-GCAGGGCTCATTCTTTTCT3’;
R:5’CCGACTGGAAGGTTTCTTTA-3’)
c) Pwl2 (F:5’-TCCGCCACTTTTCTCATTCC-3’;
R:5’- GCCCTCTTCTCGCTGTTCAC-3’)
Program PCR diawali dengan denaturasi awal pada 94°C selama 2
menit, proses amplifikasi sebanyak 35 siklus, yakni 94°C selama 30 detik,
54°C selama 30 detik, 72°C selama 1 menit, dan siklus terakhir pada pada 72°C
selama 5 menit. Elektroforesis produk PCR dilakukan pada gel agarosa 1% (w/v)
selama 40 menit untuk mengidentifikasi amplikon tiap primer sesuai ukuran
referensi (Soubabere et al.,2001; Reflinur dkk., 2005).

30
3.6. Analasis Data
Pita DNA hasil amplifikasi yang muncul untuk setiap primer dari masing-
masing isolat diskor dengan nilai 1 (ada) dan 0 (tidak ada). Selanjutnya data
ini digunakan untuk mengelompokkan isolat-isolat uji menjadi beberap haplotipe,
berdasarkan kombinasi ketiga jenis gen (Pwl2, Erg2, dan Cut1) pada isolat-
isolat yang diuji.

31
BAB IVHASIL DAN PEMBAHASAN
4.1. Hasil
4.1.1. Pengamatan Intensitas Serangan Blas Daun
Intesitas serangan blas daun dilapangan menunjukkan bahwa tingkat
serangan tertinggi pada varietas Ciliwung dengan persentase 60,66% dan 53.36%
pada dua lokasi penelitian, diikuti varietas Cigeulis dengan persentase 30.53% dan
terendah adalah varietas Ciherang 23.6%.
Tabel 3. Intensitas serangan blas daun pada dua lokasi pengambilan sampel
Lokasi VarietasIntensitas
Serangan (%)Kategori*
Bone Ciherang
Inpari 4
Ciliwung
29.95
28.34
53.36
Berat
Berat
Berat
Maros Cigeulis
Ciliwung
Ciherang
30.53
60.66
23.6
Berat
Berat
Sedang
*Kategori berdasarkan Akhasani dan Palupi (2015)
4.1.2. Isolasi dan Karakteristik Morfologi cendawan P. oryzae
Hasil isolasi cendawan patogen Pyricularia oryzae ditemukan 20 isolat yang
berasal dari 3 kabupaten yakni Kab. Bone 8 isolat yang masing diberi kode isolat
berturut PoBn1, PoBn2, PoBn3, PoBn4, PoBn5, PoBn6, PoBn7, dan PoBn8 Kab.
Maros dengan 7 Isolat dengan kode isolat berturut PoMr1, PoMr2, PoMr3, PoMr4,
PoMr5, PoMr6, PoMr7 dan PoMr8 sedangkan Kab. Gowa sebanyak 4 isolat dengan
kode isolat berturut-turut PoGw1, PoGw2, PoGw3, dan PoGw4.

32
Tabel 4. Jumlah Isolat Yang Ditemukan Pada Daerah Pengambilan Sampel
No. Kode Isolat Lokasi Pengambilan Sampel Asal Isolat
1. PoBn1 Bengo, Kab. Bone Daun
2. PoBn2 Bengo, Kab. Bone Daun
3. PoBn3 Bengo, Kab. Bone Daun
4. PoBn4 Lappariaja, Kab. Bone Daun
5. PoBn5 Lappariaja, Kab. Bone Daun
6. PoBn6 Bengo,Kab. Bone Daun
7. PoBn7 Bengo,Kab. Bone Daun
8. PoBn8 Bengo, Kab. Bone Daun
9. PoMr1 Balai Proteksi, Kab. Maros Daun
10. PoMr2 Balai Proteksi, Kab. Maros Daun
11. PoMr3 Kab. Maros Daun
12. PoMr4 Balai Proteksi, Kab. Maros Daun
13. PoMr5 Bantimurung, Kab. Maros Daun
14. PoMr6 Bantimurung, Kab. Maros Daun
15. PoMr7 Simbang, Kab. Maros Daun
16. PoMr8 Simbang, Kab. Maros Daun
17. PoGw1 Patalassang, Kab. Gowa Daun
18. PoGw2 Bontonompo, Kab. Gowa Leher
19. PoGw3 Patalassang, Kab. Gowa Leher
20. PoGw4 Moncong- Moncong, Kab. Gowa Kolar
Morfologi isolat cendawan yang di temukan pada media PDA, koloninya
berwarna abu-abu sampai abu-abu kehitaman dengan tekstur koloni beludru
(Gambar 4). Namun ada juga yang tidak teratur (Gambar 4A). Pada umumnya
miselia cendawan Po mempunyai bentuk melingkar seperti cincin konsentris yang
mengarah ke pusat (Gambar 4C).
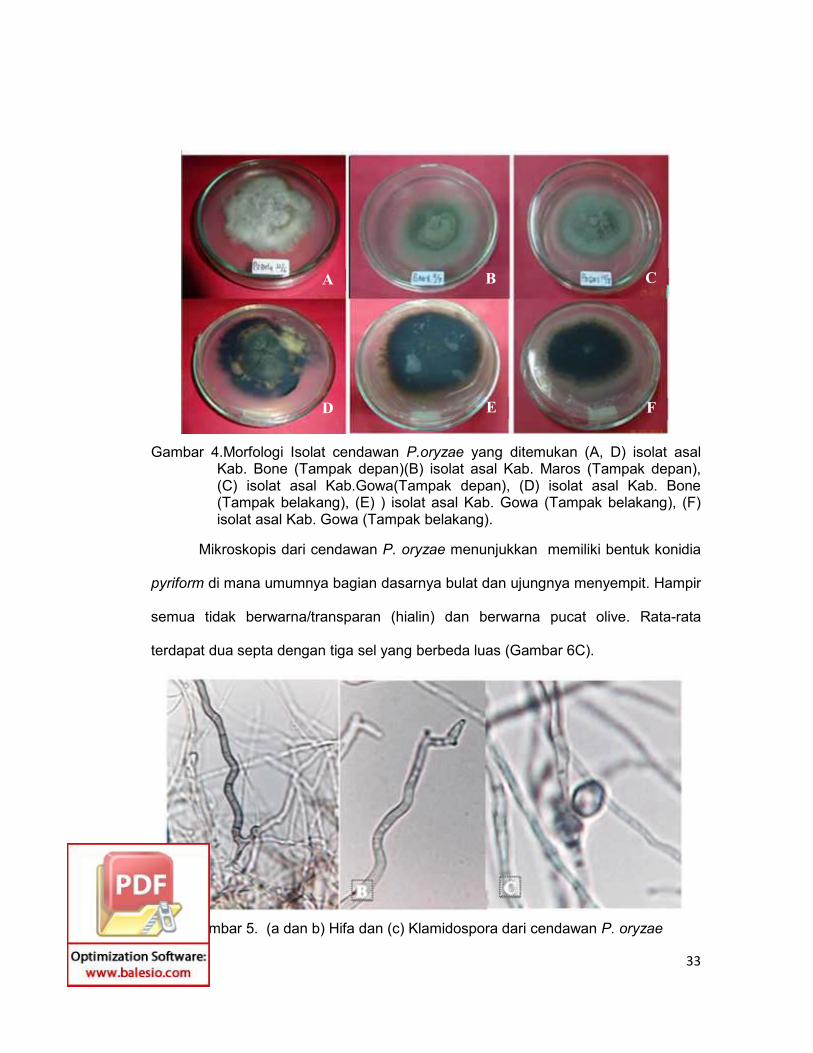
33
Gambar 4.Morfologi Isolat cendawan P.oryzae yang ditemukan (A, D) isolat asalKab. Bone (Tampak depan)(B) isolat asal Kab. Maros (Tampak depan),(C) isolat asal Kab.Gowa(Tampak depan), (D) isolat asal Kab. Bone(Tampak belakang), (E) ) isolat asal Kab. Gowa (Tampak belakang), (F)isolat asal Kab. Gowa (Tampak belakang).
Mikroskopis dari cendawan P. oryzae menunjukkan memiliki bentuk konidia
pyriform di mana umumnya bagian dasarnya bulat dan ujungnya menyempit. Hampir
semua tidak berwarna/transparan (hialin) dan berwarna pucat olive. Rata-rata
terdapat dua septa dengan tiga sel yang berbeda luas (Gambar 6C).
Gambar 5. (a dan b) Hifa dan (c) Klamidospora dari cendawan P. oryzae
A B C
D E F

