
An antigen located in the kinetochore region in metaphase and on polar microtubule ends in the...
7
Chromosoma (Berl) (1990) 99:95 101 CHROMOSOMA © Springer-Verlag1990 An antigen located in the kinetochore region in metaphase and on polar microtubule ends in the midbody region in anaphase, characterised using a monoclonal antibody Roumen Pankov*, Margot Lemieux, and Ronald Hancock Centre de Recherche en Canc6rologie de l'Universit6 Laval, H6tel-Dieu Hospital, Qu6bec, Canada GI R 2J6 Received August 11, 1989 / in revised form November 24, 1989 Accepted December 26, 1989 by P.B. Moens Abstract. We describe a new component of the kineto- chore region of Chinese hamster ovary cells, which was characterised using a monoclonal antibody (mAb). This antigen was localised on the kinetochore regions of puri- fied metaphase chromosomes, but in anaphase it was instead located on the polar microtubules in the mid- body region, where they terminate in the stembody. It was not detectable in prophase or interphase cells by immunofluorescence, but was present in the interphase nucleus as shown by immunoblotting after SDS-poly- acrylamide gel electrophoresis. The mAb recognised two polypeptides of Mr 140 000 and 155 000. The localisation of this antigen in metaphase on the kinetochore region, where the plus ends of the kinetochore microtubules are temporarily stabilised when they attach, and later in the stembody and midbody where the plus ends of the polar microtubules are stabilised in anaphase and telophase, suggests that it could play a role in stabilising the plus ends of microtubules and thus in the control of microtu- bule dynamics during mitosis. Introduction A number of polypeptide components of the kinetochore and centromere region of chromosomes have been iden- tified using specific antibodies in the serum of patients suffering from the CREST syndrome (Brenner et al. 1981 ; Cooke et al. 1987; Cox et al. 1983; Earnshaw and Rothfield 1985; Guldner etal. 1984; Kingwell and Rattner 1987; Morio et al. 1980; Hadlaczky et al. 1989). To explore further the structure of the kinetochore we have prepared monoclonal antibodies (mAbs) directed against components of this structure, and we describe here a new component whose location changes during the course of mitosis. * Present address." Department of Cytology, University of Sofia, 1421 Sofia, Bulgaria Offprint requests to : R. Hancock The kinetochore contains attachment sites for micro- tubules; these grow from the centrioles in early meta- phase, and upon attaching to a kinetochore are stabilised at this, their plus, end (Euteneuer and Mclntosh 1981; Euteneuer et al. 1983; Gorbsky et al. 1987, 1988; Hui- torel and Kirschner 1988; Koshland et al. 1988; McIn- tosh and Landis 1971; Mitchison and Kirschner 1985a, b; Mitchison etal. 1986; Spurck and Pickett-Heaps 1987; Telzer and Haimo 1981). The stabilisation is re- leased in anaphase, resulting in depolymerisation at the kinetochore end and shortening of these microtubules (Gorbsky et al. 1987, 1988; Koshland et al. 1988; Mitch- ison et al. 1986). Other (polar) microtubules grow until their plus ends meet and overlap in the stembody, in the plane of cell division (Buck and Tisdale 1962;- Krishan and Buck 1965); these probably maintain the separation of the centrioles while the kinetochore micro- tubules shorten, although experimental studies of this question have not been described. Sister chromatids are separated by the forces resulting from this cycle of growth, stabilisation, and depolymerisation of microtu- bules, but the factors which control these processes have not yet been identified. The properties of the antigen described here suggests that it could play a role in these events. Materials and methods Cells, nuclei and chromosomes. Chinese hamster ovary (CHO) cells were grown in monolayer cultures in McCoy's 5A medium with 10% foetal calf serum. Mitotic cells of > 95% purity were prepared by shaking growing cultures on a horizontal platform, 2 h after a preliminary shaking to dislodge debris and replacing the medium with fresh medium containing 0.06 Ixg/mlColcemid. Chromosomes were purified from mitotic cells as described by Gooderham and Jeppesen (1983). Nuclei were prepared from CHO cells detached with trypsin, washed in 0.15 M NaC1, 0.1 M sodium phosphate, pH 7.5 (PBS), and suspended in nuclear isolation buffer [NB; 50 mM Tris-HC1, pH 7.4, 0.25 M sucrose, 5 mM MgSO4, 1 mM phenylmethylsulphonyl fluoride (PMSF), 100 units/ml Trasylol (Miles)] by rupture in a Dounce homogeniser after addition of Triton X-100 to 0.5%. The nuclei were then centrifuged (3,000 g,
Transcript of An antigen located in the kinetochore region in metaphase and on polar microtubule ends in the...

