
Isolation of ORCTL3 in a novel genetic screen for tumor-specific apoptosis inducers

Detection of Mutagens and BaPMO Inducers inRiver Water Using the Blue CottonAdsorption Technique
Smiljana Britvic,1 Branimir K. Hackenberger,2 Davorka Jaric,2 Sandra Stepic2
1Center for Marine and Environmental Research, Rudjer Boskovic Institute, Zagreb, Croatia
2Department of Biology, J. J. Strossmayer University, Osijek, Croatia
Received 26 November 2009; accepted 16 April 2010
ABSTRACT: Induction of Mixed Function Oxidase (MFO) activity and bioactivation potential were measuredin experimental carp and in native fish species from two rivers with different pollution level (Sava and Mrezn-ica). The experimental carp were intraperitoneally exposed to various water volume equivalents of Blue Cottonextracts from both rivers. The induction of MFOmeasured as a Benzo(a)pyrene monooxygenase (BaPMO) ac-tivity was increased up to 9.3-fold in experimental carp and up to 11.3-fold in native fish from Sava River,whereas the values from Mreznica River showed only a slight increase when compared with the control(highest increase of 1.8-fold in nose carp). Accordingly, bioactivation potential using modified Ames test washigher in both experiments with Sava River. Both measured parameters in experimental carp increased in adose-dependent manner in accordance to river volume equivalents. Different induction potential of native fishspecies comparable between two rivers confirmed the known possibility of their usage in biomonitoring stud-ies. These results give qualitatively a new support to the idea of using Blue Cotton extraction technique com-bined with induction of MFO activity and Ames test in fish as a good biomarker for assessing risk of exposureto mutagens/carcinogens such as Polycyclic Aromatic Hydrocarbons (PAHs), especially due to the simplicityand cost-effectiveness of these methods. # 2010 Wiley Periodicals, Inc. Environ Toxicol 27: 146–154, 2012.
Keywords: Ames test; benzo(a)pyrene monooxygenase activity; Blue Cotton; fish; MFO; bioactivationpotential
INTRODUCTION
Majority of surface waters receive large quantities of waste
water from various sources: industrial, agricultural, and
domestic. As those waters are further used for different pur-
poses they can become a serious public health and aquatic
ecosystem problem. Many compounds detected in those
wastewaters are known mutagenic or genotoxic carcinogens
(Ohe et al., 2004). Consequently, those compounds become
the components of complex environmental mixtures that
can have adverse health effects on humans and indigenous
biota (Dearfield et al., 2002). In numerous studies, munici-
pal river water mutagenicity was reported with its character
derived from PAHs including nitro- and amino-derivates
(Sakamoto and Hayatsu, 1990; Nagai et al., 2002; Umbu-
zeiro et al., 2004).
The effective concentrations of mutagens in water can
very often be below the detection limit of analytical instru-
ments. If the bioaccumulation index of majority of muta-
gens is taken under consideration, which is quite high, then
an importance of small quantities of mutagens detection in
the waters is understandable. Thus, in such research water
samples must be concentrated and proper extraction is of
great importance for both the chemical analysis and Ames
test. In many mutagenicity investigations, as a preconcen-
tration methods XAD-2 and XAD-7 resins, Blue Cotton,
Blue Rayon, and Blue Chitin have been used (Hayatsu
et al., 1983; Sayato and Hayatsu, 1990; Nagai et al., 2002;
Correspondence to: B.K. Hackenberger; e-mail: [email protected]
Published online 1 June 2010 in Wiley Online Library
(wileyonlinelibrary.com). DOI 10.1002/tox.20625
�C 2010 Wiley Periodicals, Inc.
146

Umbuzeiro et al., 2004). Blue pigment has a ligand linked
copper phthalocyanine trisulphonate and is capable of
adsorbing aromatic compounds such as PAHs and heterocy-
clic amines (HCA) with two or more fused rings. The
adsorption capacity is strong, and the selectivity for adsorb-
ing this class of compound is high (Kobayashi and Hayatsu,
1984; Hayatsu et al., 1985). Additionally, the methods with
blue pigment show recoveries comparable to or better than
existing solid phase extraction methods, and are faster and
simpler than most of them (Skog, 2004). Therefore, they
have been extensively used in screening fresh and marine
water for the presence of mutagenic compounds (Kurelec
and Krca, 1989; Sakamoto and Hayatsu, 1990; Hayatsu,
1992; Sayato et al., 1993).
Besides the development of effective, economic, and
selective passive sampling techniques, a need for techni-
ques which fulfill the demands of monitoring trace and
ultra-trace bioavailable organic contaminants in water and
at the same time bridge the gaps between environmental an-
alytical chemistry and ecotoxicology approaches exists (Lu
et al., 2002).The coupling of bioassay assessments with
selective sampling technique yields more information than
either of these assessments individually.
The Ames mutagenicity test, with and without metabolic
activation by S9 mixture, has been frequently coupled with
blue pigment extraction technique for the detection of
mutagens in surface waters (White and Rasmussen, 1998;
Kataoka et al., 2000; Watanabe et al., 2002; Ohe et al.,
2003; Umbuzeiro et al., 2004) and drinking waters
(Schenck et al., 1998; Monarca et al., 2002) throughout the
world. As property of blue pigment is an adsorption of sub-
stances with planar polycyclic molecular structure, an
induction of cytochrome P450-dependent monooxygenase
(MFO) system in fish liver as a biomarker and bioindicator
of exposure to anthropogenic organic contaminants such as
PAHs, Polychlorinated Biphenyls (PCBs), several pesti-
cides and dioxins/furans could additionally be used (Buhler
and Wang-Buhler, 1998; Parrott et al., 2000). This can be
obtained with MFO-induct test, i.e., measurement of ben-
zo(a)pyrene monooxygenase (BaPMO) induction in the
carp liver after intraperitoneal (i.p.) application of sampled
water extracts (Kurelec et al., 1979). Previous studies were
conducted using hexane, XAD-2 or XAD-7 extracts of pol-
luted fresh and marine water samples, or dichloromethane
extracts of soil and waste from the municipal landfill. Those
extracts were i.p. injected into juvenile carp (Cyprinus car-pio) with consecutive measurement of MFO activity induc-
tion (Kurelec et al., 1982; Rijavec et al., 1981). The MFO
induct-test has been used in combination with the Ames
test (Kurelec et al., 1979, 1981) using liver homogenates
from carps i.p. injected with 3-methylcholanthrene (Protic-
Sabljic and Kurelec, 1983; Britvic et al., 1993).