34
Gambar 6. (A) Pembentukan Konidia, (B) Konidia yang masih menempeldikonidiophore dengan satu septa, (C) Konidia dengan dua septadengan tiga sel pembesaran 40x menggunakan mikroskop adapter .
4.1.3. Uji Ras isolat-isolat P. oryzae
Gejala serangan pada varietas yang tahan terhadap blas bercak yang
muncul berupa titik kecil berwarna coklat tua dan ukuran bercak tidak berkembang
(Gambar 7 B dan C) . Pada varietas yang rentan, tampak bercak berwarna abuabu
keputih-putihan berbentuk belah ketupat dengan kedua ujung bercak meruncing
(Gambar 7E) Ukuran bercak pada varietas rentan (R) akan berkembang dan
menyatu hingga seluruh bagian daun sedangkan pada varietas sangat rentan (SR)
dalam kondisi yang lembab, daun kemudian akan membusuk dan akhirnya tanaman
mati.

35
Gambar 7. Gejala serangan penyakit blas pada tanaman padi differensial setelah
inokulasi
Penentuan ras dari cendawan P. oryzae berdasarkan hasil dari pengamatan
intesitas serangan pada tujuh varietas diferensial ditemukan total 12 ras cendawan
P.oryzae. Kecamatan Bengo dan Kecamatan Lappariaja dijadikan lokasi
pengambilan sempel untuk mewakili Kabupaten Bone, Kecamatan Bantimurung,
Simbang, dan Maros Baru Untuk mewakili Kabupaten Maros sedangkan Kabupaten
Gowa diwakili oleh Kecamatan Patallasang, Kecamatan Bontonompo, dan
Moncong-moncong. Total 5 ras yang berbeda ditemukan pada kabupaten Bone,
Kabupaten Maros sebanyak 6 ras yang berbeda dan kabupaten Gowa ditemukan 3
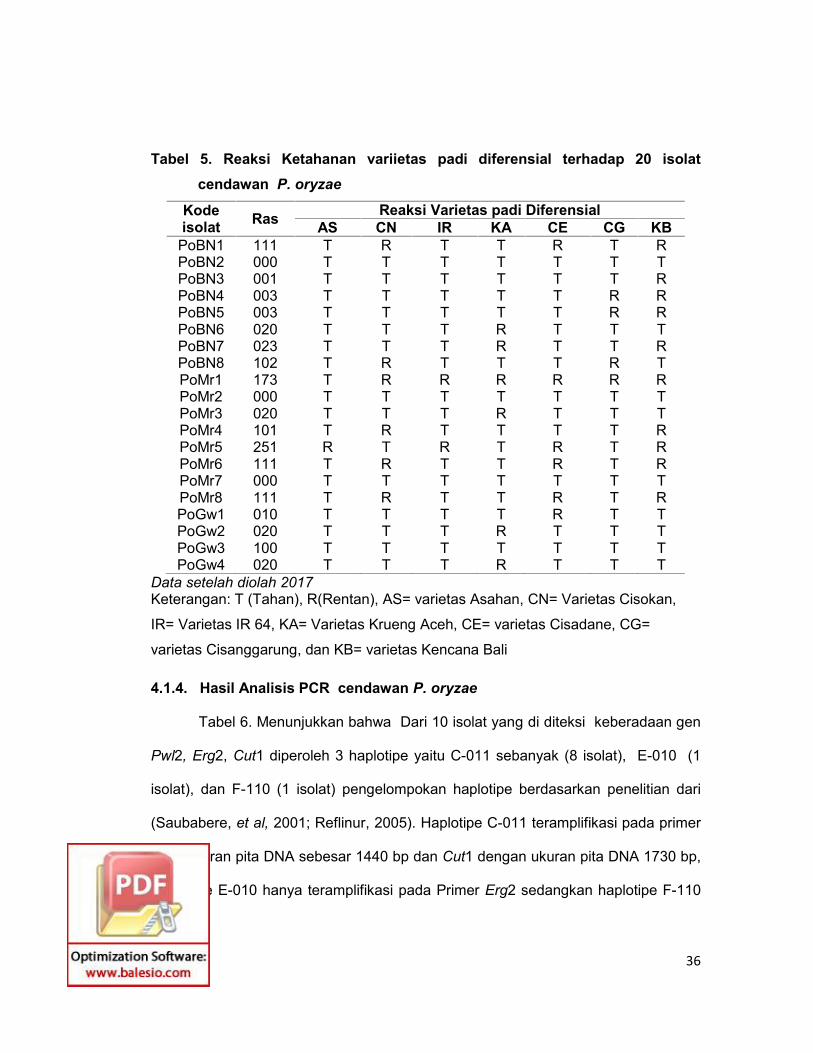
ras yang berbeda. Dari hasil pengujian ras (Tabel 5) menunjukkan bahwa ras
020,023 ditemukan pada semua lokasi,sedang ras 001, 100,102,111, 173 dan 251
hanya di temukan pada satu lokasi pengambilan sampel.
36
Tabel 5. Reaksi Ketahanan variietas padi diferensial terhadap 20 isolatcendawan P. oryzae
Kodeisolat Ras Reaksi Varietas padi Diferensial
AS CN IR KA CE CG KBPoBN1 111 T R T T R T RPoBN2 000 T T T T T T TPoBN3 001 T T T T T T RPoBN4 003 T T T T T R RPoBN5 003 T T T T T R RPoBN6 020 T T T R T T TPoBN7 023 T T T R T T RPoBN8 102 T R T T T R TPoMr1 173 T R R R R R RPoMr2 000 T T T T T T TPoMr3 020 T T T R T T TPoMr4 101 T R T T T T RPoMr5 251 R T R T R T RPoMr6 111 T R T T R T RPoMr7 000 T T T T T T TPoMr8 111 T R T T R T RPoGw1 010 T T T T R T TPoGw2 020 T T T R T T TPoGw3 100 T T T T T T TPoGw4 020 T T T R T T T
Data setelah diolah 2017Keterangan: T (Tahan), R(Rentan), AS= varietas Asahan, CN= Varietas Cisokan,
IR= Varietas IR 64, KA= Varietas Krueng Aceh, CE= varietas Cisadane, CG=
varietas Cisanggarung, dan KB= varietas Kencana Bali
4.1.4. Hasil Analisis PCR cendawan P. oryzae
Tabel 6. Menunjukkan bahwa Dari 10 isolat yang di diteksi keberadaan gen
Pwl2, Erg2, Cut1 diperoleh 3 haplotipe yaitu C-011 sebanyak (8 isolat), E-010 (1
isolat), dan F-110 (1 isolat) pengelompokan haplotipe berdasarkan penelitian dari
(Saubabere, et al, 2001; Reflinur, 2005). Haplotipe C-011 teramplifikasi pada primer
Erg2 ukuran pita DNA sebesar 1440 bp dan Cut1 dengan ukuran pita DNA 1730 bp,
Haplotipe E-010 hanya teramplifikasi pada Primer Erg2 sedangkan haplotipe F-110

