
Depletion of anaphase-promoting complex or cyclosome (APC/C) subunit homolog APC1 or CDC27 of...
22
1 DEPLETION OF APC/C SUBUNIT HOMOLOG APC1 OR CDC27 OF Trypanosoma brucei ARRESTS THE PROCYCLIC FORM IN METAPHASE BUT THE BLOODSTREAM FORM IN ANAPHASE Praveen Kumar and C. C. Wang From The Department of Pharmaceutical Chemistry, UCSF, San Francisco, CA, USA 94143-2280 Running Title: Role of APC/C subunits in cell cycle regulation of T. brucei. Address correspondence to: C.C. Wang, Department of Pharmaceutical Chemistry, UCSF, San Francisco, CA, USA 94143-2280, Tel. 415-476-1321; Fax. 415-476-3382; E-Mail: [email protected] Abstract The anaphase-promoting complex or cyclosome (APC/C) is a multi-protein subunit E3 ubiquitin ligase complex that controls segregation of chromosomes and exit from mitosis in eukaryotes. It triggers elimination of key cell cycle regulators such as securin and mitotic cyclins during mitosis by polyubiquitinating them for proteasome degradation. Seven core subunit homologs of APC/C (Apc1, Apc2, Apc11, Cdc16, Cdc23, Cdc27 and Doc1) were identified in Trypanosoma brucei genome database. Expression of six of them was individually ablated by RNA interference (RNAi) in both the procyclic and bloodstream forms of T. brucei. Only the Cdc27 and Apc1 depleted cells were enriched in the G2/M phase with inhibited growth. Further studies indicated that T. brucei Apc1 and Cdc27 failed to complement the corresponding deletion mutants of budding yeast. But their depletion from procyclic-form T. brucei enriched cells with two kinetoplasts and an enlarged nucleus possessing short metaphase like mitotic spindles, suggesting that Apc1 and Cdc27 may play essential roles in promoting anaphase in the procyclic form. Their depletion from the bloodstream form, however, enriched cells with two kinetoplasts and two nuclei connected through a microtubule bundle suggesting a late anaphase arrest. This is the first time functional APC/C subunit homologs were identified in T. brucei. The apparent differential activities of this putative APC/C in two distinct developmental stages suggest an unusual function. The apparent lack of functional involvement of some of the other individual structural subunit homologs of APC/C may indicate the structural uniqueness of T. brucei APC/C. Trypanosoma brucei is an ancient protist that causes African sleeping sickness. It has a complex life cycle that alternates between the mammalian host and insect vector. It undergoes a series of morphological, metabolic and biochemical changes to adapt to the distinct living environments. The dividing procyclic form in insects and bloodstream form in mammals have sequential G1, S, G2, and M phases (1). There is a single mitochondrion in each cell that contains a DNA complex, the kinetoplast, and divides synchronously with the nucleus (2). The identification and characterization of proteins that regulate T. brucei cell cycle progression have been the main focus of our recent studies. Among the limited number of cyclin homologs and cdc2-related protein kinases (CRKs) identified in the Trypanosome Genome Database, we found CycB2/CYC6 to be the only cyclin and CRK3 the sole protein kinase involved in regulating the G2/M transition in both procyclic and bloodstream forms of T. brucei (3,4). This unusually simple scheme of G2/M passage regulation prompted us to look further into it with the expectation of establishing a simple model for in-depth understanding of the mechanisms of G2/M checkpoint control. The detailed features of mitosis in T. brucei have not yet been well delineated due to technical difficulties in synchronizing the cell growth. Consequently, important discrete stages occurring during mitosis such as the prophase, metaphase, anaphase, telophase, etc., have not yet been positively identified in this organism. It is known that the function of an E3 ubiquitin ligase, the anaphase-promoting complex or cyclosome (APC/C), is closely associated with mitotic exit in eukaryotes. It is a multi-subunit complex that promotes degradation of proteins by forming a tripartite complex along with the target protein and the ubiquitin-conjugating enzyme E2 (5). It polyubiquitinates important cell cycle regulatory JBC Papers in Press. Published on July 1, 2005 as Manuscript M504326200 Copyright 2005 by The American Society for Biochemistry and Molecular Biology, Inc. by guest, on January 24, 2013 www.jbc.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Depletion of anaphase-promoting complex or cyclosome (APC/C) subunit homolog APC1 or CDC27 of...

1
DEPLETION OF APC/C SUBUNIT HOMOLOG APC1 OR CDC27 OF Trypanosoma bruceiARRESTS THE PROCYCLIC FORM IN METAPHASE BUT THE BLOODSTREAM FORM IN
ANAPHASEPraveen Kumar and C. C. Wang
From The Department of Pharmaceutical Chemistry, UCSF, San Francisco, CA, USA 94143-2280Running Title: Role of APC/C subunits in cell cycle regulation of T. brucei.
Address correspondence to: C.C. Wang, Department of Pharmaceutical Chemistry, UCSF, San Francisco,CA, USA 94143-2280, Tel. 415-476-1321; Fax. 415-476-3382; E-Mail: [email protected]
AbstractThe anaphase-promoting complex or cyclosome(APC/C) is a multi-protein subunit E3 ubiquitinligase complex that controls segregation ofchromosomes and exit from mitosis ineukaryotes. It triggers elimination of key cellcycle regulators such as securin and mitoticcyclins during mitosis by polyubiquitinatingthem for proteasome degradation. Seven coresubunit homologs of APC/C (Apc1, Apc2,Apc11, Cdc16, Cdc23, Cdc27 and Doc1) wereidentified in Trypanosoma brucei genomedatabase. Expression of six of them wasindividually ablated by RNA interference(RNAi) in both the procyclic and bloodstreamforms of T. brucei. Only the Cdc27 and Apc1depleted cells were enriched in the G2/M phasewith inhibited growth. Further studiesindicated that T. brucei Apc1 and Cdc27 failedto complement the corresponding deletionmutants of budding yeast. But their depletionfrom procyclic-form T. brucei enriched cellswith two kinetoplasts and an enlarged nucleuspossessing short metaphase like mitoticspindles, suggesting that Apc1 and Cdc27 mayplay essential roles in promoting anaphase inthe procyclic form. Their depletion from thebloodstream form, however, enriched cells withtwo kinetoplasts and two nuclei connectedthrough a microtubule bundle suggesting a lateanaphase arrest. This is the first timefunctional APC/C subunit homologs wereidentified in T. brucei. The apparentdifferential activities of this putative APC/C intwo distinct developmental stages suggest anunusual function. The apparent lack offunctional involvement of some of the otherindividual structural subunit homologs ofAPC/C may indicate the structural uniquenessof T. brucei APC/C.
Trypanosoma brucei is an ancient protistthat causes African sleeping sickness. It has acomplex life cycle that alternates between themammalian host and insect vector. It undergoes aseries of morphological, metabolic andbiochemical changes to adapt to the distinct livingenvironments. The dividing procyclic form ininsects and bloodstream form in mammals havesequential G1, S, G2, and M phases (1). There is asingle mitochondrion in each cell that contains aDNA complex, the kinetoplast, and dividessynchronously with the nucleus (2).
The identification and characterization ofproteins that regulate T. brucei cell cycleprogression have been the main focus of our recentstudies. Among the limited number of cyclinhomologs and cdc2-related protein kinases(CRKs) identified in the Trypanosome GenomeDatabase, we found CycB2/CYC6 to be the onlycyclin and CRK3 the sole protein kinase involvedin regulating the G2/M transition in both procyclicand bloodstream forms of T. brucei (3,4). Thisunusually simple scheme of G2/M passageregulation prompted us to look further into it withthe expectation of establishing a simple model forin-depth understanding of the mechanisms ofG2/M checkpoint control. The detailed features of mitosis in T. bruceihave not yet been well delineated due to technicaldifficulties in synchronizing the cell growth.Consequently, important discrete stages occurringduring mitosis such as the prophase, metaphase,anaphase, telophase, etc., have not yet beenpositively identified in this organism. It is knownthat the function of an E3 ubiquitin ligase, theanaphase-promoting complex or cyclosome(APC/C), is closely associated with mitotic exit ineukaryotes. It is a multi-subunit complex thatpromotes degradation of proteins by forming atripartite complex along with the target protein andthe ubiquitin-conjugating enzyme E2 (5). Itpolyubiquitinates important cell cycle regulatory
JBC Papers in Press. Published on July 1, 2005 as Manuscript M504326200
Copyright 2005 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest, on January 24, 2013w
ww
.jbc.orgD
ownloaded from

