
Дерматоглифика в антропологии. СПб.: Нестор-история,...
379
Г.Л. Хить, И.Г. Широбоков, И.А. Славолюбова ДЕРМАТОГЛИФИКА В АНТРОПОЛОГИИ
-
Upload
kunstkamera -
Category
Documents
-
view
2 -
download
0
Transcript of Дерматоглифика в антропологии. СПб.: Нестор-история,...

Г.Л. Хить, И.Г. Широбоков, И.А. Славолюбова
ДЕРМАТОГЛИФИКА
В АНТРОПОЛОГИИ

Электронная версия книги подготовлена и
отредактирована ее авторами
с учетом опечаток, выявленных
в печатном варианте
*

РОССИЙСКАЯ АКАДЕМИЯ НАУК
ИНСТИТУТ ЭТНОЛОГИИ И АНТРОПОЛОГИИ им. Н.Н. МИКЛУХО-МАКЛАЯ
Г.Л. Хить, И.Г. Широбоков, И.А. Славолюбова
ДЕРМАТОГЛИФИКА
В АНТРОПОЛОГИИ
Нестор-ИсторияСанкт-Петербург
2013

УДК 572.08+572.2 ББК 28.71
X52
Рецензенты д.б.н. М.А. Негашева,
к.и.н. Н.А. Лейбова
Книга утверждена к печати на заседании Ученого совета ИЭА РАН
Ответственный редактор И.Г. Широбоков
Х52 Хить Г.Л., Широбоков И.Г., Славолюбова И.А.Дерматоглифика в антропологии / отв. ред. И.Г. Широбоков - СПб.: Нестор- история, 2013. - 376 с.ISBN 978-5-4469-0184-5
В книге рассмотрены различные аспекты исследований антропологии популяций и рас по дерматоглифическим данным. Специальные разделы работы посвящены методическим аспектам дерматоглифики: описаны морфологические и расово-диагностические свойства признаков, в том числе впервые предлагаемых к изучению, определено их таксономическое значение для дифференциации популяций различного уровня, описаны получившие широкое распространение в антропологии и оригинальные статистические методы популяционного анализа дерматоглифических данных. В отдельных главах рассмотрены результаты анализа серий по признакам одной руки, возможности применения данных дерматоглифики к археологическим материалам, межрасовые вариации половых различий признаков кожного рельефа, соотношение картин популяционной дифференциации, полученных в рамках разных систем антропологических признаков.
УДК 572.08+572.2 ББК 28.71
ISBN 978-5-4469-0184-5
© Г.Л. Хить, 2013 © И.Г. Широбоков, 2013 © И.А. Славолюбова, 2013

СОДЕРЖАНИЕ
Введение (Г.Л. Хить, И.Г. Широбоков)
Очерк истории исследований дерматоглифики рас и популяций (И.Г. Широбоков)
Раздел I. Методические и методологические основы анализа
Глава 1. Эмбриогенез и генетика гребешковой кожи кисти (И.А. Славолюбова)
Глава 2. Методика получения отпечатков, описания признаков и первичной статистической обработки данных (И.А. Славолюбова)
Глава 3. Методы статистического анализа дерматоглифических данных (Г.Л. Хить, И.Г. Широбоков)
Глава 4. Система корреляций элементов дерматоглифики. Выделение группы ключевых признаков (Г.Л. Хить)
Глава 5. Радиальная ориентация пальцевых узоров и ее значение для расовой дифференциации (И.Г. Широбоков)
Глава 6. Половые различия признаков дерматоглифики (Г.Л. Хить)
Глава 7. Палеодерматоглифика (И.Г. Широбоков)
Раздел II. Расовые, этнические и популяционные аспекты дерматоглифических исследований
Глава 8. Внутрирасовое распределение признаков дерматоглифики и их комплексов. Ключевые признаки и их свойства. Расовые градиенты и расовые комплексы (Г.Л. Хить)
8
12
24
40
68
78
92
112
158
170
3

Глава 9. Характеристика и таксономия больших рас и их подразделений (Г.Л. Хить)
Глава 10. Бимануальное распределение и асимметрия дерматоглифических признаков: популяционный аспект (И.Г. Широбоков)
Глава 11. Дерматоглифика и исторические процессы(Г.Л. Хить, И.Г. Широбоков)
Глава 1 2. Соотношение данных дерматоглифики и других систем антропологических признаков (Г.Л. Хить, И.Г. Широбоков)
Заключение (Г.Л. Хить, И.Г. Широбоков)
Библиография
Summary
Приложения
I. Heet H.L. Abstract of a thesis «Dermatoglyphics of the USSR peoples»
II. Список дерматоглифических серий, находящихся на хранении в ИЭА РАН и МАЭ РАН
Список сокращений
254
274
288
308
318
342
350
364
374
222
4

CONTENTS
Introduction (H.L. Heet, I.G. Shirobokov)
Historical outline of the studies in racial and population dermatoglyphics (I. G. Shirobokov)
Part I. Methodical and methodological approaches of the research
Chapter 1. Ridged skin of hand: Embryology and genetics (I.A. Slavoljubova)
Chapter 2. Methods of printing, description and elementary statistical analysis (I.A. Slavoljubova)
Chapter 3. Methods of multivariate statistical analysis for dermatoglyphic data (H.L. Heet, I. G. Shirobokov)
Chapter 4. Correlation analysis of dermatoglyphic elements. Selection of the set of key traits (H.L. Heet)
Chapter 5. Radial orientation of finger patterns and its value for differentiation of races (I.G. Shirobokov)
Chapter 6. Sexual differences in dermatoglyphics (H.L. Heet)
Chapter 7. Paleodermatoglyphics (I. G. Shirobokov)
Part II. Racial, ethnic and population aspects of dermatoglyphic studies
Chapter 8. Intra-racial distribution of dermatoglyphic traits and complexes. Key traits and their characteristics. Racial gradients and racial complexes (H.L. Heet)
Chapter 9. Characteristics and taxonomy of major races and their subdivisions (H.L. Heet)
Chapter 10. Bimanual distribution and asymmetry of dermatoglyphic traits: Inter-population research(I.G. Shirobokov)
24
40
68
78
92
112
158
170
222
8
12
254
5

Chapter 11. Dermatoglyphics and historical studies (H.L. Heet, I.G. Shirobokov) 274
Chapter 12. Relations between dermatoglyphics and other branches of physical anthropology (H.L. Heet,I.G. Shirobokov) 288
Conclusion (H.L. Heet, I.G. Shirobokov) 308
Bibliography 318
Summary (in English) 342
Appendix
I. Heet H.L. Abstract of the thesis «Dermatoglyphics of thepeoples of the USSR» (in English) 350
II. List of dermatoglyphic collections hold in IEA RAS andMAE RAS 364
List of abbreviations 374
6

%

«Природа держит нас на почтительном расстоянии от всех ее тайн и уступает нам лишь сведения о немногих поверхностных качествах объектов, скрывая от нас те силы и законы, которыми всецело определяются действия этих объектов»
Дэвид Юм
Введение
Современная антропология отличается широким диапазоном направлений (эволюционная антропология, морфология, физиология различных возрастных, половых, профессиональных и др. групп человечества, антропогенетика, расоведение, этническая и популяционная антропология и др.). Признаки, описывающие кожные узоры кисти и стопы, составляют емкую информационную систему, которая имеет выход не только в антропологию, но также и во многие другие области знания и практики (эмбриология, медицина, в т.ч. клиническая, анатомия, криминалистика, спорт, судебная экспертиза). Все эти аспекты, имеющие свою историю и достижения, оставлены за рамками данной работы.
Название книги, представленной вниманию читателя, по не зависящим от авторов обстоятельствам оказалось значительно шире ее содержания. В ней, помимо ряда общих вопросов, изложены результаты многолетнего изучения рельефа папиллярных узоров кисти в расовом, этническом и популяционном аспектах.
Начало изучения кожных узоров кисти у различных групп населения мира относится к концу XIX в. и связано с именами Артура Колльмана и Фрэнсиса Гальтона. Однако и к началу XXI века специалистам в области дерматоглифики из разных стран так и не удалось прийти к общему мнению относительно программы исследований и выработать единый методический подход, позволяющий использовать необходимые данные из публикаций коллег. Наибольшей популярностью в настоящее время пользуется программа, характеризующая почти исключительно папиллярные узоры подушечек пальцев. Как правило, она включает в себя вычисление частот основных типов пальцевых узоров и гребневой счет. Признаки ладони в большинстве работ выпадают из поля зрения специалистов либо сводятся к расчетам гребневого счета на
8

разных участках. Вместе с сужением программы исследований, в дерматоглифике произошло общее сокращение числа работ, посвященных популяционной и этногенетической проблематике. Большинство работ, касающихся этих тем, отличается узким подходом к изучению материала, что, в первую очередь, выражается в использовании небольшого в территориальном отношении круга сравнительных серий. Такой подход не позволяет рассматривать данные в историческом контексте, вынуждая исследователей фактически ограничиваться описанием выявленных между сериями морфологических различий.
В книге описана система методических приемов, хорошо зарекомендовавших себя в работах по этнической и расовой дерматоглифике отечественных специалистов на всех этапах исследования, рассмотрена проблема дифференциации популяций разного таксономического уровня, проанализированы возможности применения дерматоглифики в изучении человеческих популяций в разных аспектах. В целом этот круг проблем может быть обозначен как дерматоглифические исследования в расоведении. Такая формулировка обязывает подробнее остановиться на понятии терминов «расоведение» и «раса», принятых авторами. Во второй половине XX века в силу известных социально-исторических причин (в первую очередь, идеологизации биологических различий между группами людей и установления искусственных связей между правами человека и его происхождением) в европейской и американской физической антропологии термин «расоведение» обрел исключительно негативные коннотации, вероятно, даже более негативные, нежели «раса». Оба термина практически вышли из употребления в зарубежной научной антропологической литературе.
В практике российских антропологов под «расоведением» традиционно понимается раздел физической антропологии,изучающий проблемы классификации человеческих рас и популяций, их состава и истории формирования, а также влияния на эти процессы различных факторов. В «Антропологическомсловаре», подготовленном коллективом ведущих московских антропологов, расоведение кратко определяется как «разделантропологии, посвященный всестороннему изучению рас человека» (Антропологический словарь, 2003, с.232). Под расой в свою очередь понимается «общебиологический термин, используемый систематиками для обозначения внутривидовых систем популяций, обладающих достаточным фенотипическим и генетическим
9

сходством, позволяющим отличить их от других подобных систем» (там же, с.231). Раса является характеристикой совокупности популяций, а не индивидуумов, системой, открытой для обмена генами с другими совокупностями; расовые признаки не являются абсолютными и меняются с течением времени (Бунак, 1938; Проблема расы..., 2002).
Исследование дерматоглифических материалов базируется на статистическом анализе признаков, значительная часть которых имеет дискретную природу. Пальцевые и ладонные узоры не позволяют устанавливать расовую принадлежность человека; индивидуальная вариабельность дерматоглифических признаков у человека исключительно велика. Однако различия в частотах признаков в популяциях разного ранга (от локальных выборок до больших рас) позволяют исследователям ставить и решать обратную задачу - определять предположительную степень генетического родства и различий между человеческими общностями разных таксономических уровней.
Дифференцирующая способность популяционной и расовой дерматоглифики, проанализированная в данной книге, базируется на результатах изучения:
1. группы взаимонезависимых признаков, высокое таксономическое значение которых для разграничения групп разного происхождения и таксономического ранга для различных уровней сопоставления было показано в серии специальных исследований;
2. исторически сложившихся, выявленных на обширном материале комплексов признаков в различных этнических и расовых выборках мира;
3. дивергенции расовых групп, этносов и популяций под воздействием исторических, географических и прочих факторов, определивших к настоящему времени степень сходства и различий таксономических единиц разного ранга.
В данной работе впервые на русском языке после выхода в свет монографии Т.Д. Гладковой (1966) предлагаются и описываются дерматоглифические методы анализа в расовом и популяционном аспектах для всех этапов работы - от методов снятия отпечатков и до приемов анализа дерматоглифических данных. Основной целью авторов является демонстрация спектра возможностей применения дерматоглифики в современных исследованиях расоведческого направления. Авторы обобщили
10

основные результаты исследований отечественных и ряда зарубежных специалистов. В книге рассмотрены дифференциация рас и их подразделений, проблема половых различий признаков, возможности применения дерматоглифики при изучении археологических объектов, связи между признаками кожного рельефа и признаками других антропологических систем, дифференцирующая способность дерматоглифических признаков, ранее не привлекавших внимание антропологов, и ряд других вопросов биологического и исторического характера.
В Приложение включен англоязычный вариант автореферата книги Г.Л. Хить «Дерматоглифика народов СССР», вышедшей 30 лет назад (Хить, 1983). Опубликованные в ней данные неоднократно использовались зарубежными исследователями при подготовке публикаций, нередко, к сожалению, без ссылок на полученные автором сходные выводы. Публикация англоязычного варианта автореферата позволит зарубежным коллегам получить представление об исследовании в широком масштабе дерматоглифики населения Северной Евразии и соседних областей.
Заключает книгу список дерматоглифических коллекций, находящихся на хранении в Институте этнологии и антропологии им. Н.Н. Миклухо-Маклая РАН (Москва) и Музее антропологии и этнографии им. Петра Великого (Кунсткамера) РАН (Санкт- Петербург). Заинтересованный специалист может вступить в деловые и научные контакты с руководством указанных учреждений и получить возможность работать с этими сериями, часть из которых переведена в цифровой вариант.
Авторы выражают глубокую признательность коллективу отдела физической антропологии ИЭА РАН за стимулирующее обсуждение предварительного текста монографии,A.О. Афанасьевой за помощь в подготовке материалов, Е.В. Дороничевой за переводы частей текста на английский язык, Б.А. Козинцеву за разработку статистических программ для анализа дерматоглифических данных, А.Г. Козинцеву за ценные замечания к тексту книги, завотделом антропологии МАЭ РАНB.И. Хартановичу за предоставленную возможность многократных и продолжительных командировок И.Г. Широбокова в Москву для работы над книгой с соавторами.
11

Очерк истории дерматоглифических исследований рас и популяций.
Систематическое изучение дерматоглифических особенностей различных по происхождению популяций началось немногим более ста лет назад. Однако знание о том, что отпечатки пальцевых узоров могут быть использованы для идентификации личности, применялось в древнем Китае уже в 7 в. н.э. Давнюю историю имеет использование отпечатков пальцев в качестве личной подписи также в Индии и Японии (Гейндль, 1927). Развитие расовой дерматоглифики тесно связано с всплеском интереса к изучению антропологии рас в целом и было невозможно без понимания сложного характера формирования расовых признаков.
Предпосылками для развития дерматоглифики стали исследования XVII-XVIII вв. в области анатомии, переживавшей в это время период подъема. Этот этап связан с именами англичанина Нейемии Грю, немца Говарда Бидло, итальянца Марчелло Мальпиги, а также Якоба Гинце и Альбинуса, описавших особенности кожного рельефа и строение узоров на пальцах и ладонях человека. В начале XIX в. чешский анатом Ян Пуркинье предложил первую в Европе классификацию пальцевых узоров, включающую в себя девять основных типов, и дал каждому из них специальное название.
С 80-х гг. XIX века на первый план выступает изучение отпечатков пальцевых узоров для криминалистической экспертизы (Торвальд, 1991). Центром исследований кожного рельефа кисти становится Азия. Англичанин Уильям Гершель, длительное время работавший в Калькутте в качестве чиновника Ост-Индийской компании, начал использовать отпечатки пальцев для идентификации личности. Фактически он объявил себя автором метода дактилоскопии, хотя, по всей вероятности, техника была им заимствована, поскольку уже практиковалась среди индийцев (Гейндль, 1927). По инициативе Гершеля почти в течение трех десятилетий дактилоскопия с успехом применялась при выплате пенсий, а также для идентификации преступников в одной из тюрем. Однако его предложение ввести дактилоскопию по всей Бенгалии было отвергнуто властями. В статье, опубликованной в 1880 г. в
12

журнале «Nature», Гершель справедливо указывал, что отпечатки не позволяют отличать мужчин от женщин, а также индийцев от европейцев (Herschel, 1880).
В том же году в «Nature» вышла заметка другого англичанина, Генри Фолдса, проживавшего в Японии и занявшегося изучением отпечатков папиллярных узоров после того, как его внимание привлекли оттиски ногтевых фаланг пальцев на древней керамике (Faulds, 1880). Фолдс указал, что отпечатки пальцев уникальны для каждого человека и, будучи оставленными на месте преступления, могут быть использованы для выявления преступника. В отличие от Гершеля он считал необходимым снимать отпечатки всех 10 пальцев. Он также отметил, что отпечатки пальцев обезьян очень близки отпечаткам пальцев человека, и предположил, что их изучение может пролить свет на вопрос об их родственных связях. Фолдс собрал отпечатки японцев и представителей других этнических групп с целью их сравнения, но результаты так и не опубликовал. Как Гершель, так и Фолдс интересовались антропологическими аспектами изучения папиллярных узоров в той степени, в какой это могло помочь идентификации личности. По всей видимости, убедившись в невозможности определения этнической и расовой принадлежности человека по пальцевым узорам, они не развивали эту тему в дальнейшем. Практического значения работы Гершеля и Фолдса так и не получили. Еще в начале XX века в судах данные дактилоскопической экспертизы не признавались в качестве надежного источника информации (Гейндль, 1927).
Определенный вклад в историю дерматоглифики внесла вышедшая в свет в 1883 г. книга Артура Колльмана «Осязательный аппарат руки человеческих рас и обезьян». Этот автор занимался изучением эмбрионального развития папиллярных узоров и пришел к выводу, что узоры закладываются на четвертом и полностью формируются к шестому месяцу внутриутробного развития плода человека (Galton, 1892, p.58). Колльмана можно считать одним из первых исследователей, которые целенаправленно занимались изучением дерматоглифических различий между основными расами человечества. Следует, правда, оговориться, что его данные не были репрезентативными (по 2-3 индивида из групп китайцев, японцев, турок, армян, австралийцев и мулатов). Несколько большие серии ему удалось получить для узоров на подошвах стоп. Основной
13

целью Колльмана было выявление в папиллярных узорах представителей «низших рас» (как он их понимал) обезьяньих признаков. Никаких определенных результатов в этом отношении он не достиг (Wilder, 1904, p.244-245).
Работы Гершеля и Фолдса обратили на себя внимание выдающегося английского ученого Фрэнсиса Гальтона. Изучение папиллярных узоров интересовало его в первую очередь с позиции антрополога, но итоги его работы сыграли существенную роль в становлении современной дактилоскопии. Он разработал новую классификацию дерматоглифических признаков, в основу которой были положены различия между узорами по количеству трирадиусов. Результаты были опубликованы в книге «Finger prints» в Лондоне в 1892 г. Фактически в ней были заложены основы современной дерматоглифики, обосновывались стабильность и неизменность папиллярных узоров в течение жизни человека, их вариативность и возможность классификации.
Гальтон проанализировал данные об англичанах, валлийцах, евреях, басках и ряде африканских групп и был вынужден отвергнуть мысль о существовании специфических расовых типов узоров. Он также пришел к твердому выводу, что невозможно вынести научно обоснованное суждение об этнической принадлежности человека по его кожным узорам. Гальтон не обнаружил значимых статистических отличий между расами: разница по частотам типов узоров составила величину, не большую, чем между группами, относящимися к одной расе. Некоторые различия (однако, статистически также незначимые) удалось выявить только при анализе больших рас (Galton, 1892, p. 18, 192193). С другой стороны, по мнению Гальтона, африканские группы демонстрируют большую простоту узоров: папиллярные гребешки расположены параллельно чаще, чем у европейцев, и с одинаковой периодичностью (там же, p. 196).
Попытки наметить эволюционную линию развития для различных типов папиллярных узоров часто предпринимались исследователями конца XIX - первой половины XX столетия. Африканские группы a priori рассматривались как наиболее примитивные в этом отношении, европеоидные популяции - как наиболее прогрессивные. Некоторые исследователи полагали, что завитки являются более примитивным узором, чем петли и дуги, поскольку у высших млекопитающих (сумчатых, полуобезьян)
14

фиксируются узоры, сходные с первыми. Данкмейер указал, однако, что завитковые узоры расположены у полуобезьян на межпальцевых подушечках, а не на ногтевых фалангах. У высших приматов и человека завитковые узоры чаще встречаются на пальцах, чем на ладонях. У пигмеев фиксируется наиболее низкий процент завитков. По мнению Данкмейера, данное обстоятельство свидетельствует против того, что завитки являются примитивным типом узора (Dankmeijer, 1938). Фрэнсис Гальтон был приверженцем взглядов на европейца как носителя наиболее прогрессивных генетических признаков. Однако, являясь основателем евгеники, убежденным в зависимости интеллектуальных и психических качеств человека от его наследственных признаков, он так и не смог выстроить «эволюционную» линию для популяций различного происхождения.
Несмотря на то, что попытки популяционных исследований проводились и ранее, родоначальником этнической дерматоглифики считается американский исследователь Гаррис Уайлдер. Он первым указал, что для выявления расовых различий требуются выборки достаточно большого объема (не менее 100 человек) и отметил высокую индивидуальную изменчивость признаков в любой расе. Он первым обратился к систематическому изучению расовых вариаций узоров ладони и подошвенной поверхности стопы, а также ввел в оборот комплекс признаков, получивших название главных ладонных линий. Результаты исследований Уайлдера, основанные на данных изучения отпечатков европейцев, американских индейцев, африканцев, китайцев и японцев, позволили утверждать, что средние величины признаков разграничивают выборки, относящиеся к разным расам, но этнически близкие группы разделить сложно (1904; 1913; 1922). Высокая вариабельность признаков у европеоидов объяснялась исследователем как результат включения в их состав широкого круга различных этнических групп. Обнаружив, что у индейцев майя вариабельность признаков ниже, чем у европеоидов, Уайлдер предположил, что низкая изменчивость свойственна «примитивным» расам. Кроме того, он полагал, что для «чистой» расы должен быть характерна какая-то одна формула ладонных линий с небольшими вариациями, связанными с различиями в пропорциях разных областей ладони (Wilder, 1904, p.291). Такой взгляд на антропологическую характеристику человеческих сообществ находился в соответствии с типологической концепцией расы. Однако подход, при котором
15

каждый индивидуум наделяется всеми отличительными морфологическими признаками своей расы, плохо согласуется с данными дерматоглифики, большая часть признаков которой имеет дискретный характер. Насколько известно авторам, в более поздних работах Уайлдер не пытался развивать эту идею. Вероятно, именно популярность типологической концепции заставляла исследователей даже в 1930-е гг., спустя полвека после выхода в свет статей Гершеля и Фолдса, оговаривать в публикациях, что установление расовой и этнической принадлежности конкретного человека по отпечаткам его пальцев и ладоней невозможно (см. например, Cummins, 1935, p.829). Работы Уайлдера, его коллеги и супруги Инесс Уиппл, а также генетические исследования норвежского ученого Кристины Бонневи определили развитие дерматоглифики в первой четверти XX века (Plato, Garruto, 1990, p.4-5).
Становление дерматоглифики как науки связано с именем Гарольда Камминса, впервые предложившего этот термин в 1926 г. С этого же года получила начало традиция публикаций результатов дерматоглифических исследований в престижном «American Journal of Physical Anthropology». Интерес Камминса к расовым различиям нашел выражение в серии работ по дерматоглифике различных популяций Земного шара: европейцев, американцев, населения Западной Африки, индейцев, австралийцев и др.
Наиболее важным условием для изучения расовых различий, по мнению Камминса, является правильный отбор выборок. Им были сформулированы три критерия для проведения последнего:
1. число индивидов в каждой выборке должно соответствовать величине, статистически значимой для суждения о сходстве или различии;
2. необходимо разделять мужские и женские выборки, поскольку в дерматоглифике существуют половые различия;
3. в маленьких выборках желательно исключать отпечатки родственных индивидов, чтобы на характеристике выборки не сказались специфические черты семейной группы (Cummins, 1935, p. 831).
В ранних работах Камминс находился под влиянием исследований Уайлдера (Mavalwala, 1975). Позднее он выработал собственный подход к изучению дерматоглифических признаков и совместно с Чарльзом Мидло опубликовал основополагающий методический труд «Finger prints, palms and soles» (Cummins, Midlo, 1943; 1961). Описанный в нем метод классификации пальцевых и
16

ладонных узоров и сегодня используется большинством исследователей.
Вместе с тем, в 1960-70-е гг. получает развитие альтернативная топологическая система классификации Л. Пенроза и Д. Лэш, созданная с целью облегчить проведение медикогенетических исследований в дерматоглифике. Эта система построена на допущении, что комплекс папиллярных узоров можно описать двумя наиболее существенными признаками: петлей и трирадиусом (Penrose, 1963; 1965). Система подвергалась критике за то, что не учитывает признаки, охваченные в «классической» программе (Jantz, Chopra, 1983).
Особый вариант классификации пальцевых узоров был предложен Андре де Вилде. Классификация также построена на топологическом принципе, но учитывает детальные различия между узорами. Система включает иерархическую классификацию, содержащую пять основных типов узоров, каждый из которых состоит из несколько подклассов, в общей сложности представленных 90 различными вариантами (De Wilde et al., 1981). Однако в популяционной дерматоглифике эта классификация не получила широкого признания и не может считаться альтернативой классификации Г. Камминса - Ч. Мидло.
В России первые работы, связанные с изучением папиллярных линий, относятся к началу XX века. П.С. Семеновский сопоставил частоты пальцевых узоров у различных народов и фактически положил начало изучению популяционной дерматоглифики. Исследователь собрал огромную (22 тыс. чел.) серию пальцевых отпечатков русских (Семеновский, 1923; 1927). Заслуженнуюизвестность приобрели исследования М.В. Волоцкого, внесшего существенный вклад в изучение проблемы наследования папиллярных узоров и методику исследования отпечатков. Независимо от Г. Камминса им был предложен признак, отражающий суммарное количество дельт в пальцевых узорах рук - дельтовый индекс (иногда называемый в зарубежных статьях индексом Волоцкого) (Волоцкой, 1937a; 1937б).
В 1966 году Т.Д. Гладковой было опубликовано первое на русском языке методическое руководство «Кожные узоры кисти и стопы обезьян и человека». Основная часть книги является кратким переложением содержания монографии Г. Камминса и Ч. Мидло, изданной в 1943 году. Оригинальная часть включает обобщающую сводку некоторых данных по дерматоглифике отдельных народов
17

мира, а также раздел, посвященный особенностям папиллярных узоров у приматов.
Дальнейшее развитие дерматоглифических исследований в России в области расоведения связано преимущественно с именем Г.Л. Хить. Опираясь на собранные в экспедициях материалы многочисленных монголоидных и европеоидных популяций, проживавших на территории СССР, а также на данные других авторов, Г.Л. Хить провела анализ связей между абсолютным большинством дерматоглифических признаков, предлагаемых к изучению в соответствии с классификацией Г. Камминса (за исключением гребневого счета и углов). В результате исследовательницей была выделена группа из шести основных, или ключевых, признаков, не связанных между собой значимыми корреляциями на внутригрупповом уровне, в числе которых: дельтовый индекс, индекс Камминса, одиночный низкий осевой трирадиус ладони, частота истинных узоров на гипотенаре и тенаре, добавочные межпальцевые трирадиусы. По этой программе были рассмотрены обобщенные характеристики трех больших рас (европеоидной, монголоидной и негроидной), а также населения разных регионов СССР, включая многие этнические и локальные группы Евразии (Хить, 1969; 1973;1975; 1983; Прокудина, 1974; Хить, Кейта, 1981).
В 1983 году вышла монография Г.Л. Хить «Дерматоглифика народов СССР». В ее основу легли данные кожного рельефа 614 мужских и женских выборок европеоидного, монголоидного и метисного происхождения (около 54 тыс. чел.), из которых почти 70% составили материалы автора (405 выборок, около 36 тыс. чел.). В книге рассмотрены основные закономерности внутрирасовой и межрасовой дифференциации, географически-популяционная изменчивость признаков и их комплексов в масштабе страны и в историко-географических регионах, дифференциация этносов и локальных рас и другие проблемы. Эта работа, публикация которой была задержана на шесть лет, стимулировала ряд исследований, которые были выполнены в разные годы аспирантами, сотрудниками и стажерами отдела антропологии ИЭ АН СССР (далее ИЭА РАН): Д.О. Ашиловой (Калмыкия), О.Б. Бабаковым (Туркмения), В.И. Богдановой (Ленинград), Н.А. Долиновой (Прокудиной, Никольской, Москва), Л. Кавгазовой (Болгария), Б. Кейта (Мали), И.П. Ладария (Абхазия), К.Б. Сихимбаевой
18

(Казахстан), Л.И. Тегако (Белоруссия), И.Г. Широбоковым (Санкт- Петербург).
За прошедшие четверть века по сравнению с предшествующим периодом опубликовано сравнительно небольшое количество обобщающих работ по расовой и этнической дерматоглифике народов России и сопредельных стран.1 Следует отметить ряд монографических изданий, посвященных отдельным регионам и этническим группам. Большой материал по дерматоглифике белорусов и населения сопредельных территорий собран и систематизирован Л.И. Тегако (1989). Исследованию папиллярных линий кисти у армян посвящена монография Н.Р. Кочар (1989). Дерматоглифика народов Средней Азии подробно изучена Г.К. Ходжайовой (2007): рассмотрена дифференциация попризнакам дерматоглифики на территориальном, этническом и расовом уровнях, выявлены различия в распределении признаков между историко-культурными областями, а также между городским и сельским населением. На основе данных дерматоглифики прослежена этническая история казахов в публикации О. Исмагулова, К. Сихымбаевой, А. Исмагуловой (2007), которые проанализировали материалы по локальным группам и провели сравнение казахов и других тюркоязычных народов. Исследованиям некоторых групп североазиатских монголоидов посвящена монография O.K. Галактионова (2008). Как и зарубежные публикации, указанные работы значительно отличаются между собой по программе и методам анализа.
Интерпретация результатов в дерматоглифике нередко вызывает затруднения из-за того, что характер наследования комплекса кожных узоров, полигенный в своей основе, до сих пор во многом остается неясным. Существенный вклад в изучение генетических основ и морфогенеза кожного рельефа пальцев внесла И.С. Гусева (1989; 2010). В настоящее время модель, предложенная и недавно усовершенствованная исследовательницей, наиболее удачно объясняет механизм наследования пальцевых узоров.
К сожалению, из зарубежных публикаций последних десятилетий почти невозможно извлечь методически сопоставимые сравнительные данные. Исключение составляют частоты типов пальцевых узоров и гребневой счет, которые являются обязательным элементом большинства работ. В 2000-е гг. из
1 Подробнее об отечественных исследованиях в области расовой и этнической дерматоглифики см. в Главе 10.
19

публикаций почти полностью исчезли данные по частотам добавочных межпальцевых трирадиусов и осевого трирадиуса t. Крайне редко публикуются данные по частотам типов окончаний главных линий, на основании которых можно было бы рассчитать приблизительную величину индекса Камминса. Частоты истинных узоров на гипотенаре и тенаре нередко приводятся суммарно со следами узоров, что также не позволяет использовать эти данные в сравнительном анализе.
Очевидно, что редукция программы анализируемых признаков связана с возрастающим влиянием генетики на дерматоглифические исследования. Немалую роль также играют отсутствие общепринятых стандартов при работе с дерматоглифическим материалом, трудности определения и анализа дискретно- варьирующих признаков. Вероятно, именно стремлением к большей точности определений и унификации объясняется преобладание публикаций, включающих количественные показатели (различные варианты гребневого счета), в то время как данные по описательным признакам отступают на второй план. В то же время нельзя не отметить, что ряд «классических» признаков, например, окончания главных ладонных линий и разные варианты осевых трирадиусов, являются искусственными конструкциями, рубрикация которых (в отличие от типов пальцевых узоров) является исключительно результатом договоренности специалистов. Эти признаки исчезают из публикаций, даже несмотря на то, что имеют высокую таксономическую ценность для дифференциации популяций различного ранга и происхождения.
К началу XXI века дерматоглифические исследования в области расоведения постепенно утратили свои позиции, о чем свидетельствует исключительная редкость появления работ соответствующей тематики в ведущих антропологических изданиях мира. Наиболее активно популяционные исследования сегодня проводятся в странах Азии, Латинской Америки, Африки, а также в Испании. В других европейских странах, а также в США наибольшее количество публикаций посвящено генетическим аспектам изучения дерматоглифики, связям папиллярных линий и различных типов заболеваний, которые могут быть использованы при диагностике последних.
К сожалению, в некоторых случаях методическая программа исследований уступает разработкам даже первой половины XX века. Нередко анализу подвергаются исключительно частоты пальцевых
20

узоров, а межпопуляционная изменчивость ладонных признаков игнорируется. Некоторые китайские и африканские авторы объединяют при анализе мужские и женские выборки и игнорируют половые различия, ссылаясь при этом на т.н. «европейско- американский стандарт» (Zhang et al., 2010; Osunwoke et al., 2008; и др.).
Благодаря усилиям отечественных и зарубежных специалистов российские антропологи располагают одной из самых больших по объему и территориальному охвату базой данных по дерматоглифике современного населения значительной части ойкумены. Дерматоглифика остается открытой для дальнейшего совершенствования подходов к материалам, в т.ч. аналитического аппарата, а возможности ее применения в антропологическом анализе еще далеко не исчерпаны.
21

%

Раздел I.
МЕТОДИЧЕСКИЕ И МЕТОДОЛОГИЧЕСКИЕ ОСНОВЫ ДЕРМАТОГЛИФИКИ

Глава 1. Эмбриогенез и генетика гребешковой кожи кисти
1.1. Эмбриогенез
Гребешковая кожа происходит из наружного зародышевого листка и формируется в течение 3-6-го месяцев пренатального периода (Гусева, 1982; Низимбетова, 1959; Bonnevie, 1929; Penrose, O’Hara, 1973). Развитие гребешковой кожи входит в процесс дифференцировки частей и тканей тактильного органа с общими закономерностями роста и развития (Гусева, 1986). Образованию эпидермальных гребней предшествует формирование волярных возвышений. Кожные узоры возникают на этих подушечках.
Верхние конечности у эмбриона закладываются в виде бугорков - выростов туловища. У пятинедельного эмбриона кисть имеет форму лопасти, на которой около 6,5 недель развития образуются пять зубцов, соответствующих пальцам. Эпителий на этом этапе, в основном, однослойный и тонкий.
В том же возрасте развиваются 2-4-я межпальцевые подушечки (Рис. 1.1, 1.2). К началу 7-й недели появляется центральное возвышение ладони (Рис.1.1В), которое исчезает в течение этой недели (Cummins, 1929).
ГИНПРисунок 1.1. Стадии развития кисти и волярных подушечек у эмбриона: А-D (по: Cummins, 1929).Figure 1.1. The development stages of hand and volar pads: A-D (from: Cummins, 1929).
На 8-й неделе образуются апикальные подушечки, все одновременно или от 1-го пальца к 5-му в соответствии с кранио- каудальным (или радиально-ульнарным для кисти) градиентом развития (Cummins, 1929). К концу 8-й недели у эмбриона
24

завершается дифференцировка пальцев рук (Рис.1.1С). Пальцы имеют разное эмбриональное происхождение: 1 -3-й и радиальная часть 4-го пальца формируются из нижних шейных миотомов, а ульнарная часть 4-го и 5-й палец - из 8-го шейного и 1 грудного (Пэттен, 1959).
Рисунок 1.2. Кисть эмбриона с пальцевыми и ладонными подушечками:А - схема (Cummins, 1929), Б - электронно-микроскопическоеизображение (Carlson, 1999): 1-5 - пальцевые подушечки, Th - тенар, I-IV - межпальцевые подушечки, Hy - гипотенар, Hd - гипотенар дистальный, Hp - гипотенар проксимальный.Figure 1.2. Hand of a human fetus with finger and palmar pads: A - scheme (Cummins, 1929), Б - electron microscope image (Carlson, 1999): 1-5 - finger pads, Th - thenar, I-IV - interdigital pads, Hy - hypothenar, Hd - distal hypothenar, Hp - proximal hypothenar.
Гипотенар, тенар и 1-я межпальцевая подушечка поднимаются в середине 8-й недели. Они закладываются дистальной и проксимальной частями, которые быстро сливаются друг с другом. Волярные возвышения у плода соответствуют подушечкам на генерализованной конечности млекопитающих. В некоторых случаях появляются дополнительные возвышения на основных фалангах пальцев, гомологи которых встречаются у приматов (Cummins, 1929; Whipple, 1904). Волярные подушечки развиваются одновременно с флексорными складками ладоней и пальцев рук (Kimura, Kitagawa, 1986; Kimura, Schaumann, 1988). В эпителии плода 8-9 недель выделяются три слоя: базальный, шиповатый и
25

зернистый (Гусева, 1982). На этом этапе основные нервные стволы рук уже сформированы, и появляются кровеносные сосуды глубокой сети кожи. К 2,5 месяцам нервные волокна врастают в поверхностные слои кожи (Калантаевская, 1972).
Рисунок 1.3. Схема формирования апикальной подушечки в течение 6-17 недель внутриутробного развития без учета увеличения размеров пальца (по: Wertheim, Maceo, 2002).Figure 1.3. Scheme of formation of 6-17-weeks fetus apical pad nonmetering increase of finger sizes (from: Wertheim, Maceo, 2002).
Подушечки кисти вздуваются и достигают максимального развития в 9-10 недель внутриутробной жизни. Пальцевые возвышения в этом возрасте полностью занимают дистальные фаланги и могут выходить за их границы. Увеличение подушечек является следствием пролиферации герментативного слоя эпидермиса, умножения в дерме фибриллярных волокон и скопления межтканевой жидкости (Гусева, 2010). Затем постоянные подушечки начинают редуцироваться, а дополнительные и центральное возвышение к 2,5 месяцам исчезают (Гусева, 1982; Babler, 1991; Cummins, 1929). Таким образом, в онтогенезе волярные подушечки проходят три стадии: появление, рост и редукция (Рис. 1.3, 1.4).
Регрессивные изменения подушечек соответствуют началу формирования кожных гребней. В 9-10 недель в эпидермисе образуются скопления клеток, которые позже разрастаются в направлении дермы (Рис.1.5А). Эти углубления (первичные выросты эпидермиса) увеличиваются и у плода 10-13-ти недель образуют заметные выступы - будущие протоки потовых желез (Рис.1.5Б).
26

Рисунок 1.4. Схема изменений размеров и формы апикальной подушечки (вид сбоку): 1 - выступание подушечки (7-10-я недели), 2 - рост подушечки выше кончика пальца (11-я неделя); 3 - редукция подушечки (12-13 недели), 4 - завершающая стадия (14-я неделя) (по: Seidenberg- Kajabova et al., 2010).Figure 1.4. Scheme of alteration of apical pad’s size and shape (lateral view): 1 - bulging of pad (7-10 weeks), 2 - growth of pad’s area of tip (11 week), 3 - pad’s reduction (12-13 weeks), 4 - closing stage (14 week) (from: Seidenberg- Kajabova et al., 2010).
Рисунок 1.5. Поперечный срез волярной кожи плода: А - 10.5 недель (участки пролиферации клеток эпидермиса), Б - 10.5 - 16 недель (первичные гребни), В - около 16 недель (вторичные гребни) (по: Babler, 1991), Г - 16 недель (прорастающие в дерму протоки потовых желез) (по: Гусева, 1986).Figure 1.5. Transverse section of fetus volar skin: A - 10.5 weeks (area of epidermis cells’ proliferation), Б - 10.5 - 16 weeks (primary ridges), B - about 16 weeks (secondary ridges) (from: Babler, 1991), Г - 16 weeks (sudoriferous glands’ ducts intergrowing in inner skin) (from: Guseva, 1986)
27

Они прорастают в подкожную жировую клетчатку, извиваются и формируют потовые железы (Рис.1.5Г), устья которых около 14-ти недель открываются вдоль срединной линии гребней через равные промежутки (Babler, 1991).
В возрасте 4-5 мес. пренатального онтогенеза между первичными выростами или на их боковой поверхности образуются вторичные выступы, не являющиеся зачатками потовых желез, а определяющие глубину борозд между гребнями (Рис. 1.5В).
В период быстрого увеличения площади ладони формируются дополнительные элементы кожного рельефа - минуции: разветвления гребней, островки и др. (Hale, 1949). В дерме в то же время дифференцируются сосочковый и сетчатый слои.
К концу плодного периода между первичными и вторичными выростами эпидермиса появляются рудиментарные выступы третьего порядка (Калантаевская, 1953).
В начальный период гребнеобразования в эпидермис проникают свободные нервные окончания. Позже (на 3 -4-м месяце пренатального развития) в основании дермы и в подкожной жировой клетчатке формируются тельца Фатера-Пачини (Гусева, 1986; Калантаевская, 1972; Отелин, 1965; Отелин, Машанский, Миркин, 1976).
Образование кожных гребней распространяется в двух направлениях: от пальцев к ладони и по радиально-ульнарному градиенту (Hale, 1952) (Рис.1.6). У плода 15-17 недель гребни покрывают всю пальцевую подушечку, и можно различить типы эпидермальных рисунков. Узоры образуются из маленького резко очерченного центра на вершине апикальной подушечки (Gould, 1948). В случае петлевых или завитковых узоров одна система гребней распространяется из центра, другая - от ногтевого валика в проксимальном направлении, позже третья - со сторонымежфалангового сустава - в дистальном направлении (Рис.1.7, 1.8). Там, где три системы гребешков сходятся, образуется трирадиус (дельта). Раннее развитие первичных гребней приводит кформированию завитков, а позднее - дуг (Babler, 1977, 1978).
Тип образующегося узора, по-видимому, зависит от высоты подушечки и толщины эпидермиса. Дуги формируются на плоских пальцевых возвышениях, а сложные узоры - на хорошо очерченных вздутых подушечках (Гусева, 1986; Bonnevie, 1929). Есть указанияна ведущую роль ширины пальцевой подушечки в развитиигребневого рельефа (Babler, 1987). Симметричные узоры (завитки и
28

дуги) образуются на симметричных возвышениях, а петли - на асимметричных (Рис.1.9). Ориентация петель зависит от толщины эпидермиса с радиальной и ульнарной стороны пальца (Babler, 1987). Неправильности рельефа поверхности волярных подушечек влекут за собой развитие следов узоров (Cummins, 1926).
Рисунок 1.6. Первичные зоны образования кожных гребней на волярной поверхности кисти (по: Cummins, Midlo, 1961).Figure 1.6. Primary areas of skin ridges formation in the volar area of a hand (from: Cummins, Midlo, 1961).
Рисунок 1.7. Схема развития кожных гребней на пальцах плода (по: Cummins, Midlo, 1961). А - завиток, Б - дуга.Figure 1.7. Scheme of skin ridges formation on fetus digitals (from: Cummins, Midlo, 1961). А - whorl, Б - arch.
29

Рисунок 1.8. Формирующиеся гребневые узоры: А - дуга на пальце плода 20-ти недель, Б - завиток на пальце плода 16-17-ти недель, В - участок дельты на 3-й межпальцевой подушечке ладони плода 24-х недель (по: Гусева, 1986).Figure 1.8. Formation of ridge patterns: A - arch on a 20-weeks fetus tip, Б - whorl on a 16-17-weeks fetus tip, В - area of delta on palmar III interdigital pad of a 24-weeks fetus (from: Guseva, 1986).
Рисунок 1.9. Зависимость папиллярных узоров от симметричности пальцевой подушечки: А, Б - дуга и завиток на симметричных подушечках, В - петля на асимметричной подушечке (с модификацией по: Wertheim, Maceo, 2002).Figure 1.9. Correlation between papillary patterns and symmetry of finger pad: A, Б - arch and whorl on symmetric pads, В - loop on asymmetric pad (modified from: Wertheim, Maceo, 2002).
Хотя папиллярные линии плода образуются в процессе эпидермально-дермальных взаимодействий (Гусева, 2010), ведущую роль в этом процессе играет эпидермис, поскольку он способен к самоорганизации и формированию сложных гребневых узоров в культурах клеток (Green, Thomas, 1978).
На 5-м месяце у плода разрастаются кровеносные сосуды подсосочковой сети и артериолы сосочков (Калантаевская, 1972).
30

Около 18-ти недель внутриутробного развития плоские гребни поднимаются над поверхностью кожи (Bonnevie, 1929). В старческом возрасте гребни уплощаются и истончаются (Рис.1.10), но конфигурация кожных узоров остается неизменной на протяжении всей жизни человека. Базальный слой эпидермиса сохраняет способность к гребнеобразованию в постнатальном онтогенезе и при повреждении кожи регенерирует, восстанавливая детали папиллярного рельефа (Гусева, 1986).
Рисунок 1.10. Схема изменений строения папиллярных гребней от эмбрионального периода (слева) до старости (справа) (по: Wertheim, Maceo, 2002).Figure 1.10. Scheme of papillary ridges’ formation from embr yonal period (on the left) till senility (on the right) (from: Wertheim, Maceo, 2002).
К 22-24 неделям формирование папиллярных узоров и гистогенез кожи в основном завершается. В это время слои дермы и эпидермиса (за исключением рогового слоя, образование которого только начинается) хорошо выражены. Развиты кровеносные сосуды и нервы.
После 22-24-й недели и до рождения кожа созревает как тактильный орган. Формируется защитный роговой слой эпидермиса. У плода 5-6-го месяца появляются тельца Вагнера- Мейснера (Гусева, 1982; Калантаевская, 1972; Отелин, Машанский, Миркин, 1976). Увеличивается число нервных окончаний в подкожной жировой клетчатке и в сосочках дермы. Нервные окончания распределены по радиально-ульнарному и проксимальнодистальному градиентам. Например, телец Фатера-Пачини существенно больше в 1 -3-м пальцах, чем в 4-ом и 5-ом. Кожа дистальных фаланг снабжена нервными окончаниями лучше, чем кожа основных и средних фаланг (Гусева, 1982).
Отмечена связь морфологии гребневого рельефа с характером иннервации кожи плода. Во-первых, установлена зависимость между активностью дифференцировки эпидермиса и
31

насыщенностью кожи нервными окончаниями (Загорученко, 1973). Во-вторых, топографические особенности рецепторного аппарата обнаруживают параллелизм со строением кожных узоров. При дуговых узорах средние и боковые ветви нервов соприкасаются, а при петлевых и завитковых - между ветвями есть резкий разрыв. В случае двухцентровых узоров основная ветвь нервного окончания разделена на два пучка (Bonnevie, 1929). С использованием электронно-микроскопической техники подтверждена роль сенсорных аксонов и капиллярно-нервных пар в гребнеобразовании (Dell, Munger, 1986; Hirsch, Schweichel, 1973).
Папиллярные узоры связаны общим морфогенезом с особенностями строения кровеносных сосудов (Усоев, 1976; Bonnevie, 1929(1); Cummins, 1926). Тем не менее, развитие сосудистой системы не подходит на роль детерминанты формирования кожных узоров: рисунок капиллярной сети очень изменчив и образуется в большей степени под воздействием внешних, а не генетических влияний (Канаев, 1959).
При исследовании морфогенеза гребневой кожи нельзя не учитывать влияния факторов билатеральной, лучевой, а также флуктуирующей изменчивости. Билатеральная асимметрия в распределении узоров такова: завитки на пальцах, узоры на гипотенаре и 3-й межпальцевой подушечке, поперечно направленные ладонные линии чаще встречаются на правой руке (Гладкова, 1959). Петли на пальцах, узоры на тенаре и 4 -й межпальцевой подушечке, косо расположенные ладонные линии и карпальный трирадиус - на левой руке.
Возможно, билатеральная изменчивость дерматоглифических признаков вызвана моторной асимметрией рук, хотя эту точку зрения поддерживают не все исследователи (Гладкова, 1966; Leche, 1933). Обычно у правшей несколько больше завитков на правых руках, петель - на левых, у левшей - наоборот (Newman, 1934). Обратные соотношения дерматоглифических характеристик у право- и леворуких выражены на ладонях слабее, чем на пальцах. У правшей главные ладонные линии заканчиваются дистальнее на правых руках, а у левшей - на левых (Keith, 1924). Отмечена повышенная узорность 2-й межпальцевой подушечки левой ладони у леворуких (Cromwell, Rife, 1942).
Распределение частот признаков на пальцах кисти и разных участках ладоней у народов мира обнаруживает общие закономерности. Подушечки ладони чаще других заняты
32

бездельтовыми узорами, завитки встречаются редко. Узорность гипотенара в большинстве групп выше, чем тенара и 1 -й межпальцевой подушечки. Во всех популяциях на пальцах рук преобладают ульнарные петли и завитки. Редко встречаются дуги и, особенно, радиальные петли. Завитковые узоры чаще всего образуются на 1-м и 4-м пальцах рук, дуговые - на 2-м и 3-м, ульнарные петлевые - на 5-м и 3-м пальцах, а радиальные - на 2-м (Волоцкой, 1937; Гладкова, 1966; Cummins, Midlo, 1961).
Причины такого избирательного «предпочтения» узорами определенных пальцев до сих пор не получили четкого объяснения. Определяющим моментом могут быть функциональная неравнозначность (Гусева, 1986) или различия в размерах пальцев (Хайруллин, Никитюк, Крикун, 2005), что подтверждается изменениями дерматоглифики у больных с врожденными дефектами кисти (Мглинец, 1991). Возможно, неодновременное начало (с 11,5 недель) и разный объем двигательной активности пальцев плода, вызывающей натяжение и расслабление кожи, влияют на процесс формирования гребневого рельефа (Humphrey, 1964; Kimura, Kitagawa, 1986).
Развитие пальцевых узоров по времени совпадает с началом выраженного роста конечностей в длину. Рост кисти считают одним из важнейших факторов морфогенеза гребневого рельефа (Хайруллин, Никитюк, Крикун, 2005). Зафиксированы корреляции кожных узоров с размерами пальцев. Тип узора связан с шириной подушечки дистальной фаланги пальца плода: чем уже подушечка, тем более сложный узор на ней развивается (Babler, 1987, 1990). Размеры и форма пальцев демонстрируют значимые связи с типом папиллярных узоров и в постнатальном онтогенезе человека (Хайруллин, Никитюк, Крикун, 2005).
Характеристики пальцевой дерматоглифики сопряжены с темпом внутриутробного развития, подобные связи выявляются и после рождения. При интенсивном росте и скорости созревания организма (по зрелости скелета и зубов) суммарный гребневой счет повышен (Багдасарян, 1974). Ассоциации степени развития вторичных половых признаков с дельтовым индексом, гребневым счетом и узорностью ладоней у девочек положительны и достоверны. Возраст менархе демонстрирует обратную связь с этими дерматоглифическими характеристиками (Комиссарова, Сазонова, Карелина, 2005; Славолюбова, 2008).
33

Для реализации генетической программы развития гребневого рельефа необходимо поддержание определенного гормонального фона, поскольку именно эндокринная система в пренатальном периоде играет большую роль в дифференцировке и росте тканей плода (Василенко, 2005; Губарева, 2001; Кобозева, Рассохин, 1984). Вероятно, эндокринный статус матери и плода, зависящий от наследственных и от средовых влияний, воздействует на формирование дерматоглифического рисунка (Зубович, 1989; Таболин, Лукина, 1975). В возрасте девяти недель у плода начинают функционировать половые железы, гормоны которых могут влиять на митотическую активность герментативного слоя эпидермиса (Бодемер, 1971). Возможно воздействие плацентарного гормона роста (Дильман, 1986).
Связи гормональных показателей и признаков дерматоглифики в некоторой степени сохраняются и в постнатальном онтогенезе. Так, у подростков и молодых мужчин отмечена достоверная положительная связь уровня тестостерона, а также соотношения тестостерона и эстрадиола с завитками, двойными петлями, количеством трирадиусов, ульнарным гребневым счетом и отрицательная - с дугами (Василенко, 2005; Джанибекова, 2002). Корреляции показателей кортизола, антагониста тестостерона, имеют противоположный характер (Василенко, 2005). Уровень эстрадиола обнаруживает отрицательную связь с продольным направлением главной ладонной линии А и высоким расположением осевого трирадиуса ладони.
Средовые влияния, способные существенно изменить строение кожных узоров, действуют на плод в период образования узоров или незадолго до него (Гусева, 1988). Это могут быть инфекционные заболевания, например, внутриутробная краснуха, загрязненный воздух, радиационное воздействие и проч. (Крикун, 2006; Пэттен, 1959).
Итак, формирование гребневого рельефа происходит в середине пренатального периода под действием многих внутренних и внешних, локальных и общих факторов. В дальнейшем кожные узоры не изменяются на протяжении всей жизни человека.
34

1.2. Наследование дерматоглифических признаков
Генетические проблемы дерматоглифики до сих пор остаются неразрешенными. В настоящее время нет однозначных ответов на вопросы о хромосомной локализации, количестве генов, определяющих развитие структур гребневой кожи, и механизме наследования узоров.
Предположение о наследственной природе гребневого рельефа кожи возникло еще на заре формирования дерматоглифики (Faulds, 1880; Wilder, 1902). Родоначальник науки Ф. Гальтон указывал на большее сходство пальцевых узоров в парах близнецов по сравнению с сиблингами (Galton, 1892, 1895). С развитием дерматоглифики эволюционировали представления о степени и характере генетических влияний на эпидермальный рельеф (Божченко и др., 2002). Сначала исследователи отрицали наследственную природу кожных рисунков из-за того, что в ряду поколений узоры не передавались на гомологичные пальцы, а их распределение не соответствовало менделевскому (Locard, Seneth, 1906; Stockis, 1908: цит. по Cummins, Midlo, 1961). Результаты дальнейших работ привели к признанию жесткой детерминированности пальцевых узоров. По современным же представлениям уровень наследственных влияний на формирование папиллярных узоров и составляющих их элементов сильно варьирует: от 12 до 98% (Ройтман, Липовецкая, 1969; Holt, 1968; Loesch, 1971). Таким образом, одни признаки дерматоглифики почти полностью генетически предопределены, а другие в большей мере зависят от внешних воздействий, в первую очередь внутриутробных.
В целом, узоры на пальцах кисти наследуется теснее, чем на ладони. Наследственные влияния на суммарный гребневой счет пальцев составляют 98%, на дельтовый индекс - до 87% (Ройтман, Липовецкая, 1969). Связи тотальных фенотипов (сочетание узоров без учета занятых ими пальцев) у родителей и детей достигают 89%. Завитки детерминированы более жестко, чем петли и, особенно, дуги (Гусева, 2010). Генетическую основу имеет форма пальцевых узоров (Bonnevie, 1924; Mueller 1930, 1931), например,эллиптический вариант завитка доминирует над циркулярным. Доминантное наследование выявлено для сложных узоров (Семеновский, 1926, 1927).
Узоры ладони тоже в разной степени зависят от генетических и средовых воздействий. Преимущественно генами определяются
35

радиальная петля гипотенара, петля 2-й межпальцевой подушечки, краевой и карпальный осевой трирадиусы, комбинации петель, открытых к краям ладони и общая узорная интенсивность. Индекс наследования, вычисленный в ходе семейных исследований, максимален для комбинации петель (0,86) и минимален в этой группе характеристик для петли на 2-й межпальцевой подушечке (0,62) (Loesch, 1971). Почти полностью зависят от внутриутробной среды ульнарная и карпальная петли на гипотенаре, а также пальцевые трирадиусы (Okajima, Iwayanagi, Inouye, 1982). Остальные узоры ладони характеризуются средним уровнем наследования (Loesch, 1971).
Отдельные признаки дерматоглифики (радиальная петля на гипотенаре, петля на 2-й межпальцевой подушечке и низкий осевой трирадиус), возможно, контролируются единичными генами, в случае петель - в гомозиготной рецессивной форме (Loesch, 1971).
Ход главной ладонной линии А контролируется полигенной системой с аддитивным эффектом без связи с полом (Glanville, 1965).
Гребневой рельеф стопы, по сравнению с кистью, в меньшей степени наследственно обусловлен. Халлюкальная область, тем не менее, подвержена значительному влиянию генетических факторов. Узор 1 -го пальца стопы зависит, в основном, от средовых воздействий (Reed, Sprague, Kang et al., 1975).
Генетика папиллярных узоров дистальных фаланг пальцев рук изучена подробнее, чем других участков гребневой кожи. Однако ни одна из гипотез о механизмах наследования не может объяснить всей сложности передачи пальцевых узоров в линиях родственников (Гусева, Сорокина, 1998).
Ранние генетические исследования, как уже было указано, опирались на ошибочные предположения о передаче узоров с пальцев родителей на гомологичные пальцы детей (Elderton, 1920; Gruneberg, 1928; Pospisil, 1967). Но узоры на одноименных пальцах рук не всегда совпадают даже у монозиготных близнецов: локальное сходство по количеству дельт меньше тотального, иногда существенно (Волоцкой, 1936, 1937). Очевидно, в генетических исследованиях нужно учитывать дерматоглифику всех пальцев рук индивида (Войтенко, Полюхов, Колодченко, 1979; Гусева, 1986; 2010).
По мнению большинства авторов, в основе узоров пальцевых подушечек и составляющих их элементов лежит полигенная система
36

(Волоцкой, 1937; Маккьюсик, 1967; Рицнер, 1970; Рицнер, Шехтер, 1975; Holt, 1961). В одних гипотезах наследственно обусловленными считают тип и/или ориентацию узора (Elderton, 1920; Gruneberg, 1928), в других - количественные признаки и элементы кожных рисунков: гребневой счет, положение центра или количество дельт (Волоцкой, 1937; Bonnevie, 1924; 1931).
Заметное влияние на развитие генетики признаков дерматоглифики оказала гипотеза К. Бонневи (Bonnevie, 1929, 1931). В результате посемейных, близнецовых и эмбриологических исследований автор пришла к выводу о наследственной детерминации закладки центра пальцевого узора и гребневого счета, а не типа и ориентации узора. Было выдвинуто предположение о существовании трех двуаллельных генетических факторов, определяющих толщину эпидермиса на всех пальцах кисти, а также на пальцах ульнарной (4-5) и радиальной (1-3) сторон. В основе изменчивости пальцевых узоров, по мнению К. Бонневи, лежат сочетания аллелей указанных генов. Тип и ориентация узора определяются формой эмбриональной подушечки пальца, водной насыщенностью эпидермиса и зависят от ветвления нервов. Гипотеза не нашла подтверждения в отношении локальных генетических факторов и конкретных изменений гребневого счета в зависимости от сочетаний аллелей генов, но указала направление поиска общих принципов наследования дерматоглифических признаков.
Развитием подобного подхода стала «трехполигенная типологическая модель» наследования узоров дистальных фаланг пальцев, предложенная и совершенствуемая И.С. Гусевой (1982; 1986; 2008; 2009; 2010). По мнению автора, развитие пальцевых дерматоглифов происходит под контролем системы независимых двуаллельных генных локусов с полным доминированием, действующих в разные периоды морфогенеза. Раньше других активизируется ген fs (friction skin), регулирующий развитие кожных гребней. Затем включаются в работу гены, определяющие вздутость пальцевой подушечки и детерминирующие типы узоров (дуги, завитки или петли без учета направления). Ген W регулирует водную насыщенность дермы, ген A - замедляет скорость деления клеток ростового слоя эпидермиса. По мнению И.С. Гусевой эти гены индуцируются прорастающими в кожу кровеносными сосудами и свободными нервными окончаниями. Вариации толщины слоев кожи определяют развитие вариантов узоров.
37

Сложные узоры наследуются по аутосомно-доминантному типу, а простые - по рецессивному. Поскольку пальцы рук находятся в разных условиях, гены A и W проявляются на каждом из пальцев по-разному. Ульнарное и радиальное направление узоров определяет особый генетический фактор L. Гребневая ширина и расстояние от дельты до центра наиболее генетически зависимы, в то время как гребневой счет является производным от них признаком.
Хотя рассмотренная модель наследования пальцевых узоров считается наиболее обоснованной (Божченко и др., 2002), на больших выборках она не подтверждается (Ящук, 1985). Приближение эмпирического распределения фенотипов к теоретически ожидаемому достигается объединением петель и завитков в тип сложных узоров (Гусева, 2010).
Существует точка зрения, что основные типы пальцевых дерматоглифов, гребневой счет и дельты детерминируются генами A, L и W (Рицнер и др., 1971). Гены A и L подавляют действие W, а ген W интенсифицирует A и L.
Попыткой объяснения общих законов наследования узорности и механизмов дельтообразования на пальцах рук является «двухкомпонентная пороговая модель» (Войтенко, Полюхов, Колодченко, 1979). В системе, детерминирующей этот процесс, предполагают спирализующий (ульнарный) и деспирализующий (радиальный) компоненты. Первый формирует завитки, второй - дуги. Под действием двух компонентов образуется ульнарная петля, представляющая допороговый узор. Радиальная петля развивается при максимально выраженном радиальном компоненте. При всей оригинальности гипотеза не подтверждается на семейном материале и страдает многими недостатками. Исходно неверна идея о ведущей роли дельтообразования в формировании типов узоров, вызывает вопросы ориентация спирализующего и деспирализующего компонентов и выбор ульнарной петли в качестве допорогового варианта (Гусева, 1986; 2010).
Вопрос о влиянии половых хромосом на формирование пальцевых узоров в настоящее время дискутируется. По данным ряда исследователей уровень корреляции количественных признаков в парах родственников свидетельствует в пользу аутосомной локализации генов, детерминирующих пальцевые узоры (Волоцкой, 1936; Ройтман, Липовецкая, 1972; Holt, 1968). Другие авторы признают участие половых хромосом в формировании кожного
38

рельефа дистальных фаланг пальцев (Cummins, Midlo, 1961; Penrose, Loesh, 1970). Полагают, что в X-хромосомах находятся гены, определяющие развитие потовых желез - первичных элементов гребешковой кожи (Гусева, Сорокина, 1998). Есть данные, указывающие на возможность сцепления с Х-хромосомой генов, детерминирующих общее количество дельт на дистальных фалангах пальцев рук (Никитюк, Коган, Гальперина, 1986; Спасов, Докумов, Генкова, 1978), завитковые и дуговые варианты узоров. Возможно, гены, определяющие формирование пальцевых дерматоглифов, локализуются в аутосомах, но половые хромосомы оказывают на них модифицирующее действие (Penrose, 1963).
Генетика гребневого рельефа средних и основных фаланг пальцев кисти практически не изучена. Единичные близнецовые исследования указывают на высокий уровень наследования (90-93%) узоров средних и основных фаланг (Ploetz-Radmann, 1937). Эти признаки, видимо, наследуются по полигенному типу с пенетрантностью, по-разному проявляющейся по системам фаланг и вариантам узоров. Половые хромосомы оказывают модифицирующее влияние на узорность, что проявляется в накоплении дуговых рисунков у женщин (проксимальных на средних и дистальных на основных фалангах), а волнообразных - у мужчин (Шпак, 2006).
Завершая краткий обзор исследований генетических основ признаков кожного рельефа, следует констатировать, что папиллярные узоры наследуются в разной мере, и в основе большинства признаков лежит полигенная система. «Формирование дерматоглифических признаков происходит в условиях сложного эпистатического и комплементарного взаимодействия нескольких пар родительских генов, влияния внешней и внутренней сред. Механизм взаимодействия и степень выраженности каждого из указанных факторов в настоящее время научно не объяснены» (Фандеева, 2010, с.8).
39

Г лава 2. Методика получения отпечатков, описания признаков и первичной статистической
обработки данных
2.1. Получение отпечатков
Существуют разные техники получения отпечатков ладоней и пальцев. Обычно оттиски делают с помощью типографской краски (Cummins, Midlo, 1961). Этот способ наиболее экономичен, кроме того, отпечатки сохраняются десятки лет. К числу недостатков метода можно отнести этические проблемы, необходимость пачкать руки испытуемым и возможность возникновения аллергических реакций на краску и скипидар.
Для получения отпечатков нужно иметь черную или синюю типографскую краску, скипидар, бумагу средней плотности и шероховатости формата А4, стеклянные пластинку и палочку, подушечку-подстилку (25х28 см) (обтянутые тканью 5-6 слоев фланели на фанерном основании), типографский резиновый валик. Работу нужно проводить в хорошо проветриваемом помещении.
Криминалисты делают оттиски в специальных дактилоскопических картах. В биологических исследованиях на листе бумаги в верхнем левом углу записывают сведения об испытуемом: фамилию, имя, отчество, пол, возраст, место рождения, национальность родителей, бабушек и дедушек. Затем лист укладывают на подушечку.
Стеклянной палочкой краску наносят на стеклянную пластину в 2-3 местах (загустевшую краску предварительно разводят скипидаром до состояния сметаны) и равномерно раскатывают валиком. Ладонную поверхность кисти прокрашивают валиком от карпальной складки до кончиков пальцев.
Испытуемого просят положить кисть на лист бумаги ульнарным краем. Исследователь осторожно опускает ее на бумагу прокрашенной поверхностью и одним движением (чтобы не смазать оттиск) продавливает середину тыльной стороны ладони, затем большой палец немного наклоняет ульнарно, нажимая на ногтевую фалангу, чтобы пропечаталась апикальная подушечка. Затем кисть испытуемого поднимают одной рукой вверх, а второй придерживают бумагу, стараясь не смазать отпечаток. Оттиски
40

правой и левой кистей можно выполнять на одном листе бумаги с двух сторон. Особое внимание следует уделять получению четкого изображения проксимального края ладони, в том числе карпальной складки.
Для улучшения качества отпечатков ладони можно использовать аппарат Антонюка (1975), который представляет собой широкий валик с ручкой, закрепленный на прямоугольном основании. На валик укладывают лист бумаги, на который обследуемый устанавливает запястье. При вращении валика с помощью ручки по нему прокатывается кисть вместе с листом бумаги и таким образом получается оттиск. Недостаток метода состоит в том, что ульнарный край ладони и большой палец таким способом пропечатываются хуже, чем на подушечке.
Оттиски ногтевых фаланг делают рядом с отпечатком кисти в строго определенном порядке: от первого пальца к пятому. Подушечки пальцев прокрашивают валиком тремя движениями, захватывая боковые поверхности фаланги. Исследователь прокатывает ногтевую фалангу пальца обследуемого с радиальной на ульнарную поверхность и поднимает его палец вверх (проксимально отпечаток должен захватывать межфаланговую складку, а дистально - доходить до ногтя). В процессе процедуры испытуемый должен держать руку расслабленно, но помогать легкими вращательными движениями, не продавливая палец.
При необходимости получить оттиски средних и основных фаланг пальцы прокрашивают и прокатывают целиком (вплоть до пястно-фаланговых складок).
После взятия отпечатков краску с ладоней снимают ваткой, пропитанной скипидаром, а затем смывают мылом. В полевых условиях часто обходятся без скипидара.
Многочисленные способы, призванные сменить метод получения оттисков с помощью типографской краски, не нашли широкого применения из-за худшего качества и меньшего срока сохранности отпечатков. Речь идет о методах с использованием различных красителей, пергаментной или фотографической бумаги (Бунак, 1941; Cummins et al., 1929; Schott, 1928; Strong, 1929, цит. по Гладкова, 1966).
С развитием компьютерной техники были сконструированы сканеры пальцев и ладоней, с помощью которых можно получить изображения отпечатков ладонной поверхности кисти, не прибегая к ее окрашиванию (Арутюнян, 2010; Бурлай, 2009; Задорожный, 2004;
41

Новикова, Бурлай, Спиридонов, 2009; Хрулев, Аполлонова, Спиридонов, 2005). Использование этих приборов снимает все минусы метода типографской краски. Кроме того, полученные таким образом изображения гребневого рельефа рук можно обрабатывать с помощью компьютерных программ, позволяющих с большой долей вероятности определять типы узоров или гребневой счет без визуального контроля исследователя.
Безусловно, дальнейшее совершенствование компьютерных технологий облегчит сбор материала и первичную обработку данных по дерматоглифике.
2.2. Описание признаков
2.2.1. ПАЛЬЦЕВЫЕ УЗОРЫ
Дистальные фаланги пальцевДля определения типов узоров на дистальных фалангах
пальцев (пальцевых подушечках) обычно используют классификацию Ф. Гальтона (Galton, 1892) и некоторые узоры схемы Э. Генри (Henry, 1934).
В основу разделения главных типов узоров в классификации Ф. Гальтона положено количество дельт или трирадиусов. Эти элементы узора формируются в тех местах, где сходятся три разнонаправленных потока папиллярных линий. Дельта в строгом понимании - треугольник, образованный кожными гребнями. Трирадиус в идеале состоит из трех гребней (радиантов), расходящихся из одной точки под равными углами (Рис. 2.1В). Нередко последнее условие не выполняется, и трирадиус устроен более сложно. Если гребни раздваиваются или не соединяются друг с другом, точкой трирадиуса считается участок гребня, расположенный ближе к центру. Когда в центре области трирадиуса находится короткий гребень (островок), он считается точкой трирадиуса.
Радианты, расходящиеся от трирадиуса, обозначают как проксимальный, дистальный и краевой, уходящий в направлении ульнарного или радиального краев пальца (Рис.2.1А, Б).
Завиток (whorl, W) - самый сложный из пальцевых узоров, в идеале образован концентрическими линиями и имеет две дельты, редко более (Рис.2.1А). У типичного завитка проксимальные и дистальные радианты соединяются, формируя замкнутую фигуру
42

(скелет узора), внутри которой находится узорное поле (Рис.2.1А). Область над узорным полем называется дистальной поперечной системой гребней, а под ним - проксимальной. Сердцевина или центр узора в типичных завитках может выглядеть, как островок, короткий отрезок, крючок, либо окружность, эллипс или два сцепленных крючка. Точка центра - наиболее центрально расположенный элемент сердцевины относительно скелета узора.
9 Ю
А 1- ^ Б В
Рисунок 2.1. Строение пальцевых узоров. А - завиток, Б - петля, В - трирадиус; 1 - проксимальная трансверзальная система, 2-проксимальный радиант, 3 - краевой радиант, 4 - дельта, 5 - скелет узора, 6 - поле узора, 7 - сердцевина узора, 8 - дистальный радиант, 9 - дистальная трансверзальная система, 10 - голова петли (по: Cummins, Midlo, 1961).Figure 2.1. Formation of finger patterns. A - whorl, Б - loop, В - triradius; 1 - proximal transversal system, 2 - proximal radiant, 3 - boundary radiant, 4 - delta, 5 - pattern frame, 6 - pattern area, 7 - pattern core, 8 - distal core, 9 - distal transversal system, 10 - head of a loop (from: Cummins, Midlo, 1961)
Петля (loop, L) - однодельтовый узор. Дистальный радиант трирадиуса загибается назад, образуя голову петли, и заканчивается на боковом краю пальца (Рис.2.1Б). В типичной петле центр образован одним или несколькими прямыми гребнями, либо гребнем в форме шпильки. В петле с одним коротким гребнем в качестве сердцевины узора точкой центра считается дистальный конец этого гребня; если гребней два, выбирают более удаленный от трирадиуса. Когда гребней несколько, при четном их количестве работает указанное правило для 2-х гребней, а при нечетном - выбирают лежащий в середине. Если гребень сердцевины узора
43

изогнут, центральной считается наиболее удаленная от дельты точка изгиба. Ориентация петли зависит от того, в какую сторону она открывается. Различают ульнарные (Lu, U) и радиальные (Lr, R) петли (Рис. 2.2).
Наиболее простым узором без трирадиусов является дуга (arch, A) (по сути - это отсутствие узора). Гребни дуги лежат поперек фаланги пальца, немного изгибаясь дистально (Рис.2.2, простая дуга).
U 1 2 3 4 5 R
6 7 8 9 10
Рисунок 2.2. Типы пальцевых узоров. Обозначения: 1 - простая дуга, 2 - шатровая дуга, 3 - ульнарная петля, 4 - радиальная петля, 5 - двойная петля, 6 - истинный завиток, 7 - спиральный завиток, 8 - центральный карман, 9 - латеральный карман, 10 - случайный узор.Figure 2.2. The types of finger patterns. Designation: 1 - simple arch, 2 - tented arch, 3 - ulnar loop, 4 - radial loop, 5 - twin loop, 6 - true whorl, 7 - spiral whorl, 8 - central pocket, 9 - lateral pocket, 10 - accidental pattern.
Классификация Э.Генри (Henry, 1934) более детальна, чем система Ф. Гальтона (1892): помимо количества трирадиусов в ней учитываются степень выраженности и особенности положения элементов узора (Рис.2.2).
Класс дуг подразделяется на простые и шатровые дуги. Гребни простой дуги (A) идут поперек фаланги пальца, несколько изгибаясь в дистальном направлении.
По средней оси шатровой дуги (tented arch, T), или недалеко от нее, папиллярные линии образуют фигуру типа трирадиуса. Дистальный радиант этого «трирадиуса» обрывается, и сверху его огибают гребни поперечной дистальной системы, образующие
44

крутой подъем. В некоторых случаях дистальный радиант резко загибается проксимально. В русской литературе для обозначения шатровой дуги нередко используют неудачный термин «Т-образная дуга» (от «t-formed arch»), не отражающий форму рисунка.
В некоторых дугах есть элементы узора, сходные с редуцированными завитками или петлями и незавершенные трирадиусы. Такие переходные узоры рекомендуется относить к классу дуг, если гребневой счет (см. ниже) в завитке или петле меньше двух. Существует условное правило, согласно которому истинным считается узор, образованный не менее чем 2 гребешковыми линиями, вложенными одна в другую (не считая линий, образующих рамку). Все остальные случаи определяются как след узора (см. далее).
Петли (L) (по Э. Генри) разделяют на простые и переходные. Простая петля (plain loop) образована непрерывным рядом гребней, каждый из которых формирует петельку. В переходной петле (transitional loop) правильный ход гребней нарушен, и рисунок похож на более сложный, но до конца не сформированный узор: завиток, латеральный карман, двойную петлю илицентральный карман.
Петли можно детализировать, подразделив их по размерам (гребневому счету) и форме узорного поля, по наклону к оси пальца, структуре центрального фрагмента и положению его относительно трирадиуса.
Двудельтовые и многодельтовые узоры, относящиеся к классу завитков, разделены на завитки и сложные (составные) узоры.
Завиток или истинный завиток (whorl, W) - концентрический узор, гребни которого образуют окружности, эллипсы, или спирали. В зависимости от положения радиантов различают замкнутый, внешний и внутренний завитки.
В замкнутом завитке (meet whorl) проксимальные радианты соединяются, либо между ними располагается не больше двух папиллярных линий (Рис.2.3: 4, 8). Когда проксимальный радиант, выходящий из левого трирадиуса (на отпечатке) расположен снаружи от правого трирадиуса и отделен от него тремя или более гребнями, завиток называется внешним (outside whorl; Рис.2.3: 13, 15), если изнутри - внутренним (inside whorl; Рис.2.3: 9, 11).
Очевидно, что логичнее прослеживать радианты не из левого трирадиуса, а из радиального или ульнарного (Cummis, Midlo, 1961).
45

I- •j' ^ Y т V ^ Y T
3 0 3] 32 35 3 4 35 36 3 ? 3 8
Рисунок 2.3. «Семейное древо» пальцевых узоров (по: Cummins, Midlo, 1961). Figure 2.3. A “genealogical tree” of fingerprint types (from: Cummins, Midlo, 1961).
46

Составные узоры - двудельтовые (многодельтовые) узоры, образованные двумя или несколькими типичными узорами. К этому классу относят центральные карманы, латеральные карманные петли (латеральные карманы), двойные петли и случайные узоры.
Латеральная карманная петля (lateral pocket, LP) образована двумя соединенными петлями, одна из которых огибает другую. Обе петли открываются в одну сторону. Для того чтобы определить направление петель, прослеживают гребни, выходящие из центра каждой петли. Возможны ульнарная (LP1) или радиальная (LP1) ориентация латеральных карманов.
Двойная петля (twin loops, TL) - рисунок из двух обхватывающих друг друга петель, заканчивающихся на ульнарном и радиальном краях пальца. Двойная петля направления не имеет.
Центральный карман (centralpocket, CP) представляет собой завиток небольших размеров, лежащий в узорном поле петли. В зависимости от направления различают ульнарный (CPU) и радиальный (CPR) центральные карманы. В случае необходимости можно разделить центральные карманы по форме центра, ходу типовых линий и гребневому счету.
Группа случайных узоров (асЫdental, Acc) очень разнородна: она включает в себя любые сочетания простых узоров, кроме описанных выше. Для обозначения таких вариантов, помимо символа Acc, можно использовать последовательную запись всех рисунков, образующих узор.
Варианты узоров имеют непрерывное распределение и многие из них занимают промежуточное положение между разными типами кожных рисунков (Рис.2.3).
Г ребневой счетДля количественной оценки узора существует понятие
«гребневой счет». Способ подсчета гребней диктуют различные цели исследований. В этнической дерматоглифике обычно используют гребневой счет между трирадиусом и центром узора. Эти точки соединяют прямой линией и подсчитывают гребни, попавшие на пересечение с ней. Трирадиус и центр узора не входят в гребневой счет. Обрывающиеся концы гребней, не доходящих до линии подсчета, недоразвившиеся гребни и случайные точки между гребнями в подсчет не включают. Если линия пересекает место бифуркации, учитываются оба гребня (Рис.2.4).
47

Простые и шатровые дуги не имеют гребневого счета (он равен нулю). В дугах, переходных к петлям, гребневой счет варьирует от нуля до двух; в петлевых узорах его величина больше двух. В завитках гребневой счет можно посчитать от двух трирадиусов. Внутренняя точка будет общей для линий, проведенных с двух сторон, если центр представлен островком или коротким гребешком. Когда сердцевина завитка образована кружком, эллипсом, или двумя гребнями в форме серпов, для каждой линии фиксируют центр, ближайший к соответствующему трирадиусу (Рис.2.4Б). Центры составных узоров соединяют с наиболее близко расположенными к ним трирадиусами так, чтобы линии не пересекались. При наличии трех трирадиусов в узоре определяют три гребневых счета.
Рисунок 2.4 Схема определения гребневого счета петли (А) и завитка с двумя центрами (Б): 1-8 - гребни, составляющие гребневой счет (по: Mavalwala, 1978).Figure 2.4. Scheme of defining of the ridge count of a loop (A) and whorl with two cores (Б): 1-8 - ridges used for calculating ridge count (from: Mavalwala, 1978).
До некоторой степени гребневой счет отражает величину узора, хотя следует также учитывать и различия в ширине кожных гребней. При одинаковых размерах узоры могут отличаться по гребневому счету из-за различий в положении центра узора. В двойных петлях и латеральных карманах гребневой счет скорее отражает размеры фрагментов, чем целого узора. Г. Камминс иЧ. Мидло (Cummins, Midlo, 1961) предложили в качестве общего центра для двойной петли использовать точку посередине между
48

центрами обеих петель, однако, на практике этот способ не применяют. Для характеристики размеров узоров у индивида используют тотальный (или средний) гребневой счет - сумма величин гребневого счета на десяти пальцах рук (или средний для одного пальца), причем для двух- и многодельтовых узоров берут максимальный гребневой счет (Penrose, 1968). Абсолютный гребневой счет - сумма величин гребневого счета от каждого трирадиуса на пальцах рук человека, - интегративный показатель сложности и величины узоров.
2.2.2. ЛАДОННЫЕ УЗОРЫ
Сгибательные складки и ладонные подушечкиНадежными ориентирами для оценки расположения
дерматоглифических структур являются главные сгибательные (флексорные) складки ладони. Они постоянны и сходно ориентированы у разных людей.
Снизу ладонь очерчивает дистальная часть браслетной (карпальной) складки, обычно совпадающая с границей гребешковой кожи (Рис.2.5А). В случае заметного несоответствия этих структур ориентируются на проксимальный уровень гребневой кожи. Сверху ладонь ограничена проксимальными частями пястнофаланговых складок. Область ладони вокруг 1 -го пальца, окружена радиальной продольной складкой (складкой 1 -го пальца). Дистальная поперечная складка (трехпальцевая) начинается от ульнарного края ладони и обычно заканчивается под 3 -м пальцем.Проксимальная поперечная складка (пятипальцевая)направляется от радиального края ладони к ульнарному, обычно не достигая его.
В середине ладони находится центральное углубление, вокруг которого располагаются возвышения (ладонные подушечки) разной величины и степени выраженности. Четыре межпальцевые подушечки (I-IV) лежат на уровне межпальцевых промежутков (1-4) (Рис.2.5Б). Самая вздутая область - тенар (Th) - занимает проксимально-радиальную четверть ладони и соответствует мышцам, 1 -го пальца. Тенар очерчен радиальной продольной складкой вместе с плоской 1 -й межпальцевой подушечкой. Эти подушечки описывают вместе (Th/I), поскольку граница между ними слабо различима и, кроме того, они могут быть заняты общим узором.
49

Рисунок 2.5. Топография ладони: А - сгибательные складки, Б - ладонные подушечки (по: Cummins, Midlo, 1961).Figure 2.5. The palmar topography: A - flexion crease, Б - palmar pads (from: Cummins, Midlo, 1961).
50
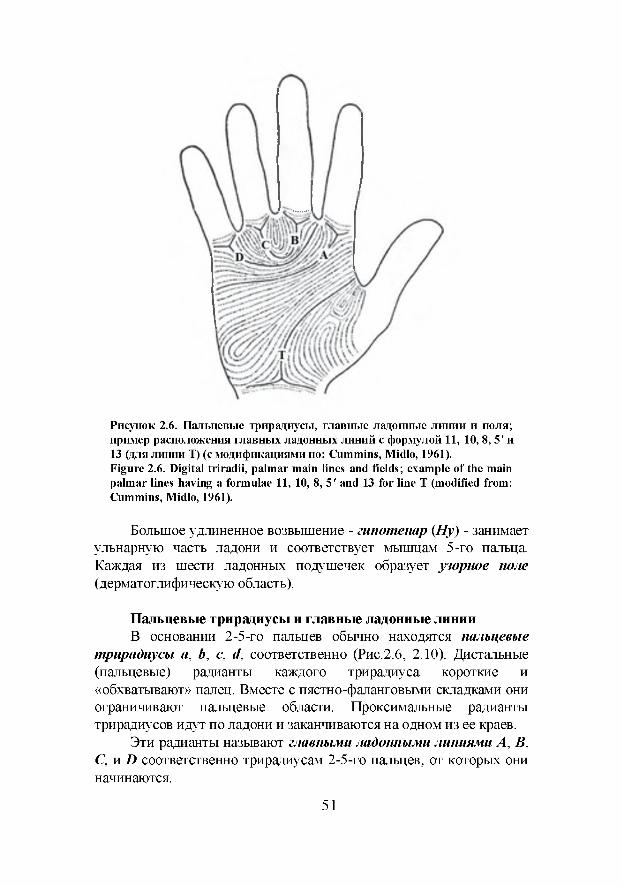
Рисунок 2.6. Пальцевые трирадиусы, главные ладонные линии и поля; пример расположения главных ладонных линий с формулой 11, 10, 8, 5' и 13 (для линии Т) (с модификациями по: Cummins, Midlo, 1961).Figure 2.6. Digital triradii, palmar main lines and fields; example of the main palmar lines having a formulae 11, 10, 8, 5' and 13 for line T (modified from: Cummins, Midlo, 1961).
Большое удлиненное возвышение - гипотенар (Hy) - занимает ульнарную часть ладони и соответствует мышцам 5-го пальца. Каждая из шести ладонных подушечек образует узорное поле (дерматоглифическую область).
Пальцевые трирадиусы и главные ладонные линииВ основании 2-5-го пальцев обычно находятся пальцевые
трирадиусы a, b, c, d, соответственно (Рис.2.6, 2.10). Дистальные (пальцевые) радианты каждого трирадиуса короткие и «обхватывают» палец. Вместе с пястно-фаланговыми складками они ограничивают пальцевые области. Проксимальные радианты трирадиусов идут по ладони и заканчиваются на одном из ее краев.
Эти радианты называют главными ладонными линиями A, B, C, и D соответственно трирадиусам 2-5-го пальцев, от которых они начинаются.
51