37
teramplifikasi pada dua primer yaitu Pwl2 dan Erg2 dengan masing-masing ukuran
pita DNA 900bp dan 1440 bp.
Tabel 6. Pengelompokan haplotip cendawan Pyricularia oryzae berdasarkankeberadaan tiga gen virulensi
Kode isolatKeberadaan amplikon
HaplotipePwl21 Erg22 Cut13
PoBn1 0a 1 1 C-011
PoBn2 0 1b 1 C-011
PoBn3 0 1 1 C-011
PoGw1 0 1 1 C-011
PoGw3 0 1 1 C-011
PoMr2 0 1 1 C-011
PoMr4 0 1 1 C-011
PoMr6 0 1 0 C-011
PoMr7 1 1 0 F-110
PoMr8 0 1 0 E-010
Keterangan: 1= 900 bp, 2= 1440 bp, 3 =1730 bp, a= tidak ada pita DNA, b=ada pita
DNA
Hasil dari elektroforesis berdasarkan primer penyandi gen spesifik
menunjukkan bahwa keberadaan ketiga primer virulensi Cut1, Erg2, dan Pwl2
tersebut tidak teramplifikasi pada semua isolat yang dikarakterisasi. Variasi
amplikon pada isolat cendawan yang diamati beserta ukuran gen virulensi tercantum
pada Gambar 8- 10.
Produk amplifikasi menggunakan primer Cut1 memiliki ukuran pita sebesar
1730 bp, Primer Erg2 menghasilkan pita sebesar 1440 bp dan dengan primer Pwl2
menghasilkan pita sebesar 900 bp. Tidak ada isolat yang menghasilkan produk
amplifikasi dengan 3 primer. Hanya isolat PoMr7 yang memiliki produk sebesar 900
bp dengan primer Pwl2 sedangkan 9 isolat isolat lainnya tidak teramplifikasi.

38
Gambar 8 . Pola pita hasil amplifikasi DNA genomik cendawan P. oryzae dengangen Pwl2. Sumur paling kiri adalah DNA ladder 1 kb.
Gambar 9. Pola pita hasil amplifikasi DNA genomik cendawan P. oryzae dengangen Erg2. Sumur paling kiri adalah DNA ladder 1 kb.
Gambar 10. Pola pita hasil amplifikasi DNA genomik cendawan P. oryzae dengangen Cut1. Sumur paling kiri adalah DNA ladder 1 kb.

39
4.2. PEMBAHASAN
Tabel 3. Rerata intensitas serangan blas di lapangan menunjukkan bahwa
intensitas tertinggi pada varietas Ciliwung di dua lokasi memiliki persentase
serangan tertinggi bila dibandingkan dengan varietas Ciherang. Hal ini dikarenakan
kondisi lingkungan di area pengambilan sampel terdapat beberapa varietas padi
yang yang ditanam oleh petani sehingga kemungkinan penyebaran patogen terjadi
dari varietas lain yang terlebih dahulu terserang penyakit blas varietas yang diamati
melalui bantuan angin. Hal ini sesuai dengan peneltian Hasti (2016), menyatakan
bahwa beberapa varietas dapat menjadi pemicu munculnya penyakit blas dimana
vaietas tersebut sudah terlebih dahulu sehingga mengakibatkan varietas lain
disekitarnya terserang penyakit.
Selain pengaruh dari varietas, faktor lain yang dapat mempengaruhi
intensitas serangan penyakit blas adalah inang alternatifnya. Santoso dan Nasution
(2008), menyatakan bahwa patogen blas selain menginfeksi tanaman padi juga
dapat merusak serealia lain seperti gandum, sorgum, dan lebih dari 40 spesies
graminae Gulma yang banyak terdapat di lahan sawah yaitu Echinochloa colona dan
Digitaria cilaris adalah inang P. oryzae yang berperan sebagai sumber spora pada
periode bera. Miselia patogen tersebut juga dapat bertahan selama setahun pada
jerami sisa panen (Teng et al. 1991). Suwandi dkk. (2016), melaporkan sisa-sisa
jerami tanaman sakit di lapangan dapat menjadi sumber inokulum bagi tanaman
musim berikutnya.

40
Berdasarkan deskripsi varietas padi tahun 2009, varietas Ciliwung hanya
tahan terhadap penyakit yang disebabkan oleh bakteri, begutu pun dengan varietas
Inpari, varietas Cegeulis dan varietas Ciherang. Hal ini tidak sesuai dengan keadaan
dilapangan dimana penyakit blas yang disebabkan oleh cendawan P. oryzae dapat
menyerang keempat varietas tersebut. Adapun Intensitas serangan varietas-
varietas pada dua lokasi pengamatan dimana pada setiap lokasi berbeda
persentase tingkat serangannya. Penggunaan varietas semula tahan terhadap
penyakit blas hanya dapat bertahan selama beberapa musim saja keadaan ini terjadi
adanya proses adaptasi, mutasi genetik dan patogen dapat membentuk ras-ras baru
yang lebih virulen. Sesuai pendapat (Purnomo, 2002), Patogen P. oryzae sangat
mudah untuk membentuk ras baru yang lebih virulen dan dominan ras patogen
sangat mempengaruhi ketahanan varietas. Penggunaan varietas-varietas semula
tahan akan menjadi rentan setelah ditanam dibeberapa musim tanam dan varietas
yang tahan pada satu tempat rentan ditempat lain artinya bahwa penggunaan
varietas tahan sangat dibatasi oleh waktu dan tempat (Santoso dan Nasution,
2009). Menurut Yuliani dan Maryana (2014), melaporkan bahwa perbedaan dan
perubahan reaksi suatu varietas terhadap blas disebabkan oleh adanya perbedaan
dan perubahan ras antar lokasi dan ras yang dominan di suatu wilayah.
Pengamatan intensitas serangan pada varrietas Ciherang pada dua lokasi
pengamatan persentase serangan antara 23.6 % sampai 28.34%. jika dibandingkan
dengan intensitas serangan pada varietas-varietas yang diamati. Persentase
serangannya pada var. Ciherang tergolong rendah artinya varietas tersebut tahan
terhadap penyakit blas daun. hal ini sesuai peneltian Akhsani dan Palupi (2015),