2
proteins such as cyclin B and Pds1/securin, targetsthem for proteolysis by the 26S proteasome andpromotes chromosome segregation, transitionfrom metaphase to anaphase and exit from mitosis(6,7). APC/C has at-least two very important rolesduring metaphase to anaphase transition in ametazoan. It activates the cysteine-proteaseseparase by ubiquitinating its inhibitorPds1/securin. The activated separase then destroysthe cohesion between sister chromatins andenables the mitotic spindle to pull them to oppositespindle poles (8,9). Secondly, APC/C ubiquitinatesmitotic cyclins and thereby inactivates cyclin-dependent kinase 1 (Cdk1, designated Cdc28 inbudding yeast), which is required for exit frommitosis and initiation of cytokinesis. Disruption ofAPC/C functions by knocking down expression ofits subunits leads to cell cycle arrest at themetaphase to anaphase transition (5). The failurein chromosome segregation arrests the cells at theG2/M boundary with short metaphase like mitoticspindles. There are 13 core subunits of APC/C inSaccharomyces cerevisiae, (6,7,10), which form acore of the particle and remain tightly associatedthroughout the cell cycle. There are also twoAPC/C activator proteins Cdc20 and Cdh1 and anE2 protein Ubc4 that associate with APC/C atspecific points during the cell cycle forpolyubiquitination of specific proteins destined forproteasome degradation (11). Cdc20 activatesAPC/C specifically for promoting anaphase,whereas Cdh1 activates the complex after themitotic exit (11). In the current study, weidentified an APC/C equivalent in T. brucei, and 7core subunit homologs in the genome database.The expression of six of them was ablated fromboth procyclic and bloodstream forms of cells withRNA interference (RNAi) and the cells wereexamined for mitotic arrest and accumulation inthe metaphase. The results indicated that only theloss of Apc1 or Cdc27 homolog led to mitoticarrest. The procyclic form was apparently blockedin the metaphase, whereas the bloodstream formwas more likely stopped in the late anaphase.Thus, the APC/C machinery appears to performdistinctive functions at different stages ofdevelopment in T. brucei.
MATERIALS AND METHODS
Cell culture—The procyclic-form T. brucei strain29-13 (12) was cultured in Cunningham’s mediumat 26 °C supplemented with 10% fetal bovineserum (Atlanta Biological). G418 (15 µg/ml) andhygromycin B (50 µg/ml) were added to theculture medium to maintain the T7 RNApolymerase and tetracycline repressor geneconstructs within the cells.
The bloodstream form T. brucei strain 90-13 (12) was cultivated at 37 °C in the HMI9medium supplemented with 10% fetal bovineserum and 10% serum plus (JRH Biosciences)(13). G418 (2.5 µg/ml) and hygromycin B (5µg/ml) were also added to the culture medium tomaintain the T7 RNA polymerase and tetracyclinerepressor constructs.
RNAi interference—RNAi experiments weredesigned for six of the APC/C core subunithomologs (see results). The sequences used forRNAi were as follows; nucleotide numbers 1723-2174 for Apc1, 623-961 for Apc2, 1-246 forApc11, 562-933 for Cdc16, 832-1311 for Cdc23and 906-1353 for Cdc27. The corresponding 250-500 bp DNA fragments were amplified by PCRusing a pair of primers with added XhoI andHindIII linkers to the ends and sub-cloned into thetetracycline-inducible pZJM vector by replacingthe a-tubulin fragment in it (14). The resultingRNAi constructs, with the inserts between twoopposing T7 promoters and two opposingtetracycline operators, were linearized with NotIand introduced into T. brucei cells byelectroporation for integration into the rDNAspacer region in the chromosome. Transfection ofthe procyclic form T. brucei was performedessentially according to a procedure describedpreviously (15,16). After phleomycin selection,single transfected cells were cloned on semi solidagarose plates, cultivated under phleomycin andinduced by tetracycline to synthesize the doublestranded RNA (dsRNA) encoded by the cDNAinsert. The dsRNA degrades the correspondingmRNA in T. brucei, and thus blocks theexpression of the encoding gene (3).
The bloodstream-form T. brucei wastransfected by a previously described procedurewith some modifications (17). Briefly, a sample of1x107 log-phased cells were harvested, washedonce and suspended in 0.5 ml of cytomix buffer(18) containing 100 µg of the linearized pZJM
by guest, on January 24, 2013w
ww
.jbc.orgD
ownloaded from