Chromosoma (Berl) (1990) 99:95 101 C H R O M O S O M A © Springer-Verlag 1990
An antigen located in the kinetochore region in metaphase and on polar microtubule ends in the midbody region in anaphase, characterised using a monoclonal antibody Roumen Pankov*, Margot Lemieux, and Ronald Hancock
Centre de Recherche en Canc6rologie de l'Universit6 Laval, H6tel-Dieu Hospital, Qu6bec, Canada GI R 2J6
Received August 11, 1989 / in revised form November 24, 1989 Accepted December 26, 1989 by P.B. Moens
Abstract. We describe a new component of the kineto- chore region of Chinese hamster ovary cells, which was characterised using a monoclonal ant ibody (mAb). This antigen was localised on the kinetochore regions of puri- fied metaphase chromosomes, but in anaphase it was instead located on the polar microtubules in the mid- body region, where they terminate in the stembody. It was not detectable in prophase or interphase cells by immunofluorescence, but was present in the interphase nucleus as shown by immunoblot t ing after SDS-poly- acrylamide gel electrophoresis. The mAb recognised two polypeptides of Mr 140 000 and 155 000. The localisation of this antigen in metaphase on the kinetochore region, where the plus ends of the kinetochore microtubules are temporari ly stabilised when they attach, and later in the s tembody and midbody where the plus ends of the polar microtubules are stabilised in anaphase and telophase, suggests that it could play a role in stabilising the plus ends of microtubules and thus in the control of microtu- bule dynamics during mitosis.
Introduction
A number of polypeptide components of the kinetochore and centromere region of chromosomes have been iden- tified using specific antibodies in the serum of patients suffering f rom the CREST syndrome (Brenner et al. 1981 ; Cooke et al. 1987; Cox et al. 1983; Earnshaw and Rothfield 1985; Guldner etal . 1984; Kingwell and Rat tner 1987; Morio et al. 1980; Hadlaczky et al. 1989). To explore further the structure of the kinetochore we have prepared monoclonal antibodies (mAbs) directed against components of this structure, and we describe here a new component whose location changes during the course of mitosis.
* Present address." Department of Cytology, University of Sofia, 1421 Sofia, Bulgaria
Offprint requests to : R. Hancock
The kinetochore contains a t tachment sites for micro- tubules; these grow from the centrioles in early meta- phase, and upon attaching to a kinetochore are stabilised at this, their plus, end (Euteneuer and Mclntosh 1981; Euteneuer et al. 1983; Gorbsky et al. 1987, 1988; Hui- torel and Kirschner 1988; Koshland et al. 1988; McIn- tosh and Landis 1971; Mitchison and Kirschner 1985a, b; Mitchison etal . 1986; Spurck and Pickett-Heaps 1987; Telzer and Haimo 1981). The stabilisation is re- leased in anaphase, resulting in depolymerisation at the kinetochore end and shortening of these microtubules (Gorbsky et al. 1987, 1988; Koshland et al. 1988; Mitch- ison et al. 1986). Other (polar) microtubules grow until their plus ends meet and overlap in the stembody, in the plane of cell division (Buck and Tisdale 1962;- Krishan and Buck 1965); these probably maintain the separation of the centrioles while the kinetochore micro- tubules shorten, al though experimental studies of this question have not been described. Sister chromatids are separated by the forces resulting f rom this cycle of growth, stabilisation, and depolymerisation of microtu- bules, but the factors which control these processes have not yet been identified. The properties of the antigen described here suggests that it could play a role in these events.