41
dimana var. Ciherang merupakan varietas yang tahan terhadap penyakit blas daun.
Suganda dkk (2016), juga melaporkan bahwa intensitas serangan blas pada var.
Ciherang di lokasi endemik serangannya lebih rendah pada blas daun tetapi
serangannya lebih tinggi pada blas leher. Sehingga dapat dikatakan bahwa
intensitas serangan blas bergantung dari ketahanan varietas.
Karakter morfologi dari berbagai jenis isolat P. oryzae menghasilkan variasi
berkenaan dengan warna koloni yang bervariasi dari warna abu-abu sampai hitam
dan menghasilkan koloni margin yang halus dan kasar. Namun, tidak ada variasi
yang berhubungan dengan konidia. Dimana konidia pyriform selalu hialin sampai
pucat olive, 2-septa dengan 3 ruang. Karakter ini adalah kesepakatan seperti yang
dijelaskan oleh( Shirai, 1896); (Srivastava et al, 2014). Kariaga et al (2016),
melaporkan bahwa miselium dari cendawan P.oryzae bercabang dan hialin.
Konidiofor bertingkat sederhana jarang bercabang yang cukup panjang dan septasi,
berwarna coklat muda dan sedikit menebal di dasar, dentikel juga ditemukan di
ujungnya. Konidia terpasang secara sympodial di ujung konidiofor umumnya
pyriform sampai obclavate.
Cendawan Pyricularia oryzae yang ditemukan memiliki warna koloni yang
cepat berubah dari gelap menjadi putih selama disubkultur berulang-ulang di
laboratorium. Hal ini sesuai dengan pendapat Valent and Chumley (1991), yang
menyatakan bahwa isolat-isolat cendawan blas padi cenderung tidak stabil
penampakan koloninya, fertilitas, dan patogenisitasnya selama disubkultur berulang-
ulang di laboratorium

42
Pengujian ras (fenotipe) telah banyak dilakukan di berbagai belahan dunia.
Di Indonesia, pengujian secara fenotipe umumnya dilakukan dengan pemantauan
perkembangan populasi patogen blas di lapangan, yang didasarkan pada reaksi
virulensi terhadap satu set varietas diferensial Indonesia (Mogi et al. 1991).
Berdasarkan hasil pengujian ras berdasarkan satu set varietas diferensial Indonesia
ditemukan 12 ras dari 20 isolat yang diuji. Banyaknya macam ras dalam 1 lokasi
bekisar 2-4 macam. Ras 020 merupakan ras yang paling dominan dengan 4 isolat,
urutan kedua adalah ras 111(3 isolat) terdapat pada dua lokasi yakni Bone dan
Maros. Ras 020 hanya dapat menginfeksi varietas Krueng Aceh dimana nilai
penentu rasnya adalah 20. Adapun ras 111 hanya menginfeksi varietas Cisokan,
Ciadane, dan Kencana Bali (Lestari dkk., 2016). Setiap individu ras yang mempunyai
sifat virulensi yang berbeda (Zeigler et al. 1994). Dominasi ras P. oryzae di suatu
wilayah dengan wilayah lain yang berbeda memungkinkan varietas padi di suatu
wilayah tahan tetapi rentan di wilayah lain. Informasi sebaran ras P. oryzae sangat
diperlukan untuk memprediksi kesesuaian varietas yang akan dilepas (spesifik
lokasi).
Varietas Asahan merupakan varietas dengan ketahanan paling tinggi yakni
tahan (T) terhadap 19 isolat dari 20 isolat yang diuji, varietas IR 64 memiliki reaksi
Tahan terhadap 18 isolat, varietas Cisanggarung bereaksi tahan terhadapat 16
isolat, varietas Krueng Aceh bereaksi tahan terhadap 14 isolat , sedangkan varietas
Cisokan dan Cisadane bereaksi tahan terhadap 13 isolat yang diuji. Varietas
Kencana Bali memliki spectrum ketahanan yang paling rendah yakni hanya bereaksi
tahan terhadap 9 isolat yang diuji hal ini sesuai dengan dengan penelitian dari

43
Lestari dkk (2016), bahwa padi IR 64 dan Asahan merupakan varietas dengan
spektrum ketahanan yang paling besar dan Varietas Kencana Bali merupakan
varietas dengan tingkat ketahanan paling rendah dengan reaksi rentan (R) sampai
sangat rentan (SR). Kencana Bali selalu digunakan sebagai tanaman cek rentan
dalam pengujian blas di Indonesia (Mogi et al. 1991).
Padi Varietas Kencana bali merupakan salah satu anggota padi diferensial di
Indonesia yang digunakan untuk uji patotipe (ras fisiologi). Padi tersebut rentan
terhadap semua (27) ras Pyricularia asal padi yang ada di Indonesia. Sedangkan
padi Cisokan moderat resisten, yaitu resisten terhadap delapan ras atau dengan
kata lain rentan terhadap serangan 21 ras Pyricularia asal padi yang ada di
Indonesia (DBPT Deptan 1992). Padi IR64 dapat dikategorikan moderat resisten
yang setara dengan padi Cisokan berdasarkan tingkatan resistensi padi diferensial
Indonesia yang digunakan untuk menentukan ras fisiologi Pyricularia asal padi (Mogi
et al. 1991).
Pada penelitian ini ditemukan juga ras 000 asal isolat Kabupaten Maros dan
Kabupaten Bone. Isolat dengan ras 000 tidak mampu menginfeksi ketujuh varietas
padi diferensial. Ras 000 juga ditemukan dalam penelitan Lestari dkk (2016) dan
Listiyowati (2012), tidak virulennya isolat yang diuji kemungkin adanya mutasi
(perubahan fisiologis selama penyimpanan), atau karena faktor lain yang belum
dapat ditentukan. Patogenisitas isolat-isolat cendawan blas padi cenderung tidak
stabil selama disubkultur berulang-ulang di Laboratorium (Valent & Chumley., 1991).

44
Hasil reaksi ketahan terhadap 7 varietas padi Diferensial Indonesia
teridentifikasi ras 001, 003, 023 dan 102 spesifik hanya ditemukan di kabupaten
Bone, sedangkan di kab. Maros ditemukan ras spesifik yaitu ras 173, 101, dan 251.
Beragamnya ras yang ditemukan di dua lokasi karena banyaknya isolat yang diuji
hal ini sesuai dengan pendapat Lestari dkk (2016), yang menyatakan bahwa
banyaknya sampel dan beragamnya varietas/galur pada lokasi pengambilan sampel
sehingga mempengaruhi keragaman ras tersebut.
Ras 001 dalam pengujian ini hanya diteridentifikasi di kabupata Bone. Ras
001 juga teridentifikasi dalam penelitian Amir dkk (2000) dan pada penelitan Lestari
dkk. (2016), dimana ras 001 mempunyai tingkat patogenitas yang paling rendah
virulensinya karena hanya dapat menginfeksi satu dari tujuh varietas diferensial
indonel yaitu varietas Kencana Bali, namun penyebaran ras tersebut sangat luas
dan mampu bertahan lama di lapangan (Sari 2008; Utami 2000). Ras 173 dalam
pengujian ras juga ditemukan pada 1 isolat asal Kab. Maros. Ras fisiologi 173
merupakan ras yang patogen pada semua padi diferensial Indonesia, kecuali
Asahan (DBPT Deptan 1992). Ras 173 merupakan ras yang selalu ada pada seruluh
lokasi endemik penyakit blas dan bersifat virulensi yang tinggi (Utami dkk ., 2000).
Amir dkk. (2000), melaporkan bahwa ras 173 adalah ras yang dapat menyerang
varietas IR 64.
Total 12 ras yang ditemukan di tiga kabupaten di Sulawesi Selatan yaitu ras
000, 001, 003, 010, 020,023, 100,101, 102, 111, 173 dan 251. Ras 001, 003 dan
173 merupakan ras yang selalu ada pada setiap musim tanam (Amir dkk., 2000).
Dimana ras ini selalu muncul pada setiap musim tanam di Lampung antara tahun