3
DNA construct. Electroporation was carried out ina 4-mm cuvette using the Gene Pulser (Bio-Rad)with parameters set as follows: 1.7-kV voltage,400-ohm resistance, and 25-microfaradcapacitance. The cells were transferred to a 24-well plate in HMI9 medium immediately afterelectroporation and incubated at 37°C for 24 h.The transfectants were then selected with theaddition of 2.5µg/ml phleomycin, and single cellwas cloned by limiting dilution on soft agaroseplates. For induction of RNAi, the clonedtransfectants were cultured in the presence of 1.0µg/ml tetracycline. Cell numbers in time sampleswere counted with a microscope using ahemocytometer.
Semi-quantitative RT-PCR— To monitor theeffect of tetracycline-induced RNAi on geneexpression, semi-quantitative RT-PCR wasconducted by a previously described procedure oncellular RNA from cells 3 days after tetracyclineinduction (3). Total RNA was extracted from T.brucei cells using the TRIzol reagent (AmershamBiosciences). First-strand cDNA was generatedfrom DNaseI-treated RNA samples using anoligo(dT)15 primer and Moloney murine leukemiavirus-reverse transcriptase (Promega). PCR wasthen performed using the first-strand cDNA and apair of gene-specific primers that differs from theprimer pair used in generating the original RNAiconstruct (sequences available upon request).
Fluorescence-activated cell sorting (FACS)analysis—Cell samples for FACS analysis wereprepared as described previously (18) and FACSanalysis was conducted by a reported procedure(Tu and Wang, 2004). The propidium iodide (PI)-stained cell samples from the analysis were alsoexamined with a Nikon phase-contrast andfluorescence microscope for a tabulation ofnumbers of nuclei and kinetoplasts in individualcells and a count of cells with differentmorphologies from a population of 200 cells.
Complementation of Saccharomyces cerevisiaedeletion mutants — Full-length genes encodingyeast Cdc27 (yCdc27), yeast Apc1 (yApc1) and T.brucei Cdc27 (tCdc27), and T. brucei Apc1(tApc1) were each amplified by PCR and sub-cloned into the vector pGADT7 (Invitrogen). Thisvector was modified by adding an additional NdeI
site at position #1480 and removing fragment#1480-1964 by Nde I digestion. Constructscontaining yCdc27, tCdc27 were each introducedinto the yeast Tet-promoter Hughes (yTHC) strainYSC1180-7428818 (obtained from OpenBiosystem of Huntsville, AL) (19) cells grown inYPD medium with 10 mg/ml doxycycline andincubated for 3 days at 37 °C. yApc1 and tApc1were each transfected into a MATa apc1-1 yeaststrain kindly provided by Dr. W. Zachariae of theMax Planck Institute, Germany (20). Cells weregrown in YPD medium at 37 °C for 3 days.
Labeling of mitotic spindles with KMX-1—Mitoticspindles in T. brucei were labeled with mousemonoclonal antibody KMX-1 (a kind gift from Dr.Keith Gull, Oxford University), (21), whichpreferentially recognizes b-tubulin in the mitoticspindle. Immunofluorescence protocol was exactlyfollowed as described earlier (22). Slides weremounted in Vectashield in the presence of 1 µg of4,6-diamino-2-phenylindole (DAPI) per ml andexamined with a fluorescence microscope.
RESULTS
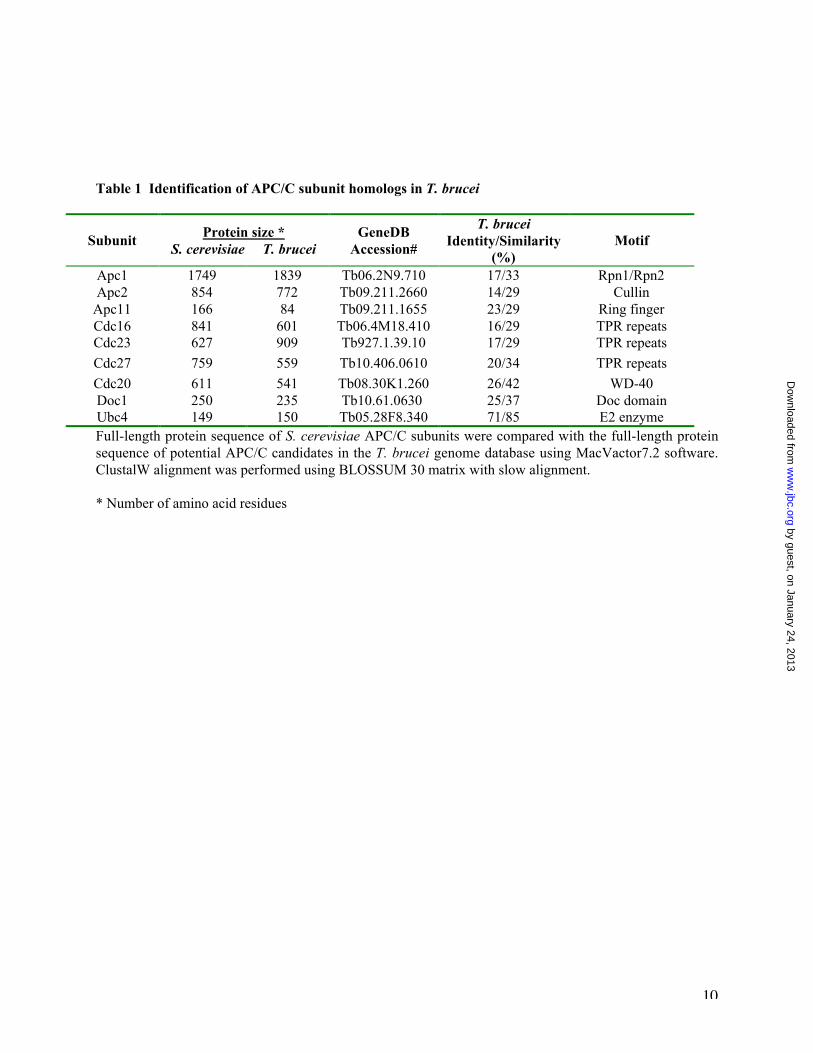
Identification of homologs of APC/C subunits inthe genome database of T. brucei—The 10 coresubunits of APC/C in S. cerevisiae include Apc1,Apc2, Apc4, Apc5, Apc9, Cdc16, Cdc23, Cdc26,Cdc27 and Doc1 (6,7,10). They and the twoAPC/C activator proteins Cdc20 and Cdh1 plusthe E2 enzyme Ubc4 were each used as a query toscreen the Sanger Trypanosome GenomicDatabase, which resulted in identification of 7 coresubunit homologs; Apc1, Apc2, Apc11, Cdc16,Cdc23, Cdc27 and Doc1, one activator proteinCdc20 and a Ubc4 homolog (Table 1). Thepresence of two hallmark proteins of APC/C,Apc2 and Apc 11 (20,23), the three tetratricopeptide repeat (TPR) proteins Cdc16, Cdc23,Cdc27 (24) and Doc1, a subunit promotingsubstrate binding and polyubiquitination (25),suggests a functioning, albeit simpler, APC/C in T.brucei. This is further supported by the presenceof a Cdc20 and a Ubc4 homolog, which isconsistent with APC/C functioning as ananaphase-promoting complex in the trypanosome(11).
by guest, on January 24, 2013w
ww
.jbc.orgD
ownloaded from

