
Deoxyribonucleic acid methyl transferases 3a and 3b associate with the nuclear orphan receptor...
14
Deoxyribonucleic Acid Methyl Transferases 3a and 3b Associate with the Nuclear Orphan Receptor COUP-TFI during Gene Activation Rozenn Gallais, Florence Demay, Peter Barath, Laurence Finot, Renata Jurkowska, Re ´ my Le Gue ´ vel Fre ´ de ´ rique Gay, Albert Jeltsch, Raphae ¨ l Me ´ tivier, and Gilles Salbert Unite ´ Mixte de Recherche 6026 (R.G., F.D., P.B., L.F., R.L.G., F.G., R.M., G.S.), Centre National de la Recherche Scientifique, Universite ´ Rennes I, Institut Fe ´ de ´ ratif de Recherche 140 Ge ´ nomique Fonctionnelle Agronomie Sante ´ , Equipe Information et Programmation Cellulaire; Equipe Endocrinologie Mole ´ culaire de la Reproduction, Campus de Beaulieu, 35042 Rennes Cedex, France; and International University Bremen (R.J., A.J.), 28759 Bremen, Germany Transcriptional activation of silent genes can require the erasure of epigenetic marks such as DNA meth- ylation at CpGs (cytosine-guanine dinucleotide). Ac- tive demethylation events have been observed, and associated processes are repeatedly suspected to involve DNA glycosylases such as mCpG binding do- main protein 4, thymine DNA glycosylase (TDG), Demeter, and repressor of silencing 1. A complete characterization of the molecular mechanisms oc- curring in metazoan is nonetheless awaited. Here, we report that activation of the endogenous vitronec- tin gene in P19 cells by the nuclear receptor chicken ovalbumin upstream promoter-transcription factor I (COUP-TFI) is observed in parallel with the recruit- ment of TDG and p68 RNA helicase, two components of a putative demethylation complex. Interestingly, when activated, the vitronectin gene was loaded with DNA methyltransferases 3a and 3b (Dnmt3a/b), and a strand-biased decrease in CpG methylation was de- tected. Dnmt3a was further found to associate with COUP-TFI and TDG in vivo, and cotransfection ex- periments demonstrated that Dnmt3a/b can en- hance COUP-TFI-mediated activation of a methyl- ated reporter gene. These results suggest that Dnmt3a/b could cooperate with the orphan receptor COUP-TFI to regulate transcription of the vitronectin gene. (Molecular Endocrinology 21: 2085–2098, 2007) D NA METHYLATION ON cytosine is an epigenetic mark found in vertebrates, plants, fungi, and, to a lesser extent, insects. Methylation, occurring mainly at CpGs (cytosine-guanine dinucleotide), has profound effects on gene activity. Indeed, it has been recog- nized that hypomethylated DNA is found in active genes, whereas hypermethylated genes are silent (1, 2). Furthermore, methylated CpGs recruit specific pro- teins that in turn recruit histone modifiers involved in chromatin condensation (1). The patterns of DNA methylation are established and maintained by DNA methyl transferases (Dnmt) that can act either on hemimethylated CpGs generated throughout the rep- lication of DNA (Dnmt1 or Dnmt3a and 3b), or de novo on unmethylated DNA (Dnmt3a and 3b). Maintenance methylation occurs during the S phase, with this cou- pling allowed by a physical interaction between Dnmt and the replication machinery. Conversely, mecha- nisms regulating de novo methylation are poorly un- derstood. However, as described for the oncogenic promyelocytic leukemia-retinoic acid receptor fusion protein (3), targeting could be achieved through direct interaction of the methyltransferases with sequence- specific DNA-binding proteins (4, 5). It has long been thought that DNA methylation is a stable mark that cannot be enzymatically removed, with multiple rounds of cell division without Dnmt- mediated remethylation required to completely erase this epigenetic mark from DNA. However, in the past recent years, numerous examples of active demethyl- ation events that would not rely on cell division were described (6–9). Active demethylation requires enzy- matic machineries that are able either to remove the methyl group branched on cytosines, or to replace the methylated nucleotides by unmethylated ones (10). Although 5-methylcytosine DNA glycosylases have been shown in plants (11–14), the data available from metazoan species are highly controversial and no de- methylase has been yet formally identified. Nonethe- less, a recent study demonstrated that active demeth- ylation in mammalian cells involves DNA strand breaks suggesting that DNA glycosylases and repair systems First Published Online June 19, 2007 Abbreviations: aa, Amino acids; AP, apurinic/apyrimidic; BER, base excision repair; 8-Br-cAMP, 8-bromo-cAMP; ChIP, Chromatin immunoprecipitation; CMV, cytomegalovi- rus; CpG, cytosine-guanine dinucleotide; Dnmt, DNA methyl transferases; DTT, dithiothreitol; ER, estrogen receptor; GST, glutathione S-transferase; HA, hemagglutinin; PKC, protein kinase C; Re-ChIP, sequential ChIP; SDS, sodium dodecyl sulfate; TDG, thymine DNA glycosylase; TPBSI, PBS/1% Tri- ton; wt, wild type. Molecular Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving the endocrine community. 0888-8809/07/$15.00/0 Molecular Endocrinology 21(9):2085–2098 Printed in U.S.A. Copyright © 2007 by The Endocrine Society doi: 10.1210/me.2006-0490 2085
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Deoxyribonucleic acid methyl transferases 3a and 3b associate with the nuclear orphan receptor...