45
2000 hingga 2004. Ras 001,100, 101 juga teridentifikasi dibeberapa lokasi di
Indonesia seperti Sumatera (Kayu Agung, Lampung), Kalimantan Tengah
(Dadahup), Bali (Tabanan) dan Jawa Barat (Kuningan) berdasarkan hasil monitoring
tahun 2007 hingga 2008 (Santoso dan Nasution, 2009).
Hasil dari elektroforesis dari 10 isolat cendawan P. oryzae dari tiga daerah
asal Kab. Bone, Kab. Gowa dan Kab. Maros ditemukan tiga haplotipe yaitu
haplotipe C-011 (8 isolat), E-010 (1 isolat) dan F-110 (1 isolat). Haplotipe C-011
merupakan haplotipe yang paling banyak ditemukan pada semua Isolat asal Bone,
Gowa dan Maros sedangkan haplotip F-110 dan E-010 ditemukan paling sedikit dan
hanya ditemukan pada isolat asal Kab. Maros hal ini sesuai penelitian Reflinur
(2005). Adanya perbedaan haplotipe pada isolat antar satu lokasi yakni Kab. Maros
menunjukkan adanya keragaman genetik. Keragaman genetik pada lokasi asal Kab.
Maros dipengaruhi oleh pengaruh lingkungan seperti suhu dan kelembaban. Hal ini
seperti yang dilaporkan oleh Chen et al (1995), melaporkan bahwa keragaman
pada sidik jari DNA genom cendawan P. oryzae dipengaruhi oleh agroekologi pada
lokasi baik pada lokasi yang sama maupun berbeda.
Gen Erg2 dengan ukuran fragmen 1440 bp teramplifikasi pada semua isolat
yang dikarakterisasi, gen Cut1 ukuran pragmen 1730 bp teramplifikasi pada 8 isolat
yang dikarakterisasi. Gen Erg2 berperan sebagai penyandi metabolit sekunder pada
cendawan menjadi target antifungal pada sel tanaman (Keon et al., 1994)
sedangkan Gen Cut1 merupakan lokus menjadikan gen cutinase berfungsi dalam
mendegradasi lapisan kutikula tanaman (Sweigard et al., 1992). Gen Pwl2 hanya

46
teramplifikasi pada satu isolat yakni PoMr7. Pwl2 dikenal sebagai gen avirulen yang
bersifat spesifik inang (Valent dan Chumley, 1994).
Isolat yang tidak teramplifikasi pada Cut1 dan Pwl2 mungkin disebabkan oleh
mutasi, yaitu dengan keberadaan aktivitas semacam elemen transposon. Mutasi dan
rekombinasi merupakan sumber utama bagi cendawan patogen tumbuhan dalam
menghasilkan variasi genetik, seperti halnya pada kebanyakan organisme (Burdon
dan Silk 1997). Dalam penelitian Lestari dkk (2014), melaporkan bahwa isolat yang
mengandung satu atau dua gen virulen tidak stabil seperti F-110 kemungkinan
sering mengalami mutasi spontan yang berpengaruh terhadap virulensinya cukup
tinggi.
Dari hasil analisis molekuler tidak ditemukan perbedaan genetik pada isolat
asal Kab. Bone dan Kab. Gowa. Dimana hanya ditemukan haplotip C-011. Hal ini
sesuai dengan penelitian dari Chuwa et al (2014), melaporkan bahwa dari delapan
isolat dengan rasnya yang berbeda tidak ditemukan adanya keragaman dalam hal
genetiknya.

47
BAB VKESIMPULAN DAN SARAN
5.1. Kesimpulan
Hasil isolasi cendawan dari tanaman padi di Kabupaten.Bone, Maros dan
Gowa ditemukan 20 isolat yang karakter morfologi dan mikroskopisnya identik
dengan P. oryzae. Berdasarkan variasi reaksi varietas tanaman padi differensial,
diperoleh 12 ras patogen Pyricularia oryzae pada tiga lokasi pengambilan sampel.
Ras 020 merupakan ras yang paling dominan dan ditemukan pada tiga lokasi.
Deteksi secara molekuler menggunakan marka terkait gen virulensi terhadap 10
isolat yang diuji, ditemukan adanya 3 haplotype yaitu C-011, F-110 dan E-010,
namun haplotype C-011 yang paling dominan.
5.2. Saran
Perlu dilakukan penelitian lebih lanjut mengenai ras-ras patogen P. oryzae
dengan menggunakan jumlah isolat yang lebih besar dari daerah lainnya, sehingga
data tentang penyebaran ras patogen dapat diketahui lebih mendetail.

48
DAFTAR PUSTAKA
Agrios GN. 1996. Ilmu penyakit tumbuhan. Edisi Ketiga. Busnia M, dan Martoredjo T(penerjemah). Gadjah Mada University Press. Yogyakarta.
Akhsani,.N, dan Palupi,.P.J.2015. Pengaruh Waktu Terhadap Intensitas PenyaktiBlas dan Keberadaan Spora Pyricularia grisea (Cooke) Sacc Pada LahanPadi Sawah (Oryza sativa) Di Kecamatan Samarinda Utara. Jurnal ZIRAA’AHvol 40(2):p114-122
Amir M. dan M.K. Kardin, 1991. Pengendalian penyakit jamur. Buku 3. hlm. 825-844.Dalam Edi Sunaryo, Djoko S. Damardjati, dan Mahyudin Syam (Ed.). Padi.Badan Penelitian dan Pengembangan Pertanian. Pusat Penelitian danPengembangan Tanaman Pangan, Bogor.
Amir M, Nasution A, dan Santoso. 2000. Inventarisasi ras Pyricularia oryzae didaerah Sukabumi Jawa Barat musim tanam 1995-1998. Prosiding NasionalXV dan Seminar Ilmiah PFI 16-18 September 1999. UNSOED. Purwokerto.
Amir M, dan Nasution A. 1993. Status dan pengendalian blas di Indonesia. ProsidingSimposium Penelitian Tanaman Pangan III. Pusat Penelitian danPengembangan Tanaman pangan. Badan Penelitian dan PengembanganPertanian. Bogor 23-25 Agustus 199.
Annamalai, P., H. Ishii, D. Lalithakumari, and R. Revathi. 1995. Polymerase chainreaction and its application in fungal diseases diagnosis. J. PlantDis.Prot.102:91 104.
Barnett, H.L. dan B.H. Barry. 1998. Illustrated genera of imperfect fungi. APS Press.Minnesota, America
Bonman JM. 1992. Durable resistance to rice blast disease-environmentalinfluences. Euphytica 63: 115-123.
Bourett TM and Howard RJ. 1990. In vitro development of penetration structure inthe riceblast fungus Magnaporthe grisea. Can. J. Bot. 68: 329–342.
Burdon JJ, Silk J. 1997. Sources and patterns of diversity in plant-pathogenic fungi.Phytopathology 87:664-669
Chada, S. and T. Gopalkrishnan.2005. Genetic diversity of Indian isolat of rice blastpathogen (M. grisea) using moleculer markers. Curr. Sci.88: 1466-1469.
Chen D. 1993. Population structure of Pyricularia grisea (Cooke) Sacc. In twoscreeningsite and quantitative characterization of major and minor resistancegenes. A thesis doctor of philosophy. University of the Philippines at LosBanos. p.161