Линию прослеживают от гребня или островка, максимально близко расположенного к центральной точке трирадиуса. В случае, когда два гребня в равной мере могут рассматриваться в качестве проксимального радианта (Рис. 2.7 K, L, P), началом главной линии считается радиальный гребень. По ходу главной линии гребень прерывается и переходит на следующий, расположенный по прямой от оборвавшегося участка. Если явного продолжения нет, то линию переносят на соседний гребень, касающийся или расположенный ближе других к концу прервавшегося гребня. В случае, когда линия идет прямо, а соседние гребни находятся от нее на одинаковом расстоянии, предпочтительнее выбрать гребень с радиальной стороны.
Рисунок 2.7. Варианты строения пальцевых трирадиусов; кружочками отмечены проксимальные радианты (по: Cummins, Midlo, 1961).Figure 2.7. Formation variants of fingers triradii; proximal radiants are circuled (from: Cummins, Midlo, 1961).
Когда гребень главной ладонной линии изгибается, ее продолжают по направлению вогнутости.
Как правило, главные ладонные линии не пропечатываются внутри флексорных складок. Их достраивают эмпирически по прямой от обрыва гребня главной линии. На практике же линия А образует подобие ступеньки в месте пересечения со сгибательной складкой и продолжается несколько ниже ее теоретически предполагаемого хода. Для более точного определения окончания линии А рекомендуется повернуть отпечаток таким образом, чтобы линия А заняла вертикальное положение. Тогда ее направление определить значительно легче, и вероятность систематической ошибки завышения окончания линии (высокая при обычном способе определения) уменьшается.
52

Осевые трирадиусыОбычно осевые трирадиусы располагаются между тенаром и
гипотенаром по оси 4-й пястной кости (4-го пальца), между проксимальным краем и центром ладони. В редких случаях осевых трирадиусов нет вообще. Например, вертикальные папиллярные линии между проксимальными частями гипотенара и тенара могут расходиться в стороны у карпальной складки, не образуя трирадиуса (parting, p).
Обычно рядом с проксимальным краем ладони присутствует один осевой трирадиус. Его дистальный радиант длинный и может рассматриваться, как главная линия T (Рис.2.6). В случаях, когда трирадиусов несколько, линию Т прослеживают от наиболее проксимально расположенного трирадиуса. Самостоятельного таксономического значения линия Т не имеет, но ее ориентация помогает более точно проследить окончание линии А.
В зависимости от уровня, на котором находится трирадиус, различают низкий (t), промежуточный (t') и центральный варианты (t"). Для определения типа осевого трирадиуса (Sharma, 1964) отрезок между проксимальной и дистальной границами папиллярной зоны ладони, ориентированный по оси 4-го пальца, делят пополам, а нижнюю половину еще на три равные части (Рис.2.8). Трирадиусы в нижней трети относят к карпальным, в средней - к промежуточным, в верхней (и выше) - к центральным. В случае, когда осевой трирадиус лежит на рубеже между выделенными зонами, его в равной мере можно отнести к одному из двух типов трирадиусов.
Осевые трирадиусы t, t', t" находятся в пределах не более 1-1,5 см в латеральном направлении от оси четвертого пальца. Любой трирадиус, выходящий за эту границу, обозначается как боковой (tb) и обычно окаймляет узор на гипотенаре.
Менее точным является определение типа осевого трирадиуса по величине угла atd (между осевым трирадиусом и пальцевыми трирадиусами a и d). Величина угла atd зависит не только от уровня, на котором находится осевой трирадиус, но и от расположения пальцевых трирадиусов a и d , а также от размеров ладони и возраста испытуемых (David, 1981). При величине этого угла меньше 41° трирадиус относится к низким, от 41° до 60° - к промежуточным, больше 60° - к центральным (Geipel, 1961).
Л. Пенроз дает иную градацию величин угла; впрочем, его классификация была разработана в основном на клиническом
53

материале и не получила широкого применения в популяционных исследованиях.
Добавочные межпальцевые трирадиусыДобавочные трирадиусы обычно встречаются в 4 -м, в 3-м и
редко - во 2-м межпальцевых промежутках. Они лежат между пальцевыми трирадиусами или ниже их (Рис.2.9).
Рисунок 2.8. Способы диагностики типов осевых трирадиусов: А - по Sharma, 1964, Б - по Geipel, 1961 (с модификациями по: Гладковой, 1966).Figure 2.8. Approachs to a identification of axial triradii types: A - from Sharma, 1964, Б - from Geipel, 1961 (modified from: Gladkova, 1966).
Окончания главных ладонных линийДля описания окончаний главных линий края ладони делят на
14 полей (Рис.2.10, Табл.2.1). Поле 1 диагностируют, когда линия заканчивается в проксимальной части тенара. Поле 2 - если линия соединяется с карпальным осевым трирадиусом t или входит в линии его рамки. Поле 3 находится на ульнарном крае гипотенара ниже 4-го поля. В классической методике поле 4 точечное - это середина ульнарного края папиллярной области ладони (между 5 -й метакарпально-фаланговой и браслетной складкой), измеренная по продольной оси кисти. В связи со сложностью точной фиксации границ ульнарного края ладони рекомендуется расширить это поле на 4 мм с каждой стороны (Хить, 1983).
54

Рисунок 2.9. Добавочные трирадиусы a' и d' (по: Penrose, 1968). Figure 2.9. Accessory palmar triradii a' and d' (from: Penrose, 1968).
Рисунок 2.10. Схема расположения ладонных полей: A - по Cummins, Midlo, 1961; Б - в модификации по: Хить, 1983.Figure 2.10. The scheme showing position of palmar fields: A - from Cummins, Midlo, 1961; Б - modification from: Heet, 1983.
Под дистальной поперечной бороздой до верхней границы поля 4 расположена область 5', над складкой - 5". Если складка не доходит до края ладони, вся область кверху от дистального края поля 4 принимается за поле 5, которое делится на две равные части - 5' и 5". Когда главная ладонная линия заканчивается точно на границе обеих частей 5-го поля, рекомендуется фиксировать в формуле 5'.
55

Таблица 2.1. Краткое описание ладонных полей (по: Cummins, Midlo, 1961 в модификации Г.Л. Хить).
Table 2.1. Concise description of palm fields (from: Cummins, Midlo, 1961, modified by G.L. Heet).
Поле Описание1 Проксимальный край тенара. Ульнарная граница - карпальный
трирадиус t, или самый нижний из осевых трирадиусов.2 Карпальный трирадиус t и его горизонтальные радианты3 Проксимальный и ульнарный края гипотенара. Радиальная
граница - вертикальный радиант трирадиуса t. Верхняя граница - точка на 4 мм ниже середины ульнарного края ладони, нижняя - ульнарный радиант трирадиуса t
4 Середина ульнарного края ладони ±4 мм.5' Участок ульнарного края ладони от верхней границы 4-го поля до
дистальной поперечной складки.5" Участок ульнарного края ладони между дистальной поперечной
складкой и пястно-фаланговой складкой 5-го пальца.6 Пальцевый трирадиус d и его радианты7 4-й межпальцевый промежуток между трирадиусами d и c.8 Пальцевый трирадиус c и его радианты9 3-й межпальцевый промежуток между трирадиусами c и b.10 Пальцевый трирадиус b и его радианты11 2-й межпальцевый промежуток между трирадиусами b и a.12 Пальцевый трирадиус а и его радианты13 1-й межпальцевый промежуток между радиальным радиантом
трирадиуса a и пястно-фаланговой складкой 1-го пальца.
Следует отметить произвольную модификацию определения поля 4 и разделения частей поля 5, применяемую без пояснений в работах Х. Бреме и Р. Дженца (H. Brehme, R. Jantz). Как объяснил при личном контакте с Г.Л. Хить в 1991 г. первый - автор описаний многочисленных коллекций и публикаций - за 5" он принимал классический вариант этого поля, далее равный по длине участок книзу от складки считал полем 5'. Такой же по протяженности отрезок в проксимальном направлении считался полем 4, а следующий за ним - до ульнарного радианта трирадиуса t - полем 3. При этих условиях происходит систематическое завышение индекса Камминса, усредняющего окончания линий А и D, что становится особенно заметным в группах с более горизонтальным ходом линии А и значительно снижает возможности сравнительного анализа.
56

Поля 6, 8, 10, 12 диагностируют в тех случаях, когда главная ладонная линия заканчивается в пальцевых трирадиусах d, c, b и a, соответственно, или сливается с радиантами этих трирадиусов.
Четвертый, 3-й и 2-й межпальцевый промежутки образуют 7-е, 9-е и 11 -е ладонные поля. Поле 13 занимает 1 -й межпальцевый промежуток между радиальным дистальным радиантом трирадиуса a и пястно-фаланговой складкой 1-го пальца.
Обозначения полей, в которых оканчиваются главные ладонные линии, составляют формулу этих линий. Запись ведется от линии D к A; цифры разделяются точками. Окончание линии T (вертикальный радиант осевого ладонного трирадиуса) обычно не включают в формулу.
Встречаются сложные для интерпретации варианты расположения и окончания главных ладонных линий. Отсутствие пальцевого трирадиуса и главной ладонной линии (С, редко В) отмечается в формуле символом «О». Обрыв линии (С, редко В) диагностируют, когда пальцевый трирадиус присутствует, но его проксимальный радиант очень короткий. В таком случае конфигурация гребней напоминает шатровую дугу, но выпуклую проксимально, а не дистально, как на пальцевых подушечках. Если проксимальный радиант едва намечен, в формуле вместо окончания ладонной линии ставят символ «x», если он прослеживается более отчетливо - «X».
Третий вариант несформированных главных линий обозначают номером поля, из которого она выходит. При этом линия изгибается назад и соединяется сама с собой или с пальцевым радиантом трирадиуса, от которого она стартует. Например, для линии С записывают окончание в поле 8.
При соединении главных линий друг с другом они приходят в четные поля. Линии могут сливаться на ограниченном участке, а их окончания быть независимыми. Если линия раздваивается, в формуле записывают двойное обозначение через косую черту (на первом месте рекомендуется указывать окончание, наиболее удаленное от начала линии). Когда на каком-то участке линии разделяют не более двух гребней, фиксируют сначала фактическое окончание линии, которое учитывается при подсчете, а потом в скобках возможный (соседний) вариант.
Проксимальные радианты добавочных трирадиусов d' и a' продолжаются в длинные линии, имитирующие D и A и определяющие ход гребней в дистальной части ладони. В таких
57

случаях в формуле указывают поля, где заканчиваются эти дополнительные линии. Рекомендуется сначала фиксировать окончание линии в более удаленном от дельты поле. Если основные радианты пальцевого и межпальцевого трирадиусов не соединяются друг с другом, обозначения ладонных полей разделяют тире, если соединяются - косой чертой. При вычислении индекса Камминса учитывается более высокое окончание для линии D и более низкое окончание для линии А.
Распределение частот встречаемости полей, в которых заканчиваются все четыре главные ладонные линии, громоздко и его сложно использовать в масштабных исследованиях дерматоглифики многих групп. Некоторые авторы, сторонники гипотезы наследования признаков комплексами, указывают процент наиболее часто встречающейся в группе формулы окончаний главных линий (например, 11.9.7.5, 9.7.5'.3 и др.).
Значительно чаще применяется индекс Камминса или главных ладонных линий, в который входят усредненные окончания линий A и D, определяющие общее направление гребней на ладони. Для этого поля ладони получают новую нумерацию (Табл.2.2). Для вычисления индекса поля ладони объединяют в два ряда: проксимально-ульнарный для окончаний линии А идистальный - для линии D. Цифры, соответствующие символам полей, в которых оканчиваются эти линии, суммируются. Например, когда линии А приходит в поле 5", а линия D - в поле 11, индекс равен 12 (6+6). Если линия А заканчивается в поле 1, а линия D в поле 7, величина индекса - 3 (1+2).
С целью генерализации окончаний главных ладонных линий их объединяют в типы. Для линии А выделено три типа: 1(1+2),3(3+4) и 5(5'+5"+6). Для линии D - 7(7+8+X+0), 9(9+10) и11(11+12+13). Окончания линии В разделяют на дистальный (6+7+8+9+10), ульнарный (3+4+5'+5") варианты и отсутствие или редукция (О+Х). Для линии С предложены ульнарный (4+5'+5"+6+7), радиальный (9+10+11+12+13), проксимальный (8+Х) типы и вариант «отсутствие» (0) (Plato, 1976).
Узорные поляКожные рисунки на каждом узорном поле описывают
отдельно. Обозначения всех узоров составляют узорную формулу. Узорные поля фиксируют в следующем порядке: гипотенар, тенар/I межпальцевая подушечка, П, III, IV межпальцевые подушечки. В
58

случае сомнений относительно типа узора рядом с основной записью в скобках указывается альтернативный вариант, не используемый при подсчетах. Когда в узорной области находятся два дискретных узора, их символы записывают в формуле через косую черту.
Таблица 2.2. Соответствие символов ладонных полей обозначениям для вычисления индекса главных ладонных линийTable 2.2. Conformity of symbols of palmar fields to designations for calculation of the Main Line Index
Проксимально-ульнарный ряд (для линии А)
Символы ладонных полей 1 2 3 4 5' 5" 6 7
Новые обозначения полей 1 2 3 4 5 6 7 8
Дистальный ряд (для линии D)
Символы ладонных полей 6 7 8 9 10 11 12 13
Новые обозначения полей 1 2 3 4 5 6 7 8
Г ипотенар. Кожные гребешки гипотенара могут формировать простые дуги, открытые поля, мультипликации и следы узоров. Истинными узорами считаются спирали, завитки, составные узоры, петли и шатровые дуги (Рис.2.11).
Чаще всего кожные гребни гипотенара ориентированы косопоперечно. Обычно книзу количество гребешков заметно увеличивается, и они расходятся веером по гипотенару из точки близ центра ладони. Такой ход папиллярных линий называют мультипликацией (М).
Рисунок гипотенара морфологически связан с осевыми трирадиусами, радианты которых образуют границы или проникают внутрь узора. Ульнарный радиант промежуточного осевого трирадиуса (t') разделяет гипотенар на дистальную и проксимальную части. В случае присутствия указанного трирадиуса символы узоров дистального, а затем проксимального участков записываются в узорную формулу через косую черту.
Истинные завитки (W) образованы концентрическими гребнями, но в отличие от завитков на пальцах, сопровождаются тремя трирадиусами. При этом ульнарный (краевой) трирадиус (f)
59

иногда отсутствует, чаще при плохо пропечатанной ладони. По направлению гребней можно предположить, что точка трирадиуса лежит вне области ладони. Такие же «потенциальные» трирадиусы могут сопровождать петли и шатровые дуги. Завиток, центр которого образован спиральными гребнями или сдвоенными петлями, обозначается Ws.
Петли (L) на гипотенаре морфологически не отличаются от пальцевых, но открываются не только на ульнарном и радиальном краях ладони, но и на карпальном (проксимальном) крае: ульнарные (L“), радиальные (Lr) и карпальные (Lc) петли соответственно.
При необходимости можно разделить узоры по размеру: маленькие завитки с гребневым счетом не больше 8 обозначают строчной буквой w , а маленькие петли - l.
Шатровая дуга (T) гипотенара по строению похожа на пальцевую, только вершину дуги ограничивает трирадиус. Дуга называется радиальной, если ее основание обращено радиально (Tr).
Ход гребней радиальной дуги исключает присутствие осевых трирадиусов. Основание ульнарной шатровой дуги (T ) направлено ульнарно-дистально, а карпальной (T ) - ульнарно-карпально.
Простые дуги (A) сопровождаются трирадиусом в области вершины и ориентированы так же, как шатровые дуги (Au, Ar, Ac). Простые дуги являются формой открытых полей (O) - редких узоров, гребни которых лежат прямо и параллельно.
Следы узоров (V), как правило, представлены либо группой резко сходящихся в одну точку гребней, либо системой прямых параллельных линий, отличных по направлению от окружающих. В отличие от истинных узоров их следы обычно не имеют круто изгибающихся гребней.
В случае, когда след все же демонстрирует сходство с определенным узорным типом, перед знаком V пишут символ соответствующего узора (например, LuV).
Перечисленные узоры гипотенара могут образовывать парные комбинации, состоящие из простых узоров: два истинных узора, истинный узор и дуга или открытое поле, две дуги или два открытых поля и т.д. В формуле вначале записывают узор расположенный дистально; обозначения узоров разделяют косой чертой.
Как правило, узоры разграничивает ульнарный радиант промежуточного трирадиуса, но они выглядят обособленными и без этого разделения: гребни одного узора не заходят в площадь другого (Рис.2.11).
60

Рисунок 2.11. Узоры гипотенара (обозначения в тексте). Figure 2.11. Types of hypothenar (see text for designations).
61

Особый рисунок, состоящий из двух петель, в котором гребни перетекают из одной петли в другую, называется спиралью, или S- узором . Необычное сочетание узоров, которое нельзя описать в терминах исходных типов, обозначают Y (шатровая дуга с основанием, по бокам изогнутым к вершине).
Внутри петли, завитка, реже других гребневых структур гипотенара могут находиться другие узоры, придавая им сложный характер. Например, радиальная петля может содержать центральный фрагмент в форме завитка, карпальная дуга - ульнарную петлю и проч. Если гребневой счет такого фрагмента превышает 8 единиц, он оценивается, как самостоятельный узор гипотенара (Lr/W, A c/Lu). Если гребневой счет внутреннего узора не больше 8 - он отмечается в формуле строчной буквой после основного узора (Lrw, Aclu).
В радиальной области гипотенара изредка встречается паратенарный узор (обозначение P-th) - узкая вертикальная дистально открытая петля посередине ладони. Она фиксируется в формуле как рисунок гипотенара, хотя не ясно, связана ли с ним морфологически.
Тенар и 1-я межпальцевая подушечка. Гребневой рельеф тенара и 1-й межпальцевой области анатомически тесно связаны и описываются вместе. Когда узор трудно отнести к той или иной области, его обозначают одним символом. Если положение узоров можно дифференцировать, в формуле отмечают два символа, разделенные косой чертой (первым оценивают узор тенара). Часто обе подушечки заняты открытым полем (О). Формирующие его гребни образуют пологую дугу или угол, открытые радиально. Истинные узоры (петли (L) и завитки (W)) встречаются нечасто. Ориентация петель такова, что почти всегда на тенаре они направлены карпально, а на 1-й подушечке - радиально. Следы узора на тенаре и 1-й межпальцевой подушечке относят к истинным узорам, если в обеих зонах они формируют рисунок, похожий на две петли (радиальную и карпальную), разделенные системой поперечных гребней (фигура Беттмана). Узор или след узора с центром в середине тенара относится и к тенару, и к 1 -й межпальцевой области. Возможны комбинации рисунков: узор или след узора в области тенара и открытое поле на 1 -й подушечке; узор или след узора в 1-й межпальцевой области и открытое поле на
62

тенаре; узор или след узора и на тенаре, и на 1 -й межпальцевой подушечке.
Второе, третье и четвертое межпальцевые поля (подушечки). Межпальцевые поля II, III, IV располагаются между пальцевыми трирадиусами a и b, b и c, с и d соответственно. В случаях, когда пальцевый трирадиус отсутствует или сильно смещен, межпальцевые поля можно разделить по средней точке в основании пальца. Гребни межпальцевых подушечек формируют открытое поле (О), в том числе его разновидность - мультипликацию (М), след узора (V) или истинные узоры (петли и завитки), а также составные узоры.
Завитки (W, w) чаще всего маленькие и сопровождаются дополнительными трирадиусами. Петли обычно открываются в ближайший межпальцевый промежуток, т.е. дистально. Поскольку другая ориентация петель бывает редко, в формуле направление петель не указывают. Петля, не сопровождающаяся добавочным трирадиусом, обозначается L или l, если она маленькая (не больше 8-ми гребешков). Аналогичным образом петля с добавочным трирадиусом фиксируется как D или d.
Нередко следы узора сформированы круто сходящимися гребнями, иногда эллипсоидальными. В обрамлении изогнутого потока гребней могут находиться параллельные вертикальные гребешки.
Открытые поля представлены дугообразными линиями и мультипликациями - многочисленными гребнями около пальцевых трирадиусов на уровне расширенной части узорного поля.
Если в одном межпальцевом поле находятся два рисунка, они отмечаются в формуле через косую черту, начиная с узора, расположенного радиально. Двойной записью отмечают в формуле варианты, когда узор (или след узора) смещен в сторону от центра подушечки и не занимает ее по всей ширине (O/L, L/O и т.д.). Чаще всего это отмечается для IV подушечки.
В некоторых медицинских, конституциональных и генетических исследованиях используется топологическая система дерматоглифики (Penrose, 1965), в которой учитывают только два базовых элемента гребневого рельефа: трирадиусы и петли. Суммарное количество трирадиусов или петель отражает общую узорность ладони и кисти в целом (Рис.2.12). В этнической дерматоглифике метод не нашел широкого применения, так как его
63

использование приводит к потере информации о распределении узоров по зонам ладони. Между тем, именно по указанным характеристикам наблюдаются основные различия между популяциями.
Г ребневой счет ладониОбычно определяют гребневой счет между пальцевыми
трирадиусами a и b , поскольку они всегда присутствуют на ладони, и чаще всего разделены открытым узором. Трирадиусы в подсчет не включаются. В некоторых работах используют гребневой счет между трирадиусами b и c, c и d, трирадиусами d или a и осевым трирадиусом t .
Рисунок 2.12. Варианты узорности кисти: а - 4 трирадиуса, 0 петель; б - 18 трирадиусов, 14 петель (по: Penrose, 1965).Figure 2.12. Variants of palmar papillars: a - 4 triradii, 0 loops; б - 18 triradii, 14 loops (from: Penrose, 1965).
64

Рисунок 2.13. Классификация минуций (по: Cummins, Midlo, 1961). Figure 2.13. Classification of minutiae (from: Cummins, Midlo, 1961)
2.3. Минуции (minutiae)
В судебной медицине и для идентификации личности используют мелкие элементы кожного рельефа (минуции). Их популяционные различия не изучены.
Выделяют семь основных вариантов минуций. Островок (island) или точка (point) - маленький округлый гребешок с одной потовой порой (Рис.2.13). Короткий гребешок (short ridge) - включает от двух до пяти потовых пор. Вилка (fork), реже Y- структура (Y-formation) - бифуркация гребня. Ограда (enclosure), озеро (lake) или кольцо (ring) - замкнутая фигура. Гребенка (comb)- фигура, в которой три или более параллельных гребешков соединяются с гребнем, лежащим к ним почти под прямым углом. Конец (end) - участок обрыва гребня. Вставочная линия (interstitial line) - узкий гребешок, ширина которого обычно меньше половины ширины окружающих гребней. Вставочная линия может быть ниже остальных гребней и крайне редко несет потовые поры. Эту минуцию не включают в гребневой счет, если на ней не видны потовые поры. Дисплазия (dysplasia) - участки несформированных или дезорганизованных гребешков. По степени нарушений кожного рельефа различают гипоплазию - недоразвитие и аплазию - отсутствие эпидермальных гребней. Более подробные классификации минуций учитывают детали, состоящие из простых элементов, а также редко встречающиеся признаки (Ивашков, 1999).
65

2.4. Первичная статистическая обработка
Первичная обработка данных может производиться как с помощью компьютерных программ (что предпочтительнее), так и ручным способом, с помощью калькулятора. Индивидуальные данные записываются на листе в горизонтальной строке в следующем порядке: № индивидуума (п/п) - главные ладонные линии (D, C, B, A) - осевой трирадиус ладони - узоры на гипотенаре (Hy), тенаре (Th/I), II-IV межпальцевых подушечках - типы узоров на пальцах от I к V. На левой половине листа отмечаются признаки левой руки, на правой - правой. Обязательно обозначаются серия или группа, дата и автор сбора материалов, пол и число индивидов, место исследования, год и автор обработки серии. Ниже приводится образец дерматоглифического бланка для групповых данных при обработке материала вручную (Табл. 2.3)2. В каждой графе обозначаются абсолютные и относительные (в процентах от числа наблюдений) частоты. В графах линии А и D приведены новые номера полей, на которые следует умножать абсолютные частоты признаков при вычислении индекса Камминса. Полученные величины суммируются и делятся на число ладоней.
Средние баллы окончаний главных ладонных линий A и D вычисляются путем умножения абсолютных частот типов окончаний на 1, 3, 5 в первом случае, и 7, 9, 11 во втором, суммированием и делением полученных значений на число ладоней (равное 2n).
2 В Таблице 2.3 представлен бланк, заполненный на примере обработки данных для группы чувашей из г.Чебоксар (n=76; пол: женский; дата обследования: январь 1991 г.; исследователи: И.А. Славолюбова, Е.Г. Лебедева).
66

O n'О
Поля (новые номера)
DПоля
с Типы пальцевых узоровL R L+R, % L R L+R, % L L, % R R, % L+R, %
7(2) 16 8 24 15.79 5' 4 1 5 3.29 А+Т 21 5.53 16 4.21 37 4.878(3) 1 1 2 1.32 5" 12 7 19 12.50 R 19 5.00 15 3.95 34 4.479(4) 34 27 61 40.13 6 1 1 2 1.32 и 220 57.89 208 54.74 428 56.3210(5) 4 3 7 4.60 7 22 14 36 23.68 R+U 239 62.89 223 58.68 462 60.7911(6) 21 36 57 37.50 8 2 0 2 1.31 W 120 31.58 141 37.11 261 34.3412(7) 0 1 1 0.66 9 21 46 67 44.08 п 380 100.00 380 100.00 760 100.00
п 76 76 152 100.00 10 0 1 1 0.66 Dlio= 100*A/L= 8.011с 7.01 8.51 7.76 X 11 4 15 9.87 (L+2W)/10=12.95
Поля (новые номера)
А 0 3 2 5 3.29100*A/W=14.18 100*W/L= 56.49
L R L+R, % п 76 76 152 100.00
1(1) 14 2 16 10.53Поля
В Осевые трирадиусы ладони2(2) 4 - 4 2.63 L R L+R, % L R L+R, %3(3) 45 43 88 57.89 3 3 1 4 2.63 0 1 1 2 1.324(4) 2 7 9 5.92 4 0 0 0 0.00 t 47 44 91 59.875 '(5) 11 22 33 21.71 5' 26 16 42 27.63 t' 15 12 27 17.765" (6) 0 1 1 0.66 5" 22 19 41 26.97 t" 7 6 13 8.556(7) 0 1 1 0.66 6 4 3 7 4.61 tt' 2 7 9 5.92
п 76 76 152 100.00 7 21 36 57 37.50 tt" 3 5 8 5.26Типы линии А L R L+R, % 8 0 1 1 0.66 t't" 1 0 1 0.66
1(1+2) 18 2 20 13.16 п 76 76 152 100.00 t't' 0 1 1 0.663(3+4) 47 50 97 63.81 Истинные узоры на
ладонных подушечкахtt't" - - - -
5(5+6+7) 11 24 35 23.03 - - - -
мА 2,82 3.58 3.20 100.00 L R L+R, % n 76 76 152 100.00
Типы линии DD Ну 26 29 55 36.18 Добавочные межпальцевые трирадиусы
L R L+R, % тм 10 5 15 9.87 L R L+R, %7(+8+Х+0) 17 9 26 17.11 II 0 3 3 1.97 IV 4 4 8 5.26
9(+10) 38 30 68 44.73 III 7 25 32 21.05 III 1 0 1 0.6611 (+12) 21 37 58 38.16 IV 26 22 48 31.58 II 0 4 4 2.63
MD 9.11 9.74 9.43 100.00 X 5 8 13 8.55
н чSS 5_н 2 •К. £ W -• то.с
оСГС1
со.
н>>
с/5
§
инмс
Таблица 2.3. Д
ерматоглифический бланк
ИЭА РАН
(составитель

Г лава 3. Методы статистического анализа дерматоглифических данных.
Для определения степени близости и различий выборок на разных таксономических уровнях, а также оценки дифференцирующего значения конкретных признаков в дерматоглифике широкое применение получил ряд статистических методов. Часть этих методов используется исследователями также при анализе выборок при работе с другими системами антропологических признаков, часть включает в себя сугубо специфические приемы анализа, разработанные специально для изучения дерматоглифических материалов.
3.1. Метод межгрупповой ш калы
Метод межгрупповой шкалы в дерматоглифике был впервые предложен и введен в исследовательскую практику Г.Л. Хить (1973; 1975; 1983). Он основан на применении группы ключевых признаков и состоит из трех статистических приемов: вычисления обобщенных дерматоглифических расстояний между выборками, построения полигонов и расчета условной величины расовых комплексов.
Обобщенные дерматоглифические расстояния (вдальнейшем - ОДР) между любыми двумя выборками представляют собой среднюю стандартизованную разницу по пяти основным признакам (за исключением Th/I), либо по шести (с учетом последнего признака). Вычисляемые без учета знака разницы между двумя группами в величинах каждого признака выражаются в процентах евразиатской шкалы (амплитуды), служащей стандартом. Максимальное и минимальное значения шкалы установлены эмпирическим путем по данным более 600 мужских и женских популяционных выборок с территории Северной Евразии от Фенноскандии до Китая (Табл. 3.1).
Расчет ОДР проводится по формуле:IPlKXArPllOB,! ^ |1с<Л|-1с<В(| ̂ ^ |HyiA|-Hy(Bl| ̂ |ДМТ|А|-ДМТ<В||
68

где A и B - выборки, между которыми производится вычисление обобщенного расстояния, a d - евразиатская амплитуда популяционных значений признака.
Таблица 3.1. Евразиатская шкала вариаций (d) и лимиты популяционных величин ключевых признаков (по: Хить, 1983).Table 3.1. Eurasian scale of variations (d) and limits of population values of key traits (from: Heet, 1983).
Dli0 Ic t Hy ДМТ Th/I
в ? в ? в ? в ? в ? в ?minmax
10.8715.98
9.5616.33
6.489.68
6.629.35
43.895.2
36.094.0
7.345.7
10.755.4
2.446.6
1.245.0
0.027.5
0.924.0
d 5.11 6.77 3.20 2.73 51.4 58.0 38.4 44.7 44.2 43.8 27.5 23.1
Таким образом, ОДР - это расстояние, которое выражает величину различий в процентах евразиатской шкалы, приходящуюся на каждый признак.
Для вычисления матрицы ОДР для серии исследуемых выборок Б.А. Козинцевым написана статистическая программа «DERM». Матрицы ОДР подвергались кластерному анализу парногрупповым методом с взвешиванием.
На основании попарных различий между группами строились вариационные кривые распределения частот ОДР на разных таксономических уровнях. Выборочные кривые распределения ОДР на территориальном уровне в пределах этносов и на уровне этнических групп в регионах свидетельствуют о сходной дифференциации выборок, принадлежащих европеоидной и монголоидной расам. Наиболее отчетливым образом это сходство проявляется при суммировании кривых каждого таксономического уровня по всем регионам бывшего СССР. Использование параметров суммарного вариационного ряда (x и s) позволило произвести ранжирование величин ОДР на малую, среднюю и большую категории в пределах каждого уровня для выборок европеоидов и монголоидов. По таблицам нормального интеграла вероятности для площади кривой Ф(и) были вычислены границы указанных категорий (малые, средние и большие ОДР), каждая из которых охватывает около 33% наблюдений. Крайние категории был подразделены на подкатегории. Указанные процедуры вычислений и оценка параметров вариационных рядов и индексов
69

производились для каждого пола отдельно с использованием соответствующей евразиатской амплитуды. В отдельных случаях, в разделах, посвященных половому диморфизму и анализу рас и их подразделений, применялась совмещенная шкала, охватывающая «мужские» и «женские» лимиты евразиатской амплитуды (Табл. 3.2).
Таблица 3.2. Лимиты значений ключевых признаков в рамках совмещенной для обоих полов шкалы вариаций.Table 3.2. Limits of key traits values within the range of variability combined for both sexes.
Признак Dl10 Ic t Hy ДМТ Th/Imin 9.56 6.48 36.0 7.3 1.2 0.0max 16.33 9.68 95.2 55.4 46.6 27.5
d 6.77 3.20 59.2 48.1 45.4 27.5
Установлен средний уровень дифференциации (средняя величина ОДР) для каждого пола с учетом таксономического уровня сопоставления. Это дает возможность оценивать анализируемую величину ОДР на фоне региональной или тотальной (охватывающей обе большие расы в пределах Евразии) дифференциации (Табл. 3.3). Выявлена иерархия уровней: на этническом уровне население наиболее разнородно, на уровне локальных рас - наиболее гомогенно (вероятно, за счет резкого ослабления выборочного эффекта при объединении популяций). Внутриэтническая популяционная дифференциация уступает межэтнической всюду (в Восточной Европе, на Кавказе, в Сибири), кроме Средней Азии, где территориальные группы в пределах этноса оказываются более разнородными, чем этнические группы в пределах региона (Хить, 1983, с.28-38).
Градации обобщенных дерматоглифических расстояний для разных таксономических уровней сопоставления, позволяющие оценить степень различий между выборками в масштабе евразиатской шкалы, приведены в Таблице 3.4. Установленные категории позволяют сохранять постоянный масштаб оценки попарных ОДР. Изменение лимитов евразиатской амплитуды, которая почти исчерпывает мировую изменчивость признаков, не отражается на этой оценке (ОДР в этом случае попадают в разряд крайне высоких и крайне низких значений); лишь при сравнении полигонов, отражающих комбинации признаков в группе,
70

происходит деформация некоторых полигонов или выход радиуса за пределы окружности (см. далее).
Поскольку метод межгрупповой шкалы отражает расстояния между группами в процентах евразиатской шкалы, принятой за стандарт, эта разбивка ОДР на категории позволяет контролировать масштаб различий между любыми группами независимо от их общего набора, таксономического ранга и расовых градиентов признаков. С другой стороны, как и все способы статистического анализа, метод межгрупповой шкалы имеет определенные недостатки: таксономическая неравноценность признаков в этой системе расчетов не учитывается. Для контроля полученных результатов используются дополнительные статистические приемы.
Таблица 3.3. Средние величины ОДР на разных таксономических уровнях (по: Хить, 1983).Table 3.3. Average values of generalized dermatoglyphic distances (GDD) for different taxonomic levels (from: Heet, 1983)
РегионТерриториальный
(внутриэтнический)уровень
Этническийуровень
Уровеньлокальных
расВосточная Европа в - 11.9 в - 14.1 в - 7.5
? - 9.7 ? - 13.3 ? - 7.0Кавказ в - 13.7 в - 16.4 в - 8.8
? - 10.9 ? - 13.7 ? - 7.1Средняя Азия в - 13.4 в - 10.1 в - 4.9
? - 15.6 ? - 13.8 ? - 4.8Северная и Центральная в - 12.6 в - 14.8 в - 9.9Азия ? - 11.2 ? - 14.3 ? - 9.5Суммарно в - 12.6 в - 15.4 в - 8.8
? - 12.6 ? - 13.9 ? - 8.2
Примечание: величины ОДР сравнимы лишь в пределах пола.Note: values of GDD are comparable only within the bounds of same sex.
Построение круговых полигонов позволяет визуально оценить различия между выборками по комбинации ключевых признаков и классифицировать их даже при большом количестве групп. Для того чтобы все признаки стали соизмеримыми, необходимо найти положение группы на евразийской шкале вариаций относительно минимума, после чего найденная величина
71
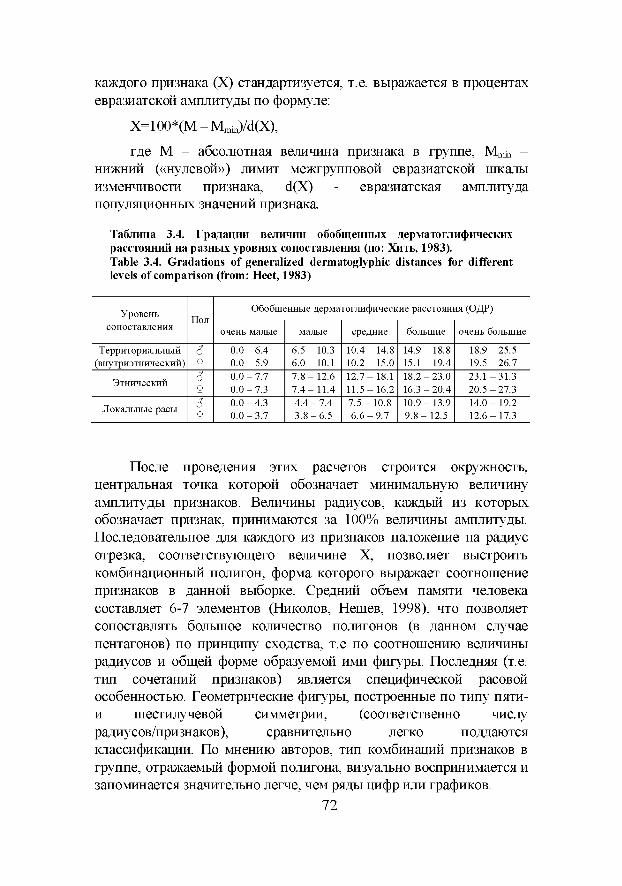
каждого признака (Х) стандартизуется, т.е. выражается в процентах евразиатской амплитуды по формуле:
Х=100*(М - Mmm)/d(X),
где М - абсолютная величина признака в группе, Mmin - нижний («нулевой») лимит межгрупповой евразиатской шкалы изменчивости признака, d(X) - евразиатская амплитуда популяционных значений признака.
Таблица 3.4. Градации величин обобщенных дерматоглифических расстояний на разных уровнях сопоставления (по: Хить, 1983).Table 3.4. Gradations of generalized dermatoglyphic distances for different levels of comparison (from: Heet, 1983)
Уровень ПолОбобщенные дерматоглифические расстояния (ОДР)
сопоставления очень малые малые средние большие очень большие
Территориальный(внутриэтнический)
в?
0.0 - 6.4 0.0 - 5.9
6.5 - 10.3 6.0 - 10.1
10.4 - 14.8 10.2 - 15.0
14.9 - 18.8 15.1 - 19.4
18.9 - 25.5 19.5 - 26.7
Этнический в?
0.0 - 7.7 0.0 - 7.3
7.8 - 12.6 7.4 - 11.4
12.7 - 18.1 11.5 - 16.2
18.2 - 23.016.3 - 20.4
23.1 - 31.3 20.5 - 27.3
Локальные расы в?
0.0 - 4.3 0.0 - 3.7
4.4 - 7.4 3.8 - 6.5
7.5 - 10.86.6 - 9.7
10.9 - 13.9 9.8 - 12.5
14.0 - 19.2 12.6 - 17.3
После проведения этих расчетов строится окружность, центральная точка которой обозначает минимальную величину амплитуды признаков. Величины радиусов, каждый из которых обозначает признак, принимаются за 100% величины амплитуды. Последовательное для каждого из признаков наложение на радиус отрезка, соответствующего величине X, позволяет выстроить комбинационный полигон, форма которого выражает соотношение признаков в данной выборке. Средний объем памяти человека составляет 6-7 элементов (Николов, Нешев, 1998), что позволяет сопоставлять большое количество полигонов (в данном случае пентагонов) по принципу сходства, т.е по соотношению величины радиусов и общей форме образуемой ими фигуры. Последняя (т.е. тип сочетаний признаков) является специфической расовой особенностью. Геометрические фигуры, построенные по типу пяти- и шестилучевой симметрии, (соответственно числу радиусов/признаков), сравнительно легко поддаются классификации. По мнению авторов, тип комбинаций признаков в группе, отражаемый формой полигона, визуально воспринимается и запоминается значительно легче, чем ряды цифр или графиков.
72

3.2. Расовые градиенты и расовые комплексы
Понятие расового градиента означает направление изменения признака от меньших к большим величинам на уровне больших рас и их подразделений. Такие градиенты могут быть установлены для сопоставления любых пар рас при использовании достаточного количества наблюдений, приходящихся на каждый признак. При малом числе выборок градиенты могут оказаться недостоверными и дать ложную картину направления различий. Аналогичное содержание имеют географические градиенты, фиксирующие направление различий между северными и южными, западными и восточными и иными парными комбинациями сопоставляемых субрасовых таксономических единиц.
В дерматоглифике твердо установлены расовые градиенты основных признаков для хорошо изученных европеоидной и монголоидной рас (Хить, 1973; 1983), а во внутрирасовом масштабе- для их северных и южных подразделений (Хить, 1983; 2010) (Табл. 3.5). Достаточно надежно определены также расовые градиенты для негроидной расы при сопоставлении ее с монголоидами и европеоидами (Хить, Кейта, 1981). Менее благоприятна ситуация в отношении остальных рас, характеристики которых базируются на значительно меньшем, а иногда и малом числе выборок (за исключением дельтового индекса - наиболее хорошо изученного признака во всех расах).
Расовые градиенты на уровне сопоставления европеоидов и монголоидов, а также географические градиенты во внутрирасовом масштабе могут быть использованы для вычисления расовых комплексов, отражающих содержание (в процентах) различных расовых компонентов в выборке: европеоидно-монголоидногокомплекса (ЕМК), прежде обозначавшегося как монголоидный, или восточный, комплекс (МК, ВК) (Хить, 1975; Хить, 1983), а также североевропеоидного (СЕК), северомонголоидного (СМК) и других, в зависимости от целей исследования.
Таким образом, расовый комплекс - это выраженное в процентах евразиатской шкалы условное содержание( приходящееся на признак) в выборке какого-либо расового компонента на фоне другой расы. Для его вычисления за минимальный лимит (соответствующий 0%) принимается тот лимит евразиатской амплитуды, величина которого более характерна для «фоновой» расы (например, европеоидов). За максимальный лимит (100%)
73

принимается величина второго лимита амплитуды, более соответствующая расовому компоненту, который предполагается выявить (например, монголоидный). Затем находится разница по каждому из пяти ключевых признаков между выборочным значением и минимальным лимитом. Далее усреднением суммы частных стандартизованных (т.е. выраженных в процентах межгрупповой амплитуды признаков) величин указанной разницы вычисляется значение ЕМК. В этой аббревиатуре первая буква обозначает «фоновую» (европеоидную) расу, вторая - выявляемый расовый компонент (монголоидный).
Таблица 3.5. Градиенты изменчивости ключевых признаков на разных таксономических уровнях (по: Хить, 2010).Table 3.5. Gradients of key traits variation in different taxonomic levels (from: Heet, 2010)
Уровень,Раса
Признак
Межрасовый уровень
(европеоиды - монголоиды)
Внутрирасовый уровень С 1
Ю
Европеоиды МонголоидыDI10 i iIc i tt t tHy i tДМТ i tTh/I ^ (?) i t
Поставив обратную задачу, можно вычислить монголоидноевропеоидный комплекс (МЕК), т.е. условную долю европеоидного компонента. Оба комплекса находятся в обратной зависимости и дополнительны по отношению друг к другу, составляя в сумме 100%. Величина ЕМК в Северной Евразии, т.е. условная доля «монголоидности», приходящаяся в среднем на признак, варьирует в мужских группах от 20 до 80%, никогда не снижаясь до 0% и не достигая 100% (для этого требуется невыполнимое условие, чтобы значения признаков в группе точно совпадали бы со значениями минимальных или максимальных лимитов межгрупповой шкалы). Эмпирическая величина ЕМК может быть пересчитана в стандартизованную, с учетом евразийской амплитуды показателя
74

(Хить, 1986). В пределах каждого из основных регионов Северной Евразии, а также в пределах европеоидной и монголоидной рас популяционные величины ЕМК распределяются по нормальному типу с различной степенью трансгрессии, но суммарная евразиатская кривая имеет две вершины, что не позволяет использовать параметры объединенного ряда (Хить, 1983, с.43-50).
Таким образом, величина расового комплекса вычисляется с учетом расовых градиентов изменчивости ключевых признаков и лимитов их евразиатской амплитуды, значения которых приведены в Таблице 3.1. Например, для монголоидов характерно сочетание высоких величин дельтового индекса и частоты проксимального трирадиуса t с низкими значениями индекса Камминса, узорности гипотенара и частоты добавочных трирадиусов, следовательно, величина ЕМК для мужской выборки A будет вычисляться по формуле:
Р1.о(д>-10.87 + 9.68-1с<л>+<)л1-43 .8< 45.7-Hy.Ai ̂ 46.6-ДМТ,л,
Аналогичным образом вычисление величины СЕК для этой выборки производится по формуле:
l5.98-Dl.o,A)+9.68-[c(A>+t(A).43.8+45.7-Hy,A> ̂46.6-ДМТ,л,
Необходимо также помнить, что по величине любого расового комплекса невозможно строго судить о доле расовых компонентов в выборке. Принцип вычисления комплексов в каждом случае предполагает наличие в анализируемой группе лишь двух расовых компонентов - фонового и искомого. Это обстоятельство накладывает определенные ограничения на применение данного статистического приема, который не может использоваться при большем или меньшем числе компонентов, вошедших в состав группы. Однако, зная величины всех ключевых признаков у всех рас и их подразделений и соответствующие градиенты, можно вычислить все возможные комплексы для каждой из них. Суммируя эти величины и разделив полученное число на количество рас, можно получить расовый комплекс (РК), количественно выражающий свойственные каждой расе особенности на «общечеловеческом» фоне и сопоставить расы по степени
75

выраженности специфических расовых черт. Главное условие корректности этого показателя состоит в достаточном числе выборок, приходящихся на каждый признак каждой расы, иначе суждение о градиенте может оказаться ошибочным.
3.3. Анализ главных компонент
В качестве одного из основных статистических методов для оценки межгрупповых различий выборок, а также для оценки дифференцирующего значения дерматоглифических признаков используется метод главных компонент.
Метод применяется для уменьшения размерности исходных данных при минимальных потерях информации. Его применение для антропологических исследований было предложено и обосновано в работах В.Е. Дерябина. Строго говоря, анализ главных компонент может быть использован только для изучения внутригрупповой изменчивости; корректное исследование межгрупповой дифференциации осуществляется при помощи канонического дискриминантного анализа (Дерябин, 1983; 1998).
Некорректность применения метода главных компонент для межпопуляционного анализа связана с тем, что данный метод, в отличие от канонического анализа, не позволяет учитывать межгрупповые и внутригрупповые закономерности скоррелированности и изменчивости признаков. Рассмотрение одной лишь межгрупповой изменчивости, описываемой дисперсиями или коэффициентами корреляции, полученными на основе средних величин, является недостаточным. В ней неизбежно будет присутствовать определенный внутригрупповой компонент, который по смыслу таксономического анализа следует устранить (Дерябин, 2005). Но поскольку группа основных признаков дерматоглифики не связана внутригрупповыми корреляциями, метод главных компонент в этом случае является вполне допустимым приемом для проведения межпопуляционного анализа .
3.4. Параметрические и непараметрические критерии оценки значимости различий между выборками.
Широкое распространение для анализа значимости различий между величинами дерматоглифических признаков в выборках в практике как отечественных, так и зарубежных специалистов
76

приобрел t-критерий Стьюдента. Корректное использование t- критерия должно опираться на две исходные посылки: 1)распределение исследуемого признака должно быть нормальным; 2) генеральные совокупности, к которым относятся выборки, должны иметь одинаковые дисперсии признака (Дерябин, 2005).
Однако в дерматоглифике, как, впрочем, и в исследованиях, опирающихся на другие системы антропологических признаков, t- критерий нередко используется для оценки значимости различий между расами или группами популяций без специального обоснования. Большая часть анализируемых дерматоглифических признаков имеет дискретную природу и на индивидуальном уровне может кодироваться в двоичной системе (за исключением индексов); их внутригрупповое распределение (по индивидуальным данным) не может иметь вид нормального. Тем не менее, на межпопуляционном уровне в рамках больших рас распределение всех ключевых признаков имеет близкий к нормальному вид (см. Главу 7). В случаях, когда анализу подвергаются группы, состоящие из нескольких десятков выборок, это обстоятельство позволяет с некоторыми оговорками использовать t-критерий для анализа значимости различий между средними величинами признаков.
Все же более корректным в дерматоглифических исследованиях является применение непараметрических критериев. Оно предполагает использование частот или рангов эмпирических данных, распределение которых может соответствовать любому типу, а условие равенства дисперсий сравниваемых групп не соблюдается. В данной работе для анализа значимости различий между двумя независимыми группами популяций использовался U- критерий Манна-Уитни. При анализе внутрирасовых различий между признаками на левой и правой руке применялся критерий для зависимых выборок - T-критерий Уилкоксона.
3.5. Расчеты корреляций признаков дерматоглифики
Взаимные связи признаков дерматоглифики навнутригрупповом (биологическом) уровне оценивались почетырехпольным таблицам с применением %2, на территориальном (в пределах этносов и регионов) - по формуле Бравэ, на межрасовом - по способу рангов Спирмена. Последний, кроме того, использовался для вычисления коэффициентов корреляции между признаками дерматоглифики и других систем антропологических признаков.
77

Глава 4. Система корреляций элементов дерматоглифики. Выделение группы ключевых признаков.
4.1. Методологические основы анализа
В основу данной работы, как и предшествующих трудов Г.Л. Хить, положены принципы, разработанные советской антропологической школой, последовательное применение которых необходимо в расо- и этногенетических исследованиях (Ярхо, 1932; 1947; Дебец, 1948; Алексеев, 1969). В их число входят:
1. предпочтительное использование в анализе признаков, не связанных между собой устойчивой внутригрупповой корреляцией на уровне индивидов;
2. применение также признаков, связанных между собой на индивидуальном (биологическом) уровне сопоставления, только при соблюдении условия, что величина и направление внутри - и межгрупповых корреляций данных признаков не совпадают;
3. анализ признаков, образующих исторически сложившиеся на исследуемой территории комплексы, с учетом таксономической неравноценности первых;
4. использование признаков, дифференцирующая способность которых для разграничения популяций разного таксономического уровня является доказанной, т.е. имеющих расовые градиенты. Анализ признаков, беспорядочно варьирующих на межпопуляционном уровне сопоставления, не может привести к обоснованным историческим выводам.
Знание взаимосвязей между признаками - необходимая основа антропологического анализа, позволяющая приблизиться к пониманию их природы. Весь комплекс дерматоглифических структур у человека в норме был исследован в этом аспекте на разных уровнях - внутригрупповом (внутрипопуляционном), межгрупповом (территориальные комплексы) и на уровне больших рас (Хить, Долинова, 1996).
4.2. Биологические (внутригрупповые) корреляции
Взаимосвязи признаков на уровне индивидов были изучены в 50 дисперсных выборках (28 мужских и 22 женских). Сюда вошли
78

финны Финляндии, несколько групп русских, белорусы, карелы, армяне, баски, коми, марийцы, казанские татары, якуты, калмыки, кеты, корейцы, тибетцы, японцы, кхмеры, индейцы Центральной Америки, австралийцы, папуасы Новой Гвинеи, негры Западной и Южной Африки, а также айны Хоккайдо и урали Индии. Таким образом, исследованы основные расовые группы и часть метисного населения ойкумены.
При вычислении попарных связей по всем признакам были получены более 2 700 коэффициентов корреляции. Для удобства анализа последовательно рассмотрим следующие взаимосвязи: 1) ключевых признаков друг с другом; 2) ключевых признаков с неключевыми; 3) неключевых признаков друг с другом.
4.2.1. Корреляции в группе ключевых признаков
Эта группа признаков была выделена эмпирически на основе многолетнего тестирования расодифференцирующей способности различных дерматоглифических показателей и последовательного применения принципа «бритвы Оккама», что было обусловлено необходимостью анализа огромного материала при отсутствии компьютерной техники (Хить, 1969). Напомним, что в число шести ключевых признаков входят дельтовый индекс (DlJ0), индекс Камминса (Ic), процент низкого осевого трирадиуса ладони (t), истинных узоров на гипотенаре (Hy) и тенаре (Th/I), а также суммарная частота добавочных межпальцевых трирадиусов (ДМТ). Из них оба индекса отличаются сложной структурой (дельтовый индекс отражает количество дельт, обусловленных петлями и завитками, а индекс Камминса усредняет окончания линий A и D). Остальные четыре представляют собой элементарные, альтернативные по способу определения, признаки, фиксирующие процент наличия той или иной особенности в кожном рельефе.
Основная масса признаков не обнаруживает сколько-нибудь стойких взаимосвязей. Единичные корреляции имеют случайный характер, не повторяются у обоих полов и весьма слабые: доля значимых коэффициентов корреляции составляет 10 и 12% от общего числа попарных г для мужских и женских выборок соответственно. Лишь одна пара признаков, а именно одиночный низкий ладонный трирадиус и узорность гипотенара, скоррелированы обратной, хотя и небольшой, связью в популяциях европеоидной расы, а также у айнов, урали и кхмеров (г достигает -
79

0.3 при p<0.01). Отметим, что у австралийцев и папуасов эта связь усилена, оставаясь, впрочем, в пределах умеренных величин (до - 0.6). На долю этой корреляции приходится до 40% указанного общего числа значимых коэффициентов, обнаруженных при анализе всего материала. Таким образом, популяции европеоидной и австралоидной рас, а также отдельные группы с заметной долей этих компонентов, отличаются определенным своеобразием, заслуживающим дальнейшего изучения.
4.2.2. Корреляции между ключевыми и неключевыми признаками
Изучены связи каждого из шести ключевых признаков с семью неключевыми, в число которых входят: узоры на II, III, IV межпальцевых подушечках, а также варианты системы осевых ладонных трирадиусов (t’, t”, t°, сложные трирадиусы суммарно или СТ). Структура этих связей такова: один ключевой признак (Th/I) практически не зависит от неключевых; остальные пять, как правило, стабильно связаны с неключевыми признаками. Так, Ic положительно скоррелирован с узорностью III межпальцевой подушечки (г достигает 0.5) и отрицательно с IV (г приближается к - 0.6). Здесь отражено «поведение» важнейшей составной Ic, а именно, линии D: при ее низком окончании (в поле 7), влекущем понижение индекса, автоматически возрастает число петель на IV подушечке. При более высоком ходе линии D, в поля 10 и 11, соседняя линия С идет обычно в поле 9, образуя петлю на III подушечке. Иначе говоря, связь Ic с III подушечкой опосредована и выявляется благодаря согласованности хода линий D и С, а с IV подушечкой она происходит напрямую, хотя и имеет обратный знак. Напомним, что Ic как интегративный признак отражает прежде всего изменчивость, обусловленную линиями A и D, но при этом линия D ограничивает ход линий В и С, очерчивая их рамку, в то время как линия А наиболее независима. Стоит отметить, что линия D вносит максимальный вклад в величину Ic (г=0.8-0.9), линия А - минимальный (г=0.2-0.6). Остальные (B и С) ладонные линии также положительно скоррелированы с Ic, особенно тесно - линия B (г=0.7-0.9). Линия С наиболее свободна в своем ходе и наиболее вариативна в окончаниях, поскольку ее связь с Ic ослаблена (г=0.4- 0.7). Таким образом, все главные ладонные линии взаимосвязаны, сильнее всего - B и D, слабее всего А и С (Табл. 4.1).
80

Таблица 4.1. Внутригрупповые связи главных ладонных линий для некоторых популяций Восточной Европы (по: Хить, 1983).Table 4.1. Intra-group correlations between main line values for some East European populations (from: Heet, 1983)
ПризнакиГруппа пол 1с-А 1с-В 1с-С Ic-D А-В А-С A-D В-С B-D C-D
в 0.57 0.81 0.45 0.81 0.45 0.29 0.40 0.58 0.89 0.52Русские Холмогор
? 0.50 0.80 0.71 0.81 0.28 0.35 0.32 0.75 0.92 0.66
в 0.63 0.79 0.73 0.79 0.44 0.41 0.42 0.72 0.88 0.62Русские Кирсанова
? 0.57 0.81 0.60 0.81 0.38 0.39 0.40 0.69 0.84 0.56
Коми-зыряне ижемские6 0.64 0.73 0.57 0.78 0.39 0.38 0.42 0.56 0.90 0.55
? 0.60 0.73 0.65 0.77 0.36 0.39 0.40 0.60 0.91 0.62
в 0.55 0.83 0.50 0.85 0.40 0.36 0.43 0.54 0.92 0.54Марийцы луговые
? 0.65 0.73 0.42 0.75 0.31 0.34 0.34 0.43 0.98 0.45
6 0.54 0.85 0.52 0.83 0.40 0.36 0.39 0.53 0.96 0.56Татары Шали
? 0.47 0.80 0.54 0.80 0.30 0.23 0.30 0.58 0.93 0.53
S 0.24 0.83 0.52 0.85 0.16* 0.14* 0.17* 0.56 0.97 0.52Калмыки-торгоуты
? 0.41 0.87 0.62 0.85 0.26 0.34 0.26 0.57 1.00 0.65
Примечание. Все коэффициенты, за исключением отмеченных *, значимы при р<0.05. Объем выборок варьирует от 82 до 103 человек.All coefficients except signed * are significant (p<0.05). Size of samples ranges between 82 and 103 persons.

Таблица 4.2. Пальцевые признаки и их корреляции в выборках из больших рас (по: Хить, Долинова, 1996). Table 4.2. Finger traits values and correlations between them for samples from major races (from: Heet, Dolinova, 1996).
Группа N,пол
признаки Коэффициенты корреляции признаков
А+Т L W Dlio Dl10- А+Т
ь-11о
3 £1О
3
A + T -L A +T-W L -W
100 3 3.6 62.0 34.4 13.08 -0.60 -0.72 0.96 0.00* -0.36 -0.93Баски
108 $ 8.2 64.6 27.2 11.90 -0.73 -0.61 0.94 -0.11* -0.44 -0.86
100 3 3.0 49.0 48.0 14.50 -0.57 -0.90 0.97 0.16* -0.39 -0.97Японцы
100 $ 6.4 53.6 40.0 13.36 -0.73 -0.67 0.94 -0.02* -0.47 -0.84
85 с? 9.8 63.9 26.3 11.65 -0.76 -0.29 0.80 -0.39 -0.35 -0.73Негры педи
92 $ 16.4 60.9 22.7 10.63 -0.87 -0.06* 0.85 -0.54 -0.49 -0.47
100 3 2.2 40.5 57.3 15.51 -0.65 -0.87 0.97 0.27 -0.48 -0.97Австралийцы
100 $ 2.3 43.4 54.3 15.20 -0.54 -0.91 0.92 0.13* -0.35 -0.97
78 3 3.1 55.0 41.9 13.88 -0.52 -0.91 0.98 0.15* -0.39 -0.97Индейцы миштеки
92 $ 3.1 60.3 36.6 13.36 -0.55 -0.87 0.96 0.07* -0.34 -0.96
Примечание. * отмечены незначимые величины коэффициентов корреляции (все остальные значимы на уровнях 0.05 или 0.01).Insignificant values o f correlation coefficient are marked with * (all other values are significant)

Все варианты одиночных трирадиусов, будучи альтернативными признаками, находятся в обратной взаимозависимости. Так, проксимальный трирадиус t достаточно тесно связан с промежуточным t' (в среднем r=-0.7) и с t” (r=-0.2- 0.3). Между проксимальным трирадиусом t и суммой сложных трирадиусов (СТ) имеется обратная и тесная топологическая связь (до -0.75 у папуасов), резко слабеющая в группах, в которых сложные трирадиусы встречаются редко - например, у монголоидов.
Узорность гипотенара стойко и тесно связана с частотой осевых сложных трирадиусов ладони, величина г колеблется в пределах 0.7-0.9, достигая максимума у австралийцев (!). В группах с повышенным содержанием СТ эта корреляция возрастает, свидетельствуя о почти автоматическом появлении окаймляющих (по продольной оси ладони) трирадиусов при наличии узора на Hy.
Добавочные межпальцевые трирадиусы стойко связаны с частотой узоров на IV ладонной подушечке, являясь, наряду с истинной петлей, одной из двух составляющих последнего признака (г колеблется в пределах 0.2-0.4). Иначе говоря, здесь выявляется тенденция к прямой связи между суммарным количеством ДМТ и той - обычно преобладающей - его частью, которая приходится на долю IV подушечки.
Эта тенденция ослабляется наличием на IV подушечке, кроме добавочных трирадиусов, также истинных петель, а иногда и более сложных узоров. Поэтому в группах с малым процентом ДМТ эта корреляция отсутствует, например, у индейцев и монголоидов. Там, где суммарная частота ДМТ повышена (негры, урали), признак теснее связан с узорностью III подушечки, в чем отражается, скорее, наличие на последней собственно ДМТ и, в меньшей степени, истинных петель. В то же время частота ДМТ слабо связана с узорностью II подушечки, так как на ней никаких иных узоров, кроме образованных добавочным трирадиусом, не бывает, а содержание ДМТ, как правило, очень невелико.
Подчеркнем независимость дельтового индекса от неключевых признаков - кроме типов пальцевых узоров, на основании частот которых индекс и рассчитывается. Этот признак - один из двух с высочайшим таксономическим весом - почти полностью определяется числом завитков (r=0.8-1.0) и менее тесно связан сильной обратной корреляцией с петлями (r колеблется в пределах - 0.6 - -0.9). Эти связи были исследованы в пяти мужских и пяти женских выборках из основных расовых групп (Табл. 4.2). У негров
83

последняя из отмеченных корреляций резко ослаблена (у женщин она даже отсутствует), однако увеличена теснота связи между дельтовым индексом и содержанием дуг (r=-0.8 — 0.9), в то время как в остальных группах этот коэффициент корреляции варьирует от -0.5 до -0.7. Этот феномен объясняется большим числом дуг у негров (а дуги выпадают из подсчета DlJ0), при одновременном уменьшении доли петель, в результате чего корреляция между DlJ0 и петлями уменьшается.
4.2.3. Корреляции в группе неключевых признаков
Эти признаки разделяются на две категории. К одной относятся признаки самостоятельного ранга, т.е. не входящие в состав ключевых. Система взаимоотношений здесь такова: количество узоров на III и IV подушечках связано стойкой обратнойи, как правило, значимой корреляцией порядка -0.4-0.5; максимальная величина r обнаружена в мужской группе айнов (0.63). Узоры на II подушечке практически не скоррелированы с узорами на III и IV подушечках: коэффициенты очень малы и незначимы. Отсутствуют также систематические связи узорности II, III и IV подушечек с частотами промежуточного (t') и срединного (t'') осевых трирадиусов ладони, а также с частотой партинга: значимые корреляции единичны и не превышают 0.2.
В системе вариантов одиночных осевых трирадиусов ладони (кроме проксимального, который был рассмотрен в группе ключевых признаков) слабая отрицательная и редко встречающаяся связь обнаружена между промежуточным и срединным. Промежуточный трирадиус связан стойкой, хотя и малой по величине (до -0.2) обратной корреляцией со сложными трирадиусами ладони. Срединный трирадиус не зависит от частоты сложных трирадиусов. Наконец, говоря о партинге, т.е. отсутствии осевых трирадиусов, подчеркнем, что этот признак не коррелирует с их остальными вариантами, включая проксимальный, а также с узорностью II, III и IV ладонных подушечек. Таким образом, в этой группе неключевых признаков обнаружены лишь два (узорность наIII и IV подушечках), которые взаимно связаны обратной и стойкой корреляцией, достигающей -0.6.
Вторая категория состоит из неключевых признаков, входящих, прямо или опосредованно, составной частью в ключевые. Это типы пальцевых узоров и главные ладонные линии A, B, C и D.
84

Дуги на пальцах практически не зависят от петель (см. таблицу 4.2) и лишь в мужских группах негров и австралийцев оба признака умеренно скоррелированы по типу обратной связи (от -0.3 до -0.5). Дуги и завитки связывает умеренная обратная зависимость (от -0.3 до -0.5). Между петлями и завитками обнаружена почти полная обратная связь (как правило, r достигает -0.9, зачастую приближаясь к -1.0). Лишь у негритянок эта корреляция заметно ослаблена (r=- 0.5). Исходя из этих закономерностей, следует заключить, что при основанном на числовой таксономии подходе к анализу материала использование всех трех типов пальцевых узоров нежелательно. Самый достоверный результат будет получен, если включить в анализ лишь один из пары наиболее связанных признаков (завитки либо петли) и дуги. Предпочтительнее брать петли и дуги как наиболее независимые элементы. Также не следует использовать одновременно пальцевые узоры и дельтовый индекс, поскольку частоты всех типов узоров сильно влияют на величину индекса. Иными словами, дельтовый индекс практически исчерпывает информацию о пальцевых узорах.
Окончания главных ладонных линий B и С, B и D взаимосогласованы (см. таблицу 4.1): первая пара линий в основном характеризуется высокой (0.6-0.8), вторая - почти полной прямой связью (0.9-1.0). Линии C и D также зависят друг от друга (0.5-0.6). Линия A наиболее свободна в своем ходе, о чем свидетельствуют пониженные коэффициенты ее корреляции с тремя остальными (в основном, порядка 0.3). Таким образом, линии B, C и D образуют единый морфологический комплекс, в котором все составляющие сильно зависят друг от друга. Следовательно, в многомерном расовом анализе (во избежание дублирования информации) желательно употреблять лишь одну из этих трех линий и линию A как наиболее самостоятельный элемент. Оба требования удачно воплощены в индексе Камминса - сложном признаке, отражающем влияние как максимально связанного с остальными признака (линия D), так и наиболее свободного от корреляций (линия А).
Основные результаты этого раздела сводятся к следующему. Группа ключевых признаков состоит из практически независимых элементов: значимые коэффициенты составляют здесь всего 11% от общего числа попарных корреляций. В то же время каждый из ключевых признаков, кроме узорности на Th/I (абсолютно инертной в плане корреляций), связан с несколькими неключевыми признаками. Последние, в свою очередь, разделяются на две группы.
85

Одни элементы входят составной частью в группу интегративных ключевых признаков (Dli0 и Ic), и как уже сказано, скоррелированы с ними, но также и друг с другом. Другие же связаны с альтернативными ключевыми признаками (т.е. с t, Hy и ДМТ) и, кроме того, зависят друг от друга. Лишь один из неключевых признаков, а именно партинг, отличается полной самостоятельностью.
Заключим раздел схемой, представляющей в обобщенном виде систему корреляций признаков пальцевого и ладонного рельефа (Рис. 4.1). На ней по горизонтали располагаются ключевые признаки (один пальцевый и пять ладонных) в порядке убывания таксономической значимости от дельтового индекса к узорности тенара (см. Главу 8). Различными линиями, соответственно тесноте связей, соединены скоррелированные признаки. Отсутствие линий отражает полное отсутствие значимых связей.
Рисунок 4.1. Обобщенная схема внутригрупповых (на уровне индивидов) корреляций признаков дерматоглифики в человеческих популяциях. Линиями обозначены значимые связи: пунктирной - слабая (r<0.40), штриховой - умеренная (r=0.40-0.59), сплошной - высокая (r=0.60-1.00). Знаки связи не отражены. Квадратиками отмечены ключевые признаки. СТ - сложные трирадиусы ладони (по: Хить, Долинова, 1996, с модификациями).Figure 4.1. General scheme of intra-group (individuum level) correlations among dermatoglyphic traits in human populations. Significant relationships are marked by lines: low correlations - dotted lines (r<0.40), mediate correlations - dashed lines (r=0.40-0.59), high correlations - solid lines (r=0.60- 1.00). Key traits are marked by a squares. СТ - compound axial triradii (modified from: Heet, Dolinova, 1996).
86

Прежде всего следует отметить, что ключевые признаки отличаются взаимной независимостью. Этому правилу не подчиняется лишь узорность гипотенара и проксимальный осевой трирадиус ладони, слабо скоррелированные между собой по типу обратной связи в европеоидных и единичных выборках из других рас (по преимуществу австралоидных или обладающих заметной долей австралоидного компонента). В то же время данная связь отсутствует у монголоидов, негроидов и американских индейцев; иначе говоря, она наблюдается при повышенном содержании узоров на гипотенаре и исчезает при его понижении. Поскольку эта связь не является универсальной и не охватывает все человечество, она не отражена на рисунке.
Во-вторых, выявляются так называемые корреляционные плеяды, или скопления скоррелированных признаков. Таковы группы «дельтовый индекс - типы пальцевых узоров», «индекс Камминса - главные ладонные линии A, B, C, D», «добавочные межпальцевые трирадиусы - II, III, IV межпальцевые подушечки», «индекс Камминса - III, IV подушечки», «проксимальный трирадиус ладони - другие виды одиночных трирадиусов - сложные трирадиусы ладони», «гипотенар - сложные трирадиусы ладони».
В эти группы входят пять из шести ключевых признаков. Эти пять признаков представляют собой как бы надводную часть айсберга, воплощающего весь набор элементов дерматоглифики, и в качестве реперных точек отражают практически весь запас морфологической изменчивости кожного рельефа кисти.
В-третьих, абсолютно независимы, т.е. свободны от каких- либо связей два признака: узорность Th/I и партинг (t°).
4.3.1. Исторические (межгрупповые или территориальные) корреляции признаков
Начиная с этого таксономического уровня, речь идет уже об изменчивости не индивидов, а групп. Иными словами, совершается переход от индивидуальной изменчивости (биологической по существу) к межпопуляционной или межгрупповой, механизмы формирования которой вызваны к жизни по преимуществу причинами социального или исторического порядка. Под воздействием миграций, сопровождаемых смешением, либо, напротив, изоляции коллективов, на определенных территориях образуются комплексы или сочетания признаков, которые и
87

рассматриваются в расовом анализе в первую очередь (внутри групп, состоящих из индивидов, этих сочетаний не должно быть).
Возможности анализа территориальных корреляций ограничиваются рядом условий, в частности, методической сравнимостью данных, статистической представительностью выборок и широтой их географической локализации. В качестве примера выделения комплексов приведем результаты изучения ключевых признаков у населения основных историкогеографических регионов бывшего СССР (Табл. 4.3).
Таблица 4.3. Территориальные межгрупповые корреляции (r) ключевых признаков в пределах основных регионов Северной Евразии (мужские группы) (по: Хить, 1983).Table 4.3. Territorial inter-group correlations (r) between key traits within main regions of Northern Eurasia (from: Heet, 1983).
Парыпризнаков
Восточная Европа (80)
Кавказ(50)
Средняя Азия (51)
Сибирь(61)
Dlio - Ic -0.364** 0.293* -0.050 -0.118D l10 - t 0.107 0.139 -0.092 0.206Dlio - Hy 0.018 -0.186 0.046 -0.135Dlio - ДМТ 0.206 0.038 0.267* 0.024Dlio - Th/I 0.018 0.152 0.056 -0.042Ic - t -0.235* 0.162 -0.293* 0.172Ic - Hy -0.086 -0.035 0.550** 0.222Ic - ДМТ 0.151 0.329* 0.038 0.367**Ic - Th/I 0.127 0.265* -0.165 -0.286*t - Hy -0.256* -0.254 -0.254 -0.071t - ДМТ -0.208* 0.039 -0.013 -0.112t - Th/I 0.253* 0.309* 0.177 -0.056Hy - ДМТ 0.345** -0.160 -0.029 0.205Hy - Th/I -0.098 -0.048 -0.334* 0.127ДМТ - Th/I 0.172 0.332* -0.147 0.279*
В скобках указано число выборок; * - p<0.05; ** - p<0.01 The number of samples is shown in brackets; * - p<0.05; ** - p<0.01
Эти комплексы различны на разных территориях и достаточно стойки, причем, теснота связей между признаками умеренна и редко превышает 0.4. Так, в группах Восточной Европы понижение частоты t (оказывающегося самым «активным» признаком на данной территории) сопровождается увеличением Ic, узорности Hy и содержанием ДМТ; два последних признака изменяются
88

параллельно, а оба индекса (Dl10 и Ic) находятся в обратной взаимосвязи. Все эти корреляции выявляют два основных компонента, участвовавших в процессе расообразования - европеоидный и монголоидный. В то же время на Кавказе наблюдаем иные и своеобразные комплексы.
Повышение Ic сочетается с увеличением Dl10 и ДМТ. Повышение узорности Th/I сопровождается увеличением Ic, t и ДМТ. Основная часть этих связей (кроме пары t - Th/I) свидетельствует о наличии южного и северного компонентов европеоидного происхождения. У современных потомков древних европеоидных аборигенов Средней Азии (узбеки, таджики, туркмены, западнопамирские народы) комплексы отличаются сложностью. Они указывают не только на наличие европеоидного и монголоидного компонентов (обратная связь между Ic и t, согласованная связь Ic и Hy), но и на взаимодействие южноевропеоидного и североевропеоидного компонентов (корреляция Dl10 и ДМТ, обратная связь между Hy и Th/I).
Южный компонент характеризуется некоторой долей экваториальной (индоавстралоидной) примеси (корреляция Dl10 - ДМТ). На территории Сибири комплексы свидетельствуют о взаимодействии северных и южных групп монголоидного происхождения (корреляции между Ic и ДМТ, ДМТ и Th/I) при определенном влиянии европеоидного населения (в пользу чего может истолковываться корреляция между Ic и ДМТ). Таким образом, исторические корреляции фиксируют современные итоги процессов расообразования, которые протекали в разных регионах страны по-разному и привели к характерным сочетаниям признаков в популяциях, позволяющим судить о взаимодействии разных расовых компонентов. В этом заключается их главное отличие от биологических, т.е. внутригрупповых, корреляций: последние в гомогенных группах выявляют общие для современного человечества закономерности, содержательное истолкование которых часто затрудняется слабой изученностью генетической природы признаков кожного рельефа.
4.3.2. Межрасовые корреляции
На уровне больших рас теснота связей между признаками резко возрастает, достигая высшего порога значимости. Так, при анализе 13 европеоидных и 13 монголоидных групп все возможные
89

корреляции ключевых признаков (кроме тех, в которых участвует Th/I) обнаружили сильную поляризацию групп и выявили исторически сложившиеся комплексы, специфичные для каждой расы (Табл. 4.4). Дельтовый индекс находится в обратной связи с индексом Камминса, узорностью гипотенара и частотой ДМТ; кроме того индекс Камминса прямо скоррелирован с обоими последними признаками и обратно - с проксимальным трирадиусом ладони. В свою очередь, t находится в обратной связи с узорностью Hy и ДМТ; оба последних признака положительно связаны друг с другом.
Таблица 4.4. Межрасовые корреляции (по Спирмену) ключевых признаков дерматоглифики при сопоставлении европеоидных и монголоидных выборок (по: Хить, 1983)Table 4.4. Inter-race correlations (after Spearman’s rank correlation coefficients) of key traits between Caucasoid and Mongoloid samples (from: Heet, 1983)
Признаки Мужчины (26 групп) Женщины (26 групп)DI10 — Ic -0.62** -0.51**DI10 — t **
О
0.51**
О 0 — H y -0.58** -0.39*DI10 — ДМТ -0.44* -0.19DI10 — Th/I 0.23 0.27Ic — t -0.60** -0.42**Ic — Hy 0.71** 0.80**Ic — ДМТ 0.76** 0.64**Ic — Th/I 0.08 0.17t — Hy -0.48* -0.34t — ДМТ -0.60** -0.36t — Th/I 0.35 0.13Hy — ДМТ 0.78** 0.71**Hy — Th/I 0.18 0.23ДМТ — Th/I 0.04 0.32
Примечание: * - p<0.05; ** - p<0.01. Каждая раса представлена 13 выборками на пол; мужчины и женщины в каждой группе исследовались параллельно.Note: * - p<0.05; ** - p<0.01. Every one of the races includes 13 samples for each sex; males and females are separately studied in each of the groups.
90

Яркость выделившихся контрастных расовых комплексов такова, что даже при условии, что расовый состав исследованных выборок неизвестен, уже на этой стадии анализа можно сделать вывод о наличии европеоидного и монголоидного компонентов. Примечательно, что из всех ключевых признаков лишь один (Th/I) абсолютно инертен, не участвуя в формировании характерных расовых комбинаций.
Следует отметить, что эти же комплексы выделились в Восточной Европе по данным локальных групп, на что указывалось в предыдущем разделе. Из этого следует вывод о наличии метисного компонента в составе восточно-европейского населения.
Без достаточного количества методически сравнимых данныхо выборках из других больших рас невозможно исследовать проблему межрасовых корреляций в целом. Несомненно, однако, что расовые комплексы, выделенные другими методами анализа (среднерасовые величины признаков и тестирование различий между расами, комбинационные полигоны признаков, результаты факторного анализа и т.д.) реально существуют, зачастую свидетельствуя о еще более резкой дивергенции рас, чем это было обнаружено на примере сравнения европеоидов и монголоидов.
Выводы:
• Исследованием биологических (внутригрупповых, на уровне индивидов) корреляций признаков дерматоглифики, проведенном на большом и разнообразном в расовом отношении материале, подтверждена правомерность выделения группы ключевых признаков. Эти признаки, во-первых, взаимонезависимы и, во- вторых, отражают основную часть изменчивости всего дерматоглифического комплекса, так как связаны с остальными элементами строения кожных узоров кисти.
• Исторически сложившиеся в разных историкогеографических регионах Северной Евразии комплексы различаются между собой, что объясняется участием разных компонентов в расогенезе населения этих территорий.
• На уровне больших рас характерные комплексы признаков достигают максимальной выраженности.
91

Глава 5. Радиальная ориентация пальцевых узоров и ее значение для расовой дифференциации.
Изучение пальцевых узоров традиционно играет ведущую роль в работах по популяционной дерматоглифике. В отечественных исследованиях основное внимание уделяется анализу распределения дельтового индекса — комплексного признака, отражающего среднее число дельт на человека в популяции и косвенно — частоты узоров различных типов. Признак впервые был предложен М.В. Волоцким (Волоцкой, 1937а). На обширном материале его высокая таксономическая ценность была показана И. Швидецкой (Schwidetzky, 1962), Т.Д. Гладкова (1966), Г.Л. Хить (1983), Г.Л. Хить и Н.А. Долиновой (1996). В зарубежных публикациях основное место отводится изучению вариаций частот встречаемости в популяциях отдельных типов узоров. Единой программы исследований не существует. Так, например, сегодня можно встретить работы, где пальцевые узоры группируются по пяти (Nagy, Pap 2005), шести (Cheng et al., 2009) и даже девяти (Nithin et al., 2009) типам. Независимо от набора признаков, как в российских, так и зарубежных исследованиях особое внимание уделяется анализу частот ульнарных и радиальных петель, однако, ориентация других типов узоров, как правило, игнорируется.
Между тем, специалисты, занимающиеся изучением наследования дерматоглифических признаков, давно уделяют внимание как различным типам, так и ориентации пальцевых узоров без учета их типа (см. историю исследований наследственной передачи узоров в: Kucken, 2007; Гусева, 2008). Разработан метод определения ориентации сложных типов узоров, основанный на различиях в гребневом счете и принятый исследователями разных стран (например: Micle et al., 1982; Zhou, Gu, 2004; и др.).
Рассмотрим способы выделения ульнарных, радиальных и симметричных групп среди узоров трех основных типов пальцевых узоров, подробно описанные еще в работе М.В. Волоцкого (1937б) и сейчас применяемые в исследовательской практике.
Дуги (бездельтовые узоры) считаются ульнарно ориентированными, если вогнутая сторона узора открыта в карпально-ульнарном направлении, и ориентированными радиально, если она открыта в карпально-радиальном направлении. Если
92

вогнутая сторона не имеет заметных отклонений в ульнарную или радиальную сторону, такой узор относится к типу симметричных. К радиальным петлям относятся однодельтовые узоры, которые обращены открытой стороной в радиальном направлении. Соответственно к ульнарным узорам относятся петли, обращенные открытой стороной в ульнарном направлении. Наконец, для завитков и сложных типов узоров ориентация определяется по той стороне, которая содержит меньшее число линий между центром и дельтой, то есть на основании различий по гребневому счету.
Кроме того, М.В. Волоцкой выделил в завитках два типа направления вращения узора: радиальный («линии проходят в верхней части узора в направлении от большого пальца к мизинцу») и ульнарный («линии проходят в верхней части узора в направлении от мизинца к большому пальцу»). «Во всех случаях ток линий спиралей прослеживается в направлении от центра к периферии, другими словами в направлении развертывания спирали» (Волоцкой, 1937а, с.153).
Из дерматоглифических сводок следует, что частота ульнарно ориентированных узоров (петель, завитков и дуг) значительно выше, чем радиально направленных узоров (Гусева, 1986). В антропологических исследованиях об этом косвенно свидетельствуют данные по частотам встречаемости ульнарных петель, практически у всех популяций мира многократно превышающей частоту встречаемости радиальных петель (см. например, сводку данных в: Хить, Долинова, 1990). Несмотря на наличие большого количества работ, посвященных изучению гребневого счета, исследования, в которых рассматривается таксономическая ценность ориентации пальцевых узоров для антропологической дифференциации, почти отсутствуют. С. Микле отмечает общий характер распределения разных типов ориентации по пальцам в изученных выборках (Micle et al., 1982, p. 491). И.С. Гусева провела расчет частот носительства, т.е. процента людей с фенотипически выраженным данным признаком в популяции, для симметричных, ульнарных и радиальных узоров в 15 различных группах Евразии (всего исследовано более 5 400 человек). Данные таблицы как будто не дают оснований считать, что частоты носительства имеют направленный географический характер изменчивости. В работе не говорится также о каких-либо различиях между европеоидными и монголоидными популяциями по этим признакам (Гусева 1986, с. 108-109).
93

Целесообразность принятого метода определения типа ориентации в дуговых и петлевых узорах не вызывает сомнений, и все же определение ориентации сложных узоров по гребневому счету не является единственно возможным способом. Подсчет гребешковых линий является трудоемкой работой, не отвечающей принципу единства методического подхода, поскольку для петлевых и дуговых узоров ориентация определяется по единственной или более открытой стороне узора и с гребневым счетом напрямую не связана. Ориентация сложных типов узоров может определяться по особенностям скелета узора, обусловленным расположением дельтообразующих линий. Именно такой способ предлагается к рассмотрению в данном разделе. Узор считался радиально (ульнарно) ориентированным, если дельтообразующие линии радиально (ульнарно) расположенной дельты попадают в область охвата линий ульнарно (радиально) расположенной дельты. Областью охвата дельты считалась вся площадь поля, ограниченная линиями, образующими дельту.
На Рисунке 5.1 изображены варианты определения ориентации для типов узоров с разным количеством дельт. Номера 1 -6 соответствуют завиткам и сложным узорам, номер 7 — радиальной петле, 8 и 9 — радиальным шатровой и простой дугам. Под номером 1 представлен радиально ориентированный завиток, закрученный радиально, под номером 2 — радиально ориентированный завиток без определенного направления вращения, под номером 3 — радиально ориентированный завиток, закрученный ульнарно. Номер4 — это редкий случай трехдельтового узора. Поскольку в области охвата дельты, расположенной с ульнарной стороны, находятся две другие дельты, такой узор считается ориентированным радиально. Под номером 5 представлен завиток, проксимальная линия ульнарной дельты которого охватывает ток проксимальной линии другой дельты, тогда как дистальные линии обеих дельт сливаются между собой. Такие случаи приняты достаточными для того, чтобы считать ориентацию узора радиальной. Под номером 6 изображен завиток, который, если исходить из расстояний между дельтами и центром, следовало бы считать ориентированным радиально. Направление вращение линий в узоре имеет радиальную направленность, однако, ни одна из дельт узора не попадает полностью в область охвата линий другой. Такой случай определен как радиально закрученный завиток, без выраженной ориентации. Под номером 7 представлена радиальная петля, под номерами 8 и 9
94

- радиальные дуги (шатровая и простая соответственно), открытые врадиальном направлении.
6 7 8 9
Рисунок 5.1. Некоторые варианты радиально ориентированных узоров (по: Широбоков, 20096). 1-5, 7-9 - пальцевые узоры с радиальным типом ориентации; 6 - завиток без выраженной ориентации и радиальным направлением вращения. Подробные пояснения даны в тексте.Figure 5.1. Some variants of radial-oriented finger patterns (from: Shirobokov, 2009). 1-5, 7-9 - finger patterns with radial-oriented frame, 6 - whorl with radial direction of rotation, orientation of pattern frame is indistinct. Detailed explanatories in the text.
Все случаи, изображенные на Рисунке 5.1, за исключением варианта номер 6, учитывались при подсчете частоты встречаемости пальцевых радиальных узоров (FRP - finger radial-oriented patterns). Общая частота встречаемости радиальных узоров на ногтевых фалангах пальцев подсчитывалась по простой формуле:
FRP=100 * (Ar+Tr+Lr+Wr)/(A+T+L+W),
где Ar - число радиально ориентированных простых дуг; Tr - число радиальных шатровых дуг; Lr - число радиальных петель (традиционно обозначаемых в работах как R); Wr - число радиально ориентированных завитков (без учета направления вращения спиралей); A, T, L, W - число узоров тех же типов без учета ориентации. Аналогичным образом подсчитывалась частота
95