Deoxyribonucleic Acid Methyl Transferases 3a and3b Associate with the Nuclear Orphan ReceptorCOUP-TFI during Gene Activation
Rozenn Gallais, Florence Demay, Peter Barath, Laurence Finot, Renata Jurkowska, Remy Le GuevelFrederique Gay, Albert Jeltsch, Raphael Metivier, and Gilles Salbert
Unite Mixte de Recherche 6026 (R.G., F.D., P.B., L.F., R.L.G., F.G., R.M., G.S.), Centre National dela Recherche Scientifique, Universite Rennes I, Institut Federatif de Recherche 140 GenomiqueFonctionnelle Agronomie Sante, Equipe Information et Programmation Cellulaire; EquipeEndocrinologie Moleculaire de la Reproduction, Campus de Beaulieu, 35042 Rennes Cedex, France;and International University Bremen (R.J., A.J.), 28759 Bremen, Germany
Transcriptional activation of silent genes can requirethe erasure of epigenetic marks such as DNA meth-ylation at CpGs (cytosine-guanine dinucleotide). Ac-tive demethylation events have been observed, andassociated processes are repeatedly suspected toinvolve DNA glycosylases such as mCpG binding do-main protein 4, thymine DNA glycosylase (TDG),Demeter, and repressor of silencing 1. A completecharacterization of the molecular mechanisms oc-curring in metazoan is nonetheless awaited. Here,we report that activation of the endogenous vitronec-tin gene in P19 cells by the nuclear receptor chickenovalbumin upstream promoter-transcription factor I(COUP-TFI) is observed in parallel with the recruit-
ment of TDG and p68 RNA helicase, two componentsof a putative demethylation complex. Interestingly,when activated, the vitronectin gene was loaded withDNA methyltransferases 3a and 3b (Dnmt3a/b), and astrand-biased decrease in CpG methylation was de-tected. Dnmt3a was further found to associate withCOUP-TFI and TDG in vivo, and cotransfection ex-periments demonstrated that Dnmt3a/b can en-hance COUP-TFI-mediated activation of a methyl-ated reporter gene. These results suggest thatDnmt3a/b could cooperate with the orphan receptorCOUP-TFI to regulate transcription of the vitronectingene. (Molecular Endocrinology 21: 2085–2098, 2007)
DNA METHYLATION ON cytosine is an epigeneticmark found in vertebrates, plants, fungi, and, to a
lesser extent, insects. Methylation, occurring mainly atCpGs (cytosine-guanine dinucleotide), has profoundeffects on gene activity. Indeed, it has been recog-nized that hypomethylated DNA is found in activegenes, whereas hypermethylated genes are silent (1,2). Furthermore, methylated CpGs recruit specific pro-teins that in turn recruit histone modifiers involved inchromatin condensation (1). The patterns of DNAmethylation are established and maintained by DNAmethyl transferases (Dnmt) that can act either onhemimethylated CpGs generated throughout the rep-lication of DNA (Dnmt1 or Dnmt3a and 3b), or de novoon unmethylated DNA (Dnmt3a and 3b). Maintenance
methylation occurs during the S phase, with this cou-pling allowed by a physical interaction between Dnmtand the replication machinery. Conversely, mecha-nisms regulating de novo methylation are poorly un-derstood. However, as described for the oncogenicpromyelocytic leukemia-retinoic acid receptor fusionprotein (3), targeting could be achieved through directinteraction of the methyltransferases with sequence-specific DNA-binding proteins (4, 5).