Materials and methods
Cells, nuclei and chromosomes. Chinese hamster ovary (CHO) cells were grown in monolayer cultures in McCoy's 5A medium with 10% foetal calf serum. Mitotic cells of > 95% purity were prepared by shaking growing cultures on a horizontal platform, 2 h after a preliminary shaking to dislodge debris and replacing the medium with fresh medium containing 0.06 Ixg/ml Colcemid. Chromosomes were purified from mitotic cells as described by Gooderham and Jeppesen (1983). Nuclei were prepared from CHO cells detached with trypsin, washed in 0.15 M NaC1, 0.1 M sodium phosphate, pH 7.5 (PBS), and suspended in nuclear isolation buffer [NB; 50 mM Tris-HC1, pH 7.4, 0.25 M sucrose, 5 mM MgSO4, 1 mM phenylmethylsulphonyl fluoride (PMSF), 100 units/ml Trasylol (Miles)] by rupture in a Dounce homogeniser after addition of Triton X-100 to 0.5%. The nuclei were then centrifuged (3,000 g,
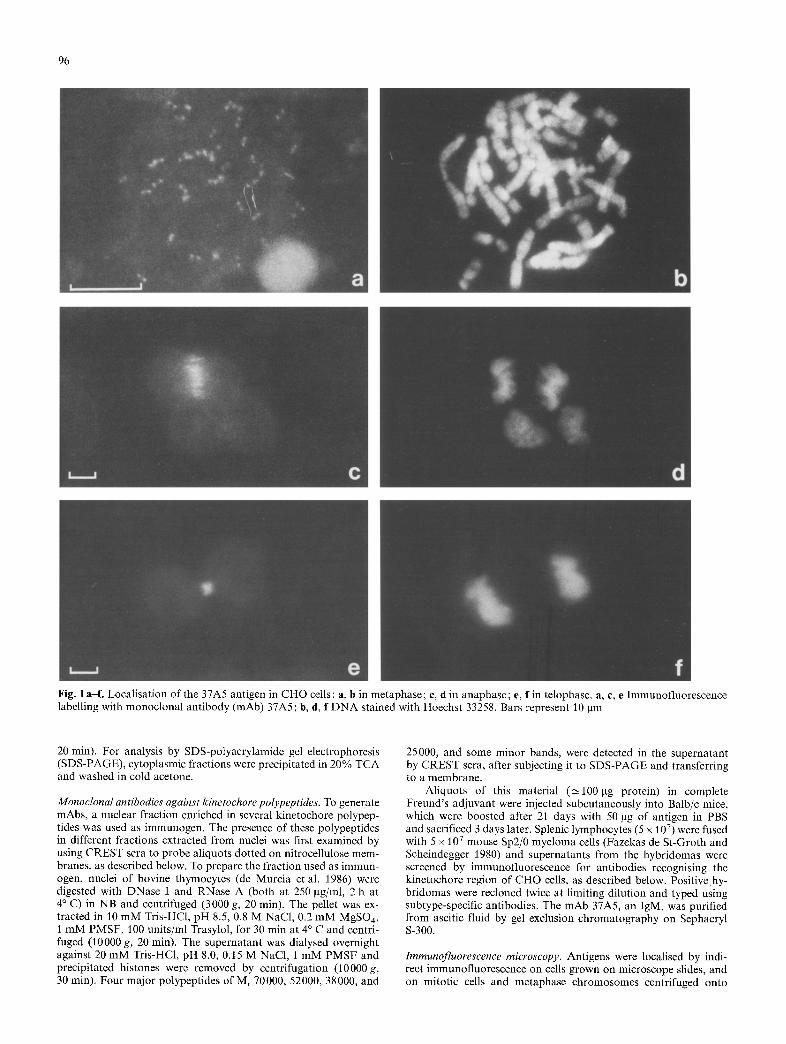
96
Fig. 1 a-f. Localisation of the 37A5 antigen in CHO cells: a, b in metaphase; e, d in anaphase; e, f in telophase, a, e, e Immunofluorescence labelling with monoclonal antibody (mAb) 37A5; b, d, f DNA stained with Hoechst 33258. Bars represent 10 gm
20 min). For analysis by SDS-polyacrylamide gel electrophoresis (SDS-PAGE), cytoplasmic fractions were precipitated in 20% TCA and washed in cold acetone.
Monoclonal antibodies against kinetochore polypeptides. To generate mAbs, a nuclear fraction enriched in several kinetochore polypep- tides was used as immunogen. The presence of these polypeptides in different fractions extracted from nuclei was first examined by using CREST sera to probe aliquots dotted on nitrocellulose mem- branes, as described below. To prepare the fraction used as immun- ogen, nuclei of bovine thymocytes (de Murcia et al. 1986) were digested with DNase I and RNase A (both at 250 gg/ml, 2 h at 4 ° C) in NB and centrifuged (3000 g, 20 min). The pellet was ex- tracted in 10 mM Tris-HC1, pH 8.5, 0.8 M NaC1, 0.2 mM MgSO4, 1 mM PMSF, 100 units/ml Trasylol, for 30 min at 4 ° C and centri- fuged (10000 g, 20 min). The supernatant was dialysed overnight against 20 mM Tris-HC1, pH 8.0, 0.15 M NaC1, 1 mM PMSF and precipitated histones were removed by centrifugation (10000 g, 30 min). Four major polypeptides of Mr 70000, 52000, 38000, and
25000, and some minor bands, were detected in the supernatant by CREST sera, after subjecting it to SDS-PAGE and transferring to a membrane.
Aliquots of this material ( ~ 1 0 0 g g protein) in complete Freund's adjuvant were injected subcutaneously into Balb/c mice, which were boosted after 21 days with 50 gg of antigen in PBS and sacrificed 3 days later. Splenic lymphocytes (5 x 107) were fused with 5 x 107 mouse Sp2/0 myeloma cells (Fazekas de St-Groth and Scheindegger 1980) and supernatants from the hybridomas were screened by immunoftuorescence for antibodies recognising the kinetochore region of CHO cells, as described below. Positive hy- bridomas were recloned twice at limiting dilution and typed using subtype-specific antibodies. The mAb 37A5, an IgM, was purified from ascitic fluid by gel exclusion chromatography on Sephacryl S-300.