49
Chen,. D., R.,S.,Zeigler, H. Leung, and R.J. Nelson. 1995. Population Structure ofPyricularia grisea at two screening Sites in the Philippines. Phytophatology85: 1011-1020.
Chumley FG, and Valent B. 1990. Genetic analysis of melanin deficient, nonpathogenic mutants of Magnaporthe grisea. Mol. Plant-Microbe Interact.3:135-369
Couch BC and Kohn LM. 2002. A multilocus gene genealogy concordant with hostpreference indicates segregation of a new species, Magnaporthe oryzae,from M. grisea. Mycologia, 94, 683-693.
Chuwa.,J.,C, Mabagala,.R.,B, Reuben., Mnyuku. S.O.W. Phatogenic Variation andMolecular Characterization of Pyricularia oryzae, Causal Agent of Rice BlastDisease in Tanzania. IJSR: 2319-7064
[DBPT Deptan] Direktorat Bina Perlindungan Tanaman, Departemen Pertanian.1992. Penyakit Padi [Laporan Akhir]. Jakarta: Kerjasama Teknis Indonesia-Jepang Bidang Perlindungan Tanaman Pangan (ATA-162), BPT Deptan.
Dean RA,Talbot NJ, Ebbole DJ, Farman ML, Mitchell TK, Orbach MJ, Thon M,Kulkarni R, Xu JR, Pan H, Read ND, Lee YH, Carbone I, Brown D, Oh YY,Donofrio N, Jeong JS, Soanes DM, Djonovic S, Kolomiets E, Rehmeyer C, LiW, Harding M, Kim S, Lebrun MH, Bohnert H, Coughlan S, Butler J, Calvo S,Ma LJ, Nicol R, Purcell S, Nusbaum C, Galagan JE, Birren BW. 2005 Apr 21.The genome sequence of the rice blast fungus . Nature. 434(7036):980-986.
Fukuta, Y., Xu, D., Kobayashi, N., Jeanie, M., Yanoria, T., Hairmansis, A., and N.Hayashi. 2009. Genetic characterization of universal differential varieties forblast resistance developed under the IRRI-Japan Collaborative ResearchProject unsing DNA markers in rice (Oryza sativa L.). p.35-68. In YoshimichiFukuta, Casiana M. Vera Crus and N. Kabayashi (Ed.). Development andCharacterization of Blast Resistance Using Differential Varieties in Rice.JIRCAS Working report No. 63. Tsukuba, Japan.
[FRACT] Fungicide Resistance Action Committee. 2008. List of plant pathogenicorganisms resistant to disease control agents. [terhubung berkala].http://www.frac.info. [31 Okt 2011].
Hans Y, Bonos SA, Clarke BB and Meyer WA .2003. Inoculation techniques forselection of gray leaf spot resistance in perennial ryegrass. USGA Turfgrassand Environmental Research online, 2:19.
Hasti., D.P.P, 2016. Dampak Varietas Campuran Terhadap Kelimpahan CendawanTanah dan Intensitas Penyakit Blas (Pyricularia grisea).UniversitasHasanuddin. [Thesis]

50
[IRRI] International Rice Research Institute. 1996. International EvaluationSystemforRice.4editionhttp://www.knowledgebank.irri.org/images/docs/ricestandard-evaluation-system.pdf [diakses 4 juni 2016 ].
Kahmann R, and Basse C. 1997. Signaling and development in pathogenic fungi-new strategies for plant protection. Trends in Plant Sci. 2: 366–36.
Karthikeyan V, Rajarajan R., and Gnanamanickam SS. 2013. Virulencecharacteristic analysis and identification of new phatotypes of rice blastfungus (Magnaporhte grisae) from India.Life Science Feed 2(1): 7-12.
Kato, H., Sasaki, T., and Koshimizy.1970. Potential for conidium formation of P.oryzae in cesions on leaves and penicles. Phytophatology 60: 608-612.
Keon J.P., C.S. James, and S. Court. 1994. Isolation of Erg2 gene, encoding steroldelta 8 to delta isomerase, from the rice blast fungus Magnaporthe grisea andits expression in the maize smut pathogen Ustilago maydis. Curr. Genet. 25:531-537
Keon JPR, James CS, Court S, Daintree CB, Bailey AM, Burden RS, Bard M,Hargreaves JA. 1994. Isolation of the gene, encoding sterol Δ Δ 7 isomerase,from the rice blast fungus and its expression in the maize smut pathogen CurrGenet. 25(6):531-537.
Kobayashi, N., Ebron, L. A., Fujita, D., and Y. Fukuta. 2009. Identification of blasresistance genes in IRRIbred rice varieties by segregation analysis based ondifferential system. p.69-86. In Yoshimichi Fukuta, Casiana M. Vera Crus andN. Kabayashi (Ed.). Development and Characterization of Blast ResistanceUsing Differential Varieties in Rice. JIRCAS Working report No. 63. Tsukuba,Japan
Koizumi, S. 2009. Monitoring of blas races to ensure durability of blast resistance inJapanese rice cultivars. p.1-9. In Yoshimichi Fukuta, Casiana M. Vera Crusand N. Kabayashi (Ed.). Development and Characterization of BlastResistance Using Differential Varieties in Rice. JIRCAS Working report No.63. Tsukuba, Japan.
Lestari, A. Widyastuti,U., Enggarini, W., 2016. Uji Virulensi 100 Isolat CendawanBlas (Pyricularia oryzae Cavara) terhadap Satu Set Varietas Padi DiferensialIndonesia. Jurnal Agrotek Indonesia 1 (1) : 37 – 46 (2016)
Lestari, P., Trijatmiko, R.T., Reflinur, Warsun, A., Tasliah, Ona, I., Vera Cruz, C., andM. Bustaman.2011. Mapping quantitative trait loci conferring blast resistancein upland indica rice (Oryza sativa L.). J. Crop Sci. Biotech. 14(1): 57-63.
Lestari, P. , Wawan, Tri P. Priyatno,Enggarini, W. Reflinur,dan Suryadi, Y.2014.Isolasi,Identifikasi,danKarakterisasi Cendawan Blas Pyricularia oryzaeHasil Rejuvenasi. Buletin Plasma Nutfah Vol.20 (1): 19-26.

51
Leung H, Shi Z. 1994. Genetic regulation of sporulation in the rice blast fungus. Didalam : Zeigler RS, Leong SA, Teng PS, editor. Rice blast disease. ManilaPhilippines : CAB International-IRRI. hlm 65-68.
Listiyowati, S. 2012. Keragaman Genetik Dan Mikroevolusi Pyricularia Grisea AsalRumput.IPB: Hal 80 [disertasi]
Mogi,S. Sugandi, B., Edwina, R., Cahyadi. 1991. Establishment of the DifferentialVariety Series for Pathogenic Race Identification of Rice blast Fungus andDistribution of Race Based On The new Differential Indonesia.Karawang:Jatisari. Indonesia. Rice Disease Study Group.
Namai T. 2011. Race differentiation of the rice blast fungus, Pyricularia oryzae, andenvironmentally friendly control of rice blast disease. J General Plant Pathol:doi: 10.1007/s10327-011-0328-8.
Nasution A, Amir M, Rachmawati SR, Zalkia S. 1992. Populasi ras jamur Pyriculariaoryzae Cav. dari daerah Karang Agung dan Tamanbogo. Seminar TahunanBalai Penelitian dan Pengembangan Tanaman Pangan Bogor. Inpress.
Nasutian A., Santoso, dan Sudir. 2013. Pemetaan ras blas (Pyricularia grisea) yangmenyerang padi sawah di daerah Jawa Barat. Buku 2. hlm. 1093-1104.Dalam Abdulrachman, S., Pratiwi, G.R., Ruskandar, A., Nuryanto, B., Usyati,N., Widyantoro, Guswara, A., Samita, P., Mejaya, M. J., (Ed.). ProsidingSeminar Nasional Inovasi Teknologi Adaptip Perubahan Iklim GlobalMendukung Surplus 10 Juta ton Beras Tahun 2014. Badan Penelitian danPengembangan Pertanian Kementerian pertanian
Neto, J. J. D., G. R. dos Santos, L. M. dos Anjos, P. H. N. Rangel, and M. E.Ferreira. 2010. Hot spots for diversity of Magnaporthe oryzae physiologicalraces in irrigated rice fields in Brazil. Pesq. agropec. bras. [online].45 (3):252-260.
Otsuka H, Tamari K and Agasawara N, 1965. Variability of Pyricularia oryzae inculture. In the Rice Blast Disease. John Hopkins press, Baltimore. Pp 69–109.
Ou SH. 1979. A Handbook of rice diseases in the tropics. 3rd ed. International RiceResearch Institute, Los Banos, Philippines. p. 17-2.
Ou SH. 1985. Rice disease. Commonwealth Mycological Institute. Kew Surrey.England. P. 125-132.
Prabawa, P, S. Yulianah I., dan Basuki N., 2014. Uji Ketahanan 10 Genotip PadiMerah (Oryza Sativa L.) Terhadap Penyakit Blas Daun (Pyricularia OryzaeCav.) Ras 173. Jurnal Produksi Tanaman, Vol 3 (6):hlm. 496 – 502
Purnomo, B. 2002. Pengantar Epidemiologi Penyakit Tanaman. Bengkulu (ID):(diktat) Faperta Unib