4
Cdc27 and Apc1 are the only subunits required forT. brucei growth—Six of the APC/C core subunithomologs in T. brucei were chosen for RNAianalysis; Apc1, Apc2, Apc11, Cdc16, Cdc23 andCdc27. We did not knock down Doc1 because it isa nonessential subunit in budding yeast and thedeletion mutant of Doc1 in yeast can grownormally (25). Apc1 is the largest subunit ofAPC/C with its motif sharing similarity with Rpn1and Rpn2 of the 19S cap of proteasome (26-30).Tetrad analysis of the S. cerevisiae Apc1-1temperature sensitive mutant showed that, at non-permissive temperature, it failed to exit frommitosis and was arrested at G2/M-phase (28).Apc2 carries a cullin domain and is associatedwith Apc11 and plays a major role in recruitingthe E2 enzyme (31-33). Apc11 is the smallestsubunit in APC/C. It carries a ring H2-fingerdomain, mediating protein-protein interactions andinteracting with the cullin domain of Apc2 (34).Tetrad analysis indicated that both Apc11 andApc2 are also essential genes in S. cerevisiae(31,35). Cdc16, Cdc23 and Cdc27 are related TPRproteins with molecular weights ranging from 70to100 kDa and 9-10 copies of TPR (24,32).Temperature sensitive mutants of these genes in S.cerevisiae blocked cell cycle at G2/M phase withlarge budded cells that failed to proceed frommetaphase to anaphase at 37°C (36). Thus, basedon this information from yeast, a knock down ofeach of the six chosen APC/C core subunithomologs in T. brucei was expected to result in agrowth arrest in the G2/M phase.
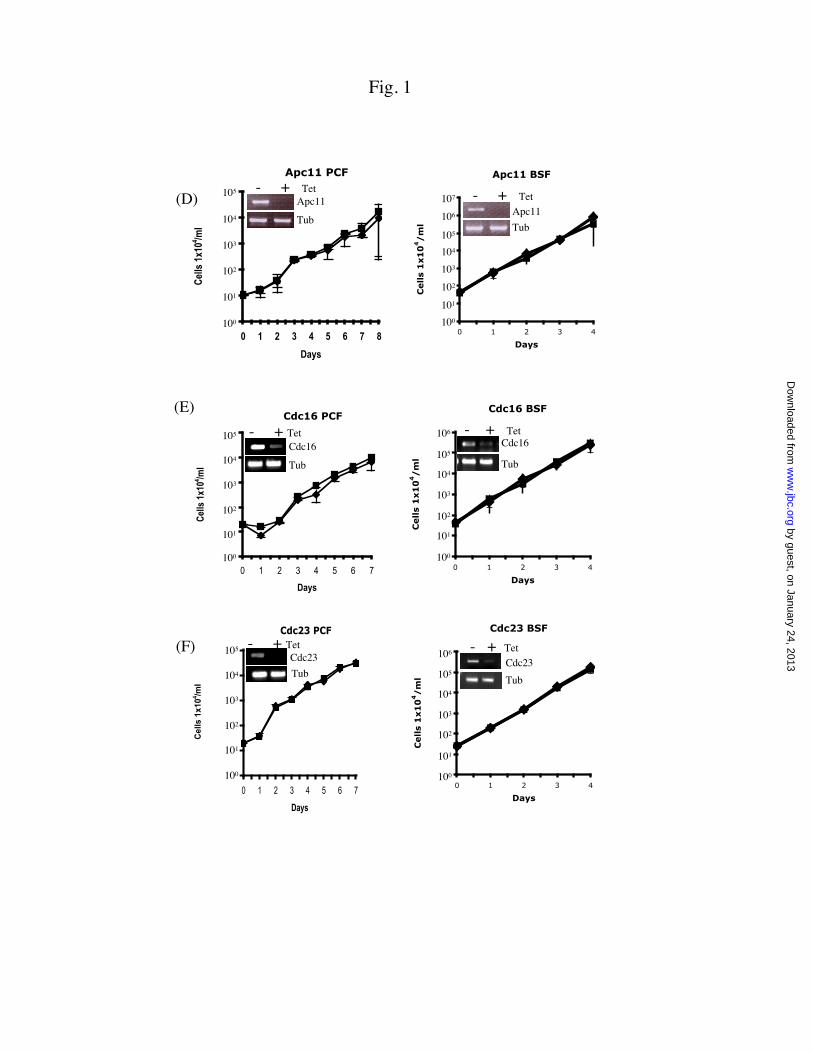
The RNAi experiments were performedon both the procyclic form and bloodstream formof T. brucei. Semi-quantitative RT-PCR wascarried out on cellular RNA samples three daysafter tetracycline induction (3). The results,presented among the insets in Figures 1A-F, showthat, in every instance, the level of mRNA wassignificantly diminished after a three-day RNAiwhen compared with that of a-tubulin mRNAincluded as a sampling control. The RNAitechnique thus successfully down-regulated theexpression of specific mRNAs in both forms of T.brucei.
The potential effect from knocking downthe expression of each of the six APC/C coresubunit genes on the proliferation of T. brucei wasexamined (Figures 1A-F). Only the knockdownsof Cdc27 (Figure 1A) and Apc1 (Figure 1B) were
followed by significant growth inhibition, whereasdecreased expression of Apc2 (Figure 1C), Apc11(Figure 1D), Cdc16 (Figure 1E) and Cdc23(Figure 1F) all failed to register any detectableeffect on the growth of both forms of T. brucei.Quantitative analysis of cell numbersdemonstrated that the growth of Cdc27-depletedprocyclic-form was completely stopped two daysinto RNAi induction, while the Cdc27-depletedbloodstream form assumed a growth rate about 1%of the un-induced control (Figure 1A). For Apc1depletion, the procyclic form grew at a 10-foldreduced rate, whereas the bloodstream form grewat approximately 1% of the control (Figure 1B).
Assuming that the cause of inhibitedgrowth was attributed to a compromised APC/Cfunction in T. brucei, Cdc27 and Apc1 may bethen playing essential roles in the complex whileApc2, Apc11, Cdc16 and Cdc23 could be eitherdispensable or not included in the complex. Athird possibility could be that their expressionsneed to be ablated together for a phenotype. Wechose to learn first whether T. brucei Cdc27 andApc1 can complement the functions of theircounterparts in S. cerevisiae and to find out if thegrowth arrest caused by Cdc27 and Apc1depletions in T. brucei is the same or similar tothat observed after APC/C depletion in yeast andmammalian cells (24,28).
T. brucei Cdc27 and Apc1 do not exhibitcomplementary functions in S. cerevisiae—T.brucei Cdc27 and Apc1 have sequence identitiesof 20% and 17% with the Cdc27 and Apc1 in S.cerevisiae, respectively (Table1). These relativelylow degrees of sequence homology cast somedoubt on whether they could complement theCdc27 and Apc1 functions in yeast. Thus, thecomplementation studies were first carried out inS. cerevisiae strain YSC1180-7428818 with Cdc27expression under doxycycline regulation.Addition of doxycycline turned off the expressionof endogenous Cdc27 gene and made the cellgrowth dependent on the ectopically introducedone. When the yeast Cdc27 gene was introduced,the cells were able to grow normally underdoxycycline (Figure 2A). However, when T.brucei Cdc27 gene was introduced into these cells,they failed to grow under the drug pressure (Figure2A), indicating that T. brucei Cdc27 cannotcomplement the function of yeast Cdc27 gene.
by guest, on January 24, 2013w
ww
.jbc.orgD
ownloaded from