It has long been thought that DNA methylation is astable mark that cannot be enzymatically removed,with multiple rounds of cell division without Dnmt-mediated remethylation required to completely erasethis epigenetic mark from DNA. However, in the pastrecent years, numerous examples of active demethyl-ation events that would not rely on cell division weredescribed (6–9). Active demethylation requires enzy-matic machineries that are able either to remove themethyl group branched on cytosines, or to replace themethylated nucleotides by unmethylated ones (10).Although 5-methylcytosine DNA glycosylases havebeen shown in plants (11–14), the data available frommetazoan species are highly controversial and no de-methylase has been yet formally identified. Nonethe-less, a recent study demonstrated that active demeth-ylation in mammalian cells involves DNA strand breakssuggesting that DNA glycosylases and repair systems
First Published Online June 19, 2007Abbreviations: aa, Amino acids; AP, apurinic/apyrimidic;
BER, base excision repair; 8-Br-cAMP, 8-bromo-cAMP;ChIP, Chromatin immunoprecipitation; CMV, cytomegalovi-rus; CpG, cytosine-guanine dinucleotide; Dnmt, DNA methyltransferases; DTT, dithiothreitol; ER, estrogen receptor; GST,glutathione S-transferase; HA, hemagglutinin; PKC, proteinkinase C; Re-ChIP, sequential ChIP; SDS, sodium dodecylsulfate; TDG, thymine DNA glycosylase; TPBSI, PBS/1% Tri-ton; wt, wild type.
Molecular Endocrinology is published monthly by TheEndocrine Society (http://www.endo-society.org), theforemost professional society serving the endocrinecommunity.
0888-8809/07/$15.00/0 Molecular Endocrinology 21(9):2085–2098Printed in U.S.A. Copyright © 2007 by The Endocrine Society
doi: 10.1210/me.2006-0490
2085

are used (15). Among the candidate proteins, the T:Gmismatch DNA glycosylase (TDG) copurifies with thep68 RNA helicase (p68) as well as small RNAs, withina complex isolated from chicken embryos, whichshows a 5-methylcytosine DNA glycosylase activity(16–19). This enzyme is involved in DNA repair pro-cessed through the base excision repair (BER) mech-anism. In BER, TDG first recognizes U:G or T:G mis-matches generated by spontaneous deamination ofcytosines and methylcytosines, respectively. T or Ubases are then flipped out of the DNA molecule andcleaved by TDG, generating abasic sites [apurinic/apyrimidic (AP)] that are then repaired by the sequen-tial recruitment of an AP endonuclease, DNA polymer-ase �, and a DNA ligase (20). From the work of Zhu etal. (19), it is postulated that TDG can flip out 5MeC andcut the modified base. Nonetheless, 5MeC still estab-lishes normal pairing with G and is not likely to distortDNA (21, 22). This could explain the very low efficiencyof the reaction when compared with T or U removalfrom mismatches (19).
TDG has been shown to interact with a number oftranscription factors, and especially with nuclear re-ceptors (23–28). The physical association of retinoicacid receptor or estrogen receptors (ER) with TDGresults in higher transcriptional activation of reportergenes; and, at least for ER, such transcriptional coac-tivation does not require a functional glycosylase cat-alytic domain (23). On the other hand, Jost et al. (25)reported that the interaction of ER with TDG increasesthe 5-methylcytosine DNA glycosylase activity of theenzyme. These studies indicate that TDG is a multi-functional protein that could be involved in both DNArepair and transcriptional regulation. Correspondingly,the fact that the histone acetyl transferase CBP caninteract with and acetylate TDG is likely to reflect amolecular switch regulating the functional commit-ment of the glycosylase. Acetylation of TDG inhibits itsassociation with the AP endonuclease, suggestingthat it could decrease TDG repair activity (26). Anothermolecular switch could be triggered by the sumoyla-tion of TDG because this posttranslational modifica-tion dramatically reduces TDG binding to DNA andabrogates G:T mismatch processing (29). However,the processes regulating TDG sumoylation are yet un-known and it is still unclear whether sumoylated TDGretains its transcriptional activities.