ImmunojTuorescence microscopy. Antigens were localised by indi- rect immunofluorescence on cells grown on microscope slides, and on mitotic cells and metaphase chromosomes centrifuged onto
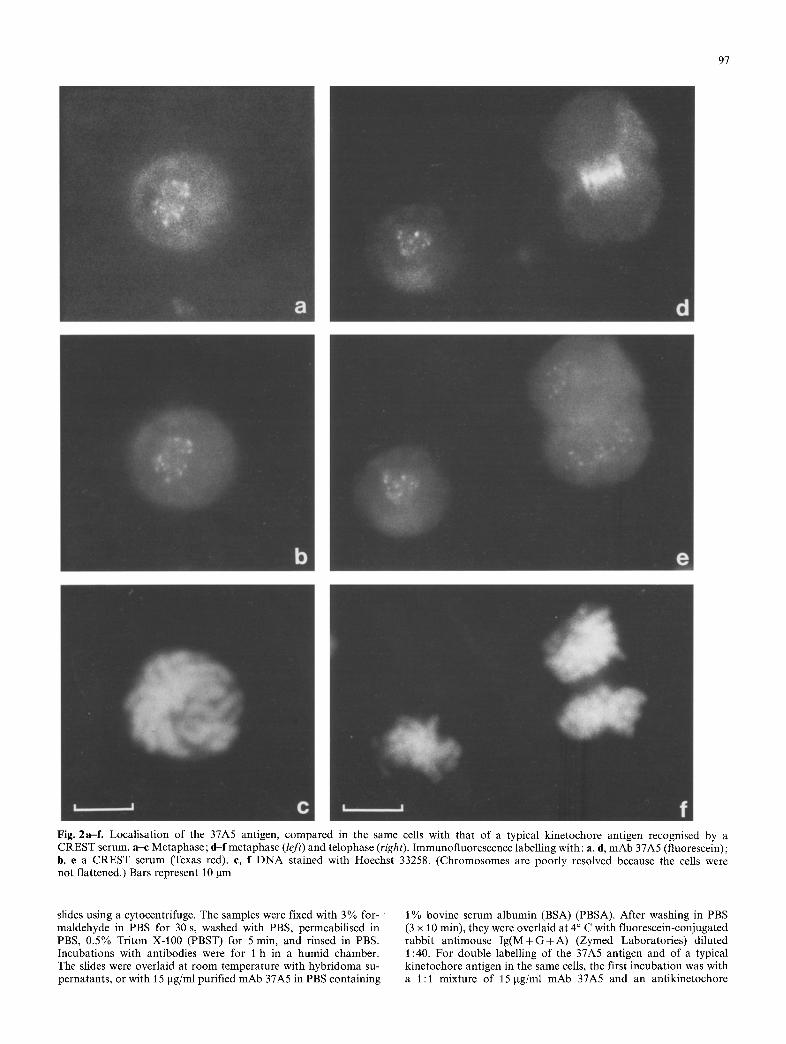
97
Fig. 2a-f . Localisation of the 37A5 antigen, compared in the same cells with that of a typical kinetochore antigen recognised by a CREST serum, a-c Metaphase; d-f metaphase (left) and telophase (right). Immunofluorescence labelling with: a, d, mAb 37A5 (fluorescein); b, e a CREST serum (Texas red). c, f DNA stained with Hoechst 33258. (Chromosomes are poorly resolved because the cells were not flattened.) Bars represent 10 lam
slides using a cytocentrifuge. The samples were fixed with 3% for- maldehyde in PBS for 30 s, washed with PBS, permeabilised in PBS, 0.5% Triton X-100 (PBST) for 5 min, and rinsed in PBS. Incubations with antibodies were for 1 h in a humid chamber. The slides were overlaid at room temperature with hybridoma su- pernatants, or with 15 gg/ml purified mAb 37A5 in PBS containing
1% bovine serum albumin (BSA) (PBSA). After washing in PBS (3 x 10 min), they were overlaid at 4 ° C with fluorescein-conjugated rabbit antimouse I g ( M + G + A ) (Zymed Laboratories) diluted 1:40. For double labelling of the 37A5 antigen and of a typical kinetochore antigen in the same cells, the first incubation was with a 1:1 mixture of 15 gg/ml mAb 37A5 and an antikinetochore

98
CREST serum diluted 1 : 1000, followed by a 1 : 1 mixture of fluo- rescein-conjugated antimouse antibodies (above) and Texas red- conjugated rabbit antihuman Ig(M+G+A), diluted 1:40. After washing in PBS (3 x 10 min) the preparations were mounted in 50% glycerol, 0.15 M NaC1, 0.1 M glycine, pH 8.6 containing the fluorescent DNA stain Hoechst 33258 (10 gg/ml) and examined in a Zeiss fluorescence microscope using appropriate filters.