The primary objective of this research was to investigate
the potential usage of the Blue Cotton adsorption technique
as the method for detection and concentration of MFO-
inducing lipophilic xenobiotics and premutagens in envi-
ronmental monitoring. For this purpose, two experiments
were conducted: (i) a laboratory experiment with juvenile
carp intraperitoneally exposed to Blue Cotton extracts
(MFO inducting experiment) and (ii) a field experiment
with native fish specimens from two investigated rivers
(Sava and Mreznica). Fish hepatic S9 was used to measure
MFO induction as the BaPMO activity and bioactivation
potential as the Ames-microsomal test in both experiments.
Additionally, BaPMO activity in S9 from juvenile carp was
measured with addition of a-naphtoflavone (ANF), a well
known inhibitor of aromatic hydrocarbon-inducible forms
of cytochrome P450.
MATERIALS AND METHODS
Sampling Sites
A volume of 20 L of water was collected on five adjacent
sublocations from Sava River at the power plant Zagreb II
cooling water pumping station and from Mreznica River
upstream from the city of Karlovac. A total of 100 L was
collected from each river. The quality of water from each
of these rivers was defined by its load of industrial and
domestic waste, expressed in population equivalents (p.e.)
(Sava river 2,400,000 p.e. and Mreznica river 2000 p.e.)
and according to physicochemical parameters (Sava river
III-V category and Mreznica river I-II category) (Vouk and
Malus, 2005; Environmental Health 1993). Recently,
Grung et al. (2007) identified several PAHs (alkylated
naphtalenes, acenaphtene, dibenzofuran, fluorine, alkylated
fluorine, and phenanthrene/anthracene) using reverse phase
HPLC fractionation technique in the wastewater effluent
from Zagreb which are released in the Sava River. The
water samples were transported to the laboratory and
extracted within 24 h with the Blue Cotton technique.
Experimental Organisms
For MFO inducting experiment, carp (Cyprinus carpio) 1year old, sexually immature, weighing 22 6 5 g were
obtained from the fish farm Draganici and were kept for up
to 3 months in laboratory basins with a flow of dechlori-
nated, well aerated tap water.
Native fish from Sava and Mreznica rivers were caught
by angling. Fish species caught were: Danubian roach
(Rutilus pigus virgo)(nSava 5 8, nMreznica 5 6), roach
(Rutilus rutilus)(nSava 5 6, nMreznica 5 5), chub (Leuciscuscephalus)(nSava 5 7, nMreznica 5 6), nose carp (Chondros-toma nasus)(nSava 5 8, nMreznica 5 5), and bleak (Alburnusalburnus)(nSava 5 8, nMreznica 5 6). All fish specimens used
in the experiment were sexually immature. The liver was sep-
arated immediately after the catch and transported in liquid
nitrogen to the laboratory and stored at2808C until analysis.
147BLUE COTTON COUPLED WITH MFO INDUCTION AND AMES TEST
Environmental Toxicology DOI 10.1002/tox

The livers of both native fish and carp were used for mea-
surement of BaPMO activity and for the preparation of S9
fraction in the Ames Salmonella assay. Livers were homoge-
nized with Potter-Elvehjem homogenizer (Sartorius, Goetin-
gen, Germany) in sucrose buffer (0.05 M Tris, 0.25 M sucrose
in 1% KCl, pH 5 7.6) at a tissue weight/buffer volume ratio
of 1:5, centrifuged at 90003 g for 20 min at 48C (S9 fraction)
and resulting homogenates were stored at2808C until use.
Preparation of Blue Cotton WaterConcentrates
Blue Cotton fibers were washed and tested for mutagenicity
activity with the TA100 strain and when the extracts
showed negative results they were used in the experiment
(Kummrow et al., 2003).
Water samples from Sava and Mreznica rivers were
passed through a set of five glass columns (200 mm 3 20
mm) with 2 g Blue Cotton each (a total of 10 g of Blue Cot-
ton per sample) at a flow rate of 70 mL/min at room tem-
perature. Afterwards, the Blue Cotton was carefully
removed from columns and washed with 50 mL of distilled
water per gram in order to remove the nonadsorbed com-
pounds and solids. The water was removed with aspiration
and the residual water was wiped out with a paper towels.
Then the adsorbed materials were extracted by gently shak-
ing the Blue Cotton in 50 mL/g Blue Cotton of methanol-
concentrated ammonia (50:1, v/v) for 1 h (Kira et al.,
1989). The eluants were combined and reduced to the vol-
ume of 6 mL in a rotary evaporator and further used for i.p.
application in carp (MFO inducting experiment).
Measurement of BaPMO Activity
In a MFO inducting experiment, an aliquot of the Blue Cot-
ton extract (equivalent to 1, 3, and 6 L of river water) was
evaporated to dryness and redissolved in 4.0 mL of corn
oil. The extracts prepared in this way were i.p. applicated to
experimental carp on a body weight basis (20 mL/kg). For
a positive control carp were i.p treated with 50 mg/kg of
3-methylcholanthrene (3-MC) and for the negative control
only corn oil was administered. Seven specimens were used
per treatment (C1, C2, 1, 3, and 6 L equivalents); all treat-
ments were done in triplicate. After 3 days, carp were sacri-
ficed and liver was prepared for determination of BaPMO ac-
tivity by measuring 3-hydroxy-benzo(a)pyrene (3-OHBaP)
concentration. BaPMO activity was determined in liver post-
mitochondrial fractions (S9) according to the method of
Nebert and Gelboin (1968). The inhibitory effect on the
BaPMO activity was measured in each sample in parallel af-
ter addition of 1024 M ANF in the reaction mixture. The
activities were expressed as pmol of BaPOH per mg of pro-
tein per min. Proteins were determined according to the
method of Lowry et al. (1951).