Here, we investigated the recruitment of variousfactors on the vitronectin gene during activation by thenuclear orphan receptor chicken ovalbumin upstreampromoter-transcription factor I (COUP-TFI) in P19 em-bryonal carcinoma cells (30). We found that, besidesthe increased level of TDG association upon transcrip-tion activation, COUP-TFI binding to the promoter en-hanced the recruitment of Dnmt3a, Dnmt3b, and p68.These changes in protein loading of the vitronectingene occur while the promoter becomes undermethy-lated and the gene is actively transcribed. Our datasuggest that Dnmt3a/b, p68 and TDG may be involvedin the dynamic process of transcription activation.
RESULTS
The Endogenous Vitronectin Gene PromoterBecomes Hypomethylated When Activated byCOUP-TFI
To investigate the mechanisms involved in COUP-TFI-mediated transcriptional activation of the vitro-nectin gene, we generated P19 cell lines expressingCOUP-TFI tagged with the hemagglutinin (HA)epitope. Stable transfectants were selected forG418 resistance and for their ability to express de-tectable levels of HA::COUP-TFI fusion protein. Thetranscription of the transgene is under the controlof the cytomegalovirus (CMV) promoter that canbe super-activated when cells are treated with 8-bromo-cAMP (8-Br-cAMP). This provides the oppor-tunity to control the levels of HA::COUP-TFI in P19cells (Fig. 1A), and thus to avoid the retinoic acidtreatment which is used to induce endogenousCOUP-TFI gene expression (31) to a similar level(30). When cells were exposed to 8-Br-cAMP, aninduction of the transgene was detected after 4 h,followed by the induction of the vitronectin geneafter 10 h, whereas PO expression remained con-stant (Fig. 1B). Neither COUP-TFI nor vitronectinendogenous genes were induced in the control celllines when treated with 8-Br-cAMP (data notshown). To determine the potential variations in thelevel of CpG methylation during COUP-TFI-inducedactivation, we ran bisulfite sequencing of the prox-imal promoter (32). This method allows the precisedetermination of the methylation status of all CpGspresent in a given genomic DNA fragment. We per-formed these analysis on genomic DNA extractedfrom control or 8-Br-cAMP-treated cells expressingCOUP-TFI for 48 h. As a control, similar treatmentswere performed on pCR3.1 P19 cells. The results ofthese experiments, summarized in Fig. 1C, showed:1) that the vitronectin gene promoter is hypermethy-lated in undifferentiated P19 cells, correlating withits transcriptionally silent state; 2) that 8-Br-cAMPon its own does not modify CpG methylation levels;and 3) that COUP-TFI expression is correlated to adecrease in CpG methylation levels (Fig. 1C). Fur-thermore, when comparing the strand-specificmethylation patterns, it was apparent that hypom-ethylation occurred on the transcribed (bottom)strand, with the coding (top) strand remaininglargely unaffected. When comparing �/� 8-Br-cAMP situations in a two-sided t test, methylationvaried significantly only in the case of the bottomstrand in HA::COUP-TFI expressing cells (Fig. 1C). Evenif both sequenced strands did not originate from thesame initial DNA molecules, these data suggest a bias instrand methylation. In summary, the vitronectin geneproximal promoter is hypermethylated when silent, andCpGs become hypomethylated when transcriptionoccurs.