SDS-PAGE and immunodetection of polypeptides on gels'. Fractions from -~ 10 v cells were solubilised by addition of threefold concen- trated sample buffer, heated in a boiling water bath for 3 min, and subjected to SDS-PAGE in 1 mm thick minigels containing 7.5% acrylamide in a Protean II apparatus (BioRad) using the procedures, gel solutions, and buffers specified by the manufactur- er. Gels were either stained with Coomassie Blue, or the polypep- tides were transferred electrophoretically to nitrocellulose mem- branes (Schleicher and Schuell) (Towbin et al. 1979) which were probed with antibodies. To block nonspecific reactions the mem- branes were incubated for 1 h at room temperature in 10 mM Tris- HC1, pH 7.4, 0.15 M NaCI (TBS) containing 5% w/v powdered skimmed milk, and then for 3 h with 50 gg/ml mAb 37A5 in TBS, 0.05% Tween 20, 1% w/v skimmed milk (TTBS). They were washed in TTBS, incubated for 1 h with lzSI-labelled (Hunter and Greenwood 1962) rabbit antimouse Ig(M + G+ A) (Zymed Labo- ratories) in TTBS, and exposed to X-ray film at -80 ° C.
Results
Localisation of the 37A5 antigen by immunoJIuorescence
The antigen described here was located on the kineto- chore region of the chromosomes in metaphase cells (Fig. 1 a). Labelling of purified chromosomes confirmed that the antigen was located on this region (see below). In contrast, in anaphase cells the antigen (referred to as the 37A5 antigen) was no longer situated in the kine- tochore region, but instead on the polar microtubules in the stembody, where they terminate and overlap (Buck and Tisdale 1962; Krishan and Buck 1965) (Fig. 1 c), and in telophase cells it remained localised in a compact manner in this region (Euteneuer and McIntosh 1980; Fuge 1977; Inou6 1981; Mullins and McIntosh 1982; Sellitto and Kur iyama 1988) (Fig. 1 e). The localisation was identical in mitotic cells collected without exposure to colcemid. No immunofluorescence signal above background could be detected in interphase or prophase cells.
Such a change of localisation during the course of mitosis is not shown by kinetochore antigens recognised by CREST sera (Brenner et al. 1981 ; Cooke et al. 1987; Cox et al. 1983; Earnshaw and Rothfield 1985; Guldner et al. 1984; Kingwell and Rat tner 1987; Morio e ta l . 1980; Hadlaczky et al. 1989) with one exception (Fritzler et al. 1987), nor by other mAbs generated during our work. This contrast could be demonstrated by double immunofluorescence labelling of mitotic cells with mAb 37A5 and a CREST serum (Fig. 2d, e).
Localisation of the 37A5 antigen on the kinetochore region of metaphase chromosomes
Immunofluorescence labelling of purified metaphase chromosomes showed that the 37A5 antigen was local-
/l ll/ Fig. 3a-f. Localisation of the 37A5 antigen on both kinetochore regions of purified chromosomes, a, e, e DNA stained with Hoechst 33258; b, d, f immunofluorescence labelling with mAb 37A5. Bar represents 10 gm
ised on the kinetochore region of both chromatids (Fig. 3). An identical pat tern was seen in chromosomes purified further by centrifugation through a glycerol gra- dient (Gooderham and Jeppesen 1983) but the intensity was lower (not shown), suggesting that the antigen is partially dissociated under these conditions, a conclusion supported by immunoblot t ing of polypeptides separated by SDS-PAGE (Fig. 4).
Characterisation of the 37A5 antigen by SDS-PAGE
The mAb 37A5 recognised two polypeptides of Mr 140000 and 155000 in metaphase chromosomes (Fig. 4b, lane 3). These were less abundant in chromo- somes further purified by centrifugation through a gly- cerol gradient (Fig. 4b, lane 5), in agreement with the lower immunofluorescence signal f rom this material (see above). These polypeptides were also present in inter- phase nuclei (Fig. 4b, lane 1) but were not detectable in the cytoplasm of interphase or mitotic cells (Fig. 4b, lanes 2, 4).