5
Similar results were observed when T.brucei Apc1 gene was used to rescue the yeasttemperature sensitive Apc1-1 mutant (MATa,apc1). Apc1 of T. brucei failed to complement thefunction of yeast A p c 1 at the restrictivetemperature, whereas the exogenously introducedyeast Apc1 could (Figure 2B). Thus, neitherCdc27 nor Apc1 from T. brucei can replace theircounterparts in yeast. Though somewhatanticipated from the poor sequence alignment, thenegative results raised some concern over the realfunctions of Cdc27 and Apc1 homologs in T .brucei, which, we hope, could be answered byfurther characterizing the phenotypes of Cdc27and Apc1 RNAi knockdown mutants of T. brucei.
The Cdc27 and Apc1-depleted cells are arrested inthe G2/M phase—Time samples were collectedduring the RNAi experiments, stained with PI andanalyzed with FACScan (Figure 3). The controlcells containing an empty pZJM vector have 52-55% of the cells remaining in G1, 25-26% in Sand 20-23% in G2/M phase during the ensuing 3days of incubation. The Cdc27-depleted procyclicform had a 15 % increase in G2/M cells and acorresponding decrease in G1 cells without muchchange in S-phase cells, whereas the Cdc27-depleted bloodstream form exhibited 25%enhancement in G2/M, 35% reduction in G1 and10% increase in S-phase cells (Figure 3A, lefthand panel). For the Apc1 depleted procyclicform, there was about 10% increase in G2/M, asimilar decrease in G1 and a 5% decrease in S-phase cells. The Apc1 depleted bloodstream formhas approximately 15% increase in G2/M, 15%reduction in G1 and virtually no change in S-phasecells (Figure 3B, left hand panel). The histogramsin the right hand panels of Figures 3A and 3Bpresent the raw data from FACScan anddemonstrate an unambiguous shift from G1 toG2/M phase during the 3 day RNAi induction.This enrichment of G2/M cells in both forms of T.brucei depleted of either Cdc27 or Apc1 suggeststhat the two proteins could function as subunits ofan APC/C-like complex in T. brucei.
Distinctive morphological changes betweenthe procyclic and bloodstream forms depletedof Cdc27 and Apc1 —The Cdc27 and Apc1-depleted cells stained with PI were examined
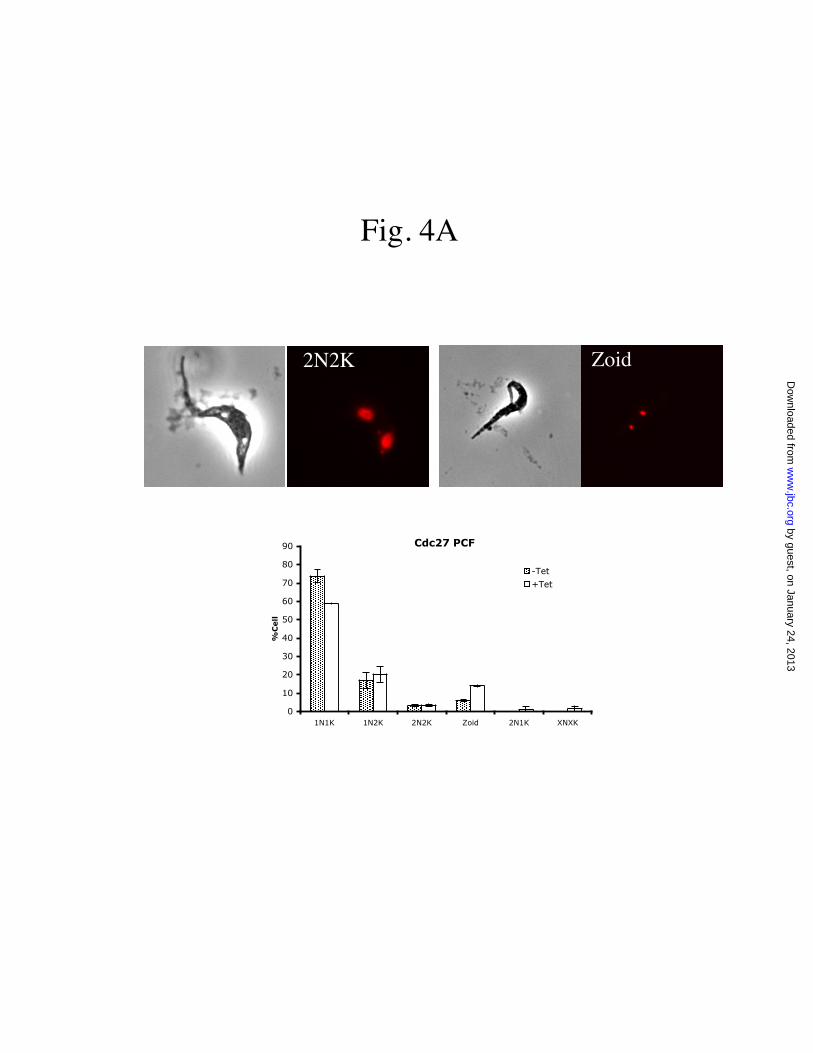
under a fluorescence microscope for aberration inintracellular numbers of nuclei and kinetoplasts(Figure 4). There was a significant drop in thecells with a single nucleus and a single kinetoplast(1N1K) reflecting a reduced population of G1cells. Cells with one nucleus and two kinetoplasts(1N2K) were slightly increased, suggesting anenrichment of G2/M cells.
In the procyclic form, the population oftwo nuclei-two kinetoplasts (2N2K) cellsremained unchanged by the knockdowns and thetwo nuclei were always well separated from eachother (Figures 4A and 4B). There was, however,an emergence of anucleate cells (0N1K, zoids) upto 10 to 15% of the population. This is anticipatedfrom our previous observation that for theprocyclic form arrested in G2/M through aknockdown of CycB2/CYC6 or CRK3, asubstantial number of the cells could still proceedinto cytokinesis and cell division resulting in theformation of zoids (3,4). The appearance of zoidsin the present study could be regarded as anadditional proof that the cells are arrested in G2/Mby depleting Cdc27 or Apc1.
The bloodstream form arrested in G2/Mby cyclin or kinase knockdown does not producezoids, but, instead, accumulates cells with onenuclear aggregate and multiple kinetoplasts (3,4).Such a population increase was, however, notobvious in the Cdc27 or Apc1-depletedbloodstream form. Instead, a sharp rise in thepopulation of 2N2K cells became apparent(Figures 3B and 3D). This increase could reach 35to 40% of the total population. At a closerexamination, however, the two nuclei in each cellwere found always closely associated with eachother with the two closely associated kinetoplastslocated at a distance toward the posterior end(Figures 4B and 4D), suggesting that they have notquite yet exited from mitosis (see below).The mitotic spindles in Cdc27 or Apc1-depletedcells differ between procyclic and bloodstreamforms—In order to resolve this mystery, westained the cells with KMX-1 monoclonalantibody known to stain preferentially b-tubulin inmitotic spindles (22). We observed in the Cdc27-and Apc1-depleted procyclic form an increase ofcells with short mitotic spindles from 10% to 19%and 17%, respectively (Figures 5A, 5B and 6).These cells were likely arrested in the metaphaseand their populations were enhanced as a
by guest, on January 24, 2013w
ww
.jbc.orgD
ownloaded from

6
consequence of Cdc27 (Figure 6A) or Apc1(Figure 6B) depletion. In Figure 5C, there is azoid in the process of being generated. Themother cell is arrested in the metaphase by thedepletion of Cdc27, but a zoid is formed in spite ofit, illustrating that cytokinesis and cell division arenot stopped by the metaphase arrest of procyclic-form T. brucei.
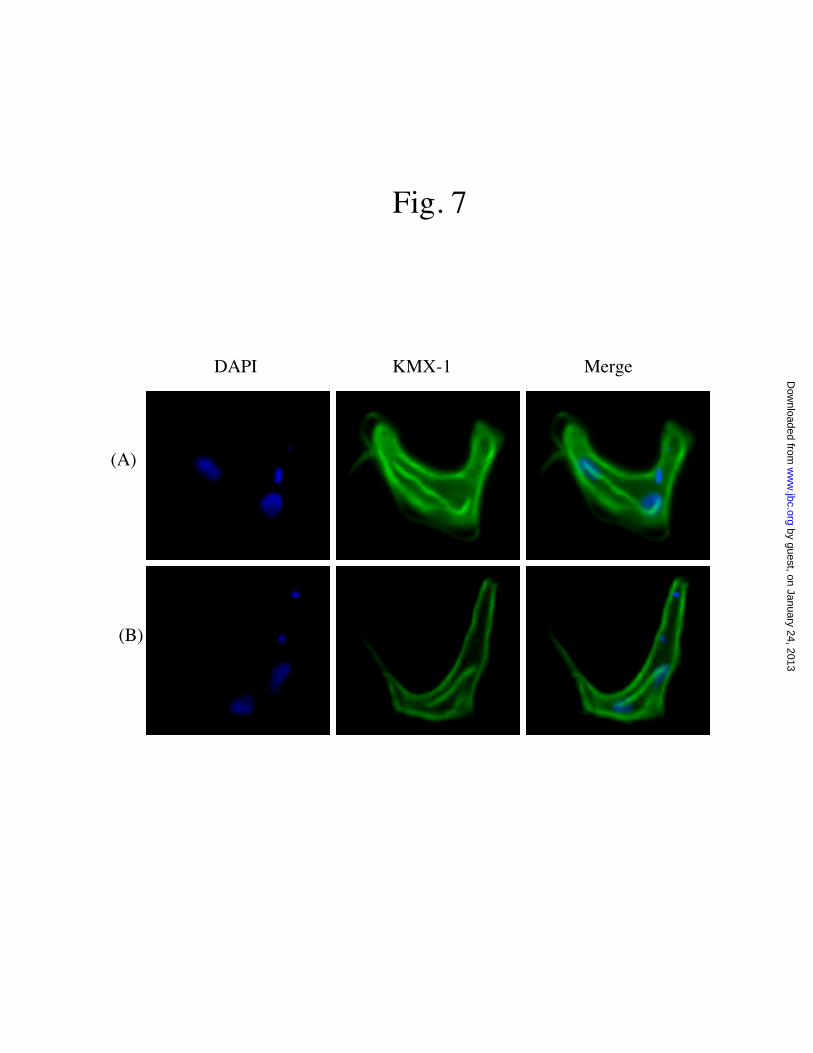
For the Cdc27 and Apc1-depletedbloodstream form, KMX-1 staining revealed thatthe two nuclei in the 2N2K cells are notcompletely separated but linked by a bundle ofmicrotubules which could be the elongatedspindles one observes in the late anaphase (Figures7A and 7B). This is illustrated in greater details inFigure 7B, in which two apparent nuclear DNAclusters are connected by a bundle ofmicrotubules, whereas the two kinetoplasts areboth located at the posterior end of the cell. Thus,deficiency of APC/C function in the bloodstreamform may have arrested the cells in late anaphaseinstead of metaphase.
DISCUSSIONIn this study, we have selectively
downregulated the expression of six core APC/Csubunit homologs by RNAi in both the procyclicand bloodstream forms of T. brucei to investigateif they play essential roles in APC/C-mediated cellcycle progression. Only depletion of the Cdc27 orApc1 homolog led to a G2/M arrest. The failure ofthese two homologs in complementing theircounterparts in yeast further suggested that T.brucei may have a structurally unique APC/C withits Cdc27 and Apc1 subunits playing essentialfunctions but incapable of replacing theircounterparts in yeast due to structural distinctions.The lack of a phenotype from depleting the otherAPC/C subunits in T. brucei could mean that someof them may have overlapping structural andfunctional roles. They may require a combinedknockdown, e.g., Apc2 with Apc11 or Cdc16 withCdc23, in order to register an effect. A search forthe remainder of the core subunits in this complex,if any, using perhaps the yeast two-hybrid assay(37) and TAP tag affinity chromatography (38),may help our future understanding of thecomposition of this protein complex. To the bestof our knowledge, there has not yet been anyAPC/C ident i f ied among eukaryot icmicroorganisms other than budding and fission
yeast. A dissection of the T. brucei APC/C couldcast some light on the structural as well asfunctional evolution of this complex.
Meanwhile, the identification of a Cdc20homolog with a 25% sequence identity and 37%similarity with the yeast Cdc20 and the apparentabsence of a Cdh1 homolog from T. brucei (seeTable 1) suggest that the APC/C in this protistcould have the sole function of promotinganaphase without involving the Cdh1-activatedfunction beyond mitotic exit (11). The presence ofa Ubc4 homolog in T. brucei (see Table 1) with71% identity and 84% similarity with yeast Ubc4(see Table 1) suggests a highly conserved E2ubiquitin-conjugating enzyme performing thefunction of a donor of ubiquitin for T. bruceiAPC/C. This well-conserved E2 suggests alsostructural conservation of APC/C in interactingwith Ubc4 (39).
Depletion of Cdc27 and Apc1 from theprocyclic form of T. brucei resulted in G2/M cellsthat possessed primarily a single nucleus with ashort metaphase like mitotic spindle. The spindleswere short, rhomboid and pointed toward the endswith DNA compressed more at the spindleconsistent with the formation of a metaphase plate.Spindle poles were sharp and pointed. Theseresults are consistent with those observed in S.cerevisiae (24,28,40) and human (Homo sapiens)Cdc27 mutants (41,42) or the Apc1 conditionalmutants (26,28) in which the cells are specificallyarrested at the metaphase to anaphase transition.
For the bloodstream form depleted ofCdc27 or Apc1, however, the G2/M cells wereenriched in an apparent 2N2K morphology. Themitotic spindles increased in size and becamestretched (Figures 7A & 7B). Two closelyassociated nuclei were visible and apparentlymoving toward the opposite poles, whereas thenewly formed kinetoplast was located at posteriorend of the cell (Figures 4B & 4D). The spindlesformed microtubule bundles at the middle of thetwo separated nuclei present on the opposite poles.This is apparently the final stage of the mitosisafter which two daughter cells should separatefrom each other to complete the mitosis. But theeventual separation has never occurred. Thearrested state could likely represent the lateanaphase.
In eukaryotic cells, expression of non-degradable cyclin B inhibits CDK inactivation,
by guest, on January 24, 2013w
ww
.jbc.orgD
ownloaded from