встречаемости радиальных узоров в популяции для каждого пальца кисти.
В некоторых случаях, при классификации сложных узоров (спиралевидные завитки, двойные петли и др.) можно также указывать тип направления вращения, который не всегда совпадает с ориентацией узора. Частота таких узоров также учитывалась отдельно, в работе она обозначается как Wrs (номера 1, 5 и 6 на Рис.5.1). Случаи, в которых радиальная ориентация узоров сочетается с ульнарным направлением вращения линий узора, обозначается как Wu (номер 3 на Рис. 5.1).
Таким образом, может быть выделен целый комплексов признаков, отражающих ориентацию пальцевых узоров без учета их типов, а также для класса завитковых (сложных) узоров. Частоты данных признаков никогда не являлись предметом специального изучения в этнической антропологии.
Предлагаемый способ определения ориентации узора, как и способ, основанный на гребневом счете, привязан к положению дельт. Однако, в отличие от последнего, он более прост при первичной обработке дерматоглифических серий. В некоторых случаях, когда из-за плохого качества отпечатка гребневой счет не может быть подсчитан для обеих сторон, его ориентация все же может быть легко определена по особенностям расположения дельтообразующих линий. Кроме того, в предлагаемом варианте дельты не имеют самостоятельного значения и рассматриваются как точки, ток линий относительно положения которых значительно различается на разных участках узора. Именно особенности тока линий определяют как тип узора, так и его ориентацию.
Если ни одна из дельт не попадает полностью в зону охвата линий, образующих другую дельту, ориентация такого узора считается невыраженной. Как показывает практика, сложные узоры с невыраженной ориентацией встречаются достаточно часто, в то же время частота симметричных узоров при таком способе расчета близка к нулю. Не являются редкими варианты узоров, в которых дельтообразующие линии раздваиваются на каком-то участке или обрываются (Рис. 5.2). В таких случаях зона охвата линий дельты, более отдаленной от центра, определялась по максимальной площади, а зона охвата дельты, ближе расположенной к центру - по минимальной.
Таким образом, в альтернативных случаях ориентацию узора предлагается определять по тем вариантам, которые минимизируют
96

частоту невыраженных типов. Во избежание ошибок скелет узора рекомендуется очерчивать иглой.
Рисунок 5.2. Область охвата дельтообразующих линий. На развилке или при обрыве линии зона охвата дистальной линии дельты A определяется по более широкой площади. Линии, образующие дельту В, полностью оказываются в зоне охвата первой дельты.Figure 5.2. The scope area of delta-formative lines. In a fork or line break, the scope zone of distal lines of delta A is defined in a wider area. The lines forming delta B appear completely within a scope zone of the first delta.
Как было показано ранее на материалах мужских выборок, пальцевое распределение частот радиально ориентированных узоров, определяемых в соответствии с предложенным способом, имеет высокое таксономическое значение для дифференциации больших рас, даже несмотря на некоторую формальность приемов определения ориентации (Широбоков, 2009б). Однако, дальнейшая практика обнаружила, что определение Wrs и Wu в некоторых случаях может вызывать затруднение, что отражается на расчете их частот. По этой причине оба признака не рассматриваются в данном разделе.
К настоящему времени получены данные по частоте встречаемости радиальных узоров для 30 мужских и 30 женских выборок различного происхождения с территории Евразии и Западной Африки общей численностью 5 480 человек (Табл. 5.1).
Внутригрупповые корреляции признаков.
Если учитывать, что ориентация узора в определенной степени может быть связана с его типом (так, например, редко встречаются дуги с выраженной ульнарной или радиальной ориентацией), при
97

оценке таксономического значения признака актуальной задачей является анализ внутригрупповых связей между частотой радиальных узоров и величиной дельтового индекса. Слабая положительная связь (г=0.26-0.36) между данными признаками действительно была обнаружена, в том числе на материалах выборок, максимально отличающихся друг от друга по дерматоглифическим показателям (Табл. 5.2).
Таблица 5.1. Частота радиальных узоров в 30 популяциях Евразии и Западной Африки.Table 5.1. Frequency of finger radial-oriented patterns in 30 populations from Eurasia and Western Africa.
№Мужчины Женщины
n I II III IV V n I II III IV V1 70 2.1 45.7 8.6 2.9 1.4 68 2.9 38.2 4.4 2.2 0.02 104 3.8 47.1 6.7 3.8 0.5 100 8.5 35.5 10.0 2.0 0.03 100 5.0 44.5 11.5 4.0 1.5 99 6.6 36.9 10.6 6.1 1.04 96 3.7 39.5 7.4 1.6 0.0 83 3.6 32.5 3.6 2.4 0.05 103 2.9 49.0 10.2 2.9 1.9 101 5.4 38.6 5.4 2.5 0.06 99 10.1 41.9 11.1 1.5 0.5 59 5.1 35.6 10.2 0.8 1.77 80 6.3 52.5 10.0 3.1 0.0 71 6.3 35.2 2.8 2.1 0.08 98 4.1 45.4 7.7 4.1 0.5 100 5.0 41.0 12.0 1.5 0.09 104 7.7 40.4 10.1 4.8 1.4 92 2.7 45.1 9.2 2.2 0.0
10 100 9.0 40.5 11.5 6.0 1.5 99 11.6 44.9 7.6 4.0 0.511 93 11.3 45.7 9.7 4.8 0.0 99 5.6 38.4 8.1 2.5 0.012 91 8.2 34.1 14.3 2.7 0.0 97 13.4 39.2 7.2 2.1 0.513 109 14.7 41.7 10.6 5.0 0.5 97 7.2 39.7 9.8 4.1 0.514 112 12.1 49.6 15.6 7.6 1.3 105 10.5 44.3 16.7 12.4 1.915 107 11.2 43.5 12.1 10.3 0.9 104 20.2 39.4 15.4 10.6 1.416 131 12.6 46.2 19.5 9.5 2.3 136 14.3 42.6 15.8 11.8 2.217 59 10.2 38.1 11.9 4.2 0.0 40 10.0 37.5 6.3 11.3 0.018 34 10.4 40.3 16.4 11.9 0.0 39 14.0 34.5 12.8 14.0 1.319 94 9.6 38.8 25.0 12.8 0.5 106 12.7 42.5 22.2 17.9 2.420 32 7.8 42.2 12.5 12.5 1.6 45 5.6 38.9 22.2 16.7 1.1
Положительно связанными оказываются DlJ0 и величина FRP на радиальной стороне кисти (I-III пальцы), тогда как частота радиальных узоров на ульнарной стороне (IV-V пальцы) и DlJ0 не обнаруживают значимых корреляций ни в одной из выборок. В то же время частоты радиальных узоров на радиальной и ульнарной сторонах кисти не связаны между собой. Устойчивая, но низкая
98

положительная связь фиксируется между частотами радиальных узоров на II и III пальцах. Остальные признаки не показывают устойчивых значимых корреляций.
Таблица 5.1. Продолжение.Table 5.1. Continued.
№Мужчины Женщины
n I II III IV V n I II III IV V21 95 11.1 41.6 13.7 10.5 0.0 142 9.2 43.4 10.2 9.2 0.522 177 3.4 53.1 19.2 5.6 0.6 67 13.4 48.5 11.9 1.5 0.723 71 14.8 39.4 15.5 14.1 2.8 184 9.2 39.7 16.3 10.1 1.124 117 9.0 36.8 19.7 10.7 2.1 65 7.7 44.6 16.2 8.5 1.525 115 9.1 35.7 16.5 14.3 3.5 154 6.5 34.1 15.3 14.0 2.626 53 2.8 37.7 12.3 14.2 2.8 106 7.1 31.1 16.5 11.8 2.427 113 17.3 28.3 7.5 2.2 0.0 40 21.3 31.3 11.3 2.5 0.028 127 13.8 28.0 7.1 3.9 1.2 39 20.5 19.2 1.3 3.8 1.329 120 15.0 26.7 7.1 1.7 0.8 55 22.7 31.8 14.5 0.9 0.030 43 17.4 27.9 10.5 2.3 1.2 41 17.1 24.4 3.7 1.2 0.0
I, II, III, IV, V - частота встречаемости радиальных узоров на отдельных пальцах; FRP - общая частота встречаемости радиально ориентированных узоров. Нумерация групп: 1 - финны; 2 - мордва-шокша; 3 - коми-зыряне; 4 - карелы; 5 - вепсы; 6 - латыши; 7 - эстонцы; 8 - русские; 9 - мари луговые; 10 - азербайджанцы; 11 - армяне; 12 - персы; 13 - белуджи; 14 - корейцы; 15 - нанайцы; 16 - дунгане; 17 - ульчи; 18 - орочи; 19 - эвенки; 20 - нганасаны; 21- качинцы; 22 - якуты; 23 - алтай-кижи; 24 - ненцы; 25 - манси; 26 - ханты; 27 - фульбе; 28 - банмана; 29 - манинке; 30 - малийцы (сбор.)I, II, III, IV, V - frequency of radial-orinted patterns on appropriate finger tips; FRP - general frequence of finger radial-oriented patterns. Numeration of groups: 1- Finns; 2 - Mordvinians; 3 - Komi; 4 - Karelians; 5 - Vepsians; 6 - Letts; 7 - Estonians; 8 - Russians; 9 - Mari; 10 - Azerbaijanians; 11 - Armenians; 12 - Persians; 13 - Baluchi; 14 - Koreans; 15 - Nanay; 16 - Dungani; 17 - Ulchi; 18 - Orochi; 19 - Evenks; 20 - Nganasany; 21 - Kachin; 22 - Yakuts; 23 - Altay Kizhih; 24 - Nenets; 25 - Mansi; 26 - Khanty; 27 - Fulbe; 28 - Banmana; 29 - Maninke; 30- Malians (random sample).
Таким образом, рассмотренный ряд признаков, выражающих радиальную ориентацию пальцевых узоров, отличается относительной независимостью. В выборках обнаружена лишь слабая положительная корреляция между частотой радиальных узоров и дельтовым индексом.
Анализ корреляций частоты радиальных узоров и пяти ключевых признаков в серии карел Медвежьегорского района
99

Карелии показал наличие значимых связей между частотой радиальных узоров и узорностью Th/I на правой руке, а также с узорностью Hy на левой руке (Табл. 5.3).
Таблица 5.2. Внутригрупповые коэффициенты корреляции между дельтовым индексом и некоторыми пальцевыми признаками без учета бимануальной асимметрии (результаты анализа четырехпольной таблицы; мужчины).Table 5.2. Intra-group correlation coefficients between PII (Dl10) and some finger traits without accounting of bimanual asymmetry (results of the analysis of fourfold table; males)
Сопоставляемые Русские Нанайцы Фульбе
признаки (98 чел.) (107 чел.) (113 чел.)Dl10 - FRP (I-III) 0.379* 0.321* 0.290*
DI10 - FRP (IV-V) 0.125 0.157 0.153
DI10 - FRP (I-V) 0.361* 0.256* 0.290*
FRP (I-III) - FRP (IV-V) -0.074 0.093 0.183
Сопоставляемые Азербайджанцы Корейцы Манинка
признаки (100 чел.) (112 чел.) (120 чел.)FRP (I) - FRP (II) 0.129 0.139 0.046
FRP (I) - FRP (III) 0.079 0.118 0.179
FRP (I) - FRP (IV-V) 0.124 0.055 -0.040
FRP (II) - FRP (III) 0.294* 0.267* 0.201*
FRP (II) - FRP (IV-V) -0.223* 0.110 0.043
FRP (III) - FRP (IV-V) 0.048 0.283* 0.289*
Dl10 - дельтовый индекс; FRP (I, II, III, IV-V) - частота радиальных узоров на соответствующих пальцах рук; * - P<0.05.Dl10 - pattern intensity index; FRP (I, II, III, IV-V) - frequency of radial- orinted patterns on appropriate finger tips; * - P<0.05.
В обоих указанных случаях корреляции слабые и, по всей вероятности, имеют случайный характер. При анализе связей без учета бимануального распределения величин признаков значимые корреляции между частотой радиальных узоров и ключевыми признаками у карел не были выявлены. В целом на внутригрупповом уровне тип ориентации пальцевых узоров не
100

зависит от основных ладонных признаков, а связь радиальности с пальцевыми узорами характеризуется невысокими коэффициентами.
Таблица 5.3. Внутригрупповые корреляции частоты радиальных узоров и основных признаков в группе сегозерских карел (96 мужчин).Table 5.3 Intra-group correlation coefficients between FRP and key traits in the Segozero Karelian group (96 males).
FRP DI10 Ic t Hy ДМТ Th/I
Левая рука 0.036 0.019 0.102 -0.232* 0.102 0.056
Правая рука 0.116 -0.139 0.121 -0.080 -0.123 0.206*
Обе руки -0.060 -0.081 0.162 -0.153 -0.099 0.159
* - P<0.05
Межгрупповой анализ
Для анализа дифференцирующего значения пальцевого распределения частот радиальных узоров применялся метод главных компонент. Статистической обработке были подвергнуты данные 30 указанных выше мужских выборок по 4 признакам: средней частоте встречаемости радиально ориентированных узоров на I, II, III, а также суммарной частоте радиальных узоров на ульнарной стороне (IV-V). Данные по частотам узоров на IV и V пальцах были объединены, поскольку радиальные узоры встречаются на них крайне редко и в значительной части исследованных групп частота FRP (V) равна 0, что не позволяет провести корректный статистический анализ.
В результате были выделены две значимые главные компоненты, суммарно отражающие более 81% общей изменчивости. Первую компоненту (охватывающей около 42% общей вариабельности признаков) определяют частоты радиально ориентированных узоров на III и IV-V пальцах. Согласно нагрузкам признаков по первой компоненте, частота встречаемости радиальных узоров на указанных пальцах имеет общий вектор изменчивости. Во второй компоненте, охватывающей 39% дисперсии, наибольшее дифференцирующее значение имеет частота встречаемости радиально ориентированных узоров на I и II пальцах. При повышении в группах частоты радиальных узоров на I пальце их частота на II пальце уменьшается.
101

Расположение групп в пространстве первых двух ГК показано на Рисунке 5.3.
Выборки занимают на графике области, в целом соответствующие их относительной географической локализации на уровне частей света. Первая компонента дифференцирует группы по оси запад - восток, а вторая отражает различия между популяциями по оси юг - север.
Рисунок 5.3. Положение 30 мужских выборок в пространстве I и II главных компонент. Нумерация групп: 1 - финны; 2 - мордва-шокша; 3- коми-зыряне; 4 - карелы; 5 - вепсы; 6 - латыши; 7 - эстонцы; 8 - русские; 9 - мари луговые; 10 - азербайджанцы; 11 - армяне; 12 - персы; 13 - белуджи; 14 - корейцы; 15 - нанайцы; 16 - дунгане; 17 - ульчи; 18 - орочи; 19 - эвенки; 20 - нганасаны; 21 - качинцы; 22 - якуты; 23 - алтай- кижи; 24 - ненцы; 25 - манси; 26 - ханты; 27 - фульбе; 28 - банмана; 29 - манинке; 30 - малийцы (сбор.)Figure 5.3. The position of 30 male samples on the first 2 principal components. Numeration of groups: 1 - Finns; 2 - Mordvinians; 3 - Komi; 4 - Karelians; 5 - Vepsians; 6 - Letts; 7 - Estonians; 8 - Russians; 9 - Mari; 10 - Azerbaijanians; 11 - Armenians; 12 - Persians; 13 - Baluchi; 14 - Koreans; 15- Nanay; 16 - Dungani; 17 - Ulchi; 18 - Orochi; 19 - Evenks; 20 - Nganasany; 21 - Kachin; 22 - Yakuts; 23 - Altay Kizhih; 24 - Nenets; 25 - Mansi; 26 - Khanty; 27 - Fulbe; 28 - Banmana; 29 - Maninke; 30 - Malians (random sample).
102

Выборки, принадлежащие к разным расовым группам, образуют на графике обособленные скопления, трансгрессия между которыми практически отсутствует. Исключение составляет малочисленная группа ульчей (№17), оказавшаяся в поле значений, характерных для европейских групп. Вместе с тем, положение групп не показывает строгого соответствия их географической локализации и не соответствует относительной близости, известной по данным других систем антропологических признаков. Лишь расположение групп европеоидного кластера в значительной мере согласуется с их изменчивостью в меридиональном направлении. Монголоидные группы, для которых характерны наиболее высокие частоты радиально ориентированных узоров на ульнарной стороне кисти, по первой компоненте достаточно четко различаются от негроидных и европеоидных популяций, которым свойственны средние или низкие частоты радиальных узоров.
Вторая компонента определяет специфичность европеоидных групп, для которых характерна низкая частота радиальных узоров наI пальце.
Итоги анализа женских выборок в значительной степени сходны с этими результатами (Рис. 5.4).
У женщин первые две главные компоненты суммарно охватывают около 80% общей изменчивости признаков. Расположение серий в пространстве первой компоненты определяют частоты FRP на III и IV-V пальцах. Группы разделяются в соответствии с их дифференциацией на уровне больших рас. В отличие от мужских выборок, отчетливое сближение с европейскими группами демонстрируют не только ульчи, но и качинцы с якутами. Впрочем, выраженная трансгрессия между монголоидными и европеоидными группами отсутствует.
Как для мужских, так и для женских выборок результаты кластерного анализа по первым двум главным компонентам свидетельствуют о том, что по признакам, отражающим пальцевое распределение радиальных узоров, европеоидные и монголоидные популяции Евразии более сходны между собой, чем любая из них с африканскими группами. Последние занимают относительно обособленное положение и характеризуются наиболее специфическим комплексом признаков (Рис. 5.5).
103

Рисунок 5.4. Положение 30 женских выборок в пространстве I и II главных компонент. Нумерация групп: 1 - финны; 2 - мордва-шокша; 3- коми-зыряне; 4 - карелы; 5 - вепсы; 6 - латыши; 7 - эстонцы; 8 - русские; 9 - мари луговые; 10 - азербайджанцы; 11 - армяне; 12 - персы; 13 - белуджи; 14 - корейцы; 15 - нанайцы; 16 - дунгане; 17 - ульчи; 18 - орочи; 19 - эвенки; 20 - нганасаны; 21 - качинцы; 22 - якуты; 23 - алтай- кижи; 24 - ненцы; 25 - манси; 26 - ханты; 27 - фульбе; 28 - банмана; 29 - манинке; 30 - малийцы (сбор.)Figure 5.4. The position of 30 female samples on the first 2 principal components. Numeration of groups: 1 - Finns; 2 - Mordvinians; 3 - Komi; 4 - Karelians; 5 - Vepsians; 6 - Letts; 7 - Estonians; 8 - Russians; 9 - Mari; 10 - Azerbaijanians; 11 - Armenians; 12 - Persians; 13 - Baluchi; 14 - Koreans; 15- Nanay; 16 - Dungani; 17 - Ulchi; 18 - Orochi; 19 - Evenks; 20 - Nganasany; 21 - Kachin; 22 - Yakuts; 23 - Altay Kizhih; 24 - Nenets; 25 - Mansi; 26 - Khanty; 27 - Fulbe; 28 - Banmana; 29 - Maninke; 30 - Malians (random sample).
Радиальные узоры во всех случаях преобладают на втором пальце. Однако формулы встречаемости радиальных узоров в кластерах различаются. В монголоидных и европеоидных популяциях Евразии преобладающей является формула II>III>I>IV>V, тогда как для населения Западной Африки характерен специфический вариант распределения - с выраженным смещением частот на радиальную сторону кисти: II>I>III>IV>V.
104

Рисунок 5.5. График распределения суммарных частот радиально ориентированных узоров по гомологичным пальцам обеих рук в популяциях, относящихся к трем основным расовым стволам. Е - европеоиды; М - монголоиды; Н - негроиды. I-V - частоты радиальных узоров на гомологичных пальцах рук.Figure 5.5. A graph showing the distribution of summary frequencies of radial-oriented finger patterns in homologous fingers of both hands in different populations: E - Caucasoids; M - Mongoloids; H - African aboriginals. I-V - frequencies of radial-oriented patterns in homologous fingers.
Обе обобщенные выборки европеоидов и монголоидов (как мужчины, так и женщины) отличаются от негроидов в первую очередь более высокими частотами радиальных узоров на II пальце и более низкими частотами радиальных типов на I пальце. Монголоидные группы отличаются от негроидов по частотам радиальных узоров на всех пальцах. Европеоидная и монголоидная выборки демонстрируют значимые различия по частотам радиальных узоров на III и IV пальцах, более низким в европеоидных группах.
Ранее в дерматоглифике высказывалась мысль о том, что негроиды и европеоиды являются генетически наиболее близкими между собой популяциями, выделившимися на относительно поздних стадиях дифференциации человечества, которым предшествовало обособление монголоидного, а до того - австралоидного комплекса признаков (Thoma, 1974). Однако данные по ключевым признакам не позволяют объединять негроидов и европеоидов в единый западный ствол и свидетельствуют об обособленном положении негроидов. Результаты анализа в большей
105

степени соответствуют предположению о том, что негроиды являются наиболее специализированной расой человечества (Хить, Кейта, 1981). Современные данные генетики не оставляют никаких сомнений в том, что население Африки в максимальной степени различается с популяциями остальных частей Земного шара (Zhivotovsky, 2003). В дерматоглифике это предположение было основано на значительных различиях по основным признакам, отражающим особенности как пальцевого, так и ладонного кожного рельефа при сравнении африканских популяций с европеоидными и монголоидными группами Евразии. Введение данных по серии новых дерматоглифических признаков подтверждает выводы о крайней специфичности негроидного комплекса. (Отметим, что этот вывод пока доказан только для популяций Западной Африки. Данные по южным группам негроидов отсутствуют, но, по всей вероятности, специфичность дерматоглифического комплекса населения, проживающего к югу от Сахары, должна только возрастать).
Межгрупповые корреляции.
На межгрупповом уровне значимые корреляции между отдельными пальцевыми признаками выявлены как в монголоидных, так и европеоидных выборках Евразии (Табл. 5.4). Для африканских популяций корреляции не рассчитывались ввиду небольшого числа выборок.
Как в монголоидных, так и в европеоидных группах одинаково тесно и положительно связаны средняя частота радиальных узоров (FRP) и частота радиальных узоров на II пальце (r=0.77). Эта связь имеет исключительно математическое объяснение, так как радиальные узоры в подавляющем большинстве случаев фиксируются именно на II пальце.
В монголоидных группах дельтовый индекс связан значимой положительной корреляцией с частотой радиальных узоров на IV пальце (r=0.64). Как было показано выше, на внутригрупповом уровне FRP на ульнарной стороне кисти не зависит от величины дельтового индекса. Следовательно, данная корреляция может объясняться исключительно вариациями исторически сложившихся комплексов в монголоидных популяциях.
106

Таблица 5.4. Межгрупповые коэффициенты корреляции между некоторыми пальцевыми признаками в монголоидных (справа от диагонали) и европеоидных (слева от диагонали) выборках Евразии.Table 5.4. Inter-group correlation coefficients among some finger traits in Mongoloids (to the right of diagonal) and Caucasoids (to the left of diagonal).
Dl10 FRP (I-V) I II III IV V
Dl10 - -0.066 -0.054 -0.258 0.428 0.635* 0.352FRP (I-V) -0.280 - -0.071 0.772** 0.501 -0.397 0.282
I 0.894** 0.097 - -0.3810.167
0.231 0.188
II -0.554* 0.767** -0.344 - 0.141 -0.402 0.429III 0.471 -0.226 0.479 -0.469 - 0.234 0.118IV 0.572* 0.448 0.407 0.012 0.088 - 0.145V -0.169 0.333 -0.313 0.14 0.092 0.293 -
* - P<0.05; ** - P<0.01
В европеоидных выборках связь между дельтовым индексом и частотой радиальных узоров на отдельных пальцах теснее, чем в монголоидных. При этом коэффициенты корреляции дельтового индекса с радиальными узорами на I и IV пальцах имеют положительное значение (r=0.89 и 0.57 соответственно), а связь индекса с частотой признака на II пальце отрицательное (r=-0.55). Вероятно, последнее обстоятельство вызвано понижением частоты радиальных узоров на II пальце в группах южных европеоидов, для которых в этом отношении характерна тенденция к сближению с негроидными выборками. В целом корреляции между пальцевыми признаками у европеоидов усилены и более многообразны, чем в монголоидных группах.
Средняя частота встречаемости и средняя частота носительства пальцевых узоров c радиальной ориентацией
Средние частоты встречаемости FRP в монголоидной, европеоидной и негроидной выборках значимо различаются между собой (Рис. 5.6). Исключение составляют женские выборки европеоидов и негроидов, при этом различия между каждой из них и монголоидной выборкой сохраняются на значимом уровне.
107

% d9
~т “Г
“Г□= L-j-jX X
^ У
Е М Н Е М Н
Рисунок 5.6. Средняя частота встречаемости и вариабельность значений FRP в европеоидной (Е), монголоидной (М) и негроидной (Н) обобщенных выборках.Figure 5.6. Average frequencies and variability of FRP values in Caucasoid (Е), Mongoloid (М) and Negroid (Н) combined samples.
Картина межрасовой дифференциации, основанная на средневыборочных частотах носительства радиальных узоров, носит менее выраженный характер. Напомним, что частота встречаемости признака отражает его процентное содержание в среднем на одного индивида в популяции (т.е. в среднем на десять пальцев), а частота носительства показывает процент людей, обладающих данным признаком (в независимости от того, зафиксирован признак на одном пальце или десяти), к общему числу людей в изучаемой выборке.
По частоте носительства радиальных узоров значимые различия между монголоидной и европеоидной выборками отсутствуют, но тенденция к повышенной частоте признака в монголоидных популяциях сохраняется.
Евразийские выборки (европеоиды и монголоиды) отличает более высокий процент людей с радиально ориентированными пальцевыми узорами относительно западноафриканских популяций, специфичность которых сохраняется при обоих способах расчета (Рис. 5.7). В отличие от мужчин, у женщин различия между частотами носительства в европеоидных и монголоидных группах значимы, тогда как между европеоидными и негроидными группами значимые различия не обнаруживаются. Отсутствие различий связано в первую очередь с крайне высокой вариабельностью
108

признака у женщин Западной Африки - как по частоте носительства, так и по частоте встречаемости радиальных узоров.
80 ■
70 ■
60
50 -
40 ■
% g ?- г Л х R тУ У □ L
X "
_ L
Е М Н Е М Н
Рисунок 5.7. Средние частоты носительства и вариабельность значений FRP в европеоидной (E), монголоидной (М) и негроидной (Н) обобщенных выборках.Figure 5.7. Average frequencies of persons and variability of FRP values in Caucasoid (Е), Mongoloid (М) and Negroid (Н) combined samples.
В целом частоты встречаемости признака позволяют более строго дифференцировать популяции, относящиеся к трем различным расам, нежели частоты носительства. Это утверждение относится как к мужским, так и женским группам. Несмотря на то, что при обоих способах расчета у женщин были выявлены значимые различия между монголоидной и европеоидной выборками, именно по частоте встречаемости радиальных узоров отличия носят наиболее выраженный характер (p<0.00005 для частот встречаемости, p<0.005 для частот носительства).
Следовательно, на межпопуляционном уровне для территории Евразии можно говорить о различиях, выраженных в первую очередь в степени фенотипической выраженности радиальной ориентации узоров в расчете на одного носителя признака. В то же время общий процент индивидуумов, обладающих данным признаком, по отношению к общему числу индивидуумов в популяции имеет примерно одинаковую величину как в монголоидных, так и европеоидных группах.
Различия дерматоглифических показателей населения Евразии и Западной Африки носят более выраженный характер и связаны как
109

с разными типами пальцевого распределения, характеризующими население двух континентов, так и относительным числом людей, являющихся носителями признака. Эти обстоятельства, безусловно, связаны с резко выделяющимися частотами генов, определяющих особенности формирования кожного рельефа кисти взападноафриканских популяциях на фоне евразийских величин.
Предлагаемые новые признаки дерматоглифики, отражающие радиальный тип ориентации узоров и его пальцевое распределение, обладают высокой таксономической ценностью для дифференциации популяций Старого Света. В западноафриканских группах выявлен наиболее специфический комплекс признаков, характеризующийся абсолютным преобладанием радиальных узоров на пальцах радиальной стороны кисти при общей низкой частоте носительства радиального типа узоров. Для монголоидныхпопуляций, напротив, характерны максимальные частоты встречаемости радиальных узоров и значительный процент радиальных узоров на пальцах ульнарной стороны кисти. Европеоидные группы по частотам встречаемости и частотамносительства радиальных узоров занимают промежуточноеположение между негроидными и монголоидными популяциями, а по типу пальцевого распределения радиальности в значительной степени сближаются с последними.
Средняя частота встречаемости радиально ориентированных узоров (FRP) и формула пальцевого распределения данного признака имеют высокое таксономическое значение для межрасовой дифференциации. По этой причине средняя частота радиальных узоров заслуживает включения в число основных диагностических признаков. На внутригрупповом уровне она имеет слабую положительную связь с величиной дельтового индекса, что не может оказать существенного влияния на результаты межпопуляционных сопоставлений. Этот вывод базируется пока на данных выборок из трех больших рас - европеоидной, монголоидной и негроидной.
110

%

Г лава 6. Половые различия признаков дерматоглифики
При изучении различий между признаками в группах мужчин и женщин как отечественными, так и зарубежными исследователями часто используется термин «половой диморфизм». Однако, в действительности, даже самые сложные дактилоскопические схемы позволяют разграничивать отпечатки рук мужчин и женщин лишь на уровне вероятностной статистики (Божченко, 2004; Gutierrez- Redomero et al., 2013). В дерматоглифике не выявлены фенотипические элементы ни по одному из признаков, которые могли бы быть связаны с половой принадлежностью индивидов. Поэтому, безусловно, правильнее говорить о направленных, статистически значимых половых различиях признаков дерматоглифики, и следует оговориться, что в данной работе термин «половой диморфизм» используется именно в этом расширенном толковании.
Среди многочисленных работ, затрагивающих проблему полового диморфизма по признакам дерматоглифики и отличающихся большим разнообразием подходов и методов, к сожалению, практически нет обобщающих исследований. Степень изученности разных признаков несравнима, выводы разных авторов часто противоречат друг другу. Некоторые стороны проблемы полового диморфизма вообще не рассматривались. Тем не менее, можно выделить общее направление половых различий у разных рас и популяций. Так, степень сложности пальцевого рельефа и соответствующие индексы у мужчин, как правило, выше, чем у женщин, так же, как индекс Камминса. Частота узоров на II межпальцевой подушечке также выше у мужчин, а на IV подушечке, так же, как и на гипотенаре напротив, выше у женщин. В отношении остальных признаков, насколько можно судить по отрывочным данным, четкие закономерности общего характера не выявлены.
Ниже излагаются основные результаты специального исследования полового диморфизма дерматоглифических признаков у человека (Хить, Долинова, 1997). Систематическое изучение сравнительной дифференциации полов на разных таксономических уровнях (человечество в целом, большие расы и их подразделения, различные уровни в пределах европеоидной и монголоидной рас -
112

селенной,3 территориальный, этнический) было выполнено с соблюдением главного условия - с единых методических позиций и с помощью современных приемов обработки анализа данных. Использовались лишь материалы параллельно исследованных мужских и женских выборок, что обеспечивает полную достоверность заключений.
Половая изменчивость была изучена во всех основных расах человечества. Главным источником материала послужила монографическая сводка мировых данных (Хить, Долинова, 1990), а также отдельные публикации. К сожалению, в большинстве работ программа исследования не охватывает все признаки. Поэтому материалы для анализа были отобраны таким образом, чтобы, по крайней мере, в отношении основных признаков, была соблюдена принадлежность цифровых данных одним и тем же выборкам.
Поскольку в большинстве публикаций описывается пальцевый рельеф, эта группа признаков в таблицах всюду представлена максимальным числом выборок. Разрыв между наибольшим и наименьшим числом выборок, приходящимся на рассматриваемые признаки в пределах расы, временами исключительно велик. Поэтому искусственным образом количество групп было сокращено (в основном по пальцевым узорам и Dl10) с тем, чтобы сгладить этот разрыв.
В общей сложности изучено 716 мужских и 716 женских параллельно обследованных выборок. Большинство из них относится к европеоидной и монголоидной расам (370 и 230 групп соответственно), остальные расы представлены гораздо меньшим числом выборок: негроиды (континентальная Африка к югу от Сахары) - 52, австралоиды (аборигены Австралии и Океании) - 37, американоиды (индейцы обоих континентов) - 27 выборками. Приведенные цифры максимальны и относятся к величине дельтового индекса.
Во внутрирасовом масштабе в качестве основной модели для изучения половых различий послужили территориальные (локальные, племенные выборки). Причиной послужило прежде всего обилие соответствующего материала, относящегося к европеоидам и монголоидам. Это позволило установить основные закономерности полового диморфизма и сравнить данный таксономический уровень с остальными внутрирасовыми уровнями
3 Здесь и далее: слово «селенной» используется для краткого обозначения уровня отдельных сельских поселений.
113

(селенным и этническим). Выяснилось, что территориальный уровень отражает в суммарном виде закономерности, характерные для всех уровней внутри этих двух рас, и в этом смысле представляют собой усредненную внутрирасовую модель. Таким образом, сравнение всех больших рас в аспекте полового диморфизма на основе материала территориальных групп населения методически представляется вполне корректным. Важную роль сыграл тот факт, что данный таксономический уровень чаще всего представлен в зарубежных публикациях, описывающих население Южной Азии, Африки, Америки.
В пределах европеоидной и монголоидной рас материал исследовался по указанным уровням и в регионах - в Европе (без учета групп смешанного, пришлого или спорного происхождения, т.е. башкир, удмуртов, марийцев, чувашей, астраханских татар, калмыков, ногайцев, ненцев, саамов), на Кавказе, в Средней Азии (использовались данные о таджиках, туркменах, узбеках, народах Западного Памира) и в Передней Азии. Монголоидные популяции рассматривались по отдельности в пределах Северной и ЮгоВосточной Азии.
Основное внимание уделялось шести ключевым признакам кожного рельефа кисти и вариантам отсутствия и редукции линии С. В первоисточнике (Хить, Долинова, 1997) проанализированы также другие неключевые признаки: дуги на пальцах (A+T), пальцевые петли (R+U) и завитки (W), средний балл окончания главных ладонных линий А и D, варианты осевых ладонных трирадиусов - промежуточный (t'), срединный (t''), партинг (t°), а также узорность ладонных подушечек (II, III, IV)
Для выявления основных закономерностей полового диморфизма внутри расы, как упоминалось, были использованы выборочные данные разных таксономических уровней на примере европеоидной и монголоидной рас. Сравнительный анализ всех рас проводился с использованием данных территориально-племенного уровня. В обоих случаях анализа строились вариационные ряды по выборочным средним и вычислялись основные параметры распределения (x, s, v). При числе выборок меньше 20 применялся способ малого числа наблюдений. Корреляции между параметрами вычислялись по Бравэ (r) и Спирмену (rs).
Для выявления различий между мужскими и женскими выборками использовались два показателя: абсолютный иотносительный. Абсолютная половая разница (АПР) вычислялась в
114

единицах измерения без учета знака (т.е. без учета направления различий между полами). Относительная разница, или коэффициент полового диморфизма (КПД), выражалась как частное от деления величины признака в мужской группе на величину признака в женской группе. Значимость различий тестировалась при помощи t- критерия Стьюдента. Для определения гомогенности человечества в целом по данным обобщенных расовых групп применялся ранговодисперсионный анализ по Фридману.
6.1. Корреляция между полами по величине признаков дерматоглифики.
Уровень больших расСреднерасовые величины признаков были подсчитаны по
вариационным рядам выборочных средних для каждого пола в отдельности со взвешиванием на число выборок. Обнаружено, что по всем ключевым признакам, а также по варианту отсутствия линии С (Со) между полами существует тесная высокозначимая корреляция (rs колеблется от 0.99 до 1.00, что соответствует вероятностным уровням от 0.05 до 0.01). Наименьшая степень соответствия полов найдена в отношении узорности Th/I, а полное отсутствие связи - по частоте партинга.
Внутрирасовый уровеньУ европеоидов разных регионов наибольший параллелизм в
значениях признаков у мужчин и женщин отмечен по обоим индексам и ДМТ, причем, связь умеренная (в основном r колеблется от 0.3 до 0.9) и обычно значимая. Другая половина ключевых и часть неключевых признаков у мужчин и женщин варьируют независимо. Для монголоидов Сибири также характерна умеренная и значимая связь величин признаков у обоих полов, увеличенная по специфическому «сибирскому» признаку (t). Негроиды отличаются полным отсутствием параллелизма в значениях индекса Камминса у полов, в то время как остальные признаки обнаруживают умеренную или повышенную корреляцию. У индейцев Америки величины обоих индексов у мужчин и женщин скоррелированы почти полностью, в то время как по остальным ключевым признакам корреляция отсутствует либо незначима. У австралоидов все ключевые признаки варьируют у обоих полов параллельно: степень связи в основном достаточно высока и значима. Неключевые
115

признаки, входящие составной частью в ключевые, обнаруживают те же закономерности.
Основные выводы:
• В пределах человечества как целого на уровне его составных частей (больших рас) существует высокая и значимая связь величин одноименных признаков у мужчин и женщин.• Внутри расы на уровне выборок наблюдается разная степень соответствия между полами по величинам признаков: коэффициент корреляции колеблется от 0.02 до 0.98. Ключевые признаки и их составляющие, как правило, скоррелированы у обоих полов более тесно по сравнению с другими признаками.• Корреляция между полами по большинству признаков максимально низка у европеоидов, наиболее велика у австралоидов и индейцев. Монголоиды приближаются к двум последним расам.
6.2. Закономерности внутрирасового распределения частот признаков дерматоглифики в мужских и женских группах
При анализе использовались данные о территориальных группах в пределах каждой расы. Распределение частот ключевых признаков при достаточном числе наблюдений имеет в целом нормальный или близкий к нему вид (Рис. 6.1).
По всем признакам без исключения наблюдается сильная трансгрессия кривых, иногда распределение почти полностью совпадает. Однако существуют при этом явные различия между полами, неодинаково выраженные у разных рас. Так, по дельтовому индексу максимальное расхождение мужской и женской кривых характеризует европеоидов и негроидов, в меньшей степени - монголоидо. В то же время у австралоидов и американоидов направленных различий между полами практически нет. По индексу Камминса наиболее дифференцированными по полу расами являются монголоиды и американоиды, тогда как в пределах негроидной и европеоидной рас оба пола различаются слабее.
Все расы, о которых имеются сопоставимые данные, обнаруживают явные различия полов в отношении проксимального трирадиуса ладони; у европеоидов этот эффект более сглажен, хотя модальный класс распределения у полов не совпадает. Характерные различия между полами, связанные с более высокой частотой Hy у
116

женщин, отмечены у европеоидов, монголоидов и американоидов, между тем как у негроидов и австралоидов эти различия невелики и неопределенны.
Распределение частот ДМТ хорошо дифференцирует мужские и женские группы всех рас, кроме американоидной (данных о которых мало, причем в каждой выборке признак чаще встречается у мужчин).
Рисунок 6.1. Распределение частот признаков у мужчин и женщин (территориальный уровень). По горизонтали - величина признака, по вертикали - частота (%). Сплошная линия - мужчины, штриховая - женщины. Обозначение рас: Е - европеоиды, М - монголоиды, Н - негроиды, Ам - американоиды, Ав - австралоиды.Figure 6.1. Distribution of traits frequencies in males and females (territorial level). Horizontal axis - trait value, vertical axis - frequencies (%). Data for males are marked by Solid line - males, dotted line - femails. Designations of races: E - Caucasoids; М - Mongoloids; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
117

Что касается узорности Th/I, то мужские кривые вполне отчетливо, хотя и менее явно в сравнении с ДМТ, различаются с женскими. У мужчин узорность Th/I несколько выше, за исключением негроидов и австралоидов, у которых различия между полами неопределенны. По варианту отсутствия линии С половые различия значимы у европеоидов, в то время как у остальных рас разнонаправленная дифференциация полов отсутствует (так же, как и по варианту Cx у европеоидов и монголоидов). В отношении остальных признаков наблюдаются в основном явные различия между мужчинами и женщинами, за редкими исключениями.
О 16 32 48Th/I
Рисунок 6.1. Продолжение.Figure 6.1. Continued.
118

Модальные классы распределения частот обычно не совпадают, как и протяженность вариационного ряда. О направлении различий разнополых групп в выраженности признаков подробно будет сказано далее; здесь лишь отметим, что не существует общего правила для всех рас по остальным признакам, наиболее отчетливо дифференцирующим мужские и женские группы.
6.3. Сравнительная вариабельность мужских и женских выборок
Как было показано выше, внутрирасовое распределение популяционных средних ключевых признаков у мужчин и женщин характеризуется сходным и близким к нормальному типом при сильной трансгрессии кривых. Вместе с тем, обнаруживаются определенные различия между полами: классы модальных частот у мужчин и женщин не совпадают, а протяженность вариационного ряда различается. Степень сходства полов по разным признакам для разных рас неодинакова. Это явление отчетливее наблюдается в группе ключевых признаков, где половые различия выражены сильнее по сравнению с неключевыми.
В пределах расы гетерогенность мужских и женских выборок, как правило, неодинакова. В одних случаях более разнородны женщины (европеоиды, монголоиды, австралоиды), в других - мужчины (индейцы Америки). Для негроидов характерно неустойчивое соотношение вариабельности полов. Все выше отмеченное относится к ключевым признакам: по неключевым признакам картина сравнительной вариабельности мужских и женских выборок менее определенна (Табл. 6.1).
6.4. Абсолютный показатель половых различий
Внутрирасовый уровень
Обширный материал, относящийся к европеоидной и монголоидной расам, был организован в рамках трех таксономических уровней: селенного, территориального иэтнического. Основные параметры вариационных рядов выборочных величин АПР, выраженных в единицах измерения признаков, представлены в Таблице 6.2.
119

Таблица 6.1. Основные параметры вариационных рядов ключевых признаков дерматоглифики у мужчин и женщин в пределах больших рас (территориальный уровень изменчивости).Table 6.1. Main parameters of variational series of key traits within the bounds of major races (territorial level of variation).
Dlio Ic t Hy ДМТ Th/I
ЕВРОПЕОИДЫ
n 3 101 ? 3 108 ? 3 98 ? 3 99 ? 3 98 ? 3 99 ?
X 12.61 12.52 8.53 8.42 60.4 57.4 32.6 35.2 19.2 16.5 9.8 8.1
s 0.63 0.67 0.35 0.32 6.83 6.61 5.53 5.95 5.91 5.43 3.76 3.89
V 5.0 5.6 4.1 3.8 11.3 11.5 17.0 16.9 30.7 32.9 38.4 47.8
МОНГОЛОИДЫ
n 3 127 ? 3 124 ? 3 92 ? 3 120 ? 3 107 ? 3 117 ?
X 14.40 13.85 7.65 7.50 77.7 71.7 21.4 25.1 12.6 10.8 9.8 7.4
s 0.85 0.92 0.59 0.57 9.34 10.04 6.58 7.87 5.52 4.92 4.72 3.91
V 5.9 6.8 7.7 7.6 12.0 14.0 30.7 31.4 43.7 45.6 48.4 53.1
примечание: n - число параллельно ооследованных мужских и женских выоорок. мвропеоиды - суммарная группа Европы и Кавказа. Монголоиды - суммарная серия Северной и Южной Азии. Note: n - number of samples (every one includes males and females). Caucasoids are presented by random sample from Europe and Caucasus. Mongoloids are presented by total group from North and South Asia.

Таблица 6.1. Продолжение. Table 6.1. Continued.
D l io I c t н у ДМТ Th/I
НЕГРОИДЫ
n 3 49 ? 3 29 ? (? 30 ? 3 56 ? с ? 47 ? с ? 53 ?
X 12.78 12.14 7.23 7.03 56 49.7 23.4 23.3 39 34.5 15.1 15.0
s 0.9 0.90 0.44 0.34 11.1 11.9 7.9 6.0 10.3 6.0 5.7 8.4
V 6.8 7.4 6.1 4.8 20.2 23.9 33.8 25.6 25.0 17.3 37.9 55.8
АВСТРАЛОИДЫ
n 3 39 ? 3 28 ? <? 10 ? с ? 29 ? с ? 26 ? с ? 23 ?
X 15.76 15.30 8.00 8.05 63.1 53.7 22.9 22.6 13.6 10.8 18.8 20.2
s 1.32 1.30 0.76 0.85 4.1 12.4 7.8 9.9 7.9 6.4 10.8 14.4
V 8.4 8.5 9.5 10.6 6.4 23.1 34.2 44.0 57.6 59.5 57.6 71.4
АМЕРИКАНОИДЫ
n 3 27 ? 3 22 ? c ? 6 9 с ? 9 9
00 9 с ? 9 9X 13.54 13.19 7.20 7.13 51.3 47.2 12.6 13.5 10.4 6.3 25.6 22.1
s 1.27 1.63 0.53 0.52 15.1 12.5 4.8 5.0 6.9 3.9 8.1 6.3
V 9.4 12.4 7.4 7.3 29.4 26.4 38.3 37.2 66.7 62.1 31.6 28.5

Таблица 6.2. Основные параметры вариационных рядов абсолютных половых разниц по ключевым признакам и результаты тестирования значимости АПР по t-критерию Стьюдента.Table 6.2. Main parameters of variational series of ASD (absolute sexual differences of key traits) and results of testing significance o f ASD by Student’s test.
Раса, Дельтовый индекс Индекс Камминса Осевой трирадиус tУровень n X s t n X s t n X s tE - c 100 0.81 0.54 15.0 102 0.32 0.26 12.4 92 8.9 5.98 14.5E - т 163 0.81 0.57 18.1 159 0.34 0.24 17.9 150 9.3 6.92 16.4Е - э 107 0.68 0.48 14.6 105 0.28 0.20 14.3 94 7.4 5.19 13.8E сумм. 370 0.77 0.54 27.4 366 0.32 0.24 25.5 336 8.6 6.23 25.3M -c 20 0.54 0.48 5.0 20 0.38 0.22 7.7 17 9.3 7.15 5.4M - t 132 0.65 0.44 17.0 124 0.27 0.23 13.1 102 7.0 5.25 13.5М -э 78 0.61 0.38 14.2 74 0.28 0.22 10.9 65 6.1 3.71 13.2M сумм. 230 0.63 0.43 22.2 218 0.28 0.23 18.0 167 6.8 4.96 17.7H - т 52 0.78 0.52 10.8 37 0.37 0.32 7.0 32 10.0 7.11 8.6Ав - т 37 0.54 0.35 9.4 29 0.26 0.20 7.0 11 10.6 8.78 4.0Am - t 27 0.69 0.46 7.8 22 0.22 0.18 5.7 6 8.0 4.27 4.6
Обозначение рас: E - европеоиды (Кавказа и Европы), М - монголоиды (северные и южные суммарно), Н - негроиды, Ам - американоиды, Ав - австралоиды. Уровни: с - селенной, т - территориальный, э - этнический. Notation of races: Е - Caucasoids (from Caucasus and Europe), M - Mongoloids (North and South Asia), H - African aboriginals, Am - American aboriginals, Ав - Australian aboriginals. Levels: с - settlements, т - territorial, э - ethnical.

Таблица 6.2.Table 6.2. Continued. Таблица 6.2. Продолжение. Table 6.2. Continued.
Раса,Уровень
Узоры на Ну ДМГ Узоры н аТ М
п X S t п X S t п X S tЕ - с 96 5.7 4.95 11.3 62 5.4 4.35 9.8 97 3.8 2.65 14.1
Е - т 156 6.2 5.03 15.4 149 5.9 4.35 16.6 151 4.1 2.97 17.0
Е - э 105 4.3 3.53 12.5 93 5.7 4.36 12.6 105 3.0 2.55 12.0
Е сумм. 357 5.5 4.67 22.2 304 5.7 4.34 22.9 353 3.7 2.80 24.8
М -с 20 5.8 3.80 6.8 19 4.3 3.28 5.7 20 3.7 3.13 5.3
М - т 118 6.0 4.45 14.6 111 4.0 3.01 14.0 115 3.6 3.34 11.6
М -э 76 4.8 3.90 10.7 56 3.4 2.42 10.5 65 3.2 2.31 11.2
М сумм. 214 5.6 4.23 19.4 206 3.6 2.86 18.1 200 3.5 3.02 16.4
Н - т 60 4.5 3.60 9.7 46 8.4 6.45 8.8 57 5.3 4.88 8.2

В пределах европеоидной расы максимальные различия полов отмечены на территориальном уровне, минимальные - на этническом. Среди монголоидов наибольшая величина АПР характеризует уровень отдельных поселений, минимальная - этнический. В методическом плане этот момент важен, так как при сопоставлении выборок разного таксономического ранга результаты могут быть неверными, отражая отмеченный факт.
В пределах европеоидной расы выборки территориального и этнического уровней по t-критерию Стьюдента значимо различаются по величине АПР всех ключевых признаков, что делает недопустимым суммирование территориальных и среднеэтнических выборок при анализе полового диморфизма (Табл. 6.2). В то же время в монголоидных выборках значимые различия между разными таксономическими уровнями не обнаруживаются.
В пределах каждой из больших рас материал был изучен не только по таксономическим уровням, но и по регионам. Таблица 6.3 может служить в качестве опорной при определении величины показателя конкретной европеоидной или монголоидной выборки.
Уровень больших рас
Практически у всех признаков, в том числе ключевых, частоты АПР в пределах основных рас распределение характеризуется резко выраженной концентрацией максимальных частот в области малых величин. Единичные исключения, как правило, наблюдаются в малочисленных расовых группах, чаще всего у американоидов и австралоидов. В прямой связи с типом распределения находится степень совпадения кривых, относящихся к различным расам, с теми же оговорками по поводу малочисленных групп.
Различия в величине признаков у мужских и женских групп разных рас приведены в Таблице 6.4. Величины АПР в основном варьируют независимо и ненаправленно, значения коэффициентов вариации АПР высоки и варьируют от 54 до 106%. Для всех рас, кроме американоидов, характерна максимальная или почти максимальная стабильность АПР по величине Dl10. Обратным свойством отмечена узорность Th/I (у монголоидов, австралоидов и негроидов она наиболее нестабильна). Как правило, вариабельность АПР неключевых признаков значительно превосходит таковую ключевых признаков. Систематические различия между расами в отношении вариабельности АПР и между признаками внутри расы не наблюдаются, за редкими исключениями.
124

Таблица 6.3. Основные параметры вариационных рядов абсолютных половых разниц по ключевым признакам внутри подразделений европеоидной и монголоидной рас.Table 6.3. Main parameters of variational series of ASD (absolute sexual differences of key traits) within Mongoloid and Caucasoid races (different levels of variation).
Раса, уровень Dlio Ic tn x s n x s n x s
ЕВРОПЕОИДЫЕвропа - с 40 0.73 0.54 40 0.28 0.24 39 7.9 5.44Европа - т 82 0.70 0.46 80 0.28 0.20 75 8.2 5.84Европа - э 40 0.62 0.39 39 0.28 0.21 32 8.0 5.10Кавказ - с 30 0.77 0.46 32 0.37 0.29 24 9.8 5.53Кавказ - т 24 0.94 0.66 24 0.42 0.24 23 6.7 6.20Кавказ - э 42 0.76 0.60 42 0.31 0.21 42 7.2 5.70Передняя Азия - т 7 0.69 0.25 6 0.31 0.22 - - -Передняя Азия - э 15 0.75 0.49 14 0.20 0.19 10 5.1 3.34Средняя Азия - с 30 0.95 0.59 30 0.31 0.25 29 9.6 6.70Средняя Азия - т 50 0.95 0.68 49 0.39 0.28 48 11.3 7.85Средняя Азия - э 10 0.59 0.18 10 0.23 0.17 10 8.8 4.70
МОНГОЛОИДЫСеверная Азия - т 88 0.70 0.51 88 0.29 0.25 87 7.1 5.38Северная Азия - э 52 0.59 0.41 51 0.29 0.22 51 5.9 3.25Южная Азия - т 44 0.54 0.26 36 0.20 0.16 15 6.3 4.51Южная Азия - э 26 0.65 0.33 23 0.26 0.23 14 6.0 4.35
Раса, уровеньУзоры на Hy ДМТ Узоры на Th/I
n x s n x s n x sЕВРОПЕОИДЫ
Европа - с 38 4.2 3.65 21 4.4 3.47 38 3.3 2.30Европа - т 76 5.9 5.02 75 5.1 3.75 76 4.0 3.20Европа - э 38 3.6 2.90 32 5.3 4.22 38 2.1 1.46Кавказ - с 28 6.2 5.59 13 4.7 3.54 29 5.1 2.92Кавказ - т 24 5.3 4.67 24 6.9 4.73 23 4.7 2.78Кавказ - э 42 5.7 4.25 41 5.9 4.47 42 3.9 2.80Передняя Азия - т 6 7.1 4.91 5 1.0 1.05 5 2.5 2.09Передняя Азия - э 15 2.7 2.56 10 8.2 4.89 15 3.5 3.20Средняя Азия - с 30 7.2 5.34 28 6.5 5.10 30 3.0 2.35Средняя Азия - т 50 6.8 5.26 45 7.0 4.80 47 4.2 2.73Средняя Азия - э 10 3.6 1.80 10 3.9 2.57 10 2.3 2.39
МОНГОЛОИДЫСеверная Азия - т 87 6.6 4.65 85 4.2 3.30 87 3.9 3.62Северная Азия - э 53 5.0 4.10 50 3.2 2.24 51 3.5 2.42Южная Азия - т 31 4.4 3.07 26 3.4 1.70 28 2.7 2.09Южная Азия - э 23 4.3 3.44 6 4.8 3.26 14 1.8 1.26
Примечание: n - число выборок, x - величина АПР, s - среднее квадратическое отклонение. Обозначение уровней как в Табл. 6.2.Note: n - number of samples, x - ASD’s value, s - average quadratic deviation. Levels: c - settlements, т - territorial, э - ethnical.
125

Таблица 6.4. Среднерасовая величина и вариабельность АПР (по рядам выборочных средних территориального уровня).Table 6.4. Average racial value and variability of ASD (in series of selected average values for territorial groups).
ПризнакиЕвропеоиды Монголоиды НегроидыАПР v АПР v АПР v
Dlw 0.81 70.5 0.65 68.4 0.78 66.5Ic 0.34 71.3 0.27 86.2 0.37 86.4t 9.3 74.5 7.0 75.2 10.0 71.1Hy 6.2 81.5 6.0 82.0 4.5 79.5ДМТ 5.9 74.2 4.0 75.5 8.4 76.8Th/I 4.1 72.3 3.6 92.6 5.3 92.3Модуль I 12.0 9.8 13.6Модуль II 12.2 10.3 14.5
ПризнакиАвстралоиды Американоиды sАПР v АПР v АПР
Dlw 0.54 64.8 0.69 67.3 0.69Ic 0.26 76.8 0.22 80.3 0.29t 10.6 82.6 8.0 53.7 9.0Hy 4.7 93.2 6.4 77.7 5.6ДМТ 5.2 63.5 5.0 88.5 5.7Th/I 5.1 105.6 6.6 60.5 4.9Модуль I 11.0 11.0 11.7Модуль II 12.3 13.2 12.8
Примечание: E - общечеловеческая невзвешенная средняя АПР. Модуль I и II - модуль АПР по пяти и шести ключевым признакам соответственно.Note: E - universal unweighted average of ASD. Модуль I и II - ASD’s module calculated for 5 and 6 key traits, respectively.
Рассмотрим подробнее вопрос о различиях между расами по степени дифференциации полов в отношении отдельных признаков и их суммы. С этой целью величины АПР выражались в процентах евразийской амплитуды соответствующих признаков, охватывающей оба пола (См. Главу 3). Этот прием позволяет сравнивать расы как по отдельным признакам, так и по среднему стандартизованному показателю - модулю АПР. Последний вычислялся усреднением частных стандартизованных (т.е. выраженных в процентах евразиатской амплитуды признака) величин АПР для разного числа ключевых признаков - пяти и шести (в последнем случае учитывались узоры на Th/I). Индивидуальные графики рас по стандартизованной величине АПР говорят прежде всего о том, что размах различий между расами сужен в отношении
126

обоих индексов и узорности Hy, последовательно увеличивается по проксимальному осевому трирадиусу t и ДМТ и достигает максимума по узорности Th/I (Рис. 6.2).
АПР
О -!■ I-------1------ 1------ 1------ 1------ 1 I-------1------ 1-------1------ 1------ 11)1,и Ic I Н> Д М Т Th/I Dim Ic t Ну Д М Т Th/1
Рисунок 6.2. Сопоставление больших рас по стандартизованной величине АПР и модулю АПР. Примечание: a) профили рас по величине АПР: по горизонтали - признаки, по вертикали - стандартизованная величина АПР; б) то же для человечества в целом (невзвешенная мировая средняя); в) среднерасовая величина модуля АПР по ключевым признакам. Слева - без учета, справа - с учетом Th/I; n - число признаков; Е - человечество в целом. Обозначение рас: Е - европеоиды,М - монголоиды, Н - негроиды, Ам - американоиды, Ав - австралоиды. Figure 6.2. Comparison among major races based on data of standardized values of ASD and ASD module. Note: a) racial profiles on ASD values: traits are posited on abscissa axis, standardized values of ASD are posited on ordinate axis; б) same data for mankind in whole (unweighted world average); в) racial average of ASD module based on 5 key traits (without Th/I, on the left) or 6 key traits (on the right). Designation of races: E - Caucasoids; М - Mongoloids; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals, Е - mankind in whole.
Вместе с тем, вырисовывается общий тип кривой для всех рас, что свидетельствует о единых закономерностях. Наибольшее уклонение от этого типа характеризует негроидов, у которых по всем ключевым признакам, кроме Hy, значения АПР экстремальны или субэкстремальны. Противоположное свойство присуще монголоидной расе, которая по половине числа ключевых признаков имеет наименьшую величину АПР (t, ДМТ, Th/I). Что касается австралоидов, то эта раса наиболее (или почти самая) мономорфная в отношении обоих индексов и Hy, но в то же время самая диморфная по трирадиусу t.
127

Индейцы характеризуются минимальными различиями полов по индексу Камминса и максимальными различиями в отношении Hy и, особенно, по типично «индейскому» признаку - Th/I. Европеоиды ни в одном случае не занимают крайней позиции, но сближаются с негроидами (DlJ0) и монголоидами (Th/I). Степень сходства рас имеет сетеобразный характер, меняясь от признака к признаку.
Переходя к анализу различий между полами по сумме признаков, рассмотрим модуль АПР. По его величине выделяются группировки в схеме расположения рас в отношении друг к другу, а также к мировой стандартной величине модуля, полученной усреднением частных среднерасовых значений. Как видно из Таблицы 6.4, средние различия между полами, приходящиеся на один признак, составляют для человечества в целом 12-13% евразиатской шкалы изменчивости.
Оценивая величину модуля, можно сказать, что в ряду рас негроиды характеризуются наиболее, а монголоиды - наименее выраженными различиями между полами (независимо от числа анализируемых признаков). Европеоиды располагаются примерно посередине между ними, положение остальных рас меняется в зависимости от набора признаков. Так, по сумме ключевых признаков без учета Th/I австралоиды и американоиды обнаруживают одинаковую степень дифференциации полов и тяготеют к монголоидам. С учетом Th/I австралоиды сближаются с монголоидами, а индейцы - с негроидами. Таким образом, включение Th/I существенно меняет соотношение вариантов восточного ствола по модулю АПР.
Человечество в целом
Среднерасовые показатели АПР были протестированы на гомогенность с помощью рангово-дисперсионного анализа по Фридману. В результате различия между всеми расами, рассмотренными одновременно по комплексу ключевых признаков, оказались незначимыми. Таким образом, в отношении дифференциации полов все расы следует признать выборками из одной совокупности. «Общечеловеческая» криваястандартизованной величины АПР по комплексу признаков показывает наименьшие различия полов в отношении Ic и Dl10, далее следуют Hy, ДМТ, t, Th/I (см. Рис. 6.2). «Общечеловеческий» модуль
128

АПР, суммирующий все признаки, естественным образом занимает срединное положение на шкале вариаций среднерасовых значений. Наиболее близкой к мировой средней является европеоидная раса, наиболее далеки от нее, при этом в равной степени, негроиды и монголоиды.
Основные выводы:
• В пределах европеоидной и монголоидной рас наименьшая величина АПР характеризует этнический уровень. Оба пола максимально различаются среди европеоидов на территориальном уровне, а среди монголоидов на уровне селенных выборок.
• При межрасовых сопоставлениях обнаружено отсутствие четкой связи между величиной АПР и ее межгрупповой вариабельностью. Коэффициенты вариации АПР ключевых признаков высоки (54-106%) и еще более увеличены у неключевых.
• Между расами существуют различия в степени дифференцированности полов. Максимально мономорфны монголоиды, наиболее диморфны негроиды. Американоиды и австралоиды сближаются с монголоидами. Европеоиды характеризуются промежуточным уровнем половых различий. Включение в анализ узорности Th/I сближает характеристики австралоидов и американоидов с негроидами.
• Распределение величины АПР по признакам обнаруживает сходство между всеми расами, яснее всего проявляющееся в суммарной общечеловеческой выборке. Различия между полами увеличиваются в следующем порядке: Ic, DlJ0, Hy, ДМТ, t, Th/I.
6.5. Относительный показатель полового диморфизма (КПД)
В данном разделе рассматриваются основные параметры вариационных рядов коэффициентов полового диморфизма на разных таксономических уровнях.
Внутрирасовый уровень. Выборки европеоидов и монголоидов как самые многочисленные анализируются на селенном, территориальном и этническом уровнях. Основные параметры вариационных рядов КПД ключевых признаков содержатся в Таблице 6.5.
129

Таблица 6.5. Основные параметры вариационных рядов коэффициента полового диморфизма (КПД) на внутрирасовом уровне (ключевые признаки).Table 6.5. Main parameters of variability of CSD (coefficients of sexual differences of key traits) in intra-racial level.
Раса, Dlio Ic tУровень n x s v n x s v N x s V
Е - с 98 1.06 0.06 6.0 100 1.02 0.04 4.4 88 1.14 0.15 13.5
E - т 157 1.06 0.06 5.6 154 1.02 0.04 4.4 144 1.17 0.18 15.6
Е - э 106 1.05 0.05 4.9 103 1.03 0.04 3.4 94 1.11 0.14 12.2
E сумм. 361 1.06 0.06 5.5 357 1.02 0.04 4.1 326 1.14 0.16 14.3
М - с 19 1.04 0.05 4.8 20 1.04 0.04 4.2 17 1.10 0.16 14.4
М - т 126 1.04 0.05 5.2 124 1.03 0.04 4.4 91 1.09 0.13 10.3
М - э 78 1.03 0.04 4.4 74 1.02 0.04 4.5 65 1.08 0.09 8.4
М сумм. 223 1.04 0.05 4.9 218 1.03 0.04 4.4 173 1.09 0.11 10.1
Н - т 49 1.06 0.06 5.8 37 1.03 0.07 6.5 28 1.14 0.20 17.3
Ав - т 39 1.03 0.05 4.2 29 0.99 0.04 3.9 10 1.09 0.20 18.4
А м ! т 27 1.04 0.06 5.5 22 1.01 0.04 4.1 6 1.10 0.17 15.5
Раса, Узоры на Hy ДМТ Узоры на Th/IУровень n x s v n x s v n x s v
Е - с 94 0.96 0.22 23.4 59 1.30 0.55 42.1 93 1.07 0.58 54.4
E - т 148 0.97 0.24 24.6 144 1.20 0.47 39.4 145 1.41 1.20 85.2
Е - э 105 0.99 0.19 19.4 93 1.26 0.36 29.1 107 1.15 0.63 54.7
E сумм. 347 0.97 0.22 22.7 296 1.24 0.46 36.8 345 1.24 0.92 73.9
М - с 20 0.90 0.23 25.3 19 1.09 0.67 61.1 20 1.35 0.67 49.7
М - т 117 0.90 0.30 33.0 109 1.26 0.58 46.4 111 1.51 1.01 67.1
М - э 75 0.89 0.23 26.0 57 1.26 0.69 54.8 65 1.41 0.60 34.4
М сумм. 212 0.90 0.27 30.0 185 1.26 0.61 48.7 196 1.46 0.86 59.2
Н - т 57 1.05 0.32 30.4 45 1.33 0.35 26.6 53 1.29 0.74 57.5
А в ! т 22 1.10 0.44 40.1 23 1.25 0.74 59.5 16 0.88 0.25 28.4
Ам - т 8 0.90 0.46 50.8 7 1.25 0.85 68.0 9 1.23 0.47 37.9
Примечание: Е - европеоиды; М - монголоиды; Н - негроиды; Ав - австралоиды; Ам - американоиды. Уровни: с - селенной; т -территориальный; э - этнический; сумм. - суммарно.Note: E - Caucasoids, M - Mongoloids, H - African aboriginals, Лм - American aboriginals, Ав - Australian aboriginals. Levels: c - settlements, т - territorial, э - ethnical; cумм. - combined sample.
130

Прежде всего следует обратить внимание на сходство обеих рас в соотносительной величине КПД. Минимальные значения КПД характеризуют оба индекса. Далее идет последовательное увеличение параметра, достигающее максимума в отношении узорности Th/I и варианта Сх . Кроме того, по всем признакам на всех уровнях в обеих расах значение признака у мужчин выше, чем у женщин (КПД больше 1).
Исключение составляют узорность Ну у обеих рас и частота отсутствия линии С у монголоидов (КПД меньше 1). Что касается соотношения уровней, то в пределах европеоидной расы определенной закономерности нет: по одним признакам наибольший КПД имеют территориальные выборки (t, Th/I, C0), по другим - селенные (Hy, ДМТ, Cx). Этнический уровень ни в одном случае не отмечен максимальной величиной КПД. Эти же наблюдения относятся к монголоидам.
В целом отмечается сходство обеих рас по иерархии таксономических уровней, отчетливо проявляющееся в отношении большинства признаков. В пределах монголоидной расы не обнаружено значимых различий между таксономическими уровнями по величине КПД. Среди европеоидов значимо различаются территориальный и этнический уровни по Ic и t, а также селенной и территориальный по Th/I. Таким образом, в отношении дифференциации полов иерархическая структура у европеоидной расы более выражена, чем у монголоидной. Сравнивая европеоидов и монголоидов на одноименных таксономических уровнях, отметим, что величина КПД в пределах европеоидной расы варьирует бессистемно (Таблица 6.6).
Можно отметить лишь несколько большую дифференциацию полов у южных европеоидов по индексу Камминса по сравнению с северными. Кроме того, в этнических и территориальных группах европеоидов Средней Азии наиболее сильно выражены половые различия в отношении трирадиуса ладони t. Европеоиды Кавказа обнаруживают исключительно большую величину КПД узорности Th/I в территориальных, а также ДМТ - в этнических группах. Также не направленными и бессистемными являются различия между северными и южными монголоидами; в одних случаях первые сильнее дифференцированы по полу (Ic), в других - слабее (ДМТ).
131

Таблица 6.6. Основные параметры вариационных рядов коэффициентов полового диморфизма (КПД) в пределах подразделений европеоидной и монголоидной рас.Table 6.6. Main parameters of variability of CSD (coefficients of sexual differences of key traits) within Mongoloid and Caucasoid races (different taxonomic levels of variation).
Раса, уровень Dl10 Ic tn x 1 s n 1 x s n 1 x 1 s
ЕВРОПЕОИДЫЕвропа - с 40 1.06 0.06 40 1.02 0.05 38 1.14 0.14Eвропа - т 82 1.05 0.05 80 1.02 0.04 75 1.15 0.17Европа - э 39 1.05 0.04 37 1.03 0.03 32 1.13 0.11Кавказ - с 28 1.06 0.07 30 1.03 0.04 21 1.13 0.16Кавказ - т 23 1.07 0.05 23 1.04 0.05 22 1.11 0.19Кавказ - э 42 1.06 0.06 42 1.03 0.04 42 1.09 0.16Передняя Азия - т 7 1.05 0.02 6 1.03 0.03 - - -Передняя Азия - э 15 1.04 0.06 14 1.01 0.03 10 1.08 0.09Средняя Азия - с 30 1.06 0.07 30 1.02 0.04 29 1.13 0.16Средняя Азия - т 45 1.06 0.07 45 1.03 0.05 43 1.20 0.17Средняя Азия - э 10 1.04 0.02 10 1.03 0.02 10 1.15 0.09
МОНГОЛОИДЫСеверная Азия - т 87 1.04 0.06 87 1.03 0.05 84 1.09 0.12Северная Азия - э 52 1.03 0.04 52 1.04 0.04 51 1.08 0.09Южная Азия - т 39 1.02 0.04 37 1.02 0.03 7 1.09 0.10Южная Азия - э 26 1.04 0.04 22 0.99 0.04 14 1.09 0.08Раса, уровень Узоры на Hy ДМТ Узоры на Th/I
n x s n x s n x sЕВРОПЕОИДЫ
Европа - с 38 0.99 0.16 21 1.44 0.62 37 1.23 0.61Eвропа- т 74 0.98 0.24 75 1.21 0.41 74 1.49 1.22Европа - э 38 0.99 0.17 31 1.27 0.38 40 1.22 1.51Кавказ - с 26 0.99 0.26 9 1.10 0.31 26 0.85 0.48Кавказ - т 23 0.96 0.22 23 1.14 0.62 23 1.67 1.70Кавказ - э 42 0.96 0.20 42 1.23 0.33 42 1.05 0.77Передняя Азия - т 6 0.84 0.12 5 1.01 0.06 5 0.94 0.37Передняя Азия - э 15 0.96 0.10 10 1.29 0.49 15 1.23 0.58Средняя Азия - с 30 0.85 0.20 29 1.27 0.52 30 1.07 0.57Средняя Азия - т 45 0.89 0.20 41 1.25 0.51 43 1.22 0.82Средняя Азия - э 10 0.89 0.06 10 1.27 0.29 10 1.17 0.34
МОНГОЛОИДЫСеверная Азия - т 87 0.88 0.30 85 1.23 0.63 84 1.48 1.11Северная Азия - э 52 0.89 0.19 51 1.24 0.64 51 1.40 0.64Южная Азия - т 30 0.92 0.24 24 1.35 0.36 27 1.61 0.64Южная Азия - э 23 0.83 0.25 6 1.39 0.97 14 1.41 0.48
Уровни: с - селенной; т - территориальный; э - этнический. Levels: c - settlements, т - territorial, э - ethnical.
132
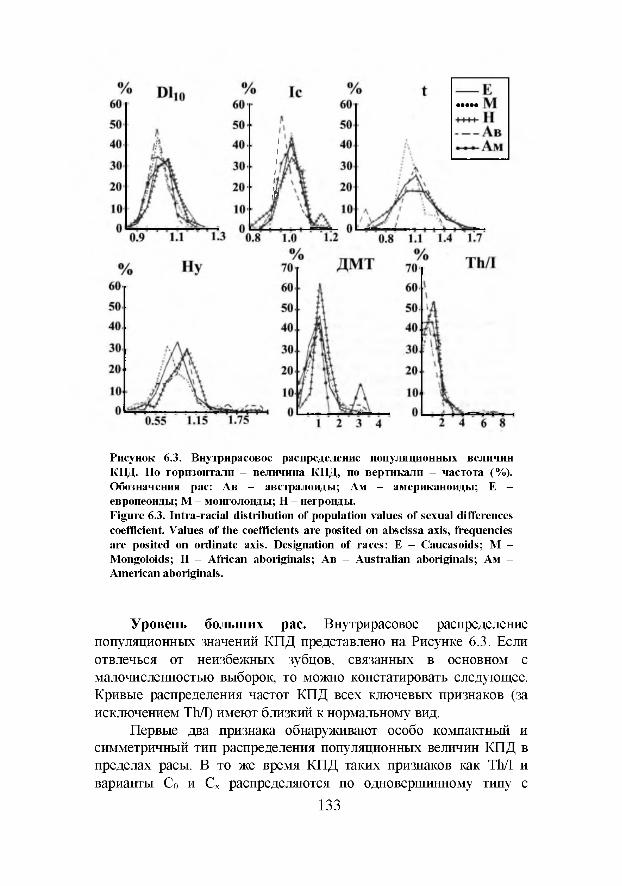
Рисунок 6.3. Внутрирасовое распределение популяционных величин КПД. По горизонтали - величина КПД, по вертикали - частота (%). Обозначения рас: Ав - австралоиды; Ам - американоиды; Е - европеоиды; М - монголоиды; Н - негроиды.Figure 6.3. Intra-racial distribution of population values of sexual differences coefficient. Values of the coefficients are posited on abscissa axis, frequencies are posited on ordinate axis. Designation of races: E - Caucasoids; М - Mongoloids; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
Уровень больших рас. Внутрирасовое распределение популяционных значений КПД представлено на Рисунке 6.3. Если отвлечься от неизбежных зубцов, связанных в основном с малочисленностью выборок, то можно констатировать следующее. Кривые распределения частот КПД всех ключевых признаков (за исключением Th/I) имеют близкий к нормальному вид.
Первые два признака обнаруживают особо компактный и симметричный тип распределения популяционных величин КПД в пределах расы. В то же время КПД таких признаков как Th/I и варианты С0 и Сх распределяются по одновершинному типу с
133

выраженной правосторонней асимметрией, а у австралоидов распределение КПД узорности Th/I имеет параболический вид. Между расами наблюдаются нерегулярные взаимоотношения по степени сходства кривых КПД. В одних случаях расы тесно сближены (варианты линии С), в других различия между расами вполне отчетливы, как и взаимное сходство некоторых из них. Сильнее всего различия между расами выражены в кривых распределения КПД узорности Ну.
Среднерасовая величина КПД по DlJ0 колеблется от 1.03 до1.06, по Ic - от 0.994 до 1.03, по трирадиусу ладони t - от 1.09 до 1.17, по узорности Ну - от 0.90 до 1.10, по ДМТ - от 1.20 до 1.33. Огромны различия между расами в колебаниях среднерасовых значений КПД узорности Th/I (0.88-1.51!). КПД варианта линии С0 варьирует от 0.72 до 1.04. Наблюдается отчетливая иерархия признаков в отношении величины КПД. В пределах каждой расы величина КПД возрастает в последовательности: Ic - Dl10 - проксимальный трирадиус t - ДМТ. Что касается узорности Th/I, то величина ее КПД максимальна в ряду признаков у монголоидов, в то время как у негроидов и американоидов она почти равна КПД по ДМТ. Таким образом, в рамках общей для всех рас тенденции отчетливо вырисовывается иерархия указанных ключевых признаков по величине относительных различий между полами. В этот ряд не вписывается КПД узорности гипотенарной и тенарной подушечек, о которых речь пойдет дальше.
Перейдем к сравнительной характеристике рас по величине КПД. Как следует из Рисунка 6.4 и Таблицы 6.7, для всех рас характерно преобладание величины признака у мужчин (КПД больше 1), за редкими исключениями (узорность Ну и Th/I).
По узорности гипотенара человечество разделяется на две полярные группы. В одну входят экваториальные расы (негроиды и австралоиды), которым свойственно преобладание величины признака у мужчин. В другую группу с обратным соотношением входят остальные расы. В случае с Th/I отчетливо обособляются австралоиды (КПД=0.88), в то время как для остальных рас характерны сильное преобладание признака у мужчин.
4 Среднерасовая величина КПД на одну сотую не достигает 1 лишь у австралоидов. Учитывая небольшое число этих выборок и незначительность отклонения КПД от 1, мы можем допустить здесь влияние случайного выборочного эффекта на показатель.
134

Таблица 6.7. Среднерасовая величина КПД (по рядам выборочных средних территориального уровня).Table 6.7. Average racial value and variability of CSD (series of sample averages for territorial groups).
Признаки Европеоиды Монголоиды Негроиды
Dl10 1.06 1.04 1.06Ic 1.02 1.03 1.03t 1.17 1.09 1.14
Hy 0.97 0.90 1.05ДМТ 1.20 1.26 1.33Th/I 1.41 1.51 1.29
Признаки Австралоиды Американоиды Е
Dl10 1.03 1.04 1.05Ic 0.99 1.01 1.02t 1.09 1.10 1.12
Hy 1.10 0.90 0.98ДМТ 1.25 1.25 1.26Th/I 0.88 1.23 1.26
Примечание: E - невзвешенная мировая средняя КПД.Note: E - universal unweighted average of ASD.
При сравнении рас по величине КПД набора ключевых признаков легко заметить, что обе экваториальные расы, как правило, полярны; так негроиды по большинству элементов кожного рельефа характеризуются максимальными или субмаксимальными значениями КПД (оба индекса, трирадиус t, ДМТ). У австралоидов величина КПД всех ключевых признаков минимальна. Остальные расы чаще всего занимают промежуточное положение между этими вариантами. Следует отметить, что монголоиды и американоиды систематически сближаются по величине КПД большинства ключевых признаков.
Представление о средней величине КПД по комплексу признаков можно получить, применив модуль КПД - величину, полученную усреднением частных значений КПД отдельных признаков. Для сопоставления рас сравнимы лишь модули, вычисленные с использованием тех признаков, тенденции полового диморфизма которых одинаковы у всех рас. Таким образом из числа основных признаков исключается узорность Ну и Th/I (Рис. 6.4).
135
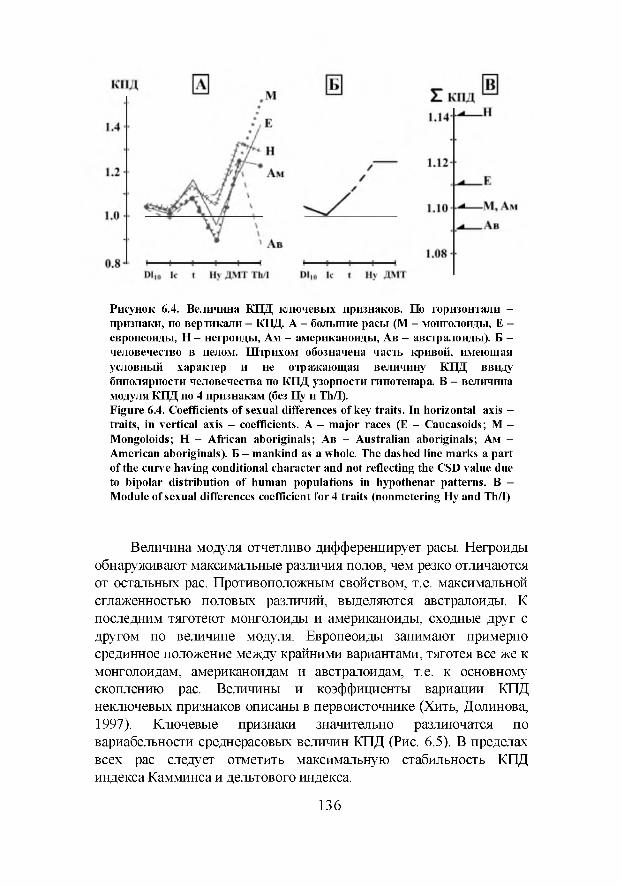
Рисунок 6.4. Величина КПД ключевых признаков. По горизонтали - признаки, по вертикали - КПД. А - большие расы (М - монголоиды, Е - европеоиды, Н - негроиды, Ам - американоиды, Ав - австралоиды). Б - человечество в целом. Штрихом обозначена часть кривой, имеющая условный характер и не отражающая величину КПД ввиду биполярности человечества по КПД узорности гипотенара. В - величина модуля КПД по 4 признакам (без Ну и Th/I).Figure 6.4. Coefficients of sexual differences of key traits. In horizontal axis - traits, in vertical axis - coefficients. A - major races (E - Caucasoids; М - Mongoloids; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals). Б - mankind as a whole. The dashed line marks a part of the curve having conditional character and not reflecting the CSD value due to bipolar distribution of human populations in hypothenar patterns. B - Module of sexual differences coefficient for 4 traits (nonmetering Hy and Th/I)
Величина модуля отчетливо дифференцирует расы. Негроиды обнаруживают максимальные различия полов, чем резко отличаются от остальных рас. Противоположным свойством, т.е. максимальной сглаженностью половых различий, выделяются австралоиды. К последним тяготеют монголоиды и американоиды, сходные друг с другом по величине модуля. Европеоиды занимают примерно срединное положение между крайними вариантами, тяготея все же к монголоидам, американоидам и австралоидам, т.е. к основному скоплению рас. Величины и коэффициенты вариации КПД неключевых признаков описаны в первоисточнике (Хить, Долинова, 1997). Ключевые признаки значительно разлиючатся по вариабельности среднерасовых величин КПД (Рис. 6.5). В пределах всех рас следует отметить максимальную стабильность КПД индекса Камминса и дельтового индекса.
136

1 2 3 4 5 6 1 2 3 4 5 6 1 2 3 4 5 6
Рисунок 6.5. Внутрирасовая вариабельность КПД (А) и абсолютных величин признаков (Б) в пределах каждого пола. По горизонтали - признаки, по вертикали - величина межгруппового коэффициента вариации КПД.Figure 6.5. Intra-racial variability of CSD (A) and variability of traits (Б) within the bounds of every sex. In horizontal axis - traits, in vertical axis - the inter-group variation coefficients.
Изменчивость КПД остальных признаков увеличена и возрастает в порядке: проксимальный трирадиус t - узорность Hy - ДМТ - узорность Th/I. Отдельные случаи нарушения этой схемы характеризуют американоидов, у которых вариабельность КПД узорности Hy максимальна в ряду признаков. Аналогичным образом в пределах австралоидной расы наиболее вариабелен КПД ДМТ, в то время как стабильность этого показателя в отношении узорности Th/I резко увеличена (V его КПД уменьшен). Полным антиподом австралоидов по Th/I являются европеоиды, которым свойственна максимальная вариабельность его КПД (коэффициент вариации достигает 85%).
Подводя итог, отметим, что стабильность коэффициента полового диморфизма признаков дерматоглифики внутри расы (определяемая уменьшением его коэффициента вариации) находится в прямой и, видимо, причинной связи с вариабельностью соответствующих признаков в пределах каждого пола. Чем меньше признак варьирует внутри расы, тем более стойки взаимоотношения полов по данному признаку, т.е. тем меньше коэффициент вариации соответствующего КПД (Рис. 6.5). И наоборот, чем сильнее внутрирасовая популяционная изменчивость признака, тем большей нестойкостью (выражающейся в увеличении V) характеризуется величина его КПД. Иными словами стабильность расового
137

комплекса обеспечивается не только суженной внутрирасовой вариабельностью признаков, но и повышенной стойкостью величины различий между полами. В этом смысле группа ключевых признаков резко выделяется на фоне неключевых, обнаруживая максимальную стабильность расового комплекса.
Человечество в целом.
Общие для человечества закономерности в распределении величин КПД отдельных признаков выявляются суммированием индивидуальных кривых всех рас (Рис.6.4). Все суммарные кривые одновершинны, и большинство их имеет распределение, близкое к нормальному; особенно это относится к обоим индексам, t трирадиусу ладони, а также узорности Hy и ДМТ. В двух последних случаях присутствует некоторая правосторонняя асимметрия. Еще более асимметрично мировое распределение КПД узорности Th/I и вариантов линии С. Мировые средние величины КПД признаков, полученные усреднением суммы среднерасовых значений, варьируют от 0.92 (вариант С0) до 1.26 (ДМТ и Th/I) и, как правило, превышают единицу5.
Из ключевых признаков в мировой выборке лишь узорность Hy имеет КПД ниже 1 (0.98). Величина эта - абстрактная, так как получена суммированием частных значений ниже единицы (европеоиды, монголоиды и американоиды) и превышающих единицу (негроиды и австралоиды) и, таким образом, объединяет противоположные тенденции в половом диморфизме. Мировая средняя КПД в ряду остальных ключевых признаков минимальна по обоим индексам - Ic и Dl10 (1.02 и 1.05 соответственно), заметно возрастает в отношении проксимального трирадиуса ладони (1.12) и максимальна по ДМТ и Th/I (1.26 в обоих случаях). Общечеловеческая модель КПД ключевых признаков, полученная на основе указанных цифр, представлена на Рисунке 6.4 и демонстрирует описанную иерархию признаков. Очевиден разрыв между минимальной и максимальной величинами КПД. Все признаки сильнее выражены у мужчин по сравнению с женщинами, за исключением Ну. Поскольку относительно последнего признака
5 Среднерасовые значения КПД(х) вычислены по вариационным рядам каждой расы в отдельности. Вычисление показателя по суммарным (мировым) рядам частот КПД было бы некорректным, так как в последних существенно преобладают европеоидные и монголоидные выборки.
138