The livers from native fish specimens of both rivers
were extracted and S9 fraction was prepared (at least five
specimens per species). BaPMO activity was determined in
liver postmitochondrial fractions (S9) according to the
method of Nebert and Gelboin (1968). The activities were
expressed as pmol of BaPOH per mg of protein per min.
Proteins were determined according to the method of
Lowry et al. (1951).. For the negative control intact experi-
mental carp was used.
Ames Test
The potential of liver postmitochondrial fractions (S9) from
the experimental carp induced with i.p. injection of Blue
Cotton extracts or from natural fish specimens for bioacti-
vation of precarcinogen BaP was measured with a modified
Ames test (Britvic and Kurelec, 1986). Bioactivation poten-
tial was expressed as the net increase of the number of
revertants activated from BaP (10 lg/plate) with S9 (2 mg/
protein/plate) over the spontaneous number (125 6 6) of
Salmonella typhimurium TA100 revertants. Plates were
incubated upside down at 378C and colonies (His1 rever-
tants) were counted with automatic colony counter after 48
h. Each sample was tested in triplicate and all data were
presented as averages of revertants numbers. As positive
controls, methyl methanesulphonate (MMS) 625 lg/platewas used to monitor the sensitivity of the bacterial strain
TA100 and BaP 10 lg/plate for the activity of carp liver
S9. Additionally, for a positive control carp were i.p treated
with 50 mg/kg of 3-methylcholanthrene (3-MC) and for the
negative control only corn oil was administered. There was
no positive and negative control for native fish species. Val-
ues of mutagenicity ratio (MR) were obtained by dividing
the number of revertant colonies in sample plates by the
spontaneous number of revertants. A MR [2 with a clear
dose response was defined as mutagenic, whereas a MR\2
was defined as a lack of mutagenicity.
Statistical Analysis
Prior to comparison among various groups of fish and corre-
lation analysis, the data were tested for normality (Shapiro-
Wilk and Kolmogorov-Smirov tests) and homogeinity of
variance (Levene’s test). The BaPMO activities were trans-
formed using the logarithmic function f(x) 5 log(1 1 x).The Ames test results (number of revertants) were trans-
formed using the square root function f(x) 5 Hx. Thesetransformations resulted in homogenous variance and nor-
mal distribution of data enabling the usage of parametric
statistical methods. For comparison of groups treated with
different equivalent of water, single-way analysis followed
by Dunnet’s or Tukey’s posttest was used. Kruskal-Wallis
followed by Dunn’s posttest and Spearman rank correlation
yielded similar results as parametric methods.
148 BRITVIC ET AL.
Environmental Toxicology DOI 10.1002/tox

RESULTS
To investigate the potential usage of Blue Cotton extraction
method for measurement of MFO induction and bioactiva-
tion potential in river monitoring research several experi-
ments were conducted.
BaPMO Activity in Experimental Carp
Results of the MFO inducting experiment showed that the
xenobiotics extracted from the Sava River induced BaPMO
activity in experimental carp in a dose-dependent manner,
according to the river water volume equivalent [Fig. 1(a)].
Namely, the amount of xenobiotics in 1 L, 3 L, and 6 L of
extract equivalents from the Sava River caused 3.9, 5.3,
and 9.3-fold increase of the BaPMO activity (12.4 6 2.3,
16.8 6 4.1, and 29.9 6 1.8 pmol BaPOHmgprot21min21,
respectively) when compared with the basal BaPMO activ-
ity (3.2 6 0.3 pmol BaPOHmgprot21 min21), i.e., negative con-
trol. Those values expressed as 3-MC toxicity equivalents
would correspond to activities induced with i.p. injection of
19.3 mg/kg, 26.2 mg/kg, and 46.6 mg/kg 3-MC, respectively.
The increase obtained with 6 L river water equivalent was
close to maximum induction obtained with 50 mg kg21 of
3-MC (32.1 6 3.8 pmol BaPOHmgprot21 min21), i.e., positive
control. In contrast, Blue Cotton water extracts from Mreznica
River, as expected, did not significantly induce BaPMO activ-
ity in experimental carp. The highest induction of only
1.2-fold over control (4.5 6 0.3 pmol BaPOHmgprot21 min21)
was obtained with 6 L river water volume equivalent. The
other two water volume equivalents did not show statistically
significant difference between themselves or with the control
(P\0.05).
In the experiment with ANF, the induction of BaPMO
with extracts from Sava River decreased significantly [Fig.
1(b)]. Obviously, highest difference, when compared with
the previous experiment, was with the 6 L extract of both
rivers showing that when larger volumes of water are
extracted the quantity of MFO inducers increased. Even
though, no statistically significant difference was observed
between 3 and 6 L extract of Sava River and positive con-
trol, and with all three extract quantities of Mreznica River
and negative control. Six liter water extract from the Sava
River (9.3 6 0.8 pmol BaPOHmgprot21min21) elicited
stronger BaPMO induction than the positive control (8.3 62.2 pmol BaPOHmgprot
21 min21).
BaPMO Activity in Native Fish
Apart from laboratory, a field experiment with native fish
populations from both rivers was conducted. In all fish spe-
cies native to polluted Sava River, when compared to
untreated juvenile carp used as a negative control, an
increase in BaPMO activity was measured (from 1.3
to 11.3-fold) (Fig. 2). Simultaneously, of all fish species
native to the Mreznica River only nose carp showed a slight
increase in BaPMO activity when compared to control (1.8-
fold). Interestingly, this species showed the highest induction
of BaPMO activity in both rivers (11.3 and 1.8-fold) (Fig. 3).