2086 Mol Endocrinol, September 2007, 21(9):2085–2098 Gallais et al. • Dnmt3 and COUP-TFI Activate Transcription

kDa
A
7550
8-Br-cAMP (24 h) - + - +
pCR 3.1COUP-TFI/HA
HA::COUP-TFI
8-Br-cAMP (h) 0 4 10 24 34 48
Vitronectin
COUP-TFI
PO
B
C
PCR3.1(P = 1)
COUP-TFI/HA (P = 0.83)
8-Br-cAMP
+-
PCR3.1(P = 0.13)
COUP-TFI/HA (P = 0.02)
8-Br-cAMP
+-
Vitronectin
+1
- 600 - 500 - 400 - 300 - 200 - 100
+1
- 600 - 500 - 400 - 300 - 200 - 100
Vitronectin
Fig. 1. COUP-TFI Induces Hypomethylation of the Vitronectin Gene Promoter DNAA, Western blot analysis of transgene expression with HA antibody. High levels of COUP-TFI protein were detected in
HA::COUP-TFI expressing P19 cells treated with 1 mM 8-Br-cAMP for 24 h, whereas low but detectable levels were observed inthe absence of 8-Br-cAMP. B, Semiquantitative RT-PCR analysis of COUP-TFI and vitronectin expression in stable cell linestreated with 1 mM 8-Br-cAMP. PO is a ribosomal phosphoprotein gene used as a control. C, DNA was prepared from stable P19cells treated with 1 mM 8-Br-cAMP for 48 h, and the DNA methylation status of individual CpG sites was determined bysequencing sodium bisulfite-modified DNA. The positions of each CpG, as well as COUP-TFI binding sites, are indicated on thescheme by black dots and gray rectangles, respectively. For each condition, strand-specific PCR products were subcloned, and10–11 clones were subjected to nucleotide sequence analysis. For each clone, the methylation status is depicted by black andwhite squares indicating methylated and unmethylated CpGs respectively. Squares above the scheme show methylation statusof the top strand (5�–3�), whereas bottom strand (3�–5�) methylation is depicted below the scheme. To determine the statisticalsignificance of the methylation changes observed, the total methylation at all CpGs was calculated for each clone. Thedistributions obtained for the upper and lower strand in absence and presence of 8-Br-cAMP were compared using a two-sidedt test. P values are indicated in parentheses.
Gallais et al. • Dnmt3 and COUP-TFI Activate Transcription Mol Endocrinol, September 2007, 21(9):2085–2098 2087

COUP-TFI Expression Increases TDG, p68, andDnmt3a/b Mobilization onto the Vitronectin GenePromoter in Vivo
TDG DNA glycosylase has been suggested to be partof a demethylation complex together with the p68 RNAhelicase (p68), and is known to interact with nuclearreceptors (16–19, 23). Because we observed a loss ofDNA methylation during activation of the vitronectingene, a COUP-TFI-mediated loading of the TDG onto
the vitronectin promoter was tested in vivo by chro-matin immunoprecipitation (ChIP) assays using chro-matin extracts of P19 cells expressing COUP-TFI ornot, after a 24 h 8-Br-cAMP treatment. Oligonucleo-tides were designed to amplify the proximal promoter�252/�53 fragment of the vitronectin gene and theproximal promoter �279/�84 region of the PO controlgene (Fig. 2A). First, we showed that the activation ofthe vitronectin gene by COUP-TFI correlates with thebinding of the orphan receptor to the promoter in vivo
Vitronectin PO
pCR3.1
COUP-TFI/H
A
α Dnmt1
α Dnmt2
pCR3.1
COUP-TFI/H
A
α Dnmt3a
α Flag
5
10
15
20
25Vitronectin
% O
f Inp
uts
% O
f Inp
uts 0
PO
α Dnmt1
α Dnmt2
α Dnmt3a
α Flag
α Dnmt3b
-252 -53
+1NR2 NR1
C
Vitronectin
A
B
pCR3.1
COUP-TFI/H
A
Inputs (15%)
α COUP-TFI
α P-RNAPol II
α diMeK9 H3
α HA
α HP1
α TDG
α Flag
Vitronectin PO
pCR3.1
COUP-TFI/H
A
0
10
20
30
40
50pCR3.1
COUP-TFI/HA
Vitronectin
PO
α COUP-T
FI
α P-RNA Pol
II
α diMeK
9 H3
α HA
α HP1
α TDGα Flag
% O
f Inp
uts
% O
f Inp
uts
D E
-279 -84
+1
PO
α Dnmt3b
0
10
20
30
40
pCR3.1
COUP-TFI/HA
5
0
Inputs (15%)
Fig. 2. Expression of COUP-TFI Favors TDG and Dnmt3 Loading on the Vitronectin Promoter in VivoChIP experiments performed using chromatin prepared from P19 stable cell lines expressing or not HA::COUP-TFI and treated
for 24 h with 1 mM 8-Br-cAMP. A, Schematic representation of the PO and vitronectin gene promoters indicating COUP-TFIbinding sites (NR1 and NR2, black boxes) as well as the positions of the oligonucleotides used for PCR and QPCR (open arrows).B and D, The recruitment of COUP-TFI (�COUP-TFI and �HA), phosphorylated RNA Pol II (�P-RNA Pol II), dimethyl histone H3(�diMeK9 H3), Heterochromatin Protein 1 � (�HP1�), TDG (�TDG), Dnmt1 (�Dnmt1), Dnmt2 (�Dnmt2), Dnmt3a (�Dnmt3a), andDnmt3b (�Dnmt3b) to the vitronectin and control PO gene promoters was monitored through immunoprecipitation with theindicated antibodies after cross-linking of proteins to DNA, and subsequent amplification of the vitronectin gene �252/�53fragment and PO gene �279/�84 fragment by PCR. C and E, Quantitative analysis of the ChIP signal through real-time PCR.Results are expressed as a percentage of the signal obtained with the input chromatin DNA.