52
Reflinur, Bustaman. M., Widyastuti U., dan Aswidinnoor H. 2005. Keragaman genetikcendawan Pyricularia oryzae berdasarkan primer spesifik gen virulensiJurnal Bioteknologi Pertanian, Vol. 10(2): 55-60.
Reflinur. 2005. Keragaman genetik cendawan Pyricularia grisea berdasarkan primerspesifik gen virulensi dan interaksinya terhadap gen ketahanan padi [tesis].Bogor : Pascasarjana, Institut Pertanian Bogor. 42 hlm.
Santoso, A. Nasution, D.W. Utami, I. Hanarida, A.D. Ambarwati, S. Mulyopawiro,dan D. Tharreau. 2007. Variasi genetik dan spectrum virulensi pathogen blaspada padi asal Jawa Barat dan Sumatera. Jurnal Penelitian PertanianTanaman Pangan 26(3): 150-155.
Santoso dan A. Nasution. 2008. Pengendalian penyakit blas dan penyakit cendawanlainnya. Buku Padi 2. Hlm. 531563. Dalam Darajat A.A., A. Setyono, A.K.Makarim, dan A. Hasanuddin (Ed.). Padi Inovasi Teknologi. Sukamandi: BalaiBesar Penelitian Tanaman Padi
Santoso, dan Nasution A. 2009. Pengendalian penyakit blas dan penyakit cendawanlainnya. Dalam Inovasi Teknologi Produksi Padi. Buku 2. Balai BesarPenelitian Tanaman Padi. Badan Penelitian dan Pengembangan Pertanian.p: 531-563.
Santoso, dan Nasution A. 2012. Pengendalian penyakit blas dan penyakit cendawanlainnya. Dalam Inovasi Teknologi Produksi Padi. Buku 2. Balai BesarPenelitian Tanaman Padi. Badan Penelitian dan Pengembangan Pertanian.p: 531-56.
Scardaci SC, Webster RK, Greer CA, Hill JE, William JF, Mutters DM, Brandon RG,McKenzie KS, and Oster JJ. 1997. Rice blast: A new disease in California. J.Agr. Fact. Sheet Ser. 1:2-5
Semangun, H. 1991. Penyakit-Penyakit Tanamaan Pangan Di Indonesia.Yogyakarta: Gadja Mada University Press. P.449
Shirai M,. 1896. Notes on plants Collected in Suruga, Totomi, Yamato and Kii.Botanical Magazine Tokyo. Vol 10:111-114
Skamnioti P, Gurr SJ. 2007 Aug. cutinase2 mediates appressorium differentiationand host penetration and is required for full virulence. Plant Cell. 19:2674-2689.
Soetarto, Jasis A, Subroto SWG, Siswanto M, dan Sudiyanto E. 2001. Sistemperamalan dan pengendalian organisme pengganggu tanaman (OPT)mendukung sistem produksi padi berkelanjutan. Dalam Las et al. (eds.).Implementasi Kebijakan Strategis untuk Meningkatkan Produksi Padiberwawasan Agribisnis dan Lingkungan. Pusat Penelitian danPengembangan Tanaman pangan. hal. 247.

53
Soubabere, O., V. Jorge, J.L. Notteghem, M.H. Lebrun, and D.Tharreau. 2001.Sequence characterized amplified regionmarkers for the rice blastfungus,Magnaporthe grisea.Mol. Ecol. Notes 1(1-2):11-12
Srivastava,.D, Shamim,.Md, Kumar., D, Pandey., P, Khan,.N.A, and Singh,. K.N.2014. Morphological and Molecular Characterization of Pyricularia oryzaeCausing blast Disease in Rice (Oryza sativa) from North india. IJRS. Volume4[7]:4
Sudir, Yuliani D., Nasution A., Nuryanto B. 2013.Pemantauan penyakit utama padisebagai dasar skrining ketahanan varietas dan rekomondasi pengendalian dibeberapa daerah sentra produksi padi di Jawa. Laporan Hasil PenelitianBalai Besar Penelitian Tanaman Padi, Sukamandi. p33.
Suwandi, H. Hamidson, dan A. Muslim. 2016. Penekanan penyakit blas leher malaipadi menggunakan ekstrak kompos jerami padi. Jurnal Fitopatologi Indonesia12(3):104–108
Sweigard, J.A., F.G. Chumley, and B. Valent. 1992. Disruption of Magnaporthegrisea cutinase gene. Mol. Gene Genet. 232: 183-190.
Tasugi, H and L. Yoshida.1959. Relation Between Rice Blast Resistance and SuhueEnvironment. Ann. Phytopth. Soc. Japan
TeBeest, D.O., C. Guerber, and M. Ditmore. 2007. Rice blast. The Plant HealthInstructor. DOI. 10.1094/PH 11-20070313-07 APSnet.http://www.apsnet.org/edcenter/nintropp/lessons/fungi/ascomycetes/Pages/RiceBlast.aspx Cited on 27 August 2016.
Teng, P.S., H.W. Klein-Gebbinck, and H. Pinnschmidt. 1991. An analysis of the blastpathosystem to guide modeling and forcasting. pp.1-30. Los Banos,Philippines. International Rice Research Institute.
Utami DW. 2000. Evaluasi sifat ketahanan terhadap tiga ras uji penyakit blas(Pyricularia grisea) pada spesies padi liar Oryza rufipogon dan populasitanaman BC2 F turunannya [tesis]. Bogor: Pascasarjana, Institut PertanianBogor.
Utami, D.W. Aswidinnoor, H., Moelyopawiro, S., Hanarida, I., dan Reflinur.2006.Pencarian Alel-Alel Baru untuk Gen-gen Penting Toleran Cekaman Biotik danAbiotik pada Padi. Laporan Hasil Penelitian. Bogor.
Valent, B. and F.G. Chumley. 1994. Avirulence genes and mechanisms of geneticinstability in rice blast fungus. p. 111-134. In R.S. Zeigler, S.A. Leong, andP.S. Teng (Eds.). Rice Blast Disease. Wallingford (UK): CABInternationalIRRI, Manila, Philippines.
Wakman W., Rahamma S., and Hasanuddin A., 1981. Rice blast disease outbreakin Sinjai and Bulukumba, South Sulawesi, Indonesia . Research Institute for

54
Food Crops, South Sulawesi, Indonesia. In International Rice ResearchInstitute. 1981. vol 6(6): Filipina.
Winarti L. 1992. Hubungan antara keadaan cuaca dengan intensitas penyakit blas(P. oryzae) dan pengaruhnya terhadap hasil padi gogo di KP Tamanbogo,Lampung Tengah. Tesis Fakultas Biologi Universitas nasional Jakarta.
Yuliani. D. dan Maryana. Y.E.,2014. Integrasi Teknologi Pengendalian Penyakit Blaspada Tanaman Padi di Lahan Sub-Optimal [prosiding]:Palembang
Ziegler, R.S. 1998. Recombination in Magnaporthe grisea. Ann. Phytopathology36:249-275.
Zeigler RS, Tohme J, Nelson R, Levy M, and Correa-Victoria FJ. 1994. Lineageexclusion : A proposal for linking blast population analysis to resistancebreeding. Rice Blast Disease. CAB International IRRI 267-292.
.