Fig. 1. BaPMO activities in carp i.p. induced with Blue Cot-ton river water volume extracts from Sava River ( ) 1 L, ( )3 L, and ( ) 6 L and Mreznica River ( ) 1 L, ( ) 3 L, and ( ) 6L, and BaPMO activities for positive control (i.p. inductionwith 3-MC) ( ) and negative control (only corn oil) ( ), (A)without addition of ANF and (B) with addition of ANF. Statis-tically significant differences are compared to negative con-trol: P\0.05 (*), P\0.01 (**) and P\0.001 (***).
Fig. 2. BaPMO activities in native fish specimens caught inSava ( ) and Mreznica ( ) River.
149BLUE COTTON COUPLED WITH MFO INDUCTION AND AMES TEST
Environmental Toxicology DOI 10.1002/tox

From the fish species caught in the Sava River lowest
BaPMO activity was recorded in Danubian roach (4.1 6 0.4
pmol BaPOHmgprot21 min21). From the Mreznica River, except
of nose carp, all tested fish species had similar BaPMO activ-
ities (from 2 6 0.3 pmol BaPOHmgprot21 min21 to 2.7 6 0.6
pmol BaPOHmgprot21 min21) and they were all lower than the
negative control (3.26 0.3 pmol BaPOHmgprot21 min21).
Ames Test
The increase of the potential for bioactivation of BaP to S.typhimurium TA100 mutagens was proportional with the
increase of river water volume equivalent, i.e. the increase
was dose dependent, for both rivers (Table I). The only two
samples that were not statistically significant (P \ 0.05)
when compared with control were 1 L and 3 L extracts
from Mreznica River, as the mean number of revertants
was 137 6 3 and 138 6 7, respectively. Postmitochondrial
fractions (S9) of carp liver pretreated with volume extracts
(1, 3, and 6 L) from Sava River increased the bioactivation
potential of BaP to TA100 mutagens and calculated muta-
genicity ratio (MR) was 3.0, 5.0, and 9.0, respectively. At
the same time, S9 from carps induced with Blue Cotton
extracts from Mreznica River did not induce liver potential
to bioactivate BaP to bacterial mutagens using TA100
strain, the highest MR (1.5) was calculated for the 6 L
equivalent (Table I). Postmitochondrial fractions (S9) from
native fish species generated similar bioactivation as S9
from Blue Cotton induced carp (Table II). Namely, none of
the fish species from Mreznica River increased the potential
for bioactivation of BaP to S. typhimurium TA100 muta-
gens, and the number of revertants after test was more or
less the same as the spontaneous number of revertants. On
the other hand, S9 from most of the Sava River fish species
increased the bioactivation potential of BaP to S. typhimu-rium TA100 mutagens. Only Danubian roach had calcu-
lated MR below 2. The MR of other tested species ranged
from 2.8 to 7.4.
TABLE I. Results of the Ames test using Salmonella TA100 strain for the carp liver S9 previously induced with differentBlue Cotton river water volume equivalents from Sava and Mreznica rivers
Mean Number of Revertants/Plate, Standard Deviation (SD), and Mutagenic Ratio (MR)
DoseSava Mreznica 1 Con 2 Con MMS (2S9) BaP (1S9)
RWE/l Mean SD MR Mean SD MR Mean SD Mean SD Mean SD Mean SD
1 381 9 3.0 137 3 1.1 1085 23 125 6 2000 16 1025 14
3 631 12 5.0 138 7 1.1
6 1125 26 9.0 186 4 1.5
Fig. 3. Correlation between number of revertants in Ames test and BaPMO activity innative fish species from (A) Sava and (B) Mreznica River.
150 BRITVIC ET AL.
Environmental Toxicology DOI 10.1002/tox
DISCUSSION
In the past few decades, many researches have been con-
ducted to assess chemical pollution and consequent poten-
tial ecological hazard of surface freshwaters such as rivers
and lakes. The need for an appropriate and efficient bioas-
say or a set of tests for evaluation of surface waters on
potential hazard to human and the water environment was
the main actuator of this researches. Although, many mod-
ern sophisticated techniques has been developed the need
for a cost-effective methods should not be neglected. In this
work we investigated further potential and efficacy of two
well known tests, MFO inducting experiment and Ames
test, in combination with Blue Cotton extraction technique
to assess the possible potential mutagenicity of two rivers.
Similarly to carp exposed to Blue Cotton extracts, exposure
of carp to a flow of polluted Sava river water for 29 days
resulted in rapid 9.5-fold increase in the BaPMO activity
when compared to untreated carp (Britvic et al., 1993).
Simultaneously, BaPMO in Blue Cotton extract induced
liver was measured with the addition of ANF, a specific in-
hibitor for fish CYP1A (Stegeman and Hahn, 1994). ANF
is known to inhibit CYP1A activity in rainbow trout (Taka-
hashi et al., 1995; Hodson et al., 2007) and in cultured fish
hepatocytes (Manik et al., 2000). In this work, ANF has
decreased measured BaPMO activity in a dose-dependent
manner. The stronger BaPMO induction with 6 L water
extract from the Sava River than the induction elicited with
3-MC in positive control could be explained with the sur-
plus of PAHs causing the induction kinetics to override the
ANF inhibitory effect, or possibly the AhR agonists present
in the complex mixture of river water may have caused syn-
ergism (Basu et al., 2001; Grung et al., 2007). The inducing
potency of a complex mixture of 16 PAHs can be about 2-
fold higher than predicted from their individual induction
equivalency factors (Till et al., 1999). The induction of
BaPMO activity from natural fish specimens from both riv-
ers showed the correlation with the induction of carp
exposed to Blue Cotton extracts. Namely, highest induction
in fish from Sava River was 11.3-fold, and from Mreznica
River 1.8-fold when compared to control. While an
enhanced BaPMO activity in fish from Sava River may
indicate the presence of unknown xenobiotics causing pro-
found pathophysiological and molecular responses, values
from Mreznica River fish could as well be the normal val-
ues of MFO activity in all native fish species living in
unpolluted waters (Britvic et al., 1993) and are suggesting
the absence of xenobiotics with the BaPMO-inducing prop-
erty. Indeed, previous study of BaPMO activity in the popu-
lation of fish living in the polluted Sava river and fish living
in two less polluted Kupa and Krka rivers, showed high
correlation between the level of pollution by communal and
industrial waste and the level of BaPMO induction (Kezic
et al., 1983).