общечеловеческая тенденция полового диморфизма отсутствует, разные расы следует рассматривать раздельно. По этой же причине невозможно применить дисперсионный анализ Фридмана для комплекса всех ключевых признаков. Если же применить эту процедуру к признакам со сходной тенденцией полового диморфизма у всех рас (величина признака у мужчин превышает таковую у женщин), то по четырем важнейшим ключевым признакам обнаруживается гомогенность человечества.
Основные выводы:
• Относительный показатель полового диморфизма на разных таксономических уровнях в пределах европеоидной и монголоидной рас колеблется ненаправленно. Так же неопределенно варьируют величины КПД по регионам в пределах каждой расы.
• Общее правило для всех рас человечества состоит в преобладании величины ключевых признаков у мужчин. Исключение составляют два признака - узорность Ну и Th/I. Человечество отчетливо разделяется на две неравные части по величине КПД узорности Ну: в обеих экваториальных расах (негроиды и австралоиды) КПД превышает единицу, в то время как у остальных рас ее не достигает. Кроме того, австралоиды являются единственной расой, у которой КПД узорности Th/I ниже единицы.
• Степень взаимного сходства рас по величине КПД разных признаков различна. Все расы сходны в отношении иерархии признаков по величине КПД, которая возрастает в последовательности: Ic - Dl10 - t - ДМТ. По комплексу признаков наибольшая дифференциация полов характерна для негроидной расы. Наиболее мономорфны австралоиды. Монголоиды и американоиды тяготеют к австралоидам, а европеоидная раса занимает почти срединное положение между крайними вариантами, сближаясь все же по модулю КПД с восточным расовым стволом.
• Внутрирасовая вариабельность популяционных величин КПД обнаруживает ту же иерархию признаков, что и на межрасовом уровне. Изменчивость КПД увеличивается в ряду ключевых признаков от Ic и Dl10 к Th/I.
• Несмотря на все отмеченные различия между расами, по итогам рангово-дисперсионного анализа среднерасовых значений КПД большинства ключевых признаков все расы принадлежат к единой совокупности.
139

6.6. Метод оценки величины полового диморфизма с учетом его формы
Исследователи часто сталкиваются на практике с необходимостью количественной оценки половых различий выборок. Определение различий дается субъективно и не подкрепляется специальными исследованиями проблемы. Ниже предлагается способ оценки величины КПД в мировом масштабе на основе взаимного сходства рас в отношении внутрирасового распределения частот КПД признаков (см. Рис.6.3). В основу метода положены данные рядов мирового распределения популяционных частот КПД 14 признаков (6 из них ключевые). В этих суммарных рядах подавляющая доля выборок приходится на группы европеоидного и монголоидного происхождения. Предлагаемая рубрикация имеет условный характер и, вероятно, будет изменена при накоплении данных об остальных расах.
Варианты полового диморфизма.
Установлено, что существует три разных формы или варианта полового диморфизма, определяемого отношением величины признака у мужчин в долях к таковой у женщин: а (КПД больше 1), в (КПД меньше 1) и 0 (КПД=1, т.е. различия между полами отсутствуют) (Павловский, 1981). Для всех рас в отношении каждого признака дерматоглифики, как правило, отмечаем общее свойство, которое заключается в заметном преобладании процентного содержания какой-либо из двух форм (а или в). Нулевой вариант наиболее редок. Распределение популяционных значений КПД в мировой совокупности отличается в большинстве случаев отчетливой асимметрией, что связано, прежде всего, с отмеченным выше неравновесным содержанием а и в-форм и разной численностью выборок, представляющих расу. Поэтому ранжировать ряды распределения с помощью статистических параметров невозможно, и это исключает строго объективную оценку величин КПД. Однако вполне допустим эмпирический подход, основанный на объективных посылках. По смыслу КПД, чем ближе его величина к единице, тем меньше степень различий между полами. Поэтому в ряду распределения КПД выделяется нулевой класс, где КПД=1. Остальные две части ряда распределения (справа и слева от нулевого класса) ранжируются на равные классы
140

или категории, в возрастающем порядке по мере удаления от нулевого класса. Это ранжирование отражает увеличение полового диморфизма при одновременном учете его формы. Так, примыкающие к нулевому классы получают название класса малых величин как при a-форме, так и при P-форме. Следующий класс в зонах обеих форм считается классом средних величин, следующий за ним - классом больших величин, и т.д. (Табл. 6.8).
Таким образом, в основу ранжирования положено равенство интервалов классов мирового вариационного ряда с учетом формы полового диморфизма6. Процентная насыщенность классов различна в пределах обеих форм. В рамках а- и в- форм малая величина КПД встречается наиболее часто, большая - реже всего.
При этом количество классов а- формы обычно больше по сравнению с в- формой, что отражает асимметричность распределения частот КПД. При наличии общей для всех рас закономерности полового диморфизма, состоящей преимущественно в преобладании одной из его форм, рубрикация осуществлялась по суммарному мировому ряду популяционных величин КПД.
Практически это относится ко всем рассмотренным признакам, за двумя исключениями. По первому из них (узорность Ну) расы отчетливо разделились на две группы в соответствии с преобладающей формой полового диморфизма. Для европеоидов, американоидов и монголоидов характерен в-диморфизм, для негроидов и австралоидов - a- форма. В связи с этим рубрикация категорий полового диморфизма узорности Ну для каждой из двух группировок проведена раздельно. Второе исключение касается отсутствия линии С (Со). Подавляющая часть человечества (кроме монголоидов) характеризуется большим (около 40%) преобладанием в-формы над a -вариантом, а среднерасовый КПД варьирует от 0.72 до 0.99.
6 Статистическая обработка обширного материала первоначально охватывала все классы распределения равномерным образом от минимальных до максимальных величин. В дальнейшем пришлось вычленить частоты с КПД=1 с тем, чтобы рассмотреть по отдельности распределение всех трех типов полового диморфизма. Вследствие этого границы первого класса a-формы немного сузились по сравнению с другими классами. Отметим, что случаи равных значений признаков у мужчин и женщин редки, и указанная процедура не меняет результаты сколько нибудь существенным образом.
141

Таблица 6.8. Оценка величины КПД при разных формах полового диморфизма ключевых признаков дерматоглификиTable 6.8. Evaluation of CSD’s value for different types of sexual dimorphism of key traits.
Форма ПД и категория КПД
Г раницы категорий
КПД
%групп
Границыкатегорий
КПД
%групп
Границыкатегорий
КПД
%групп
Dl10 (398) Ic (366) t (279)в большая X-0.90 - Х-0.89 0.8 Х-0.59 -в средняя 0.90-0.94 3.5 0.90-0.94 4.1 0.60-0.79 1.4в малая 09.5-0.99 14.3 09.5-0.99 24.9 0.80-0.99 16.10 1.00 5.2 1.00 9.3 1.00 2.9а малая 1.01-1.04 32.7 1.01-1.04 34.4 1.01-1.19 50.9а средняя 1.05-1.09 26.7 1.05-1.09 23.5 1.20-1.39 22.3а большая 1.10-1.14 13.6 1.10-1.14 1.4 1.40-1.59 5.4а очень большая 1.15-х 4.0 1.15-х 1.6 1.60-x 1.0
ДМТ (328) Hy (273): E+M+Aм
Hy (79): Н+Ав
в очень большая - - Х-0.54 5.2 Х-0.54 3.8в большая - - 0.55-0.69 10.6 0.55-0.69 5.1в средняя x-0.49 3.7 0.70-0.84 26.0 0.70-0.84 13.9в малая 0.50-0.99 26.5 0.85-0.99 27.1 0.85-0.99 20.20 1.00 0.3 1.00 2.2 1.00 0.0а малая 1.01-1.49 47.1 1.01-1.14 14.6 1.01-1.14 30.4а средняя 1.50-1.99 15.4 1.15-1.29 6.7 1.15-1.29 13.9а большая 2.00-2.49 4.6 1.30-1.44 3.3 1.30-1.44 3.8а очень большая 2.50-x 2.4 1.45-x 4.4 1.45-x 9.8
Th/I (334)в большая в средняя в малая 0а малая а средняя а большая а очень большая
x-0.991.00
1.01-1.992.00-2.993.00-3.99
4.00-x
37.8 1.2
45.9 9.7 3.0 2.4
Примечание: в скобках - число территориальных выборок суммарного ряда популяционных величин КПД, охватывающего все расы (если нет дополнительных обозначений). Расы: Е - европеоиды, М - монголоиды, Н - негроиды Африки, Ав - австралоиды, Ам - индейцы Америки.Note: in brackets - number of territorial samples of total series of CSD including races. Races: E - Caucasoids, M - Mongoloids, H - African aboriginals, Ам - American aboriginals, Ав - Australian aboriginals.
142

У монголоидов это преобладание составляет всего 8%, а в среднем значение признака у мужчин больше, чем у женщин (среднерасовый КПД=1.04). Поэтому в обоих случаях - для монголоидов и остальных рас - рубрикация произведена раздельно. Кроме того, в отношении Сх варианта имеются сравнимые методически данные лишь о европеоидах и монголоидах. По этой причине рубрикации подвергался суммарный ряд, охватывающий данные лишь этих двух рас. Следует особо отметить специфику австралоидов, у которых, в отличие от остальных рас, в-форма резко преобладает над a-формой. Несмотря на небольшое число выборок австралоидов, они были включены в анализ общечеловеческих закономерностей полового диморфизма по указанным признакам. Предложенный метод определения величины полового диморфизма, конечно, условен, а его недостатки очевидны (неравновесность представленных расовых групп, отсутствие данных о трех расах человечества в отношении Сх). Однако его применение избавляет от субъективного подхода, неизбежно сопровождающего большинство публикаций и, главное, позволяет сохранить постоянный масштаб оценки параметра, по крайней мере, до следующей попытки обобщения всех наличных материалов.
Варианты полового диморфизма и их соотношение
Соотношение процентных частот вариантов полового диморфизма анализировалось на разных таксономических уровнях (большие расы, подразделения европеоидной и монголоидной рас).
Уровень больших рас.
Как и в предыдущих случаях, в основу анализа положены данные территориальных групп. (Табл. 6.9 и Рисунки 6.6 и 6.7). Прежде всего, обнаруживается разнообразие рас как по частотам вариантов полового диморфизма, так и по их количественному соотношению. Главная и общая закономерность, характеризующая все расы, заключается в преобладании частот a-диморфизма. Нулевой вариант у всех рас редок и будет проанализирован отдельно.
Рассмотрим ключевые признаки по соотношению a- и в-форм.Дельтовый индекс. Вариант a значительно преобладает.
Однако по пропорции вариантов расы отчетливо различаются. Так,
143
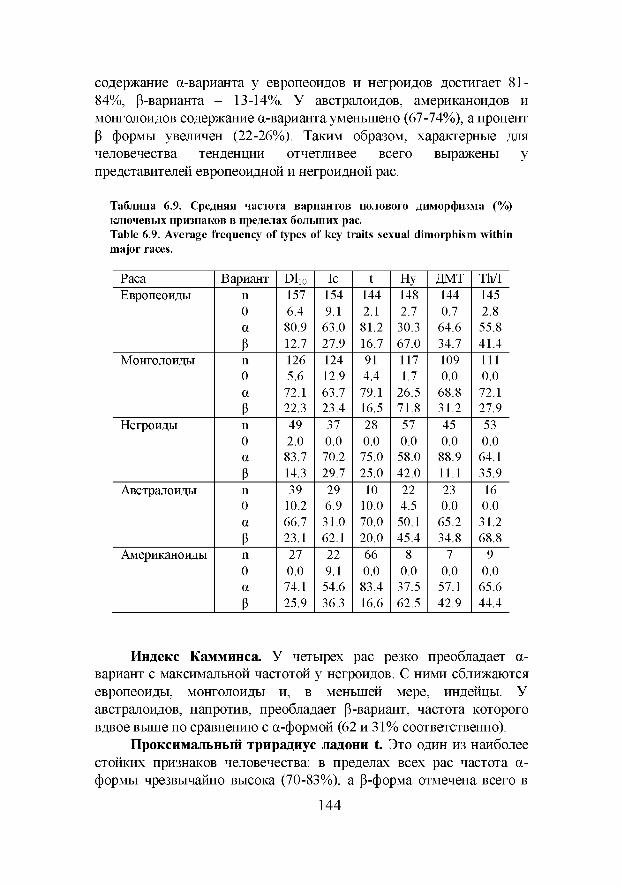
содержание а-варианта у европеоидов и негроидов достигает 81 - 84%, в-варианта - 13-14%. У австралоидов, американоидов и монголоидов содержание а-варианта уменьшено (67-74%), а процент в формы увеличен (22-26%). Таким образом, характерные для человечества тенденции отчетливее всего выражены упредставителей европеоидной и негроидной рас.
Таблица 6.9. Средняя частота вариантов полового диморфизма (%) ключевых признаков в пределах больших рас.Table 6.9. Average frequency of types of key traits sexual dimorphism within major races.
Раса Вариант Dl10 Ic t Hy ДМТ Th/IЕвропеоиды n 157 154 144 148 144 145
0 6.4 9.1 2.1 2.7 0.7 2.8а 80.9 63.0 81.2 30.3 64.6 55.8в 12.7 27.9 16.7 67.0 34.7 41.4
Монголоиды n 126 124 91 117 109 1110 5.6 12.9 4.4 1.7 0.0 0.0а 72.1 63.7 79.1 26.5 68.8 72.1в 22.3 23.4 16.5 71.8 31.2 27.9
Негроиды n 49 37 28 57 45 530 2.0 0.0 0.0 0.0 0.0 0.0а 83.7 70.2 75.0 58.0 88.9 64.1в 14.3 29.7 25.0 42.0 11.1 35.9
Австралоиды n 39 29 10 22 23 160 10.2 6.9 10.0 4.5 0.0 0.0а 66.7 31.0 70.0 50.1 65.2 31.2в 23.1 62.1 20.0 45.4 34.8 68.8
Американоиды n 27 22 66 8 7 90 0.0 9.1 0.0 0.0 0.0 0.0а 74.1 54.6 83.4 37.5 57.1 65.6в 25.9 36.3 16.6 62.5 42.9 44.4
Индекс Камминса. У четырех рас резко преобладает а- вариант с максимальной частотой у негроидов. С ними сближаются европеоиды, монголоиды и, в меньшей мере, индейцы. У австралоидов, напротив, преобладает в-вариант, частота которого вдвое выше по сравнению с а-формой (62 и 31% соответственно).
Проксимальный трирадиус ладони t. Это один из наиболее стойких признаков человечества: в пределах всех рас частота а- формы чрезвычайно высока (70-83%), а в-форма отмечена всего в
144

16-20%. По комбинации частот обоих вариантов взаимно сходны австралоиды и негроиды, с одной стороны, и все остальные расы - с другой. В последнем случае характерные для человечества тенденции выражены в максимальной форме, т.е. высокая частота a формы сочетается с очень низкой частотой в-варианта.
Dl,o Ic t Ну ДМ Т Th/I Dlio Ic * Ну ДМ Т Tli/I
Рисунок 6.6. Индивидуальные профили больших рас по частоте вариантов полового диморфизма ключевых признаков.Figure 6.6. Profiles of major races for frequencies of sexual differences variants.
Узорность гипотенара. Этот крайне специфический признак по соотношению вариантов полового диморфизма разделяет человечество на две группы. У австралоидов и, особенно у негроидов, преобладает a-форма (50-58%), все остальные расы объединяются отчетливым перевесом в-варианта (60-72%).
Добавочные межпальцевые трирадиусы. У всех рас преобладает a-вариант, особенно резко - у негроидов (около 90%). Соотношение вариантов у американоидов наиболее сбалансировано, разница в частотах составляет около 14%. Остальные расы взаимно сходны, разрыв между вариантами составляет 30-40%.
Узорность тенара. В пределах четырех рас (за исключением австралоидов) отчетливо преобладает a -форма (56-72%) над в- вариантом (28-44%). Наиболее сбалансировано соотношение вариантов у европеоидов (56 и 41% соответственно). На этом фоне
145

резко выделяются австралоиды, у которых р-форма встречается вдвое чаще по сравнению с а формой (69% и 31% соответственно).
Обратимся к анализу нулевого варианта полового диморфизма (см. Рис. 6.6 и 6.7).
Среднерасовая частота этого самого редкого типа варьирует от 0 до 13%. Минимальный лимит этой амплитуды характерен для всех ключевых признаков, т.е. по любому из них нельзя обнаружить расу, у которой выборочные значения признаков у мужчин и женщин совпадают.
Рисунок 6.7. Соотношение частот вариантов полового диморфизма ключевых признаков в пределах больших рас.Figure 6.7. Relationships among frequencies of sexual dimorphism variants within major races.
Дельтовый индекс. Среднерасовая частота 0-варианта колеблется от 0% у американоидов до 10% у австралоидов, негроиды сближаются с индейцами (2%), а европеоиды - с монголоидами (около 6%).
Индекс Камминса. Средняя частота варианта колеблется по расам от 0% у негроидов до 13% у монголоидов. Остальные три расы сближаются с монголоидами (7 -9%).
Проксимальный трирадиус ладони. Отсутствие варианта характеризует негроидов и американоидов, максимум - австралоидов (10%). У европеоидов и монголоидов среднерасовая частота признака мала (2-4%).
146

Узорность гипотенара. Нулевой вариант отсутствует у негроидов и американоидов, чаще всего встречается у австралоидов (4.5%). У монголоидов и европеоидов он отмечен в 2-3%.
Добавочные межпальцевые трирадиусы. Признак принадлежит к числу максимально дифференцирующих оба пола. Содержание нулевого варианта колеблется всего лишь от 0 до 0.7%. Последняя цифра характеризует европеоидов, у остальных рас данный вариант отсутствует.
Узорность тенара. Содержание нулевого варианта здесь распределяется аналогичным по отношению к ДМТ образом. Он отмечен в 2.8% у европеоидов и отсутствует у остальных рас.
Таким образом, наблюдаются отчетливые направленные различия между ключевыми признаками по содержанию нулевого варианта полового диморфизма. По трем из них (Dl10, Ic, t) в пределах всех рас обнаруживаются наиболее частые совпадения средних величин в мужских и женских выборках. По трем остальным (Ну, ДМТ, Th/I) частота 0-варианта приближается к минимуму. Иными словами различия между полами внутри рас по трем последним признакам встречаются наиболее часто.
Обобщая итоги сравнительного анализа частот нулевого варианта по разным признакам, отметим сетеобразное сходство рас. Однако при более детальном подходе выявляется ряд важных моментов. Так, своеобразие негроидной расы заключается в отсутствии случаев совпадения значений пяти из шести ключевых признаков в параллельных мужских и женских выборках. Лишь по дельтовому индексу наблюдается сходство полов, причем только у 2% выборок. Аналогичное свойство обнаруживают американоиды, у которых нулевой вариант отмечен лишь в отношении индекса Камминса, правда, почти с максимальной частотой (9%). По остальным ключевым признакам американоиды, как и негроиды, наиболее диморфны. Европеоиды характеризуются стойким наличием нулевой формы полового диморфизма признаков, по которым у остальных рас эта форма отсутствует (ДМТ и Th/I), то есть максимальной частотой мономорфного варианта. Монголоиды и австралоиды сближаются с европеоидами по большинству ключевых признаков.
При сравнении рас по частоте разных вариантов полового диморфизма выявлены следующие закономерности:
147

• По ключевым признакам у всех рас преобладает, как правило, а-вариант. На внутрирасовом уровне у женских выборок уменьшены величины всех основных признаков по сравнению с мужскими. Исключения из этого правила зафиксированы у монголоидов, европеоидов и американоидов в отношении узорности Ну. Кроме того, у австралоидов по Ic и узорности Th/I преобладает Р-вариант.
• Негроидная раса выделяется максимальной частотой а- варианта по четырем признакам. Ее антиподом являются австралоиды, у которых частота этого варианта минимальна также по четырем признакам.
• При сравнении рас выявляется обособленное положение негроидов и американоидов, у которых резко уменьшена частота нулевого варианта и увеличена дифференциация полов.
Субрасовый уровень. Рассмотрим соотношение а- и Р- вариантов в рамках подразделений европеоидной и монголоидной рас. В Таблице 6.10 использованы обобщенные показатели региональных подразделений каждой расы.
Дельтовый индекс. Всюду преобладает а-вариант (75-86% против 12-21% P-варианта). На Кавказе, в Европе и в Передней Азии разрыв между частотами обоих вариантов наиболее велик: «мужской» тип полового диморфизма выражен максимально четко (81-86%), «женский» - сокращен до минимума (12-14%). Европеоиды Средней Азии практически неотличимы от северных и южных монголоидов, у которых содержание P-варианта несколько увеличено (20-22%) при одновременном уменьшении частоты а- варианта (75%).
Индекс Камминса. При общем преобладании а- над Р-формой (44-66% против 22-30%) подразделения монголоидов заметно отличаются друг от друга. В южной группе соотношение вариантов (44 и 30%) более сбалансировано, чем в Северной (22 и 66%). Сибирские монголоиды сближаются с европеоидами, для которых характерно резкое преобладание а-частот (62-72%). Наиболее отчетливо этот эффект выражен на Кавказе (72 против 21%), последовательно ослабевая у населения Средней и Передней Азии и достигая минимума в Европе.
Проксимальный трирадиус ладони t. Всюду преобладает а- вариант (до 90%). Характерны различия между южными и северными монголоидами. У первых сильно преобладает а-форма
148

над в-вариантом (90 и 10% соответственно). У вторых пропорция сдвинута за счет увеличения в- частот (78 и 19%). Европеоиды разных регионов сходны по пропорции частот вариантов a (84-88%) и в (21-25%). На этом фоне выделяются кавказцы, благодаря менее значительному преобладанию a- формы над в- вариантом (66 и 31%).
Таблица 6.10. Содержание вариантов полового диморфизма ключевых признаков в подразделениях европеоидной и монголоидной рас.Table 6.10. Share of variants of sexual differences of key traits within Caucasoid and Mongoloid races.
Регион Вариант Dl10 Ic t Hy ДМТ Th/IЕВРОПЕОИДЫ
Европа n 161 157 145 150 127 1510 6.8 10.8 1.4 4.0 0.0 4.0a 81.4 61.6 84.2 38.7 73.2 60.9в 11.8 27.4 14.4 57.3 26.8 35.1
Кавказ n 93 95 85 91 74 910 3.2 7.4 3.5 1.1 0.0 1.1a 85.0 71.6 65.9 30.8 64.9 42.9в 11.8 21.0 30.6 68.1 35.1 56.0
Передняя Азия n 22 20 14 21 15 200 0.0 10.0 0.0 4.8 6.7 0.0a 86.4 65.0 85.7 19.0 53.3 45.0в 13.6 25.0 14.3 76.2 40.0 55.0
Средняя Азия n 85 85 82 85 80 830 4.7 8.2 1.2 2.4 1.2 1.2a 77.7 69.4 87.8 16.4 68.8 57.8в 17.6 22.4 11.0 81.2 30.0 41.0
МОНГОЛОИДЫСеверная Азия n 139 139 135 139 135 136
0 4.3 11.5 3.7 4.3 0.7 0.0a 74.8 66.2 77.8 24.5 70.4 64.7в 20.9 22.3 18.5 71.2 28.9 35.3
Южная Азия n 65 59 21 53 41 300 3.1 25.4 0.0 0.0 0.0 0.0a 75.4 44.1 90.5 22.6 87.8 76.7в 21.5 30.5 9.5 77.4 12.2 23.3
Узорность гипотенара. В подразделениях рас, так же, как и по среднерасовым показателям, частота a -формы резко преобладает
149

над P-вариантом. В максимальной степени это выражено у среднеазиатских и переднеазиатских европеоидов (81 и 76% соответственно). В то же время на Кавказе, и в особенности, в Европе это преобладание менее заметно. Южные и северные монголоиды сходны друг с другом по частоте вариантов, но в еще большей степени сближаются с переднеазиатскими и кавказскими европеоидами соответственно.
Добавочные межпальцевые трирадиусы. Всюду преобладает а-вариант, особенно резко у южноазитских монголоидов (88%). Компактную группу образуют европеоиды Европы, Средней Азии и Кавказа (65-73%), в скоплении которых также располагаются северные монголоиды. У европеоидов Передней Азии частоты а- и P-форм почти сбалансированы (53 и 40%), что выделяет их среди всех регионов.
Узорность тенара. У всех рас, кроме австралоидов, преобладает а-вариант (56-72%) над Р-формой (28-44%). Наиболее сбалансированы пропорции а- и P-форм у европеоидов (41 и 56% соответственно). На этом фоне резко выделяются австралоиды (69 и 31%).
Отсутствие линии С (С0). Всюду преобладает Р-форма, причем монголоиды и европеоиды отчетливо различаются. У монголоидов соотношение а- и P-вариантов более сбалансировано по сравнению с европеоидами (52-59%). Европеоиды разных регионов взаимно сходны (а=22-28; Р=69-75%), отличаясь перевесом частот Р- формы.
Редукция линии С (Сх). Как и в случае с Th/I, подразделения европеоидов различаются. В среднеазиатских и европейских группах преобладает Р-вариант (50-56%), в переднеазиатских и кавказских - а (53-59%). Северные и южные монголоиды необычайно сходны между собой (а=58%, Р=39-40%).
Отсутствие половых различий (нулевой вариант). У европеоидов равные значения признаков в группах мужчин и женщин наиболее часто отмечаются по обоим индексам, (особенно по Ic - до 11%). По остальным признакам происходит более или менее упорядоченное уменьшение содержание 0-варианта, особенно отчетливое на Кавказе. При этом на Кавказе процент нулевого варианта ниже, чем в других регионах, по всем признакам, кроме осевого трирадиуса ладони t. Различия между остальными подразделениями бессистемны, но стоит отметить, что среднеазиатские европеоиды в отношении подавляющего
150

большинства признаков занимают промежуточное положение между европейцами и кавказцами. Подразделения монголоидов хорошо вписываются в общую картину. При этом к мировой схеме соотношения вариантов по комплексу признаков ближе всего находятся южные монголоиды (если не считать вариантов линии С). В южномонголоидной группе случаи совпадающих или крайне близких значений по Ic у мужчин и женщин наиболее часты, гораздо более редки по DlJ0 и сведены к минимуму по остальным ключевым признакам. Северные монголоиды оказываются более близкими к европейской группе, чем к южным группам. В заключение раздела отметим, что для подразделений европеоидной и монголоидной рас характерны те же закономерности распределения признаков по полу, что и для соответствующих обобщенных расовых групп. Вместе с тем уточнились детали дифференциации полов в пределах расовых вариантов. Обнаружилось разграничение северных и южных вариантов европеоидной большой расы, в некоторых случаях отличающееся неожиданно резким характером. Так, на Кавказе и в Передней Азии выявлено преобладание в-варианта по узорности Th/I, в то время как для остальных европеоидов, а также для всех больших рас (за исключением австралоидов) характерна а-форма. Кроме того, в суммарной европеоидной выборке частоты а- и в- вариантов Сх одинаковы, а в подразделениях ее выявлены две группы с противоположным соотношением форм. У большинства южных европеоидов (Кавказ, Передняя Азия), так же, как и у монголоидов, негроидов и американоидов, преобладает а-вариант. В то же время в Европе и Средней Азии обнаружена беспрецедентная форма обратного соотношения вариантов (в- больше а-).
Что касается северных и южных монголоидов, то по большинству признаков они отчетливо различаются, одновременно сохраняя особенности, свойственные монголоидам в целом. Сибирские монголоиды часто обнаруживают сходство с европеоидами Европы по пропорции вариантов полового диморфизма.
Кроме того, на субрасовом уровне у европеоидов и монголоидов по частоте отсутствия полового диморфизма прослеживаются те же закономерности, что и на уровне больших рас. Случаи равного значения признаков в группах мужчин и женщин наиболее часты в отношении Ic и Dl10 .
151

Кратко обобщим итоги исследования проблемы полового диморфизма признаков дерматоглифики у человека. Был использован материал, охватывающий все большие расы человечества (716 параллельно исследованных мужских и женских выборок) и применены различные независимые методы анализа.
Характеристика больших рас в аспекте полового диморфизма.
• Рангово-дисперсионным анализом по Фридману среднерасовых показателей абсолютных и относительных различий между полами выявлена принадлежность всех больших рас человечества к единой совокупности. Таким образом, по степени дифференциации полов человечество как вид гомогенно.
• В пределах всех рас среднерасовые абсолютные различия между полами, как правило, значимы на разных таксономических уровнях. Вместе с тем, необходимо отметить нестойкость величины АПР: коэффициенты межгрупповой вариации АПР ключевых признаков очень высоки (54-106%) и еще выше - в группе неключевых признаков.
• Существует определенная иерархия признаков по величине АПР: оба пола менее всего различаются по величине обоих индексов (Ic и DlJ0). Далее, в порядке возрастания АПР, следуют узорность Ну, ДМТ, осевой проксимальный трирадиус ладони, узорность Th/I.
• Общечеловеческие, т.е. видовые, тенденции полового диморфизма наиболее отчетливо проявляются при анализе относительных различий между полами (по коэффициенту полового диморфизма, или КПД). Подавляющее большинство ключевых и связанных с ними неключевых признаков кожного рельефа характеризуются преобладанием величины признака у мужчин. Внешне это явление имеет аллометрический характер, однако причинные связи между размерами тела, с одной стороны, и частотой/типом узора, с другой, отсутствуют, и таким образом, природа обнаруженного явления остается неясной. В общей картине преобладания величин признаков у мужчин выявляется отчетливая биполярность человечества в отношении двух ключевых признаков - узорности Hy и Th/I. Так, у тропических рас (негроиды и австралоиды) частота узоров на Ну у мужчин выше, чем у женщин, в то время как у остальных рас наблюдается обратное соотношение. Что касается узорности Th/I, то у всех больших рас она выше у
152

мужчин, за исключением австралоидной, где признак чаще отмечен у женщин.
• С учетом специфики Hy и Th/I человеческие расы отчетливо различаются по средней величине КПД. Максимальная диморфность свойственна негроидной расе, а наиболее гомогенны австралоиды, к которым тяготеют американоиды и монголоиды. Европеоидной расе присуща умеренная дифференциация полов. Тропический ствол, включающий обе экваториальные расы по соматологической классификации, не может быть охарактеризован по признакам кожного рельефа как генетически единый, так как составляющие его расы - негроиды и австралоиды - полярны по степени полового диморфизма.
Половой диморфизм на уровне субрасовых подразделений.
Были рассмотрены разные таксономические уровни по материалам монголоидной и европеоидной рас. Этнический уровень в обоих случаях отмечен минимальными различиями между полами по абсолютным величинам признаков. Абсолютные половые различия на разных таксономических уровнях у европеоидов неодинаковы в отношении почти всех ключевых признаков. У монголоидов статистически значимые различия между уровнями АПР не обнаружены. В то же время систематические различия между таксономическими уровнями по относительному показателю полового диморфизма (КПД) у обеих рас отсутствуют. Последнее свидетельствует о предпочтительности использования КПД по сравнению с АПР.
Методическое значение проведенного исследования в связи с практическим приложением выводов.
• Приведенные характеристики вариационных рядов внутрирасового распределения выборочных значений абсолютных и относительных различий между полами позволяют статистически оценить степень полового диморфизма полного набора ключевых признаков в любой группе. Применительно к европеоидам и монголоидам такая оценка может быть произведена на любом из трех уровней (селения, территориальной группы, этноса). Кроме того, предлагаемая рубрикация величин КПД признаков позволяет оценить половые различия по форме (типы а-, в- и нулевой) и
153

количественно (большая, средняя, малая степени) в пределах каждой формы, с учетом специфики некоторых рас в отношении отдельных признаков.
• На внутрирасовом уровне величины признаков одновременно исследованных мужских и женских групп не обнаруживают стойкого параллелизма. Однако изменчивость ключевых признаков и их составляющих, как правило, у обоих полов более тесно согласована по сравнению с другими признаками. Степень взаимосвязи величин признаков мужских и женских выборок неодинакова у разных рас и максимально низка у европеоидов. Эти наблюдения свидетельствуют о некорректности использования суммарных данных обоих полов в антропологических исследованиях; на что неоднократно указывалось в ряде публикаций, в том числе в основополагающей монографии Г. Камминса и Ч. Мидло (1943).
В заключение рассмотрим соотношение картин дифференциации мужских и женских групп по комплексу ключевых признаков. Различные способы статистического анализа позволяют выделять на одном и том же материале кластеры, частично совпадающие по составу. Попытка сопоставления результатов дифференциации полов на разных таксономических уровнях по комплексу ключевых признаков привела к следующим результатам.
Территориальные группы мужчин и женщин в пределах одиннадцати этносов Северной Евразии дифференцированы абсолютно несходным образом: матрицы их ОДР дают малые и незначимые показатели связи (Табл. 6.11). Лишь у удмуртов наблюдается явная тенденция к сходству (r=0.5). Это может быть связано с их метисным монголоидно-европеоидным происхождением и соответственно биполярностью формообразующих процессов, которая проявляется у обоих полов и не перекрывается выборочным эффектом.
На уровне этносов в пределах историко-географических регионов обнаруживается согласованная дифференциация полов (r колеблется от 0.3 до 0.7). При этом европеоидные по типу этносы южных регионов (Кавказа и Средней Азии) показывают умеренную степень связи (r=0.4), монголоиды Сибири - минимальную (r=0.3), а в Восточной Европе - максимальную согласованность (r=0.7). Следовательно, при относительной расовой однородности населения оба пола на этническом уровне в пределах регионов скоррелированы
154

друг с другом в общей картине варьирования слабо, хотя и значимо. Там же, где можно предполагать наличие двух заведомо разных расовых компонентов (этносы Волго-Камья с весомой долей монголоидности, с одной стороны, и этносы остальной части восточноевропейского региона, где абсолютно преобладает европеоидный компонент, с другой), параллелизм изменчивости мужчин и женщин повышается почти вдвое. Вероятно, по той же причине резко увеличен показатель корреляции мужской и женской матриц ОДР всех локальных рас Северной Евразии (до 0.8). Он достигает максимума при переходе на уровень больших рас, где расовая разнородность наиболее велика и проявляется параллельно по данным обоих полов (r=0.9).
Таблица 6.11. Степень соответствия дифференциации одноименных мужских и женских выборок по комплексу признаков (корреляция матриц ОДР)Table 6.11. Correlation of dermatoglyphic traits complex in male and female samples (correlations of GDD’s matrixes).
Группа Число выборок rВнутриэтнический уровень
Русские 19 0.12Карелы 12 0.12Таджики 7 -0.01Туркмены 21 -0.03Удмурты 8 0.48*Казахи 23 0.18Татары Сибири 10 0.27Калмыки 10 0.03Тувинцы 6 0.17Эвенки 5 0.23Чукчи 6 0.17
Этнический уровень (в регионах)Европейская часть России 25 0.65*Кавказ 40 0.43*Средняя Азия 11 0.44*Сибирь 40 0.27*
Уровень локальных расСеверная Евразия 20 0.81*
Уровень больших расБольшие расы ойкумены 5 0.92*
* отмечены значимые величины коэффициента корреляции (r) от p<0.05155

Таким образом, согласованность дифференциации мужских и женских выборок на разных таксономических уровнях неодинакова. В пределах этноса, среди локальных групп, как правило, действует выборочный эффект, затушевывая фактор расы и приводя в итоге к несходной картине варьирования обоих полов. Начиная с этнического уровня, явившегося мощным фактором вызванной в основном историческими причинами дивергенции населения в регионах, мы наблюдаем отчетливое сходство в итоговой картине дифференциации мужских и женских групп. Особого развития это совпадение достигает там, где снят этнический барьер и мат ериал организован по принципу принадлежности к расе (локальной или большой). В последнем случае степень совпадения близка к максимальной. Иначе говоря, там, где проявляется в основном фактор расы (биологический по существу), оба пола дают пример согласованной изменчивости в рассматриваемом таксономическом поле, хотя и в этом случае она не является исчерпывающе полной.
156

%

Г лава 7. Палеодерматоглифика
В 1981 году греческим специалистом К. Бартсокасом для обозначения дерматоглифических исследований, объектом которых являются археологические (керамика, пергамент, фрагменты предметов органического происхождения) и антропологические (мумии) материалы, был предложен термин «палеодерматоглифика» (Bartsocas, 1982).
Термин получил признание среди исследователей ряда европейских стран, и на протяжении последних десятилетий изучение отпечатков пальцев на археологическом материале неоднократно становилось темой отдельных работ, в том числе экспериментального направления (Cummins, 1941; Kamp et al. 1999; Сыроватко, Елистратов, 2002; Branigan et al., 2002; Панкратова, 2008; и др.). В начале XXI века с целью согласованного изучения отпечатков пальцев на фрагментах древней керамики и создания общей базы данных, группой, объединившей европейских исследователей из Чехии, Швеции и Великобритании, было организовано научное общество «Ancient Fingerprint Society» (Astrom, 2007, p.2). Под эгидой той же группы в 2007 году выходит первый (и к настоящему времени единственный) номер журнала «Journal of ancient fingerprints». Основной задачей исследований является определение при помощи дактилоскопических методик пола и возраста древних гончаров (объектом изучения, как правило, является керамика), минимальной численности лиц, оставивших отпечатки на одном объекте, а также установление принадлежности отпечатков на разных объектах (сосудах) одному и тому же человеку (ремесленнику).
Однако в отличие от дерматоглифистов и дактилоскопистов, археолог имеет дело почти исключительно с неполными отпечатками пальцев и ладоней древних людей. У него отсутствует возможность верифицируемого соотнесения фрагментов узоров с конкретными пальцами, а также отпечатками человека определенного пола и возраста в разные исторические эпохи. Эти обстоятельства в значительной степени сужают тот круг методик, который может быть применен для изучения археологических материалов. Определение пола и возраста человека по отпечаткам в криминалистических исследованиях опирается на комплекс
158

признаков (не только графических, но и химических), а также на дополнительную контекстную информацию, недоступную при работе с археологическими объектами.
Как правило, для вероятностного установления пола и возраста человека по отпечаткам на керамических изделиях в археологии используются показатели, связанные с плотностью папиллярных линий на определенном участке пальцевых подушечек. Применение этих признаков основывается на том статистическом факте, что для отпечатков пальцев женщин характерны более тонкие и плотно расположенные папиллярные линии, чем для мужчин, а для детей более тонкие гребешки, чем для врослых.
Одним из таких показателей является плотность папиллярных линий, рассчитываемая на единицу площади узора. Традиционно рассчитывается количество папиллярных линий, располагающихся на отрезке площадью 5х5 мм. В качестве альтернативного показателя используется средняя ширина папиллярной линии, рассчитываемая как расстояние от центра/начала одного гребешка до центра/начала соседнего.
В отечественной криминалистической литературе наиболее широкую известность получили следующие расчеты. У детей в возрасте 8-12 лет в среднем на отрезке узора длиной 5 мм располагается 12-13 папиллярных линий, у подростков 13-17 лет - 10-12 линий, у взрослых людей - 9-10 линий. Межполовые различия не учитываются. Расчеты эти носят приблизительный характер и могут в значительной степени варьировать в зависимости от конституциональных особенностей конкретного человека. У очень полных людей может встречаться 6-7 линий / 5 мм, у худощавых - 10-11 линий / 5 мм. (Титов, 2002, с. 11; Моисеева, 2000, с. 28, 154). Именно эта схема легла в основу статей отечественных археологов, занятых изучением возможностей определения пола и возраста древних гончаров по отпечаткам на керамических сосудах (Сыроватко, Елистратов, 2002; Гей, 2007).
Однако некоторыми исследователями отмечается ненадежность приведенных данных. Во-первых, связи между плотностью папиллярных линий и возрастом человека были установлены на небольшом статистическом материале. Во-вторых, в них совершенно не учитываются различия в плотности линий, фиксируемые на разных участках даже одного узора и одного индивида (Хазиев, 1984). Кроме того, в действительности исследованиями установлены значимые различия в плотности
159

узоров между мужскими и женскими выборками. Плотность линий возрастает от базиллярной (проксимальной) области через центральную к дистальной части узора. У женщин она варьирует в среднем от 9.5 до 12.7 линий / 5 мм, у мужчин - от 8 до 10.7 линий / 5 мм (Божченко, 2004, с.44).
Кроме того, специальные исследования показали, что определенные различия в плотности линий фиксируются между ульнарной и радиальной сторонами пальцев, между типами узоров, между разными пальцами, между левой и правой руками, между пальцевыми и ладонными узорами (Cummins et al., 1941; Ohler, Cummins, 1942; Gutierrez-Redomero et al., 2008). Так, еще в начале 1940-х гг. Г. Камминс с коллегами установили плотность папиллярных линий для разных участков ладоней и пальцев и пришли к выводу о наличии значимых различий между ними. Материалами послужили отпечатки 200 студентов в возрасте от 18 до 30 лет, а также 100 студенток в возрасте от 16 до 28 лет. Для всех участков узоров у женщин в целом фиксировался более высокий показатель плотности узоров. Плотность узоров на пальцах рук, как у мужчин, так и у женщин, возрастает по формуле: I<II<III<V<IV. Плотность папиллярных линий на ладони ниже, чем на пальцах, а на пальцах правой руки ниже, чем на пальцах левой (Cummins et al., 1941; Ohler, Cummins, 1942). Кроме того, недавно испанскими коллегами было показано, что количество папиллярных линий на единицу площади отдельно взятого узора как в мужских, так и в женских выборках, возрастает в порядке: проксимальная сторона - ульнарная сторона - радиальная сторона. Причем для обоих полов размах средних значений для разных областей пальцевых узоров составляет около 4.5 папиллярных линий / 25 мм2 (Gutierrez- Redomero et al., 2008).
Результаты исследований однозначно свидетельствуют о том, что приведенные выше данные для установления возраста не могут быть использованы в качестве надежного источника информации не только при работе с археологическими материалами (представленными фрагментарными отпечатками папиллярных узоров, точную локализацию которых далеко не всегда возможно установить), но и в современной дактилоскопии.
Зарубежные исследователи, занимающиесяпалеодерматоглифическими исследованиями, часто используют другой показатель плотности линий - среднюю ширину папиллярных линий (в дальнейшем MRB - mean ridge breadth). При
160

этом в работах делается акцент на собственные экспериментальные данные. Измерения проводятся с помощью калипера с точностью до 0.01 мм. Как указывалось выше, измерение MRB может проводиться двумя способами - от центра одного гребня до центра соседнего, либо от начала одного гребня до начала соседнего. Рассмотрим результаты исследований группы археологов, которые применили второй способ как более удобный при работе с рельефными отпечатками. Для расчета величин признака измерялось общее расстояние от центра узора к его периферии для 10 гребешков (от начала первого до начала одиннадцатого), а затем полученная величина делилась на 10 (Kamp et al. 1999). Анализировались отпечатки на глиняных фигурках животных, а также на керамических сосудах, изготовленных одной из групп американских индейцев пуэбло. Средняя величина MRB для отпечатков на фигурках животных составила 0.37 мм. При этом MRB 50% отпечатков варьирует от 0.33 до 0.41 мм. Чтобы рассчитать примерную плотность папиллярных линий на единицу площади в принятых масштабах, разделим 5 мм на полученные значения MRB и получим 13-14 папиллярных линий с размахом, охватывающим половину проанализированных отпечатков, от 12 до 15 линий. Для сосудов средняя величина MRB составила 0.49 мм, что соответствует плотности папиллярных линий 10 гребешков / 5 мм (с соответствующим размахом изменчивости от 0.44 до 0.53, т.е. от 9 до 11 гребешков / 5 мм). С учетом усадки керамики при обжиге (достигающей 7.5-10% объема) реконструированная средняя плотность составила 12 линий / 5 мм для фигурок животных и 9 линий / 5 мм для керамических сосудов. Полученные данные позволили авторам работы утверждать, что фигурки животных изготавливались в основном детьми 11 -13 лет, а сосуды - взрослыми (Kamp et al. 1999, p. 314).
Несколько дальше в исследовании продвинулись чешские исследователи М. Кралик и В. Новотный (Kralik, Novotny, 2003). Последние протестировали различные формулы, в том числе предложенные Кэмп с соавторами, для установления возраста людей, оставивших отпечатки, а также межполовые различия по MRB, опираясь на собственные эмпирические данные. Материалами послужили две небольшие выборки: 56 школьников и 50 взрослых (23 мужчины и 27 женщин). В результате был построен график, показавший четкую зависимость между MRB и возрастом человека и согласующийся с данными К. Кэмп. Судя по графику при MRB от
161

0.4 до 0.5, невозможно различить отпечатки женщин, мужчин и подростков. Различия по полу до 20 лет не проявляются. MRB менее 0.39 мм (более 12.5 линий / 5 мм) встречались только у детей и подростков примерно 15 лет. MRB выше 0.52 мм (менее 9.5 линий /5 мм) были отмечены только у взрослых мужчин. При этом средняя величина MRB у мужчин составила 0.494 мм (около 10 линий / 5 мм), у женщин - 0.450 мм (11 линий / 5 мм) (Kralik, Novotny, 2003, p.22-23). Недостатком исследования является небольшая численность проанализированных отпечатков, и все же авторам удалось подтвердить наличие значимых корреляций между возрастом и толщиной гребешков, а также выделить крайние значения признака, которые с высокой долей вероятности позволяют устанавливать половозрастные характеристики людей по отпечаткам.
Однако действительно ли возможно определить возраст и пол человека, если придерживаться четкой методической системы и опираться на представительные выборки по различным половозрастным группам современного населения, отбросив пограничные значения признаков? Ответ на этот вопрос позволяет дать сравнительный анализ результатов работ, посвященных половым различиям в плотности папиллярных линий и проведенных независимыми группами исследователей из разных стран.
Как уже упоминалось выше, специалистами было показано наличие значимых различий по плотности папиллярных линий между мужскими и женскими выборками. Для женщин характерны более тонкие и более плотно расположенные папиллярные линии, чем для мужчин. Неоднократно предпринимавшиеся попытки определить плотность папиллярных линий, при которой половая принадлежность человека могла бы быть определена с высокой степенью вероятности, привели к неоднозначным итогам. Результаты некоторых исследований приведены в Таблице 7.1.
Представленные данные показывают, что плотность папиллярных линий как дифференцирующий по полу признак обладает высокой межгрупповой изменчивостью. Размах среднепопуляционных значений у женщин составляет 6.4 линий / 25 мм2 (от 12.6 у афроамериканок до 19.0 у аргентинок), у мужчин - 6.1 линии / 25 мм2 (от 10.9 у афроамериканцев до 17.0 у аргентинцев). Коэффициент межгрупповой вариации плотности папиллярных линий составляет 14.9% у женщин и 18.7% у мужчин.
162

По результатам исследований выделяются, по меньшей мере, три варианта пограничных значений плотности папиллярных линий, использование которых, казалось бы, позволяет исследователям определять с высокой степенью вероятности пол человека.
Первый вариант фиксируется на индийских, американских, малайзийских и китайских материалах. Пограничное значение плотности, при котором пол человека не может быть определен, составляет здесь примерно 12 линий / 25 мм2 (Nayak et al., 2010a; 2010b; Acree, 1999). Второй вариант фиксируется на южноиндийских материалах и соответствует плотности 13 -14 линий / 25 мм2 (Nithin et al., 2011; Gungadin, 2007). Наконец, третий вариант обнаружен в выборках испанских исследователей. Средние значения плотности папиллярных линий здесь столь высоки, что почти позволяют усомниться в достоверности результатов проведенного исследования - 16-17 линий / 25 мм2 для Испании и17-18 линий / 25 мм2 для Аргентины (Gutierrez-Redomero et al., 2008; 2013).
Таблица 7.1. Средние значения плотности папиллярных линий на участке 5х5 мм в мужских и женских выборках некоторых стран (по материалам взрослого населения)Table 7.1. Average values of fingerprint ridge density within a 5x5 mm square in male and female samples from some countries (data for adults).
Авторы Регионисследований
Женщины(чел.)
Мужчины(чел.)
Acree, 1999США (негроиды)
США (европеоиды)12.6 (100) 13.3 (100)
10.9 (100) 11.1 (100)
Gungadin, 2007Южная Индия
(Карнатака)14.6 (250) 12.8 (250)
Gutierrez-Redomero et al., 2008
Испания 17.9 (100) 16.2 (100)
Nayak et al., 2010a; Nayak et al., 2010b
ИндияКитай
Малайзия
14.2 (100)14.2 (100) 13.6 (50)
11.0 (100) 11.7 (100) 11.4 (50)
Nithin et al., 2011 Южная Индия 14.1 (275) 12.6 (275)
Agnihotri et al. 2012Маврикий
(индо-маврикийцы)14.0 (100) 11.4 (100)
Gutierrez-Redomero et al., 2013 Аргентина
19.0 (100) 18.5 (93)
17.0 (100) 16.7 (100)
163

Следует подчеркнуть, что во всех указанных случаях в качестве материалов исследования использовались отпечатки взрослых людей. Плотность папиллярных линий определялась для участка 5х5 мм, определенном для радиальной области в дистальном направлении от центра узора. Для расчетов использовалась средняя плотность линий для всех 10 пальцев. Детали методики четко оговариваются в специальном разделе каждой из указанных в таблице статей.
Маловероятно, что в сложившейся картине значимую роль сыграли различия в методических подходах. Некоторое влияние на результаты анализа, по-видимому, оказали конституциональные особенности и частоты типов пальцевых узоров обследованных групп. Однако наиболее важной причиной расхождений являются популяционные различия выборок по анализируемому признаку.
Несложно заметить, что размах средних значений как в мужской, так и в женской выборке в территориальном отношении перекрывает изменчивость средней плотности папиллярных линий для различных возрастных групп по данным как отечественных криминалистов, так и чешских коллег (Рис. 7.1).
Это утверждение справедливо, даже если учитывать некорректность прямого сопоставления цифр в работах, в которых в качестве основного показателя используется MRB, плотность папиллярных линий на отрезке 5 мм и плотность папиллярных линий на участке 5х5 мм. Различия могут быть устранены при помощи простых арифметических действий с небольшой погрешностью в результатах.
Работающий с археологическими материалами исследователь не может восстановить полную дерматоглифическую характеристику изучаемых по отпечаткам индивидов. Более того, он не обладает достоверными знаниями о средней для конкретной древней популяции плотности папиллярных узоров у взрослых мужчин и женщин. Археолог не может делать обоснованные выводы о поле и возрасте древних гончаров, опираясь на анализ отпечатков пальцев в современных группах. Когда исследователи уверяют, будто, опираясь на результаты анализа отпечатков, им удалось показать, что определенные керамические изделия на памятнике изготавливались подростками, иногда указывая при этом их точный возраст до года (см. например: Moran, 2007, p. 18), упрофессиональных дерматоглифистов это вызывает лишь недоумение.
164

линий /25 мм2
24.0 Сf 923.022.0 ▲21.0 т20.019.0 I18.0 Дети 8-12 л ет Только дети17.016.015.0
Д ети 13-17 л ет Ж енщ ины
14.0 Мужчины
13.0 В зрослы е
12.0 С11.0 В10.09.08.0 . A
Рисунок 7.1. Внутри- и межпопуляционная изменчивость плотности папиллярных линий на участке 25 мм2, половые различия и возрастные категории признака: A - с учетом данных криминалистов разных стран, только взрослые (указанных в Таблице 1); B - по пересчитанным данным российских криминалистов, которые используют археологи (например: Сыроватко, Елистратов, 2002); С - по пересчитанным данным из: Kralik, Novotny, 2003.Figure 7.1. Intra- and inter-population variability of mean ridge density, sexual differences and age categories: A - data of criminologists from different countries, only adults (listed in Table 1); B - corrected data of Russian criminologists applied by archaeologists (example: Сыроватко, Елистратов 2002); С - corrected data from Kralik, Novotny 2003.
В современных палеодерматоглифических исследованиях доказуемых оснований делить керамические изделия на изготовленные детьми и изготовленные взрослыми в действительности не больше, чем оснований делить эти же сосуды как на изготовленные мужчинами и женщинами. Представляется, что половозрастные определения не могут основываться на столь
165

ненадежном и изменчивом признаке как плотность папиллярных линий.
При этом нет никаких оснований сомневаться в том, что плотность папиллярных линий пальцевых и ладонных узоров уменьшается с возрастом и ростом организма, как и в том, что для отпечатков пальцев женщин в среднем характерны более тонко и плотно расположенные папиллярные линии, чем для мужчин. Однако существует целый ряд причин, препятствующих созданию универсальной схемы определения пола и возраста человека.
Средняя плотность папиллярных линий зависит, по меньшей мере, от семи факторов: 1. от локализации на конкретном пальце; 2. от типа узора и его участка; 3. от пола; 4. от возраста; 5. от конституциональных особенностей; 6. от длины тела; 7. от популяционных особенностей (антропологического типа).
Вероятно, для археологических материалов все же может быть выстроена более обоснованная система признаков, определяющих половозрастные характеристики. Необходимыми условиями для этого являются создание общей базы данных отпечатков пальцев и ладоней для разных регионов и эпох, более строгий отбор материалов, а также учет археологического контекста находок. Однако даже в этом случае весьма высока вероятность того, что результаты исследований не оправдают усилий, затраченных на их проведение.
Tеоретически издание такой базы данных частично может опираться на результаты исследования кожного покрова мумий, которые могут рассматриваться в качестве особого объекта изучения в палеодерматоглифике (Kralik, Nejman, 2007). Кожный покров мумифицированных кистей после специальной обработки может быть приведен в состояние, пригодное для снятия отпечатков и проведения исследований папиллярных узоров (Schmidt et al., 2000; Fields, Molina, 2008). Находки мумий известны практически для всех частей света. Помимо знаменитых египетских мумий, большое количество мумифицированных останков различной степени сохранности обнаружено в торфяных болотах Северной и Западной Европы, погребальных катакомбах Италии, монастырях Чехии и Японии, памятниках древних археологических культур на территории современных Перу и Чили и др. Большую известность в научных кругах приобрели таримские мумии II тысячелетия до н.э. - нач. I тыс. н. э., сохранившиеся в засушливых условиях пустыни
166

западных районов Китая (см. например, сборник: Humanmummies..., 1996).
Изучение мумий представляет несомненный интерес для специалистов, занимающихся исследованием различных сторон быта, особенностей внешнего облика, диеты, болезней и генетических особенностей древнего человека. Однако результаты исследований узоров кожи мумий вряд ли могут оправдать затраченные на подготовку и проведение анализа усилия. К сожалению, сохранность большинства известных мумий все же не позволяет получить полное описание папиллярных узоров индивида, а по редким исключениям из этого правила невозможно составить представление о характерных для конкретной древней популяции значениях дерматоглифических признаков и их изменчивости.
167

%

Раздел II.
РАСОВЫЕ, ЭТНИЧЕСКИЕ И ПОПУЛЯЦИОННЫЕ АСПЕКТЫ
ДЕРМАТОГЛИФИЧЕСКИХ ИССЛЕДОВАНИЙ

Глава 8.Внутрирасовое распределение признаков дерматоглифики и их
комплексов. Ключевые признаки и их свойства. Расовые градиенты и расовые комплексы.
Из многих известных схем расового деления человечества был выбран модифицированный (сокращенный) вариант классификации К.С. Куна (Coon, 1965), позволяющий дать обобщенную характеристику пяти основных расовых групп человечества. В рамках этой классификации были рассмотрены все наличные литературные и оригинальные данные Г.Л. Хить.
При описании признаков выдерживалась следующая схема. В первую очередь рассматривались ключевые признаки, далее - все остальные, сгруппированные в последовательности от пальцевых к ладонным. В программу исследования вошли классические признаки дерматоглифики кисти, за исключением гребневого счета, сгибательных борозд и минуций, а также углов.
В общей сложности описываются 24 признака кожного рельефа: дельтовый индекс (Dl10), индекс Камминса (Ic),проксимальный трирадиус ладони (t), узорность гипотенара (Hy), добавочные межпальцевые трирадиусы (ДМТ), узорность тенара / первой межпальцевой подушечки (Th/I), пальцевые бедельтовые (A), однодельтовые (U и R) и двудельтовые (W) узоры, индекс Пола (100*A/L), индекс Данкмейера (100*A/W), индекс Фуругаты (100*W/L), средний балл окончания линии A, средний балл окончания линии D, варианты отсутствия, редукции и тип 8 линии С, промежуточный (t') и срединный (t'') ладонные трирадиусы, партинг (t°), узорность II, III и IV межпальцевых ладонных подушечек, частота радиальных пальцевых узоров (FRP).
Основу анализа составили данные свыше 1400 мужских и женских выборок из разных регионов. Число мужских выборок, как правило, значительно больше, чем женских, поэтому далее в тексте приводятся цифры, относящиеся к мужчинам. Однако в таблицах и на графиках дана также характеристика соответствующих женских групп. Расовые различия описываются по данным о мужчинах, женские группы привлекаются для сравнения лишь в случае несовпадения тенденций варьирования у обоих полов (Хить, Долинова, 1990; 2002).
170

8.1. Ключевые признаки
Дельтовый индекс (Dl10)
Признак хорошо разграничивает большинство рас и имеет одновершинное распределение (Рис. 8.1). Кривые всех расовых групп трансгрессируют.7 Наиболее близки между собой кривые европеоидов и негроидов, смещенные в область малых значений признака. Взаимно сходны также кривые американоидов и монголоидов, занимающие срединное положение. Кривая австралоидов тяготеет к зоне высоких величин признака. Наименьший дельтовый индекс характерен для негроидов (средняя величина 12.8), далее следуют европеоиды (13.1), американоиды (13.5), монголоиды (14.4) и австралоиды (15.8). Статистические различия между всеми расами значимы. Внутрирасовая популяционная вариабельность признака невелика (V=5.9-9.4); наиболее разнородны индейцы Америки, наиболее гомогенны монголоиды.
Среди европеоидных вариантов минимальной узорной интенсивностью отличаются европейцы, максимальной - население Передней и Средней Азии. Кавказцы занимают промежуточное положение между крайними вариантами. Среднее число дельт достигает в Европе 12.6, на Кавказе 13.3, в Передней и Средней Азии - 13.9 и 14.0, соответственно. Внутрирасовая межгрупповая вариабельность европеоидов невелика, что доказывается небольшими колебаниями коэффициентов вариации (V=4.7-5.6).
Закономерно различаются между собой монголоиды Северной и Южной Азии: у первых узорность интенсивность пальцев немного ниже. Об этом говорят как кривые распределения признаков, так и средние значения его (14.2 и 14.6, соответственно). Южноазиатские монголоиды более гомогенны по сравнению с северными (V=4.8 и 6.3 соответственно).
7 Следует отметить, что трансгрессия (в различной степени) характерна для распределения частот всех без исключения дерматоглифических признаков.
171

Рисунок 8.1. Дельтовый индекс. Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды. Figure 8.1. Intra-racial distribution of population values of pattern intensity index (left) and average racial values of the trait (right).E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
172

Индекс Камминса (Ic)
В трех расовых группах (европеоиды, негроиды и индейцы) признак распределяется по одновершинному типу (Рис. 8.2). У монголоидов и австралоидов отмечается двувершинное распределение. Кривая европеоидов наиболее сдвинута в область высоких значений признака, противоположным свойством отмечены кривые негроидов и американских индейцев. Срединное положение между крайними вариантами занимают монголоиды и австралоиды, причем последние обнаруживают определенную близость к европеоидам. Наиболее низкий индекс в среднем отмечается у американоидов и негроидов (7.2), а самый высокий - у европеоидов (8.6). Промежуточное положение занимают монголоиды (7.7). По данным о женщинах негроиды характеризуются чуть меньшей величиной признака (7.0) по сравнению с индеанками (7.13). Различия между расами варьируют у мужчин от 0.3 до 1.44 единицы индекса и значимы. Внутри расы признак варьирует в узких пределах, коэффициент межгрупповой изменчивости составляет 4.6 - 9.5. Европеоиды - наиболее гомогенная с этой точки зрения раса, австралоиды максимально разнородны.
Европеоидные варианты группируются следующим образом: и кривые распределения популяционных частот, и среднерасовые величины признака выделяют переднеазиатскую и кавказскую группы, обладающих повышенными значениями признака (8.9-8.7), противоположным свойством отличаются среднеазиатские европеоиды (8.5). Население Европы располагается посередине между кавказскими и среднеазиатскими популяциями (8.6).
Европейцы не различаются на статистически значимом уровне с европеоидами Кавказа и Средней Азии. Во всех остальных случаях сопоставления европеоиды разных регионов отчетливодифференцированы: абсолютная разница в величине признака варьирует от 0.2 до 0.4 единиц индекса. Внутрирасоваямежгрупповая изменчивость признака у всех расовых групппримерно одинакова (V=4.2-4.8).
Кривые распределения признака у северных и южныхмонголоидов близки к нормальному типу, причем первая отчетливо смещается в область средних и повышенных значений признака, в то время как вторая тяготеет к области малых величин. Таким образом, в Сибири индекс заметно выше, чем в Южной Азии.
173

Рисунок 8.2. Индекс Камминса. Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды. Figure 8.2. Intra-racial distribution of population values of main line index (left) and average racial values of the trait (right).E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
174

В связи с этим обстоятельством находится уже отмеченная двувершинность обобщенной «монголоидной» кривой. Обе группы монголоидов значимо различаются на самом высоком уровне. Северные монголоиды более гомогенны по сравнению с южными: соответствующие межгрупповые коэффициенты вариации равны 4.9 и 6.9.
Проксимальный одиночный трирадиус ладони (t)
Распределение признака имеет близкий к нормальному характер там, где число выборок в расовой группе достаточно велико (европеоиды, монголоиды), а также у австралоидов (Рис. 8.3). Европеоиды и сходные с ними австралоиды располагаются в основном в области средних величин, в то время как негроидная кривая имеет две вершины и наиболее смещена в область низких значений признака. Кривая распределения признака у монголоидов смещена в область высоких значений. К сожалению, сравнимые данные об американоидах слишком малочисленны, что не позволяет исследовать тип распределения выборочных средних как в данном, так и в остальных случаях, однако для среднерасовой характеристики этот признак использован. Максимальной среднерасовой частотой низкого трирадиуса ладони отличаются монголоиды (76%), минимальной - американские индейцы (51.3%). С индейцами сближаются негроиды (56%). Австралоиды и европеоиды взаимно сходны и характеризуются промежуточной частотой признака по сравнению с крайними вариантами (63 -65%).
Тестирование различий по этому признаку не выявило расхождений между европеоидами и австралоидами. Во всех остальных случаях различия между расами колеблются от 5 до 14% и значимы.
Показатель внутрирасовой межпопуляционной изменчивости характеризует индейцев и негроидов как самые гетерогенные расы (коэффициенты вариации достигают 29 и 20%, соответственно). Монголоиды и европеоиды заметно более гомогенны и практически не различаются в этом отношении (V=11.6 и 12.9).
Среди европеоидных вариантов заметно выделяются среднеазиатские популяции, кривая распределения частот у которых смещена в область высоких величин. Кривые остальных подразделений европеоидов практически совпадают.
175

t % 6 ?
30 ' ' SO' '' 70' 90
Рисунок 8.3. Проксимальный трирадиус ладони. Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды.Figure 8.3. Intra-racial distribution of population values of palmar proximal triradius (left) and average racial values of the trait (right).E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
Наименьшую среднюю величину признака имеют кавказцы (около 61%), с которыми сближаются переднеазиатские группы и европейцы (64-65%). Максимальной частотой характеризуются среднеазиатские популяции (72%). Все варианты европеоидов
176

значимо различаются между собой по средней частоте низкого трирадиуса. Внутрирасовая гетерогенность европеоидов разных регионов примерно одинакова: коэффициент межгрупповойвариации признака колеблется от 10 в Средней Азии до 14% в Передней Азии.
У северных монголоидов признак развит сильнее по сравнению с южными монголоидами (78 и 70%, соответственно). Обе группы монголоидов дифференцированы примерно в равной степени: коэффициент межгрупповой вариации составляет около 11%.
Узорность гипотенара (Ну)
Признак всюду распределяется по одновершинному, причем у монголоидов, негроидов и европеоидов - практически по нормальному типу (Рис. 8.4). Единственное исключение составляют австралоиды, у которых кривая распределения имеет два отчетливых зубца, отражающих, по-видимому, расовую гетерогенность группы. Расовые различия выражены отчетливо во всех случаях сопоставления.
Наименьшая узорность характерна для американоидов, максимальная - для европеоидов: средние частоты равны 13 и 32%, соответственно. Австралоиды, монголоиды и негроиды взаимно различаются слабо, но закономерно. Средняя частота признака составляет у австралоидов и негроидов около 23, у монголоидов - 22%. Коэффициент внутрирасовой межгрупповой вариации достаточно высок (31-38%) у всех рас, кроме европеоидной (18%). Таким образом, европеоиды отличаются исключительной гомогенностью в отношении этого признака.
Характерны различия между европеоидными вариантами. Минимальную узорность гипотенара имеют среднеазиатские популяции (29%), далее следуют европейцы (32), максимально признак выражен на Кавказе и в Передней Азии (около 35%). Коэффициенты внутрирасовой межгрупповой вариации признака колеблются от 15 до 18% и, следовательно, в этом отношении европеоидные варианты почти неразличимы.
У северных монголоидов признак встречается чаще, чем на юге (23 и 18%, соответственно). При этом первые значительно гомогеннее вторых: внутрирасовые межгрупповые коэффициенты вариации составляют 27 и 39%.
177

Рисунок 8.4. Узорность гипотенара. Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды. Figure 8.4. Intra-racial distribution of population values of hypothenar patterns (left) and average racial values of the trait (right). E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
Добавочные межпальцевые трирадиусы (ДМТ)
Признак распределяется в основном по одновершинному типу, с правосторонней асимметрией и хорошо разграничивает расы и их
178

подразделения (Рис. 8.5). Максимальным своеобразием отличаются негроиды, имеющие самое высокое содержание ДМТ (в среднем 39%!). Реже всего признак отмечен у американоидов, монголоидов и австралоидов, у которых средние частоты чрезвычайно сходны (10 - 14%). Во всех остальных случаях средние различия между расами колеблются от 2 до 26%. Признак заметно варьирует внутри расы: коэффициенты вариации групповых средних колеблются в пределах 25-67%. Наиболее гетерогенны индейцы Америки. Минимальная вариабельность признака (V=25%) характерна для негроидов, не имеющих аналогий по степени внутрирасовой гомогенности.
Среди европеоидов признак максимально выражен в Передней Азии (средняя частота достигает 26%), минимально - в Европе (в среднем до 18%). Кавказские и среднеазиатские популяции более близки к переднеазиатскому населению (24-25%). По средней частоте ДМТ население Европы сильно и статистически значимо отличаются от остальных вариантов. Внутрирасовая межгрупповая изменчивость европеоидных вариантов приблизительно одинакова (V=32-38%), за исключением среднеазиатского региона, где содержание признака варьирует в более узких пределах (V=26%).
У северных монголоидов признак встречается немного чаще по сравнению с южными, средние составляют 13 и 11%. В обоих подразделениях монголоидов признак варьирует одинаково: межгрупповые коэффициенты вариации составляют 43-44%.
Тенар / первая межпальцевая подушечка (Th/I)
Признак распределяется по одновершинному типу с правосторонней асимметрией всюду, за исключением малочисленных австралоидов, дающих резко асимметричное «ступенчатое» распределение (Рис. 8.6). По кривым распределения частот и по средним величинам (9.9%) европеоиды и монголоиды абсолютно идентичны. Средняя величина признака максимальна у индейцев Америки (около 26%). Промежуточное положение между крайними вариантами занимают австралоиды (19%) и негроиды (15%). По внутрирасовой вариабельности рассмотренные расовые группы заметно неоднородны. Наиболее гетерогенны монголоиды (V=50%) и австралоиды (58%). У негроидов и европеоидов признак варьирует почти в одинаковой степени (V=38%). У американских индейцев внутрирасовая вариабельность признака заметно сужена по сравнению с остальными расами (V=31.6%).
179

ДМТ % (3 d ?
1 1 ' ' 17 ' ' ' 33 ' ' ' 4 9 ' ' ' 65
Рисунок 8.5. Добавочные межпальцевые трирадиусы. Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды. Figure 8.5. Intra-racial distribution of population values of accessory interdigital triradii (left) and average racial values of the trait (right).E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
180

Рисунок 8.6. Узорность тенара/первая межпальцевая подушечка. Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды. Figure 8.6. Intra-racial distribution of population values of thenar / interdigital I area patterns (left) and average racial values of the trait (right).E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
181

Таким образом, отмеченная выше максимальная величина признака, характерная для индейцев, составляет их стойкую расовую особенность. Европеоидные варианты на первый взгляд мало отличимы друг от друга по содержанию признака. Средняя частота его колеблется от 9 до 12%, максимальная выраженность отмечается в Средней Азии, минимальная - в Европе.
Межгрупповая вариабельность признака в пределах указанных групп европеоидов приблизительно одинакова (V=34-36%), за исключением Кавказа, более гетерогенного в этом отношении (V=43%).
Монголоидные варианты по данному признаку кажутся сходными: в Северной Азии средняя частота составляет 11, а в Южной - 7%. Признак заметно сильнее варьирует у южноазиатских монголоидов по сравнению с североазиатскими (V=54 и 46%, соответственно).
8.2. Неключевые признаки
8.2.1. Пальцевый рельеф
Бездельтовый узор (А + Т)
Распределение частот признака имеет практически нормальный вид с правосторонней асимметрией, идентичный у европеоидов и негроидов (Рис. 8.7). У австралоидов признак имеет резко выраженное асимметричное распределение. Сходное асимметричное распределение с концентрацией частот в области низких величин характеризует монголоидов и американоидов. Минимальная величина средней арифметической признака характеризует австралоидов (1.2%). Далее следуют монголоиды (3.2), американоиды (4.5), европеоиды и негроиды (6 и 6.4%, соответственно).
Практически абсолютно совпадают среднерасовые характеристики у негроидов и европеоидов, с одной стороны, а также монголоидов и австралоидов, с другой (разница составляет 0.2-0.3%). Признак отличается значительной внутрирасовой вариабельностью. Наиболее разнородны американоиды и австралоиды (V=79-88%). Наиболее гомогенны европеоиды и негроиды (V=41-45%). Монголоиды более сходны в этом смысле с американоидами (V=68%), хотя и более гомогенны.
182

Рисунок 8.7. Бездельтовые узоры (А+Т). Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды. Figure 8.7. Intra-racial distribution of population values of simple and tented archs (left) and average racial values of the trait (right). E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
183

Для европеоидных вариантов характерно одновершинное распределение с небольшой тенденцией к правосторонней асимметрии. Наименьшие частоты свойственны передне- и среднеазиатским европеоидам.
По распределению средних величин отмечается такая последовательность: Передняя Азия и Средняя Азия (4.5%), Кавказ (5.4%), Европа (6.8%). Абсолютная величина различий колеблется от 0 до 2.3%. Максимально различаются европеоиды Европы и Средней Азии. Отметим, что наиболее гомогенны по данному признаку европеоиды Европы: коэффициент межгрупповойвариации равен 35.6. Европеоиды остальных регионов более гетерогенны (V=41-45%) и взаимно сходны.
Резко асимметричное распределение с преобладанием низких частот характерно для подразделений монголоидов. У североазиатской группы бездельтовый узор встречается вдвое чаще (4%), чем у южноазиатской (2.1). Сибирские монголоиды по содержанию признака несколько гетерогеннее южноазиатских: коэффициенты межгрупповой вариации равны 60 и 56.1%, соответственно.
Однодельтовый узор (R + U)
Суммарное количество радиальных и ульнарных петель у мужчин распределяется по одновершинному типу с тенденцией к левосторонней асимметрии (Рис. 8.8). Лишь в австралоидной группе распределение имеет двувершинный тип. Кривые негроидов и европеоидов почти полностью совпадают.
Минимальное количество петель характеризует австралоидов (в среднем 42.6%). Далее следуют монголоиды (49.2%), американоиды (53.7%), европеоиды (57.4%) и негроиды (62%). Абсолютная разница в содержании петель между большими расами колеблется от 4 до 19%. Наиболее гомогенны по содержанию признака европеоиды и негроиды (коэффициенты межгрупповой вариации равны 10.5 и 11%) С ними сближаются монголоиды (V=13.4%). Сильнее всего признак варьирует у американоидов и австралоидов (V равны 18.1 и 27.4 соответственно). Среди европеоидных вариантов, которые характеризуются симметричным распределением, минимальное количество узоров наблюдается в Средней Азии (51.3%). Далее следуют Передняя Азия (54.6%), Кавказ (56.9%) и Европа (59.6%).
184
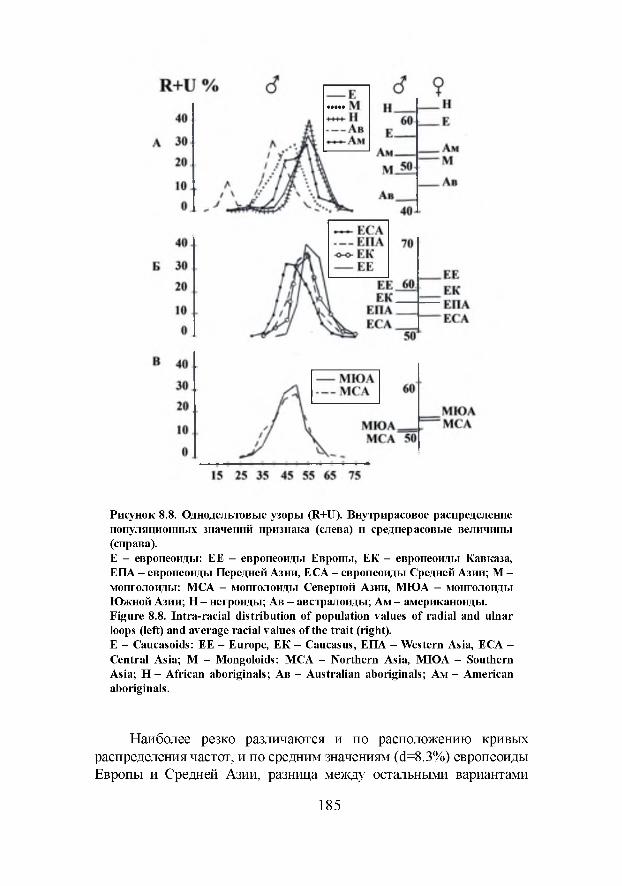
Рисунок 8.8. Однодельтовые узоры (R+U). Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды. Figure 8.8. Intra-racial distribution of population values of radial and ulnar loops (left) and average racial values of the trait (right).E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
Наиболее резко различаются и по расположению кривых распределения частот, и по средним значениям (d=8.3%) европеоиды Европы и Средней Азии, разница между остальными вариантами
185

составляет 2.3-5.6%. Для монголоидных вариантов также характерно нормальное и почти совпадающее распределение, небольшие различия можно обнаружить лишь на уровне средних величин: в Южной Азии частота признака незначительно выше, чем в Северной Азии (49.5 и 49%, соответственно). Северные монголоиды чуть более разнородны по содержанию признака, чем южные; коэффициент межгрупповой вариации составляет 13.9 и 12.8%.
Двудельтовый узор (W)
Как и в предыдущем случае, распределение имеет одновершинный симметричный тип, за исключением кривой у австралоидов, имеющей две вершины (Рис. 8.9). Минимальное количество завитков характерно для негроидов (средняя величина 31.4%) и европеоидов (36.9%). Далее следуют американоиды (41.7%), монголоиды (47.4%) и австралоиды (57.6%). Средние различия между расами составляют 5-26%. Наиболее сближены европеоиды с американоидами и негроидами, наиболее резко и значимо различаются негроиды и австралоиды. Кривые распределения признака у европеоидов разных регионов, как правило, одновершинны и симметричны и отчетливо дифференцируют эти варианты. Наименьшую величину признака имеет население Европы (33.9%), далее следуют популяции Кавказа (37.9%), Передней Азии (41.7%) и Средней Азии (44%). Абсолютные различия среднерасовых значений минимальны между передне- и среднеазиатскими популяциями (d=2.3%). Во всех остальных случаях сопоставлений разница между вариантами составляет 4-10%. Внутрирасовая вариабельность признака у европеоидов разных регионов, в общем, сходна: коэффициент межгрупповой вариабельности составляет 13.3-17.5. Взаимно сходны по степени вариабельности признака популяции Кавказа и Передней Азии, с одной стороны (V=16.4-17.5%), Средней Азии и Европы, с другой. Среди монголоидных вариантов признак встречается немного реже у северных монголоидов (46.7%) по сравнению с южными (48.3%). Кривые распределения частот близки к нормальным и почти совпадают, хотя североазиатская кривая несколько смещена в область малых величин. Сибирские монголоиды немного более гетерогенны по сравнению с южноазиатскими (V равны 16.7 и 13.1, соответственно).
186

с? 6 ?
15 25 35 45 55 65 75
Рисунок 8.9. Двудельтовые узоры (W). Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды. Figure 8.9. Intra-racial distribution of population values of whorls (left) and average racial values of the trait (right).E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
187

Индекс Полла (100*A/L)
Распределение признака имеет вид одновершинной кривой с многочисленными отдельными зубцами и тенденцией к правосторонней асимметрии (Рис. 8.10). Лишь у австралоидов признак распределяется с сильной положительной асимметрией. Расовые различия выражены нечетко ввиду наложения почти всех кривых друг на друга и выявляются лишь благодаря средним значениям. Минимальную пропорцию дуг по отношению к петлям имеют австралоиды (2.2), далее следуют монголоиды (6.6), американоиды (9), негроиды (10.1) и европеоиды (10.4) Внутрирасовая гетерогенность сильнее всего выражена у австралоидов (V=81%), наиболее гомогенны европеоиды и негроиды (V равен 42 и 45%, соответственно). Срединное положение между этими двумя крайними вариантами занимают американоиды и монголоиды, сходные друг с другом (V равен 59 и 62%).
Европеоиды мало различаются по типу распределения частот индекса. Минимальную среднюю величину имеют передне- и среднеазиатские популяции (8.2 и 8.6). Далее следуют популяции Кавказа (9.4) и Европы (11.5). Все варианты европеоидов, за исключением среднеазиатского, дифференцированы почти в одинаковой мере (V варьирует от 38 до 41%). Среднеазиатские популяции отличаются повышенным разнообразием (V=48%). Различия между монголоидами более отчетливы: в Южной Азии величины индекса гораздо меньше по сравнению с Северной. Соответствующие средние составляют 4.2 и 8.2. При этом сибирские популяции немного разнороднее, чем южноазиатские: коэффициент межгрупповой вариации признака равен 54 и 52, соответственно.
Индекс Данкмейера (100* A/W)
Расположение кривых распределения частот сходно с описанным в предыдущем случае (Рис. 8.11). Тенденции расовых различий прослеживаются лишь по средним величинам: признак имеет минимальную величину в группе австралоидов (2.2), ближе всего к которым стоят монголоиды (7.8). Далее следуют американоиды (12.3), европеоиды (17.3) и негроиды (20.2). Две последние группы выделяются на фоне остальных также повышенной гомогенностью по данному признаку: коэффициент межгрупповой вариации составляет 56 и 50%, соответственно.
188

Рисунок 8.10. Индекс Полла (100*A/L). Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды. Figure 8.10. Intra-racial distribution of population values of Poll’s index (left) and average racial values of the trait (right).E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
189

Рисунок 8.11. Индекс Данкмейера (100*А/W). Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды. Figure 8.11. Intra-racial distribution of population values of Dankmeijer’s index (left) and average racial values of the trait (right).E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
190

В то же время наибольшей разнородностью характеризуются австралоиды (V=73%) и особенно индейцы (V=77%). Среди европеоидов минимальной величиной признака отличаются средне - и переднеазиатские группы (10.3 и 10.6, соответственно), с которыми сближаются кавказские популяции (14.1). В Европе индекс максимален (21.0). По внутрирасовой вариабельности признака систематические различия между европеоидами разных регионов не обнаружены, коэффициент межгрупповой изменчивости колеблется от 44 в Европе до 53 на Кавказе. Как и по предыдущему индексу, признак хорошо разграничивает северо- и южноазиатские монголоидные популяции, если исходить из кривых распределения и средних значений. Более компактное распределение и малые значения (4.5) индекса характерны для Южной Азии по сравнению с Северной (9.3). В Сибири признак варьирует сильнее, чем в Южной Азии; V составляет 63 и 58%, соответственно.
Индекс Фуругаты (100*W/L)
Признак распределяется по одновершинному типу с заметной тенденцией к правосторонней асимметрии (Рис. 8.12). Наименьшие величины характеризуют негроидов (средняя составляет 52.2), с которыми сближаются европеоиды (65.8). Повышенной концентрацией завитков относительно петель отличаются американоиды (84.8), монголоиды (99.1) и в особенности австралоиды (162.0!). В пределах расы у монголоидов, европеоидов и негроидов признак варьирует умеренно, V составляет 30-32%. Напротив, более гетерогенны австралоиды (50.0) и монголоиды (44.0%).
Среди подразделений европеоидов и по кривым распределения, и по средним значениям наиболее сходны передне- и среднеазиатские группы (77.7 и 86.1, соответственно), у которых завитки максимально часты по отношению к петлям. В Европе индекс минимален (57.6). Кавказские популяции сближаются с переднеазиатскими (69.5). На Кавказе, помимо этого, наблюдаем повышенную гетерогенность популяций по данному признаку (V=30%), в то время как европеоиды остальных регионов дифференцированы немного слабее и почти одинаково (V=24-26%).
191
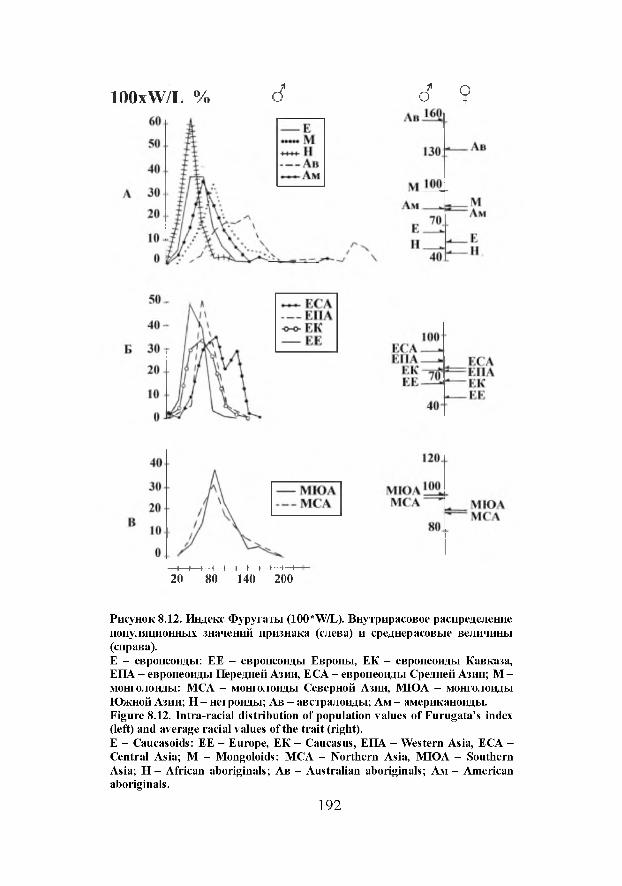
IOOxW/L % <$ J ?
— I---- 1 t i » -t —<---- t - H ---- 1----120 80 140 200
Рисунок 8.12. Индекс Фуругаты (100*W/L). Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды. Figure 8.12. Intra-racial distribution of population values of Furugata’s index (left) and average racial values of the trait (right).E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
192