2088 Mol Endocrinol, September 2007, 21(9):2085–2098 Gallais et al. • Dnmt3 and COUP-TFI Activate Transcription

(Fig. 2, B and C). Additionally, this binding correlateswith the detection of an activated RNA Pol II ChIPsignal (phospho-polymerase, P-RNA Pol II), as well aswith the loss of the dimethylated K9 histone H3 signaland the release of Heterochromatin Protein 1� fromthe promoter, two marks of heterochromatin (Fig. 2, Band C). Finally, although already present in the ab-sence of COUP-TFI, the amount of TDG recruited tothe vitronectin promoter was clearly enhanced whenCOUP-TFI was expressed (Fig. 2, B and C). Thesedata suggest the existence of a functional relationshipbetween the nuclear receptor and the glycosylase.Conversely, the active PO gene, which transcription isnor differentiation dependent nor COUP-TFI depen-dent, did not show variations in the recruitment ofthese various proteins upon COUP-TFI expression(Fig. 2, B and C). Because we had shown that CpGmethylation decreased during the activation of thevitronectin gene, we also examined the presence ofDnmt1, Dnmt2, Dnmt3a, and Dnmt3b on the vitronec-tin promoter by ChIP assays, and found that Dnmt3bwas present on the silent gene (Fig. 2, D and E).Importantly, the recruitment of Dnmt3a and Dnmt3bwas enhanced when the promoter was active (Fig. 2, Dand E). Through sequential ChIP (Re-ChIP) experiments,
we then analyzed the simultaneous presence of COUP-TFI, TDG, p68, the activated RNA Pol II, Dnmt3a, andDnmt3b on the vitronectin promoter. TDG was not foundassociated with COUP-TFI in Re-ChIP assays, suggest-ing that these two proteins are not simultaneouslypresent in DNA-bound complexes (Fig. 3). Nonetheless,Re-ChIP experiments demonstrated that p68 was re-cruited to the activated vitronectin promoter and found inboth COUP-TFI- and TDG-bound chromatin fractions(Fig. 3A). Interestingly, p68 was not found associatedwith the vitronectin promoter in the TDG-bound fractionof chromatin when the gene was not transcribed, sug-gesting that p68 association with TDG-bound chromatincould be dependent on transcription, and that COUP-TFIcould mediate the assembly of a p68/TDG complex onthe active gene. Re-ChIP experiments further suggestedthat Dnmt3a and Dnmt3b could be associated to bothCOUP-TFI and TDG on transcriptionally active chroma-tin, and that TDG and Dnmt3 are found on inactive chro-matin as well.