Furthermore, various fish species revealed different lev-
els of BaPMO induction. This species differences in the
magnitude of the response may be influenced by both bio-
logical variables (sex, age, spawning status, size, and
genetic strain) and habitat variables (temperature, pH, and
water flow) (Stegeman and Chevion, 1980; Koivusaari
et al., 1981; Luxon et al., 1987; Lindstrom-Seppa, 1985;
Pedersen et al., 1976; Jimenez and Burtis, 1989). Addition-
ally, large species differences in the induction measured
during field studies may be due to variations in exposure,
accumulation or sensitivity to inducers (Fuchsman et al.,
1999). In this study none of the investigated fish species
where exclusive carnivores or herbivores, their calculated
trophic level ranged from 2.0 6 0.00 (nose carp) to 3.03 60.46 (chub) (Froese and Pauly, 2000). Hence, no unambigu-
ous conclusions could be made from the MFO induction
and species feeding habits or contact with sediments. All
aforementioned leads to large variations in induction capa-
bilities of MFO enzymes in various fish species using very
similar protocols and inducing agents (Kloepper-Sams and
Benton, 1994; Focardi et al., 1995; Smith and Gagnon,
1998). A nose carp (Chondrostoma nasus) had the highest
level of BaPMO activity in both Sava (36.2 6 4.2
BaPOHmgprot21 min21) and Mreznica (5.8 6 0.4
BaPOHmgprot21 min21) rivers (Fig. 2). Previous studies on
this species from Sava River also showed high BaPMO
activities at different sampling sites; with the highest being
69.7 6 9.8 BaPOHmgprot21 min21 measured in liver S9 from
nose carp (Britvic and Kurelec, 1999). The largest differ-
ence between rivers was observed in chub as BaPMO activ-
ity was even 15.5 times higher in the Sava River. However,
in the study of Krca et al. (2007), where a set of biomarkers
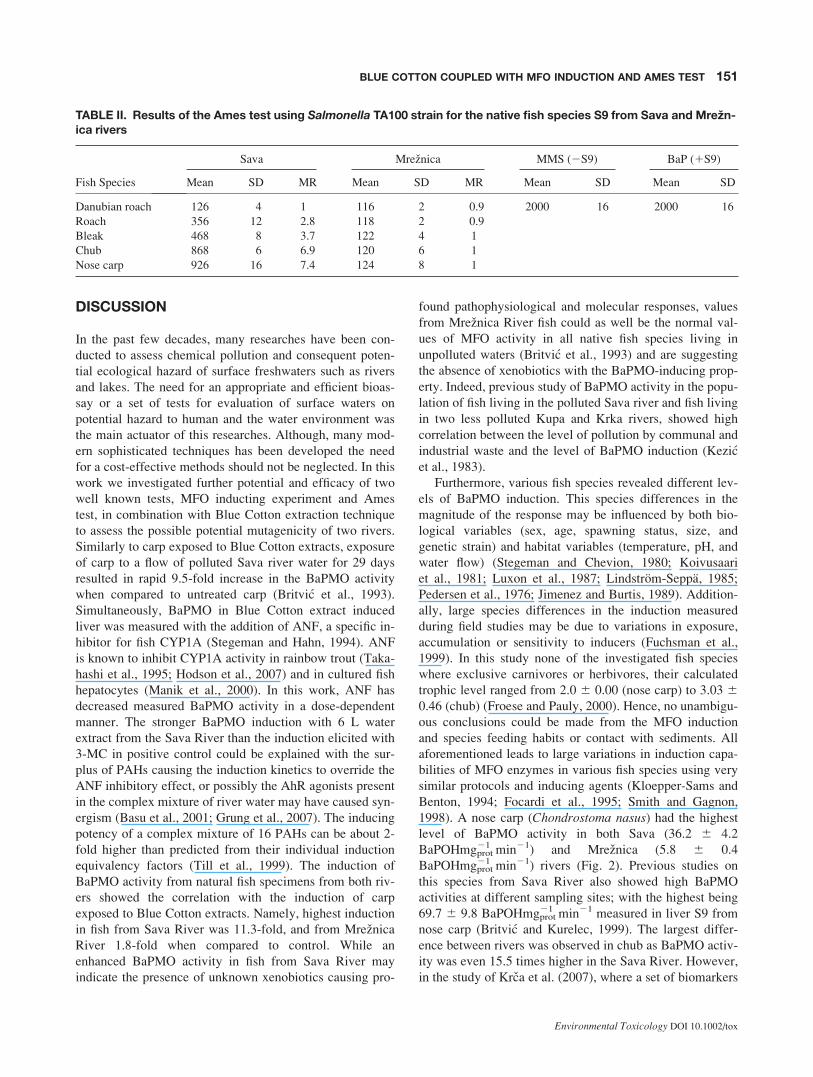
TABLE II. Results of the Ames test using Salmonella TA100 strain for the native fish species S9 from Sava and Mrezn-ica rivers
Fish Species
Sava Mreznica MMS (2S9) BaP (1S9)
Mean SD MR Mean SD MR Mean SD Mean SD
Danubian roach 126 4 1 116 2 0.9 2000 16 2000 16
Roach 356 12 2.8 118 2 0.9
Bleak 468 8 3.7 122 4 1
Chub 868 6 6.9 120 6 1
Nose carp 926 16 7.4 124 8 1
151BLUE COTTON COUPLED WITH MFO INDUCTION AND AMES TEST
Environmental Toxicology DOI 10.1002/tox

were measured in chub, this species was found to be a valu-
able indicator of environmental pollution. A correlation
was found for the results of Ames test and BaPMO mea-
surement in native fish specimens from Sava River (R 50.96) (Fig. 3). When liver S9 from native fish specimens
was used in the Ames test, only Danubian roach from Sava
River increased its potential to bioactivate BaP less than a
twofold. And this species also had the lowest BaPMO activ-
ity induction in the MFO inducting experiment. On the
other hand, no correlation was found for the results from
Mreznica River (R 5 0.62) (Fig. 3). All tested fish species
from the Mreznica River had MR around one, having
almost the same number of revertants as the number of
spontaneous revertants. These results are in accordance
with previously conducted experiments with native fish
specimens from Sava and Korana Rivers (Britvic et al.,
1993). In the experiment with carps the number of revertants
increased proportionally with the volume of river water in
the Blue Cotton extract from Sava River, and from Mreznica
River the number of revertants increased only with 6 L
extract. The liver potential to bioactivate BaP using the TA
100 strain was increased fourfold in the experiment with
carp exposed to Sava River water for 29 days (Britvic et al.,
1993). All three calculated MR’s for Sava River were higher
than two (3, 5, and 9, respectively). Even though the MR
from two separate experiments testing Sava River water
were greater than two, suggesting the positive mutagenic
activity (Fujimoto et al., 2007; Umbuzeiro et al., 2007), other
experiments with 32P-postlabeling analyses of DNA adducts
in chub, bream and barbel failed to show any pollution-
related DNA adducts (Kurelec et al., 1989).