Различия между монголоидами разных регионов почти неощутимы: кривые распределения практически совпадают, а средняя величина индекса в Южной Азии чуть выше по сравнению с Северной (99.6 и 98.4, соответственно). В Сибири признак варьирует сильнее (39%), чем в Южной Азии (32%).
8.2.2. Ладонный рельеф
Среднее окончание линии A (MA)
В случаях, когда число выборок достаточно велико, распределение признака внутри расы близко к нормальному (европеоиды, монголоиды, негроиды). У австралоидов кривая имеет две отчетливые вершины (Рис. 8.13). Тенденции различий между расами выражены явственно: у американоидов наиболее низкое окончание (2.81), у австралоидов - максимально высокое (3.74); с австралоидами сближаются европеоиды (3.67). Монголоиды занимают срединное положение между крайними вариантами (3.28), в то время как негроиды более тяготеют к европеоидам и австралоидам (3.47). Абсолютная разница между среднерасовыми значениями признака варьирует от 0.1 до 0.9.
Внутрирасовая вариабельность признака исключительно мала, величина коэффициента межгрупповой вариации колеблется от 5.4 до 9.0. В эти пределы попадают все расы, кроме австралоидной. Разнородность последней заметно выше (V=13.8).
Обращаясь к европеоидным вариантам, отметим совмещение кривых распределения частот у европеоидов Европы и Средней Азии, с одной стороны, Кавказа и Передней Азии - с другой.
По среднерасовым значениям самое низкое окончание линии А наблюдается в Средней Азии, наиболее высокое - в Передней Азии. Европа и Кавказ практически неразличимы и располагаются посередине между крайними вариантами, разница между которыми составляет всего 0.2 балла (3.61 и 3.80, соответственно). Примечательно, что взаимоотношения подразделений европеоидов по данным о мужчинах и женщинах неодинаковы: минимальной частотой признака в первом случае обладают среднеазиатские европеоиды, а во втором - европейцы. Подразделения европеоидной расы дифференцированы практически в одинаковой мере: величина коэффициента внутрирасовой межгрупповой вариации составляет 7.2-8.7. Наиболее однородны популяции Передней Азии и Кавказа.
193

Рисунок 8.13. Средний балл окончания линии А. Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды. Figure 8.13. Intra-racial distribution of population values of main line A termination’s symbols (left) and average racial values of the trait (right).E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
194

При сравнении северных и южных монголоидов выявлено почти полное совпадение кривых распределения частот признака и более заметная растянутость «южноазиатской» кривой. В целом, у популяций Северной Азии линия А заканчивается немного ниже, чем в Южной (3.25 и 3.42, соответственно). В Южной Азии признак варьирует вдвое больше, чем в Северной (V равны 12.8 и 5.5).
Среднее окончание линии D (MD)
Признак хорошо разграничивает расы и распределяется, в основном, по одновершинному типу у всех рас, кроме монголоидной (Рис. 8.14). Наиболее низкие частоты характерны для негроидов (8.42). Американоиды, австралоиды и монголоиды образуют сравнительно компактную группу (8.89, 9.01 и 9.18,соответственно). Самые высокие частоты и наибольшая средняя величина признака характеризует европеоидов (9.69). Абсолютные различия в величине среднего балла D варьируют значительно - от 0.12 до 1.20. Европеоидная раса наиболее гомогенна по сравнению с остальными, если судить по величине коэффициента межгрупповой вариации признака (V=2.5). Все остальные расы дифференцированы заметно сильнее и почти в равной степени (V равен 4.1-5.4). Наиболее гетерогенны австралоиды.
Европеоидные варианты различаются слабо, но закономерно. В Средней Азии признак имеет наименьшую величину (9.56). Во всех остальных подразделениях варьирует от 9.70 (Европа) до 9.78 (Передняя Азия). Абсолютные различия в средней величине признака не дифференцируют кавказские и переднеазиатские популяции (d=0.03), а также европейские и кавказские (d=0.05). В остальных случаях разница колеблется от 0.08 до 0.22. У европеоидов разных регионов признак мало варьирует (V=2.3-2.6). Монголоидные варианты сильно различаются и по кривым распределения популяционных частот (кривая южных монголоидов смещена в область очень малых значений), и по средним значениям признака.
Крайне важно отметить, что различия между монголоидными вариантами по окончанию линии А имеют противоположное направление. Трудно судить, в какой степени эта картина отражает реальную ситуацию: число выборок, касающихся линии А, почти вдвое меньше по сравнению с окончанием линии D, так что здесь мог проявиться эффект случайности.
195

Рисунок 8.14. Средний балл окончания линии D. Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды. Figure 8.14. Intra-racial distribution of population values of main line D termination’s symbols (left) and average racial values of the trait (right).E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
196

В Южной Азии среднее значение признака составляет 8.69, в Северной - 9.40. Межгрупповая вариабельность чуть выше, чем у европеоидных вариантов (V=3-3.1) и не дифференцирует северных и южных монголоидов.
Варианты окончания линии С
Отсутствие линии (С0)
Распределение частот признака имеет одновершинное распределение лишь у монголоидов и европеоидов, со значительной правосторонней асимметрией (Рис. 8.15). В остальных случаях оно характеризуется неупорядоченностью и асимметричностью, причем кривые практически накладываются друг на друга.
Средняя величина признака почти не разграничивает расы. У европеоидов, монголоидов, негроидов и австралоидов она варьирует в пределах 4-7%. Чаще всего линия С отсутствует у индейцев Америки (16%), резко выделяющихся на фоне остальных групп. Среднерасовая разница минимальна между австралоидами и негроидами (0.1%). В остальных случаях сопоставлений разница варьирует от 1.2 до 11.2%. Признак характеризуется значительно более выраженной вариабельностью по сравнению с предыдущим.
По среднерасовой величине коэффициента межгрупповой вариации монголоиды сближаются с негроидами (V=59-61%). Индейцы резко контрастируют с остальными расами, благодаря своей относительной гомогенности (V=40%).
Европеоидные варианты практически не различимы по частоте С0 (3.8-4.4%), за исключением Средней Азии, где признак отсутствует вдвое чаще (8%). Интересно, что в наиболее многочисленной группе европейцев признак распределяется наиболее асимметрично. Попарные различия между регионами невелики и не превышают 1%. Переднеазиатские и среднеазиатские европеоиды сходны, будучи наиболее гомогенны по степени межгрупповой вариабельности признака (V=47-48%). В то же время европеоиды Кавказа и Европы более гетерогенны (V=60-68%).
Кривые распределения частот у монголоидных вариантов почти совпадают. Признак в среднем выражен чуть сильнее у южных монголоидов (около 7%) по сравнению с северными (6%). В Северной Азии признак варьирует немного слабее, чем в Южной (V равны 51 и 54%, соответственно).
197
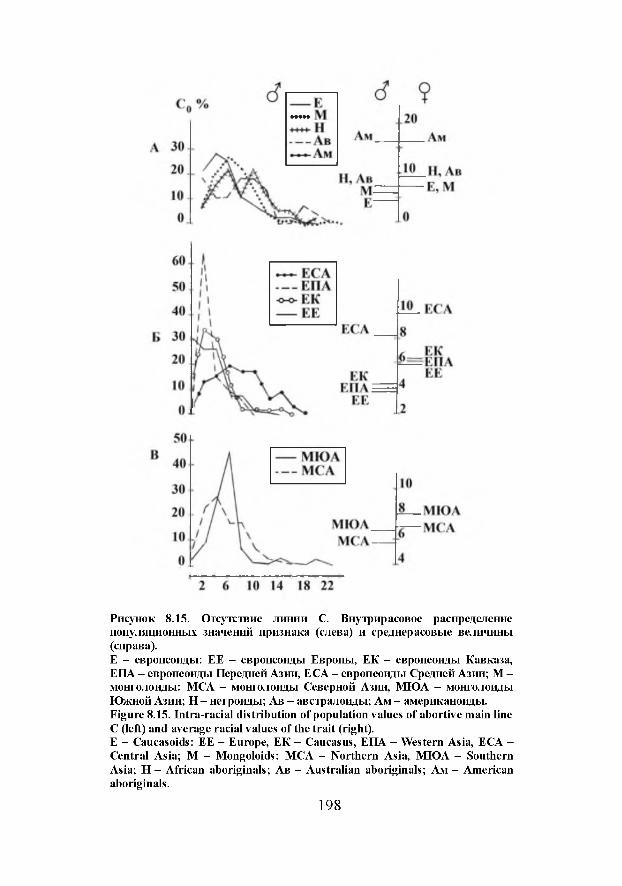
Рисунок 8.15. Отсутствие линии С. Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды. Figure 8.15. Intra-racial distribution of population values of abortive main line C (left) and average racial values of the trait (right).E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
198

Редукция линии С (Сх)
Методическая сопоставимость мировых данных этого признака невелика. Поэтому пришлось отказаться от мысли построения кривых его распределений у всех рас, за исключением европеоидной и монголоидной, большая часть данных о которых принадлежит отечественным авторам (Рис. 8.16).
Внутри монголоидной расы признак распределяется по одновершинному типу со значительной правосторонней асимметрией. «Европеоидная» кривая почти полностью трансгрессирует с «монголоидной», отличаясь от нее высокой концентрацией частот в классе минимальных величин. Среднерасовая частота признаков у европеоидов немного ниже, чем у монголоидов (10 и 11%). Кроме того, европеоиды по этому признаку заметно гомогеннее (V равны 40 и 56%, соответственно). Европейцы обнаруживают сходство с общеевропеоидной кривой распределения частот. В остальных регионах признак распределяется по условно нормальному типу с множеством зубцов, причем кривые практически совпадают. Максимальной средней частотой признака обладают кавказцы (около 14%), минимальной - европейцы (около 8%). В Передней и Средней Азии средняя величина признака составляет 11-13%. Признак варьирует в пределах регионов неодинаково. Наиболее гомогенно население Кавказа (V=31%), более разнородны популяции Средней и Передней Азии, максимально разнородны европейцы (V=74%).
У северных монголоидов распределение одновершинное, чего нельзя сказать о южных; различия между теми и другими достигают в среднем 7%. В Северной Азии признак варьирует слабее, чем в Южной (V равны 49 и 59%, соответственно). Следует отметить, что большие методические различия между авторами в определении признака лишают обнаруженные различия убедительности. Важно учесть, что небольшая выборка южных монголоидов (25 выборок), в основном, составлена по данным зарубежных исследователей.
Проксимальный тип линии С (С8)
Во всех группах распределение имеет вид кривой, вытянутой по оси абсцисс (Рис. 8.17). Минимальная средняя частота, означающая практически полное отсутствие признака, характеризует негроидов (0.3%), максимальная - европеоидов (1.2%).
199
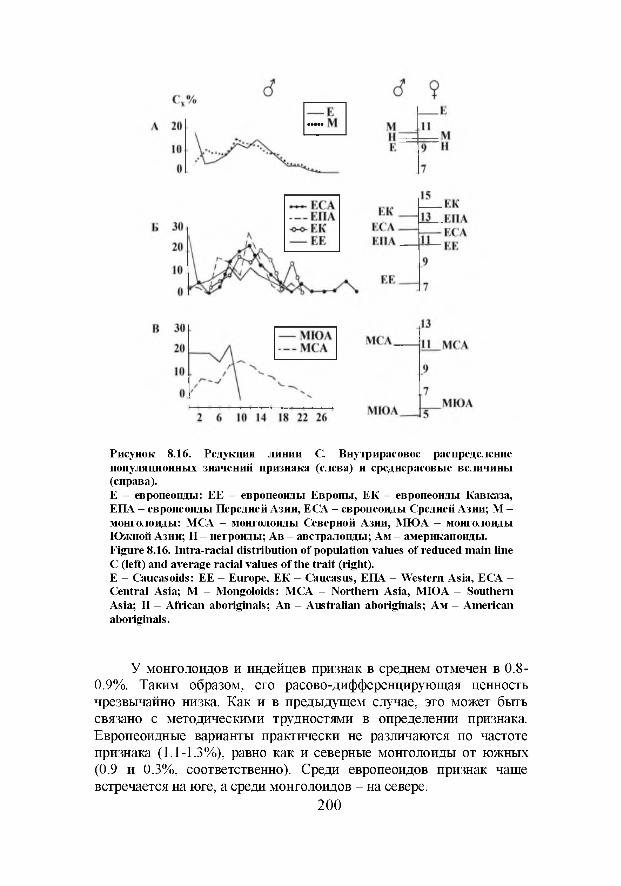
Рисунок 8.16. Редукция линии С. Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды. Figure 8.16. Intra-racial distribution of population values of reduced main line C (left) and average racial values of the trait (right).E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
У монголоидов и индейцев признак в среднем отмечен в 0.80.9%. Таким образом, его расово-дифференцирующая ценность чрезвычайно низка. Как и в предыдущем случае, это может быть связано с методическими трудностями в определении признака. Европеоидные варианты практически не различаются по частоте признака (1.1-1.3%), равно как и северные монголоиды от южных (0.9 и 0.3%, соответственно). Среди европеоидов признак чаще встречается на юге, а среди монголоидов - на севере.
200

Рисунок 8.17. Проксимальный тип линии С. Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды. Figure 8.17. Intra-racial distribution of population values of proximal type of main line C (left) and average racial values of the trait (right).E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
201

Партинг (t°)
Признак распределяется всюду по сильно вытянутой по оси абсцисс кривой (Рис. 8.18). Максимально часто он отмечается у европеоидов (в среднем до 1.5%), наиболее редко у негроидов (0.3%), у остальных рас - в 0.5-0.6% случаев. Внутрирасовая межгрупповая вариабельность исключительно велика: V равен 87200% (!). Наиболее гомогенна европеоидная раса, максимально разнородны индейцы. К европеоидам примыкают монголоиды (V=116%), далее следуют австралоиды, сходные с негроидами (V=140-152%).
Среди европеоидов максимальная частота наблюдается на Кавказе (до 2%), минимальная - в Передней Азии (до 1%). Диагностическая ценность признака для европеоидов существенно снижена благодаря несовпадению картины по данным о мужчинах и женщинах. Абсолютные различия между европеоидными вариантами колеблются от 0.2 до 0.1%. Среднеазиатские популяции не отличаются от европейских и кавказских. У европеоидов Европы признак варьирует минимально (V=69%) в сравнении с остальными вариантами (V равен 92-98%). Среди последних наиболее однородны кавказские популяции, максимально гетерогенны - среднеазиатские.
В пределах монголоидных вариантов региональные кривые почти совпадают, не различаются варианты и по средней частоте признака (0.5-0.6% в Южной и Северной Азии, соответственно). Население обоих регионов характеризуется одинаково высоким уровнем межгрупповой изменчивости (V=96-98%).
Промежуточный трирадиус (t’)
Для всех рас, за исключением американоидов, данные которых из-за малочисленности не были использованы, характерен почти нормальный (с тенденцией к правосторонней асимметрии) тип распределения признака (Рис. 8.19). Средняя частота варьирует от 14.5% у австралоидов до 41% (!) у американоидов.
Европеоиды и монголоиды абсолютно идентичны (17.6%), у негроидов средняя частота трирадиуса повышена и составляет 3560%. Различия между расами варьируют от 0 до 26%.
Система осевых ладонных трирадиусов
202
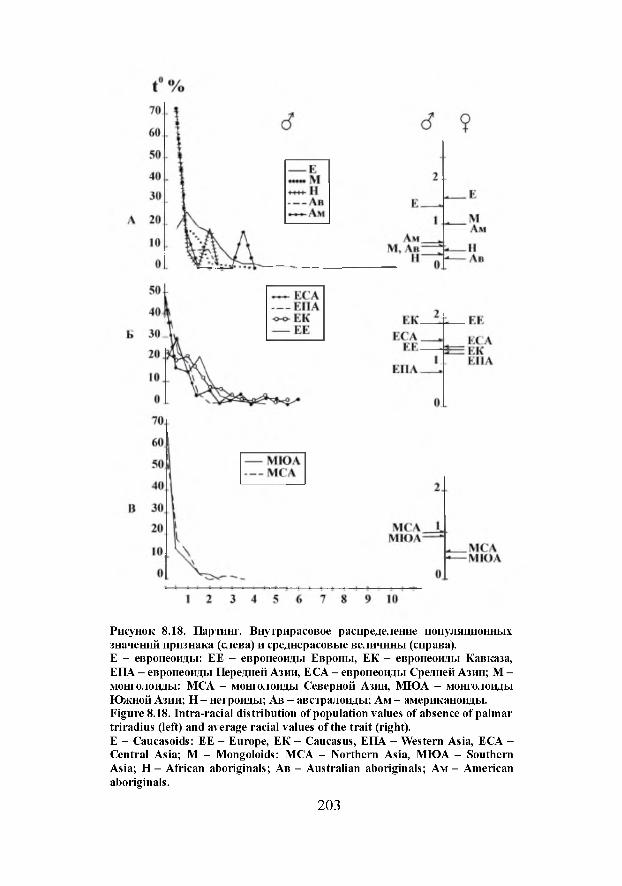
Рисунок 8.18. Партинг. Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды. Figure 8.18. Intra-racial distribution of population values of absence of palmar triradius (left) and average racial values of the trait (right).E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
203

Распределение признака у европеоидных вариантов близко к нормальному, региональные кривые практически совпадают. Среднее значение колеблется от 15% в Средней Азии до 20% на Кавказе, составляя в Европе и Передней Азии около 18%. У европейских популяций признак варьирует минимально (V=26%), в остальных регионах гетерогенность популяций заметно увеличена. (V=34-39%).
В пределах монголоидных вариантов распределение признака также сходно с нормальным, при этом кривая южных монголоидов заметно смещена в область повышенных значений. У северных монголоидов признак варьирует сильнее, чем у южных (V равны 46 и 34%, соответственно).
Срединный трирадиус (t")
У всех рас, за исключением европеоидной, признак распределяется с максимальной концентрацией в области низких значений, у европеоидов - по одновершинному типу с сильной правосторонней асимметрией (Рис. 8.20). Среднерасовая частота признака минимальна у монголоидов (около 2%), максимальна у европеоидов и австралоидов (около 4%). У индейцев признак встречается в среднем в 3%. Признак сильно варьирует внутри расы: величина коэффициента межгрупповой вариации колеблется от 69 у австралоидов до 111 у монголоидов. Европеоиды сходны в этом отношении с австралоидами (V=75%), негроиды - с монголоидами (V=176%), индейцы занимают промежуточное положение между европеоидами и монголоидами (V=91.7%).
По распределению признака среди европеоидных вариантов сближаются население Европы и Кавказа, с одной стороны, Передней и Средней Азии, с другой.
В первом случае с максимальной частотой встречаются низкие значения, но кривая деформирована за счет пониженной концентрации частот в начальном классе ряда, во втором кривая распределения имеет несколько зубцов, вероятно, возникшими из-за малой численности данных.
По среднерасовым значениям признака выделяются среднеазиатские популяции (2.8%) и их антипод - население Кавказа (4.8%). Европейские и переднеазиатские группы располагаются между этими двумя крайними вариантами и неразличимы (3.7-3.5%).
204

50
40
Б 30
20
10
0
___ МЮА-25- __ МСА
15
1 13 25 37 49
Рисунок 8.19. Промежуточный трирадиус. Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды. Figure 8.19. Intra-racial distribution of population values of intermediate palmar triradius (left) and average racial values of the trait (right).E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
205

Рисунок 8.20. Срединный трирадиус. Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды. Figure 8.20. Intra-racial distribution of population values of palmar mediate triradius (left) and average racial values of the trait (right).E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
206

Слабее всего признак варьирует на Кавказе (V=54%), максимально - в Передней Азии (V=88). В Средней Азии и в Европе коэффициент межгрупповой вариации составляет 71-82%.
Среди монголоидов обоих регионов признак имеет отчетливо гиперболическое распределение, причем, кривые почти полностью совпадают. Среднерасовые значения исключительно малы и составляют в Северной Азии 1.4, в Южной - 1.9%. Примечательно, что у южноазиатских монголоидных популяций признак варьирует значительно слабее, чем у североазиатских (V составляет 68 и 109%, соответственно).
Узорность межпальцевых подушечек
Вторая межпальцевая подушечка (II)
Данный признак учитывает исключительно частоту добавочных межпальцевых трирадиусов и окаймляемых ими петель или завитков. Распределение признака, сходное с нормальным, наблюдается у европеоидов и негроидов, то есть в тех расовых группах, которые отличаются от остальных повышенным процентом добавочных межпальцевых трирадиусов в целом (Рис. 8.21). В остальных случаях признак распределяется по гиперболе.
Кривые сильно трансгрессируют, точнее выражаясь, зачастую совмещаются друг с другом. Максимальная частота характерна для негроидов (8%). Далее следуют европеоиды (6%), монголоиды (около 4%) и австралоиды (3%). Признак минимально развит у американоидов (0.7%). Среднерасовые различия практически отсутствуют между европеоидами и австралоидами (0.5%), а в остальных случаях варьируют от 2.2 до 2.7%.
Величина коэффициента внутрирасовой межгрупповой вариации колеблется в очень широких пределах, от 54% у европеоидов, к которым тесно примыкают негроиды, до 132% у монголоидов. Австралоиды тяготеют к американоидам (V=98%), а монголоиды - к европеоидам (V=75%). Среди европеоидов распределение в основном происходит по одновершинному типу и имеет сильную правостороннюю асимметрию. Резко выделяется европейская кривая, смещенная в область малых величин, в то время как все остальные кривые практически совмещены. Соответственно этому средняя частота признака минимальна в Европе (4.8%), у остальных вариантов она почти одинакова (7.7-8.1%).
207

Рисунок 8.21. Узорность II межпальцевой подушечки. Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды. Figure 8.21. Intra-racial distribution of population values of interdigital pad II patterns (left) and average racial values of the trait (right).E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
208

В пределах монголоидных вариантов признак распределяется по гиперболическому типу, если не считать небольшого понижения частот первого класса распределения у южных монголоидов. В Северной Азии ряд частот заметно увеличен за счет класса высоких значений признака. В соответствии с этим средние расовые величины составляют 3.8% в Северной и 2.8% - в Южной Азии. И северо-, и южноазиатские монголоиды одинаково дифференцированы: коэффициент межгрупповой вариациидостигает 74%, что значительно превышает аналогичный показатель у европеоидных вариантов.
Третья межпальцевая подушечка (III)
Признак имеет сложное строение, включая как истинные петли, так и петли, образованные добавочными межпальцевыми трирадиусами. Внутри расы признак распределяется по типу, близкому к нормальному, если не считать австралоидов, распределение признака у которых образует отчетливую двувершинную кривую (Рис. 8.22). Минимальная среднерасовая величина характеризует индейцев и австралоидов (20 -21%), затем следуют монголоиды (25%) и негроиды (29%). Наиболее часто признак встречается у европеоидов (38%).
Европеоидные варианты имеют нормальный тип распределения. Кривые сильно трансгрессируют, но различия между вариантами по взаимному расположению кривых и по средним значениям очевидны и закономерны. Наиболее редок признак в Средней Азии (около 34%). В остальных случаях средняя величина колеблется от 38% в Европе до 41% в Передней Азии. Межгрупповая внутрирегиональная изменчивость признака уменьшена по сравнению с таковой узорности на II подушечке.
Слабее всего признак варьирует в переднеазиатских и кавказских популяциях (V=14-17%), затем в среднеазиатских (V=22%); максимально гетерогенны популяции Европы (V=28%).
У монголоидных вариантов распределение признака весьма условно может быть сближено с нормальным. Кривая сибирских популяций сдвинута в область высоких значений, хотя трансгрессия с южноазиатской кривой значительна. В Южной Азии признак встречается гораздо реже (16%), чем в Северной (26%).
209

Рисунок 8.22. Узорность III межпальцевой подушечки. Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды. Figure 8.22. Intra-racial distribution of population values of interdigital pad III patterns (left) and average racial values of the trait (o right).E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
210

Примечательно, что взаимоотношения монголоидных вариантов имеют тот же характер, что по предыдущему признаку. Это относится не только к средним частотам, но и к показателям вариабельности: южные монголоиды чуть гомогеннее северных (V=33-37%).
Четвертая межпальцевая подушечка (IV)
Как и предыдущий, признак имеет сложное строение, включая истинные петли и петли, образованные добавочными трирадиусами. То же можно сказать и о типе распределения частот. Кривые приближаются к нормальным у монголоидов, европеоидов и австралоидов, имея нерегулярный характер у негроидов (Рис. 8.23). Максимальной средней частотой отличаются негроиды (62%), что связано не только с низким окончанием линии D, влекущим повышенный процент петель, но и с большой частотой добавочных межпальцевых трирадиусов на IV подушечке. По аналогичным причинам высокое значение признака обнаруживают австралоиды (63%). Минимальная величина характерна для европеоидов (45%), у которых отмечается наименьшая частота петель на IV подушечке (в связи с высокой частотой окончания линии D - в 9 и 11 полях), а частота второй составной части признака, т.е. узоров, образованных добавочными трирадиусами, невелика. Сходные величины признака имеют монголоиды и американоиды (50-51%). Абсолютная разница в среднерасовых значениях варьирует от 0 (негроиды и австралоиды) до 18% (европеоиды, с одной стороны, австралоиды и негроиды, с другой).
Наиболее гомогенны по данному признаку австралоиды, у которых коэффициент межгрупповой вариации составляет 14%, с ними сближаются европеоиды (18%). Еще более гетерогенны монголоиды и негроиды (V=24%), а наиболее разнородны индейцы Америки (V=32%). Среди европеоидов частоты признака распределяются практически по нормальному типу. Кривые группируются в две пары (Европа и Кавказ, с одной стороны, Средняя и Передняя Азия, с другой), внутри каждой из которых происходит почти полное совмещение. При этом во второй паре преобладает доля высоких значений признака. Те же взаимоотношения между вариантами европеоидов отмечены и по средней величине признака: в Европе и на Кавказе она составляет 43%, в Передней и Средней Азии - 48-49%.
211

Рисунок 8.23. Узорность IV межпальцевой подушечки. Внутрирасовое распределение популяционных значений признака (слева) и среднерасовые величины (справа).E - европеоиды: ЕЕ - европеоиды Европы, ЕК - европеоиды Кавказа, ЕПА - европеоиды Передней Азии, ЕСА - европеоиды Средней Азии; М - монголоиды: МСА - монголоиды Северной Азии, МЮА - монголоиды Южной Азии; Н - негроиды; Ав - австралоиды; Ам - американоиды. Figure 8.23. Intra-racial distribution of population values of interdigital pad IV patterns (left) and average racial values of the trait (right).E - Caucasoids: ЕЕ - Europe, ЕК - Caucasus, ЕПА - Western Asia, ЕСА - Central Asia; М - Mongoloids: МСА - Northern Asia, МЮА - Southern Asia; Н - African aboriginals; Ав - Australian aboriginals; Ам - American aboriginals.
212

Наиболее гомогенной группой по данному признаку являются переднеазиатские европеоиды (V=10%); более разнородны, причем в равной степени, кавказцы и среднеазиатские популяции (V=15%). Повышенный уровень межгрупповой изменчивости характеризует население Европы (V=19%).
Монголоидные варианты четко различаются по данному признаку, распределяющемуся по нормальному типу в обоих случаях. Обе кривые трансгрессируют сравнительно мало, причем южноазиатская кривая заметно сдвинута в область высоких значений. Средняя частота признака составляет в Северной Азии 47, в Южной - 70%. В Южной Азии признак варьирует вдвое меньше, чем в Северной, соответствующие коэффициенты межгрупповой вариации составляют 11 и 21%.
Частота радиальных пальцевых узоров (FRP)
Поскольку признак введен в исследовательскую практику недавно, лимиты его изменчивости не установлены, а средние значения определены лишь приблизительно и только для трех рас: европеоидной, негроидной и монголоидной. Наибольшая частота FRP характеризует монголоидные популяции (15.9%), несколько ниже величина признака у европеоидов (13.0%). Негроиды имеют наименьшую среди трех рас среднюю величину FRP (11.0%). Признак слабо варьирует внутри рас. Так, у европеоидов и монголоидов коэффициент межгрупповой вариации FRP составляет всего 8.9%, у негроидов - 6.1%. В последнем случае к величине коэффициента следует относиться с большой осторожностью, поскольку он вычислен на данных всего четырех выборок.
Выводы:
• в пределах больших рас популяционные значения ключевых признаков распределяются по одновершинному типу, близкому к нормальному (случаи отклонения от этого типа, очевидно, связаны с малым числом выборок). В этом можно увидеть косвенное доказательство полигенной природы ключевых признаков.
• расы отчетливо различаются не только по средним значениям признаков, но и по степени их вариабельности.
• неключевые признаки варьируют внутри расы значительно шире, чем ключевые (Рис. 8.24).
213

v %150 140 130 120 110 100
90 80 70 60 50 40 30 20 10 0
Dl10 Ic t Ну ДМТ Th/I A+T L W A/LA /W W /L MA M D C0 C , t ' t" t 0 II III IV
Рисунок 8.24. Внутрирасовые коэффициенты межгрупповой вариации признаков дерматоглифики (мужчины). Обозначения: а - ключевые признаки, б - пальцевые узоры, в - пальцевые индексы, г - главные ладонные линии, д - система осевых ладонных трирадиусов, е - узорность ладонных подушечек.Figure 8.24. Intra-racial inter-group coefficients of dermatoglyphic traits’ variation (males). Designation: a - key traits, б - finger patterns, в - finger indexes, г - palmar main lines, д - axial triradii, е - palmar pads patterns.
• Важным свойством ключевых признаков является соподчиненная изменчивость. Внутрирасовая популяционная вариабельность признаков подчиняется строгому правилу. Она растет в следующем порядке: Ic- Dl10 - t - Hy - ДМТ - Th/1. У европеоидов и монголоидов это правило не нарушается, а отдельные исключения, свойственные американоидам и австралоидам, могут быть связаны с малым числом выборок, приходящимся на «выпадающие» признаки. Единственное закономерное исключение составляют негроиды, у которых вариабельность ДМТ - признака, максимальная частота которого специфична именно для негроидной расы - резко сужена в ряду признаков и рас.
• Отмеченная последовательность возрастания внутрирасовой изменчивости связана с диагностической ценностью признаков: чем
214
В

меньше признак варьирует в пределах расы, тем более стойкой характеристикой расы он является и тем меньше вероятность возникновения случайных, не свойственных расе величин признака в популяциях, составляющих расу.
8.3. Европеоидно-монголоидный комплекс (ЕМК)
Напомним, что европеоидно-монголоидный комплекс - это условное содержание (в процентах евразиатской шкалы) монголоидного компонента в выборке, дополнительного по отношению к европеоидному компоненту. Для вычисления европеоидно-монголоидного комплекса используются пять ключевых признаков: дельтовый индекс, индекс Камминса,узорность гипотенара, частота проксимального трирадиуса ладони и суммарный процент добавочных межпальцевых трирадиусов. Техника вычисления, с учетом расовых градиентов признаков, описана в Главе 3.
Характерные для рас и их вариантов значения ЕМК приведены в следующей главе. В данном разделе рассмотрен популяционный уровень изменчивости. Величина ЕМК внутри расы распределяется практически по нормальному типу. Кривые распределения частот популяционных средних трансгрессируют даже при сопоставлении контрастных в расовом отношении вариантов (население Кавказа и Сибири), хотя в последнем случае и незначительно. Кривые распределения комплекса у европеоидов Кавказа, Восточной Европы и Средней Азии сильно трансгрессируют, при этом наиболее «европеоидны» кавказцы, а в наибольшей степени сближаются с монголоидами среднеазиатские популяции. Восточноевропейское население в этом отношении занимает промежуточное положение между указанными вариантами (Хить, 1986, с.31 -42).
Картографирование значений ЕМК на огромном пространстве от Пиренеев до Дальнего Востока и Индонезии выявляет два очага концентрации разных величин показателя: малых и очень малых на Западе, больших и сверхбольших на Востоке и Юго-Востоке (Рис. 8.25). Первый (западный или европейско-кавказский) охватывает всю Европу до Волго-Уралья, Кавказ, Западную Индию и часть Средней Азии. Второй очаг (восточный или азиатский) включает азиатскую часть России, Казахстан, Киргизию, Тибет, Корею, Вьетнам, Японию и Индонезию. Иначе говоря, величины показателя маркируют европеоидные и монголоидные популяции, очерчивая их ареалы.
215

Рисунок 8.25. Вариации европеоидно-монголоидного комплекса у мужских групп Евразии. 1 - очень малые, 2 - малые, 3 - средние, 4 - большие, 5 - очень большие, 6 - сверхбольшие.
216

Figure 8.25. Variation of Mongoloid complex in Eurasian groups (males): 1 - lowest, 2 - low, 3 - mean, 4 - large, 5 - largest.
217

Все население Европы, вплоть до Волго-Уралья и севера европейской части России, достаточно гомогенно и имеет малые и очень малые значения ЕМК. Вместе с тем, у финнов восточных провинций Финляндии, некоторых групп карел и северных групп русских ЕМК возрастает; особенно велик он у саамов Финляндии и в группах европейских (тундровых) ненцев.
Отметим, что принцип вычисления ЕМК, к сожалению, искусственно увеличивает долю «монголоидности» в популяциях южных европеоидов, имеющих повышенный дельтовый индекс. Но даже при этом условии аборигены Кавказа (не отраженного на карте из-за масштаба) отличаются небольшой величиной ЕМК, распределяющейся мозаично. Евразиатский минимум обнаружен у табасаранцев Дагестана (21.2 у мужчин); величина комплекса, как правило, уменьшается в горных кавказских группах. В Закавказье комплекс варьирует мало: по данным о мужчинах - в пределах 3839. В полном соответствии с данными расовой истории региона дифференциация аборигенного населения Кавказа не связывается с монголоидным элементом. В то же время отчетливо проявляются различия в доле южноевропеоидного компонента (ЮЕК), которая увеличивается в направлении с северо-запада на юго-восток: ЮЕК на Северном Кавказе составляет 60, в Закавказье - 59 на западе, 62 в центральных областях и 67 на востоке.
Подавляющее большинство популяций казахов, киргизов и каракалпаков характеризуется увеличенной или большой долей монголоидности, органически вписываясь в картину распределения ЕМК среди азиатских монголоидных популяций и особенно тесно сближаясь с населением Алтае-Саян, составляющим прямой переход к центрально-азиатским группам.
Для Сибири характерны высокие и очень высокие значения ЕМК. Ослабление монголоидности отчетливо проявляется у южных манси, у некоторых групп сибирских татар, селькупов, чулымцев, кетов, челканцев, юго-западных тувинцев и тувинцев-тоджинцев. В то же время популяции тундрового пояса - от саамов Финляндии и европейских ненцев на западе и до народов крайнего северо-востока России, а также айны имеют высокие и очень высокие величины показателя. Еще более высокие значения характерны для дунган, китайцев, вьетнамцев, корейцев, японцев и индонезийцев.
Оба очага концентрации разных величин ЕМК смыкаются в двух регионах, где процесс метисации резко усилен, что отразилось в заметном увеличении монголоидного компонента по сравнению с
218

западным очагом. Это, с одной стороны, Поволжье и Приуралье (марийцы, удмурты, часть башкир и татар, в том числе астраханские татары, а также чуваши, ногайцы, калмыки - последние, впрочем, являются поздними переселенцами в Европу). Другой регион - Средняя Азия, где картина отличается пестротой: малые и средние величины ЕМК (таджики, часть узбеков и западнопамирских народов) перемежаются с увеличенными и большими (некоторые группы узбеков, туркмен, уйгуров). Народы Западного Памира, не обозначенные на карте из-за ее масштаба, характеризуются средними и низкими значениями ЕМК, и лишь у горанцев он достигает уровня больших величин (58.7).
В среднеазиатских популяциях наблюдаются также следы третьего расового компонента - индо-австралоидного, - в результате влияния которого возникла комбинация признаков, получившая название среднеазиатского типа, не находящего аналогий в классификации европеоидных вариантов (Хить, 1973; 1975; 1983; 1991).
Обращают на себя внимание европеоидные популяции побережья северо-западной Индии (три группы брахманов и сонколи), обладающие малыми значениями ЕМК.
Таким образом, географическая изменчивость популяционных величин европеоидно-монголоидного комплекса в Евразии иллюстрирует характерные различия между двумя большими расами, выявляет области переходного типа, соответствующие очагам интенсивной метисации, и локальные варианты внутрирасовой дифференциации по комплексу признаков кожного рельефа. Все вышеописанное опирается на данные мужских выборок Евразии. Лимиты евразиатской шкалы вариаций ЕМК для мужчин составляют 21.2 и 79.9 (амплитуда 58.7), для женщин 26.9 и 80.5 (амплитуда 53.6). Для того чтобы сделать возможным сопоставление данных по мужчинам и женщинам, для каждого пола были рассчитаны значения ЕМК по общей (совмещенной) шкале вариаций показателя (Рис. 8.26). Величины этого стандартизованного монголоидного комплекса образуют два скопления - в области малых и высоких значений, соответствующих европеоидным и монголоидным группам. В европеоидном скоплении наименьшие величины относятся к населению Кавказа и Передней Азии (курдам, белуджам, персам, евреям, арабам), далее следуют группы зарубежной Европы и восточноевропейские выборки. Следующее звено образуют среднеазиатские народы,
219

причем узбеки и уйгуры занимают промежуточное положение между европеоидами и монголоидами, а таджики тяготеют к европеоидным группам. В переходной зоне располагаются также народы Поволжья и Приуралья, более «монголизированные» по сравнению со среднеазиатскими группами. При этом марийцы и башкиры оказываются ближе к монголоидным выборкам.
Рисунок 8.26. Вариации стандартизованного монголоидного комплекса в этнических группах Евразии. По оси абсцисс - мужчины, по оси ординат - женщины. Пунктирная диагональ соответствует полной корреляции данных. 1 - европеоиды Кавказа; 2 - европеоиды Европы; 3 - европеоиды Средней Азии; 4 - европеоиды Передней Азии; 5 - монголоиды Сибири; 6 - монголоиды Юго-Восточной Азии; 7 - народы Поволжья, Прикамья и Приуралья (татары, удмурты, чуваши, марийцы, башкиры); 8 - саамы Финляндии.Figure 8.26. Variation of standardized Mongoloid complex in Eurasian ethnic groups. The values for males are posited on abscissa axis, the values for females are posited on ordinate axis. Caucasoids: 1 - Caucasus, 2 - Europe, 3 - Central Asia, 4 - Western Asia; Mongoloids: 5 -Siberia, 6 - Southeastern Asia; 7 - ethnic groups from Volga-Urals regions; 8 - Sami from Finland.
220

Среди монголоидов сибирские группы, как правило, характеризуются менее выраженными расовыми чертами по сравнению с монголоидами Юго-Восточной Азии. Однако расовый комплекс отчетливо проявляется у народов Центральной Сибири, тундрового пояса и Дальнего Востока.
Минимальную величину монголоидного комплекса находим у чулымцев и селькупов, наиболее высокие значения - у вьетнамцев, японцев и китайцев. В последнем случае комплекс вычислен по четырем признакам, так как данные о ДМТ в публикации отсутствуют.
221

Глава 9. Характеристика и таксономия больших рас и их подразделений
В развитии науки о человеке в последние полвека наблюдается тенденция к обобщению фактических данных в эволюционном аспекте и построению различных таксономических схем и классификаций. Несколько таких попыток было сделано и в дерматоглифике (Thoma, 1974; Plato, 1976; Rothammer et al., 1977; Heet, Keita, 1979; Хить, Кейта, 1981; Хить, Долинова, 1990, 2002). Результаты оказались сопоставимыми лишь частично, что в значительной мере объясняется различиями в методических подходах авторов (разные программы исследований, объем и сравнимость данных, способы анализа).
Рассмотрим дерматоглифическую дифференциацию человечества как на уровне таксонов высокого ранга (большие или основные расы, региональные варианты европеоидов и монголоидов), так и на уровне популяций, представляющих собой выборки из этих рас. Специально подчеркнем, что в нашу задачу не входило создание собственно дерматоглифической классификации, основанной на методе числовой таксономии (хотя в разделе о популяциях будут показаны определенные возможности решения проблемы). Для этого необходимы гораздо более многочисленные и методически сравнимые материалы, чем те, которые оказались доступными. Но подойти к освещению проблемы соотношения дерматоглифических и расово-соматологических данных можно и по-другому, организовав дерматоглифический материал в рамках соматологической классификации. Заранее можно сказать, что специфика системы кожных гребешков как самостоятельного источника генетической информации неизбежно проявится в тех или иных результатах, так же, как и общая генетическая основа, определяющая расовые морфологические комплексы по обеим системам признаков у одного и того же объекта - человечества и его подразделений.
Основные результаты, описанные в данном разделе, опубликованы (Хить, Долинова, 2002). Анализ проводился в двух вариантах. В первом случае исследовался «классический» набор из пяти ключевых признаков (DlJ0, Ic, t, Hy, ДМТ); во втором привлекались также данные по узорности Th/I, отражающей прежде
222

всего малую методическую сопоставимость данных разных исследователей.
Многомерный анализ выполнялся двумя методами: межгрупповой шкалы и главных компонент. В первом случае обобщенное дерматоглифическое расстояние (ОДР) вычислялось, по мере необходимости, в процентах мужской, женской и совмещенной межгрупповой евразийской шкалы вариаций популяционных средних обоих полов. Матрицы ОДР подвергались кластерному анализу парногрупповым способом с взвешиванием. Различия параметров тестировались по t критерию Стьюдента. Для характеристики хорошо изученных рас (европеоидной и монголоидной) были привлечены все пригодные данные, вследствие чего число мужских и женских выборок оказалось различным. В остальных случаях использовались лишь материалы параллельного обследования обоих полов: это позволяет проконтролировать влияние выборочного эффекта на среднерасовые показатели и выявить основные тенденции, свойственные расе. Остальные сведения о материале содержатся в других разделах
9.1. Дифференциация больших рас
Материал был организован в рамках расовой классификации К. Куна (Coon, 1965), отчасти модифицированной Г.Л. Хить и Н.А. Долиновой (Хить, Долинова, 1990). Анализ базировался на среднерасовых показателях, полученных по вариационным рядам внутрирасовых популяционных значений признаков. Число наблюдений в пределах расы по разным признакам сильно варьирует, достигая максимума в отношении DlJ0. В общей сложности рассмотрены 1322 выборки (756 мужских и 566 женских), среди которых преобладают европеоиды и монголоиды.
В Главе 8 отмечалось, что вариационные ряды всех ключевых признаков имеют нормальный или близкий к нему тип и трансгрессируют у разных рас, однако степень трансгрессии и среднерасовые величины могут сильно различаться. Диагностическая роль ключевых признаков подтверждена статистически. Как правило, различия между расами значимы. В редких случаях разница недостоверна (между негроидами и американоидами по Ic и t; между негроидами и монголоидами по Ну; между монголоидами и австралоидами по ДМТ; между монголоидами и европеоидами по Th/1). Эти закономерности
223

выявлены на материале мужских групп, число которых в пределах европеоидной и монголоидной рас значительно выше, чем женских.
Сравнение величин признаков и их комбинаций выявляет своеобразие каждой расы (Табл.9.1, 9.2). Так, европеоидной расе в целом свойственно небольшое число дельт на пальцах, умеренное содержание t и ДМТ, повышенное значение Ic и узорности Hy (Рис.9.1, 9.2). Монголоидная раса характеризуется противоположной комбинацией признаков: повышенные значения FRP, DlJ0 и t сочетаются с умеренными или малыми величинами отдельных признаков. Негроиды в целом отличаются экстремально высокой частотой ДМТ при малых или очень малых величинах остальных признаков. Индейцы выделяются крайне малыми величинами всех признаков, за исключением умеренного DlJ0. Для австралоидов характерно сочетание высоких Dl10 и Th/I при средних значениях остальных признаков.
ЕЕ ЕК ЕПА ЕСА МСА МЮА
Рисунок 9.1. Комбинации признаков дерматоглифики в больших расах и субрасах (мужские группы). Центр окружности соответствует минимуму, величина радиуса - амплитуде евразийской межгрупповой шкалы популяционных средних, принятой за 100%. Обозначения рас даны в примечании к таблице 9.2. Аналогичные формы полигонов характеризуют женские группы.Figure 9.1. Combinations of key traits values in major races and subraces (male groups). Center of circle conforms to minimum value, radius size conforms to Eurasian inter-group range of population averages. Polygons of key traits’ values in female groups have similar shapes. Е - Caucasoids; М - Mongoloids; Н - African aboriginals, Ав - Australian aboriginals, Ам - American aboriginals. Sub-races: ЕЕ, ЕК, ЕПА, ЕСА - Caucasoids of Europe, Caucasus, Western Asia and Central Asia, respectively; МСА, МЮА - Mongoloids of North and South Asia, respectively.
224

? %30 6
Б
---- АмА м ___ 20
ЕСАА в ---- ___Ав ЕК >
МСА, ЕПА^Н ___
10
__ Н ЕЕ/,МЮА/
Е Мм — Е
. 0 .
МЮА
Рисунок 9.2. Частота истинных узоров на Th/I в больших расах (А) и в подразделениях монголоидной и европеоидной рас (Б).Figure 9.2. Frequency of thenar true patterns in major races (A) and in subraces of Mongoloids and Caucasoids (Б).
Эти комбинации (т.е. пропорциональные соотношения) признаков в пределах каждой расы явились результатом длительного развития составляющих расу популяций под влиянием сложных причин, прежде всего исторического характера, вызвавших миграции, смешения, изоляцию коллективов. По-видимому, они имеют очень древнее происхождение.
Иногда тип комбинации красноречиво свидетельствует о родстве, в то время как абсолютные различия в величинах признаков резко дифференцируют группы. Например, индейцы Америки по форме полигона напоминают редуцированный вариант австралоидной расы, а по величине ОДР в большей мере сближаются с монголоидами. В других случаях тип комбинации может маскировать истоки расовой группы: своеобразный тип сочетаний признаков в обобщенной среднеазиатской группе (это потомки древнейших европеоидных аборигенов региона) не позволяет отнести эту расовую группу к европеоидам, но по результатам таксономических расчетов она все же туда включается, однако, на достаточно низком уровне сходства.
Следует еще раз отметить принципиальное различие между исторически сложившимися комплексами признаков (историческими корреляциями) и биологическими(внутригрупповыми, на уровне индивидов) корреляциями. Вторые не дублируются первыми. Более того, оба типа комплексов не должны совпадать, так как одни формируются на биологической основе, а другие - под воздействием исторических причин.
225

Таблица 9.1. Ключевые признаки дерматоглифики и FRP у основных расовых групп человечества (невзвешенные средние вариационных рядов признаков)Table 9.1. Values of key traits and FRP in major racial groups of mankind (unweighted average of variational series).
№ Dl10 Ic t Hy ДМТ Th/I FRP
n x n x n x n x n x n x n x
1. s 427 13.06 388 8.64 403 65.1 362 32.2 355 20.6 362 9.9 13 13.0
? 275 12.42 243 8.45 220 58.2 232 35.4 219 19.1 232 9.5 13 11.1
2. S 214 14.39 188 7.66 151 76.5 178 21.5 152 12.7 178 9.9 13 15.9
? 176 13.93 170 7.51 141 71.5 165 25.7 146 10.9 165 8.2 13 15.8
3. S 49 12.78 29 7.23 30 56.0 53 23.4 47 39.0 53 15.1 4 11.0
? 49 12.14 29 7.03 30 49.7 53 23.3 47 34.5 53 15.0 4 11.4
4. S 39 15.76 28 8.00 10 63.1 23 22.9 26 13.6 23 18.8 - -
? 39 15.30 28 8.05 10 53.7 23 22.6 26 10.8 23 20.2 - -
5. S 27 13.54 22 7.20 6 51.3 9 12.6 8 10.4 9 25.6 - -
? 27 13.19 22 7.13 6 47.2 9 13.5 8 6.3 9 22.1 - -
Примечание.: n - число выборок; х - невзвешенная средняя; 1 - европеоиды; 2 - монголоиды; 3 - негроиды; 4 - австралоиды; 5 - американоиды.Источник: Хить, Долинова, 2002; FRP - частота встречаемости радиальных узоров; признак не включен в многомерный анализ, т.к. введен в практику исследования недавно (Широбоков, 2009б) и приводится здесь для полноты расовых характеристик.Группы: 1) Европа (без метисных популяций Приуралья, Поволжья и Севера), Кавказ, Передняя Азия, Средняя Азия (узбеки, таджики, западнопамирские народы, туркмены). В дальнейшем, с целью усиления присущих расе свойств, из суммарной группы европеоидов были исключены среднеазиатские популяции, метисные по происхождению и обладающие специфическими сочетаниями признаков (Хить, 1983; 1991). Характеристика анализируемой далее суммарной группы европеоидов Европы, Кавказа и Передней Азии (359 мужских и 222 женских выборок) выглядит так: Dl10 - 12.88 и 12.20; Ic - 8.66 и 8.47; t - 63.6 и 57.2; Ну - 32.8 и 35.6; ДМТ - 20.0 и 18.4; Th/1 - 9.6 и 9.1, соответственно; 2) Сибирь, Южная и Восточная Азия; 3) без мальгашей и популяций к северу от Сахары; 4) аборигены Австралии и Океании, 5) индейцы.Note: n - number of samples; x - unweighted average; 1 - Caucasoids; 2 - Mongoloids; 3 - African aboriginals, 4 - Australian aboriginals, 5 - American aboriginals.
226

Таблица 9.2. Показатели внутрирасовой межгрупповой изменчивости ключевых признаков дерматоглифики.Table 9.2. Parameters of intra-racial inter-group variability of key traits.
Расоваяпол
Dl10 Ic t Hy ДМТ Th/1группа s v s v s v s v s v s v
Е: ЕЕ м 0.65 5.2 0.58 4.4 7.46 11.5 5.68 17.8 5.58 31.7 3.38 36.5
ж 0.68 5.7 0.35 4.1 8.31 14.5 5.96 17.3 4.44 28.3 3.86 45.5EK м 0.75 5.6 0.40 4.6 7.20 11.8 5.61 16.2 8.45 34.9 4.19 42.6
ж 0.72 5.8 0.32 3.7 7.09 12.4 5.75 15.5 7.02 32.0 5.45 52.1ЕПА м 0.67 4.9 0.42 4.7 9.01 14.1 5.06 14.5 9.73 38.1 3.78 33.6
ж 0.70 5.3 0.31 3.6 7.04 13.0 7.75 19.8 10.13 41.0 3.60 41.6ЕСА м 0.66 4.7 0.41 4.8 7.57 10.5 5.25 18.0 6.31 26.3 3.99 34.4
ж 0.95 7.1 0.42 5.1 8.82 14.4 7.51 21.5 6.04 28.1 4.07 37.1Сумм. м 0.87 6.7 0.40 4.6 8.40 12.9 5.80 18.1 7.47 36.2 3.81 38.5
ж 0.93 7.5 0.36 4.2 8.37 14.4 6.46 18.2 6.70 35.1 4.42 46.4М: МСА м 0.94 6.3 0.39 4.9 8.77 11.3 6.14 26.8 5.74 44.0 4.98 45.7
ж 0.96 6.9 0.38 4.9 9.32 12.9 6.95 25.4 6.18 54.7 4.15 46.0МЮА м 0.70 4.8 0.49 6.9 7.35 10.5 6.95 39.0 4.69 43.2 3.83 53.6
ж 0.80 5.6 0.50 7.2 8.37 12.7 8.10 38.7 4.49 50.3 3.19 57.2Сумм. м 0.85 5.9 0.60 7.8 8.91 11.6 6.74 31.3 5.63 44.4 4.97 50.4
ж 0.94 6.7 0.54 7.1 9.45 13.2 7.75 30.1 5.98 54.9 4.20 51.2Н м 0.87 6.8 0.44 6.1 11.10 20.2 7.92 33.8 10.26 25.0 5.73 37.9
ж 0.90 7.4 0.34 4.8 11.90 23.9 5.96 25.6 5.98 17.3 8.36 55.8Ав м 1.32 8.4 0.76 9.5 4.06 6.4 7.82 34.2 7.85 57.6 10.84 57.6
ж 1.30 8.5 0.85 10.6 12.43 23.1 9.93 44.0 6.44 59.5 14.43 71.4Ам м 1.27 9.4 0.53 7.4 15.10 29.4 4.84 38.3 6.91 66.7 8.07 31.6
ж 1.63 12.4 0.52 7.3 12.46 26.4 5.01 37.2 3.94 62.1 6.31 28.5
Примечание. Расы (здесь и далее): - Е - европеоиды, М - монголоиды, Н - негроиды, Ав - австралоиды, Ам - американоиды. Субрасы: ЕЕ, ЕК, ЕПА, ЕСА - европеоиды Европы, Кавказа, Передней Азии и Средней Азии, соответственно; МСА, МЮА - монголоиды Северной и Южной Азии, соответственно.Note. Races (hereinafter): Е - Caucasoids; М - Mongoloids; Н - African aboriginals, Ав - Australian aboriginals, Ам - American aboriginals. Subraces: ЕЕ, ЕК, ЕПА, ЕСА - Caucasoids of Europe, Caucasus, Western Asia and Central Asia, respectively; МСА, МЮА - Mongoloids of North and South Asia, respectively.
227

Несмотря на неповторимость комбинаций признаков, свойственных каждой расе, монголоиды и австралоиды объединяются определенным сходством, имея высокие и очень высокие значения Dli0, пониженные величины Hy и ДМТ. Обе расы резко различаются по t и Th/1. Первый признак реже отмечен у австралоидов и чаще - у монголоидов, по второму картина противоположна. Американоиды по типу сочетаний сближаются с монголоидами и, особенно, австралоидами, отличаясь от них резким снижением значений всех признаков, кроме Th/1. По величине последнего признака американоиды занимают первое место среди всех рас. Негроиды и европеоиды не обнаруживают никакого сходства с остальными расами и, кроме того, предстают как почти полные антиподы. Признаки, имеющие среднюю или низкую встречаемость у европеоидов, сильно выражены у негроидов (ДМТ, Th/1), и наоборот (1с, t, Hy, FRP); один из важнейших диагностических признаков - дельтовый индекс, - однако, практически не разграничивает обе расы. Отметим также, что европеоиды и монголоиды максимально различаются по комбинациям и величинам всех ключевых признаков, кроме одного: средняя частота узорности на Th/I у них почти одинакова.
Кластеризация матриц ОДР по пяти ключевым признакам (без Th/1, выполненная трижды, для каждого пола в отдельности и обоих полов одновременно, обнаружила, прежде всего, максимальное сходство мужчин и женщин в пределах расы (Табл. 9.3; Рис. 9.3).
Во всех случаях анализа монголоиды и австралоиды наиболее сближены, к ним присоединяются американоиды. Негроиды и европеоиды последовательно включаются в этот кластер (каждая раса - на уровне, превышающем средние расстояния, что свидетельствует о самостоятельном ее положении в общей системе). Отметим, что дендрограмма, построенная по результатам кластерного анализа, полностью определяется типом последнего и отражает общую картину дифференциации; отдельные детали ее могут не соответствовать действительности. В первую очередь, это относится к положению негроидной и европеоидной рас.
Из Рисунка 9.3 следует, что европеоиды отмечены несколько большим своеобразием по сравнению с негроидами. Между тем, данные Таблицы 9.3 показывают, что негроидная раса несколько опережает европеоидную по степени обособленности от остальных рас (среднее ОДР для негроидов 24.8 у мужчин и 25.0 у женщин, для европеоидов 23.7 и 24.6, соответственно).
228

Рисунок 9.3. Результаты кластеризации матрицы ОДР (а) и компонентного анализа (б) больших рас с использованием пяти ключевых признаков. Здесь и далее черными кружками обозначены мужчины, белыми - женщины. Пунктирная вертикаль соответствует средней арифметической (х) матрицы попарных ОДР, штрихом выделены границы доверительного интервала на уровне 0.05. Обозначение расы, стоящее между кружками, относится к обоим полам. ОДР для каждого пола вычислялось на основе соответствующей амплитуды; при одновременном анализе данных обоих полов использована совмещенная амплитуда. Названия групп даны в примечании к Таблице 9.2.Figure 9.3. Results of clusterization of GDD matrix (a) and principal component analysis (б) for major races (5 key traits). The males are marked by black circles, the females are marked by white circles (hereinafter). Average (x) calculated from GDD matrix is posited on dotted vertical axis, confidence interval (p<0.05) is marked by dash. GDD for every sex was calculated by using of corresponding amplitude. Matched amplitude was used in the analysis of data on both sexes. Designations of groups are pointed in note for Table 9.2.
229

Таблица 9.3. Матрица ОДР между большими расами при одновременном анализе мужских и женских выборокTable 9.3. Matrix of GDD between major races (analysis of male and female samples)
Расы Е М Н Ав Ам Среднеепол м ж м ж м ж м ж м ж ОДР*
Ем - 7.2 23.0 19.9 24.0 27.4 19.7 22.6 27.9 30.1 23.7
ж 6.3 - 26.4 23.4 24.0 23.9 22.9 21.7 27.0 29.2 24.6
Мм 19.3 22.5 - 6.5 26.7 30.0 11.7 14.1 18.6 22.9 20.0
ж 17.4 20.0 6.4 - 23.7 27.0 13.7 14.8 15.6 20.0 21.3
Нм 23.4 23.6 25.4 23.9 - 7.3 27.4 26.1 21.1 23.3 24.8
ж 26.1 23.5 28.1 26.6 6.2 - 30.7 27.8 20.8 21.1 25.0
Авм 22.0 24.9 15.1 17.8 25.1 27.9 - 6.2 21.2 25.5 20.0
ж 25.2 24.8 18.0 19.6 24.8 26.3 6.0 - 15.7 20.0 21.4
Амм 32.9 32.5 25.0 23.5 25.0 23.8 21.8 16.3 - 5.1 22.2
ж 32.6 32.2 26.5 25.0 23.7 21.9 23.3 17.8 6.3 - 22.9
Справа - ОДР по пяти, слева - по шести ключевым признакам. Обобщенное дерматоглифическое расстояние вычислено с использованием евразийской популяционной шкалы, охватывающей вариации признаков у обоих полов. Знаком * отмечено среднее ОДР по 5 признакам для каждой расы от остальных.GDD calculated for 5 key traits is presented to the right of diagonal, one calculated for 6 key traits - to the left of diagonal. The generalized dermatoglyphic distance was computed using the Eurasian population range covering variation of traits values using the simultaneous data for both sexes.
По итогам компонентного анализа значимы две первые главные компоненты. На их долю приходится около 80% общей изменчивости (Табл. 9.4). Первая ГК по вкладу в изменчивость не слишком заметно (на 3-6%) превосходит вторую, т.е. факторы дифференциации в обоих случаях действуют почти равномерно. Основная нагрузка по первой ГК в убывающем порядке приходится на три признака: Ic, Hy, t, по второй ГК - на Dli0 и ДМТ. Таким образом, все рассматриваемые признаки активно участвуют в дифференциации рас, но Ic оказывается среди наиболее значимых, в отличие от Dlio. В пространстве двух первых ГК европеоиды,
230

негроиды и американоиды находятся в трех разных квадрантах, австралоиды и монголоиды - в четвертом квадранте (ем. Рис. 9.3). Отметим, что по значениям второй ГК американоиды сходны с монголоидами и австралоидами. Раздельный анализ полов дает ту же картину.
Таблица 9.4. Факторная структура. Коэффициенты корреляции ключевых признаков больших рас с ГК при одновременном анализе мужских и женских групп.Table 9.4. Correlation coefficients of key traits in major races and principal components using the simultaneous data for both sexes.
Признак n=5 n=6ГК I ГК II ГК III ГК I ГК II ГК III
Dl10 0.277 -0.841** 0.102 -0.163 0.880** -0.060Ic 0.900** 0.111 -0.384 0.741* 0.331 0.564t 0.707** -0.246 0.620 0.644* 0.515 -0.550Hy 0.780** 0.601 -0.059 0.938** -0.176 0.259ДМТ -0.197 0.824** 0.384 0.206 -0.826** -0.198Th/1 - - - -0.927** 0.106 0.297
Собственноечисло
2.034 1.821 0.694 2.769 1.874 0.818
Дисперсия,% 40.67 36.43 13.87 46.16 31.24 13.64
Примечание: n - число признаков; * P<0.05; ** P<0.01 Note: n - number of traits; * P<0.05; * * P<0.01
Обратимся к результатам анализа всех шести ключевых признаков, с привлечением Th/1. Этот последний признак может считаться специфически американоидно-австралоидным, так как его максимальные частоты (свыше 25%) характерны для индейцев, особенно центральноамериканских (до 40%), а повышенные - для австралоидных популяций. У негроидов величина признака ниже (около 15%), чем у указанных рас, но значительно выше, чем у монголоидов и европеоидов; обе последние расы практически неразличимы и имеют в среднем около 10% узорности (см. Табл.9.1).
Кластеризация матрицы ОДР по-прежнему обнаруживает максимальное сходство австралоидов с монголоидами, но в остальном дендрограммы мужчин и женщин различаются (Рис. 9.4).
231

Рисунок 9.4. Результаты кластерного анализа матрицы ОДР (а) и компонентного анализа (б) больших рас, выполненных с использованием шести ключевых признаков. Названия групп даны в примечании к таблице 9.2.Figure 9.4. Results of cluster analysis of GDD matrix (a) and principal component analysis (б) for major races (6 key traits). Designations of groups are pointed in note for Table 9.2.
При анализе данных мужчин и обоих полов, рассмотренных одновременно, достаточно неожиданным образом к австралоидомонголоидному кластеру присоединились европеоиды, заместив собой в схеме американоидов.
Практически самостоятельное положение заняли негроиды и американоиды, т.к. обе расы объединяются на уровне почти больших различий. Таким образом, единственный признак (Th/1) оказался способен перевесить влияние целого комплекса признаков и существенно изменил форму таксономического древа.
232

По итогам компонентного анализа, как и в предыдущем случае, значимы лишь первые две ГК, отражающие в сумме те же 77% общей изменчивости, но разница между обеими компонентами заметно увеличена - до 15-16%. Максимальная нагрузка приходится по первой компоненте на Ну, t, Ic и Th/1, по второй - на Dl10 и ДМТ. Иными словами, все шесть признаков связаны с дифференциацией рас, причем пять из них сохранили свою долю участия в ней, а Th/1 вошел в число наиболее активных признаков, обеспечив усиление изменчивости, отражаемой первой ГК. В пространстве двух первых ГК каждая из рас обособлена и располагается в отдельном квадранте. Лишь австралоиды и американоиды занимают общий квадрант, локализуясь, правда, в его противоположных областях. Австралоиды по второй ГК сближаются с монголоидами, а по первой занимают переходную позицию между ними и американоидами. Женщины и мужчины в пределах расы максимально сходны.
9.2. Cубрасовая дифференциация монголоидной и европеоидной рас
Многочисленные данные о популяциях были распределены по историко-географическим регионам; население каждого из них имеет специфические особенности как в строении лица и тела, так и в отношении других признаков, включая кожные узоры. В общей сложности рассмотрены 6 субрасовых вариантов: 4 европеоидных (Европа, Кавказ, Передняя Азия, Средняя Азия) и 2 монголоидных - Северная и Южная Азия (Табл. 9.1 и 9.5, Рис.9.1).
Европеоиды. С Севера на Юг, от Европы через Кавказ к Передней Азии возрастают величины Dl10, Ic, Ну и ДМТ при параллельном уменьшении t; узорность Th/1 увеличивается лишь в мужских группах. Важно отметить, что при этом сохраняется типичное для европеоидов сочетание признаков. Особняком стоит среднеазиатский вариант, который отличается от прочих комбинацией и величинами признаков, свидетельствующими о заметной доле инорасовой примеси, в основном монголоидного происхождения. Две черты среднеазиатской выборки, которые не могут объясняться этой примесью (повышение значений ДМТ и Th/I), по всей вероятности, связаны со следами индо-австралоидного влияния (Хить, 1983, 1991). Определяя доли расовых компонентов,
233
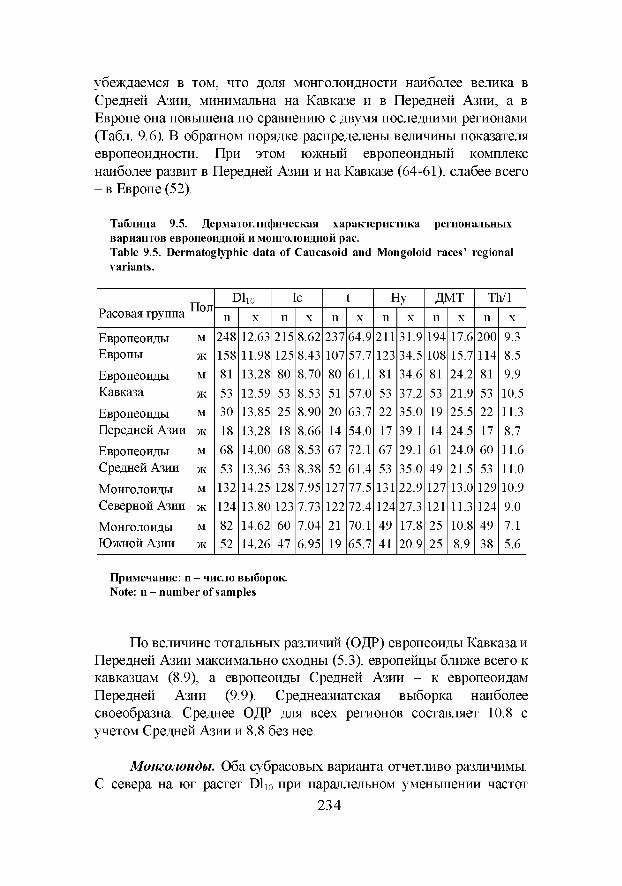
убеждаемся в том, что доля монголоидности наиболее велика в Средней Азии, минимальна на Кавказе и в Передней Азии, а в Европе она повышена по сравнению с двумя последними регионами (Табл. 9.6). В обратном порядке распределены величины показателя европеоидности. При этом южный европеоидный комплекс наиболее развит в Передней Азии и на Кавказе (64-61), слабее всего - в Европе (52).
Таблица 9.5. Дерматоглифическая характеристика региональных вариантов европеоидной и монголоидной рас.Table 9.5. Dermatoglyphic data of Caucasoid and Mongoloid races’ regional variants.
Расовая группа ПолDl10 Ic t Hy ДМТ Th/1
n x n x n x n x n x n x
Европеоиды м 248 12.63 215 8.62 237 64.9 211 31.9 194 17.6 200 9.3Европы ж 158 11.98 125 8.43 107 57.7 123 34.5 108 15.7 114 8.5
Европеоиды м 81 13.28 80 8.70 80 61.1 81 34.6 81 24.2 81 9.9Кавказа ж 53 12.59 53 8.53 51 57.0 53 37.2 53 21.9 53 10.5
Европеоиды м 30 13.85 25 8.90 20 63.7 22 35.0 19 25.5 22 11.3Передней Азии ж 18 13.28 18 8.66 14 54.0 17 39.1 14 24.5 17 8.7
Европеоиды м 68 14.00 68 8.53 67 72.1 67 29.1 61 24.0 60 11.6Средней Азии ж 53 13.36 53 8.38 52 61.4 53 35.0 49 21.5 53 11.0
Монголоиды м 132 14.25 128 7.95 127 77.5 131 22.9 127 13.0 129 10.9Северной Азии ж 124 13.80 123 7.73 122 72.4 124 27.3 121 11.3 124 9.0
Монголоиды м 82 14.62 60 7.04 21 70.1 49 17.8 25 10.8 49 7.1Южной Азии ж 52 14.26 47 6.95 19 65.7 41 20.9 25 8.9 38 5.6
Примечание: n - число выборок.Note: n - number of samples
По величине тотальных различий (ОДР) европеоиды Кавказа и Передней Азии максимально сходны (5.3), европейцы ближе всего к кавказцам (8.9), а европеоиды Средней Азии - к европеоидам Передней Азии (9.9). Среднеазиатская выборка наиболее своеобразна. Среднее ОДР для всех регионов составляет 10.8 с учетом Средней Азии и 8.8 без нее.
Монголоиды. Оба субрасовых варианта отчетливо различимы. С севера на юг растет DlJ0 при параллельном уменьшении частот
234

остальных ключевых признаков. Основная часть этих отличий вполне объяснима заметной долей европеоидной примеси в составе сибирских монголоидов, в основном живущих в Западной и Южной Сибири, где процесс метисации с древнейших времен был весьма интенсивным (уменьшение Dl10, увеличение Ic, ДМТ, Ну). Однако высокий процент t - стойкая черта северных монголоидов и одновременно маркер монголоидной расы в целом - непонятным образом сохранился в этой ситуации; видимо, у древних аборигенов Сибири он был еще выше.
Таблица 9.6. Расовые комплексы в мужских группах субрасовых вариантов европеоидной и монголоидной рас.Table 9.6. Racial complexes in male groups of Caucasoid and Mongoloid races’ regional variants.
Субрасовые варианты ЕМК МЕК Южный комплексЕвропеоиды: Европа 42.0 58.0 51.8
Кавказ 38.3 61.7 60.6Передняя Азия 39.4 60.6 63.9Средняя Азия 49.3 50.7 55.2
Монголоиды: Северная Азия 64.2 35.8 58.0Южная Азия 72.1 27.9 71.6
Комплексы: ЕМК - европеоидно-монголоидный (условный показатель «монголоидности»); МЕК - монголоидно-европеоидный (показатель «европеоидности», дополнительный по отношению к ЕМК); оба рассчитываются исходя из наличия двух указанных расовых компонентов. Южный комплекс допускает наличие лишь одного из компонентов и выявляет выраженность южного компонента в пределах каждой расы. Техника вычислений, расовые и географические градиенты, а также издержки метода описаны в Главе 3.Complexes: ЕМК - conditional score of “Mongoloid quantity”; МЕК - conditional score of “Mongoloid quantity” (complementary to ЕМК); «Южный комплекс» (Southern complex) expresses “southern quantity” within the bounds of every race. Computing technique, racial and territorial gradients, restrictions of the method are described in Chapter 3.
В то же время данный признак у южных монголоидов встречается заметно реже, отражая общий древний компонент, лежащий в основе южных монголоидов и австралоидов. Австралоидный компонент проявляется у южных монголоидов не только в уменьшении t, но и в увеличении Dl10. И монголоидность, и
235

южномонголоидные черты в южноазиатской группе резко усилены (достигая примерно 72% евразиатской шкалы на каждый признак), по сравнению с сибирской (см. Табл. 9.6). ОДР между вариантами монголоидов составляет 13.7 и 15.7 для мужчин и женщин, соответственно, что заметно превышает соответствующие показатели у подразделений европеоидной расы.
Субварианты монголоидов и европеоидов в таксономии больших рас
Рассмотреть региональные варианты всех рас оказалось невозможным - как из-за малой численности выборок (американоиды, австралоиды), так и ввиду отсутствия критерия разбивки (негроиды). Тем не менее, субварианты монголоидов и европеоидов как наиболее изученных рас были рассмотрены на фоне остальных (Рис. 9.5). Выяснилось, что в принципе серьезных изменений в таксономии рас не возникло.
На дендрограмме ОДР по-прежнему наиболее обособлены европеоиды и негроиды; монголоиды и австралоиды образуют кластер, с которым сливаются американоиды. На мужской дендрограмме уточнилось положение австралоидов, разделивших северных и южных монголоидов в их субкластере, однако на женской и общей для обоих полов дендрограммах австралоиды заняли традиционное положение, примкнув к монголоидам. Варианты европеоидов всюду образуют компактные кластеры, в пределах которых по данным раздельного анализа полов наиболее своеобразны европейцы, в отличие от тесно сблизившихся южных европеоидов (Кавказа и Передней Азии), к которым примкнули европеоиды Средней Азии.
Таким образом, субрасы европеоидов с южной основой объединяются сходством, о чем свидетельствует величина ОДР. Однако при совместном анализе обоих полов ярко проявляется первостепенная роль половых различий в дифференциации вариантов европеоидной расы: прежде всего, объединяются половые группы, и лишь затем выделяется европеоидный кластер. Наблюдаются также два исключения: среднеазиатские женщины, оказавшиеся в «мужском» кластере ввиду максимального сходства с мужчинами Кавказа, и европейские мужчины.
236

Рисунок 9.5. Варианты европеоидной и монголоидной рас в системе остальных рас человечества (при анализе данных по пяти ключевым признакам). Названия групп даны в примечании к Табл. 9.2. В пространстве двух главных компонент обведены скопления, выделенные кластеризацией данных главных компонент.Figure 9.5. Variants of Caucasoid and Mongoloid races relative to other races of mankind (analysis of data on 5 key traits). Designations of groups are pointed in note for Table 9.2. The clouds of groups are marked by clusterization of data on first two principal components.
В последнем случае мужская и женская половины выборки сближены в наибольшей степени; это же явление отмечено для всех остальных рас, в том числе монголоидных вариантов. Существование различий в расовом составе мужчин и женщин и более весомый южноевропеоидный компонент у вторых были
237

констатированы при изучении туркмен (Долинова, Хить, 1989). Как видим, этот феномен характеризует всю среднеазиатскую группу европеоидов.
По результатам компонентного анализа австралоиды систематически располагаются между северными и южными монголоидами. Европеоиды образуют отдельное скопление. В нем среднеазиатский вариант уклоняется к северным монголоидам, что связано с увеличением монголоидного компонента в Средней Азии. Однако при суммарном анализе данных обоих полов среднеазиатские женщины отдаляются от остальных женских выборок, приближаясь к мужским группам европеоидов Кавказа и Европы.
Американоиды и негроиды изолированы. Статистически, при кластеризации компонентных нагрузок, выделены следующие кластеры: 1. негроиды; 2. европеоиды; 3. монголоиды, австралоиды и американоиды (см. Рис. 9.5). С привлечением шестого признака (Th/1) картина меняется существенным образом (Рис.9.6). Это касается как дендрограммы по ОДР, так и расположения групп в пространстве двух первых главных компонент. Статистически, по нагрузкам на компоненты выделяются кластеры: 1. негроиды; 2. европеоиды и монголоиды; 3. австралоиды и американоиды.
Следовательно, использование Th/1 в многомерном анализе приводит к объединению близких по его частоте монголоидов и европеоидов, резко различающихся по сочетанию значений остальных ключевых признаков.
Дифференциация популяций
На этой стадии анализа необходимо было резко ограничить количество рассматриваемых групп: все расы должны бытьпредставлены одинаковым числом выборок. Лишь в этом случае исключается вероятность деформации таксономического пространства. Ориентируясь на число оказавшихся доступными выборок с полным набором данных о ключевых признаках у индейцев (как наименее исследованной расы), получаем заданный максимум в 5 групп на расу. С учетом пола общее число выборок составило 50. В методическом отношении материалы сравнимы, т.к. свыше двух третей данных принадлежат Г.Л. Хить и Н.А. Долиновой, остальные - исследователям, применяющим сходные приемы описания отпечатков (Х Бреме, М. Поспишилу, Б. Кейта).
238

Рисунок 9.6. Варианты европеоидной и монголоидной рас в системе остальных рас человечества (при анализе данных по шести ключевым признакам). Названия групп даны в примечании к Таблице 9.2. В пространстве двух главных компонент обведены скопления, выделенные кластеризацией данных главных компонент.Figure 9.6. Variants of Caucasoid and Mongoloid races relative to other races of mankind (analysis of data on 6 key traits). Designations of groups are pointed in note for Table 9.2. The clouds of groups are marked by clusterization of data on first two principal components.
239

16/ТМ 7/ТМ 8/ТЧ 1 9 /Т \ 2 0 /Т \
Рисунок 9.7. Круговые полигоны признаков дерматоглифики 50 популяций ойкумены. Радиус окружности равен евразийской амплитуде популяционных средних мужских групп. Нумерация групп как в таблице 9.7. Расовые группы: 1-10 - европеоиды, 11-20 - монголоиды, 21-30 - австралоиды, 31-40 - американоиды, 41-50 - негроиды. Деформация полигонов 35-38 обусловлена выходом значений некоторых признаков за минимальные лимиты амплитуды.Figure 9.7. Circle polygons of key traits for 50 groups from populated universe. Radius size conforms to Eurasian amplitude of population male averages. Groups are numbered as in Table 9.7. Racial groups: 1-10 - Caucasoids, 11-20 - Mongoloids, 21-30 - Australian aboriginals, 31-40 - American aboriginals, 41-50 - African aboriginals.
240

Материал охватывает северные и южные группы европеоидов, монголоидов и американоидов. В отношении остальных рас пришлось ограничиться негроидами Западной и Южной Африки, австралийцами, папуасами Новой Гвинеи и полинезийцами. Некоторые, в основном индейские, выборки малы по объему (атапаски, трио, уастеки), что отразилось на результатах анализаХарактеристика групп содержится в Таблице 9.7 и Рисунке 9.7. Отметим, что среднерасовые значения признаков, полученные по данным о популяциях, приведенные в таблице, в подавляющем большинстве случаев близки к характеристикам больших рас, основанным на громадном по объему материале и приведенным в Таблице 9.1. Что касается комбинативной изменчивости, то полигоны дают возможность оценить не только тип сочетаний, но и величину признаков. Как правило, одноименные мужские и женские выборки сходны по комбинациям признаков.
Различия обычно имеют направленный характер и связаны с феноменом полового диморфизма, основные закономерности которого являются общими для всех рас (см. Главу 6). Величины всех ключевых признаков в норме у женщин меньше, чем у мужчин, за исключением узорности гипотенара. Последний признак сильнее развит именно у женщин (но у негроидов и австралоидов - у мужчин).
На уровне выборок отступления от правил встречаются чаще, однако в данном случае указанные закономерности прослеживаются вполне отчетливо, особенно у популяций монголоидной и европеоидной рас. В группе индейцев значения признаков иногда выходят за пределы установленной евразийской шкалы, что приводит к деформации полигонов. В целом же полигоны популяций представляют собой вариации среднерасовых типов (см. Рис. 9.1).
Не иллюстрируя, кратко опишем результаты кластерного анализа матриц межпопуляционных ОДР по пяти признакам. При одновременном учете данных о мужчинах и женщинах выявлены три основных кластера: 1 - европеоиды (все группы) и большинство австралоидов (включая австралийцев), образующих субкластер; 2 - монголоиды, американоиды и австралоиды (сюда входят все монголоиды, большинство индейцев, образующих субкластер, и единичные группы австралоидов); 3 - негроиды (исключительно негроидные выборки, образующие компактное и наиболее изолированное скопление).
241

Таблица 9.7. Дерматоглифическая характеристика 50 групп населения ойкуменыTable 9.7. Dermatoglyphic data o f 50 groups from populated universe.
Раса Группа N, пол Dlio Ic t Hy ДМТ Th/1Е 1.Русские 2939 м 12.60 8.56 63.1 30.8 17.4 8.3В 2. Русские 2468 ж 11.79 8.44 61.5 28.2 12.8 7.7Р 3. Финны 1532 м 11.91 8.68 65.8 26.7 13.8 8.1О 4. Финны 122 ж 11.91 8.21 58.2 31.6 12.3 6.1П 5. Армяне 413 м 13.65 9.09 58.8 32.4 26.6 11.8Е 6. Армяне 403 ж 12.76 8.53 52.5 38.2 22.4 5.5О 7. Азербайджанцы 343 м 14.02 8.87 64.7 32.4 26.4 14.0И 8. Азербайджанцы 356 ж 13.09 8.64 58.9 38.2 24.2 7.2Д 9. Чехи 1610 м 12.70 8.90 63.4 32.3 23.7 13.0Ы 10. Чехи 608 ж 12.03 8.50 60.8 34.9 24.0 11.0
М 11.Эвенки 298 м 15.30 8.07 70.4 25.9 8.4 19.1О 12.Эвенки 380 ж 14.97 7.80 63.3 24.2 9.6 12.7Н 13.Буряты 365 м 15.44 7.81 80.3 19.2 9.4 10.4Г 14.Буряты 327 ж 14.74 7.44 76.4 20.6 9.8 11.3О 15.Чукчи 355 м 14.11 7.60 72.8 20.1 8.2 7.9Л 16.Чукчи 405 ж 13.84 7.36 66.6 29.1 5.5 6.2О 17.Корейцы 257 м 14.50 6.62 68.8 26.3 10.6 9.3И 18.Корейцы 229 ж 13.82 6.78 63.4 28.4 7.8 7.6Д 19.Кхмеры 80 м 14.28 6.64 73.8 14.4 16.2 1.2Ы 20.Кхмеры 71 ж 13.93 6.96 73.9 19.7 19.7 4.2
А 21.Австралийцы 174 м 15.69 8.77 64.8 25.0 13.4 11.9В 22.Австралийцы 141 ж 15.33 8.92 56.7 25.9 14.9 11.0С 23.Бунди 103 м 14.27 7.88 61.6 34.5 18.0 35.9Т 24.Бунди 91 ж 14.43 8.10 61.0 30.8 24.2 34.7Р 25.Амеле 89 м 15.70 8.22 62.4 23.6 14.6 19.1А 26.Амеле 26 ж 15.86 8.56 61.2 22.9 11.7 30.8Л 27.Хаганаи 79 м 14.32 8.15 65.9 29.7 3.8 7.7О 28.Хаганаи 46 ж 14.48 8.26 53.3 26.7 12.8 26.7И 29.Полинезийцы 56 м 15.41 6.43 58.9 25.0 3.6 12.5ДЫ
ЗО.Полинезийцы 53 ж 15.83 6.77 43.4 30.2 2.8 9.4
Примечание см. далее
242
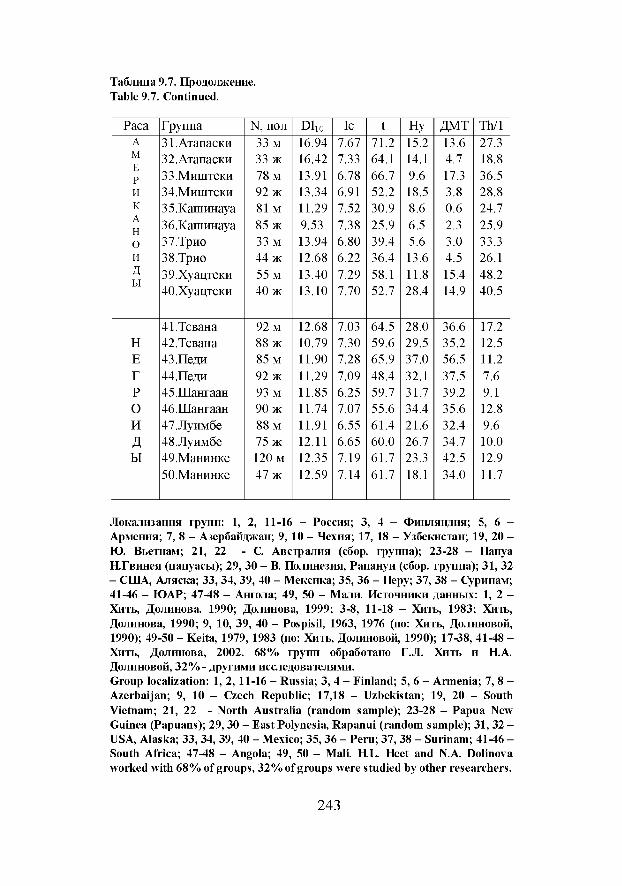
Таблица 9.7. Продолжение. Table 9.7. Continued.
Раса Группа N, пол Dl10 Ic t Hy ДМТ Th/1А 31.Атапаски 33 м 16.94 7.67 71.2 15.2 13.6 27.3МЕР
32.Атапаски 33 ж 16.42 7.33 64.1 14.1 4.7 18.8ЗЗ.Миштеки 78 м 13.91 6.78 66.7 9.6 17.3 36.5
И 34.Миштеки 92 ж 13.34 6.91 52.2 18.5 3.8 28.8К 35.Кашинауа 81 м 11.29 7.52 30.9 8.6 0.6 24.7АН Зб.Кашинауа 85 ж 9.53 7.38 25.9 6.5 2.3 25.9О 37.Трио 33 м 13.94 6.80 39.4 5.6 3.0 33.3И 38.Трио 44 ж 12.68 6.22 36.4 13.6 4.5 26.1ДЫ 39.Хуацтеки 55 м 13.40 7.29 58.1 11.8 15.4 48.2
40.Хуацтеки 40 ж 13.10 7.70 52.7 28.4 14.9 40.5
41.Тсвана 92 м 12.68 7.03 64.5 28.0 36.6 17.2Н 42.Тсвана 88 ж 10.79 7.30 59.6 29.5 35.2 12.5Е 43.Педи 85 м 11.90 7.28 65.9 37.0 56.5 11.2Г 44.Педи 92 ж 11.29 7.09 48.4 32.1 37.5 7.6Р 45.Шангаан 93 м 11.85 6.25 59.7 31.7 39.2 9.1О 46.Шангаан 90 ж 11.74 7.07 55.6 34.4 35.6 12.8И 47.Луимбе 88 м 11.91 6.55 61.4 21.6 32.4 9.6Д 48.Луимбе 75 ж 12.11 6.65 60.0 26.7 34.7 10.0Ы 49.Манинке 120 м 12.35 7.19 61.7 23.3 42.5 12.9
50.Манинке 47 ж 12.59 7.14 61.7 18.1 34.0 11.7
Локализация групп: 1, 2, 11-16 - Россия; 3, 4 - Финляндия; 5, 6 - Армения; 7, 8 - Азербайджан; 9, 10 - Чехия; 17, 18 - Узбекистан; 19, 20 - Ю. Вьетнам; 21, 22 - С. Австралия (сбор. группа); 23-28 - ПапуаН.Гвинея (папуасы); 29, 30 - В. Полинезия, Рапануи (сбор. группа); 31, 32 - США, Аляска; 33, 34, 39, 40 - Мексика; 35, 36 - Перу; 37, 38 - Суринам; 41-46 - ЮАР; 47-48 - Ангола; 49, 50 - Мали. Источники данных: 1, 2 - Хить, Долинова. 1990; Долинова, 1999; 3-8, 11-18 - Хить, 1983; Хить, Долинова, 1990; 9, 10, 39, 40 - Pospisil, 1963, 1976 (по: Хить, Долиновой, 1990); 49-50 - Keita, 1979, 1983 (по: Хить, Долиновой, 1990); 17-38, 41-48 - Хить, Долинова, 2002. 68% групп обработано Г.Л. Хить и Н.А. Долиновой, 32% - другими исследователями.Group localization: 1, 2, 11-16 - Russia; 3, 4 - Finland; 5, 6 - Armenia; 7, 8 - Azerbaijan; 9, 10 - Czech Republic; 17,18 - Uzbekistan; 19, 20 - South Vietnam; 21, 22 - North Australia (random sample); 23-28 - Papua New Guinea (Papuans); 29, 30 - East Polynesia, Rapanui (random sample); 31, 32 - USA, Alaska; 33, 34, 39, 40 - Mexico; 35, 36 - Peru; 37, 38 - Surinam; 41-46 - South Africa; 47-48 - Angola; 49, 50 - Mali. H.L. Heet and N.A. Dolinova worked with 68% of groups, 32% of groups were studied by other researchers.
243
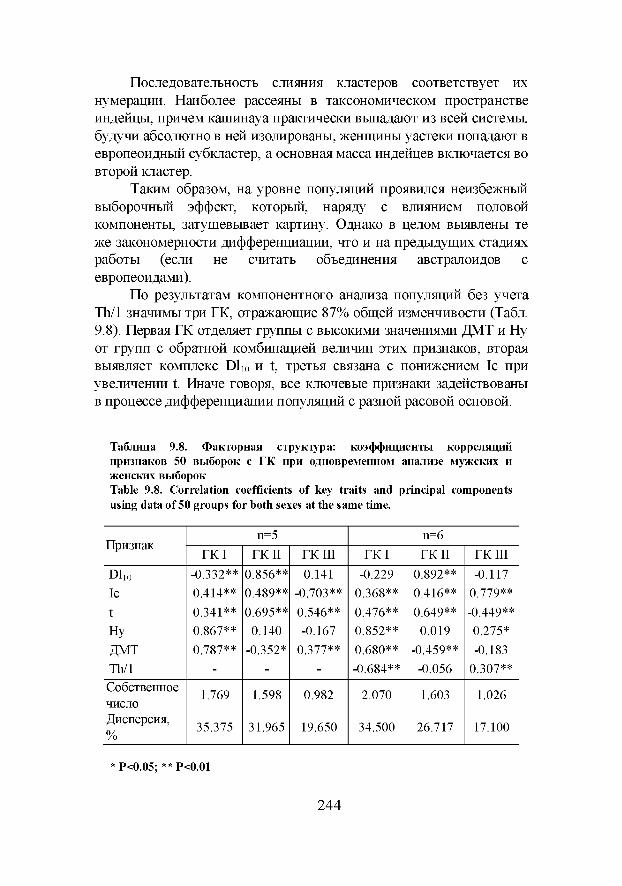
Последовательность слияния кластеров соответствует их нумерации. Наиболее рассеяны в таксономическом пространстве индейцы, причем кашинауа практически выпадают из всей системы, будучи абсолютно в ней изолированы, женщины уастеки попадают в европеоидный субкластер, а основная масса индейцев включается во второй кластер.
Таким образом, на уровне популяций проявился неизбежный выборочный эффект, который, наряду с влиянием половой компоненты, затушевывает картину. Однако в целом выявлены те же закономерности дифференциации, что и на предыдущих стадиях работы (если не считать объединения австралоидов с европеоидами).
По результатам компонентного анализа популяций без учета Th/1 значимы три ГК, отражающие 87% общей изменчивости (Табл. 9.8). Первая ГК отделяет группы с высокими значениями ДМТ и Ну от групп с обратной комбинацией величин этих признаков, вторая выявляет комплекс Dl10 и t, третья связана с понижением Ic при увеличении t. Иначе говоря, все ключевые признаки задействованы в процессе дифференциации популяций с разной расовой основой.
Таблица 9.8. Факторная структура: коэффициенты корреляцийпризнаков 50 выборок с ГК при одновременном анализе мужских и женских выборокTable 9.8. Correlation coefficients of key traits and principal components using data of 50 groups for both sexes at the same time.
Признакn=5 n=6
ГК I ГК II ГК III ГК I ГК II ГК III
Dl10 -0.332** 0.856** 0.141 -0.229 0.892** -0.117Ic 0.414** 0.489** -0.703** 0.368** 0.416** 0.779**t 0.341** 0.695** 0.546** 0.476** 0.649** -0.449**Hy 0.867** 0.140 -0.167 0.852** 0.019 0.275*ДМТ 0.787** -0.352* 0.377** 0.680** -0.459** -0.183Th/1 - - - -0.684** -0.056 0.307**Собственноечисло
1.769 1.598 0.982 2.070 1.603 1.026
Дисперсия,%
35.375 31.965 19.650 34.500 26.717 17.100
* P<0.05; ** P<0.01
244