7
spindle disassembly and cytokinesis withoutaffecting chromosome segregation, which resultedin late anaphase-like-arrest (43,44). Thus itappears that the function of APC/C in thebloodstream form of T. brucei is required only fordegrading mitotic cyclin for mitotic exit but notfor Pds1/securin degradation for chromosomesegregation. The two forms of T. brucei havebeen shown to have distinct cell cycle regulations.Although depletion of CycB2/CYC6 or CRK3 ledto G2/M arrest in both forms of T. brucei (3,45),the procyclic form is arrested at the G2/M phase
with one somewhat enlarged nucleus, whereas thebloodstream form continues to enter G1 phase foranother round of nuclear cycle resulting in amultinuclear aggregate. Thus, a lowered mitoticCDK activity through knocking down eitherCycB2/CYC6 or CRK3 is relatively ineffective inplacing the bloodstream form under mitotic arrest,whereas an apparent over-expression of mitoticCDK through APC/C knockdown stops it in thelate anaphase. These differences in regulation ofmitosis in bloodstream-form and procyclic form ofT. brucei will be the subject of future experiments.
REFERENCES
1. Woodward, R., and Gull, K. (1990) J Cell Sci 95 ( Pt 1), 49-572. Ploubidou, A., Robinson, D. R., Docherty, R. C., Ogbadoyi, E. O., and Gull, K. (1999) J
Cell Sci 112 ( Pt 24), 4641-46503. Tu, X., and Wang, C. C. (2004) J Biol Chem 279, 20519-205284. Li, Z., and Wang, C. C. (2003) J Biol Chem 278, 20652-206585. Zachariae, W., and Nasmyth, K. (1999) Genes Dev 13, 2039-20586. Peters, J. M. (2002) Mol Cell 9, 931-9437. Harper, J. W., Burton, J. L., and Solomon, M. J. (2002) Genes Dev 16, 2179-22068. Michaelis, C., Ciosk, R., and Nasmyth, K. (1997) Cell 91, 35-459. Ciosk, R., Zachariae, W., Michaelis, C., Shevchenko, A., Mann, M., and Nasmyth, K.
(1998) Cell 93, 1067-107610. Yoon, H. J., Feoktistova, A., Wolfe, B. A., Jennings, J. L., Link, A. J., and Gould, K. L.
(2002) Curr Biol 12, 2048-205411. Visintin, R., Prinz, S., and Amon, A. (1997) Science 278, 460-46312. Wirtz, E., Leal, S., Ochatt, C., and Cross, G. A. (1999) Mol Biochem Parasitol 99, 89-
10113. Hirumi, H., and Hirumi, K. (1989) J Parasitol 75, 985-98914. Wang, Z., Morris, J. C., Drew, M. E., and Englund, P. T. (2000) J Biol Chem 275, 40174-
4017915. Li, Z., and Wang, C. C. (2002) J Biol Chem 277, 42686-4269316. Li, Z., Zou, C. B., Yao, Y., Hoyt, M. A., McDonough, S., Mackey, Z. B., Coffino, P., and
Wang, C. C. (2002) J Biol Chem 277, 15486-1549817. Morris, J. C., Wang, Z., Drew, M. E., Paul, K. S., and Englund, P. T. (2001) Mol
Biochem Parasitol 117, 111-11318. Mutomba, M. C., To, W. Y., Hyun, W. C., and Wang, C. C. (1997) Mol Biochem
Parasitol 90, 491-50419. Hughes, T. R., Marton, M. J., Jones, A. R., Roberts, C. J., Stoughton, R., Armour, C. D.,
Bennett, H. A., Coffey, E., Dai, H., He, Y. D., Kidd, M. J., King, A. M., Meyer, M. R.,Slade, D., Lum, P. Y., Stepaniants, S. B., Shoemaker, D. D., Gachotte, D., Chakraburtty,K., Simon, J., Bard, M., and Friend, S. H. (2000) Cell 102, 109-126
by guest, on January 24, 2013w
ww
.jbc.orgD
ownloaded from