Discussion
The 37A5 antigen is localised on the kinetochore region in metaphase, as shown by immunofluorescence label- ling of isolated chromosomes (Fig. 3). It appears to be in the the diffuse outer corona region of the kinetochore (Alov and Lyubskii 1977; Rieder 1982; Ris and Witt 1981) according to preliminary observations we have made by immunoelectron microscopy. This is the region where tubulin is bound (Mitchison and Kirschner 1985a) and where the kinetochore microtubules attach (Valdivia and Brinkley 1985). As shown by fluorescence microscopy (Figs. 1, 2) the antigen is no longer detect- able on the kinetochore region in anaphase, but is now localised on the polar microtubules in the stembody, where they overlap and terminate. In prophase and inter- phase cells the immunofluorescence signal is neither lo-
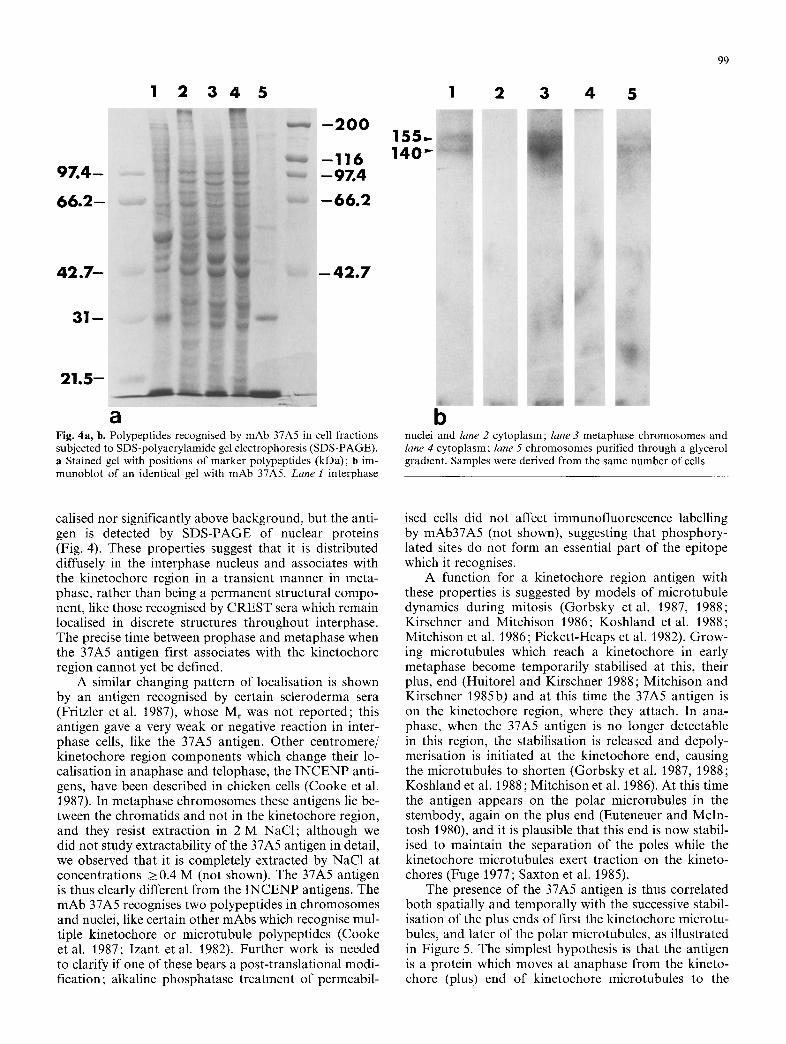
1 2 3 4 5
- 2 O O
- 1 1 6 - 9 Z 4
- 6 6 . 2
- - 4 2 . 7
a Fig. 4a, b. Polypeptides recognised by mAb 37A5 in cell fractions subjected to SDS-polyacrylamide gel electrophoresis (SDS-PAGE). a Stained gel with positions of marker polypeptides (kDa); b im- munoblot of an identical gel with mAb 37A5. Lane 1 interphase
155 140
1 2 3 4 5
99
b nuclei and lane 2 cytoplasm; lane 3 metaphase chromosomes and lane 4 cytoplasm; lane 5 chromosomes purified through a glycerol gradient. Samples were derived from the same number of cells
calised nor significantly above background, but the anti- gen is detected by SDS-PAGE of nuclear proteins (Fig. 4). These properties suggest that it is distributed diffusely in the interphase nucleus and associates with the kinetochore region in a transient manner in meta- phase, rather than being a permanent structural compo- nent, like those recognised by CREST sera which remain localised in discrete structures throughout interphase. The precise time between prophase and metaphase when the 37A5 antigen first associates with the kinetochore region cannot yet be defined.
A similar changing pattern of localisation is shown by an antigen recognised by certain scleroderma sera (Fritzler et al. 1987), whose Mr was not reported; this antigen gave a very weak or negative reaction in inter- phase cells, like the 37A5 antigen. Other centromere/ kinetochore region components which change their lo- calisation in anaphase and telophase, the INCENP anti- gens, have been described in chicken cells (Cooke et al. 1987). In metaphase chromosomes these antigens lie be- tween the chromatids and not in the kinetochore region, and they resist extraction in 2 M NaC1; although we did not study extractability of the 37A5 antigen in detail, we observed that it is completely extracted by NaC1 at concentrations > 0.4 M (not shown). The 37A5 antigen is thus clearly different from the INCENP antigens. The mAb 37A5 recognises two polypeptides in chromosomes and nuclei, like certain other mAbs which recognise mul- tiple kinetochore or microtubule polypeptides (Cooke et al. 1987; Izant et al. 1982). Further work is needed to clarify if one of these bears a post-translational modi- fication; alkaline phosphatase treatment of permeabil-
ised cells did not affect immunofluorescence labelling by mAb37A5 (not shown), suggesting that phosphory- lated sites do not form an essential part of the epitope which it recognises.