Отметим, что столь популярный у специалистов Dl10, привлекаемый даже для выделения двух основных расовых стволов (Алексеев, 1974; Чебоксаров, Чебоксарова, 1985), по первой ГК оказывается наименее значимым признаком, а по третьей вообще не имеет никакого влияния на дифференциацию, в отличие от Ic.
В пространстве ГК самые компактные и изолированные скопления популяций характеризуют негроидов (Рис.9.8); то же относится к европеоидам, с которыми, правда, вплотную сближаются новогвинейские папуасы бунди. Монголоиды, австралоиды и индейцы образуют следующее скопление. Трио и кашинауа отдалены от остальных индейцев.
В генерализованном виде эта картина представлена Рисунком 9.9, на котором обведены ареалы больших рас в соответствии с расовой принадлежностью популяций и обозначены расовые центроиды. Здесь по-прежнему, как и на генерализованном уровне (большие расы и их субварианты), негроиды и европеоиды наиболее обособлены; третье скопление образуют австралоиды, монголоиды и американоиды, ареалы которых перекрываются, причем особенно сильно трансгрессируют австралоиды и монголоиды.
Особо отметим беспрецедентную гетерогенность индейцев, чей ареал исключительно велик и исчерпывает половину амплитуды по первой и всю амплитуду - по второй ГК. Что касается расовых центроидов, то по ним наблюдается полное совпадение с результатами кластерного анализа матриц ОДР больших рас, характеристики которых базируются на огромном материале (см. Таблицу 9.1).
Кластерным анализом данных трех первых ГК были выделены четыре основных кластера (Рис. 9.8). Первый объединяет все европеоидные выборки и единичные, в основном женские группы других рас (уастеков, австралийцев, новогвинейских папуасов бунди и хагахаи), а также мужскую выборку папуасов бунди. Второй кластер включает всех монголоидов, большинство австралоидов и половину индейцев. Третий кластер, резко расходящийся с двумя первыми, состоят исключительно из негроидных выборок. Абсолютно изолированы во всей системе индейцы двух племен - кашинауа и трио, образующие отдельное «суперамериканоидное» скопление (четвертый кластер). Во втором кластере выделяются два субкластера, условно обозначенные как северный (монголоиды Сибири, атапаски, австралийцы и часть папуасов) и южный (южные популяции монголоидов, миштеки и уастеки).
245

Рисунок 9.8. Результаты компонентного анализа 50 популяций на основе пяти ключевых признаков (без Th/1). Заливкой обозначены мужские группы, контуром - женские. Скопления популяций выделены по результатам статистического анализа, выполненного на основе пяти ключевых признаков (кластеризацией данных трех главных компонент). В монголоидно-австралоидно-американоидном кластере чертой разделены субкластеры, выделенные тем же способом. Заливкой обозначены мужские группы, контуром - женские.Figure 9.8. Results of principal component analysis of data on 50 groups (5 key traits, nonmetering Th/I). Male groups are marked by a flow, female groups are marked by a outline. Results of clusterization of data based on the analysis of three principal components (5 key traits). Male groups are marked by a flow, female groups are marked by an outline.
Таким образом, анализ взаимного расположения популяций, выполненный по пяти ключевым признакам дважды, притом вне рамок расовой принадлежности (кластеризацией матрицы тотального показателя различий, или ОДР, и кластеризацией координат групп в пространстве двух ГК), привел к выделению одних и тех же скоплений, объединивших выборки в соответствии с их расовой принадлежностью. Отдельные случаи несовпадений объясняются как выборочным эффектом, так и большей мощностью факторного анализа, учитывающего векторы дифференциации.
246

Рисунок 9.9. Ареалы больших рас (в рамках соматологической расовой классификации) и расовые центроиды по данным компонентного анализа 50 популяций с использованием пяти ключевых признаков (мужчины).Figure 9.9. Areals of major races (within the bounds of racial somatological classification) and racial centroids computed on results of principal component analysis for 50 groups (5 key traits, males).
Обратимся к факторному анализу всех шести ключевых признаков (Рис. 9.10). Негроидные и европеоидные популяции по- прежнему образуют отдельные скопления в области положительных значений первой ГК. Основная часть монголоидов, перемежаясь австралоидными выборками, приблизилась к европеоидам, разместившись в переходной зоне первой ГК. Остальные австралоиды и индейцы заняли половину поля первой ГК, то есть расширили занимаемое пространство. Очертив расовые ареалы в плоскости двух первых ГК и обозначив расовые центроиды, получаем практически ту же картину, что и по результатам анализа пяти признаков, с той разницей, что американоиды обрели отдельный ареал, не пересекающийся с областями, занятыми монголоидами и австралоидами (Рис. 9.11).
247

X Н О Е А М V А в □ А м
ГК II (2 7 % )
3
3 CD <9=----------------
□ с
i #
*$
-3
ГК I(3 5 % )
Рисунок 9.10. Результаты компонентного анализа 50 популяций по шести ключевым признакам.Figure 9.10. Results of principal component analysis of data on 50 groups (6 key traits).
Однако применением не типологического, а статистического критерия (кластерного анализа по данным всех трех значимых компонент, отражающих в сумме 78% изменчивости) получена иная схема дивергенции. Выделяются пять кластеров (Рис. 9.12).
После слияния двух первых (европеоидного и австралоидного) произошло последовательное подключение монголоидов, негроидов и американоидов. Американоиды максимально отдалились от прочих вариантов.
Следует отметить, что европеоидный, негроидный и американоидный кластеры не включают популяции иного происхождения, а в остальных случаях группы иной расовой принадлежности присутствуют в кластерах наряду с нуклеарными популяциями.
248

Рисунок 9.11. Расовые ареалы и расовые центроиды по результатам компонентного анализа 50 популяций с использованием шести ключевых признаков.Figure 9.11. Areals of major races (within the bounds of racial somatological classification) and racial centroids computed on results of principal component analysis for 50 groups (6 key traits).
Так, в монголоидном кластере находятся также полинезийцы и атапаски, а в австралоидном - эвенки и женщины уастеки. Кроме того, американоидный кластер неполон по составу и включает мужчин и женщин индейцев трио, кашинауа, миштеков и мужскую группу уастеков. Следовательно, при учете Th/1 на уровне популяций компонентный анализ выделяет в целом те же скопления, что и без данного признака, но детализирует картину и подчеркивает обособленность основной части американоидов, размещая остальные группы индейцев среди монголоидов и австралоидов.
Субрасовые варианты европеоидной и монголоидной рас исследовались также на уровне популяций, с учетом их зонального расселения (80 выборок северного и южного происхождения).
249
Рисунок 9.12. Дифференциация 50 групп по шести ключевым признакам (результаты кластерного анализа данных трех первых ГК). Обозначения соответствуют принятым в Таблице 9.7.Figure 9.12. Results of clusterization of data based on the analysis of three principal components for 50 groups (6 key traits). Groups are numbered as in Table 9.7.
250

В результате статистического анализа обнаружено, что расовый фактор играет главную роль, разделяя популяции в соответствии с их расовой принадлежностью, а географический - второстепенную.
Различия между северными и южными вариантами в пределах каждой расы имеют второстепенный характер, однако по абсолютной величине они почти достигают уровня межрасовых различий. Монголоидные выборки обнаружили большую гомогенность по сравнению с европеоидами. Северные выборки из обеих рас различаются в меньшей степени, чем южные, что может объясняться длительными контактами между ними. По этой причине у северян-европеоидов отчетливо фиксируется северомонголоидная примесь, а у сибирских монголоидов - североевропеоидная (в широком смысле, включая восточноевропейское влияние). Обнаружено также, что женские выборки обеих рас различаются слабее, чем мужские (Хить, 2010).
Подведем итоги. Дерматоглифическая дифференциация человечества исследовалась на каждом из двух таксономических уровней двумя независимыми методами и по разному числу ключевых признаков. В первом случае, на уровне больших рас и субвариантов, материал был организован в рамках расово- соматологической классификации. Во втором анализировались популяционные выборки как самостоятельные таксоны, а конечные результаты оценивались дважды - как с помощью типологического критерия (в соответствии с расовой принадлежностью выборок), так и посредством статистического критерия (выделение скоплений кластерным анализом матриц ОДР и по данным анализа ГК) с последующей расовой идентификацией выборок, составляющих скопления. Выяснилось, что на обоих уровнях действуют одни и те же формообразующие факторы, приводя к устойчивому выделению кластеров и в целом к сходной картине расположения таксонов. На уровне больших рас этот процесс более канализован и отражает увеличенную долю общей изменчивости признаков по сравнению с уровнем популяций (по обеим ГК 77 и 67%, соответственно).
Суммируя результаты анализа всех уровней исследования, следует отметить, что каждая из рас характеризуется своеобразным сочетанием признаков и занимает самостоятельное положение в таксономическом пространстве. Наиболее специализированы негроиды и, в меньшей степени, европеоиды. Определенным
251

сходством отмечены австралоиды и монголоиды; американоиды тяготеют к этим двум расам (в большей степени - к монголоидной) и крайне резко различаются с остальными. Таким образом, подтверждается идея о выделении трех стволов - западного (европеоиды), южного (негроиды Африки) и восточного (монголоиды, австралоиды, американоиды), высказанная ранее (Хить, Кейта, 1981; Хить, Долинова, 1990). В описательной форме такое разделение рас по признакам кожного рельефа впервые отметил К. Плато (Plato, 1976).
Если исходить из гипотезы генетического единства человечества и дихотомической модели эволюционного древа при равной скорости эволюции рас по разным признакам, а таксономические расстояния связывать со временем дивергенции, то древнейшими расовыми ветвями древа следует считать негроидную и европеоидную, а самыми молодыми и сравнительно недавно разделившимися ветвями - австралоидов и монголоидов. Тогда нужно признать, что американоиды выделились из очень разнородного восточного ствола раньше, чем дифференцировались австралоиды и монголоиды. В действительности же не все перечисленные посылки могли иметь место, так что последнее слово по праву должно принадлежать палеоантропологии, а также генетике, воссоздающим процесс эволюции человечества во времени и пространстве на основе наиболее надежных источников.
В данном разделе представлены схемы, охватывающие все человечество и полученные на основе полного набора ключевых признаков дерматоглифики. Такие схемы, к сожалению, отсутствуют в краниологии, остеологии, расовой соматологии и одонтологии, несмотря на то, что накопленные материалы, значительно превосходят по объему и разнообразию использованные нами дерматоглифические данные. Это задача еще ждет своих исследователей.
Обнаруженные расхождения результатов анализа разных таксономических уровней неизбежны. Они отражают прежде всего неполное совпадение расово-соматологической идерматоглифической дифференциации человечества на уровне больших рас и популяций, усугубляемое недостаточностью числа популяционных выборок. Самыми корректными представляются результаты анализа больших рас, дополнительно свидетельствующие об адекватности расово-соматологической
252

классификации, в рамках которой был организовандерматоглифический материал.
Выводы, касающиеся всех рас, кроме американоидной, представляются надежными. Они базируются на четырехкратном пересчете среднерасовых величин, выполнявшемся по мере поступления новых материалов, но в результате характеристики практически не изменились. Однако данные об индейцах (в том, что касается отдельных признаков у расы в целом и популяционного уровня в особенности) нельзя считать достаточнопредставительными и методически полностью сравнимыми.Поэтому соответствующие заключения имеют ориентировочный характер.
253

Г лава 10. Бимануальное распределение и асимметрия дерматоглифических признаков:
популяционный аспект
Проблема бимануальной асимметрии неоднократно становилась предметом специальных исследований в дерматоглифике. Внимание специалистов привлекают проблемы симметрии и асимметрии различных типов ладонных и пальцевых узоров, гребневого счета, окончаний главных ладонных линий, связей признаков с полом, особенностей наследственной передачи и значения для медицинской диагностики (Гладкова, 1966; Гусева, Антонюк, 1969; Тегако, 1989; Micle, Kobyliansky, 1992; Sanders, Waters, 2001; Гусева, 2011; и др.). Обзор публикаций последних лет показывает, что различные аспекты исследования асимметрии папиллярных узоров входят в число немногочисленных актуальных тем в зарубежной дерматоглифике (Salzano, Benevides, 2005; Sengupta, Karmakar, 2006; Yousefi-Nooraie, Mortaz-Hedjri, 2008; King et al., 2009; и др.).
Результаты немногочисленных работ, посвященных популяционным аспектам асимметрии признаков, противоречивы и касаются почти исключительно пальцевых узоров. По мнению одних исследователей, явления симметрии и асимметрии пальцевых узоров мало отличаются у разных народов мира (Гладкова, 1962; Salzano, Benevides, 2005). По другим данным, симметрия по пальцевым узорам у монголоидов выше, чем у европеоидов (Гусева, 1986).
Расхождения в результатах, по всей вероятности, связаны с различиями в методических подходах к анализу материалов, а также с набором выборок, представляющих ту или иную расовую группу. Необходимо отметить, что понятие бимануальной асимметрии в дерматоглифике является относительным. Если принимать во внимание тончайшие различия между гребешковыми линиями, минуции, то придется признать, что ни людей, ни тем более человеческих популяций с абсолютно идентичными узорами папиллярных линий на гомологичных пальцах рук или различных участках ладоней просто не существует. Результаты исследований бимануальной асимметрии в значительной мере определяются
254

выбором системы дерматоглифических признаков и способа расчета величины последних.
Традиционно по классификации, впервые предложенной Л. Ван Валеном, выделяется три вида асимметрии: направленная, флуктуирующая и антисимметрия (Van Valen, 1962). В дерматоглифике направленная асимметрия определяется по значительным, генетически детерминированным различиям в частотах признака на гомологичных пальцах или участках ладоней. Флуктуирующая асимметрия встречается при незначительных ненаправленных отклонениях частот признаков от строгой билатеральной симметрии. Наконец, антисимметрия определяется по отрицательной связи между наличием признака на левой и правой руках.
Представленные ниже результаты межгрупповых исследований позволили выделить среди ключевых признаков два вида асимметрии: направленный (для Dl10, FRP, fc и Th/I) и флуктуирующий (для ДМТ, Hy и t). Однако следует заранее оговорить, что явления симметрии и асимметрии в данной главе понимаются в расширенном толковании, а способы их определения могут вызвать справедливое раздражение у генетиков. Для оценки бимануальной асимметрии использовались различные показатели соотношения средневыборочных значений признаков на левой и правой руках.
Показатели и даже типы асимметрии дерматоглифических признаков на разных таксономических уровнях далеко не во всех случаях связаны между собой. Теоретически можно представить популяцию, в которой для большинства индивидов характерно асимметричное распределение признака, однако соотношение суммарных величин последнего на левой и правой руках в группе будет близко к единице. В действительности такое расхождение на разных таксономических уровнях может возникнуть в случае, если на индивидуальном уровне для признака характерен тип флуктуирующей асимметрии, либо бимануальная антисимметрия. С другой стороны, если для какого-либо признака на уровне индивидов характерен направленный тип асимметрии, это будет проявляться и на других уровнях сопоставления.
255

10.1. Бимануальное распределение ключевых признаков у представителей трех больших рас
Межгрупповое сопоставление ключевых признаков дерматоглифики с учетом их бимануального распределения, насколько известно, до сих пор не становилось предметом специального исследования. Исключением является небольшой раздел в монографии Л.И. Тегако (1989). Исследователем отмечено, что в выборках европеоидов и монголоидов дельтовый индекс, индекс Камминса и узорность гипотенара как у мужчин, так и у женщин, имеют более высокую величину на правой руке. Низкий осевой трирадиус t, узоры на тенаре и частота ДМТ чаще встречаются на левой руке. По результатам анализа для всех пальцевых и ладонных признаков были выявлены значимые различия между левой и правой руками (Тегако, 1989, с.68-72). Несмотря на введенный в оборот обширный дерматоглифический материал, закономерности межпопуляционного распределения собственно показателей асимметрии не изучались.
Материалами для написания данной главы послужили неопубликованные данные, охватывающие 20 параллельно исследованных мужских и женских выборок (8 европеоидных, 8 монголоидных и 4 негроидных) численностью 3651 человек.
Установлено, что в мужских выборках величина обоих индексов, а также частота радиальных пальцевых узоров выше на правой руке, частота узорности тенара выше на левой. Для остальных признаков выделить строгие закономерности не удалось. В большинстве случаев частота проксимального трирадиуса у европеоидов выше на левой руке, у монголоидов - на правой. Добавочные межпальцевые трирадиусы, напротив, у европеоидов чаще встречаются на правой руке, у монголоидов - на левой. В обоих случаях узорность гипотенара, как правило, выше на левой руке (Табл. 10.1).
Результаты расчета T-критерия Уилкоксона свидетельствуют о существовании у мужского населения Старого Света (европеоидов, монголоидов и негроидов, анализируемых суммарно) на межгрупповом уровне значимых бимануальных различий по большинству основных признаков, за исключением узорности гипотенара, частот низкого осевого трирадиуса и ДМТ.
Те же различия проявляются при отдельном рассмотрении выборок в рамках европеоидной и монголоидной рас.
256

Таблица 10.1. Распределение частот встречаемости основных признаков и FRP на левой и правой руках в 20 популяциях Евразии и Западной Африки (мужчины).Table 10.1. Distribution of key traits frequencies on left and right hands in 20 male groups from Eurasia and Western Africa.
№Левая рука Правая рука
Dl10 Ic t Hy ДМТ Th/I FRP Dl10 Ic t Hy ДМТ Th/I FRP
1 s 5.39 7.02 82.7 26.9 12.5 15.4 12.0 5.95 8.82 69.2 27.9 13.5 6.7 14.8
? 5.58 8.33 63.2 31.6 6.3 8.4 10.3 5.96 9.01 61.1 33.7 5.3 3.2 10.5
2 s 6.41 8.06 72.0 31.0 19.0 16.0 9.1 6.91 10.10 64.0 30.0 27.0 4.0 17.4
? 5.69 7.49 52.3 30.8 6.2 15.4 9.2 6.05 9.82 52.3 27.7 12.3 7.7 12.2
3 s 6.04 8.22 78.9 36.8 18.9 17.9 13.5 6.27 9.35 74.7 33.7 13.7 4.2 15.3
? 5.89 7.35 71.8 32.4 9.9 7.0 9.0 6.01 9.13 62.0 33.8 15.5 5.6 9.6
4 s 5.96 8.35 54.5 37.4 17.2 11.1 11.4 6.48 9.13 59.6 25.3 13.1 2.0 13.3
? 5.85 8.05 52.0 34.0 11.0 10.0 9.4 6.27 8.89 55.0 34.0 11.0 3.0 14.4
5 s 6.71 8.19 65.0 32.0 32.0 23.0 12.4 7.06 9.68 60.0 34.0 30.0 5.0 15.0
? 6.48 7.74 61.0 41.0 27.0 8.0 15.4 6.57 9.78 63.0 45.0 26.0 5.0 12.1
6 s 6.45 7.26 46.8 41.4 18.9 4.5 13.8 6.67 9.51 50.5 36.0 24.3 1.8 14.8
? 6.45 7.26 46.8 41.4 18.9 4.5 12.1 6.67 9.51 50.5 36.0 24.3 1.8 9.7
7 s 6.55 8.40 49.5 29.0 39.8 19.4 11.2 7.10 9.97 46.2 34.4 40.9 9.7 12.5
? 6.43 8.33 50.5 45.4 20.6 17.5 11.1 6.81 10.08 49.5 38.1 26.8 12.4 13.4
8 s 7.19 7.80 57.3 38.2 37.3 14.5 12.7 7.55 10.15 51.8 39.1 38.2 6.4 16.3
? 6.92 7.74 50.0 45.0 43.0 16.0 13.4 6.81 9.64 54.0 48.0 35.0 10.0 11.5
9 s 6.80 6.12 75.7 29.0 8.4 15.0 12.7 7.29 8.16 74.8 32.7 3.7 2.8 18.5
? 7.00 5.52 66.3 35.6 4.8 5.8 15.4 7.30 7.70 68.3 34.6 4.8 4.8 19.4
10 s 7.38 5.99 63.4 19.8 10.7 17.6 15.3 7.80 8.32 74.8 21.4 13.0 3.8 20.8
? 7.24 5.71 65.7 30.0 10.7 13.6 14.5 7.49 8.09 71.4 27.9 16.4 5.0 19.7
11 s 6.73 6.00 74.6 22.0 0.0 10.2 11.5 7.10 8.22 71.2 20.3 5.1 5.1 14.2
? 6.35 6.75 60.0 30.0 15.0 37.5 12.0 7.30 8.73 70.0 17.5 7.5 12.5 14.0
12 s 7.60 7.07 60.4 30.2 12.5 29.2 13.6 8.14 9.22 68.8 22.9 9.4 13.5 21.1
? 7.19 5.86 76.4 29.1 5.5 14.5 14.5 7.44 8.15 74.5 18.2 6.4 5.5 24.5
13 s 7.67 7.63 75.5 32.7 12.2 10.2 15.6 8.16 8.92 79.6 14.3 6.1 4.1 15.0
? 7.58 6.56 79.2 39.6 2.1 8.3 12.9 7.75 10.88 83.3 22.9 2.1 0.2 20.9
14 s 6.62 6.90 81.0 28.0 18.0 15.0 14.9 6.85 9.14 83.0 24.0 12.0 5.0 15.8
? 6.30 6.17 60.0 31.0 13.0 13.0 14.7 6.37 8.57 66.0 27.0 9.0 5.0 14.3
15 s 7.20 6.84 83.0 22.0 10.0 9.0 11.5 7.79 8.47 86.0 17.0 6.0 3.0 21.2
? 6.97 7.58 77.0 28.0 8.0 18.0 13.7 7.46 8.36 82.0 24.0 10.0 6.0 16.7
16 s 7.53 7.73 78.8 45.5 15.9 19.7 14.5 7.93 8.74 74.2 29.5 15.9 8.3 16.8
? 7.31 7.73 71.2 32.8 10.4 20.0 15.4 7.54 8.59 72.0 25.6 10.4 8.8 16.0
257

Таблица 10.1. Продолжение. Table 10.1. Continued.
№ПОЛ
Левая рука Правая рука
Dl10 Ic t Hy ДМТ Th/I FRP Dl10 Ic t Hy ДМТ Th/I FRP
17 3 6.06 7.10 60.0 14.0 46.0 16.0 11.0 6.38 8.17 61.0 19.0 48.0 9.0 11.2
? 6.21 7.13 56.4 25.6 30.8 15.4 13.0 5.97 7.69 56.4 20.5 33.3 12.8 13.5
18 3 5.95 6.72 67.0 21.0 45.0 23.0 9.9 6.13 7.85 60.0 19.0 40.0 11.0 11.7
? 5.61 7.16 50.0 18.4 31.6 21.1 8.7 5.66 7.97 42.1 15.8 34.2 13.2 9.7
19 3 6.30 7.01 71.0 22.0 44.0 18.0 10.0 6.54 8.13 67.0 23.0 46.0 4.0 10.5
? 6.44 7.19 56.3 21.9 31.3 21.9 15.3 6.59 8.34 59.4 21.9 25.0 9.4 12.7
20 3 6.20 6.65 68.9 23.6 36.8 26.4 13.5 6.72 7.49 63.2 27.4 38.7 5.7 10.2
? 6.07 7.32 54.4 23.5 47.1 17.6 13.2 6.19 8.28 57.4 23.5 48.5 2.9 5.4
Нумерация групп: 1 - вепсы; 2 - латыши; 3 - эстонцы; 4 - русские; 5 - азербайджанцы; 6 - армяне; 7 - персы; 8 - белуджи; 9 - нанайцы; 10 - дунгане; 11 - ульчи; 12 - эвенки; 13 - нганасаны; 14 - качинцы; 15 - якуты; 16 - ненцы; 17 - фульбе; 18 - банмана; 19 - манинке; 20 - малийцы (сбор.).Numeration of groups: 1 - Vepsians; 2 - Letts; 3 - Estonians; 4 - Russians; 5 - Azerbaijanians; 6 - Armenians; 7 - Persians; 8 - Baluchi; 9 - Nanay; 10 - Dungani; 11 - Ulchi; 12 - Evenks; 13 - Nganasany; 14 - Kachin; 15 - Yakuts; 16 - Nenets; 17 - Fulbe; 18 - Banmana; 19 - Maninke; 20 - Malians (random sample).
Таким образом, четыре признака из семи (DlJ0, Ic, Th/I, FRP) обладают направленным типом асимметрии, тогда как асимметрия в бимануальном распределении остальных признаков (ДМТ, Hy, t) носит случайный (флуктуирующий) характер. Статистический анализ данных негроидных выборок не проводился из-за их небольшой численности.
Направления различий между величинами признаков в женских выборках в целом аналогичны наблюдаемым у мужчин. Оба индекса имеют большую величину на правой руке в большинстве групп. То же наблюдение относится к бимануальному распределению частот ДМТ и FRP, хотя и с несколько большим числом исключений. Для узорности Hy и Th/I, напротив, характерны более высокие частоты на левой руке.
По результатам расчета T-критерия Уилкоксона у женского населения Старого Света на межгрупповом уровне сопоставления выявлены значимые билатеральные различия по большинству
258

признаков, за исключением частот ДМТ и проксимального осевого трирадиуса t (в последнем случае различия близки к значимым, p<0.055). У европеоидов, кроме указанных признаков, не являются значимыми также различия по частоте узорности гипотенара и FRP. У монголоидов картина сходна с общечеловеческой.
С целью оценки значения бимануального распределения признаков для дифференциации популяций была проведена серия анализов методом главных компонент по тем же материалам, представляющих три больших расы. Анализировались значения 6 ключевых признаков и FRP отдельно для левой и правой рук, а также без учета бимануального распределения.
10.1.1. Анализ главных компонент признаков на левойруке
Положение мужских и женских групп в пространстве первых двух компонент определяется разными комбинациями признаков. Однако в обоих случаях различия между европеоидными и негроидными выборками, с одно стороны, и монголоидными выборками, с другой, выступают на первое место (Рис. 10.1). Суммарно первые две компоненты охватывают 57.4% общей изменчивости признаков у мужчин, и 60.9% - у женщин. В целом группы, относящиеся к трем разным большим расам, составляют обособленные скопления в пространстве первых двух ГК. Исключение составляет мужская выборка вепсов и женская выборка нанайцев, которые по второй ГК сближаются с группами негроидов в большей степени, чем с европеоидами и монголоидами, соответственно.
Разнополые группы характеризуются относительным параллелизмом в изменчивости признаков. Так, первые ГК у мужчин и женщин связаны значимой отрицательной (-0.79), вторые ГК - значимой положительной связью (0.66). При этом величины коэффициентов корреляции между изменчивостью исходных показателей значимы для всех признаков, за исключением Th/I, и возрастают в порядке: FRP - t - Hy - Ic - ДМТ - DlJ0.
По признакам левой руки по результатам T-теста Уилкоксона значимые различия между мужчинами и женщинами на межпопуляционном уровне были зафиксированы для трех признаков из семи: дельтового индекса (p<0.009), частоты осевого трирадиуса (p<0.004) и частоты ДМТ (p<0.03).
259

Рисунок 10.1. Положение 20 мужских (слева) и женских (справа) групп в пространстве первых двух главных компонент по результатам анализа семи признаков левой руки. Нумерация групп: 1 - вепсы; 2 - латыши; 3 - эстонцы; 4 - русские; 5 - азербайджанцы; 6 -армяне; 7 - персы; 8 - белуджи; 9 - нанайцы; 10 - дунгане; 11 - ульчи; 12 - эвенки; 13 - нганасаны; 14 - качинцы; 15 - якуты; 16 - ненцы; 17 - фульбе; 18 - банмана; 19 - манинке; 20 - малийцы (сбор.).Figure 10.1. Positions of 20 male (left) and female (right) groups on the first two principal components (data on 7 traits for left hand). Numeration of groups: 1 - Vepsians; 2 - Letts; 3 - Estonians; 4 - Russians; 5 - Azerbaijanians; 6 - Armenians; 7 - Persians; 8 - Baluchi; 9 - Nanay; 10 - Dungani; 11 - Ulchi; 12 - Evenks; 13 - Nganasany; 14 - Kachin; 15 - Yakuts; 16 - Nenets; 17 - Fulbe; 18 - Banmana; 19 - Maninke; 20 - Malians (random sample).
10.1.2. Анализ главных компонент признаков на правойруке
В отличие от предыдущего варианта анализа, положение как мужских, так и женских групп в пространстве первых двух компонент определяется одинаковыми комбинациями признаков (Рис.10.2). Первая компонента дифференцирует группы по частотам радиальных пальцевых узоров, добавочных межпальцевых трирадиусов и низкого осевого трирадиуса, а также величине дельтового индекса Вторая компонента разграничивает выборки по величине индекса Камминса и узорности гипотенара. Выборки группируются в соответствии с их расовой принадлежностью, без трансгрессии между скоплениями. Женская выборка нганасан располагается в поле «африканских» значений по второй
260

компоненте, однако максимально резко отличается от негроидов по первой ГК. Суммарно первые две компоненты охватывают около 67% общей изменчивости признаков у мужчин, и 64% - у женщин. Таким образом, межпопуляционная изменчивость ключевых признаков по признакам правой руки имеет несколько более канализированный характер, чем по признакам левой.
Дифференциация по признакам правой руки обнаруживает также больший параллелизм между разнополыми выборками, нежели по признакам левой. Коэффициент корреляции между вновь полученными признаками мужских и женских групп для правой руки в пространстве первой компоненты составляет 0.91, в пространстве второй ГК - 0.69, т.е. выше, чем для левой. При этом корреляции между вариационными рядами всех исходных признаков у мужчин и женщин на правой руке значимы и выше, возрастая в ряду: Th/I - FRP - Ic - t - Hy - ДМТ - Dli0. Значимыми являются межполовые различия по тем же признакам, что и для левой руки: по величине дельтового индекса (p<0.0007), частоты осевого трирадиуса (p<0.008) и ДМТ (p<0.03).
Рисунок 10.2. Положение 20 мужских (слева) и женских (справа) групп в пространстве первых двух главных компонент по результатам анализа семи признаков правой руки. Нумерация групп соответствует подписи к Рис. 10.1Figure 10.2. Positions of 20 male (left) and female (right) groups on the first two principal components (data on 7 traits for right hand). Groups are numbered as in note of Figure 10.1.
Различия между европеоидными и монголоидными группами в большей степени проявляются именно по признакам правой руки (Табл. 10.2). Более выраженная разграничительная способность
261

признаков правой руки частично может быть объяснена статистическими причинами. Четыре из шести признаков, различия по которым между монголоидами и европеоидами значимы, имеют большую величину на правой руке.
Таблица 10.2. Уровень значимости различий между европеоидами и монголоидами по средневыборочным значениям признаков левой и правой руки.Table 10.2. Significance value of distinctions between Caucasoids and Mongoloids (average values of traits on left and right hands).
Признаки левой рукиDlio Ic t Hy ДМТ Th/I FRP
P (tf) 0.004 0.005 - - 0.003 - -M (E) 6.34 7.91 63.3 34.1 24.4 15.2 12.0V (E) 0.09 0.11 0.16 0.28 0.62 0.36 0.15
M (M) 7.19 6.79 74.0 28.6 11.0 15.7 13.7V (М) 0.06 0.10 0.11 0.29 0.49 0.42 0.12
P (? ) 0.02 0.007 0.02 0.04 - - 0.03M (E) 6.16 7.79 56.0 37.7 17.9 10.9 11.2V (E) 0.08 0.12 0.16 0.23 0.75 0.51 0.18
M (M) 6.99 6.49 69.5 32.0 8.7 16.3 14.1V (М) 0.06 0.13 0.11 0.12 0.51 0.60 0.08
Признаки правой рукиDli0 Ic t Hy ДМТ Th/I FRP
P (tf) 0.007 0.005 0.004 0.005 0.003 - 0.05M (E) 6.75 9.59 59.5 32.5 25.1 5.0 14.9V (E) 0.10 0.09 0.16 0.26 0.68 0.54 0.22
M (M) 7.63 8.65 76.5 22.8 8.9 5.7 17.9V (М) 0.06 0.05 0.08 0.27 0.49 0.63 0.16P (? ) 0.004 0.02 0.0008 0.005 0.02 - 0.002M (E) 6.39 9.48 55.9 37.0 19.5 6.1 11.7V (E) 0.10 0.10 0.17 0.31 0.70 0.58 0.32
M (M) 7.33 8.63 73.4 24.7 8.3 6.0 18.2V (М) 0.06 0.11 0.08 0.22 0.52 0.59 0.20
P(tf) - уровень значимости различий между расами для мужских групп; P (?) - уровень значимости различий между расами для женских групп; M и V(E) - среднее значение и коэффициент вариации признака у европеоидов; M и V(M) - среднее значение и коэффициент вариации признака у монголоидов.P(tf) - Significance value of distinctions in male groups; P (? ) - Level of significance of differences between female groups; M и V(E) - average and variation coefficient in Caucasoid groups; M и V(M) - average and variation coefficient in Mongoloid groups.
262

При этом чаще всего чем меньше значение признака, тем больше коэффициент его межгрупповой вариации, и признак в большей степени подвержен случайным отклонениям. Впрочем, это правило оказывается не таким строгим при детальном рассмотрении. Средние значения FRP выше на правой руке как у европеоидов, так и у монголоидов, коэффициенты вариаций признака также выше на правой руке. У тех же рас частоты узорности гипотенара и ДМТ выше на левой руке, однако, различия значимы только для признаков на правой.
Таким образом, можно предположить, что признаки правой руки несут более надежную информацию о межрасовой дифференциации, чем признаки левой, и причина этого не может быть полностью объяснена статистическими эффектами. Возникает вопрос: является ли дифференцирующая способность ключевых признаков правой руки более высокой, чем при традиционном расчете усредненных величин признаков обеих рук (без учета их бимануального распределения)?
10.1.3. Анализ главных компонент признаков без учета бимануального распределения
Наибольшее значение для дифференциации мужских и женских серий в пространстве первой компоненты имеют частоты ДМТ, FRP, t, а также Dl10, в пространстве второй - частота узорности Hy и Ic, а у женщин, помимо указанных признаков, - частота Th/I (Рис.10.3). Суммарно первые две компоненты охватывают 64.9% и 69.9% общей изменчивости признаков в мужских и женских группах соответственно, что в большей степени согласуется с результатами анализа признаков правой руки. При этом скопления выборок соответствуют их принадлежности трем большим расам, а трансгрессии значений отсутствуют.
Коэффициенты корреляции между вновь полученными признаками у мужчин и женщин составляют 0.87 для первой ГК, и - 0.81 - для второй, т.е. также близки к значениям, полученным для признаков правой руки. Связи между изменчивостью ключевых признаков в разнополых группах значимы для всех показателей, кроме Th/I, и возрастают в порядке FRP - t - Ic и Hy - ДМТ - Dl10. Коэффициенты корреляции в большей степени сближаются с соответствующими показателями для правой руки или имеют
263

промежуточную между признаками отдельно рассмотренных рук величину.
Межполовые различия, как и в предыдущих случаях, значимы по величине дельтового индекса (p<0.0006), частоте осевого трирадиуса (p<0.004) и ДМТ (p<0.02). Кроме того, значимыми оказываются различия по частоте узорности гипотенара (p<0.03) - признаке, не имеющем дифференцирущего значения для пола при отдельном рассмотрении его частот на разных руках.
I K II (27.2% )1 8 Ну (-0.89)
И Ic (-0.85)а17 О П
19 20
4 2 3■ ■ ■
■-1 о
11□
14□
12□
16
dГ К II (25.6% )Ну (0.88)Th/I (-0.70)Ic (0.66)
13□ i r -'Ш
ГК I (37.7% )ДМ Т (-0.88) FRP (0.86) 1(0.71)D110 (0.67)
■ ЕВРОПЕОИДЫ □ МОНГОЛОИДЫа н е г р о и д ы
на—20
18а
8§#7 4 2
17 19и а
14□
11□
□13
9161c D
□ □□
10ГК I (44.3% )FRP (0.93) t (0.89)D ll0 (0.84) ДМТ (-0.68)
12□-1
Рисунок 10.3. Положение 20 мужских (слева) и женских (справа) групп в пространстве первых двух главных компонент по результатам анализа ключевых признаков без учета бимануального распределения. Нумерация групп соответствует подписи к Рис. 10.1Figure 10.3. Positions of 20 male (left) and female (right) groups on the first two principal components (data on 7 traits nonmetering bimanual asymmetry). Groups are numbered as in note of Figure 10.1.
По совокупности показателей ключевые признаки дерматоглифики (без учета их бимануального распределения) обладают примерно равной дифференцирующей способностью с признаками правой руки, превышая разграничительное значение признаков левой. Кроме того, усредненные показатели отличаются согласованною изменчивостью параллельно исследованных мужских и женских выборок. Межполовые различия признаков, определяемых суммарно для обеих рук, проявляются более отчетливо, чем в случае учета бимануального распределения их значений. Отметим также, что во всех вариантах анализа частота узорности Th/I оказывается наименее значимым признаком для
264

дифференциации выборок различной расовой принадлежности, а ладонные признаки менее значимы, чем пальцевые (Dl10 и FRP). Эти различия могут быть связаны как с особенностями генетического наследования пальцевых и ладонных узоров, так и с методическими подходами к определению дерматоглифических признаков. (Например, узорность гипотенара может быть сравнительно менее эффективным признаком, чем FRP, только по той причине, что она является комплексным признаком, составные элементы которого, т.е. радиальные петли, ульнарные петли, завитки и др., в действительности отчетливо разграничивают расы, но имеют разные направления изменчивости на межпопуляционном уровне).
10.2. Коэффициенты асимметрии основных признаков
Для оценки влияния степени выраженности асимметрии дерматоглифических признаков на межпопуляционную дифференциацию в пределах трех больших рас был применен метод главных компонент, в качестве исходных данных для которого использовались коэффициенты асимметрии семи признаков. В действительности эти показатели могут считаться коэффициентами асимметрии только при широком толковании термина, поскольку они не выражают средневыборочное соотношение индивидуальных значений признаков на левой и правой руках. Во всех случаях коэффициенты асимметрии рассчитывались как отношение средней величины признака на левой руке по отношению к средней величине признака на правой (Табл. 10.3).
Во всех мужских и женских группах коэффициент асимметрии имеет величину меньше единицы для обоих индексов и частоты радиальных пальцевых узоров (за редкими исключениями) и больше единицы - для частоты узорности тенара. Таким образом, эти цифры можно считать отражающими общечеловеческие тенденции. Для остальных трех признаков общие закономерности не были выявлены. Частота проксимального осевого трирадиуса, как правило, выше в большинстве групп на левой руке, однако, для каждой из трех больших рас фиксируются случаи обратного соотношения. Изменчивость признака свидетельствует лишь о наличии половых различий и только по пальцевым признакам.
Величины коэффициентов асимметрии ДМТ и узорности гипотенара беспорядочно варьируют в выборках всех трех рас. у женщин, независимо от расы, относительные различия между
265
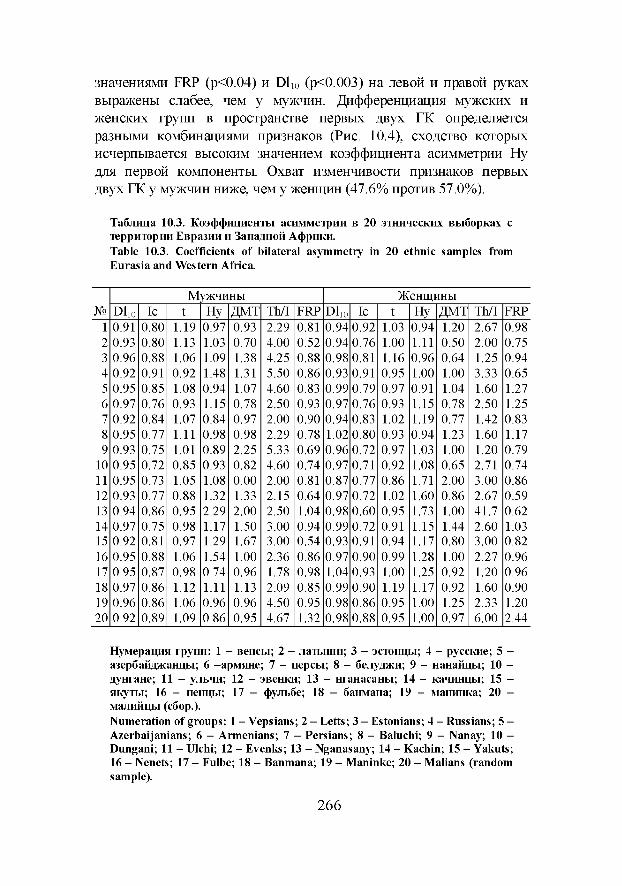
значениями FRP (p<0.04) и Dl10 (p<0.003) на левой и правой руках выражены слабее, чем у мужчин. Дифференциация мужских и женских групп в пространстве первых двух ГК определяется разными комбинациями признаков (Рис. 10.4), сходство которых исчерпывается высоким значением коэффициента асимметрии Hy для первой компоненты. Охват изменчивости признаков первых двух ГК у мужчин ниже, чем у женщин (47.6% против 57.0%).
Таблица 10.3. Коэффициенты асимметрии в 20 этнических выборках с территории Евразии и Западной Африки.Table 10.3. Coefficients of bilateral asymmetry in 20 ethnic samples from Eurasia and Western Africa.
№Мужчины Женщины
Dl10 Ic t Hy ДМТ Th/I FRP Dl10 Ic t Hy ДМТ Th/I FRP1 0 91 0.80 1 19 0.97 0 93 2.29 0 .81 0 94 0 92 1.03 0.94 1.20 2 .67 0.982 0 93 0.80 1 13 1.03 0 70 4.00 0 .52 0 94 0 76 1.00 1.11 0.50 2 .00 0.753 0 96 0.88 1.06 1.09 1 38 4.25 0 .88 0 98 0 81 1 16 0.96 0.64 1.25 0.944 0 92 0.91 0 92 1.48 1 31 5.50 0 .86 0 93 0 91 0 .95 1.00 1.00 3 .33 0.655 0 95 0.85 1.08 0.94 1 07 4.60 0 .83 0 99 0 79 0 .97 0.91 1.04 1.60 1.276 0 97 0.76 0 .93 1.15 0 78 2.50 0 .93 0 97 0 76 0 .93 1.15 0.78 2 .50 1.257 0 92 0.84 1.07 0.84 0 97 2.00 0 .90 0 94 0 83 1.02 1.19 0.77 1.42 0.838 0 95 0.77 1.11 0.98 0 98 2.29 0 .78 1 02 0 80 0 .93 0.94 1.23 1.60 1.179 0 93 0.75 1.01 0.89 2 25 5.33 0 .69 0 96 0 72 0 .97 1.03 1.00 1.20 0.79
10 0 95 0.72 0 .85 0.93 0 82 4.60 0 .74 0 97 0 71 0 2.9 1.08 0.65 2 .71 0.7411 0 95 0.73 1 5.0 1.08 0 0 0 2.00 0 .81 0 78 0 77 0 6.8 1.71 2.00 3 0.0 0.8612 0 93 0.77 0 8.8 1.32 1 33 2.15 0 4.6 0 79 0 72 1 2.0 1.60 0.86 2 7.6 0.5913 0 49 0.86 0 5.9 2.29 2 0 0 2.50 1 4.0 0 98 0 60 0 5.9 1.73 1.00 41.7 0.6214 0 97 0.75 0 .98 1.17 1 50 3.00 0 .94 0 99 0 72 0 .91 1.15 1.44 2 .60 1.0315 0 92 0.81 0 97 1.29 1 67 3.00 0 .54 0 39 0 91 0 4.9 1.17 0.80 3 .00 0.8216 0 95 0.88 1 6.0 1.54 1 0 0 2.36 0 6.8 0 79 0 90 0 .99 1.28 1.00 2 .27 0.9617 0 95 0.87 0 .98 0.74 0 96 1.78 0 .98 1 04 0 93 1.00 1.25 0.92 1.20 0.9618 0 97 0.86 1 12 1.11 1 13 2.09 0 .85 0 99 0 90 1.19 1.17 0.92 1.60 0.9019 0 96 0.86 1 6.0 0.96 0 69 4.50 0 .95 0 98 0 68 0 5.9 1.00 1.25 2 3.3 1.2020 0 92 0.89 1 09 0.86 0 95 4.67 1 32 0 98 0 88 0 95 1.00 0.97 6 00 2.44
Нумерация групп: 1 - вепсы; 2 - латыши; 3 - эстонцы; 4 - русские; 5 - азербайджанцы; 6 -армяне; 7 - персы; 8 - белуджи; 9 - нанайцы; 10 - дунгане; 11 - ульчи; 12 - эвенки; 13 - нганасаны; 14 - качинцы; 15 - якуты; 16 - ненцы; 17 - фульбе; 18 - банмана; 19 - манинка; 20 - малийцы (сбор.).Numeration of groups: 1 - Vepsians; 2 - Letts; 3 - Estonians; 4 - Russians; 5 - Azerbaijanians; 6 - Armenians; 7 - Persians; 8 - Baluchi; 9 - Nanay; 10 - Dungani; 11 - Ulchi; 12 - Evenks; 13 - Nganasany; 14 - Kachin; 15 - Yakuts; 16 - Nenets; 17 - Fulbe; 18 - Banmana; 19 - Maninke; 20 - Malians (random sample).
266

ГК II (22.6%)Ic (-0.82)
' FRP (-0.77)
Ю□ 13-1
1215 D□□9
10□
<f
14□
2 11■ 8 a
16
3 <jg 171
13□ГК II (22.4%) ДМТ (-0.73) t (0.60)
■ ЕВРОПЕОИДЫ□ 1МОНГОЛОИДЫ□ НЕГРОИДЫ
20ao
ГК I <2S.0%) ]ДМТ (-0.80)
1 Ну (-0.72)11□
12□
-2
2 18a10 ш 1? 1
° e . 7 a 9n^16 5 о
14 15^8.".4 a ■ -ij
19 20aГК I (34.6%)Ну (-0.83)Ic (0.70)
■1 0 Th/I (-0.66)
3
Рисунок 10.4 Положение 20 мужских (слева) и женских (справа) групп в пространстве первых двух главных компонент по результатам анализа коэффициентов асимметрии семи признаков. Нумерация групп соответствует подписи к Рис. 10.1Figure 10.4. . Positions of 20 male (left) and female (right) groups on the first two principal components (data on asymmetry parameters of 7 traits). Groups are numbered as in note of Figure 10.1.
Однако только положение мужских выборок свидетельствует об отчетливой тенденции к разделению всех трех больших рас по степени выраженности асимметрии дерматоглифических признаков. У женщин намечена межрасовая дифференциация монголоидных и европеоидных выборок, но в целом результаты анализа вряд ли можно считать надежными, поскольку они оказались почти полностью обусловлены случайными (возможно) отклонениями значений коэффициентов асимметрии в выборках нганасан и ульчей относительно большинства привлеченных групп. Значимее корреляции между главными компонентами в мужских и женских группах отсутствуют.
10.3. Частоты носительства асимметричного распределения основных признаков
Частоты носительства асимметричного распределения основных признаков определялись как процентное соотношение числа индивидов с билатеральной асимметрией признаков по отношению к общему числу индивидов в выборке. При этом асимметрия сложных признаков (индексы, частоты радиальных узоров и ДМТ) определялась в результате сопоставления суммарных
267

величин для левой и правой рук, без учета распределения составляющих признаки элементов и особенностей их топографии. Например, если у конкретного индивида на пальцах каждой из рук суммарно было зафиксировано по 6 дельт, такой вариант рассматривался как случай носительства симметричного распределения признака, даже если на левой руке располагается четыре однодельтовых и один двудельтовый узор, а на правой - два однодельтовых, два двудельтовых и один бездельтовый.
В большинстве европеоидных и монголоидных групп ключевые признаки, выстроенные в ряд в соответствии с увеличением процента лиц - носителей асимметричногораспределения, - располагаются в порядке: Ic - Dli0 - FRP - Hy - ДМТ - t - Th/I. (Табл. 10.4). Для африканских выборок эта последовательность имеет несколько иной вид: Ic - Dl10 - FRP - ДМТ - Hy и Th/I - t.
Применение критерия Манна-Уитни позволило выявить значимые различия между монголоидами и негроидами только для частоты носительства ДМТ в мужских выборках (p<0.001). В европеоидных группах процент носителей асимметричногораспределения ДМТ, как правило, выше, чем в монголоидных. Между евразийскими выборками и африканскими значимые различия отмечены чаще. Так, у мужчин между негроидами и европеоидами выявлены различия по числу носителей асимметричногораспределения FRP (p<0.009), Dl10 (p<0.05) и ДМТ (p<0.03).
Частота носительства первых двух признаков выше в европейских выборках, последнего - в африканских. У женщин негроидные и европеоидные выборки различаются по проценту лиц с асимметричным распределением Ic, ДМТ и Hy (p<0.05), негроидные и монголоидные - по Ic (p<0.05) и ДМТ (p<0.005), а у мужчин в последнем случае также по FRP (p<0.005). У негроидов асимметрия всех ключевых признаков сравнительно редко встречается. Лишь число людей с различным соотношением значений ДМТ на разных руках в Африке значительно выше, чем среди представителей двух других больших рас.
Кроме того, при общем рассмотрении групп, независимо от их расовой принадлежности, было зафиксировано наличие половых различий по ряду признаков. Процент мужчин с асимметричным распределением Dl10 (p<0.05), ДМТ (p<0.009) и Th/I (p<0.007) в выборках, как правило, значительно выше, чем процент женщин с соответствующими признаками.
268

Таблица 10.4. Частоты носительства асимметричного распределения основных признаков в этнических группах Евразии и Западной Африки. Table 10.4. Frequencies of persons with bimanual asymmetrical key traits in ethnic groups from Eurasia and Western Africa.
Мужчины Женщины№ DI10 Ic t Hy ДМТ Th/I FRP DI10 Ic t Hy ДМТ Th/I FRP
1 64.4 79.8 13.5 20.2 17.3 12.5 49.5 73.2 62.1 25.3 26.3 4.2 5.3 48.5
2 66.0 79.0 16.0 27.0 14.0 14.0 60.6 72.3 87.7 15.4 21.5 12.3 13.8 47.5
3 58.9 76.8 12.6 26.3 13.7 15.8 53.8 67.6 78.9 18.3 18.3 11.3 4.2 38.0
4 69.7 68.7 25.3 22.2 14.1 11.1 51.0 70.0 79.0 23.0 18.0 10.0 11.0 44.0
5 61.0 76.0 15.0 22.0 30.0 18.0 54.0 65.0 85.0 14.0 32.0 22.0 5.0 50.5
6 60.0 79.0 22.0 33.0 21.0 9.0 51.6 69.4 88.3 18.9 38.7 15.3 4.5 49.5
7 66.7 75.3 16.1 35.5 26.9 16.1 41.8 58.8 77.3 21.6 34.0 21.6 9.3 53.6
8 64.5 84.5 18.2 28.2 31.8 11.8 56.0 62.0 77.0 20.0 27.0 21.0 10.0 47.4
9 66.4 86.0 15.9 22.4 8.4 14.0 58.9 62.5 87.5 21.2 22.1 7.7 2.9 55.8
10 60.3 84.7 19.1 19.8 13.0 13.7 56.5 57.9 85.7 20.0 23.6 15.7 11.4 56.5
11 64.4 86.4 16.9 15.3 5.1 8.5 49.2 72.5 87.5 25.0 12.5 10.0 10.0 40.0
12 59.4 85.4 20.8 24.0 11.5 24.0 52.1 63.6 86.4 16.4 25.5 4.5 12.7 58.5
13 55.1 75.5 12.2 26.5 14.3 10.2 50.0 68.8 89.6 8.3 25.0 0.0 8.3 55.6
14 59.0 84.0 18.0 20.0 12.0 12.0 53.7 60.0 89.0 16.0 22.0 10.0 10.0 54.1
15 61.0 78.0 9.0 17.0 12.0 6.0 55.9 58.0 70.0 10.0 14.0 3.0 8.0 46.3
16 57.6 72.7 15.2 37.1 15.2 14.4 49.6 60.8 66.4 15.2 21.6 16.0 16.0 47.7
17 60.0 71.0 13.0 17.0 27.0 13.0 34.5 74.4 66.7 20.5 10.3 20.5 7.7 45.0
18 54.0 73.0 13.0 12.0 33.0 16.0 37.8 52.6 71.1 18.4 13.2 28.9 7.9 33.3
19 49.0 76.0 16.0 13.0 29.0 14.0 46.7 68.8 62.5 15.6 18.8 25.0 25.0 50.9
20 61.3 74.5 13.2 24.5 29.2 22.6 39.5 57.4 66.2 8.8 20.6 36.8 14.7 46.3
Нумерация групп: 1 - вепсы; 2 - латыши; 3 - эстонцы; 4 - русские; 5 - азербайджанцы; 6 -армяне; 7 - персы; 8 - белуджи; 9 - нанайцы; 10 - дунгане; 11 - ульчи; 12 - эвенки; 13 - нганасаны; 14 - качинцы; 15 - якуты; 16 - ненцы; 17 - фульбе; 18 - банмана; 19 - манинка; 20 - малийцы (сбор.).Numeration of groups: 1 - Vepsians; 2 - Letts; 3 - Estonians; 4 - Russians; 5 - Azerbaijanians; 6 - Armenians; 7 - Persians; 8 - Baluchi; 9 - Nanay; 10 - Dungani; 11 - Ulchi; 12 - Evenks; 13 - Nganasany; 14 - Kachin; 15 - Yakuts; 16 - Nenets; 17 - Fulbe; 18 - Banmana; 19 - Maninke; 20 - Malians (random sample).
269

Полученные при помощи метода главных компонент результаты анализа популяционных частот носительства асимметричных узоров, представлены на Рисунке 10.5.
Положение мужских и женских групп в пространстве первых двух ГК определяется разными комбинациями признаков. Однако в обоих случаях наибольшее значение для дифференциации выборок по первой компоненте имеют индекс Камминса и частоты ДМГ и радиальных пальцевых узоров, общий охват изменчивости первых двух ГК составляет около 55%, а дифференциация расовых групп по данным обоих полов представляет собой сходную картину.
Первая компонента четко разграничивает африканские и евразийские выборки. Связь между соответствующими значениями вновь полученных признаков у мужчин и женщин статистически значима (r=-0.72). По второй компоненте у мужчин намечается тенденция к разделению групп монголоидов и европеоидов, практически невыраженная на материалах женских выборок.
Следует отметить, что между собственно величинами признаков и частотами носительства асимметричного распределения признаков у мужчин существуют высокие значимые корреляции.
Рисунок 10.5. Положение 20 мужских (слева) и женских (справа) групп в пространстве первых двух главных компонент по результатам анализа частот носительства асимметричного распределения семи признаков. Нумерация групп соответствует подписи к Рис. 10.1 Figure 10.5. . The position of 20 male (on the left) and female (on the right) groups on the first two principal components (data on frequency of persons with asymmetrical distribution of traits). Groups are numbered as in note of Figure 10.1.
270

Исключение составляют оба индекса. Для осевого проксимального трирадиуса r=-0.53 при p<0.008, для Hy r=0.73 при p<0.00006, для ДМТ r=0.87 при p<0.000001, для Th/I r=0.88 при p<0.000001, для частоты радиальных узоров r=0.71 при p<0.0001. В то же время между коэффициентами асимметрии и величинами признаков значимые корреляции имеют небольшую величину и выявлены лишь в двух случаях: для тенара (r=-0.45 при p<0.03) и частоты радиальных узоров (r=-0.50 при p<0.02). Таким образом, частоты носительства асимметричного распределения признаков не могут рассматриваться в качестве самостоятельных признаков, поскольку находятся в прямой зависимости от частоты встречаемости того или иного признака в группе. Редкая встречаемость в изученных выборках таких признаков, как узоры на тенаре и частота ДМТ, имеет определяющее значение для подсчета общего количества носителей асимметричных признаков в популяции, а случайные колебания частот исходных признаков оказывают влияние и на общие итоги исследования дифференцирующей способности частот носительства.
Основные выводы
На материалах 20 мужских и 20 женских выборок, относящихся к трем большим расам, было выявлено стабильное преобладание направленного вида асимметрии для 4 из 7 исследованных признаков: Dl10, Ic, FRP и Th/I. По этим признакам в выборках всех рас выявлена правосторонняя асимметрия, за исключением частота узорности Th/I, которая выше на левой руке. У женщин, кроме того, отчетливо выражена левосторонняя асимметрия частоты узорности Hy. Показатели бимануального распределения остальных признаков (ДМТ и t у обоих полов, а также Hy у мужчин) варьируют на межпопуляционном уровне беспорядочно, что свидетельствует о флуктуирующем виде асимметрии.
Показатели асимметрии обладают относительно низкой дифференцирующей способностью на межпопуляционном уровне сопоставления, более высокой по результатам анализа мужских выборок. В большинстве случаев, по результатам по результатам расчетов популяционных величин коэффициентов асимметрии и частот носительства асимметричного распределения признаков, монголоиды характеризуются максимально, а негроиды -
271

минимально выраженной бимануальной асимметрией признаков (за исключением ДМТ, показатели асимметрии которых выше у негроидов). Европеоиды занимают среднее положение между этими вариантами.
В ходе сопоставления результатов ряда статистических анализов, проведенных с учетом бимануального распределения межпопуляционных значений признаков, были выявлены следующие закономерности:
• Признаки правой руки и признаки, определяемые без учета бимануального распределения, обладают более выраженной разграничительной (на уровне больших рас) мощностью по сравнению с признаками левой;
• Выраженность направленных различий между полами возрастает в следующем порядке: признаки левой руки - признаки правой руки - признаки без учета бимануального распределения (усредненные характеристики);
• Наблюдается почти полный параллелизм в изменчивости признаков правой руки в мужских и женских выборках и снижение согласованной вариабельности между полами по признакам левой.
Вышеперечисленные результаты справедливы в рамках выбранной для анализа группы из семи признаков (ключевые признаки, а также FRP) и не могут быть безоговорочно отнесены ко всей системе папиллярных узоров кисти человека. Представляется, что показатели бимануальной асимметрии группы ключевых признаков относительно неэффективны для задач межрасовой дифференциации выборок. Это наблюдение, по крайней мере, частично объясняется тем, что при изучении асимметрии этих признаков из поля зрения исследователя автоматически выпадают такие важные элементы, как бимануальные различия по типам узоров (при использовании данных по дельтовому индексу, радиальным пальцевым узорам, узорности тенара и гипотенара) и по деталям ладонной топографии (при использовании индекса Камминса и частоте ДМТ), что придает вычисляемым показателям искусственный характер.
272

%

Глава 11. Дерматоглифика и исторические процессы
К началу XXI века дерматоглифические исследования в области расоведения, достигшие максимального развития в 60-80-е гг. прошлого столетия, постепенно утратили свои позиции, о чем свидетельствует исключительная редкость публикации работ соответствующей тематики в ведущих антропологических изданиях мира. В этом отношении показательным является пример «American Journal of Physical Anthropology», в котором в 70-80-е гг. публикациям результатов изучения кожного рельефа отводилось значительное место. На протяжении 1990-х гг. число таких статей неуклонно снижалось, и наконец, в материалах 2000-х гг. дерматоглифические публикации представлены единичными работами, подавляющая часть которых не имеет прямого отношения к этногенетической проблематике.
За рубежом наиболее активно дерматоглифические исследования популяций сегодня проводятся в странах Азии, Латинской Америки, Африки, но прежде всего - в странах бывшего СССР. В европейских странах, а также в США большинство публикаций посвящено генетическим аспектам изучения дерматоглифики, связям папиллярных линий и различных типов заболеваний, которые могли бы быть использованы при диагностике последних. Изучение дерматоглифических признаков, которое (в силу специфики изучаемого объекта) не имеет эмпирически верифицируемого диахронного направления, постепенно вытесняется исследованиями в области популяционной генетики.
Перефразируя слова из доклада В.В. Бунака 1972 года, касающиеся антропологии в целом, можно сказать, что дерматоглифические материалы могут быть использованы лишь для подтверждения или опровержения гипотез о путях формирования этнических групп, тогда как содержание самих гипотез определяется историческими материалами. Кроме того, отсутствие диахронного среза не позволяет антропологам заниматься надежно аргументированными реконструкциями дерматоглифических комплексов и их географических ареалов в далеком прошлом. Однако дерматоглифисты, оперируя данными об исторически сложившихся к настоящему времени комплексах признаков, имеют возможность проведения сравнительных анализов в рамках расово-
274

соматологических, лингвистических и этнических классификаций человеческих сообществ, обусловленных историческими и географическими факторами не в меньшей степени, чем признаки дерматоглифики. Вклад дерматоглифических исследований в разработку этноисторической проблематики за последние несколько десятилетий нуждается в специальном освещении как в плане обобщающих работ, так и отдельных народов или их групп.
11.1. Дерматоглифическая дифференциация в пределах языковых групп.
Изучение дерматоглифической дифференциации в пределах языковых групп Евразии неоднократно становилось целью специальных исследований. Во всех случаях картина дифференциации не согласуется с языковыми классификациями, поскольку в значительной степени является отражением расогенетических процессов на территории Евразии, никогда не связанных с лингвогенезом непосредственно. Различия между языковыми и дерматоглифическими (выделяемыми в результате статистических операций) группами являются иллюстрацией одного из основных тезисов популяционной антропологии: между языком, с одной стороны, и антропологическим типом народа, с другой, нет причинно-следственной связи.
Финно-угорская группа уральской языковой семьи
Результаты исследования 176 локальных выборок, представляющих основные финно-угорские народы, позволили прийти к выводу, что по типу комбинации ключевых признаков финно-угры образуют ряд, в пределах которого максимально европейские формы (характеризующие финнов, карел, эстонцев, мордву, коми, вепсов, венгров) через промежуточные варианты (отмеченные для марийцев, удмуртов, саамов и манси) постепенно переходят в типично сибирские (у хантов).
Различными методами в составе финно-угорских популяций выделено три типа - североевропеоидный, метисный (самый многочисленный, с увеличенной долей монголоидного компонента) и монголоидный западно-сибирский, представленный выборкой хантов. Картина дифференциации в пределах каждого пола практически полностью определяется различиями в соотношении
275

европеоидного и монголоидного компонентов. Кроме этого, в пределах форм с преобладающим европеоидным компонентом выделяются этнические группы с повышенной долей североевропеоидного компонента: карелы, финны и вепсы.Минимальные значения северного компонента выявлены у венгров и мордвы. Гетерогенность финно-угров проявляется в величине средней межэтнической ОДР, которая составляет 11.5 у мужчин и 12.5 у женщин, и в резко отличающихся типах комбинаций признаков (Хить, Долинова, 2000). Дифференциация финноугорских народов по признакам дерматоглифики служит наглядной моделью расообразовательного процесса на территории значительной части Северной Евразии, в которой ведущим элементом расогенеза является метисация.
Альтернативное исследование дерматоглифической дифференциации финно-угорских народов на том же материале, но значительно уменьшенном по объему выборок, было проведено Р. Дженцем с применением другой программы. Исследовались гребневой счет пальцевых узоров, гребневой счет междупальцевыми трирадиусами, типы пальцевых узоров, узорность гипотенара и тенара, типы окончаний основных ладонных линий, а также группа пяти ключевых признаков.
Наиболее высокий уровень корреляций данныхдерматоглифики, языка и географии был получен по результатам анализа пальцевых узоров (по величине гребневого счета и частотам разных типов). В наименьшей степени соответствуют языковой и географической дифференциации финно-угров результаты,полученные в ходе анализа данных по гребневому счету между пальцевыми трирадиусами, а также для группы ключевых признаков (Jantz et al., 1992). Следует отметить, что даже в случае анализа гребневого счета пальцев коэффициент корреляции между дерматоглифическими данными и лингвистической классификацией не превысил 0.3-0.4. Система взаимоотношений между выборками не исследовалась, подчеркивались лишь единичные факты статистического сходства между отдельными народами. На основании корреляций между дерматоглифической, языковой и территориальной дифференциацией по данным разных систем был сделан вывод о большей эффективности примененного подхода по сравнению с публикацией Г.Л. Хить (Heet, 1990).
Поскольку ни географическая локализация, ни языковая принадлежность не связаны с реальной картиной антропологической
276

дифференциации финно-угров, представляется некорректным (как это делают Дженц и Бреме) сравнивать разграничительную способность различных подходов и систем дерматоглифических признаков на данных материалах. С методической точки зрения более правильным было бы сопоставить варианты дерматоглифической и генетической дифференциации финно-угров (либо языковых групп, менее разнородных в антропологическом отношении). Впрочем, заинтересованный читатель может самостоятельно оценить сравнительную эффективность двух различных подходов при анализе одной и той же проблемы, ознакомившись с указанными оригинальными публикациями.
Тюркская группа алтайской языковой семьи
В основу наиболее полного к настоящему времени анализа дерматоглифической дифференциации тюркских народов легли данные 40 мужских этнических выборок России, Кавказа, Средней Азии, Болгарии, Молдавии, Турции и Монголии (Хить, 2009). Эта дифференциация в большинстве случаев не находит соответствия в лингвистической классификации. В некоторых случаях, однако, наблюдается частичное совпадение данных дерматоглифики и лингвистики. Так, выделяется огузский вариант тюркоязычных групп (туркмены, гагаузы, азербайджанцы, турки), относящийся к западной ветви тюркских народов. Частично сближаются между собой кыпчакские группы: башкиры и ногайцы, с одной стороны, казахи и каракалпаки - с другой.
Однако все случаи дерматоглифического объединения лингвистически родственных групп базируются на их антропологическом (расово-соматологическом) сходстве. Дерматоглифический комплекс тюркоязычных народов Евразии формировался на основе местного европеоидного, в основном южного, субстрата, на который наслаивались пласты как центральноазиатского, так и сибирского происхождения. Как и в случае финно-угров, применением разных методов были выделены три варианта: южноевропеоидный (карачаевцы, балкарцы, гагаузы, азербайджанцы, турки, кумыки), монголоидный (долганы Ямала) и метисный (куда входят все остальные группы). Последний вариант сближается с дерматоглифическими типами уральской и южносибирской рас, отличаясь от них несколько меньшей долей европеоидного компонента. По степени гетерогенности тюрки
277

превосходят финно-угров и славян (средние величины межэтнических ОДР составляют 15.8 у мужчин и 11.5 у женщин).
Результаты анализа связей между лингвистической и дерматоглифической дифференциацией тюрков, осуществленного казахскими исследователями на менее обширном материале, также показали отсутствие параллелизма между обеими системами (Исмагулов и др., 2007, с.192-199).
Северомонгольская группа языковой алтайской семьи
Изучено 10 мужских и 9 женских выборок монголоязычного населения Евразии (Хить, 1983). Монголоязычные народы в целом характеризуются чертами кожного рельефа, свойственного монголоидному расовому стволу. Вместе с тем, среди них наблюдается гетерогенность, имеющая направленный характер. Она связана с вошедшими в их состав расовыми компонентами, причем каждый из рассматриваемых народов по комплексу признаков оказывается ближе к другим, немонгольским народам, чем друг к другу. Буряты ближе всего к сибирскому населению (в первую очередь к якутам, долганам, нганасанам, эвенкам). Монголы комбинируют черты центральносибирских и дальневосточных монголоидов и максимально приближаются к общемонголоидной выборке, а в ее пределах - к населению Алтая, Тувы и Дальнего Востока. Калмыки наиболее своеобразны, имея заметную европеоидную примесь в своем составе, и сходны с хакасами и западносибирскими народами.
Разнородность монголоязычных народов может быть связана с влиянием европеоидного компонента, древний вариант которого прослеживается не только в составе калмыков, но и у части монголов, забайкальских бурят, юго-западных тувинцев, а также в обширном круге популяций Алтая, Хакасии и Западной Сибири.
Таким образом, центральноазиатская расовая общность, ядро которой оставляют монголоязычные народы, не может считаться однородной по кожным узорам кисти. Составляющие ее народы различаются как по удельному весу, так и по происхождению преобладающего монголоидного компонента и европеоидной примеси (Хить, 1983).
278

Славянская группа индоевропейской языковой семьи
Обобщающая работа по дерматоглифике восточных славян была выполнена Н.А. Долиновой (1999). В исследовании были использованы данные 78 мужских и женских параллельно изученных выборок русских, украинцев и белорусов. Результаты сопоставления восточнославянских народов свидетельствуют об их общем сходстве по признакам кожного рельефа. Средние межэтнические показатели ОДР у славян составляют 7.0 и 10.2 для мужчин и женщин соответственно, что свидетельствует об их относительной гомогенности, отчетливо проявляющейся при сравнении с финно-угорскими и особенно тюркскими народами.Н.А. Долиновой не удалось выявить направленные различия между территориальными группами восточных славян. Южно- и североевропеоидный компоненты, по ее мнению, выделяются лишь в качестве тенденции территориальной дифференциации, не согласующейся по данным анализа мужских и женских выборок.
Исследование Е.В. Балановской и О.П. Балановского, выполненное на тех же материалах, позволило скорректировать последний вывод. Картографирование результатов анализа, осуществленного методом главных компонент, показывает, что признаки дерматоглифики в выборках русских имеют выраженную географическую изменчивость по вектору север - юг (Балановская, Балановский 2007, с. 77-78, 380).
11.2. Обзор дерматоглифических исследований в области этнической истории отдельных народов
Ослабление интереса к дерматоглифическим исследованиям в области расоведения, отмеченное для многих стран, почти не затронуло отечественную антропологию: за прошедшие 25 лет опубликовано значительное количество работ по расовой и этнической дерматоглифике народов России и сопредельных стран. В их числе следует, в первую очередь, отметить монографию Г.Л. Хить и Н.А. Долиновой (1990), содержащую самую большую сводку данных, обзор и обобщение сведений по дерматоглифике народов мира. На основе материалов по репрезентативным выборкам с использованием сравнимых данных разных исследователей по комплексу ключевых признаков этими авторами разработаны таксономические схемы, охватывающие все
279

человечество (Хить, Долинова, 2002). Рассмотрен также широкий круг проблем, связанных с применением данных дерматоглифики в расоведении (Хить, 2004в).
Ниже приводится краткий обзор основных публикаций по теме за последние четверть века. Сведения об отечественных и зарубежных работах, не вошедших в данный обзор, содержатся в Таблице 11.1, которая, разумеется, не может считаться исчерпывающей.
Таблица 11.1. Список некоторых отечественных и зарубежных публикаций по расовой и этнической дерматоглифике, вышедших за последние четверть века.Table 11.1. The list of some publications dedicated to racial and ethnic dermatoglyphics over a period of 25 years.
Регион Группа Авторы Исследованные признаки
Европейская удмурты Долинова, 1989 Dl10, Ic, t, Hy, ДМТ, Th/Iчасть России
коми-зыряне,коми-пермяки
Долинова, 2005 Dl10, Ic, t, Hy, ДМТ, Th/I
чуваши Лебедева,Славолюбова,2009;Лебедева, 2009
Dl10, Ic, t, Hy, ДМТ, Th/I
пальцевые узоры (A, L, W)
Кавказ ногайцы Хить, 2003 Dl10, Ic, t, Hy, ДМТ, Th/I
курды, турки- месхетинцы, горские евреи
Хить, 2010 Dl10, Ic, t, Hy, ДМТ, Th/I
Сибирь чулымцы Хить, 1991 Dl10, Ic, t, Hy, ДМТ, Th/I
нганасаны Хить, 1992 Dl10, Ic, t, Hy, ДМТ, Th/I
якуты Хить, Долинова, 2002
Dl10, Ic, t, Hy, ДМТ, Th/I
Средняя иЦентральнаяАзия
киргизы Хить,Сихимбаева,1989
Dl10, Ic, t, Hy, ДМТ, Th/I
арабы Хить, 2010 Dl10, Ic, t, Hy, ДМТ, Th/I
народыМонголии
Дашцэвэгийн,1990
Dl10, Ic, t, Hy, ДМТ, Th/I
280

Регион Группа Авторы Исследованные признаки
Передняя Азия 13 групп Ирана (арабы, армяне, ассирийцы, туркмены, зороастрийцы, курды и др.)
Kamali, 1991
Kamali,Mavalwala, 1990
Мд, пальцевые узоры (A, Lr, Lu, W), ладонные узоры, PPI
TPPRC, RC(Hy), RC(III), RC (IV)
персы Хить, 2010 Dl10, Ic, t, Hy, ДМТ, Th/I
курды Хить, 2010 Dl10, Ic, t, Hy, ДМТ, Th/I
Южная иВосточнаяАзия
56 этнических групп Китая
Zhang et al., 2010
TFRC, FC(a-b), пальцевые узоры (A, Lr, Lu, W), истинные узоры ладони (Th/I, Hy, II, III, IV)
кхмерыВьетнама
Хить, Долинова, 2002
Dl10, Ic, t, Hy, ДМТ, Th/I
дунгане, карены Бирмы, японцы, мео, балийцы, тибетцы, флоресцы
Хить, 2010 Dl10, Ic, t, Hy, ДМТ, Th/I
группы Индии Bhasin, 2007 TFRC, пальцевые узоры (A, Lr, Lu, W), истинные узоры ладони (Th/I, Hy, II, III, IV)
кам и хань Китая
Cheng et al. 2008 Пальцевые узоры (As, At, Lr, Lu, Ws, Wd).
5 группнегритосовМалайзии
Ismail et al. 2009 TFRC, пальцевые узоры (W, Lr, Lu, A), atd, FC(a-b)
3 группы гондов Орисса (Индия)
Choudhury, 2005 Пальцевые узоры (W, L, A)
12 групп СевероЗападной Индии
Reddy et al., 2000
FRC, PPRC
2 группыСевернойБенгалии
Sen et al., 2011 Истинные узоры ладони (Th/I, Hy, II, III, IV), формулы главных ладонных линий
Европа болгары Kavgazova et al., 1999
Dl10, Ic, t, Hy, Th/I, ДМТ
281

Регион Группа Авторы Исследованные признаки
3 группы словаков
Weisensee, Sivakova, 2003
RRC
сербы, венгры,болгары,хорваты,македонцы,румыны,словаки, греки
Scheil et al., 2005
Пальцевые узоры (A, W, Lr, Lu), TFRC, истинные и следовые узоры ладони (Th/I, Hy, II, III, IV), Ic, варинты окончания линии С, FC(a-b)
литовцы Клевцова, Кучинскас, 1988
Dl10, Ic, t, Hy, Th/I, ДМТ
поляки и чехи Hajn,Gasiorowski,1999
FRC, Ta-dRC, RC(II), RC (III), RC (IV)
4 группы сардинцев
Sana et al., 2005 FRC
испанцы Rosa, Arqumbai, Fananas, 1998
FC(a-b) , узоры ладони (Th/I, Hy, II, III, IV), MD
6 групп хорват MiliCiC, Ivicevic, 2005
Пальцевые узоры (A, W, Lr, Lu), истинные и следовые узоры ладони (Th/I, Hy, II, III, IV), варианты осевого трирадиуса (t, t', t" и др.)
словенцы Milicic, Vidovic, 2005
FRC, atd, FC (a-b), FC(b-c),FC (c-d)
албанцы и турки Косово
Temaj et al., 2009
-c)(b-C(-b)
О
td, at , d)
C, (c-d
R C(
[Jn [Jn
население 17 деревень Болоньи (Италия)
Gualdi-Russo et al., 1994
Пальцевые узоры (A, W, Lr, Lu), FRC, t, t', t", FC (a-
b ), Ic
бельгийцы,датчане,англичане
Хить, 2010 Dl10, Ic, t, Hy, Th/I, ДМТ
Америка 6 групп индейцев чибча
Segura-Wang, Barrantes, 2009
Пальцевые узоры (A, W, Lr, Lu), истинные и следовые узоры ладони (Th/I, Hy, II, III, IV), TFRC, Ic, atd, FC (a-b),FC(b-c> FC (c-d)
282

Регион Группа Авторы Исследованные признаки
индейцы чороте Demarchi, Seisdedos 1996
TFRC, пальцевые узоры (W, Lr, Lu, A), истинные и следовые узоры ладони (Th/I, Hy, II, III, IV), главные ладонные линии С и D, atd, FC (a-b), FC(b-c),FC (c-d)
17 групп индейцев Юж. Америки
Demarchi, Marcellino, 1998
Пальцевые узоры (A, W, Lr, Lu),
атапаски, миштеки, кашинауа, трио, уастеки
Хить, Долинова, 2002
Dl10, Ic, t, Hy, Th/I, ДМТ
Африка берберы Sabir et al., 2005 Пальцевые узоры (As, At, Lr, Lu, Ws, Wd).
6 групп Туниса (берберы, смеш. население)
Namouchi, 2011 Пальцевые узоры (W, L, A), TFRC, PPRC
луимбе, тсвана, шангаан, педи
Хить, Долинова, 2002
Dl10, Ic, t, Hy, Th/I, ДМТ
3 группынубийцевЕгипта
Pospisil, Ferak, 1990
Пальцевые узоры (Lu, Lr, W, CP, TL, A, At), LFRC, RFRC, TRFC, типы главной ладонной линии D, узоры II
2 группы Нигерии
Osunwoke et al., 2008
Пальцевые узоры (W, Lr, Lu, A), atd, dat
Танзанийцы и кенийцы
Igbigbi, Msamati, 2005
TFRC, FC (a-b), atd, пальцевые узоры (A, Lr, Lu, W),
малавийцы Igbigbi, Msamati, 1999
TFRC, FC (a-b), atd, пальцевые узоры (A, Lr, Lu, W),
Австралия и Океания
австралийцыАрнхемленда
Cho, 2000 Пальцевые узоры (As, At, Lr, Lu, Ws, Wd, Wcp).
австралийцы Арнхемленда, папуасы Новой Гвинеи (бунди, амеле, хагахаи)
Хить, Долинова, 2002
Dl10, Ic, t, Hy, Th/I, ДМТ
283