CONCLUSIONS
The results of this investigation demonstrate that the inte-
gration of used bioassays with Blue Cotton extraction tech-
nique may provide an effective water monitoring tool. Even
though this experiment does not represent environmentally
realistic exposure scenario, measured biological effects
show that the extracted substance that caused the effect
would certainly caused it in such a scenario. This combina-
tion of extraction and biological endpoints can affect which
analytical techniques will be used for chemical analysis
and, consequently, the cost of preparation of fraction for
testing. The investigations and existence of cost-effective
methods for determination of wastewater mutagenic poten-
tial are important particularly for the developing countries.
Additionally, as in ecotoxicological studies it is essential
to assess the toxic response of indigenous fauna as indicator
species or sentinels of environmental contamination this
experiment demonstrated that the MFO-sensitivity, i.e.,
inducibility, is species-dependent.
From the results of this experiment it is evident that the
inducibility gradation was equal in the same species from
differently polluted sites (Fig. 3). Consequently, it implies
the possibility of freshwater monitoring of mutagenic com-
pounds in the sense of MFO induction comparison between
tested and reference location.
Finally, since the drinking water supply in the Sava ba-
sin relies almost exclusively on the rich resources of high-
quality groundwater, which are under the direct influence
of the Sava River (Kallqvist et al., 2008) it is important to
have easily conductible, reliable and interpretable biomoni-
toring methods.
REFERENCES
Basu N, Billiard S, Fragoso N, Omoike A, Tabash S, Brown S,
Hodson P. 2001. Ethoxyresorufin-O-deethylase induction in
trout exposed to mixtures of polycyclic aromatic hydrocarbons.
Environ Toxicol Chem 12:1244–1251.
Britvic S, Kurelec B. 1986. Selective activation of carcinogenic
aromatic amines to bacterial mutagens in the marine mussel
Mytilus galloprovincialis. Comp Biochem Physiol 85:111–114.
Britvic S, Kurelec B. 1999. The effect of inhibitors of multixeno-
biotic resistance mechanism on the production of mutagens by
Dreissena polymorpha in waters spiked with premutagens.
Aquatic Toxicol 47:107–116.
Britvic S, Lucic D, Kurelec B. 1993. Bile fluorescence and some
early biological effects in fish as indicators of pollution by
xenobiotics. Environ Toxicol Chem 12:765–773.
Buhler DR, Wang-Buhler J-L. 1998. Rainbow trout cytochrome
P450s: Purification, molecular aspects, metabolic activity,
induction and role in environmental monitoring. Comp Bio-
chem Physiol 121:107–137.
Dearfield KL, Cimino MC, McCarroll NE, Mauer I, Valcovic LR.
2002. Genotoxicity risk assessment: A proposed classification strat-
egy. Mutat Res Genet Toxicol Environ Mutagen 521:121–135.
Environmental Health—Croatian Country Report. 1993. Work-
shop on Water Related Environmental Health Aspects in the
Catchment Area of Danube. Croatian Institute of Public Health,
Zagreb. pp 1–53.
Focardi S, Fossi MC, Lari L, Casini S, Leonzio C, Meidel SK,
Nigro M. 1995. Induction of MFO activity in the antarctic fish
Pagothenia bernacchii: Preliminary results. Mar Environ Res
39:97–100.
Froese R, Pauly D, editors. 2000. FishBase 2000: Concepts,
Design and Data Sources. Los Banos, Laguna, Philippines:
ICLARM. pp 344.
Fuchsman PC, Duda DJ, Barber TR. 1999. A model to predict
threshold concentrations from toxic effects of chlorinated ben-
zenes in sediment. Environ Toxicol Chem 18:2100–2109.
Fujimoto T, Tsuchiya Y, Shibuya N, Taiyoji M, Nishiwaki T,
Nakamura K, Yamamoto M. 2007. Contamination of the Shi-
nano river water with mutagenic substances after the Niigata
Chuetsu earthquake. Tohuku J Exp Med 211:171–180.
Grung M, Lichtenthaler R, Ahel M, Tollefsen K-E, Langford K,
Thomas KV. 2007. Effects-directed analysis of organic toxi-
cants in wastewater effluent from Zagreb. Croatia. Chemo-
sphere 67:108–120.
152 BRITVIC ET AL.
Environmental Toxicology DOI 10.1002/tox

Hayatsu H. 1992. Cellulose bearing covalently linked copper
phthalocyanine trisulphonate as an adsorbent selective for poly-
cyclic compounds and its use in studies of environmental muta-
gens and carcinogens. J Chromatogr 597:37–56.