TDG, p68, and Dnmt3a Interact with COUP-TFI
Because TDG has been shown to interact with allnuclear receptors tested to date, we examined
α Dnmt3a
α Flag
pCR3.1
COUP-TFI/H
A
α HA
α TDG
α HA
Input
α TDG
α p68
α Flag
Re-ChIP
α P-RNA Pol II
Vitronectin
PO
α Dnmt3b
pCR3.1
COUP-TFI/H
A
α HA
α TDG
α HA
Input
α TDG
Re-ChIP
Vitronectin
PO
Fig. 3. p68 and Dnmts Associate with TDG or COUP-TFI as Separated Complexes onto Activated Vitronectin Gene PromoterChIP experiments were performed using chromatin prepared from P19 stable cell lines expressing or not HA::COUP-TFI and treated
for 24 h with 1 mM 8-Br-cAMP. A second immunoprecipitation (Re-ChIP) was then run on HA or TDG immunoprecipitates with theindicated antibodies before amplification of the vitronectin and PO gene promoters. �Flag antibody was used as a negative control.Note that COUP-TFI and TDG are not simultaneously found on transcriptionally active chromatin. Pol II, Polymerase II.
Gallais et al. • Dnmt3 and COUP-TFI Activate Transcription Mol Endocrinol, September 2007, 21(9):2085–2098 2089

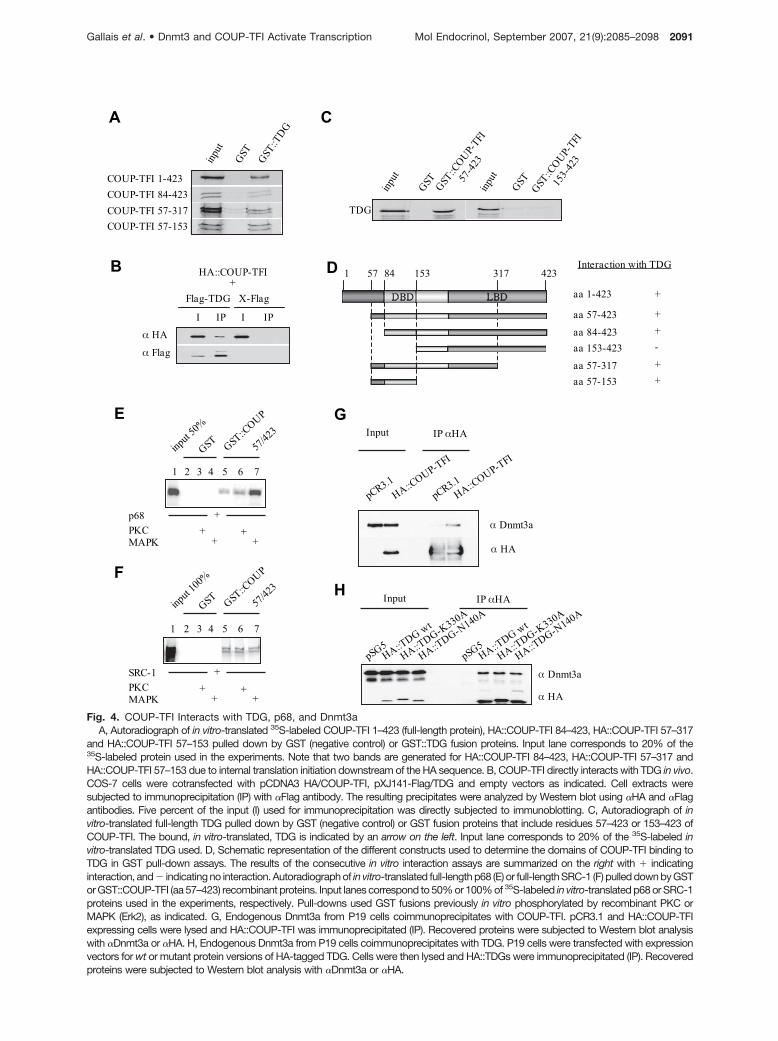
whether it could also interact with COUP-TFI. Even ifRe-ChIP data showed the exclusive engagement ofCOUP-TFI and TDG at the vitronectin promoter, suchan interaction could be involved in the observed in-creased occupancy of the vitronectin promoter byTDG upon COUP-TFI expression. This hypothesis wasfirst stressed by glutathione S-transferase (GST) pull-down experiments that demonstrated that full-lengthin vitro-translated COUP-TFI interacts with a TDG pro-duced as a GST fusion protein (Fig. 4A). This interactionwas also observed in vivo. Indeed, a Flag::TDG fusionprotein coimmunoprecipitated with HA::COUP-TFI whencoexpressed in COS-7 cells (Fig. 4B). To delineate thedomains of COUP-TFI interacting with TDG, several invitro-translated truncated COUP-TFI proteins were pro-duced and incubated with recombinant GST::TDG (Fig.4A). Reverse experiments using in vitro-translated TDGand GST::COUP-TFI fusion proteins were also run (Fig.4C). In accordance with the data shown Fig. 4C, in vitro-translated COUP-TFI 153–423 and bacterially expressedTDG did not interact in pull-down assays (data notshown). These assays determined that the DNA bindingdomain of COUP-TFI, formed by residues 84–153, inter-acts with TDG (Fig. 4D).