55
LAMPIRAN

56
Gambar lampiran 1. Survey lokasi serangan blas di kab. Bone (gejala blas padadaun padi).
Gambar lampiran 2. Survey lokasi serangan blas di kab. Maros (gejala blas padadaun padi).
Gambar lampiran 3. Survey lokasi serangan blas di kab.Gowa (gejala blas pada (A)Kolar daun, (B)leher malai dan (C) Daun padi).

57
CILIWUNG
Nomor seleksi : B4183B-PN-33-6-1-2Asal persilangan : IR38//2*Pelita I-1/IR4744-128-4-1-2Golongan : CereUmur tanaman : 117 - 125 hariBentuk tanaman : TegakTinggi tanaman : 114 - 124 cmAnakan produktif : 18 - 25 batangWarna kaki : HijauWarna batang : HijauWarna telinga daun : Tidak berwarnaWarna lidah daun : Tidak berwarnaWarna daun : Hijau tuaMuka daun : KasarPosisi daun : TegakDaun bendera : Miring sampai tegakBentuk gabah : Sedang sampai rampingWarna gabah : Kuning bersihKerontokan : SedangKerebahan : TahanTekstur nasi : PulenKadar amilosa : 22%Indeks Glikemik : 86Bobot 1000 butir : 23 gRata-rata hasil : 4,8 t/haPotensi hasil : 6,5 t/haKetahanan terhadapHama Penyakit : Tahan wereng coklat biotipe 1, 2 dan rentan wereng
coklat biotipe 3 Agak tahan terhadap hawar daun bakteristrain IV
Anjuran tanam : Baik ditanam di lahan irigasi berelevasi rendah sampai 550m dpl
Pemulia : I. Sahi, Taryat T., dan H. MaknunDilepas tahun : 1988

58
INPARI 4
Nomor Persilangan : BP2280-1E-12-2Asal persilangan : S4384F-14-1/Way Apo Buru/S4384F-14-1Golongan : CereUmur tanaman : 115 hariBentuk tanaman : SedangTinggi tanaman : 95 – 105 cmAnakan produktif : 16 anakanWarna kaki : HijauWarna telinga daun : PutihWarna lidah daun : HijauWarna daun : HijauPermukaan daun : KasarPosisi daun : TegakPosisi daun bendera : TegakWarna batang : HijauKerebahan : SedangKerontokan : SedangBentuk gabah : Panjang dan RampingWarna gabah : Kuning bersihRata-rata hasil : 6,04 t/haPotensi hasil : 8,80 t/ha GKGBobot 1000 butir : 25 gTekstur nasi : PulenKadar amilosa : 21,07 %Ketahanan terhadapHama: Agak rentan terhadap hama Wereng Batang Coklat Biotipe 1,2 dan 3Ketahanan terhadap penyakit : Agak tahan terhadap penyakit Hawar Daun Bakteri
strain III dan IV serta agak rentan strain VIII, agak tahan terhadap HawarDaun Bakteri strain IV dan VIII, agak tahan penyakit virus tungroinokulum varian 073 dan 031
Keterangan : Cocok ditanam pada lahan irigasi dengan ketinggiansampai 600 m dpl.
Pemulia : Aan A. Daradjat, dan Bambang Suprihatno.Peneliti : I.N. Widiarta, Baehaki S.E., Triny S.K., S. Dewi Indrasari,
Prihadi Wibowo, Omi Syahromi, Nafisah, Cucu GunarsihEstria Furry P.
Teknisi : Toyib S. Ma`ruf, Maman Suherman, Meru, Uan Sudjanang,

59
M. Sailan, Zaenal Arifin, Karmita, Sukanda, Suwarsa, DedeMunawar.
Pengusul : Balai Besar Penelitian Tanaman PadiAlasan utama dilepas : Lebih tahan terhadap HDB Strain IV daripada Ciherang,
hasil dan mutu sama dengan CiherangDilepas tahun : 2008
CIHERANG
Nomor seleksi : S3383-1D-PN-41-3-1Asal persilangan : IR18349-53-1-3-1-3/3 *IR19661-131-3-1- 3//4*IR64Golongan : CereUmur tanaman : 116-125 hariBentuk tanaman : TegakTinggi tanaman : 107-115 cmAnakan produktif : 14-17 batangWarna kaki : HijauWarna batang : HijauWarna telinga daun : Tidak berwarnaWarna lidah daun : Tidak berwarnaWarna daun : HijauMuka daun : Kasar pada sebelah bawahPosisi daun : TegakDaun bendera : TegakBentuk gabah : Panjang rampingWarna gabah : Kuning bersihKerontokan : SedangKerebahan : SedangTekstur nasi : PulenKadar amilosa : 23%Indeks Glikemik : 54Bobot 1000 butir : 28 gRata-rata hasil : 6,0 t/haPotensi hasil : 8,5 t/haKetahanan terhadapHama Penyakit : Tahan terhadap wereng coklat biotipe 2
Tahan terhadap hawar daun bakteri strain III dan IVAnjuran tanam : Baik ditanam di lahan sawah irigasi dataran rendah sampai
500 m dpl.Pemulia : Tarjat T, Z. A. Simanullang, E. Sumadi dan Aan A. Daradjat

60
Dilepas tahun : 2000
CIGEULIS
Nomor seleksi : S3429-4D-PN-1-1-2Asal persilangan : Ciliwung/Cikapundung//IR64Golongan : CereUmur tanaman : 115-125 hariBentuk tanaman : TegakTinggi tanaman : 100 – 110 cmAnakan produktif : 14 – 16 batangWarna kaki : HijauWarna batang : HijauWarna telinga daun : Tidak berwarnaWarna lidah daun : Tidak berwarnaWarna daun : HijauMuka daun : Agak kasarPosisi daun : TegakDaun bendera : TegakBentuk gabah : Panjang rampingWarna gabah : Kuning bersihKerontokan : SedangKerebahan : SedangTekstur nasi : PulenKadar amilosa : 23%Indeks glikemik : 64Bobot 1000 butir : 28 gRata-rata hasil : 5,0 t/haPotensi hasil : 8,0 t/haKetahanan terhadapHama Penyakit : Tahan terhadap wereng coklat biotipe 2 dan rentan biotipe 3
Tahan terhadap hawar daun bakteri strain IV•Anjuran tanam : Baik ditanam pada musim hujan dan kemarau, cocok
ditanam pada lokasi dibawah 600 meter di atas permukaa lautInstansi pengusul : Balitpa dan, BPTP LampungPemulia : Z.A. Simanullang, Aan A. Daradjat, dan N. YunaniTim peneliti : B. Suprihatno, M.D. Moentono, Ismail B.P., Atito D.,
Baehaki S.E., Triny S.Kadir dan W. S.Ardjasa.Teknisi : Toyib S.M., Edi Suwandi M.K., M. Suherman, dan Sail HanafiDilepas tahun : 2002




