8
20. Zachariae, W., Shevchenko, A., Andrews, P. D., Ciosk, R., Galova, M., Stark, M. J.,Mann, M., and Nasmyth, K. (1998) Science 279, 1216-1219
21. Birkett, C. R., Foster, K. E., Johnson, L., and Gull, K. (1985) FEBS Lett 187, 211-21822. Sasse, R., and Gull, K. (1988) J Cell Sci 90 ( Pt 4), 577-58923. Gmachl, M., Gieffers, C., Podtelejnikov, A. V., Mann, M., and Peters, J. M. (2000) Proc
Natl Acad Sci U S A 97, 8973-897824. Lamb, J. R., Michaud, W. A., Sikorski, R. S., and Hieter, P. A. (1994) Embo J 13, 4321-
432825. Carroll, C. W., and Morgan, D. O. (2002) Nat Cell Biol 4, 880-88726. Peters, J. M., King, R. W., Hoog, C., and Kirschner, M. W. (1996) Science 274, 1199-
120127. Yamashita, Y. M., Nakaseko, Y., Samejima, I., Kumada, K., Yamada, H., Michaelson,
D., and Yanagida, M. (1996) Nature 384, 276-27928. Zachariae, W., Shin, T. H., Galova, M., Obermaier, B., and Nasmyth, K. (1996) Science
274, 1201-120429. Grossberger, R., Gieffers, C., Zachariae, W., Podtelejnikov, A. V., Schleiffer, A.,
Nasmyth, K., Mann, M., and Peters, J. M. (1999) J Biol Chem 274, 14500-1450730. Lupas, A., Baumeister, W., and Hofmann, K. (1997) Trends Biochem Sci 22, 195-19631. Kramer, K. M., Fesquet, D., Johnson, A. L., and Johnston, L. H. (1998) Embo J 17, 498-
50632. Yu, H., Peters, J. M., King, R. W., Page, A. M., Hieter, P., and Kirschner, M. W. (1998)
Science 279, 1219-122233. Zachariae, W., Schwab, M., Nasmyth, K., and Seufert, W. (1998) Science 282, 1721-
172434. Ohta, T., Michel, J. J., and Xiong, Y. (1999) Oncogene 18, 6758-676635. Smith, K. N., Iwanejko, L., Loeillet, S., Fabre, F., and Nicolas, A. (1999) Yeast 15, 1255-
126736. Heichman, K. A., and Roberts, J. M. (1996) Cell 85, 39-4837. Hammarton, T. C., Clark, J., Douglas, F., Boshart, M., and Mottram, J. C. (2003) J Biol
Chem 278, 22877-2288638. Arhin, G. K., Shen, S., Ullu, E., and Tschudi, C. (2004) Methods Mol Biol 270, 277-28639. Townsley, F. M., Aristarkhov, A., Beck, S., Hershko, A., and Ruderman, J. V. (1997)
Proc Natl Acad Sci U S A 94, 2362-236740. Irniger, S., and Nasmyth, K. (1997) J Cell Sci 110 ( Pt 13), 1523-153141. Tugendreich, S., Boguski, M. S., Seldin, M. S., and Hieter, P. (1993) Proc Natl Acad Sci
U S A 90, 10031-1003542. Tugendreich, S., Tomkiel, J., Earnshaw, W., and Hieter, P. (1995) Cell 81, 261-26843. Holloway, S. L., Glotzer, M., King, R. W., and Murray, A. W. (1993) Cell 73, 1393-140244. Surana, U., Amon, A., Dowzer, C., McGrew, J., Byers, B., and Nasmyth, K. (1993)
Embo J 12, 1969-197845. Li, J., Ruyechan, W. T., and Williams, N. (2003) Biochem Biophys Res Commun 306,
918-923
FOOTNOTES
We would like to thank Dr. Keith Gull of Oxford University for providing us the KMX-1 antibody and
by guest, on January 24, 2013w
ww
.jbc.orgD
ownloaded from

9
Dr. Wolfgang Zachariae of the Max Plank Institute for the yeast apc1-1 temperature-sensitive mutant.Our thanks also go to Drs. W. Zacheus Cande of UC Berkeley and Nidhi Ahuja and the members ofWang’s lab for their critical readings of the manuscript and their constructive suggestions. This work wassupported by an NIH grant AI-21786.
Figure legends:
Fig.1 RNAi knockdown of individual APC/C core subunits in the procyclic and bloodstream forms of T.brucei. Cloned procyclic and bloodstream forms of T. brucei harboring individual APC/C RNAi plasmidswere incubated in the culture medium containing 1.0 mg/ml of tetracycline (©) or without tetracycline (n).Growth of the cells was monitored daily and plotted in a logarithmic scale. Insets depict semi-quantitative RT-PCR estimated levels of APC/C subunit mRNA three days into RNAi. Levels of a-tubulin (Tub) mRNA were measured as a sampling control. PCF, procyclic form; BSF, bloodstreamform; Tub, a-tubulin
Fig. 2 Complementation of the S. cerevisae cdc27 or apc1 mutants with T. brucei Cdc27 or Apc1 gene.Yeast cdc27 and apc1 mutants were transfected with the full-length genes of T. brucei Cdc27 and Apc1,respectively. The transfected cells were plated in various dilutions on YPD plates and incubated for threedays under different conditions. (A), Cdc27 complementation was assayed in the presence of 50mg/mldoxycycline; (B). Apc1 complementation was monitored by incubation at 37°C.
Fig. 3 FACScan analysis of Cdc27 and Apc1 depleted procyclic and bloodstream form of T. brucei.Cdc27/Apc1-depleted T. brucei cells were incubated under tetracycline for 4 days, labeled with PI andsubjected to FACScan analysis. The percentage of G1, S and G2/M-phase cells was calculated by usingsoftware ModFitLT. (A), Cdc27-depleted cells; (B), Apc1-depleted cells.
Fig. 4 Morphological distributions of APC/C subunit-depleted T. brucei cells. The cells, fixed in ethanoland labeled with PI, were visualized with a fluorescence microscope and tabulated in terms of numbers ofnuclei (N) and kinetoplasts (K) in each cell. Data are presented as mean percent (+S.E) of total cellscounted (>200) from three independent experiments. (A), Cdc27-depleted procyclic form; (B), Cdc27-depleted bloodstream form; (C), Apc1-depleted procyclic form; (D), Apc1-depleted bloodstream form.
Fig. 5 KMX-1 staining of the procyclic form of T. brucei arrested by Cdc27 depletion. Mitotic spindleswere labeled with the b-tubulin monoclonal mouse antibody, KMX-1 in Cdc27 deficient procyclic formof T. brucei (Green). The nucleus was stained by DAPI (Blue). (A) and (B) show 1N2K and cells withshort mitotic spindles in apparent metaphase. (C) demonstrates a zoid in the process of being generatedfrom a 1N1K cell that has a short mitotic spindle.
Fig. 6 Quantitative analysis of the mitotic spindles in the procyclic-form T. brucei cells with Cdc27 andApc1 depletion. The mitotic spindles were labeled with KMX-1 and the nuclei with DAPI. Cells withshort, medium sized and fully stretched mitotic spindles were tabulated and presented as mean percent(+S.E) of total cells counted (>200) from three independent experiments. (A), Cdc27-depletion; (B),Apc1-depletion.
Fig. 7 KMX-1 staining of the bloodstream form of T. brucei arrested by Cdc27 depletion. The mitoticspindles were labeled with KMX-1 whereas the nuclei by DAPI. (A) and (B) show two 2N2K cellsarrested in apparent late anaphase with elongated anaphase mitotic spindles.
by guest, on January 24, 2013w
ww
.jbc.orgD
ownloaded from
10
Table 1 Identification of APC/C subunit homologs in T. brucei
SubunitProtein size *
S. cerevisiae T. bruceiGeneDB
Accession#
T. bruceiIdentity/Similarity
(%)Motif
Apc1 1749 1839 Tb06.2N9.710 17/33 Rpn1/Rpn2Apc2 854 772 Tb09.211.2660 14/29 Cullin
Apc11 166 84 Tb09.211.1655 23/29 Ring fingerCdc16 841 601 Tb06.4M18.410 16/29 TPR repeatsCdc23 627 909 Tb927.1.39.10 17/29 TPR repeats
Cdc27 759 559 Tb10.406.0610 20/34 TPR repeats
Cdc20 611 541 Tb08.30K1.260 26/42 WD-40Doc1 250 235 Tb10.61.0630 25/37 Doc domainUbc4 149 150 Tb05.28F8.340 71/85 E2 enzymeFull-length protein sequence of S. cerevisiae APC/C subunits were compared with the full-length proteinsequence of potential APC/C candidates in the T. brucei genome database using MacVactor7.2 software.ClustalW alignment was performed using BLOSSUM 30 matrix with slow alignment.
* Number of amino acid residues
by guest, on January 24, 2013w
ww
.jbc.orgD
ownloaded from