A function for a kinetochore region antigen with these properties is suggested by models of microtubule dynamics during mitosis (Gorbsky et al. 1987, 1988; Kirschner and Mitchison 1986; Koshland etal. 1988; Mitchison et al. 1986; Pickett-Heaps et al. 1982). Grow- ing microtubules which reach a kinetochore in early metaphase become temporarily stabilised at this, their plus, end (Huitorel and Kirschner 1988; Mitchison and Kirschner 1985b) and at this time the 37A5 antigen is on the kinetochore region, where they attach. In ana- phase, when the 37A5 antigen is no longer detectable in this region, the stabilisation is released and depoly- merisation is initiated at the kinetochore end, causing the microtubules to shorten (Gorbsky et al. 1987, 1988; Koshland et al. 1988; Mitchison et al. 1986). At this time the antigen appears on the polar microtubules in the stembody, again on the plus end (Euteneuer and McIn- tosh 1980), and it is plausible that this end is now stabil- ised to maintain the separation of the poles while the kinetochore microtubules exert traction on the kineto- chores (Fuge 1977; Saxton et al. 1985).
The presence of the 37A5 antigen is thus correlated both spatially and temporally with the successive stabil- isation of the plus ends of first the kinetochore microtu- bules, and later of the polar microtubules, as illustrated in Figure 5. The simplest hypothesis is that the antigen is a protein which moves at anaphase from the kineto- chore (plus) end of kinetochore microtubules to the

100
EARLY METAPHASE
/ l \ / \
CENTRIOLE/ \
\ 0 /
/ \
ANTIGEN
A.,P.ASE / / . oo, /
Fig, 5, A schema of the current model of the dynamics of the kinetochore and polar mierotubules during mitosis, related to the localisation of the 37A5 antigen
s tembody (plus) ends of polar micro tubules and plays a role in this stabil isation. Alternat ively, it could be an epitope having a par t icular con fo rma t ion or secondary modi f ica t ion ; the p h e n o m e n o n would then be no less interesting, bu t the ant igen would represent a conforma- t ional feature tempora l ly associated with the stabilisa- t ion of mic ro tubu le plus ends. A third, trivial possibil i ty is that the ant igen is associated with mic ro tubu le plus ends t h roughou t mitosis and has no dynamic role, bu t is masked in the k inetochore region in anaphase and telophase.
In order to evaluate these models we are examining the possible ident i ty of the 37A5 ant igen with a pro te in which can stabilise isolated micro tubules in vitro, STOPa45 (Margolis et al. 1986), which has a similar Mr.
Acknowledgements. We thank E. Beaulieu (University Hospital, Quebec) and M. Fritzler and S. Kabeary (Department of Medicine, University of Calgary) for gifts of CREST sera. We thank N. Islam and H. Lambert for help and advice, P. Paquin and G. Langlois for photography, and reviewers for constructive comments. This work was supported by Grant MA-9589 from the Medical Re- search Council of Canada.
References
Alov IA, Lyubskii SL (1977) Functional morphology of the kineto- chore. Int Rev Cytol Suppl 6: 59-74
Brenner S, Pepper D, Berns MW, Tan E, Brinkley BR (198l) Kine- tochore structure, duplication, and distribution in mammalian cells: analysis by human autoantibodies from scleroderma pa- tients. J Cell Biol 91:95-102
Buck RC, Tisdale JM (1962) The fine structure of the midbody of the rat erythrocyte. J Cell Biol 13 : 109-115
Cooke CA, Heck MMS, Earnshaw WC (1987) The inner centro- mere protein (INCENP) antigens: movement from inner cen- tromere to midbody during mitosis. J Cell Biol 105:2053 2067
Cox JV, Schenk EA, Olmsted JB (1983) Human anticentromere antibodies : distribution, characterisation of antigens, and effect on microtubule organisation. Cell 35:331-339
Earnshaw WC, Rothfield N (1985) Identification of a family of human centromere proteins using autoimmune sera from pa- tients with scleroderma. Chromosoma 91 : 313-321
Euteneuer U, McIntosh JR (1980) Polarity of midbody and phrag- moplast microtubules. J Cell Biol 87:509 515
Euteneuer U, McIntosh JR (1981) Structural polarity of kineto- chore microtubules in PtK1 cells. J Cell Biol 89:338-345
Euteneuer U, Ris H, Borisy GG (1983) Polarity of kinetochore microtubules in chinese hamster ovary cells after recovery from a colcemid block. J Cell Biol 97: 202-208
Fazekas de St-Groth S, Scheindegger D (1980) Production of monoclonal antibodies: strategy and tactics. J Immunol Meth- ods 35:1M