Регион Группа Авторы Исследованные признаки
маори Cho, 1998a Пальцевые узоры (As, At, Lr, Lu, Ws, Wd, Wcp).
самоанцы Cho, 1998b Пальцевые узоры (As, At, Lr, Lu, Ws, Wd, Wcp), TFRC
4 группы полинезийцев
Katayama, 1990 Пальцевые узоры (A, Lr, Lu, W), узоры Hy и Th/I, ДМТ, типы главной ладонной линии С
микронезийцы Fox, Garruto, Gajdusek, Plato, 1990
Пальцевые узоры (A, Lr, Lu, W), типы главной ладонной линии D, узоры ладони (Th/I, Hy, II, III, IV), добавочные ладонные трирадиусы, флексорные складки
Примечания: FRC - гребневой счет; TFRC - общий гребневой счет пальцев; FC(a-b), FC(b-c), FC(c-d) - гребневой счет между пальцевымитрирадиусами a и b, b и c, c и d; PPI - общее число трирадиусов ладони; TPPRC - общий гребневой счет ладони.Note: FRC - finger ridge count; TFRC - total finger ridge count; FC(a-b), FC(b- c), FC(c-d) - finger count between triradii a and b, b and c, c and d, respectively; PPI - palmar pattern index; TPPRC - total palmar pattern ridge count.
Целый ряд работ посвящен отдельным регионам, народам или территориальным группам. Так, большой материал по дерматоглифике белорусов и населения сопредельных территорий собран и систематизирован Л.И. Тегако (1989). Основные проблемы дерматоглифики народов Средней Азии, включая характеристику и расогенетические связи аборигенных и пришлых народов, рассмотрены Г.Л. Хить (1991). Среднеазиатские народы подробно изучены Г.К. Ходжайовой (1997, 2007): рассмотренадифференциация на территориальном, этническом и расовом уровнях, выявлены различия в распределении признаков по историческим культурным областям, а также между городским и сельским населением. Детально исследованы народы Западного Памира (Бадахшана): установлены их местные, среднеазиатские корни и генетические связи с населением Индии, Афганистана и Ирана (Хить, 2005). В подробностях изучены туркменские популяции (Хить, Долинова, 1989). На основе обширных данных
284

дерматоглифики воссоздана этническая история казахов в монографии О. Исмагулова, К. Сихымбаевой, А. Исмагуловой (2007). Эти авторы проанализировали материалы по локальным группам, а также субэтническим и региональным подразделениям казахов. Проведено сравнение казахов с другими тюркоязычными народами. Некоторые группы североазиатских монголоидов рассмотрены в монографии O.K. Галактионова (2008).
Н.А. Суворовой (2005) проведены масштабные исследования башкирского народа. Изучено географическое распределение показателей дерматоглифики у башкир, проведено сопоставление территориальных, племенных и историко-географических групп башкир, рассмотрена проблема этногенеза башкирского народа.
Изучению антропологического состава и этнической истории карел посвящены работы И.Г. Широбокова (2009а; 2010а). Анализ дерматоглифических характеристик в территориальных и этнолокальных группах карел, проведенный на обширном материале, позволил выделить устойчивые комплексы признаков, связанные с различным происхождением рассмотренных групп. Установлено соответствие основных направлений изменчивости дерматоглифических комплексов у карел с соматологическими и краниологическими системами признаков.
На материале отечественных и зарубежных экспедиций рассмотрена дерматоглифика трех саамских групп Финляндии и России, выявлены восточные истоки происхождения саамов (Хить, 1991в). Проанализированы обширные материалы о кожном рельефе народов Чукотки и Камчатки (Долинова, Хить, Балуева, 2002), понтийских греков (Хить, 1990б; 2004а). В подробностяхрассмотрена дерматоглифика финноязычных народов циркумбалтийского региона (Хить, Долинова, 2001), мордвы (Хить, 2008), татарских групп Евразии (Хить, Долинова, 1995) и сибирских татар, в частности (Хить, 1990а; Томилов, Хить, 1991). Детально изучены этносы и популяции важнейшего расогенетического узла Евразии - Поволжья и Приуралья (Хить, 2004г). Введены в научный оборот данные по арабам Йемена и сокотрийцам (Хить, Шинкаренко, Наумкин, 1988; 1995; Гохман, Богданова, Чистов, 1995). В связи с проблемой происхождения уральской расы рассмотрены материалы по уграм Оби (Хить и др., 1996).
Широкий спектр проблем дерматоглифики, в том числе расовой и этнической, отражен в первом на русском языке
285

энциклопедическом словаре по дерматоглифике (Звягин, Ракитин, 2012).
К сожалению, из зарубежных публикаций последних десятилетий практически невозможно извлечь методически сопоставимые сравнительные данные. Исключение составляют частоты типов пальцевых узоров и гребневой счет, которые являются обязательным элементом практически всех работ. В 2000-е гг. из публикаций почти полностью исчезли данные по частотам добавочных межпальцевых трирадиусов и осевого трирадиуса t. Крайне редко публикуются данные по частотам типов окончаний главных линий, на основании которых можно было бы рассчитать приблизительную величину индекса Камминса. Частоты узорности гипотенара и тенара нередко приводятся суммарно со следами узоров, что также не позволяет использовать эти данные в сравнительном анализе.
Кроме того, огромное число статей принадлежит не специалистам-антропологам, а представителям смежных профессий, увлекшимся кажущейся легкостью работы с отпечатками. Эти авторы слабо знакомы с антропологической проблематикой, недостаточно ориентируются в специфике признаков кожного рельефа (включая топологические аспекты и систему взаимных корреляций) и закономерностях их варьирования в выборках разного таксономического ранга, игнорируют половые различия между выборками (в связи с чем часто суммируются данные обоих полов), и т.д.
Приведенный обзор демонстрирует существенность вклада, вносимого данными дерматоглифики при изучении процесса расогенеза как на примере огромных массивов популяций, так и отдельных народов. Остается надеяться, что традиция включения дерматоглифики в программу исследования сохранится в дальнейшем.
286

%

Г лава 12. Соотношение данных дерматоглифики и других систем антропологических признаков
Внутригрупповые корреляции.
Отсутствие существенных и устойчивых внутригрупповых (на уровне индивидов) связей между признаками разных систем - один из важнейших постулатов расового анализа, выработанных отечественной антропологической школой (Ярхо, 1932; 1947;Рогинский, Левин, 1978).
Корреляции между признаками дерматоглифики и других систем изучались в различных группах населения Евразии. У финнов Финляндии редкие значимые связи (порядка 0.2-0.3) свидетельствуют о параллельном проявлении специфического расового комплекса (например, усилении черт северных европеоидов). С другой стороны, у якутов значимые корреляции (того же порядка) более многочисленны и типы сочетаний признаков другие. Направление связей у якутов таково, что можно говорить о наличии в их составе двух расовых компонентов - с усиленной и с ослабленной монголоидной основой (Хить, 1975). Ж. Оливье на большой выборке (2 тыс. мужчин) французов обнаружил слабые значимые корреляции, позволившие выделить два комплекса: один с пониженным дельтовым индексом, более высокорослый, с более крупной головой удлиненной формы, более смуглокожий, и второй с противоположным сочетанием признаков (Olivier, 1967). При изучении сомато-дерматоглифических зависимостей в сборных группах русских юношей и девушек выявлено 25% значимых корреляций низкого уровня (0.3). Значимые связи с некоторыми размерами головы, лица (лобным и скуловым диаметрами, толщиной обеих губ) обнаруживает гребневой счет пальцев рук. Кроме того, найдены связи между типами пальцевых узоров и диаметрами головы (продольным и поперечным), высотой носа, верхней губы и шириной рта (Негашева, 2007а, б).
Набор наиболее таксономически ценных элементов дерматоглифики и одонтологии был изучен в нескольких европейских и азиатских популяциях. Так, в дисперсной группе белорусов значимые связи обнаружены в единичных случаях как в мужской, так и в женской группе (0.2-0.3). Эти нестойкие связи,
288

различные у обоих полов, говорят о слабой тенденции к параллельному проявлению в группе особенностей кожного и зубного рельефа, свойственного популяциям западного расового ствола (Тегако, 1978, с. 133-136). У венгров Закарпатья наблюдается сходная картина. В то же время у бурят и хакасов значимые связи встречаются чаще, чем у венгров и белорусов, и свидетельствуют о расовоконтрастном сочетании признаков обеих систем как у мужчин, так и у женщин (Хить, Халдеева, 1976).
В целом можно констатировать отсутствие либо наличие слабых и неустойчивых корреляций между признаками разных систем на внутригрупповом (индивидуальном) уровне. Таким образом, тесной биологической связи между этими признаками нет, все системы признаков автономны и следуют своим законам развития.
Межгрупповые корреляции.
Варианты дифференциации популяций, основанные на материалах разных антропологических систем (соматологии, одонтологии, дерматоглифики, краниологии, генетики и др.), нередко существенно различаются между собой, в том числе на высших таксономических уровнях. Традиционно в качестве причины расхождений указывается на то, что каждая система признаков имеет свою специфику, время и ареал формирования. Очевидно также, что многие анализируемые морфологические признаки в рамках любой системы являются искусственно выделенными объектами. Это обстоятельство приводит не только к методическим различиям между авторами, но и к артефактам, возникающим при сопоставлении картин изменчивости разных систем признаков.
Тем не менее, определенную степень совпадения между результатами анализа дерматоглифических и соматологических данных можно наблюдать на уровне больших рас (Хить, 1975; 1983; Хить, Долинова, 1990). Однако расово-соматологические и другие классификации не основаны на применении многомерного анализа и имеют преимущественно описательный (и часто интуитивный) характер, различаясь в существенных деталях у разных авторов. Так,А.А. Зубов придерживался дуальной схемы, разделяя человечество по строению зубной системы на два ствола - западный и восточный, причем первый включает европеоидов и негроидов, второй -
289

монголоидов, австралоидов и американоидов (Зубов, 1968, 1977). На ту же схему, основанную на данных краниологии и соматологии, опирался В.П. Алексеев, относивший, однако, австралоидов к западному стволу (Алексеев, 1974; 1985). Анализдерматоглифических данных, в том числе методами многомерной статистики (не использованной указанными авторами), привел к выделению в пределах человечества трех стволов: западного (европеоиды), южного (негроиды) и восточного (американоиды, монголоиды, австралоиды) (Хить, Кейта, 1981; Хить, Долинова, 1990; 2002). В описательной форме такое разделение рас было впервые предложено К. Плато (Plato, 1976).
12.1. Дерматоглифика и расовая соматология
При анализе расового состава обобщенных групп населения крупных историко-географических регионов бывшего СССР (европейской части, Кавказа, Средней Азии и Сибири) обнаружено совпадение дифференциации по данным обеих систем признаков (Хить, 1973; 1983). Однако на более низком таксономическом уровне, где дерматоглифический материал был организован в рамках расово-соматологической классификации и распределен по локальным расам, возникли расхождения. На материалах дерматоглифики впервые была получена методом числовой таксономии схема локальных рас, основанная на результатах многомерного анализа и охватывающая все население страны и прилегающих территорий Северной Евразии. Выявлено два основных компонента - европеоидный и монголоидный, наряду с компонентами метисного происхождения. Расхождения этой схемы с данными расовой соматологии носят как частный, так и принципиальный характер (Хить, 1983). Детальное обсуждение несовпадений дерматоглифической и соматологической картин дифференциации представляется излишним, поскольку, как указывалось выше, существующие соматологические схемы носят описательный либо интуитивный характер и, кроме того, различаются у разных авторов.
На региональном уровне была предпринята попытка сопоставления основанных на числовых расчетах схем, отражающих дифференциацию народов Кавказа по признакам дерматоглифики (данные Г.Л. Хить) и расовой соматологии (данные М.Г.
290

Абдушелишвили). Результаты кластерного анализа обеих таблиц расстояний между группами совпали лишь частично (Рис. 12.1).
Явным доказательством отражения исторических процессов на признаках обеих систем служит стойкое выделение «адыгского» кластера (в который вошли адыгейцы, черкесы, абазины, кабардинцы, абхазы на дерматоглифической дендрограмме, и адыгейцы, кабардинцы, черкесы, абазины - на соматологической), а также объединение карачаевцев и балкарцев на уровне большого сходства. В остальном обе схемы не совпадают, особенно в случае положения абхазов: по дерматоглифике они теснейшим образом сближены с кабардинцами и входят в «адыгский» кластер, в то время как по соматологии они образуют с армянами отдельный кластер, уступающий по степени обособленности лишь азербайджанцам (Хить, 2004).
Рисунок 12.1. Сопоставление дерматоглифической и соматологической дифференциации кавказских этносов по результатам кластеризации матриц ОДР и КМД (коэффициент морфологической дистанции по М.Г. Абдушелишвили) соответственно.Figure 12.1. Comparison between dermatoglyphic and somatological differentiations for Caucasoid ethnic groups (results of clusterization of GDD matrix and matrix of coefficients of morphological distances introduced by M.G. Abdushelishvili).
291

Рисунок 12.2. Вариации величины европеодно-монголоидного комплекса (основа - финно-угорская шкала изменчивости) по данным соматологии (белые кружки) и дерматоглифики (черные кружки) в мужских группах Восточной Европы и Западной Сибири (по: Хить, 1974, с учетом новых данных).Figure 12.2. Variation of Mongoloid complex values (Finno-Ugrian range) in male groups from Eastern Europe and Western Siberia based on dermatoglyphic (black circles) and somatological (white circles) data (from: Heet, 1974, new data).
292

Проведено также межпопуляционное сравнение многих народов европейской части России и Западной Сибири (Рис. 12.2). Одинаковым способом вычислялось содержание монголоидного компонента посредством расчета величины монголоидного комплекса по признакам дерматоглифики (данные Г.Л. Хить) и соматологии (данные К.Ю. Марк) с использованием «финноугорской» шкалы вариаций.
Обнаружено, что этнические группы по обоим показателям располагаются в одинаковом порядке относительно друг друга, что говорит о согласованной реакции групп на монголоидную примесь. Кроме того, вариации дерматоглифического ЕМК в пределах этносов сдвинуты в сторону увеличения, что свидетельствует о более выраженной реакции дерматоглифической системы на эту примесь в популяции. Реальным подтверждением данного феномена служит соотношение комплексов у шведов, финнов и саамов, где по обеим системам признаков параллельно обследовались одни и те же индивиды (Хить, 1974). Во всех остальных случаях речь идет не только о разных индивидах в пределах выборки, но и о разных территориальных группах народа, изученных по обеим системам признаков.
Одним из авторов данной работы при помощи вычисления ранговых коэффициентов Спирмена было проведено межгрупповое сопоставление данных дерматоглифики и соматологии по материалам локальных мужских карельских и вепсских выборок, принадлежащих к этническим группам с максимальными значениями североевропеоидного комплекса (Широбоков, 2010б). К анализу были привлечены соматологические данные М.В. Витова (1997) по карелам Петровского (Кондопожского), Сегозерского (Медвежьегорского), Олонецкого, Пряжинского, Калевальского, Кестеньгского (Лоухского) районов Карелии и Лихославльского района Тверской области, оштинским (Вологодская область) и шелтозерским вепсам, а также дерматоглифические выборки из соответствующих районов. Каждая из выборок в обеих системах признаков включает не менее 100 человек. Всего были сопоставлены данные 9 территориально-локальных групп.
Рассмотрим выявленные связи между признаками в контексте изменчивости дерматоглифического северо-европеоидного комплекса, или СЕК (Табл. 12.1). При повышении СЕК (определяемом понижением значений Ic и Dl10 и повышением частоты t) высота лица и носа, а также продольный диаметр головы
293

в карельских и вепсских группах уменьшаются, возрастает скуловой диаметр, уменьшается процент русых волос и увеличивается процент темных оттенков глаз. Соответственно, с понижением СЕК указанные признаки меняются обратным образом. При этом следует отметить, что в пределах восточнобалтийских групп складывается парадоксальная ситуация: наиболее депигментированные группы обладают наименьшей величиной СЕК. Более того, повышение доли североевропеоидного комплекса в выборках отчетливо связано с проявлением урало-лапоноидного комплекса соматологических признаков. По данным дерматоглифики вепсы характеризуются максимально выраженной в регионе величиной СЕК. С другой стороны, Г.А. Аксянова отметила некоторое сближение характеристик вепсов и саамов и определенное структурное сходство лапоноидного комплекса (при всей его специфичности) с прионежским вариантом восточнобалтийского типа (Аксянова, 2004, с.306). Близкие к вепсским значения СЕК обнаруживаются в группах южных карел, тогда как к северу Карелии комплекс уменьшается и вновь достигает высоких значений уже в саамских популяциях. Возрастание СЕК у карел следует рассматривать как следствие контактов с вепсскими группами, а низкий СЕК в группах северной Карелии, по всей вероятности, связан с отсутствием или низкой долей в их составе саамского компонента, а не присутствием каких- то элементов южноевропеоидного происхождения.
Таблица. 12.1. Значимые межгрупповые корреляции между ключевыми дерматоглифическими и некоторыми соматологическими признаками по материалам карельских и вепсских выборок (по: Широбоков, 2010б).Table 12.1. Significant inter-group correlations between key traits and some somatological features in Karelian and Vepsian groups (from: Shirobokov, 2010)
Признаки Dlio Ic t Ну ДМТ
Продольный диаметр 0.76 - - - -Скуловой диаметр - -0.67 - - -Физиономическая высота лица 0.95 - - - -Высота носа (от нижнего края бровей) - - -0.75 - -Процент русых волос (по Фишеру) - 0.87 - - -Процент темных оттенков глаз - - 0.68 - -
Коэффициенты значимы от р<0.05.
294

Связь между условным показателем «монголоидности» (ЕМК) по расово-соматологическим и дерматоглифическим признакам была изучена у населения Финляндии (Хить, 1982). В пределах 16 популяций финнов и шведов обе системы признаков оказались взаимонезависимыми (rs=0.53, p>0.05). Однако при включении в анализ двух саамских выборок корреляции между системами увеличиваются и достигают значимого уровня (rs=0.67, p<0.01). Таким образом, при учете саамских групп возникает параллелизм в реакции обеих систем признаков на присутствие инородного компонента, который свидетельствует об ассимиляции финнами и шведами части саамов, оттеснявшихся на восток и северо-восток страны.
12.2. Дерматоглифика и краниология: интеграция данных.
Воссоздание истории человеческих популяций невозможно на основании результатов анализа только одной программы антропологических признаков. Интеграция данных нескольких независимых систем позволяет учитывать случайные колебания частот признаков в популяциях. Сопоставление данных разного рода признаков, в случае если эти данные разделены небольшим отрезком времени, позволяет выявить наиболее устойчивые исторически сложившиеся антропологические комплексы и проследить основные направления изменчивости.
Наиболее исчерпывающий комплексный анализ был выполненВ.Г. Моисеевым (2004). Он сопоставил данные краниометрии, краниоскопии, одонтологии и дерматоглифики 38 евразийских групп. Было обнаружено, что по дерматоглифике абсолютное большинство выборок демонстрируют устойчивый сдвиг в восточном (т.е. в монголоидном направлении) по отношению к остальным группам. Взаимоотношения последних по признакам других систем признаков хаотичны и разнонаправлены. Отметим, что этот результат был получен автором при включении в анализ, помимо 5 ключевых признаков, обладающих универсальной расово- дифференцирующей способностью, также узорности на Th/I. Напомним, что этот признак не разделяет монголоидов и европеоидов и значительно более вариабелен по сравнению с остальными основными признаками.
Кроме того, И.Г. Широбоковым был проведен анализ 50 мужских этнических групп с территории Северной Евразии:
295

Восточной и Северо-Восточной Европы, Кавказа, Средней Азии и Сибири. Материалами для исследования послужили близкие к современности мужские краниологические серии (XVIII-начала XX века) и дерматоглифические выборки, большая часть которых относится ко второй пол. XX века (Табл. 12.2).
Таблица 12.2. Источники использованных в анализе краниометрических и дерматоглифических данных 50 этнических выборок Северной Евразии (мужчины).Table 12.2. Sources of craniometrical and dermatoglyphic data on 50 ethnic groups from Northern Eurasia (males) used in the analysis.
№ Выборка Краниометрия Дерматоглифика1 адыгейцы Алексеев, 1974 Хить, 19832 кабардинцы Алексеев, 1974 Хить, 19833 балкарцы Алексеев, 1974 Хить, 19834 абхазы Алексеев, 1974 Хить, 19835 осетины Алексеев, 1974 Хить, 19836 ингуши Алексеев, 1974 Хить, 19837 чеченцы Алексеев, 1974 Хить, 19838 аварцы Алексеев, 1974 Хить, 19839 лакцы Алексеев, 1974 Хить, 198310 даргинцы Алексеев, 1974 Хить, 198311 удины Алексеев, 1974 Хить, 198312 хевсуры Алексеев, 1974 Хить, 198313 шведы Алексеев, 1974 Хить, 198314 латыши Алексеев, 1969; Денисова, 1977 Хить, 198315 литовцы Алексеев, 1969 Хить, 198316 финны Хартанович, 1995 Хить, Долинова, 200117 карелы Хартанович, 1986; Широбоков, 2009а18 саамы Хартанович, 1980; 2004;
Алексеев, 1974Хить, 1991
19 эстонцы Алексеев, 1969 Хить, 198320 коми-зыряне Хартанович, 1991 Хить, Долинова, 200421 коми-пермяки Алексеев, 1969 Хить, Долинова, 200422 удмурты Алексеев, 1969 Долинова, 198923 марийцы Алексеев, 1969 Хить, Долинова, 200424 мордва Алексеев, 1969 Хить, 200825 татары
казанскиеАлексеев, 1971 Хить, 1983
26 чуваши Алексеев, 1971 Хить, Долинова, 200427 башкиры Юсупов, 1989 Суворова, 200928 русские Алексеев, 1969 Долинова, 199929 ногайцы Герасимова, 2003; Круц, 2003 Хить, 2003
296

Таблица 12.2. Продолжение. Table 12.2. Continued.
№ Выборка Краниометрия Дерматоглифика30 казахи Исмагулов, 1970 Хить, 198331 киргизы Миклашевская, 1959 Хить, 198332 калмыки Алексеев, 1980 Хить, 198333 теленгеты Исмагулов, 1970 Хить, 198334 тувинцы Исмагулов, 1970 Хить, 198335 монголы Исмагулов, 1970 Хить, 198336 якуты Исмагулов, 1970 Хить, 198337 шорцы Исмагулов, 1970 Хить, 198338 буряты Исмагулов, 1970 Хить, 198339 сагайцы Багашев, 2000 Хить, 198340 качинцы Багашев, 2000 Хить, 198341 селькупы Дремов, 1984 Хить, 198342 ненцы Багашев, 2000 Хить, 198343 кеты Багашев, 2000 Хить, 198344 кызыльцы Багашев, 2000 Хить, 198345 татары сибирские Багашев, 1993 Хить, 198346 телеуты Багашев, 2000 Хить, 198347 кумандинцы Багашев, 2000 Хить, 198348 тубалары Багашев, 2000 Хить, 198349 манси Багашев, 1999; Дебец, 1951 Хить и др., 199650 ханты Дебец, 1951;
Дремов, 1984; 1991Хить и др., 1996
Для анализа в рамках краниологической программы использовались 14 признаков (№№ по Мартину и др.: 1, 8, 17, 9, 45,48, 55, 54, 51, 52, 77, zm, 75(1), SS:SC), в рамкахдерматоглифической программы - 5 ключевых признаков (за исключением Th/I).
Программа межсистемного исследования в рамках применения методов главных компонент и канонического анализа была впервые предложена и апробирована на разного рода антропологических материалах мужских выборок, характеризующих население различных регионов Евразии, А.Г. Козинцевым и В.Г. Моисеевым (Козинцев, Моисеев, 1995; Моисеев, 2004).
На первом этапе данные анализируются в пределах каждой системы признаков. Частоты дерматоглифических и других описательных признаков (соматологических, одонтологических и краниоскопических) обрабатываются методом главных компонент,
297

значения измерительных (остеометрических, краниометрических) - каноническим анализом с использованием стандартной внутригрупповой корреляционной матрицы при помощи программы Б.А. Козинцева (CANON).
Затем значения вновь полученных в рамках каждой из систем признаков (т.е. координаты групп в пространстве переменных) анализируются при помощи метода главных компонент. В результате выделяются интегративные главные компоненты; положение групп в их пространстве определяется различным вкладом данных нескольких систем антропологических признаков.
На первом этапе были отобраны первые две ГК, охватывающие около 81% изменчивости признаков в рамках дерматоглифической системы, а также три канонических вектора, отражающих около 81% изменчивости признаков в рамках краниометрической системы. Значения новых признаков были вновь подвергнуты анализу методом главных компонент.
Полученные первые три интегральные главные компоненты (ИГК) суммарно отражают около 69% изменчивости исходных признаков. Данные по дерматоглифике и краниологии скоррелированы с интегральными главными компонентами примерно в равной степени (Табл. 12.3). При этом вклад дерматоглифических признаков по сравнению с краниометрическими по первым двум ИГК увеличен, тогда как третья ИГК почти полностью определяется краниометрическими признаками.
Таблица 12.3. Коэффициенты корреляции первых трех интегральных главных компонент с исходными векторами, полученными в результате анализа 50 этнических выборок Северной Евразии (мужчины).Table 12.3. Correlation coefficients of 3 integrated principal components and primary canonical vectors and primary principal components received through data analysis of 50 ethnic groups from Northern Eurasia (males).
Исходные векторы Интегральная ГК I
Интегральная ГК II
Интегральная ГК III
КВ 1 (краниометрия) 0.91 -0.30 0.10КВ 2 (краниометрия) -0.20 -0.61 -0.68КВ 3 (краниометрия) -0.40 -0.46 0.69ГК 1 (дерматоглифика) 0.97 0.02 0.05ГК 2 (дерматоглифика) -0.07 0.83 -0.08Собственное число:% общей изменчивости:
2.039.3
1.427.3
0.918.9
298

Главная компонента и канонический вектор, определяющие первую интегральную компоненту (ГК 1 и КВ 2), связаны между собой значимой положительной корреляцией (r=0.85 при p<0.0001). Обе переменные отражают различия между выборками в направлении запад - восток (что соответствует вектору «европеоиды - монголоиды») и отчетливо разграничивают европейские и азиатские группы (Рис. 12.3). Европейские группы (а также адыгейцы) и азиатские выборки по данным дерматоглифики обнаруживают значительную трансгрессию; в переходной области которой располагаются народы Приуралья и Поволжья. Между краниологическими выборками из Европы и Азии почти нет переходных форм. Исключение составляют группы башкир и ногайцев, попавшие в «сибирскую» область значений и отделившиеся от остальных поволжско-приуральских серий.
С другой стороны, кавказские группы (за исключением адыгейцев), отчетливо обособляются от всех остальных серий по признакам дерматоглифики (первая ГК), а по данным краниометрии обнаруживают существенную трансгрессию с европейскими сериями (первый КВ). Значимые связи между остальными исходными переменными, полученными по результатам анализа дерматоглифических и краниологических материалов, отсутствуют.
После процедуры интеграции данных обеих систем были получены следующие результаты (Рис. 12.4).
Первая интегральная ГК разграничивает группы по оси запад - восток. При этом трансгрессии между европейскими и азиатскими группами практически нет, за исключением башкир, оказавшихся в «западно-сибирской» зоне значений. Изменчивость в пространстве ИГК определяется примерно в равной степени как данными краниологии (г между первыми КВ и ИГК составляет 0.91), так и данными дерматоглифики (г между первыми ГК и ИГК равен 0.97). Вторая интегральная ГК определяется в первую очередь второй ГК и вторым КВ, при этом коэффициент корреляции (без учета знака) для дерматоглифического вектора значимо выше, чем для краниометрического (соответствующие г составляют 0.83 и -0.61, при p<0.03). В целом вторая ИГК отражает изменчивость групп в направлении юг - север. При этом североевропейские и североазиатские (западносибирские) группы в значительной степени сближены друг с другом.
299

Рисунок 12.3. Положение 50 этнических групп Северной Евразии в пространстве первой главной компоненты и первого канонического вектора (мужчины). Обозначения выборок: 1 - адыгейцы; 2 -кабардинцы; 3 - балкарцы; 4 - абхазы; 5 - осетины; 6 - ингуши; 7 - чеченцы; 8 - аварцы; 9 - лакцы; 10 - даргинцы; 11 - удины; 12 - хевсуры; 13 - шведы; 14 - латыши; 15 - литовцы; 16 - финны; 17 - карелы; 18 - саамы; 19 - эстонцы; 20 - коми-зыряне; 21 - коми-пермяки; 22 - удмурты; 23 - марийцы; 24 - мордва; 25 - татары казанские; 26 - чуваши; 27 - башкиры; 28 - русские; 29 - ногайцы; 30 - казахи; 31 - киргизы; 32 - калмыки; 33 - теленгеты; 34 - тувинцы; 35 - монголы; 36 - якуты; 37 - шорцы; 38 - буряты; 39 - сагайцы; 40 - качинцы; 41 - селькупы; 42 - ненцы; 43 - кеты; 44 - кызыльцы; 45 - татары сибирские; 46 - телеуты; 47 - кумандинцы; 48 - тубалары; 49 - манси; 50 - ханты. Figure 12.3. Positions of 50 ethnic groups from northern Eurasia on the first principal component and canonical vector (males). Numeration of groups: 1 - Adygei; 2 - Kabardinians; 3 - Balkars; 4 - Abkhazians; 5 - Ossetians; 6 - Ingushs; 7 - Chechens; 8 - Avars; 9 - Laks; 10 - Dargians; 11 - Udins; 12 - Khevsurs; 13 - Swede; 14 - Letts; 15 - Lithuanians; 16 - Finns; 17 - Karelians; 18 - Sami; 19 - Estonians; 20 - Komi Zyrians; 21 - Komi Permyaks; 22 - Udmurts; 23 - Mari; 24 - Mordvinians; 25 - Kazan Tatars; 26 - Chuvashs; 27 - Bashkirs; 28 - Russians; 29 - Nogai; 30 - Kazakhs; 31 - Kirghizs; 32 - Kalmyks; 33 - Telengets; 34 - Tuvinians; 35 - Mongols; 36 - Yakuts; 37 - Shors; 38 - Buryats; 39 - Sagai; 40 - Kachins; 41- Selkups; 42 - Nenets; 43 - Kets; 44 - Kyzyls; 45 - Siberian Tatars; 46 - Teleuts; 47 - Kumandins; 48 - Tubalars; 49 - Mansi; 50 -Khanty.
300

Рисунок 12.4. Положение 50 этнических групп Северной Евразии в пространстве первых двух ИГК. Обозначения выборок соответствует принятым для Рисунка 12.3.Figure 12.4. Positions of 50 ethnic groups from Northern Eurasia on the first two integrated principal components. Groups are numerated as in note of Figure 12.3
Большая часть этих групп относится к уральской языковой семье. Уралоязычные выборки (финны, карелы, саамы, эстонцы, коми-зыряне, коми-пермяки, удмурты, марийцы, мордва, ханты, манси, селькупы, ненцы) по своему положению в пространстве ИГК II на значимом уровне отличаются как от остальных групп в целом (p<0.002), так и от неуралоязычных европейских (p<0.02) и азиатских серий (p<0.002), рассмотренных отдельно.
Наиболее отчетливо различия между уралоязычными группами, с одной стороны, и неуралоязычными европейскими и азиатскими выборками, с другой стороны, проявляются по первой интегральной компоненте (p<0.0006 и p<0.000007, соответственно). Этот результат свидетельствует о наличии большей доли монголоидной примеси в уралоязычных группах относительно неуралоязычных европейцев, а также о присутствии у первых более
301

значительного европеоидного компонента, чем у неуралоязычных азиатских групп. Данное объяснение, однако, не может быть использовано для интерпретации выявленных расхождений по второй компоненте, не выявляющей значимых различий между европеоидными и монголоидными группами.
Следует также отметить значимые различия между уралоязычными и остальными выборками по результатам раздельных исходных анализов. По данным краниометрии такие различия проявляются в пространстве второго КВ (p<0.000001), по данным дерматоглифики - в пространстве второй ГК (p<0.03). Если по краниологическим данным, так же, как и результатам интегративного анализа признаков разных систем (см. например: Козинцев, Моисеев, 1995; Моисеев, 2004), своеобразие уралоязычных групп подчеркивалось исследователями и ранее, то по данным анализа ключевых признаков дерматоглифики такая картина выявлена впервые. При этом дифференциацию групп в пространстве второй ГК определяют в первую очередь дельтовый индекс и частота ДМТ. Оба признака в среднем имеют в уральских сериях меньшую величину по сравнению с их соседями.
Расово-соматологическая дифференциация групп, в отличие от языковых классификаций, является более вариативной и неоднозначной конструкцией, оставаясь традиционной темой дискуссий среди антропологов. Тем не менее, различия выявляются также при сравнении результатов анализа данных уральской и южносибирской рас, значительная роль метисации в формировании современного состава которых признается большинством исследователей. Напомним, что ранее Г.Л. Хить указывала, что дерматоглифические характеристики этих рас близки между собой (при увеличенной монголоидности в Южной Сибири), а их положение на общеевразийском фоне свидетельствует о смешанном европеоидно-монголоидном происхождении данных групп популяций (Хить, 1983; Хить и др., 1996).
Между выборками, в формировании антропологического состава которых роль уральского (саамы, марийцы, чуваши, манси, ханты, ненцы, селькупы и кызыльцы) и южносибирского (казахи, киргизы, теленгеты, шорцы, качинцы, телеуты, кумандинцы и тубалары) расовых компонентов признается большинством исследователей (независимо от взглядов на проблему происхождения указанных рас), выявлены значимые статистические различия (p<0.04 для ИГК I и p<0.03 для ИГК III). Кроме того,
302

первая группа выборок отчетливо отделяется от второй и по результатам анализа главных компонент (ключевые признаки дерматоглифики, p<0.03 для ГК I). Эти данные говорят о самостоятельном таксономическом статусе уральских популяций, происхождение которых не может быть сведено к результатам смешения двух больших рас Евразии.
Результаты интегративного анализа свидетельствуют о существовании общих направлений в изменчивости дерматоглифических и краниометрических комплексов признаков. В первую очередь эта изменчивость обусловлена взаимодействием двух больших рас и различным соотношением монголоидного и европеоидного компонентов в выборках. Вторым направлением изменчивости, отраженным сходным образом в рамках систем тех и других признаков, является дифференциация южных и северных вариантов внутри европеоидной и монголоидной рас, причем сближение северных вариантов может быть интерпретировано как свидетельство общей генетической основы населения, большая часть групп которого относится к одной языковой (уральской) семье.
Отметим, что разграничительная мощность системы краниометрических признаков по результатам проведенного анализа выше на уровне больших рас, а дерматоглифики - на субрасовом уровне (что подтверждается отчетливой дифференциацией южных и северных европеоидов, Рис.12.4).
12.3. Дерматоглифика и одонтология
Один из примеров изучения соотносительной изменчивости дерматоглифического и одонтологического комплексов связан с антропологией казахов (Сихимбаева, 1987). Их сравнение с широким кругом этнических групп обнаруживает сходную картину взаимоотношений по данным двух систем признаков (Рис.12.5).
К.Б. Сихимбаевой была также впервые выполнена интеграция данных дерматоглифики и одонтологии 18 народов Евразии с целью определения таксономического положения казахов. При этом одонтологические расстояния и одонтологический монголоидный комплекс рассчитывались по тому же принципу, что и соответствующие дерматоглифические показатели, с применением евразиатской шкалы вариаций признаков. Казахи вместе с киргизами вошли в кластер, объединяющий народы монголоидного
303

типа, оказавшись, впрочем, в этом скоплении наименее «монголизированными» (Рис.12.6).
Сопоставление данных дерматоглифики и одонтологии 13 башкирских групп выявило отсутствие статистических связей между признаками обеих систем и ненаправленную дифференциацию башкир по выбранным признакам. При этом по сравнению с одонтологической системой признаки дерматоглифики обнаружили большую разграничивающую способность при анализе как мужских, так и женских выборок. Дерматоглифика более отчетливо дифференцирует мужские группы в направлении запад - восток (Суворова, 2005, с. 18-19).
д ер м ато гл и ф и к а одон тология
Рисунок 12.5. Обобщенные расстояния, выраженные в масштабе, между казахами (в центре) и сравнительными группами по комплексу признаков кожного (ОДР, мужчины) и зубного (СТР, мужчины и женщины суммарно) рельефа (по: Сихимбаева, 1987).Figure 12.5. Generalized distances between Kazakhs (in the centre) and other groups based on dermatoglyphic (GDD, males) and odontological (total taxonomic distances, both sexes) data analysis (from: Сихимбаева, 1987)
При исследовании белорусских выборок выявлено отсутствие внутригрупповой связи между признаками дерматоглифики и одонтологии, а также несовпадение внутри- и межгрупповых коэффициентов корреляции по обеим системам. Результаты свидетельствуют, что взаимоотношения между группами сложились исторически на данной территории (Тегако, 1970).
русские
таджики
304

Рисунок 12.6. Кластеризация групп Северной Евразии по общему таксономическому расстоянию признаков дерматоглифики иодонтологии (без учета расовых градиентов признаков). СЕМК - суммарный европеоидно-монголоидный комплекс. OTP - обобщенное таксономическое расстояние (по: Сихимбаева, 1987).Figure 12.6. Clusterization of groups from northern Eurasia based on integrated taxonomic distances of dermatoglyphic and odontological traits. СЕМК - integrated Mongoloid complex. ОТР - generalized taxonomic distance (from: Сихимбаева, 1987)
12.4. Дерматоглифика и популяционная генетика
Схема дивергенции больших рас, построенная по данным дерматоглифики (см. Главу 9), во многом оказывается сходной со схемой эволюционного древа человечества по данныммикросателлитной изменчивости ДНК-маркеров. При построении последней были использованы данные 52 популяций, охватывающие 1056 человек (Zhivotovsky et al., 2003; Животовский, 2006). В обоих случаях выделяется наиболее «молодой» восточный ствол (монголоиды, американоиды, австралоиды), наибольшей
305

специфичностью отличаются европеоиды и негроиды Африки (Рис.12.7).8 При этом по данным дерматоглифики наиболее сходны монголоиды и австралоиды, по данным микросателлитной изменчивости - монголоиды и американоиды. Максимальной специфичностью по данным генетики отмечены негроиды, по данным дерматоглифики - негроиды и (в меньшей степени) европеоиды.
Рисунок 12.7. Эволюционное древо человечества по данным о микросателлитной изменчивости (по: Животовский, 2006).Figure 12.7. Evolution tree of mankind based on the microsatellite variability data (from: Zhivotovsky, 2006).
8 Аналогичные или близкие к представленной схемы дифференциации человечества можно встретить в работах и других исследователей (см. например: Jakobsson et al., 2008; Tishkoff, Kidd, 2004; и др.). Вместе с тем, как показывают данные расшифровки генома одного австралийского аборигена, австралийцы должны быть исключены из состава восточного ствола, поскольку в генеалогическом отношении противостоят общим предкам европеоидов и монголоидов (подробнее см. Козинцев, 2013).
306

%

Заключение
Гребешковые линии и образованные ими узоры кожного рельефа пальцев и ладоней рук у каждого человека составляют неповторимый рисунок. Этот рисунок под влиянием наследственных, а частично и пренатальных факторов, формируется в эмбриональный период и остается неизменным в течение всей жизни. Однако неповторимые в деталях узоры кисти поддаются классификациям, разделению на дискретные типы и количественные показатели, которые можно вычислить и обобщить для любой группы людей, отобранной по каким-либо параметрам. Ни одна человеческая раса, популяция или этническая общность не выделяется оригинальными, присущими только ей, типами или какими-либо характерными деталями узоров кожного рельефа. Вместе с тем распределение дерматоглифических признаков и их комплексов, рассчитанных для популяций разных частей ойкумены, позволяют устанавливать относительную степень сходства и различий между ними, а также выявлять географические закономерности и определять расовые компоненты в их составе.
К началу XXI века специалистам в области дерматоглифики из разных стран так и не удалось выработать общую программу исследований. Наиболее широкое распространение сегодня имеет программа, которая включает в себя вычисление частот основных типов пальцевых узоров и гребневой счет. Признаки ладони в большинстве современных работ либо выпадают из поля зрения специалистов, либо сводятся к расчетам гребневого счета. Кроме того, за последние два десятилетия в дерматоглифике произошло резкое сокращение числа работ, посвященных популяционной и этногенетической тематике.
Предлагаемая вниманию читателя книга посвящена описанию системы методических приемов, хорошо зарекомендовавших себя в работах российских специалистов по этнической и расовой дерматоглифике, проблемам дифференциации популяций разного таксономического уровня и возможностям применения дерматоглифики в изучении человеческих популяций в разных аспектах. Термин «раса» авторы используют для обозначения групп популяций, обладающих достаточным фенотипическим и генетическим сходством, позволяющим отличить их от других
308

подобных групп. Расы открыты для обмена генами с другими расами; расовые признаки не являются абсолютными, имеют высокую внутригрупповую вариабельность, способны меняться с течением времени.
Основные результаты исследования сведены в разделы, реферативное изложение которых дублируется далее в англоязычном варианте.
Ключевые признаки дерматоглифики
Все коллекции отпечатков, использованные при подготовке книги, исследовались в соответствии с правилами методики Камминса и Мидло (Cummins, Midlo, 1961). Положение осевых ладонных трирадиусов определялось по схеме А. Шармы, согласно которой низкий трирадиус t располагается в зоне нижней 1/6 части продольной ладонной оси. Выше этой границы и до верхней границы 1/3 части оси располагается зона трирадиуса t', выше этого уровня - зона трирадиуса t'' (Sharma, 1964). Поле 4 определялось не в «классическом» точечном варианте, а было расширено на 3-4 мм выше и ниже средней точки ульнарного края ладони (Хить,1983).
В 1969 г. Г.Л. Хить впервые была выказана идея о разделении дерматоглифических признаков на ключевые и неключевые. Впоследствии эта идея была подтверждена на большом материале, охватывающем основные расовые группы (европеоидов, монголоидов, негроидов, австралоидов и американоидов) и метисные популяции. В ходе исследований были вычислены попарные корреляции между большей частью дерматоглифических признаков, за исключением гребневого счета и угловых величин. Подавляющее большинство признаков обладаетрасодиагностическими свойствами, но только группа из шести ключевых признаков статистически не связана между собой на внутригрупповом индивидуальном уровне. В их число входят: индекс Камминса (Ic), дельтовый индекс (Dl10), частота проксимального осевого трирадиуса (t), узорность гипотенара (Hy), суммарная частота добавочных межпальцевых трирадиусов (ДМТ), узорность тенара и I межпальцевой подушечки (Th/I). Последний признак имеет ограниченную таксономическую ценность, так как методически трудноопределим, не разграничивает европеоидов и монголоидов, в пределах больших и локальных рас варьирует значительно сильнее, чем остальные ключевые признаки, обладает
309

чрезвычайно высокой степенью асимметрии. Недавно И.Г. Широбоковым (2009б) было показано также высокое таксономическое значение радиальной ориентации пальцевых узоров (FRP), которая слабой и неустойчивой корреляцией связана с величиной дельтового индекса. Метод определения ориентации пальцевых узоров, предложенный им, базируется на особенностях строения скелета узора (положении его более открытой стороны) и не связан напрямую ни с типом узора, ни с величиной гребневого счета.
Ключевые признаки и FRP связаны с группами неключевых элементов и отражают основную часть морфологического комплекса папиллярных узоров ладони и концевых фаланг пальцев. Ключевые признаки, в отличие от большинства неключевых, в пределах расы распределяются практически по типу нормальной кривой. Хотя кривые распределения ключевых признаков всех больших рас обнаруживают взаимные трансгрессии в разной степени, различия средних значений признаков между расами, как правило, статистически значимы.
Важным свойством ключевых признаков является соподчиненная изменчивость. Внутрирасовая популяционная вариабельность признаков подчиняется строгому правилу. Вариабельность возрастает в порядке: Ic - Dl10 - t - Hy - ДМТ - Th/I. Исключение составляют негроиды, у которых изменчивость ДМТ меньше, чем гипотенара. Эта последовательность связана обратным соотношением с диагностической ценностью признака: чем меньше он варьирует в пределах расы, тем более стойкой характеристикой расы он является и тем меньше вероятность возникновения случайных величин признака.
Половые различия признаков дерматоглифики
Межполовые различия признаков дерматоглифики носят выраженный и направленный характер, поэтому анализ суммированных данных по мужчинам и женщинам приводит к искажениям результатов исследования. По результатам анализа 716 параллельно исследованных мужских и женских выборок из пяти рас (европеоиды, монголоиды, негроиды, американоиды, австралоиды) были обнаружены следующие закономерности.
310

1. На уровне больших рас ключевые признаки обоих полов полностью скоррелированы, а в пределах расы (на популяционном уровне) эта связь ослабевает и сохраняется не во всех случаях. Величины признаков в большей степени согласованы между полами у австралоидов и американоидов, в наименьшей - у европеоидов. Ключевые признаки скоррелированы у обоих полов более тесно, чем неключевые.
2. Ключевые признаки в мужских выборках всех больших рас имеют большую величину, кроме узорности гипотенара. Последний признак чаще встречается в женских выборках у европеоидов, монголоидов и американоидов, в мужских выборках - у негроидов и австралоидов.
3. У всех рас коэффициент полового диморфизма (выражающий величину признака у мужчин в долях по отношению к таковой у женщин) возрастает в порядке: Ic - Dl10 - t - ДМТ - Th/I. (Узорность Hy не рассматривалась, так как для этого признака отсутствует общий для всех рас вектор межполовых различий). Таким же образом увеличиваются показатели изменчивости КПД данных признаков.
4. Максимальные различия между полами демонстрируют негроиды, минимальную - австралоиды. Монголоиды и американоиды сближаются в этом отношении с австралоидами. Европеоиды занимают срединное положение между австралоидами и негроидами.
Характеристика и таксономия больших рас и их подразделений
Поскольку понятие расы изначально базировалось на соматологических данных, дерматоглифический материал был организован в рамках соматологической классификации. Число наблюдений в пределах расы по разным признакам сильно варьирует, достигая максимума в отношении Dl10. В общей сложности рассмотрены 1322 выборки ойкумены (756 мужских и 566 женских), среди которых преобладают европеоиды и монголоиды. Анализ проводился двумя методами - межгрупповой шкалы и главных компонент. Монголоиды и австралоиды отмечены определенным сходством и имеют высокие и очень высокие значения Dl10 и t, а также пониженную частоту Hy и ДМТ. По типу комбинаций признаков к ним приближаются американоиды. Таким
311

образом, типологически эти три расы сходны. Две остальные расы - негроиды и европеоиды - являются почти полными антиподами: признаки, имеющие среднюю или небольшую величину у европеоидов, сильно выражены у негроидов (ДМТ, Th/I), и наоборот (Ic, t, Hy, FRP). Только дельтовый индекс практически не разграничивает обе эти расы. По результатам серии статистических анализов наиболее обособленное положение занимают негроиды и европеоиды, австралоиды и монголоиды характеризуются определенным сходством, а американоиды тяготеют к двум последним расам (в большей степени - к монголоидам). Таким образом, по комплексу признаков человечество разделяется на три расовых ствола - западный (европеоиды), южный (негроиды) и восточный (монголоиды, австралоиды и американоиды).
Бимануальное распределение и асимметрия признаков дерматоглифики
Бимануальная асимметрия ключевых признаков дерматоглифики рассматривалась на материалах 20 мужских и 20 женских параллельно изученных выборок, относящихся к трем большим расам: европеоидной, монголоидной и негроидной (всего более 3 600 человек).
Как в мужских, так и в женских выборках величина дельтового индекса, индекса Камминса и частоты радиальных узоров выше на правой руке, а узорность гипотенара, тенара и частота проксимального трирадиуса ладони, как правило, на левой.
В целом у мужского населения Старого Света на межэтническом уровне сопоставления выявлены значимые билатеральные различия по всем основным признакам, за исключением узорности гипотенара и частоты ДМТ. У женщин зафиксированы те же закономерности, однако бимануальные различия по узорности гипотенара также являются значимыми.
Показатели асимметрии ключевых признаков в пределах трех изученных рас возрастают в ряду: от обоих индексов до узорности тенара. Частота радиальных узоров обнаруживает максимально симметричное распределение на пальцах правой и левой рук.
Как в мужских, так и в женских выборках, признаки правой руки имеют более высокую таксономическую ценность для межпопуляционной дифференциации на уровне больших рас, чем признаки левой. Так, европеоидные и монголоидные выборки
312

различаются между собой по величинам всех ключевых признаков правой руки, кроме тенара, тогда как в число признаков левой руки, расовые различия по которым незначимы, входят, помимо тенара, еще три признака: проксимальный трирадиус t, узорностьгипотенара и частота FRP. При этом, как показывает корреляционный анализ данных главных компонент, между картинами дифференциации мужских и женских выборок по признакам правой руки обнаружено значительно большее сходство, чем по признакам левой.
Между мужчинами и женщинами выявлены значимые различия по степени выраженности бимануальной асимметрии по большинству признаков. Для мужчин характерны более резко выраженные различия, тогда как в женских выборках бимануальные различия носят более сглаженный характер. Единственным исключением является узорность гипотенара, бимануальные различия по которой в большей степени выражены у женщин.
Дерматоглифика и исторические процессы
Исторические и географические факторы являются определяющими для сложения комплексов антропологических признаков, определяющих степень сходства и различий человеческих популяций. Исследование описанной выше программы признаков дерматоглифики позволяет реконструировать некоторые этапы истории формирования популяций разного таксономического уровня, в т.ч. больших рас, несмотря на то обстоятельство, что аналитическая база построена на материалах, собранных в XX веке.
Неоднократно становилось целью специальных исследований изучение дерматоглифической дифференциации в пределах языковых групп. Поиски истоков генетической общности в связи с языковым сходством были произведены на обширных материалах по тюркской, финно-угорской, северомонгольской и славянской группам языков. В результате выяснилась дерматоглифическая разнородность финно-угров и тюрков, в составе которых обнаружены различные варианты, не сходные как по комбинации признаков, так и по условной доле монголоидности. Варианты связаны с различным содержанием европеоидного и монголоидного компонентов. Один из таких вариантов является общим для финно- угров и тюрков, свидетельствуя о преобладании тенденции
313

самодостаточности в процессах расо- и лингвогенеза, преодолевающих барьеры языка и расы, соответственно. В обеих языковых группах не найдено сходства результатов дерматоглифического анализа и языковой дифференциацией, за одним исключением. Финноязычные народы циркумбалтийского региона (финны, карелы, вепсы и эстонцы), соматологически представляющие северных европеоидов, и по данным дерматоглифики представляют единую совокупность. Таким образом, в этом регионе произошло совпадение по трем параметрам: языку, расовому типу и дерматоглифике. Следует также отметить, что в значительной степени однородными - в дерматоглифическом и расовом отношении - представляются народы славянской группы, при этом особенно отличаются гомогенностью восточные славяне: русские, украинцы, белорусы. Все же в большинстве случаев расогенез не связан с лингвогенезом, и картины дерматоглифической и языковой дифференциации не совпадают между собой.
Раздел содержит библиографический список отечественных и зарубежных работ по этнической и популяционной дерматоглифике, опубликованных за последние 25 лет.
Соотношение признаков дерматоглифики и других систем
Варианты дифференциации популяций, основанные на материалах разных антропологических систем (соматологии, одонтологии, дерматоглифики, краниологии, серологии и др.), нередко существенно различаются между собой, в том числе на высших таксономических уровнях. Традиционно в качестве причины расхождений указывается, что каждая система признаков контролируется разными группами генов и обладает спецификой, обязывающей учитывать природу признаков, а также время и ареал их формирования. Очевидно, одна из основных причин расхождений заключается также и в том, что многие морфологические признаки в рамках любой антропологической системы (в том числе дерматоглифики) являются искусственно выделенными объектами.
При анализе расового состава обобщенных выборок населения крупных историко-географических регионов России и сопредельных территорий Северной Евразии было обнаружено совпадение дифференциации по данным дерматоглифики и соматологии. Некоторые расхождения возникли на уровне локальных рас
314

Северной Евразии, где дерматоглифический материал был организован в рамках расово-соматологической классификации и проанализирован методом числовой таксономии. В ходе исследований было выявлено два основных компонента - европеоидный и монголоидный, наряду с компонентами метисного происхождения. Расхождения этой схемы с данными расовой соматологии (числовые схемы евразиатского масштаба для которой отсутствуют) носят как частный, так и принципиальный характер. Выявлены заметные различия между данными дерматоглифики и соматологии при исследовании этнических групп Кавказа. Были сопоставлены обе схемы, основанные, однако, на ограниченном числе этнических выборок. По данным дерматоглифики отчетливо выделились кластеры абхазо-адыгейских народов, с одной стороны, и тюркских (за исключением ногайцев) народов, с другой. Первые демонстрируют взаимную близость (за исключением абхазов) и в рамках соматологической схемы, но этим сходство картин дифференциации в разных системах признаков и ограничивается.
Обнаружено также важное свойство признаков кожного рельефа. Они демонстрируют устойчивый сдвиг в сторону увеличения европеоидно-монголоидного комплекса в пределах этнических выборок Европы и Западной Сибири по сравнению с соматологическими данными, что свидетельствует о более выраженной реакции дерматоглифической системы на монголоидный компонент в популяции. Выраженный «восточный» сдвиг дерматоглифических показателей зафиксирован также относительно краниометрической, краниоскопической и одонтологической систем признаков при интеграции данных 38 групп Северной Евразии (Моисеев, 2004, с.192).
Нельзя без сожаления отметить, что к началу XXI века интересы европейских и американских специалистов постепенно сместились в область анализа строения кожного рельефа человека в генетическом, медицинском, конституциональном и судебномедицинском аспектах, тогда как расоведение утратило прежние позиции и в настоящее время замыкает тематический список работ. Отдельные антропологи стран бывшего СССР, казалось бы, сохраняют интерес к изучению рас и их локальных вариантов по дерматоглифическим данным, однако общая численность специалистов и публикаций явно недостаточна для полноценного развития этой антропологической дисциплины.
315

Одной из главных целей авторов, которыми они руководствовались при написании этой книги, было привлечь внимание молодых специалистов-антропологов к возможностям применения дерматоглифики для анализа особенностей и этапов формирования популяций Земного шара, выявления основных компонентов в их антропологическом составе, их характеристик и истоков.
Популяционная и этническая дерматоглифика в системе антропологического знания сегодня незаслуженно (пусть и негласно) отодвинута на второй план. В обобщающих работах по антропологии отдельных народов результаты дерматоглифических исследований нередко представлены краткими упоминаниями и ссылками на литературу. Дискуссии и рецензии на публикации по дерматоглифической тематике давно исчезли со страниц российских периодических научных изданий. (Тот факт, что эта же ситуация относится к публикациям по краниологической и соматологической тематике, не уменьшает сожаления авторов). Вместе с тем, благодаря усилиям отечественных и зарубежных специалистов, сегодня мы располагаем одной из самых больших по объему и территориальному охвату базой данных по дерматоглифике современного населения значительной части ойкумены. Тем не менее, существует острая необходимость увеличения материалов, касающихся популяций тропического и субтропического поясов Земли, а также аборигенного населения Америки.
316

%

Библиография
1. Аксянова Г.А. Итоги расово-соматологического изучения финноязычных народов Северо-Запада России // Расы и народы. Вып. 30. М.: Наука, 2004. С. 292-325.
2. Алексеев В.П. Происхождение народов Восточной Европы (краниологическое исследование). М.: Наука, 1969. 324 с.
3. Алексеев В.П. Очерк происхождения тюркских народов Восточной Европы в свете данных краниологии // Вопросы этногенеза тюркоязычных народов Среднего Поволжья. Казань: АН СССР, 1971. С. 232-271.
4. Алексеев В.П. География человеческих рас. М.: Мысль, 1974а. 120 с.
5. Алексеев В.П. Краниологическая характеристика населения Восточной Фенноскандии (по материалам Г.Ф. Дебеца и автора) // Расогенетические процессы в этнической истории. М.: Наука, 1974б. С.85-105.
6. Алексеев В.П. Происхождение народов Кавказа (краниологическое исследование). М.:Наука, 1974в. 316 с.
7. Алексеев В.П. К краниологии калмыков в связи с их происхождением // Вопросы сравнительной этнографии и антропологии калмыков. Элиста, 1980. C.3-41.
8. Алексеев В.П. Географические очаги формированиячеловеческих рас. М.: Мысль, 1985. 237 с.
9. Антонюк С.А. К методике получения отпечатка ладони человека // Вопросы антропологии. 1975. Вып. 50. С.219-221.
10. Антропологический словарь. М.: Классикс Стиль, 2003. 328 с.11. Арутюнян А.Р. Информационные технологии анализа и синтеза
деформаций дактилоскопических изображенийю Автореф. дис. ... канд. техн. наук. М., 2010. 18 с.
12. Ашилова ДО. Дерматоглифическая характеристика калмыков // Проблемы современных этнических процессов в Калмыкии. Элиста. 1985. С. 121-135.
13. Багашев А.Н. Краниологический тип нижнеиртышских хантов // Обские угры (ханты и манси). Материалы к серии "Народы и культуры". М., 1991. С.29-37.
14. Багашев А.Н. Этническая антропология тоболо-иртышских татар. Новосибирск: Наука, 1993. 152 с.
318

15. Багашев А.Н. Краниологический тип пелымских манси // Вестник археологии, антропологии и этнографии. 1999. №2. С.69-77.
16. Багашев А.Н. Палеоантропология Западной Сибири. Лесостепь в эпоху раннего железа. Новосибирск: Наука, 2000. 374 с.
17. Багдасарян А.Г. Дерматоглифические корреляты ускоренного и замедленного роста и развития ребенка // Новые исследования по возрастной физиологии. М., 1974. №3. С.62- 63.
18. Балановская Е.В., Балановский О.П. Русский генофонд на Русской равнине. М.: Луч, 2007. 416 с.
19. Бодемер Ч. Современная эмбриология. М.: Мир, 1971. 446 с.20. Божченко А.П. Возможности определения идентификационно
значимых признаков человека посредством анализа дерматоглифических структур пальцев рук // Проблемы экспертизы в медицине. №4 (16). Ижевск, 2004. С.42-47
21. Божченко А.П., Ракитин В.А, Самарин А.И., Щербаков В.В. Методы дерматоглифики в идентификации личности погибших. Ростов-на-Дону: ООО Ростиздат, 2002. 160 с.
22. Бунак В.В. Раса как историческое понятие // Наука о расах и расизм. Труды Института антропологии МГУ. М., 1938. Вып.4. С.5-46.
23. Бунак В.В. Антропометрия. М.: Медгиз, 1941. 368 с.24. Бурлай Е.В. Биотехническая система дерматоглифических
исследований ладони: Автореф. дис. ... канд. техн. наук. М.,2009. 16 с.
25. Бурлай Е.В., Новикова И.В., Спиридонов И.Н. Автоматизированные дерматоглифические исследования ладони для выявления идиопатической эпилепсии // Научнотехническая конференция «Медико-технические технологии на страже здоровья». Сб. докл. 19-26 сентября 2009 г., Черногория. М.: НИИ РЛ МГТУ им. Н.Э.Баумана, 2009. С. 117-119.
26. Василенко Ю.А. Параллелизм изменений дерматоглифики, эндокринного и психологического статуса в популяции детского населения, проживающего в районах с высокой антропогенной нагрузкой: Автореф. дис. .. к.б.н. Ставрополь, 2005. 20 с.
27. Витов М.В. Антропологические данные как источник по истории колонизации Русского Севера. М.: ИЭА РАН, 1997. 202 с.
28. Войтенко В.П., Полюхов А.М., Колодченко В.П. О наследовании пальцевых узоров человека // Генетика. 1979. № 7. С. 1317-1326.
319

29. Волоцкой М. В. К вопросу о генетике папиллярных узоров пальцев (исследование 234 пар близнецов) // Труды Медикогенетического института. Т.4. М., 1936. С.404-439.
30. Волоцкой М.В. Генетика кожного рельефа: анализдельтообразования как количественного признака // Антропологический журнал. 1937а. №3. С.42-73.
31. Волоцкой М.В. Схема дактилоскопической типологии // Ученые записки МГУ. 1937б. Вып. 10: Антропология. Вып. 3. С.138-154.
32. Волоцкой М.В. Новый дактилоскопический индекс и его распределение по земному шару // Ученые Записки МГУ. 1937в. Вып. 10: Антропология. Вып. 3. С. 156-171.
33. Галактионов O.K. Дерматоглифика в популяционныхисследованиях: (анализ количественных и качественныхпризнаков в архаичных монголоидных этносах). Владивосток: Изд-во Дальневост. ун-та, 2008. 114 с.
34. Гей И.А. Орнаменты на керамике и отпечатки кончиков пальцев древних гончаров (историографический очерк) // Изучение историко-культурного наследия народов Южной Сибири. Горно- Алтайск, 2007. Вып.5. С.7-20.
35. Гейндль Р. Дактилоскопия и другие методы уголовной техники вделе расследования преступлений. М.: Государственноетехническое издательство, 1927. 320 с.
36. Герасимова М.М. Краниология калаусских ногайцев // Материалы по изучению историко-культурного наследия Северного Кавказа. Вып. IV: Антропология ногайцев. М.: Памятники исторической мысли, 2003. С. 36-68.
37. Гладкова Т.Д. Сравнительная характеристика кожного рельефа ладоней обезьян и человека: Автореф. дис. ... к.б.н. М., 1959. 19 с.
38. Гладкова Т.Д. Явления симметрии и асимметрии у человека в признаках дерматоглифики // Вопросы антропологии. 1962. Вып.10. С.44-54.
39. Гладкова Т.Д. Кожные узоры кисти и стопы обезьян и человека. М.: Наука, 1966. 151 с.
40. Гладкова Т.Д., Намсарайнайдан Л., Ящук Е.В. Дерматоглифическая характеристика монголов-халха в сравнении с некоторыми другими народами Азии. Вопросы антропологии. Вып.79. 1987. С.131-139.
41. Гохман И.И., Богданова В.И., Чистов Ю.К. Некоторые итоги антропологического изучения современного населения Южного
320

Йемена // Антропология сегодня. Вып.1. СПб.: МАЭ РАН, 1995. С.6-41.
42. Губарева Л.И. Экологический стресс. СПб.: Ланъ, 2001. 448 с.43. Гусева И.С. Генетические проблемы в дерматоглифике:
Автореф. дис. ... д.б.н. Минск, 1982. 38 с.44. Гусева И.С. Морфогенез и генетика гребешковой кожи человека.
Минск: Беларусь, 1986. 158 с.45. Гусева И.С. О наследовании пальцевых узоров человека //
Вестник антропологии. 2008. Вып.16. С.109-117.46. Гусева И.С. Эпигенетический подход к анализу особенностей
формирования и популяционного распределения пальцевых узоров рук человека // Вестник Московского университета. Серия XXIII. Антропология. 2009. №3. С.47-54.
47. Гусева И.С. Пальцевые узоры человека. Морфология. Морфогенез. Генетика. Дерматоглифика как маркер в медицинской и спортивной антропологии. Минск: ФУАинформ,2010. 336 с.
48. Гусева И.С. Симметрия-асимметрия как форма морфологической организации животных и человека // Актуальные вопросы антропологии. 2011. Вып. 6. С.88-111.
49. Гусева И.С., Антонюк С.А. Симметрия в строении кожного рельефа конечных фаланг пальцев руки человека и ее генетическая основа // Вопросы антропологии. 1969. Вып. 31. С.33-41.
50. Гусева И.С., Сорокина Т.Т. Дерматоглифика как конституциональный маркер при мультифакториальной патологии // Вопросы антропологии. 1998. Вып.89. С.99-111.
51. Дашцэвэгийн Т. Дерматоглифические особенности некоторых этнических групп Западной Монголии. // Археологические, этнографические и антропологические исследования в Монголии. Новосибирск, 1990. С. 184-192.
52. Дебец Г.Ф. Палеоантропология СССР. М., 1948. (ТИЭ. Т.17) М Л., 1948. 392 с.
53. Дебец Г.Ф. Антропологические исследования в Камчатской области. Труды Северо-восточной экспедиции (ТИЭ. Т.17). М., 1951. 264 с.
54. Денисова Р.Я. Этногенез латышей (по данным краниологии). Рига: Зинатне, 1977. 360 с.
55. Дерябин В.Е. Многомерная биометрия для антропологов. М.: МГУ, 1983. 227 с.
321

56. Дерябин В.Е. О методиках многомерного таксономического анализа в антропологии. Канонический анализ против главных компонент // Вестник антропологии. 1998. Вып.4. С.54-61.
57. Дерябин В.Е. Краткий справочник по решению типовых задач биометрической обработки антропологических данных. М.: МГУ, 2005. 223 с.
58. Джанибекова И.В. Связь тестостерона с билатеральной асимметрией у мужчин и женщин: Автореф. дис. . к.б.н. М., 2002. 22 с.
59. Дильман В.М. Большие биологические часы. Введение в интегративную медицину. М.: Знание, 1986. 256 с.
60. Долинова Н.А. Дерматоглифика удмуртов // Новые исследования по этногенезу удмуртов. Ижевск: УрО АН СССР, 1989. С.108-122.
61. Долинова Н.А. Дерматоглифика восточных славян // Восточные славяне. Антропология и этническая история. М.: Научный мир, 1999. С. 60-79.
62. Долинова Н.А. Дерматоглифика коми-зырян и коми-пермяков // Антропология коми. М.: ИЭА РАН, 2005. С. 193-211.
63. Дремов В.А. Расовая дифференциация угорских и самодийских групп Западной Сибири по данным краниологии // Проблемы антропологии древнего и современного населения севера Евразии. Л., 1984. С.106-132.
64. Дремов В.А. Краниология среднеобских хантов // Материалы к серии "Народы и культуры". Обские угры (ханты и манси). М., 1991. С.10-28.
65. Животовский Л.А Микросателлитная изменчивость в популяциях человека и методы ее изучения // Информационный Вестник ВОГиС. 2006. С.74-96.
66. Загорученко Е.А. Эмбриональное развитие и топографическая гистология кожи человека. Автореф. дис. ... д.м.н. М., 1973. 33 с.
67. Задорожный В. Идентификация по отпечаткам пальцев. Часть 1 // PC Magazine. Russian Edition. 2004. №1. С.114-119.
68. Звягин В.Н., Ракитин В.А. Дерматоглифика: энциклопедический словарь. Пермь: Пресстайм, 2012. 386 с.
69. Зубов А.А. О расово-диагностическом значении некоторых одонтологических признаков // Советская этнография. 1968. №3. С.49-59.
70. Зубович В.К. Гормональные влияния на организм новорожденного. Минск: Беларусь, 1989. 160 с.
322

71. Ивашков В.А. Особенности составления заключения эксперта при выполнении дактилоскопических экспертиз. М., 1999. 240 с.
72. Исмагулов О. Население Казахстана от эпохи бронзы до современности (палеоантропологическое исследование). Алма- Ата: Наука Казахской ССР, 1970. 239 с.
73. Исмагулов О., Сихымбаева К.Б., Исмагулова А.О. Этническая дерматоглифика казахов. Алматы, 2007. 240 с.
74. Калантаевская К.А. Эмбриология рисунка кожи у человека, его возрастные и патологические изменения // Научные известия Казахской ССР. 1953. №11. С.80-91.
75. Калантаевская К.А., Морфология и физиология кожи человека. Киев: Здоров’я, 1972, 267 с.
76. Канаев И.И. Близнецы. М.-Л.: Изд-во АН СССР, 1959. 382 с.77. Клевцова НИ., Кучинскас В.К. Дерматоглифические
особенности литовцев // Вопросы антропологии. Вып.81. 1988. С.74-88.
78. Кобозева Н.В., Рассохин А.В. Роль эндокринной системы плода в невынашиваемости беременности // Невынашивание и недонашивание беременности. М., 1984. С. 101 -106.
79. Козинцев А.Г. Расовая классификация в свете новыхгенетических данных // Радловский сборник: Научныеисследования и музейные проекты МАЭ РАН в 2012 г. СПб.: МАЭ РАН, 2013. С.64-71.
80. Козинцев А.Г., Моисеев В.Г. Об антропологическом своеобразии уралоязычных народов: сопоставление данных краниоскопии и краниометрии // Этнографическое обозрение 1995. №4. С.81-88.
81. Комиссарова Е.Н., Сазонова Л.А., Карелина Н.Р. Соматотипология и пальцевая дерматоглифика у девочек, проживающих в Северо-Западном регионе России // Тезисы докладов VI конгресса этнографов и антропологов России. СПб., 2005. С.372.
82. Кочар Н.Р. Антропология армян. Дерматоглифика и популяционная структура. Ереван: АН Армянской ССР, 1989. 103 с.
83. Крикун Е.Н. Изменчивость морфофункциональных показателей организма человека под влиянием неблагоприятных экологобиологических факторов: Автореф. дис. ... д.м.н. М., 2006. 39 с.
84. Круц СИ. Антропологическая характеристика ногайцев XVI- XVIII вв. // Материалы по изучению историко-культурного
323

наследия Северного Кавказа. Вып.ГУ: Антропология ногайцев. М.: Памятники исторической мысли, 2003. С.206-233.
85. Лебедева Е.Г., Славолюбова И.А. Дерматоглифические исследования чувашских девушек // Антропологический мониторинг населения Чувашии. Чебоксары: ЧГИГН, 2009.С.45-77.
86. Лебедева Е.Г. Изучение пальцевых узоров у чувашей // Антропологический мониторинг населения Чувашии. Чебоксары: ЧГИГН, 2009. С.77-129.
87. Маккьюсик В. Генетика человека. М.: Мир, 1967. 200 с.88. Мглинец В.А. Врожденные формы брахидактилии и
дерматоглифика // Онтогенез. 1991а. Т. 22. № 3. С.291 -299.89. Мглинец В.А Связь изменчивости дерматоглифики с длиной
пальцев при генетических нарушениях: синдром Дауна // Генетика. 1991б. Т. 27. № 3. С.541-547.
90. Миклашевская H.H. Краниология киргизов // Труды Киргизской археолого-этнографической экспедиции. 1959. T II. С.266-294.
91. Моисеев В.Г. Опыт интеграции данных четырех систем антропологических признаков (краниометрии, краниоскопии, одонтологии и дерматоглифики) // Палеоантропология. Этническая антропология. Этногенез. Сборник к 75 -летию И.И. Гохмана. СПб.: МАЭ РАН, 2004. С. 186-200.
92. Моисеева Т.О. Комплексное криминалистическое исследование потожировых следов человека. М.: Городец-издат, 2000. 224 с.
93. Негашева М.А. Дерматоглифические аспекты морфологической конституции человека // Морфология. 2007. № 5. С.88 -93.
94. Негашева М.А. Взаимосвязи соматических, дерматоглифических и психологических признаков в структуре общей конституции человека с позиций системного подхода // Морфология. 2008. Т. 133. № 1. С.73-77.
95. Низимбетова А.Н. Эмбриогенез кожи на различных участках тела человека: Автореф. ... к.м.н. Алма-Ата, 1959. 19 с.
96. Никитюк Б.А., Коган Б.И., Гальперина З.З. Влияние половых хромосом на некоторые признаки дерматоглифики // Вопросы антропологии. 1986. №76. С. 136-140.
97. Отелин А.А. Иннервация скелета человека. Атлас. М.: Медицина, 1965. 251 с.
98. Отелин А.А, Машанский В.Ф., Миркин А.С. Тельце Фатер- Пачини: Структурно-функциональные особенности. М.: Наука, 1976. 175 с.
324

99. Павловский О.М. Межгрупповая оценка полового диморфизма в оссеографических показателях // Вопросы антропологии. 1981. Вып.68. С.3-23.
100. Панкратова Л.В. Дактилоскопическое исследование отпечатков пальцев рук на позднекулайской керамике Саровского городища // Культуры и народы Северной Азии и сопредельных территорий в контексте междисциплинарного изучения: Сборник Музея археологии и этнографии Сибири им.В.М. Флоринского. Томск, 2008. Вып.2. C. 158-163.
101. Проблема расы в российской физической антропологии. М.: ИЭА РАН, 2002. 150 с.
102. Прокудина Н.А. Дерматоглифические исследования некоторых народов европейской части СССР // Новое в этнографических и антропологических исследованиях. М., 1974.
103. Пэттен Е.М. Эмбриология человека. М., Медицина, 1959. 768 с.104. Рицнер М.С. Материалы к полигенной гипотезе наследования
папиллярного узора пальцев // Вопросы антропологии. 1970. Вып.36. С.143-148.
105. Рицнер М.С., Шехтер И.А. К вопросу о генетике пальцевого гребневого счета // Вопросы антропологии. 1975. Вып.49. С.158-163.
106. Рицнер М.С., Шехтер И.А., Базилевская Е.С., Костина Н.В. Пальцевая дерматоглифика при олигофрении. Сообщение 1. Анализ папиллярных узоров // Вопросы антропологии. 1971. Вып.39. С.132-137.
107. Рогинский Я.Я., Левин М.Г. Антропология. М.: Высшая школа, 1978. 528 с.
108. Ройтман А.Б., Липовецкая Н.Г. Генетические исследования отпечатков пальцев близнецов // Генетика. 1969. Т.5. № 8. С. 151-161.
109. Ройтман А.Б., Липовецкая Н.Г. Количественные зависимости в наследовании папиллярных узоров // Вопросы антропологии. 1972. Вып.42. С.130-136.
110. Семеновский П.С. Дактилоскопия как метод регистрации. М.: Изд. Упр. Угророзыска Республики, 1923. 113 с.
111. Семеновский П.С. К вопросу о наследственности тактильных узоров // Труды Второго Всероссийского съезда судебномедицинских экспертов. Ульяновск. 1926. С.103-106.
325

112. Семеновский П.С. Распределение главных типов тактильных узоров на пальцах рук человека // Русский антропологический журнал. 1927. Т.16. Вып.1-2. С.47-63.
113. Сихимбаева К.Б. Антропологический состав населения Западного Казахстана по данным дерматоглифики и одонтологии. Автореф. дис. ... к.и.н. Москва, 1987. 23 с.
114. Славолюбова И.А. Жироотложение как конституциональный признак в антропологии: Автореф. дис. ... к.б.н. М., 2008. 24 с.
115. Спасов С., Докумов С., Генкова П. Значение Х и У хромосом для половых различий дерматоглифического рельефа // Вопросы антропологии. 1978. Вып.59. С.112-116.
116. Суворова Н.А. Проблема формирования антропологического состава башкир по данным дерматоглифики и одонтологии. Автореф. дис. ... к.и.н. М., 2005. 30 с.
117. Суворова Н.А. Дерматоглифика башкир // Антропология башкир. СПб.: Алетейя, 2009. С.217-271.
118. Сыроватко А.С., Елистратов А.Я. Об опыте определения отпечатков пальце на дьяковских керамике и мелкой глиняной пластике // Тверской археологический сборник. Вып. 5. Тверь, 2002. С.503-507.
119. Таболин В.А., Лукина Л.И. Нейроэндокринная система плод - плацента-мать // Функции надпочечников у плодов, новорожденных и грудных детей. М.: Медицина, 1975. С.71 -84.
120. Тегако Л.И. Материалы по корреляции между одонтологическими и дерматоглифическими признаками // Вопросы антропологии. Вып.38. 1971. С.133-136.
121. Тегако Л.И. Дерматоглифика населения Белоруссии.Популяционные аспекты изменчивости. Минск: Наука итехника, 1989. 181 с.
122. Титов В.И. Научные основы и методика дактилоскопической экспертизы. Барнаул: Изд-во Алтайского университета, 2002. 123 с.
123. Торвальд Ю. Век криминалистики. М.: Прогресс, 1991. 335 с.124. Усоев С.С. Связь признаков дерматоглифики с кровоснабжением
пальцев // III съезд Белорусского общества генетиков и селекционеров. Минск, 1976. С.222.
125. Фандеева О.М. Структурная органиация папиллярных узоров пальцев и их особенности в семейных группах: Автореф. дис. . д. м. н. СПб., 2010. 30 с.
326

126. Хазиев Ш.Н. Криминалистическое моделирование неизвестного преступника по его следам. Дисс. канд. юрид. наук. М., 1984.
127. Хайруллин Р.М., Никитюк Д.Б., Крикун Е.Н. Факторы изменчивости кожных узоров пальцев. Ульяновск: УлГУ, 2005. 152 с.
128. Хартанович В.И Новые материалы к краниологии саамов Кольского полуострова // Сборник МАЭ. Л., 1980. Т.36. С.35-47.
129. Хартанович В.И. Краниология карел // Антропология современного и древнего населения Европейской Части СССР. Л.: Наука, 1986. С.63-120.
130. Хартанович В.И. Новые материалы к краниологии коми-зырян // Сборник МАЭ. 1991. Т.44. (Новые коллекции и исследования по антропологии и археологии). С.108-125.
131. Хартанович В.И. Материалы к краниологии финнов // Антропология сегодня. СПб., 1995. Вып.1. С.71-80.
132. Хартанович В.И. Новые краниологические материалы по саамам Кольского полуострова // Палеоантропология, этническая история, этногенез. Сборник к 75-летию И.И. Гохмана. СПб.: МАЭ РАН, 2004. С.108-125.
133. Хить Г.Л. Дерматоглифика населения Финляндии и прилегающих областей СССР // Вопросы антропологии. 1969. Вып 32. С.163-171.
134. Хить Г.Л. Дермаоглифическая дифференциация населения СССР (Доклады советской делегации на IX МКАЭН в Чикаго, 1973). М.: Наука, 1973. С.1-20.
135. Хить Г.Л. Дерматоглифика финно-угорских народов // Расогенетические процессы в этнической истории. М., 1974. C.106-124.
136. Хить Г.Л. Расовая дифференциация населения СССР: соматологический и дерматоглифический аспекты. // Расы и народы. 1975. Вып.5. C.69-94.
137. Хить Г.Л. Расовый состав населения Финляндии по данным дерматоглифики // Финно-угорский сборник. Антропология, этнография, археология. М.: Наука, 1982. С. 148-171.
138. Хить Г.Л. Дерматоглифика народов СССР. М.: Наука, 1983. 280 с.
139. Хить Г.Л. Расовый состав населения СССР (по материалам дерматоглифики) // Расы и народы. Вып.16. 1986. С.31 -42.
327

140. Хить Г.Л. Антропологический состав и генетические связи сибирских татар по данным дерматоглифики // Антропология и историческая этнография Сибири. Омск: Омск. Ун -т, 1990а. С.14-34.
141. Хить Г.Л. Дерматоглифика греков // Новое в этнографии. М., 1990б. Вып.1. С. 114-121.
142. Хить Г.Л. Дерматоглифика чулымцев // Тюрки таежногоПричулымья: Популяция и этнос. Томск: Изд-во ТГУ,1991а. С.153-165.
143. Хить Г.Л. Расо- и этногенетические связи населения Средней Азии по данным дерматоглифики. // Проблемы этногенеза и этнической истории народов Средней Азии и Казахстана. Вып.IV. Антропология. М., 1991б. С.87-100.
144. Хить Г.Л. Саамы в дерматоглифической систематике финно- угров Евразии // Происхождение саамов (по данным антропологии и археологии). М.: Наука, 1991в. С.59-82.
145. Хить Г.Л. Дерматоглифика нганасан // Нганасаны. Антропологическое исследование. М., 1992. С.329-347.
146. Хить Г.Л. Дерматоглифика ногайцев // Антропология ногайцев. Вып.4. М., 2003. С.188-196.
147. Хить Г.Л. Дерматоглифика греков Украины и Абхазии // Греки России и Украины. СПб.: Алетейя, 2004а. С.557-558.
148. Хить Г.Л. Дерматоглифика и расогенез населения Кавказа // Древний Кавказ: ретроспекция культур. М., 2004б. С. 198-200.
149. Хить Г.Л. Дерматоглифика народов Памира // Вестник антропологии. 2005. Вып.12. С.37-44.
150. Хить Г.Л. Мордва в дерматоглифической систематике населения Восточной Европы // Вестник антропологии. Вып. 16, 2008. С.100-108.
151. Хить Г.Л. Дерматоглифика тюркоязычных народов Евразии: двадцать лет спустя // Вестник антропологии. М., 2009. С.254-263.
152. Хить Г.Л. Первичные факторы дерматоглифическойдифференциации аборигенного населения Евразии // Вестник антропологии. 2010. Вып.18. С.143-155.
153. Хить Г.Л., Долинова Н.А. Дерматоглифика туркмен // Туркмены в Среднеазиатском междуречье. Ашхабад, 1989. С.70-117.
328

154. Хить Г.Л., Долинова Н.А. Расовая дифференциация человечества (дерматоглифические данные). М.: Наука, 1990. 206 с.
155. Хить Г.Л., Долинова Н.А. Биологические и исторические корреляции признаков дерматоглифики // Вестник антропологии. 1996. Вып.1. С. 161-186.
156. Хить Г.Л., Долинова Н.А. Видовые и расовоспецифические особенности полового диморфизма у человека по признакам дерматоглифики. // Единство и многообразие человеческого рода. М.: ИЭА РАН, 1997. Ч.1. С.98-213.
157. Хить Г.Л., Долинова Н.А. Дерматоглифика и расогенез финно- угров Евразии // Антропология современных финно-угорских народов. М., 2000. С.27-99.
158. Хить Г.Л., Долинова Н.А. Дерматоглифика в расоведении: таксономия рас и популяций // Папиллярные узоры: идентификация и определение характеристик личности (дактилоскопия и дерматоглифика). М.: АК, 2002. С.136-164.
159. Хить Г.Л. Долинова Н.А., Козлов А.И., Вершубская Г.Г. Угры Оби и уральская раса: дерматоглифический аспект // Вестник антропологии. Вып.2. 1996. С.111-128.
160. Хить Г. Л., Кейта Б. Дерматоглифическая дивергенция основных расовых ветвей человечества // Расы и народы. М., 1981. Вып. 11. С.25-39.
161. Хить Г.Л., Сихимбаева К.Б. Киргизы в дерматоглифической и одонтологической стратификации южносибирской расы // Вопросы этнической истории киргизского народа. Фрунзе,1989. С. 127-141.
162. Хить Г.Л., Халдеева Н.И. Внутригрупповые связи элементов дерматоглифики и одонтологии // Вопросы антропологии. Вып.52. 1976. С.101-107.
163. Хить Г.Л., Шинкаренко В.С., Наумкин В.В. Кожные узоры кисти у сокотрийцев // Наумкин В.В. Сокотрийцы. Историкоэтнографический очерк. М. 1988. С.54-71.
164. Хить Г.Л., Шинкаренко B.C., Наумкин В.В. Дерматоглифика населения островного Йемена. // Хадрамаут: Археологические, этнографические и историко-культурные исследования. (Труды Советско-Йеменской комплексной экспедиции). М., 1995. Т.1. С.480-502.
165. Ходжайова Г.К. Этническая антропология Узбекистана: Автореф. дис. ... д.и.н. Ташкент, 1997. 47 с.
329

166. Ходжайова Г.К. Дерматоглифика народов Средней Азии. М., 2007. 207 с.
167. Хрулев А.А., Аполлонова И.А., Спиридонов И.Н. Разработка алгоритма формирования изображений пальцевых узоров // Биомедицинские технологии и радиоэлектроника. 2005. №9.С.61-66.
168. Чебоксаров Н.Н., Чебоксарова И.П. Народы, расы, культуры. М.,1985. 149 с.
169. Широбоков И.Г. Антропологический состав карел по данным дерматоглифики // Микроэволюционные процессы в человеческих популяциях. СПб.: МАЭ РАН, 2009а. С.268 -293.
170. Широбоков И.Г. Частота радиальных узоров: новыйдерматоглифический признак и его таксономическое значение для дифференциации популяций Старого Света // Вестник антропологии. 2009б. Вып.17. С.163-174.
171. Широбоков И.Г. Антропологический состав и проблема происхождения карел по данным дерматоглифики. Автореф. дис. ... к.и.н. СПб, 2010а. 26 с.
172. Широбоков И.Г. Проблема формирования антропологического состава населения севера европейской части России по данным дерматоглифики // Вестник МГУ. Серия XXIII. Антропология. Вып. 2. 2010б. С.77-88.
173. Шпак Л.Ю. К вопросу о наследовании папиллярных узоров // Научный альманах кафедры антропологии. 2006. Вып.5. С.48-67.
174. Юсупов Р.М. Краниология башкир. Л.: Наука, 1989. 200 с.175. Ярхо А.И. Против идеалистических течений в расоведении
СССР // Антропологический журнал. 1932. №1. С.9-23;176. Ярхо А.И. Алтае-Саянские тюрки (антропологический
очерк). Абакан: Хакасское обл. нац. изд-во, 1947. 148 с.177. Ящук Е.В. Проверка одной из гипотез наследования пальцевых
узоров человека // Генетика. 1985. Т. XXL. №11. С.1909-1917.178. Acree M.A. Is there a gender difference in fingerprint ridge density?
// Forensic science international. Vol.102. 1999. P.35-44.179. Arrieta М.Г, Ibarrondo M.A., Lostao C.M. Digital dermatoglyphics
in the Basque population: univariate and multivariate comparison with other Spanish populations // American Journal of Physical Anthropology. 1987. Vol.73. P.89-98.
180. Astrom P. The study of ancient fingerprints // Journal of ancient fingerprints. Vol.1. 2007. P.2-3.
330