Fig. 1
(A)
Cdc27 PCF
1.E+00
1.E+01
1.E+02
1.E+03
1.E+04
1.E+05
1.E+06
0 1 2 3 4 5 6 7
Days
Cel
ls 1
x104 /m
l
- + TetCdc27
Tub
100
101
102
103
104
105
106
Cdc27 BSF
1.E+00
1.E+01
1.E+02
1.E+03
1.E+04
1.E+05
1.E+06
0 1 2 3 4
DaysC
ell
s 1
x1
04/
ml
Cdc27Tub
- + Tet
100
101
102
103
104
105
106
(B) Apc1 PCF
1.E+00
1.E+01
1.E+02
1.E+03
1.E+04
1.E+05
0 1 2 3 4 5 6 7
Days
Cel
ls 1
x104 /m
l
- + TetApc1Tub
100
101
102
103
104
105
Apc1 BSF
1.E+00
1.E+01
1.E+02
1.E+03
1.E+04
1.E+05
1.E+06
0 1 2 3 4
Days
Cell
s 1
x1
04/
ml
Apc1Tub
- + Tet
100
101
102
103
104
105
106
Apc2 BSF
1.E+00
1.E+01
1.E+02
1.E+03
1.E+04
1.E+05
1.E+06
0 1 2 3 4
Days
Cell
s 1
x1
04/
ml
- + TetApc2Tub
100
101
102
103
104
105
106
(C)Apc2 PCF
1.E+00
1.E+01
1.E+02
1.E+03
1.E+04
1.E+05
1.E+06
0 1 2 3 4 5 6 7 8
Days
Cel
ls 1
x104 /m
l
- + !!TetApc2Tub
100
101
102
103
104
105
106
by guest, on January 24, 2013w
ww
.jbc.orgD
ownloaded from
(E)
(D)- + Tet
Apc11 PCF
1.E+00
1.E+01
1.E+02
1.E+03
1.E+04
1.E+05
0 1 2 3 4 5 6 7 8
Days
Cel
ls 1
x104 /m
lApc11
Tub
100
101
102
103
104
105
Cdc16 PCF
1.E+00
1.E+01
1.E+02
1.E+03
1.E+04
1.E+05
0 1 2 3 4 5 6 7
Days
Cel
ls 1
x104 /m
l
Cdc16
Tub
- + Tet
100
101
102
103
104
105
(F)Cdc23 BSF
1.E+00
1.E+01
1.E+02
1.E+03
1.E+04
1.E+05
1.E+06
0 1 2 3 4
Days
Cell
s 1
x1
04/
ml
Cdc23Tub
- + Tet
100
101
102
103
104
105
106
Apc11 BSF
1.E+00
1.E+01
1.E+02
1.E+03
1.E+04
1.E+05
1.E+06
1.E+07
0 1 2 3 4
Days
Cells
1x1
04/
ml
Apc11Tub
- + Tet
100
101
102
103
104
105
106
107
Cdc16 BSF
1.E+00
1.E+01
1.E+02
1.E+03
1.E+04
1.E+05
1.E+06
0 1 2 3 4
Days
Cells
1x1
04/
ml
Cdc16
Tub
- + Tet
100
101
102
103
104
105
106
Cdc23 PCF
1.E+00
1.E+01
1.E+02
1.E+03
1.E+04
1.E+05
0 1 2 3 4 5 6 7
Days
Cel
ls 1
x104 /m
l
Cdc23- + Tet
100
101
102
103
104
105
Tub
Fig. 1
by guest, on January 24, 2013w
ww
.jbc.orgD
ownloaded from

Cells
Vector
Ycdc27
Tcdc27
Doxycycline ug/ml 0 50
Fig. 2(A)
Cells
Vector
YApc1
TApc1
Temperature 25 oC 37 oC
(B)
Ts Apc1-1 cells
tetO7 regulated Cdc27 cells
by guest, on January 24, 2013w
ww
.jbc.orgD
ownloaded from

Cdc27 BSF
0
10
20
30
40
50
60
0 1 2 3
Days
% c
ell
G1
G2
S
Cdc27 PCF
0
10
20
30
40
50
60
0 1 2 3
Days
% c
ell
G1
G2
S
Fig. 3A
0
1
2
3
BSF
+Tet
PCF
-Tet +Tet-Tet
G1G2/M
G1G2/M
G1G2/M
G1G2/M
Days of RNAi
by guest, on January 24, 2013w
ww
.jbc.orgD
ownloaded from

Apc1 PCF
0
10
20
30
40
50
60
0 1 2 3
Days
% c
ell
G1
G2
S
Apc1 BSF
0
10
20
30
40
50
60
0 1 2 3
Days
% c
ell
G1
G2
S
Fig. 3B
Tet+
0
1
2
3
-Tet
G1G2/M
+Tet-Tet
PCF BSF
G1G2/M
G1G2/M
G1G2/M
Days of RNAi
by guest, on January 24, 2013w
ww
.jbc.orgD
ownloaded from
Zoid2N2K
Fig. 4A
Cdc27 PCF
0
10
20
30
40
50
60
70
80
90
1N1K 1N2K 2N2K Zoid 2N1K XNXK
%C
ell
-Tet
+Tet
by guest, on January 24, 2013w
ww
.jbc.orgD
ownloaded from

2N2K
Fig. 4B
Cdc27 BSF
0
10
20
30
40
50
60
70
80
90
100
1N1K 1N2K 2N2K Zoid 2N1K XNXK
% C
ell
s
-Tet
+Tet
by guest, on January 24, 2013w
ww
.jbc.orgD
ownloaded from

Fig. 4C
2N2K ZOID
Apc1 PCF
0
10
20
30
40
50
60
70
80
90
100
1N1K 1N2K 2N2K Zoid 2N1K XNXK
% C
ell
s
-Tet
+Tet
by guest, on January 24, 2013w
ww
.jbc.orgD
ownloaded from

Fig. 4D
2N2K 2N2K
Apc1 BSF
0
10
20
30
40
50
60
70
80
90
100
1N1K 1N2K 2N2K Zoid 2N1K XNXK
% C
ell
s
-Tet
+Tet
by guest, on January 24, 2013w
ww
.jbc.orgD
ownloaded from

Fig. 5
Zoid
DAPI KMX-1 Merge
(A)
(B)
(C)
by guest, on January 24, 2013w
ww
.jbc.orgD
ownloaded from

0
5
10
15
20
25
shortspindles
mid sizespindles
fullystrechedspindles
% c
ell
-Tet
+Tet
0
5
10
15
20
25
shortspindles
mid sizespindles
fullystrechedspindles
% C
ells
-Tet
+Tet
Fig. 6
(A) (B)
by guest, on January 24, 2013w
ww
.jbc.orgD
ownloaded from
DAPI KMX-1 Merge
Fig. 7
(A)
(B)
by guest, on January 24, 2013w
ww
.jbc.orgD
ownloaded from




