Hayatsu H, Oka T, Wakata A, Ohara Y, Hayatsu T, Kobayashi H,
Arimoto S. 1983. Adsorption of mutagens to cotton bearing co-
valently bound trisulfo-copper-phthalocyanine. Mutat Res
119:233–238.
Hayatsu H, Hayatsu T, Wataya Y, Mower HF. 1985. Fecal muta-
genicity arising from ingestion of fried ground beef in the
human. Mutat Res 143:207–211.
Hodson PV, Qureshi K, Noble CAJ, Akhtar P, Brown RS. 2007.
Inhibition of CYP1A enzymes by [a]-naphthoflavone causes
both synergism and antagonism of retene toxicity to rainbow
trout (Oncorhynchus mykiss). Aquatic Toxicol 81:275–285.
Jimenez BD, Burtis LS. 1989. Influence of environmental varia-
bles on the hepatic mixed-function oxidase system in bluegill
sunfish, Lepomis machrochirus. Comp Biochem Physiol 93:11–
21.
Kallqvist T, Milacic R, Smital T, Thomas KV, Vranes S, Tollefsen
K-E. 2008. Chronic toxicity of the Sava River (SE Europe)
sediments and river water to the algae Pseudokirchneriella sub-capitata. Water Res 42:2146–2156.
Kataoka H, Hayatsu T, Hietsch G, Steinkellner H, Nishioka S,
Narimatsu S, Knasmuller S, Hayatsu H. 2000. Identification of
mutagenic heterocyclic amines (IQ. Trp-P-1 and A alpha C) in
the water of the Danube River. Mutat Res 466:27–35.
Kezic N, Britvic S, Simmons JE, Rijavec M, Zahn RK, Kurelec B.
1983. Activity of benzo(a)pyrene monooxygenase in fish from
Sava river. Sci Total Environ 27:59–69.
Kira S, Hayatsu H, Ogata M. 1989. Detection of mutagenicity in
mussels and their ambient water. Bull Environ Contam Toxicol
43:583–589.
Kloepper-Sams PJ, Benton E. 1994. Exposure of fish to biologi-
cally treated bleacked-kraft effluent. II. Induction of hepatic
cytochrome P4501A in mountain whitefish (Prospium william-soni) and other species. Environ Toxicol Chem 13:1483–1496.
Kobayashi H, Hayatsu H. 1984. A time-course study on the muta-
genicity of smoker’s urine. Gann 75:489–493.
Koivusaari U, Harri M, Hanninen O. 1981. Seasonal variation of
hepatic biotransformation in female and male rainbow trout
(Salmo gairdneri). Comp Biochem Physiol 70:149–157.
Krca S, Zaja R, Calic V, Terzic S, Grubesic MS, Ahel M, Smital
T. 2007. Hepatic biomarker responses to organic contaminants
in feral chub (Leuciscus cephalus)—Laboratory characteriza-
tion and field study in the Sava river. Croatia. Environ Toxicol
Chem 26:2620–2633.
Kummrow F, Rech CM, Coimbrao CA, Roubicek DA, Umbuzeiro
G de A. 2003. Comparison of the mutagenic activity of XAD4
and blue rayon extracts of surface water and related drinking
water samples. Mutat Res 541:103–113.
Kurelec B, Krca S. 1989. Glucuronides in mussel Mytilus gallo-provincialis as a possible biomonitor of environmental carcino-
gens. Comp Biochem Physiol 92(C):371–376.
Kurelec B, Matijasevic Z, Rijavec M, Alaccevic M, Britivic S,
Muller WEG, Zahn RK. 1979. Induction of benzo(a)pyrenemonooxygenase in fish and the Salmonella test as tool for
detecting mutagenic/carcinogenic xenobiotics in the aquatic
environment. Bull Environ Contam Toxicol 21:799–807.
Kurelec B, Protic M, Britvic S, Kezic N, Rijavec M. 1981. Toxic
effects in fish and mutagenic capacity of water from the Sava
River in Yugoslavia. Bull Environ Contam Toxicol 26:179–
187.
Kurelec B, Protic M, Rijavec M, Britvic S, Muller WEG, Zahn
RK. 1982. Induction of benzo(a)pyrene monooxygenase in fish
after ip application of water hexane extract—A prescreen tool
for detection of xenobiotics. In: Richards NL, Jackson BL, edi-
tors. Carcinogenic Polynuclear Aromatic Hydrocarbons in the
Marine Environment. Gulf Breeze, Florida: EPA-600/9–82-023.
pp 124–136.
Kurelec B, Garg A, Krca S, Gupta RC. 1989. A natural environ-
ment surpasses polluted in inducing DNA damage in fish. Car-
cinogenesis 10:1337–1339.
Lindstrom-Seppa P. 1985. Seasonal variation of the xenobiotic
metabolizing enzyme activities in the liver of male and female
vendace Coregonus albula L. Aquatic Toxicol 6:323–331.
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. 1951. Protein
measurement with the Folin phenol reagent. J Biol Chem 193:
265–275.
Lu Y, Wang Z, Huckins J. 2002. Review of the background and
application of triolein-containing semipermeable membrane
devices in aquatic environment. Aquatic Toxicol 60:139–153.
Luxon PL, Hodson PV, Borgmann U. 1987. Hepatic aryl hydro-
carbon hydroxylase: Activity of lake trout (Salvelinus namay-cush) as an indicator of organic pollution. Environ Toxicol
Chem 6:649–657.
Manik C, Ghosh RG, Arun KR. 2000. Induction of CYP1A by car-
bofuran in primary culture of fish hepatocytes. J Biochem Mol
Toxicol 14:204–209.
Monarca S, Richardson SD, Feretti D, Grottolo M, Thruston AD
Jr, Zani C, Navazio G, Ragazzo P, Zerbini I, Alberti A. 2002.