181. Babler W.J. The prenatal origins of population differences in human dermatoglyphits. Thesis, University of Michigan. 1977.
182. Babler W.J. Prenatal selection and dermatoglyphic patterns // American Journal of Physical Anthropology. 1978. Vol.48 (1). P.21- 27.
183. Babler W.J. Prenatal Development of Dermatoglyphic Patterns: Associations with Epidermal Ridge, Volar Pad, and Bone Morphology // Collegium Anthropologicum. 1987. Vol.11 (2). Р.297 -304.
184. Babler W.J. Prenatal Communalities in Epidermal Ridge Development // Trends in Dermatoglyphic Research. Dordrecht: Kluwer Academic Press, 1990. P.54-68.
185. Babler W.J. Embryologic Development of Epidermal Ridges and Their Configurations // Dermatoglyphics: Science in Transition. New York, 1991. P.95-112.
186. Bartsocas C.S. Paleodermatoglyphics // Progress in dermatoglyphic research (Progress in clinical and biological research. Vol.84). New York, 1982. P.139-143.
187. Bhasin M.K. Genetic of castes and tribes of India: dermatoglyphics // International journal of human genetics. 2007. Vol.7(2). P.175-215.
188. Bonnevie K. Studies on papillary patterns of human fingers // Journal of Genetics. 1924. Vol.15. P. 1-111.
189. Bonnevie K. Was lehrt die Embryologie der Papillarmuster uder ihre Bedeutung als Rassen- und Familiencharakter? // Zeitschrift fur Induktive Abstammungs- und Vererbungslehre. 1929. N.2, Bb.50. P.219-274.
190. Bonnevie K. Zur genetic des quantitativen Wertes der Pappilarmuster // Zeitschrift indikt. Abstain mungs und Vererbhungslehre. 1931. Bb.59. P.160.
191. Branigan K., Papadatos Y., Wynn D. Fingerprints on early minoan pottery: a pilot study // The annual of the British school of Athens. №97. London, 2002. P.49-53.
192. Carlson B. Human Embryology and Development Biology. New York, 1999.
193. Cheng X., Li H., Gupta S., Pan S., Hou J., Jin L. Dermatoglyphic changes during the population admixture between Kam and Han Chinese // Homo. 2009. Vol.60. P.143-157.
194. Cho Ch. A finger dermatoglyphic study of Maori // Korean Journal of Biological Sciences. 1998a. Vol.2. P.277-280.
331

195. Cho Ch. A finger dermatoglyphics of the New Zealand-Samoans // Korean Journal of Biological Sciences. 1998b. Vol.2. P.507-511.
196. Cho Ch. Finger dermatoglyphics of Australian aborigines in the northern territory of Australia // Korean Journal of Biological Sciences. 2000. Vol.4 (1). P.91-94.
197. Choudhury K.R. A study of dermatoglyphics among the three groups of Kondhs of Orissa // International journal of anthropology. 2005. Vol.20. P.27-38.
198. Coon C.N. The living races of man. New York, 1965.199. Cromwell H., Rife D.C. Dermatoglyphics in relation to functional
handedness // Human Biology. 1942. Vol. 14 (4). Р.516-526.200. Cummins H. Epidermal-ridge configurations in developmental
defects, with particular reference to the ontogenetic factors which condition ridge direction // American Journal of Anatomy. 1926. Vol.38 (1). P.89-151.
201. Cummins H. The topographic history of the volar pads (walking pad, Tasfballen) in the human embryo. Contributions to embryology: Carnegie Institution of Washington. 1929. Vol.20. P.103-126.
202. Cummins H. Racial differences in finger-prints // Journal of dimma! law and a'iminology. 1935. Vol.25 (5). P.829-835.
203. Cummins H. Ancient finger prints in clay // Journal of criminal law and criminology. Vol.32. №4. 1941. P.468-481.
204. Cummins H., Midlo C. Finger prints, palms and soles: intoduction to dermatoglyphics. Philadelphia, 1943; Philadelphia, New York: Dover Publications, 1961. 300 р.
205. Cummins H., Waits W.J., McQuitty J.T. The breadths of epidermal ridges on the finger tips and palms: a study of variation // Journal of anatomy. Vol. 68. №1. 1941. P.127-150.
206. Dankmeijer J. Some anthropological data on finger prints // American Journal of Physical Anthropology. 1938. Vol.23 (4). P.377-388.
207. David T.J. The Corrected atd Angle // Human Heredity. 1981. Vol.31. P.283-285.
208. Dell D.A., Munger B.L. The early embryogenesis of papillary (sweat duct) ridges in primate glabrous skin: The dermatotopic map of cutaneous mechanoreceptors and dermatoglyphics // Journal of Comparative Neurology. 1986. Vol.244. P.511-532.
209. Demarchi D.A., Seisdedos L. Dermatoglyphics of the Chorote Indians and their relationships with other inhabitants of the Gran Chaco. Annals of Human Biology. 1996. Vol.23(5). P.363-373.
332

210. Demarchi D.A., Marcellino A.J. Dermatoglyphic relationships among South Amerindian populations // Human Biology. 1998. Vol.70 (3). P.579-596.
211. De Wilde A.G., Van Meulenaarsgraf H., Pleeging J.H., Amesz- Voorhoeve W.H.M. Classification of fingerpatterns // Collegium Anthropologicum. 1981. Vol.2. P.135-146.
212. Elderton E.M. On the inheritance of the finger-print // Biometrika 1920. Vol.13. P.57-91.
213. Faulds H. On the skin-furrows of the hand // Nature. 1880. Vol.22. P.75-84.
214. Fields R., Molina D.C. A novel approach for fingerprinting mummified hands // Journal of Forensic Sciences. 2008. Vol.53. P.952-955.
215. Fox K.M., Garruto R.M., Gajdusek D.C., Plato C.C. Dermatoglyphics of the isolated Kapingamarangese of Micronesia // // Trends in Dermatoglyphic Research. Dordrecht: Kluwer Academic Press, 1990. P.278-286.
216. Galton F. Finger-prints. London: Macmillan and Co., 1892. 310 p.217. Geipel G. Das Tastleistensystem der Hande bei Vorderindern //
Zeitschrift fur Morphologie und Anthropologie. 1961. Bd.51. №2. S.181-207.
218. Glanville E.V. Heredity and Line A of Palmar Dermatoglyphics // American Journal of Human Genetics. 1965. Vol.17. Is.5. P.420-424.
219. Gould E. A topographical study of the differentiation of the dermatoglyphics in the human fetus. Tulane University, Thesis, 1948.
220. Green H., Thomas J. Pattern formation by cultured human epidermal cells: development of curved ridges resembling dermatoglyphs // Science. 1978. Vol.200. №4348. P.1385-1388.
221. Gruneberg H. Untersuchungen uber die Asymmetrie der Tastfiguren // Zeitschrift fur Induktive Abstammungs- und Vererbungslehre. 1928. Vol.47. Is.1. P. 79-87.
222. Gualdi-Russo E., Facchini F., Brasili-Gualandi P., Gruppioni G., Pettener D. Digital and palmar dermatoglyphics and the population structure of the Upper Bologna Apennine // Annals of Human Biology. 1994. Vol.21. P.519-532.
223. Gungadin S. Sex determination from finger ridge density // Internetjournal of medical update. Vol.2 (2). 2007,http://www.geocities.com/agnihotrimed/paper01Jul-dec2007.htm
224. Gutierrez-Redomero E., Alonso C., Romero E., Galera V. Variability of fingerprint ridge density in a sample of Spanish Caucasoids and its
333

application to sex determination // Forensic science international. Vol.180. 2008. P.17-22.
225. Gutierrez-Redomero E., Sanchez-Andres A., Rivalderia N., Alonso-Rodriguez C., Dipierri J.E., Martin L.M. A comparative study of topological and sex differences in fingerprint ridge density in Argentinian and Spanish population samples // Journal of Forensic and Legal Medicine. 2013, http://dx.doi.org/10.1016/j.jflm.2012.12.002
226. Hajn V., Gasiorowski A. Quantitative values on fingers and palms in Czech and Polish populations. Biologica. 1999. Vol.37. P.107-115.
227. Hale A.R. Breadth of epidemral ridges in the human fetus and its relation to growth of the hand and foot // The Anatomical Record. 1949. Vol.105 (4). P.763-776.
228. Hale A.R. Morphogenesis of volar skin in the human fetus // The American Journal of Anatomy. 1952. Vol.91 (1). P.147-180.
229. Heet H.L. Dermatoglyphics of the Finno-Ugrians of Eurasia // Trends in Dermatoglyphic Research. Dordrecht, The Netherlands: Kluwer Academic Publishers, 1990. P.218-233.
230. Heet H.L., Keita B. Dermatoglyphic divergence of the main racial branches of Mankind // Dermatoglyphics: fifty years later. Birth defects, original article series. New York, 1979. Vol.15. P.249-260.
231. Henry E.K. Classification and uses of finger prints. London, 1934. 143 p.
232. Herschel W.J. Skin furrow of the hand // Nature. 1880. №23. P.76.233. Hirsch W., Schweichel J.U. Morphological evidence concerning the
problem of skin ridge formation // Journal of Mental Deficiency Research. 1973. Vol.17 (1). P.58-72.
234. Holt S. The genetics of dermal ridges. Springfield, C. C. Thomas, 1968. 195 р.
235. Human Mummies: A Global Survey of their Status and the Techniques of Conservation (The Man in the Ice). New York, 1996.
236. Humphrey T. Some correlations between the appearance of human fetal reflexes and the development of the nervous system // Progress in Brain Research. 1964. Vol.4. P.93-135.
237. Igbigbi P.S., Msamati B.C. Palmar and digital dermatoglyphic patterns in Malawian subjects // East African Medical Journal. 1999. Vol.76. P.668-671.
238. Igbigbi P. S., Msamati B.C. Palmar and digital dermatoglphic traits of Kenyan and Tanzanian subjects. West African Journal of Medicine. 2005. Vol.24 (1). P.26-30.
334

239. Ismail E., Razak S.A., Selamat L., Gurusamy R., Zariman H., Shahrudin M.S., Amini F., Bakar Y., Nor S., Osman M. Dermatoglyphics: Comparison between Negritos Orang Asli and the Malays, Chinese and Indian // Sains Malaysiana. 2009. Vol.38 (6). P.947-952.
240. Jakobsson M., Scholz S.W., Scheet P., Gibbs J.R., VanLiere J.M., Fung H.-C., Szpiech Z.A., Degnan J.H., Wang K., Guerreiro R., Bras J.M., Schymick J.C., Hernandez D.G., Traynor B.J., Simon-Sanchez J., Matarin M., Britton A., van de Leemput J., Rafferty I., Bucan M., Cann H.M., Hardy J.A., Rosenberg N.A., Singleton A.B. Genotype, haplotype and copy-number variation in worldwide human populations // Nature. 2008. Vol.451. P.998-1003.
241. Jantz R.L., Brehme H., Eriksson A.W. Dermatoglyphic variationamong Finno-Ugric speaking populations: methodologicalalternatives // American Journal of Physical Anthropology. 1992. Vol.89. P.1-10.
242. Jantz R.L., Chopra V.P. A comparison of dermatoglyphic methodologies in population studies // American Journal of Physical Anthropology. 1983. Vol.60. P.61-67.
243. Kamali S., Mavalwala J. Diversity of palmar pattern ridge counts in Iranian populations // American Journal of Physical Anthropology.1990. Vol.81. P.363-373.
244. Kamali S., Mavalwala J., Khaneqah A.A., Bhanu B.V. Qualitative dermatoglyphic traits as measures of population distance // American Journal of Physical Anthropology. 1991. Vol.85. P.429-450.
245. Kamp K.A., Timmerman N., Lind G., Graybill J., Natowsky I. Discovering childhood: using fingerprints to find children in archaeological record // American antiquity. Vol.64. №2. 1999. P.309-315.
246. Kavgazova L., Stoev R., Mitova Z. Dermatoglyphic characteristics of a population from the Central Rhodopes (South Bolgaria) // Anthropologischer Anzeiger. 1999. P.349-360.
247. Katayama K. Dermatoglyphics of Native Polynesians in the Cook Islands, and their biological positioning among South Pacific populations // Trends in Dermatoglyphic Research. Dordrecht: Kluwer Academic Press, 1990. P.258-277.
248. Keith H.H. Racial differences in the papillary lines of the palm // American Journal of Physical Anthropology. 1924. Vol.7. P.165-206.
335

249. Kimura S., Kitagawa T. Embryological development of human palmar, plantar, and digital flexion creases. // Anatomical Record.1986. Vol.216. P.191-197.
250. Kimura S., Schaumann B. Embryological development and prevalence of thumb flexion creases // Anatomical Record. 1988. Vol.222. P.83-89.
251. King S., Mancini-Marie A., Brunet A., Walker E., Meaney M.J., Laplante D.P. Prenatal maternal stress from a natural disaster predicts dermatoglyphic asymmetry in humans // Development and Psychopathology. 2009. Vol.21. P.343-353.
252. Kralik M., Novotny V. Epidermal ridge breadth: an indicator of age and sex in paleodermatoglyphics // Variability and evolution. 2003. Vol.11. P.5-30.
253. Kralik M., Nejman L. Fingerprints on artifacts and historical items: examples and comments // Journal of ancient fingerprints. 2007. Vol.1. P.4-15.
254. Кйcken M. Models for fingerprint pattern formation // Forensic Science International. 2007. Vol.171. P.85-96.
255. Leche S.M. Handedness and bimanual dermatoglyphic differences // The American Journal of Anatomy. 1933. Vol.53 (1). P.1-53.
256. Loesch D. Genetics of dermatoglyphic patterns on palms //Annals of human genetics. 1971. Vol.34 (3). P.277-293.
257. Mavalwala J. Harold Cummins - and the birth, growth and development of dermatoglyphics // American Journal of Physical Anthropology. 1975. Vol.42. P.177-182.
258. Mavalwala J. A Methodology for Dermatoglyphics - Finger and Palms // Dermatoglyphics. An International perspective. The Hague: Mouton, Chicago. 1978. P.19-54.
259. Micle S., Kobyliansky E. Asymmetry and diversity of dermatoglyphics // Homo. 1992. Vol.42 (1). P.21-42.
260. Micle S., Kobyliansky, Arensburg B., Nathan H. Whorl patterns on fingertips: Their classification as based on the proportionality of their ulnar and radial ridge counts // Journal of Human evolution. 1982. Vol.11 (6). P.487-491.
261. Milicic J., Ivicevic A. Dermatoglyps of the population from the island of Rab (Croatia) // International journal of anthropology. 2005. Vol.20. P.13-26.
262. Milicic J., Vidovic M. Latent structure of dermatoglyphs in the population of Selska Valley // Homo. 2005. Vol.56. P.161-172.
336

263. Moran K.S. Unintentional artefacts: fingerprinting material culture // Journal of ancient fingerprints. 2007. Vol.1. P.16-18.
264. Mueller B. Untersuchungen uber die Erblichkeit von Fingerbeerenmustern unter besonderer Berucksichtigung rechtlicher Fragestellungen // Zeitschrift fiir induktive Abstammungs-und. Vererbungslehre. 1930. №56. P.302-382.
265. Mueller B. Die Lehre von der Erblichkeit des Reliefs der Hohland und der Fingerbeeren vom gerichtlichmedizinischen Standpunkt aus. Deutsche Zeitschrift fur die Gesamte Gerichtliche Medizin. 1931. №17. P.407-425.
266. Nagy N.S., Pap M. Pattern influence on the fingers // HOMO: Journal of Comparative Human Biology. 2005. Vol.56. P.51-67.
267. Namouchi I. Anthropological significant of dermatoglyphic trait variation: an intra-Tunisian population analysis // International journal of modern anthropology. 2011. Vol.4. P.12-27.
268. Nayak C.V., Rastogi P., Kanchan T., Lobo S.W., Yoganarasimha K., Nayak S., Rao N.G., Kumar G.P., Shetty B.S.K., Menezes R.G. Sex differences from fingerprint ridge density in the Indian population // Journal of forensic and legal medicine. 2010a. Vol.17. P.84-86.
269. Nayak C.V., Rastogi P., Kanchan T., Yoganarasimha K. Kumar G.P., Menezes R.G. Sex differences from fingerprint ridge density in Chinese and Malaysian population // Forensic science international. 2010b. Vol.197. P.67-69.
270. Newman H.H. Dermatoglyphics and the problem of handedness // The American Journal of Anatomy. 1934. Vol.55 (2). P.277-322.
271. Nithin M.D., Balaraj B.M., Manjunatha B., Mestri S.C. Study of fingerprint classification and their gender distribution among South Indian population // Journal of forensic and legal medicine. 2009. Vol.16. P.460-463.
272. Nithin M.D., Manjunatha B., Preethi D.S., Balaraj B.M. Gender differentiation by finger ridge count among South Indian population // Journal of forensic and legal medicine. 2011. Vol.18. P. 79-81.
273. Ohler E.A., Cummins H. Sexual differences in breadths of epidermal ridges on finger tips and palms // American Journal of Physical Anthropology. 1942. Vol.29. №3. P.341-362.
274. Okajima M., Iwayanagi Ch., Inouye E. Absence of palmar digital triradius d in Japanese twins and their families // Japanese Journal of Human Genetics. 1982. Vol.27. P.27-34.
337

275. Olivier G. Recherches sur les dermatoglyphes digito-palmaires // Bulletin et Memoires de la Societe d’Anthropologie de Paris. 1967. XVIe ser. Vol.1.
276. Osunwoke E.A., Ordu K.S., Hart J., Esomonu C., Tamunokuro F.B. A study on the dermatoglyphic patterns of Okrika and Ikwerre ethnic groups of Nigeria // Scientia Africana 2008. Vol.7 (2). P.143-147.
277. Penrose L.S. Finger-prints, palms and chromosomes // Nature. 1963. Vol.197. № 4871. P.933-938.
278. Penrose L.S. Dermatoglyphic topology // Nature. 1965. Vol.205. №6. P.544-546.
279. Penrose L.S. Memorandum on dermatoglyphic nomenclature // Birth Defects Original Article Series. 1968. Vol.4. №3. P. 1-13.
280. Penrose L.S., Loesch D. The effect of sex chromosomes characteristics of dermal ridges on palms and fingerprints // Genetica Polonica. 1970. Vol.10. P.328-331.
281. Penrose L.S., O’Hara P.T. The development of epidermal ridges // Journal of Medical Genetics. 1973. Vol.10. P.201-208.
282. Plato C.C. Polymorphism of the C-line of palmar dermatoglyphics with a new classification of the C-line terminations // American Journal of Physical Anthropology. 1970. Vol.33. P.413-420.
283. Plato C.C. Dermatoglyphic features: a review. // Materia Medica Graeca. 1976. №4. (Suppl. 3). P.246-257.
284. Plato C.C., Garruto R.M. Historical notes on dermatoglyphics: from Purkinje to Cummins // Trends in dermatoglyphic research. (Studies in human biology. Vol.1). Dordrecht-Boston-London, 1990. P.2-9.
285. Ploetz-Radman М. Die Hautleistenmuster der interen beiden Fingerlieder der menschlichen Hand // Zeitschrift f tr Morphologie und Anthropologie 1937. Bd.36. S.281-310.
286. Pospisil M.F. Ein Beitrag zum Problem der Erblichkeit der Papillarformationen der Fingerbeeren // Acta Facultatis Rerum Naturalium Universitatis Comenianae, Anthropologica. 1967. Bd.12. P.201-213.
287. Pospisil M.F., Ferak V. The dermatoglyphics of Egyptian Nubians // Trends in Dermatoglyphic Research. Dordrecht: Kluwer Academic Press, 1990. P.234-243.
288. Reddy B.M., Chopra V.P., Karmakar B., Malhotra K.C., Mueller H. Quantitative dermatoglyphics and population structure in Northwest India // American Journal of Human Biology. 2000. Vol.12. P.315- 326.
338

289. Reed T., Sprague F.R., Kang K.W., Nance W.E., Christian J.C. Genetic analysis of dermatoglyphic patterns in twins // Human Heredity. 1975. Vol.25(4). P.263-275.
290. Rosa A., Arquimbai R., Fananas L. Quantitative and qualitative palmar dermatoglyphics in the Mediterranean population of Delta de l'Ebre (Spain) // International journal of anthropology. 1998. Vol.13. P.89-96.
291. Rothhammer F., Сhakraborty R., Llop E. A collation of marker gene and dermatoglyphic diversity at various levels of population differentiation. // American Journal of Physical Anthropology. 1977. Vol.46 (1). P.51-60.
292. Sabir B., Cherkaoui M., Baali A. Characterisation of the digital dermatoglyphic in a berber population of the high Atlas (Marrakesh, Morrocco) // International journal of anthropology. 2005. Vol.20 (34). P.277-289.
293. Salzano F.M., F.R. Benevides De Sa E. Fingerprint quantitative variation and asymmetry in Brazilian Whites and Blacks // American Journal of Physical Anthropology. 2005. Vol.40 (3). P.325-328.
294. Sanna E., Usai E., Soro M.R., Floris G. Quantitative digital dermatoglyphics in males and females of four Sardinian linguistic groups // International journal of anthropology. 2005. Vol.20 (1-2). P.1-11.
295. Sanders G., Waters F. Fingerprint asymmetry predicta within sex differences in the performance of sexually dimorphic tasks // Personality and Individual differences. 2001. Vol.31. P. 1181-1191.
296. Scheil H.-G., Schmidt H.D., Baltova S., Djordjevic D., Vulpe C., Sivakova D., Efremovska L. Dermatoglyphic Studies in Eastern and South-Eastern Europe // Anthropologischer Anzeiger. 2005. Jahrg.63.H.4. P.393-399.
297. Schmidt C.W., Nawrocki S.P., Williamson M.A., Marlin D.C. Obtaining fingerprints from mummified fingers: a method for tissue rehydration adapted from the archeological literature // Journal of Forensic Sciences. 2000. Vol.45. P.874-875.
298. Schwidetzky I. Die neue Rassenkunde. Stuttgart, 1962.299. Segura-Wang M., Barrantes R. Dermatoglyphic traits of six Chibcha-
speaking Amerindians of Costa Rica, and an assessment of the genetic affinities among populations // Revista de biologia tropical. 2009. Vol.57 (Suppl. 1). P.357-369.
300. Seidenberg-Kajabova H., Pospisilova V., Vranakova V., Varga I. An original histological method for studying the volar skin of the fetal
339

hands and feet // Biomedical papers of the Medical Faculty of the University Palacky, Olomouc, Czechoslovakia. 2010. Vol.154 (3). P.211-218.
301. Sen J., Kanchan T., Mondal N. A comparison of palmar dermatoglyphics in two ethnic Indian populations of North Bengal, India // Journal of Forensic Sciences. 2011. Vol.56 (1). P. 109-117.
302. Sengupta M., Karmakar B. Genetics of Dermatoglyphic Asymmetry in Vaidyas of West Bengal, India // Human Biology. 2006. Vol.78 (2). P.199-214.
303. Sharma A. Comparative methodology in dermatoglyphics. Delhi, 1964.
304. Temaj G., Milicic J., Juric T.S., Behluli I., Narancic N.S., Hadzisellimovic R., Nefic H. Comparative analysis of dermatoglyphic traits in Albanian and Turkish population living in Kosovo // Collegium Anthropologicum. 2009. Vol.33 (4). P.1001- 1005.
305. Thoma A. Dermatoglyphics and the origin of races // Journal of Human evolution. 1974. Vol.3. P.241-245.
306. Tishkoff S.A., Kidd K.K. Implications of biogeography of human populations for “race” and medicine // Nature Genetics. 2004. Vol.36. P.21-27.
307. Van Valen L. A study of fluctuating asymmetry // Evolution. 1962. №16. P .125-142.
308. Weisensee K.E., Sivakova D. A comparison of Slovakian Haban populations using finger ridge count data // Studies of Tribes and Tribals. 2003. Vol.1. P.73-76.
309. Wertheim K., Maceo A. The critical stage of friction ridge and pattern formation // Journal of Forensic Identification. 2002. Vol.52 (1). P.35.
310. Whipple I.L. The ventral surface of the mammalian chiridium // Zeitschrift fur Morphologie und Anthropologie. 1904. Vol.7. P.261- 368.
311. Wilder H. Palms and soles // American Journal of Anatomy. 1902. Vol.1. Is.4. P.423-441.
312. Wilder H.H. Racial differences in palm and sole configurations // American anthropologist. 1904. №6. P.244-293.
313. Wilder H.H. Racial differences in palm and sole configurations. Palm and sole prints of Liberian natives // American anthropologist. 1913. №15. P.189-207.
340

314. Wilder H.H. Racial differences in palm and sole configurations. Palm and sole prints of Japanese and Chinese // American anthropologist. 1922. №15. P.143-206.
315. Yousefi-Nooraie R., Mortaz-Hedjri S. Dermatoglyphic asymmetry and hair whorl patterns in schizophrenic and bipolar patients // Psychiatry research. 2008. Vol.157. P.247-250.
316. Zhang H., Chen Y., Ding M., Jin L., Case D.T., Jiao Y., Wang X., Bai C., Jin G., Yang J., Wang H., Yuan J., Huang W., Wang Z., Chen R. Dermatoglyphics from all Chinese ethnic groups reveal geographic patterning // PloS One. 2010. Vol.5 (1):e8783.
317. Zhivotovsky L.A., Rosenberg N.A., Feldman M.W. Features of Evolution and Expansion of Modern Humans, Inferred from Genomewide Microsatellite Markers // American Journal of Human Genetics. 2003. Vol.72. P.1171-1186.
318. Zhou J., Gu J. Modeling orientation fields of fingerprints with rational complex functions // Pattern recognition. 2004. Vol.37. P.389-391.
341

SUMMARY
By the beginning of the XXI-st century, experts in the field of dermatoglyphics from different countries have failed to develop a common research agenda. A special program which includes calculations of finger and palm patterns and finger counts is used by most researchers. In most studies palm traits fall out of the researcher’s analysis, or are reduced to an estimated ridge count. In addition, there had been a decline in the number of papers devoted to the population and ethnogenetic topics in dermatoglyphics over the past two decades.
This book is dedicated to the description of instructional techniques, proven in the works of Russian specialists on ethnic and racial dermatoglyphics, the problems of populations’ differentiation on different taxonomic levels and the possible use of dermatoglyphics in the study of human populations in different ways. The authors use the term "race" to describe groups of populations with sufficient phenotypic and genetic similarity to distinguish them from other similar groups. Races are open to exchange genes with other races; racial characteristics are not absolute, have high intra-group variability and can change over time.
Key traits of dermatoglyphics
All collections of prints used in the book were analyzed according to the Cummins-Midlo (1961) methodology. Palmar axial triradius position was determined according to A. Sharma scheme, under which low triradius t localizes in the bottom of the 1/6 of a palmar longitudinal axis. Above this limit and up to the third shaft portion is a t' triradius zone, above this level - t" triradius zone (Sharma, 1964). Field 4 was not determined in the "classical" point version, and was extended to 3-4 mm above and below the midpoint of a palmar ulnar edge.
In 1969, H.L. Heet was the first to show the division of dermatoglyphic traits on the key and non-key elements. Subsequently, the idea was tested and confirmed on a large data set, covering major racial groups (Caucasoid, Mongoloid, Negroid, Australoid and Americanoid samples), as well as metis populations. Special studies then computed pair wise correlation between most dermatoglyphic evidence, except ridge count and angle values. Most of the signs have race-diagnostic
342

properties, but only a group of six key traits are not statistically related to each other on the within-individual level, i.e., not detect intra-ties. The key traits include: the Main line index (Ic), the Pattern intensity index (Dl10), the frequency of proximal axial triradius (t), the patterned hypothenar (Hy), the cumulative frequency of additional interdigital triradius (AIT), and the patterned thenar and the I-st interdigital pad (Th/I). The latter feature is of limited taxonomic value as methodologically difficult to define, does not distinguish between Caucasoid and Mongoloid groups, and within large and local races varies considerably wider than the other key features. Subsequently, I.G. Shirobokov has also shown high taxonomic value of the radial orientation of finger patterns (FRP) that weak and unstable correlation is related to the delta index. The method of determining the orientation of the finger patterns proposed by him is based on the skeleton structure features of a pattern (the position it over the open side) and is not associated directly with any type of pattern or with the value of ridge count.
Key features and FRP are associated with groups of secondary traits and reflect the major part of the palm morphological complex of papillary patterns and terminal phalanges. Key features show almost a normal curve within a race type, unlike most non-key features do. Distribution curves of the key features of all races exhibit large mutual transgression, wherein differences between the average characteristic values of races are usually statistically significant.
An important property of the key features is the Subordinate variability. The population variability of symptoms is subject to strict rules. Typically, the variability increases, and the diagnostic value decreases in the order: Ic - Dl10 - t - Hy - AIT - Th/I. Policies associated with valuable diagnostic criteria: the smaller it varies within a race, the more resistant characteristic of a race it is and the smaller probability of random variables are characteristic.
Sexual differences in the dermatoglyphics
Sexual differences of the dermatoglyphic traits are clear and direct in their nature, and therefore the analysis of pooled data for men and women leads to the distortion of the results of the study. Analysis of the 716 investigated in male and female samples of the five races (Caucasoid, Mongoloid, Negroid, Americanoid and Australoid samples) revealed the following patterns:
343

1. Key features of both sexes are fully correlated at the large races level, and this relationship is weakened and remains not in all cases within the race (at the population level). The values of attributes are more consistent between the sexes in Australoids and Americanoids, and the lowest in Caucasoids. Key features are typically correlated both sexes more closely than non-key features.
2. Key features in the male samples of all major races tend to have larger values, except for the hypothenar patterns. The latter feature is more common in the female samples in Caucasoids, Mongoloids and Americanoids, in the male samples - in Negroids and Australoids.
3. In all races sexual dimorphism coefficient (feature quantity expressing males in fractions in relation to such female) increases in the order: Ic - Dl10 - t - AIT - Th/I. (Patterned Hy was not considered, since this feature is not shared by all races the same vector intersexual differences). Likewise, the variability of these traits increases.
4. The maximum difference between sexes is shown by Negroids and the minimum by Australoids. Mongoloids and Americanoids converge in this respect to Australoids. Caucasoids are in the position between Australoids and Negroids.
Characteristics and taxonomy of big races and their subdivisions
Since the concept of race was originally based on somatological data, the dermatoglyphic material was organized in the framework of somatologic classification. The number of observations within a race for different traits varies greatly, reaching a maximum in respect of D lw A total of 1,322 samples were examined (756 men and 566 women), which are dominated by Caucasoids and Mongoloids. The analysis was performed by two methods - the intergroup scale and the principal components. Mongoloids and Australoids marked certain similarity and have high and very high values of Dl10 and reduced frequency of Hy and AIT. By the type of traits combinations they are approached to Americanoids. Typologically these three races form a unity. Negroids and Caucasoids are almost complete opposites: features have an average or low value in Caucasoids, and are strongly expressed in Negroids (AIT, Th/I), and vice versa (Ic, t, Hy). Only the delta index is almost undistinguished between these two races. After a series of statistical tests, the most isolated position is occupied by Caucasoids and Negroids,
344

Mongoloids and Australoids are characterized by a certain similarity, and Americanoids are close to the last two races (especially to the Mongoloids).
Bimanual distribution and asymmetry of the dermatoglyphic traits
Bimanual asymmetry of the key traits is seen on the dermatoglyphic materials of 20 male and 20 female parallel studied samples belonging to the three great races: Caucasoid, Negroid and Mongoloid (over 3600 people). Both in male and female samples, the value of the delta index, Cummins index and the frequency of radial patterns are identified on the right hand, and thenar and hypothenar patterned and frequency of palm proximal triradius are identified usually on the left hand.
In general, male population of the Old World at the level of interethnic comparisons revealed significant bilateral differences in all the main traits, except for the hypothenar patterns and frequency of AIT. Women recorded the same pattern, but bimanual differences in the hypothenar pattern are also significant.
Both in male and female samples, traits of the right hand have a higher taxonomic value to the inter-population differentiation at large distances than traits of the left. Thus, Caucasoid and Mongoloid sample differ from the values of all the key features of the right hand, with the exception of the thenar, while the number of features in the left arm, the differences are not significant to include, in addition to the thenar, the proximal triradius t, patterned Hy, as well as the frequency of FRP . In this case, as shown by the correlation analysis of principal components, the differentiation between male and female samples on the grounds of the right hand revealed a much greater similarity than on the grounds of the left hand.
Significant differences in the degree of severity of bimanual asymmetry for the most traits were revealed between men and women. More pronounced differences are characteristic for men, whereas in the female samples bimanual differences are smoother in their character. The only exception is a patterned hypothenar, which bimanual differences are more pronounced in women.
345

Dermatoglyphics and historical processes
The historical and geographical factors are decisive for the complexes of anthropological traits that define the degree of similarity and difference of human populations. The study of the system of dermatoglyphic traits described above allows reconstructing some evolutional stages of populations of different taxonomic levels, including the level of major races, and population formation histores, despite the fact the analytical framework is based on the materials collected in the XX-th century.
Dermatoglyphic differentiation within language groups had been repeatedly the aim of a special research. The data for the origins of genetic commonality in connection with the language similarities were produced on vast materials of Turkic, Finno-Ugric, North-Mongolic and Slavic languages. The results showed the dermatoglyphic heterogeneity of the Finno-Ugric and Turkic groups in various options and are not similar to a combination of both groups or a conditional addition of non- Mongoloids. Variants are associated with different contents of Caucasoid and Mongoloid components. One such option is common for the Finno- Ugric and Turkic, indicating the prevalence of self-sufficiency trend in the processes of racial or lingvo-genesis, respectively overcoming race or language barriers. The results of dermatoglyphic and linguistic analysis do not show similarities in both groups, only with one exception. Finnishspeaking people of the Circum-Baltic region (the Finns, Karelians, Estonians, and Vepsians) that somatologically represent the Northern Caucasoids, represent a single population according to the dermatoglyphic data. Thus, there was a coincidence of three parameters: language, racial type and dermatoglyphics. To a large extent the uniformity of dermatoglyphic and racial traits is shown by peoples of the Slavic group, with especially distinguished homogeneity of the Eastern Slavs: Russians, Ukrainians and Belarusians. However, in most cases genesis of a race is not associated with a genesis of language, and dermatoglyphic and language results do not match.
The chapter contains a bibliography of Russian and foreign studies on ethnic and population dermatoglyphic published over the past 25 years.
346

Paleodermatoglyphics
There is no reason to doubt that the papillary lines density of finger and palm patterns gradually decreases with the increase of a human body, as well as the fact that fingerprints of females on average have more finely and densely located papillary lines than in males. However, there are a number of barriers for creation of a universal scheme for determining personal sex and age based on papillary lines imprints in archaeological materials.
The average papillary lines density depends of at least four factors: 1. particular finger; 2. pattern type and its specific area; 3. constitutional features of a person; 4. inter-population variability of the feature. The difference between the papillary lines density calculated for different skin areas are in some cases statistically significant. On the other hand, there is a significant overlapping in sampled group values between adult males or females, and between adults and children.
Correlation of the dermatoglyphic traits and traits from the other systems
The population differentiation defined on different branches of physical anthropology (somatology, odontology, dermatoglyphics, craniology, serology, and etc.), often differ significantly, including the higher taxonomic levels. Traditionally, the differentiation was based on the assumption that each system of traits is controlled by different groups of genes and has a specific character depending on the nature of the features, and time and area of their formation. Obviously, one of the main reasons for the discrepancies lies in the fact that the definition of research agenda within the morphological features of any anthropological systems is based on artificially selected objects, including dermatoglyphics studies.
Match differentiation was found according to dermatoglyphics and somatology in the analysis of population racial composition of large samples of generalized historical and geographical regions of Russia and adjacent areas of northern Eurasia. Some differences emerged on the level of local North Eurasian races, where dermatoglyphic materials were organized in the frameworks of race-somatology classification. Studies have revealed two major components - Caucasoid and Mongoloid, along with a some metis component. Discrepancies of this scheme with data of
347

racial somatology (there is absent taxometric scheme) are both specific and fundamental. According to dermatoglyphic data, a hypothetical relationship of Pontic race with the southern Russian population is confirmed. Also, it is identified a close resemblance of South Siberian and Ural races, apparently originated from Mongoloids. A specifity of variant of the Central Asian Caucasoids (Pamir-Fergana) has been demonstrated. A specific position of the Amur-Sakhalin race is shown. Noticeable differences between the dermatoglyphic and somatology data were revealed in the study of ethnic groups of the Caucasus. Both taxometric schemes, while they are based on a limited number of ethnical samples, were compared. According to the dermatoglyphic data, Abkhazo-Adyghean peoples are distinguished from the Turkic (except Nogais). Abkhazo-Adyghean peoples also show the proximity within a somatological scheme.
An important property of dermatoglyphic traits was also studied. The investigation demonstrates a steady shift in the direction of increasing the Mongoloid component within ethnic samples of Eastern Europe compared to somatological data. This certainly indicates a more pronounced reaction of a dermatoglyphic system on the admixture of Mongoloid traits in a population.
348

ПРИЛОЖЕНИЯ

I. Heet H.L. Abstract of a thesis “Dermatoglyphics of the USSR peoples” (1983)
The results present in this study are reported in Russian and foreign languages in more than forty separate publications. The monograph numbers 280 pages and consists of the main text (pages from 1 to 191) and tables as the supplement (pages from 192 to 280).
The Introduction provides a definition of ethnic and racial dermatoglyphics and a brief historical sketch. Besides the founder of this discipline, the great British biologist F. Galton (1892), other scholars who made significant contributions to dermatoglyphics are mentioned, viz. G.Wilder (1904, 1922), H. Cummins and Ch. Midlo (1943). In Russia, the population studies of dermal relief were started by P.S. Semenovsky (1927) and continued by a number of scholars whose works are discussed further.
The population dermatoglyphic research in the former USSR has been irregular. Data reported by various authors are often methodologically incomparable, and no standardized sampling program is published too. The analytical procedure remains undeveloped. Consequently, up to the present it was impossible to integrate the earlier findings on a nation-wide scale. The author having set herself this task undertook a systematic study of peoples of the Soviet Union in 19571975, collecting materials during almost annual field trips. The Methodically comparable materials reported by other researchers that make up about one third of the data are also used. The total sample exceeds 50 000 individuals from more than 600 groups.
The main methods of analysis include as the following. The geographical distribution of trait values was examined, regionally specific complexes of racial traits were identified, ethnic groups and local races within regions were classified, and generalized characteristics of two major races - Caucasoid and Mongoloid - were compared. At the first two levels (population and ethnic) dermatoglyphical data were analyzed independently, without any external system of reference; further within the frame of racial (somatic) typology, mean phonetic distances between taxa belonging to various hierarchical levels were computer-based calculated and dendrograms were constructed on the basis of distance matrices by means of a weighted pair-group method.
350

In Chapter 1, “Methodological frameworks of the analysis” ,principles outlined by Soviet specialists in race studies (Yarkho, 1947; Debets, 1948; 1951; Alexeyev, 1969) are briefly formulated including as the following. Ideally only independent traits should be used. If the traits show some degree of within-group correlation, it should be opposite to between-group correlation. Complexes of traits intercorrelated on a group level should be identified. Traits have to be ranked according to their diagnostic value at different systematic levels. The traits showing the highest values for discriminating racial types are preferred.
In the paragraph, “Interdependence of dermatoglyphical traits”, correlations between dermal elements are analyzed at three levels:
1. intra-group (purely biological) variability;2. inter-group (regional or historical) variability;3. inter-racial variability.A set of main (or key) traits is segregated. These traits should be
independent against inter-group or extra-group variability that latter is historically or regionally specific. The key traits reflect major morphological properties of the groups under comparizon and are then used for statistical comparisons.
The paragraph, “Stability of the dermal relief elements system”, is devoted to the analysis of intra- and inter-group variability of the key features. The phenomenon of coordinated or hierarchical variability is detected: within groups, within regions, and within races trait variation increases in a fixed and invariable order, viz. main line index - pattern intensity index - proximal axial palmar triradius - hypothenar patterning - accessory interdigital triradii.
This rule doesn’t hold true for racially neutral characters showing increased variability (thenar patterning, 0, x, and 8-variant of C-line).
The inverse relationship between the level of variability and the taxonomic value of traits is established: the trait having the lowest variation of a race-specific feature within a race results in a diminished probability of random fluctuation and hence in the increase of its racial diagnostic value.
It was also found that key elements of dermatoglyphics are less variable than somatic racial traits, and are more sensitive to racial admixture. Also, Population differences in dermal characters are weaker thus indicating that dermatoglyphics reflects a more archaic stage of a race formation.
351

In the paragraph, “Taxonomic evaluation of differentiation levels according to dermatoglyphics traits”, divergences between groups at various taxonomic levels are analyzed. Comparative data are contradictive, the conclusions ranging from maximal divergence at a local group level (Gadzhiev, 1971; Tausik, Rychkov, 1977) to a complete absence of differences in variation among systematic levels (Rothammer, Chakrabortti and Llop, 1977).
The Generalized Dermatoglyphical Distance (GDD), which means the difference in 5 key traits divided by the Eurasian Range of Variation (ERV), is accepted as the measure of divergence in this study.
Male and female groups are analyzed separately with the use of Sex-specific Eurasian Range (SER).
The investigation was carried out at four levels within the majorraces:
1. populations or local groups within ethnic groups;2. ethnic groups within local races;3. local races within regions;4. regions within major races.In addition, as the highest inter-race level, differences between
average characteristics of two major races, Caucasoid and Mongoloid, have been analyzed.
It is found that the divergence of taxa and the nature of differences between the systematic levels in the degree of variation for both of the major races are the same. A hierarchical ordering of levels is established. Local groups within ethnic groups differ less than do ethnic groups. Interethnic level of differences is the highest within major races and amount to 70% of the divergence between major races. Local races are much more homogeneous than any of the other taxa. Thus, it is established that the ethnical barrier is the most powerful factor of group divergence within the major races.
A method for measuring GDD is suggested based upon a normal distribution of its frequencies. Five standard categories of GDD are established for each sex and taxonomic level in particular, viz. very small, small, medium, large and extra large. This gradation performed separately for each sex and each taxonomic level constitutes a basis for the evaluation of differences among taxa.
Chapter 2, “General dermatoglyphical characteristics of the populations of the former USSR” forms an introduction for further
352

analysis. Racial typologies based on somatic traits are briefly described; dermatoglyphical data pertaining to major regions (European region, Caucasus, Central Asia, Siberia and the Far East) are summarized. Distribution curves of the trait frequencies within regions show a considerable transgression. Both inter-racial and intra-racial regional tendencies (racial gradients) are clearly discernible both graphically and analytically. On the basis of trait frequencies, polygonic graphs, reflecting specific combinations of dermatoglyphical features, are drawn. The Eastern or Mongoloid Component (EC) estimate is computed, with its value being roughly proportional to the supposed relative amount of Mongoloid genes within a group.
The maximal differences were found between the Siberian Mongoloids as a whole and the peoples of the Caucasus. Caucasoids show an extremely Caucasoid combination of traits while the peoples of the Eastern Europe and particularly Central Asia (Turkmen, Tajiks and Uzbeks) include a significant proportion of Mongoloid admixture. A total Central Asiatic sample exhibits also some specific features that are resulted exclusively from the metisation between the Mongoloids and the Caucasoids.
Chapter 3 is titled “Anthropological composition of the individual regions according to dermatoglyphics” . At the beginning of each regional section, a list of cited literature and a brief description of the materials used are present. Geographic variations of traits are examined at a territorial level, racial complexes are calculated, and clustering of ethnic groups is performed on the basis of GDD matrices. The groups are ranked in the order of increasing of EC estimates. At the last stage, local races (or local types) are analyzed. This includes numerical characterization based on the combination polygons computation of EC and the deviation from a regional mean clustering on the basis of GDD, and reconstruction of genetic links.
East Europe. The material covers 207 local (125 male and 82 female) groups numbering in the total about 19 000 individuals; 94 groups (over 8 000 people) were investigated by the first author, and the rest was sampled by other researchers. In addition, the analyzed sample includes published data on the populations of Finland, viz. Finns, Swedes and Lapps.
353

The heterogeneity of dermatoglyphical traits among the populations analyzed is seen already at the territorial level. Two subregions of unequal size may be noticed. The first includes populations inhabiting most of the territory, up to the Volga River. Those populations are Caucasoids both in somatic and dermatoglyphical traits. Among them, a group of northern populations (Finns, Karelians, Vepses, Komi-Zyrians, western Estonians and Letts, as well as Mordvinians) shows a peculiar combination of reduced pattern intensity index, reduced hypothenar patterning and diminished number of accessory interdigital triradii.
The second sub-region includes the peoples living in Volga and Kama river basins as well as in Bashkiria (Mari, Udmurts, Chuvashes, Tatars, Komi-Permyaks and Bashkirs); Kalmyks and Lapps also belong to this complex. In all the main features, these groups deviate from a standard Caucasoid type in the eastern direction. As a whole the populations of this sub-region are much more heterogeneous and diverse than those of the first sub-region.
The same differentiation is observed at the level of ethnic groups. To show this, pair wise distances were calculated and cluster analysis was applied. EC-values for both sexes and a mean EC were computed. Two widely distinct clusters are visible on the dendrograms and correspond to sub-regional groups described above. The ranking of EC-values results in the same dichotomy.
Karaims occupy a peculiar position being maximally isolated from other populations both according to EC and the ethnic dendrogram. A combination of an extremely high main line index, very high hypothenar patterning and low percent of proximal axial palmar triradius suggests a genetic affiliation of Karaims with southern Caucasoids.
At the third level of analysis - local races - somatic racial classification developed by Alexeyeva (1973) is applied. Two groups of unequal size are segregated. The first may be described as Europeans, as it includes a bulk of local races (6 of 8) with typically Caucasoid combination of traits. Baltic, Lake Beloye-Kama, Valday-Upper Dneprian, Dneprian, Central-East European and Steppe types belong to this group. A relatively isolated position among them is occupied by northern variants - Baltic and Lake Beloye-Kama - that are quite similar and differ from the other local races by an increased proportion of Mongoloid component. Other types form a very homogeneous subgroup. The Caucasoid background shows the strongest expression in the Central-East European type.
354

A second group of local racial types, which is described as Eastern group, includes Volga-Kama and Uralian variants. Their features, in the comparison to members of the first group, have an increased value of Mongoloid component. This component is considerably more pronounced in the Uralian type than in the Volga-Kama type, and the distance between these two is greater than within any other pair of types in Eastern Europe.
Thus, various levels of analysis reveal certain conformity in the pattern of racial differentiation in Eastern Europe and Fennoskandia, according to both dermatoglyphical and somatic traits. One of the main features of race formation in the territory was the interaction of numerically predominant Caucasoid populations with groups including variable proportions of Mongoloid component.
Caucasus. The material analyzed consists of 107 local groups (about 9 000 individuals), almost equally distributed by sex. Eighty five groups (over 7 000 individuals) were studied by the author.
The geographical distribution of dermatoglyphical trait frequencies shows a wide and irregular variation. Two points should be stressed. First, the Caucasus is the only region in the former USSR, where certain regularities in distribution of thenar patterning may be noted, and those regularities are similar in males and females. The highest frequencies of these traits are observed in the Greater Caucasus, mainly in Chechen- Ingushetia and southern Dagestan. Second, populations of Western and Northwestern Caucasus show several peculiar features, i.e. low values of both indexes, reduced hypothenar and thenar patterning, and a low percentage of accessory interdigital triradii. All these are deviations from a typically Caucasoid trait complex.
On the ethnic level, the picture is more complicated and irregular both typologically and on the dendrograms based on GDD-values. The latter show that Khevsurs, Pshavs and Tsakhurs are sharply distinct from the other Caucasoid groups. The bulk of Caucasoid populations are divided among two clusters that are widely divergent and hardly bear any resemblance to somatic racial types.
All aboriginals of Dagestan (except Tsakhurs and Kumycks) are very close each other and to Ingushes, Azerbaijanians and Muslim Tats of Dagestan. Mountain Jews are totally isolated from the bulk of the populations by very high distances. Adygeyans closely resemble Noghayans, and this pair has not only a small GDD but identical values
355

of EC. The latter value sharply separates them from the rest of Caucasoid peoples. This has a parallel in a Mongoloid admixture found in cranial remains of Middle Age Adygeyans from burials antedating the Mongol invasion. This admixture is attributed to Kipchak influence (Alexeyev, 1974).
The analysis of dermatoglyphical data with reference to somatic racial schemes (Abdushelishvili, 1964; Alexeyev, 1974) reveals a biological unity of Western Asian and Caucasoid local races. Towards the periphery of this area, deviations from a classical Caucasoid trait combination become more pronounced. In the north-west and west, there are apparent Pontic race traits characteristic for the populations of south European part of the former USSR. In the east, in the West Caspian region, the Caspian local race resembles Trans-Caspian variants that include a proportion of the Indo-Australoid racial elements.
Thus, the dermatoglyphic data do not contradict to racial classifications established on the basis of somatic and cranial traits. While chronological and territorial issues of the race formation should be analyzed primarily based on the paleoanthropological material, dermatoglyphics may provide some indirect data. Specifically, several Caucasoid populations belonging mainly to the Caucasionic and West Asian local races - Chechens, Avars, Ossetians, Muslim Tats, Rutuls, and Udines - dermatoglyphically display not only a great mutual similarity but also a striking affinity to some geographically and socially isolated groups of Central Asia and India - Darvaz Tajiks, Yagnobians, several groups of Shugnanians and Bartangians, and the Parsee of Bombay. One can speculate that they are remnants of an archaic dermatoglyphical complex, which was distributed over the Caucasus, northern India, Iran, Afghanistan and Central Asia and included ancient Europoids of Asia thus having a common origin. Ancient trait combinations could have survived only in areas where various factors prevented intensive race admixture, and local conditions helped to preserve archaic racial types. This complex might have originated in the Neolithic or Bronze Age when the long-headed and narrow-faced Europoids predominated on a vast territory from the Mediterranean Sea to Central Asia and north-western India.
Central Asia. The sample numbers over 10 000 persons and includes 141 local groups with some predominance of males; 109 groups (about 8 500 individuals) were investigated by the author.
356

Apart from the descendants of an ancient autochthonic population (Turkmens, Tajiks, Uzbeks, Uyghurs, Pamir aboriginals, Yagnobians), immigrant Caucasoid groups (Baluchis, Kurds, Central Asian Jews, Iranians, Arabs) and Central Asian Mongoloids (Karakalpaks, Kazakhs, Kirghizians) were studied.
The geographical distribution of traits over local population reveals a broad range of variation and utmost heterogeneity. The Central Asian population is extremely diverse, this being due not only to the presence of Mongoloids, but also sharp divergences among local groups of Caucasoid people. The latter differ, firstly, in the proportion of Mongoloid admixture (which is minimal in Yagnobians, some groups of Tajiks and Pamirians, but is more pronounced in Turkmens and especially Uzbeks and Uyghurs). The second gradient of divergence within the Caucasoids is presumably explained by an Indo-Australoid influence, in the form of an increased number of accessory interdigital triradii that is shown by Turkmens and Baluchis, while Tajiks exhibit this tendency in the lowest degree.
Both male and female samples of Central Asian Caucasoids are statistically most heterogeneous on a country scale. This heterogeneity is most accentuated in Pamirians and Tajiks. In all samples, females show more variation than males, which is opposite to those seen in other region of the former USSR. It may be concluded that specific factors in ethnic and racial history of Central Asia resulted in a steep rise of local heterogeneity.
On the ethnic level, a less complicated pattern emerges. First, Kirghizians, Kazakhs and Karakalpaks stay close to each other and differ sharply from the other groups. Mongoloid traits are less expressed in Kazakhs and most apparent in Kirghizians.
Aboriginal Caucasoids - Turkmens, Tajiks, Pamirians, and entire Uzbeks - are also close to each other as well as to Kurds and Uyghurs. All these peoples display dermatoglyphical characteristics of the southern Caucasoid type in combination with some peculiar traits, viz. increased pattern intensity index and high frequency of proximal palmar triradius. They form a specific assemblage which could be designed the Central Asiatic complex.
Peoples belonging to this group differ in a relative proportion of Mongoloid component. It is minimal in Tajiks that are close to Pamirians, in which the Mongoloid admixture is higher. Turkmens display not only a substantial Mongoloid component, but also an Indo-Australoid feature -
357

an increased percent of accessory interdigital triradii. Uzbeks as a whole are closer to Turkmens than to Tajiks. The Examination of separate Uzbek samples shows that urban Uzbeks (descendants of “Sarts”) are minimally mongolized while the maximal Mongoloid influence is seen in the descendants of “Desht-i-Kipchaks”, while the descendants of “Turks” occupy an intermediate position.
Uyghurs are most close to Uzbeks, Kurds of Turkmenia - to Turkmens. The Immigrant Caucasoid groups are quite different from the autochthonic populations and as a rule differ from each other. Most isolated are the Baluchis, who combine features of southern Caucasoid and Indo-Australoid complexes. Persians (Iranians) stand in sharp contrast to all Central Asian populations and have a direct analogy in the Khevsurs of the Caucasus. Jews find no analogies whatever. The status of Yagnobians was discussed above.
Dendrograms based on GDD-values are different for males and females. Although, there are some common features including the separation of Mongoloids from other groups, isolated position of immigrant populations within Caucasoids, mutual closeness of Tajiks, Pamirians, Turkmens, Uzbeks and Uyghurs, as well as a peculiar position of Yagnobians. This pattern of differentiation is primarily due to the rate of admixture, which constitutes a major factor of race formation in most parts of Central Asia at least during the last two millennia. EC-values sharply discriminate the descendants of Caucasoid aboriginals from Iranian, Afghan and Arab immigrants, on one side, and from Mongoloid nomads, on the other.
The relationship of local races is simpler. Trans-Caspian race and the Central Asian Inter-fluvial (Oxus - Yaxart) race are almost identical and hence likely have a common origin. They differ somewhat in values of the basic southern Caucasoid element and Mongoloid admixture, both being greater in the Trans-Caspian race. The Central Asian variant of the South Siberian race, which includes Central Asian Mongoloids, is very specific and contrasts sharply with both Caucasoid races.
Siberia and the F a r East. The sample consists of 159 groups, about equally distributed between males and females, and numbering over 16 000 individuals; 136 groups (over 13 000 persons) were studied by the author.
The geographical variation exhibits a regular pattern with marked differences among populations. In western and southern Siberia,
358

deviations from the classical Mongoloid dermatoglyphical complex are common. Other sub-regions also display certain differences which are clearly discernible at higher levels of analysis as well.
At the ethnic level, heterogeneity of Mongoloids is revealed by the results of clustering and EC-ranking. Two clusters of unequal size are segregated. The first includes populations of Altay, Tuva, Mongolia, Soviet and non-Soviet Far East (except for Oroches and Udegheys), Sakhalin and Chukotka. The second cluster of a greater size includes all the remaining Siberian groups together with Noghays, Kalmycks, Kazakhs, Kirghizians and Karakalpaks. In the second cluster, Nivkhs and Eskimos stay close to each other, and to Chelkans; in the first cluster, Selkups, Oroches and Enets are separated from other populations. While, in general, the first cluster is more homogeneous than the second cluster, at least two components may be noted - the first showing distinctly Mongoloid features, and the second incorporating a certain amount of Caucasoid admixture. Divergences between the mixed components of both clusters are mainly quantitative whereas Mongoloid components are significantly different in trait combinations. Several ethnic groups are heterogeneous, especially Selkus, Mongols, Evenky, Nivkhs, Khanti, Mansi, Touvinians, and Nenets.
At the level of local races, the material was classified according to somatic racial schemes (Yarkho, 1947; Cheboksarov, 1951; Levin, 1958; Alexeyev, Gokhman, 1983). Uralian race was found to be dermatoglyphically (and presumably genetically) close to South Siberian race. The same is true to Baikalian and Mongolo-Siberian races. Within them, Uralian and Baikalian races conform to the classical Mongoloid complex in a lesser degree than do South Siberian and Mongolo-Siberian races. Arctic Mongoloids are more similar to Mongolo-Siberian than other local races; the Far Eastern variant has analogies in South East Asia. Amuro-Sakhalinian variant is the most specific, but shows a certain similarity to the Arctic variant.
Within the Uralian race, there is a group of populations combining “super-Mongoloid” traits with an increased main line index. This combination may be regarded as a relic of an archaic Mongoloid complex which has absorbed a certain proportion of Caucasoid component. The Altay-Sayanian region is discussed separately. The Altayans and Khakassians are close to each other, while the former are less homogeneous and more mongolized than the latter. Both are similar to a generalized West Siberian sample.
359

Chapter 4, “Differentiation of racial types of the former USSR”. The findings of preceding analysis have been integrated and a geographical distribution of key traits is summarizes over the whole former USSR. Separate traits together with EC-values allow tracing, on the population level, a gradual transition from Caucasoid to Mongoloid extreme in the eastern direction.
As the result, a single system is created, incorporating data on all 20 local races: 1. Baltic; 2. Valday - Upper Dneprian; 3. Dneprian; 4. Central - Eastern European; 5. Lake Beloye - Kama; 6. Volga - Kama; 7. Steppe; 8. Pontic; 9. Caucasionic; 10. Western Asian; 11. Caspian; 12. Trans-Caspian; 13. Pamir-Fergana; 14. Uralian; 15. Southern Siberian; 16. Central Asiatic; 17. Baikalian; 18. Arctic; 19. Far Eastern; 20. Amuro-Sakhalinian. Based on this system, seven European (1 -7), four Caucasoid (8-11), two Central Asian (12 and 13) and seven Siberian and Far Eastern (14-20) variants were studied.
The analysis of trait distribution, EC-values, combination types and GDD clustering result in the similar conclusions. Two groups of racial variants are defined. They may be described as Mongoloid and Caucasoid groups according to the somatic racial attribution. Within each group, some local races form a transition to another group. The following text explains this pattern.
Caucasoids (variants 1-11, except Volga-Kama race) exhibit a regular succession with gradual changes, from the north to the south, of traits: pattern intensity index, hypothenar patterning and percentage of accessory interdigital triradii increase, while the frequency of low palmar triradius diminishes. Following groups of races may be segregated:A) Northern local races - Baltic and Lake Beloye-Kama -have a low
pattern intensity index and low frequencies of AIT and Hy combined with an increased percentage of proximal palmar triradius. The whole complex except the first trait suggests a considerable Mongoloid admixture. EC-values range from 43 to 45;
B) Central European local races (Valday-Upper Dneprian, Dneprian, Central-East European, Steppe) are quite close each other and to a generalize East European sample. EC-values are 40 to 41. It is interesting that the Pontic race, localized in the Caucasus, reveals a certain similarity to this group and forms a transition between East European and Caucasoid races;
360

C) Caucasoid local races, for which Caucasionic and West Asian traits are most typical. All races show characteristic features suggesting the South Caucasoid origin: high values of MLI and Hy, low frequency of proximal palmar triradii, increased per cent of finger delts. EC-values are very small (34-39), and would be even less if it may be technically possible to differentiate the contribution of PII to the racial source (southern Caucasoid or Mongoloid) of increased deltal number.
Caspian races are peculiar in combination of typical Caucasoid features with a high frequency of AIT and a maximal value of PII. This makes Caspian races transitional between Caucasoid and Central Asian local races. The latter - Trans-Caspian and Central Asian Interfluvial - races are mutually close, displaying Caucasoid (high MLI, increased Hy), Mongoloid (increased percentage of proximal palmar triradius) and Indo- Australoid traits (high PII, increased frequency of AIT), with EC-values varying at 49-51. In all traits reflecting the genetic heterogeneity of their component parts, both local races do not fully conform to the typical Caucasoid standard. Both have a predominant Caucasoid element falling into the same cluster as the Caucasoids on the dendrogram and showing Caucasoid EC-values.
An isolated position is occupied by the Volga-Kama local race which differs from the remaining European variants by a maximal EC- value (52) and constitutes a Caucasoid-Mongoloid transition variant in all traits, except the low pattern intensity index. On the dendrogram, this race is linked (though only by medium to large distances) with the northern Caucasoids and forms one sub-cluster with the latter; this indicates that the similarity may be due to Mongoloid admixture. EC- value within this sub-cluster is 47 which are considerably higher than the mean value (41) within the sub-cluster that includes main Caucasoid variants.
Mongoloids (variants 14-20) incorporate local races of western and southern Siberia (Uralian and South Siberian) that deviate from the Mongoloid standards. Compared to the generalized Mongoloid sample and to other variants, they show increased main line index, hypothenar patterning and AIT frequency. Both races are quite similar, Uralian having a slightly lower EC (62) than South Siberian (63) race.
361

Inner Siberian variants - Mongolo-Siberian and Balkalian - are virtually indistinguishable. EC is high, averaging 70 and 71; the same is also true for other Mongoloid races, except the Uralian and South Siberian ones.
Arctic Mongoloids are close to Balkalian and Mongolo-Siberian races and differ with a reduced PII and a very low “super-Mongoloid” Hy. Far Eastern and Amuro-Sakhalinian local races are most specific. Extremely low MLI and a reduced percent of proximal palmar triradius separate the Far Eastern race from all Siberian variants and link it with Mongoloids of South Eastern Asia. In the Amuro-Sakhalinian variant, trait combination is very unusual, with relatively low PII, increased Hy and exceedingly high frequency of proximal palmar triradius. First two of these features are most probably due to Ainu admixture. The Amuro- Sakhalinian variant is the most isolated population among all Mongoloid variants.
Summarizing the results, it should be noted that Caucasoid races undoubtedly sharing a common genetical heritage are differentiated primarily along the North to South gradient and have a large number of intermediate types. Within northern Caucasoids, it is evident a substantial Mongoloid admixture which affected all key characters except the PII. Low values of this trait are no less typical of the northern Caucasoid than a strong depigmentation.
East European and Caucasoid variants are connected by the Pontic race, which occupies an intermediate position between them. Also, Caspian race presents a link between local races of the Caucasus and Central Asia. This race reveals certain traces of Indo-Australoid influence, which are even more distinct in Central Asiatic races. Local races of Central Asia cannot be reliably attributed to any major race, as they contain at least three components, namely (in decreasing succession), Caucasoid, Mongoloid and Indo-Australoid. The latter two are stronger in the Trans-Caspian variant. Another intermediate variant is the Volga-Kama race which constitutes a direct transition from northern Caucasoid to Mongoloid forms.
Mongloid local races are somewhat more homogeneous in comparison to Caucasoid races. Uralian, Southern, Baikallian and Mongolo-Siberian are morphological variations of a single type; its characteristics are less distinct in the first two races due to a pronounced Caucasoid admixture. The mixed origin of the South Siberian race is well
362

documented by paleoanthropological materials. Dermatoglyphically, both races are virtually identical. Such a degree of similarity at a relatively high level obviously could not result from convergence, so we are forced to accept the hypothesis of a common ancestor for both these races. They diverge only in the proportion of the Caucasoid admixture which is more accentuated in Uralian than in South Siberian race.
Thus, the major part of Siberia is populated by similar and closely related groups. Peoples inhabiting western and southern frontiers of Siberia reveal distinct traces of intermixture with Caucasoids.
Of the remaining Mongoloid variants, the Arctic race is close to Inner-Siberian Mongoloids (i.e. Mongolo-Siberian and Baikalian races) and presumably is genetically related to them. The Far Eastern race has no analogies in Siberia and resembles the South East Asian Mongoloids. The Amuro-Sakhalinian variant combines features of northern and southern Asiatic Mongoloids together with evidence of Ainu admixture, and has a maximal value of the basic Mongoloid component.
363

II. Список дерматоглифических серий, находящихся на хранении в ИЭА РАН и МАЭ РАН
СЕРИИ ИЭА РАН
Этническаягруппа
n,пол
Регион Автор, время сборов
Европейская часть России и Скандинавия
Башкиры 21& Башкирия, Пермский эксп. Р.М. Юсупова18? край, Самарская обл.,
Саратовская обл., Оренбургская обл.
1983, 2001, 2004-2005. Н.А. Суворова, 2000-06
Белорусы 2&2?
Белоруссия Г.Л. Хить, 1965.
Болгары 3&3?
Молдавия, Болгария Н.А. Никольская, 1975; З. Митова, 1988;А. Варзарь, 2001.
Вепсы 4& Карелия, Н.Л. Прокудина, 1972;4? Ленинградская и
Вологодская обл.эсп. Н.Н. Цветковой, 1983-89; Г.Л.Хить, 1965;
Венгры 1&1?
Украина Г.Л. Хить, 1970
Г агаузы 1&1?
Молдавия Н.А. Никольская, 1975
Калмыки 3&3?
Калмыкия Н.А. Прокудина, 1972; К.Б. Сихимбаева, 1985.
Карелы 3&3?
Карелия Г.Л. Хить, 1965;Н.А. Прокудина, 1972
Коми- 10& Мурман. обл., Г.М. Давыдова, 1968;зыряне 7? Тюмен. обл. и
Пермский крайН.А Прокудина, 1972; Г.А. Аксянова, 1976; Н.А. Дубова, 1981.
Латыши 2&2?
Латвия Г.Л. Хить, 1965
Литовцы 2&2?
Литва Г.Л. Хить, 1965, 1972.
Марийцы 5&2?
Марий Эл А.В. Шевченко, 1970; Н.А. Прокудина, 1971.
Молдаване 1&1?
Молдавия Н.А. Никольская, 1975.
Мордва 73
Мордовия Н.А. Прокудина, 1971; Н.Н. Цветкова, 1980-е
364

Этническаягруппа
n,пол
Регион Автор, время сборов
Нагайбаки 1& Челябинская обл. А.И. Макеева, 2012
Русские1?37^, Разные области Г.Л. Хить, 1965;36? Европейской части И.И. Гохман,
Саамы 4&
России; Карелия; Якутия; Армения
Финляндия,
В.П. Алексеев, 1965; В.Д. Дьяченко, 1965; Г.М. Давыдова, 1966; Н.А. Прокудина, 1969, 1971; Н.А. Дубова, 1971, 1980-82; Н.Н. Цветкова, 1981; Е. Печенкина, 1991.
Г.Л. Хить, 1969;4? Мурмажкая обл. В. Леманн, 1974.
Саамы 1tf Финляндия Н.В. Шлыгина, 1969.(отпечаткистоп)Татары 1& Узбекистан Г.Л. Хить, 1979крымскиеТатары
1?5& Татарстан Н.А. Прокудина, 1971;
казанские 5? А.И Макеева, 2012.Татары 3& Татарстан, Н.А. Кузнецова, 1979;мишари 3? Нижегородская обл. А.И. Макеева, 2012-13.Татары 1& Белоруссия Г.Л. Хить, 1965.литовскиеТатары
1?1& Астраханская обл. Эксп. Н.А. Томилова,
астраханские 1? 1978.Татары 3& Башкирия, Татарстан А.И. Макеева, 2013кряшеныТатары
3?1& Пермский край эксп. Р.М. Юсупова
пермские 1? 2005Удмурты, 12& Удмуртия Н.А. Прокудина, 1971;Бесермяне 12? Г.В. Рыкушина, 1984.Украинцы 8& Разные области Г.Л. Хить, 1965.
Финны8?
19&УкраиныРазные провинции В. Леманн, 1974;
4? Финляндии Н.В. Шлыгина, 1967-68,
Финны 2^ Финляндия1978, 1982.В. Леманн, 1974.
(отп. стоп) Шведы 3^ Финляндия Н.В. Шлыгина, 1968.
365

Этническаягруппа
n,пол Регион Автор, время сборов
Чуваши 1& Чувашия Н.А. Прокудина, 1971.1?
Эстонцы 2& Эстония Г.Л. Хить, 1965.2?
Кавказ
Абазины 1& Карачаево-Черкессия Г.Л. Хить, 1984.шкарауа 1?Абазины 2& Карачаево-Черкессия И.П. Ладария, 1969;тапанта 2? Г.Л. Хить, 1984.Абхазы 3& Абхазия И.П. Ладария, 1969.
3?Аварцы 2& Дагестан, Г.Л. Хить, 1966-68.
2? АзербайджанАджарцы 2& Аджария Г.Л. Хить, 1968;
2? И.П. Ладария, 1969.Адыгейцы 4& Адыгея Г.Л. Хить, 1986.чемгуи 4?Адыгейцы 3& Адыгея Г.Л. Хить, 1986.бжедуги 3?Адыгейцы 4& Краснодарский край Г.Л. Хить, 1986.шапсуги 4?Адыгейцы 2& Адыгея И.П. Ладария 1969;абадзехи 2? Г.Л. Хить, 1986.Азербай- 4& Азербайджан, Г.Л. Хить, 1967;джанцы 4? Туркмения О.Б. Бабаков, 1969.Андийцы 1& Дагестан Г.Л. Хить, 1968.
1?Агулы 1& Дагестан Г.Л. Хить, 1967
1?Армяне 2& Армения Г.Л. Хить, 1968
2?Ассирийцы 1& Армения Г.Л. Хить, 1968.
1?Балкарцы 4& Кабардино-Балкария Г.Л. Хить, 1967, 1983;
4? Ф. Чотчаева, 2001.Греки- 3& Узбекистан, Украина Г.Л. Хить, 1979; эксп.эллины 3? Ю.В. Ивановой, 1988Греки- 2& Грузия, Украина Г.Л. Хить, 1979; эксп.урумы 2? Ю.В. Ивановой, 1988Грузины- 1& Грузия Г.Л. Хить, 1972.гурийцы 1?
366

Этническаягруппа
n,пол
Регион Автор, время сборов
Грузины- 2& Грузия Г.Л. Хить, 1969;имеретины 2? И.П. Ладария, 1970.Грузины- 1& Грузия Г.Л. Хить, 1968.карталинцыГрузины-
1?1& Грузия Г.Л. Хить, 1968.
кахетинцыГрузины-
1?1& Грузия И.П. Ладария, 1968.
мегрелыГрузины-
1?1& Грузия Г.Л. Хить, 1968.
мохевцыГрузины-
1?1& Абхазия Г.Л. Хить, 1983.
рачинцыГрузины-
1?1& Грузия Г.Л. Хить, 1968.
пшавыГрузины-
1?1& Грузия И.П. Ладария, 1969.
сваныГрузины-
1?1& Грузия Г.Л. Хить, 1968.
хевсурыДаргинцы
1?1&1?2&
Дагестан Г.Л. Хить, 1968.
Евреи Дагестан, Г.Л. Хить, 1968, 1983.горские 2? Кабардино-БалкарияИнгуши 1&
1?7&
Ингушетия Г.Л. Хить, 1967.
Кабардинцы Кабардино-Балкария, Г.Л. Хить, 1967, 1983,7? Адыгея 1986; Ф. Чотчаева, 2001.
Карачаевцы 2& Карачаево-Черкессия Г.Л. Хить, 1967, 1984.
Кумыки2?1& Дагестан Г.Л. Хить, 1967
Курды-1?1& Армения Г.Л. Хить, 1968.
езидыКурды-
1?1& Туркмения О.Б. Бабаков, 1969.
мусульманеЛакцы
1?1& Дагестан Г.Л. Хить, 1967
Лезгины1?2& Дагестан Г.Л. Хить, 1967
Ногайцы2?1& Дагестан Г.Л. Хить, 1967
(караногаи)Ногайцы
1?2& Карачаево-Черкессия Г.Л. Хить, 1984;
(акногаи) 2? Н.А. Суворова, 2001.
367

Этническаягруппа
n,пол
Регион Автор, время сборов
Осетины 3&3?
Сев. Осетия Г.Л. Хить, 1967.
Рутульцы 1&1?1&1?1&1?1&1?
Дагестан Г.Л. Хить, 1967
Табасаранцы Дагестан Г.Л. Хить, 1967
Талыши Азербайджан Г.Л. Хить, 1968
Таты(горскиеевреи)
Дагестан Г.Л. Хить, 1967
Турки- 2& Узбекистан Г.Л. Хить, 1979;месхетинцы 2? Г.К. Ходжайова, 1987.
Удины 1&1?
Азербайджан Г.Л. Хить, 1968.
Цахуры 1&1?
Дагестан Г.Л. Хить, 1968.
Черкесы 3&3?
Карачаево-Черкессия И.П. Ладария, 1968; Г.Л. Хить, 1984, 1986.
Чеченцы 1&1?
Чечня Г.Л. Хить, 1967.
Средняя Азия и Казахстан
Арабы 2& Узбекистан Г.Л. Хить, 1975, 1979.
Баджуйцы2?1& Таджикистан Г.Л. Хить, 1963.
Белуджи1?1& Туркмения Г.Л. Хить, 1975.
Ванчцы1?1& Таджикистан Г.Л. Хить, 1963.
Дунгане1?2& Киргизия Г.Л. Хить, 1970.
Евреи2?1& Узбекистан Г.Л. Хить, 1975
бухарскиеКазахи
1?5& Казахстан, Л. Намсарайнайдан,3? Туркмения, 1970; Г.Л. Хить, 1970;
Каракалпаки 6&
Узбекистан, Омская обл., Монголия
Узбекистан,
Н. Рысназаров, 1967; О.Б. Бабаков, 1971; эксп. Н.А. Томилова, 1976.Н. Рысназаров, 1968;
5? Туркмения О.Б. Бабаков, 1969.
368

Этническаягруппа
n,пол
Регион Автор, время сборов
Киргизы
55
40 , Киргизия Г.Л. Хить, 1963, 1970;
В.М. Кондик, 1969-70; Н.А. Прокудина, 1969; Н.А. Дубова, 1975.
Курды- 2& Туркмения, О.Б. Бабаков, 1969;мусульмане 1? Узбекистан Н.А. Дубова, 1972;
Г.Л. Хить, 1975.Персы 2& Туркмения, Г.Л. Хить, 1975, 1979(ирони) 2? УзбекистанТаджики 27^, Таджикистан, Г.Л. Хить, 1963, 1970,
27? Узбекистан 1979; О.Б. Бабаков, 1969, 1976;Е.Ф. Губенко, 1970; Н.А. Дубова, 1972-75.
Туркмены 30^, Туркмения, Г.Л. Хить, 1963;30? Таджикистан,
Узбекистан, Астраханская обл.
Н.А. Дубова, 1975, 1987; Н.А. Никольская, 1976.
Узбеки 19& Узбекистан, Г.Л. Хить, 1963, 1970,18? Туркмения, Киргизия 1979; О.Б. Бабаков,
1976; Е.Ф. Губенко, 1970; Н.А. Дубова, 1972; Н. Рысназаров, 1968.
Уйгуры 1&1?
Узбекистан Г.Л. Хить, 1970.
Шугнанцы 5&5?
Таджикистан Н.А. Дубова,Г.В. Рыкушина, 1978
Ягнобцы 1&1?
Таджикистан Г.Л. Хить, 1970.
Язгулемцы 1&1?
Таджикистан Г.Л. Хить, 1973.
Сибирь и Дальний Восток
Алтай-кижи 2& Г орный Алтай Г.Л. Хить, 1969;2? А.Р. Ким, 1977.
Бельтыры 1& Хакассия Г.Л. Хить, 1972.1?
Буряты 4& Бурятия, Монголия Г.Л. Хить, 1971;3? Л. Намсарайнайдан,1968
Долганы 1& Таймыротий н.о. В.Н. Лебедев, 1972.1?
369

Этническаягруппа
n,пол
Регион Автор, время сборов
Качинцы 2&2?
Хакассия Г.Л. Хить, 1972.
Кеты 1&1?
Красноярский край эксп. И.И. Гохмана, 1965-66.
Корейцы 2&2?
Узбекистан; Сахалинская обл.
Г.Л. Хить, 1970, 1974.
Кумандинцы 1&1?
Г орный Алтай Г.Л. Хить, 1969.
Кызыльцы 2& Хакассия; эксп. Н.А. Томилова,2? Красноярский кр. 1978; В.А. Дремов,
1980.Манси 3&
3?Тюменская обл. М.С. Акимова, 1966;
Г.М. Давыдова, 1968.Монголы 2^ Монголия Л. Намсарайнайдан,
1967-68.Нанайцы 1&
1?Хабаровский край Г.Л. Хить, 1973.
Ненцы 2&2?
Красноярский кр. Г.Л. Хить, 1962; И.П. Ладария, 1973.
Нивхи 1&3?
Хабаровский кр., Сахалинская обл.
Г.Л. Хить, 1973-74
Орочи 1&1?
Хабаровский кр. Г.Л. Хить, 1973.
Сагайцы 1&1?
Хакассия Г.Л. Хить, 1972
Селькупы 1&1?
Красноярский край эксп. И.И. Гохмана, 1965-66
Татары 16& Тюменская, Омская, эксп. Н.А. Томилова,сибирские 16? Новосибирская,
Томская, Кемеровская обл.
1976-78.
Теленгеты 1&1?1&1?1&1?1&1?1&1?
Г орный Алтай Г.Л. Хить, 1969
Телеуты Кемеровская обл. А.Р. Ким, 1978
Тубалары Г орный Алтай Г.Л. Хить, 1969
Удэгейцы Хабаровский край Г.Л. Хить, 1973
Ульчи Хабаровский край Г.Л. Хить, 1973
370

Этническаягруппа
n,пол
Регион Автор, время сборов
Хакасы 13, Хакассия Г.Л. Хить, 1972южные 1?Ханты 13, Тюменская обл. эксп. М.С. Акимовой,
1? 1960-еЧелканцы 13,
1 ОГ орный Алтай В.И. Селезнева, 1970.
Чукчи1?13,1 О
Чукотский АО М.Г. Левин, 1958
Чулымцы1?13, Томская обл. Г.А. Аксянова,1? В.А. Дремов, 1980.
Шорцы 2 3 , Хакассия; Г.Л. Хить, 1972; эксп.2? Кемеровская обл. Н.А. Томилова, 1976.
Эвенки 33 , Хабаровский край Г.Л. Хить, 1973-74;3? В.Н. Лебедев, 1974.
Эвены 13, Хабаровский край Г.Л. Хить, 1974.1?
Энцы 13, Красноярский край Г.Л. Хить, 1962.1?
Якуты 6 3 , Якутия Г.Л. Хить, 1964;6? Л.Ф. Томтосова, 1970-е;
эксп.ИЭ АНСССР, 1966.Индия
Брахманы 13 Тамил Наду С. Мэтью, 1983.айянгарыБрахманы 13 Махараштра О. Исмагулов, 1974-75.дешастхаригведиБрахманы 13 Махараштра О. Исмагулов, 1974-75читпаваныБхилы 13 Махараштра О. Исмагулов, 1974-75Бхилы паора 13 Махараштра О. Исмагулов, 1974-75Каткари 13 Бомбей О. Исмагулов, 1974-75Лингаяты 13 Майсор С. Мэтью, 1983.Маратхи 13 Махараштра О. Исмагулов, 1974-75Махары 13 Махараштра О. Исмагулов, 1974-75Парсы 13 Бомбей О. Исмагулов, 1974-75Санталы 13 Зап. Бенгалия С. Мэтью, 1983.Урали 13 Керала О. Исмагулов, 1974-75Чандрасенья 13 Махараштра О. Исмагулов, 1974-75каястхапрабху
371

Этническаягруппа
n,пол
Регион Автор, время сборов
Передняя Азия, Африка
Арабы 5 3 Сев.и Ю. Йемен В.С. Шинкаренко, 198386
Арабы 13 о-ва Абд-эль-Кури В.В. Наумкин, 1985Сокотрийцы 7 3 ,
1?о. Сокотра В.С. Шинкаренко, 1983
84Негры 13 о. Сокотра В.С. Шинкаренко, 1983
84Малийцы(банмана,манинке,сонинке,фульбе)
44 , Мали Б. Кейта, 1972-75
Суммарно 4813, 413 ? (всего 894 серии)
372

СЕРИИ МАЭ РАН
Этническаягруппа
n,пол
Регион Автор и время сборов
Тувинцы
44
40 , Тува В.И. Селезнева, 1972,
1973, 1976Арабы 183 Йемен В.И. Селезнева,
Советско-Йеменская экс., 1988, 1989, 1991
Арабы 13 Узбекистан В.И. Селезнева, 2005Узбеки 13 Узбекистан В.И. Селезнева, 2005Карелы 123, Карелия, Тверская эсп. Н.Н. Цветковой,
12? обл. 1983-89,И.Г. Широбоков, 2008
Русские 53 , Новгородская Н.Н. Цветкова, 1982,5? обл., Карелия,
Архангельск. обл.И.Г. Широбоков, 2008-11
Негры Кубы 13,1?13,1?
Куба С. Перес, 1980-е гг.
ЕвропеоидыКубы
Куба С. Перес, 1980-е гг.
Метисы Кубы 13,1?
Куба С. Перес, 1980-е гг.
373

Список сокращений
ИЭА АН СССР - Институт этнографии и антропологии Академии наук СССРИЭА РАН - Институт этнологии и антропологии Российской академии наукМАЭ РАН - Музей антропологии и этнографии им. Петра Великого (Кунсткамера) Российской Академии наукМГУ - Московский государственный университет им. М.В. Ломоносова
374

%

Научное издание
Генриэтта Леонидовна Хить Иван Григорьевич Широбоков
Ирина Анатольевна Славолюбова
Дерматоглифика в антропологии
Ответственный редактор И.Г. Широбоков Корректор И.Г. Широбоков
Оригинал-макет И.Г. Широбоков Дизайн обложки И.Г. Широбоков
Подписано в печать 18.12.2013. Формат 60 х 88 Vi6 Бумага офсетная. Печать офсетная
Усл.-печ. л. 23. Тираж 200 экз. Заказ № 3660
Издательство «Нестор-История»197110 СПб., Петрозаводская ул., д. 7
Тел. (812)235-15-86 e-mail: [email protected]; www.nestorbook.ru
Отпечатано с готового оригинал-макета в типографии «Нестор-История»
198095 СПб., ул. Розенштейна, д. 21 Тел. (812)622-01-23

ГЕНРИЭТТА ЛЕОНИДОВНА ХИТЬ, кандидат биологических наук, доктор исторических наук, главный научный сотрудник ИЭА РАН. Член Европейской ассоциации антропологов (ЕАА). Автор более 150 работ по возрастной морфологии, серологии и дерматоглифике, в том числе монографий «Дерматоглифика народов СССР» и «Расовая дифференциациячеловечества: дерматоглифические данные»(последняя в соавторстве с Н.А. Долиновой). Специалист в области популяционной, этнической и расовой дерматоглифики.
ИВАН ГРИГОРЬЕВИЧ Ш ИРОБОКОВ,кандидат исторических наук, научный сотрудник МАЭ РАН (Кунсткамера). Член Санкт-Петербургского союза ученых (СПбСУ). Автор 35 публикаций по этнической краниологии, дерматоглифике, изучению кремированных скелетных останков.
ГРИГОРЬЕВИЧ
ИРИНА АНАТОЛЬЕВНА СЛАВОЛЮБОВА,кандидат биологических наук, доцент кафедры антропологии биологического факультета Московского государственного университета им. М.В. Ломоносова (МГУ). Член Европейской ассоциации антропологов (ЕАА). Автор более 20 работ по конституциональной, медицинской антропологии и дерматоглифике. Специалист в области морфологии человека, медицинской антропологии и дерматоглифики. На протяжении ряда лет ведет практические и лекционные занятия по дерматоглифике для студентов кафедры антропологиибиологического факультета МГУ.

![С. М. Михеев. [Рец. на:] Святые князья-мученики Борис и Глеб / Н. И. Милютенко (СПб., 2006) (Библиотека христианской](https://static.fdokumen.com/doc/165x107/63140c13fc260b71020f601d/s-m-mikheev-rets-na-svyatie-knyazya-mucheniki.jpg)


