Fritzler M J, Ayer LM, Gohill J, O'Connor C, Laxer RM, Humbel R (1987) An antigen in metaphase chromatin and the midbody of mammalian cells binds to scleroderma sera. J Rheumatol 14: 291-294
Fuge H (1977) Ultrastructure of the mitotic spindle. Int Rev Cytol Suppl 6:1 58
Gooderham K, Jeppesen P (5983) Chinese hamster metaphase chromosomes isolated under physiological conditions. Exp Cell Res 144:1 14
Gorbsky GJ, Sammak P J, Borisy GG (1987) Chromosomes move poleward in anaphase along stationary microtubules that coor- dinately disassemble from their kinetochore ends. J Cell Biol 104:9-18
Gorbsky G J, Sammak P J, Borisy GG (1988) Microtubule dynamics and chromosome motion visualised in living anaphase cells. J Cell Biol 106 : 1185-1192
Guldner HH, Lakomek HJ, Bautz FA (1984) Human anticentro- mere sera recognise a 19.5 kD non-histone chromosomal pro- tein from HeLa cells. Clin Exp Immunol 58:13 20
Hadlaczky G, Praznovszky T, Rasko I, Kereso J (1989) Centro- mere proteins. 1. Mitosis specific centromere antigen recognized by anti-centromere autoantibodies. Chromosoma 97:282-288
Huitorel P, Kirschner MW (1988) The polarity and stability of microtubule capture by the kinetochore. J Cell Biol 105: 151- 159
Hunter WM, Greenwood FC (1962) Preparation of 1-131 labelled human growth hormone of high specific activity. Nature 194:495-496
Inou+ S (1981) Cell division and the mitotic spindle. J Cell Biol 91 : 131s-147s
Izant JG, Weatherbee JA, McIntosh JR (1982) A microtubule- associated protein found in the mitotic spindle and interphase nucleus. Nature 295 : 249 250
Kingwell B, Rattner JB (1987) Mammalian kinetochore/centro- mere composition: a 50 kDa antigen is present in the mamma- lian kinetochore/centromere. Chromosoma 95:403M07
Kirschner M, Mitchison T (1986) Beyond self-assembly: from mi- crotubules to morphogenesis. Cell 45:329-342
Koshland DE, Mitchison TJ, Kirschner MW (1988) Polewards chromosome movement driven by microtubule depolymerisa- tion in vitro. Nature 331:499-504
Krishan A, Buck RC (1965) Structure of the mitotic spindle in L strain fibroblasts. J Cell Biol 24:433M44
Margolis RL, Rauch CT, Job D (1986) Purification and assay of a 145-kDa protein (STOP145) with microtubule-stabilizing and motility behavior. Proc Natl Acad Sci USA 83 : 639-643

101
McIntosh JR, Landis SC (1971) The distribution of spindle micro- tubules during mitosis in cultured human cells. J Cell Biol 49 : 468-497
Mitchison TJ, Kirschner MW (1985a) Properties of the kineto- chore in vitro. I. Microtubule nucleation and tubulin binding. J Cell Biol 101:755-765
Mitchison TJ, Kirschner MW (1985b) Properties of the kineto- chore in vitro. II. Microtubule capture and ATP-dependent translocation. J Cell Biol 101:766-777
Mitchison T, Evans L, Schulze E, Kirschner M (1986) Sites of microtubule assembly and disassembly in the mitotic spindle. Cell 45:515-527
Morio Y, Peebles C, Fritzler MJ, Steigerwald J, Tan EM (1980) Autoantibody to centromere (kinetochore) in scleroderma sera. Proc Natl Acad Sci USA 77:1627-1631
Mullins JM, Mclntosh JR (1982) Isolation and characterisation of the mammalian midbody. J Cell Biol 94:654-661
de Murcia G, Huletsky A, Lamarre D, Gaudreau A, Pouyet J, Daune M, Poirier G (1986) Modulation of chromatin super- structure induced by poly(ADP-ribose) synthesis and degrada- tion. J Biol Chem 261:7011-7017
Pickett-Heaps JD, Tippit DH, Porter KR (1982) Rethinking mito- sis. Cell 29 : 729-744
Rieder CL (1982) The formation, structure, and composition of
the mammalian kinetochore and kinetochore fiber. Int Rev Cy- tol 79:1-58
Ris H, Witt PL (1981) Structure of the mammalian kinetochore. Chromosoma 82:153-170
Saxton WM, Stemple DL, Leslie RJ, Salmon ED, Zavortink M, McIntosh JR (1985) Tubulin dynamics in cultured mammalian cells. J Cell Biol 99:2175-2186
Sellitto C, Kuriyama R (1988) Distribution of a matrix component of the midbody during the cell cycle in chinese hamster ovary cells. J Cell Biol 106:431-439
Spurck TP, Pickett-Heaps JD (1987) On the mechanism of ana- phase A: evidence that ATP is needed for microtubule disas- sembly and not generation of polewards force. J Cell Biol 105:1691-1705
Telzer BR, Haimo LT (1981) Decoration of spindle microtubules with dynein: evidence for uniform polarity. J Cell Biol 89:373- 378
Towbin H, Staehelin T, Gordon J (1979) Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci USA 76:4350-4354
Valdivia MM, Brinkley BR (1985) Fractionation and initial charac- terisation of the kinetochore from mammalian metaphase chro- mosomes. J Cell Biol 101 : 1124-1134




