Mutagenicity and disinfection by-products in surface drinking
water disinfected with peracetic acid. Environ Toxicol Chem
21:309–318.
Nagai A, Kano Y, Funasaka R, Nakamuro K. 2002. Mutagenic
characteristics and contribution of polycyclic aromatic hydro-
carbons to mutagenicity of concentrates from municipal river
water by blue chitin column. J Health Sci 48:232–241.
Nebert DW, Gelboin HV. 1968. Substrate-inducible microsomal
aryl hydroxylase in mammalian cell culture. J Biol Chem
243:6242–6249.
Ohe T, White PA, DeMarini DM. 2003. Mutagenic characteristics
of river waters flowing through large metropolitan areas in
North America. Mutat Res 534:101–112.
Ohe T, Watanabe T, Wakabayashi K. 2004. Mutagens in surface
waters: A review. Mutat Res 567:109–149.
Parrott JL, van den Heuvel MR, Mark Hewitt L, Baker MA, Mun-
kittrick KR. 2000. Isolation of MFO inducers from tissues of
white suckers caged in bleached kraft mill effluent. Chemo-
sphere 41:1083–1089.
Pedersen MG, Hershberger WK, Zachariah PK, Juchau MR. 1976.
Hepatic biotransformation of environmental xenobiotics in six
strains of rainbow trout (Salmo gairdneri). J Fish Res Board
Can 33:666–675.
153BLUE COTTON COUPLED WITH MFO INDUCTION AND AMES TEST
Environmental Toxicology DOI 10.1002/tox

Protic-Sabljic M, Kurelec B. 1983. High mutagenic potency of
several polycyclic aromatic hydrocarbons induced by liver post-
mitochondrial fractions from control and xenobiotics treated
immature carp. Mutat Res 118:177–189.
Rijavec M, Britvic S, Protic M, Kurelec B. 1981. Detection of the
presence of xenobiotics in seawater samples from Rijeka Bay
applying benzo(a)pyrene monooxygenase induction. Thalassia
Jugosl 17:245–250.
Sakamoto H, Hayatsu H. 1990. A simple method for monitoring
mutagenicity of river water mutagens in Yodo river system.
Kyoto-Osaka. Bull Environ Contam Toxicol 44:521–528.
Sayato Y, Hayatsu H. 1990. A simple method for monitoring
mutagenicity of river water. Mutagens in Yodo River system,
Kyoto-Osaka. Bull Environ Contam Toxicol 44:521–528.
Sayato Y, Nakamuro K, Ueno H, Goto R. 1993. Identification of
polycyclic aromatic hydrocarbons in mutagenic adsorbates to a
copper-phthalocyanine derivative recovered from municipal
river water. Mutat Res 300:207–213.
Schenck KM, Wymer LJ, Lykins Jr BW, Clark RM. 1998. Appli-
cation of a Finnish mutagenicity model to drinking waters in
the US. Chemosphere 37:451–464.
Skog K. 2004. Blue Cotton. Blue Rayon and Blue Chitin in the
analysis of heterocyclic aromatic amines—A review. J Chroma-
togr B 802:39–44.
Smith BJ, Gagnon MM. 1998. MFO induction of three Australian
fish species. Environ Toxicol 15:1–7.
Stegeman JJ, Chevion M. 1980. Sex differences in cytochrome p-
450 and mixed-function oxygenase activity in gonadally mature
trout. Biochem Pharmacol 29:553–558.
Stegeman JJ, Hahn ME. 1994. Biochemistry and molecular biol-
ogy of monooxygenases: Current perspectives on forms, func-
tion, and regulation of cytochrome P450 in aquatic species. In:
Malins DC, Ostrander GK, editors. Aquatic Toxicology: Molec-
ular, Biochemical and Cellular Perspectives. Boca Raton, Flor-
ida: Lewis. pp 87–174.
Takahashi N, Miranda CL, Henderson MC, Buhler DR, Williams
DE, Bailey GS. 1995. Inhibition of in vitro aflatoxin B1-DNA
binding in rainbow trout by CYP1A inhibitors: [a]-naphthofla-vone, [b]-naphthoflavone and trout CYP1A1 peptide antibody.
Comp Biochem Physiol 110:273–280.
Till M, Riebniger D, Schmitz H-J, Schrenk D. 1999. Potency of var-
ious polycyclic aromatic hydrocarbons as inducers of CYP1A1
in rat hepatocyte cultures. Chem Biol Interact 117:135–150.
Umbuzeiro GA, Roubicek DA, Rech CM, Sato MIZ, Claxton LD.
2004. Investigating the sources of the mutagenic activity found
in a river using the Salmonella assay and different water extrac-
tion procedures. Chemosphere 54:1589–1597.
Umbuzeiro GA, Coimbrao CA, Kummrow F, Lobo DJA, Saldiva
PHN. 2007. Mutagenic activity assessment of Cristais river.
Sao Paulo, Brazil, using the blue rayon/Salmonella microsome
and the Tradescantia pallida micronuclei assays. J Braz Soc
Ecotoxicol 2:163–171.
Vouk D, Malus D. 2005. Impact of the Zagreb wastewater treat-
ment and disposal on sava river. In: Jugovic CJ, editor. Water
Management and Hydraulic Engineering. Wien, Austria: Nacht-
nebel. pp 323–332.
Watanabe T, Takahashi Y, Takahashi T, Nukaya H, Terao Y, Hir-
ayama T, Wakabayashi K. 2002. Seasonal fluctuation of the muta-
genicity of river water in Fukui, Japan, and the contribution of
2-phenylbenzotriazole-type mutagens. Mutat Res 519:187–197.
White PA, Rasmussen JB. 1998. The genotoxic hazards of domes-
tic wastes in surface waters. Mutat Res 410:223–236.
154 BRITVIC ET AL.
Environmental Toxicology DOI 10.1002/tox
Copyright © 2022 FDOKUMEN




