We also assayed the occurrence of an interactionbetween COUP-TFI and p68 in vitro by GST pull-downassays. These experiments evidenced the interactionbetween these two proteins (Fig. 4E). Moreover, whenrecombinant GST::COUP-TFI protein was phosphory-lated in vitro by recombinant Erk2, we observeda marked increase in its ability to interact with p68.In contrast, the in vitro phosphorylation ofGST::COUP-TFI by protein kinase C (PKC) had noeffect (Fig. 4E). As a control, both kinases did notinfluence the level of association between COUP-TFIand SRC-1 (Fig. 4F). Interestingly, we have previouslydemonstrated that MAPKs positively regulate COUP-TFI activity (33). Such a regulation could thus be linkedto the enhanced recruitment of p68 by phosphorylatedCOUP-TFI, as shown in the case of the ER�/p68 in-teraction (34). When HA-tagged COUP-TFI, TDG, TDGN140A (glycosylase defective; Ref. 35), and TDGK330A (sumoylation defective; Ref. 29) were used tofish out endogenous Dnmt3a from P19 cells by coim-muprecipitation assays, we found that Dnmt3a couldinteract with the orphan receptor as well as with thewild-type (wt) and mutated glycosylase (Fig. 4, G andH). Furthermore, Flag-Dnmt3b coimmunoprecipitatedwith COUP-TFI and TDG (data not shown). Togetherwith our Re-ChIP experiments, these interaction datasupport the hypothesis that COUP-TFI/p68, COUP-TFI/Dnmt3a-3b, TDG/p68 and TDG/Dnmt3a-3b, couldbe assembled as complexes on chromatin.
Next, we investigated the output of COUP-TFI/TDGinteraction in terms of DNA recognition. Indeed, thefact that TDG interacts with the orphan receptorthrough its DNA binding domain suggested that sucha complex would not be able to bind DNA. We first ranEMSA with probes containing one COUP-TFI bindingsite from the vitronectin promoter, directly adjacent to
a CpG (position �164, relative to the transcription startsite) found to be heavily methylated in the absence ofCOUP-TFI and methylated to a much lower extentfollowing the expression of COUP-TFI (see Fig. 1).Three probes were used: one with a hemimethylatedCpG (5MeC), one with a T:G mismatch in the CpG(with the T in the upper strand), and one with nomodification (Control). These experiments used re-combinant full-length TDG and COUP-TFI [amino ac-ids (aa) 57–423] purified to homogeneity. The assaysshowed that COUP-TFI binds equally well to the threeprobes, whereas TDG was better retained on the mis-match-containing probe (Fig. 5). The high stability ofTDG on a mismatch probe has already been describedand is explained by the fact that TDG cleaves themismatch oligonucleotide in conditions used forEMSA and establishes a very strong interaction withthe abasic site produced by the reaction (35). Whenboth proteins were coincubated with the probes, nosuper-retarded complex appeared, suggesting thatTDG may not interact with COUP-TFI when bound toDNA. This was further suggested by the exclusion ofTDG from the mismatch probe observed when in-creasing the amounts of COUP-TFI (Fig. 5). In accor-dance with Re-ChIP data, these experiments suggestthat TDG and COUP-TFI independently bind to DNA,with their respective binding likely exclusive. Thesedata also indicate that a preformed COUP-TFI/TDGcomplex may be disrupted by DNA binding of one orboth partners. Then it could be hypothesized that suchcomplexes may allow the glycosylase to load and scanCOUP-TFI targeted promoters.
Dnmt3 Enhance COUP-TFI Activation of aMethylated Reporter Gene
To examine the functional significance of these variouscomplexes, we next run glycosylase assays as well ascotransfection experiments. In glycosylase assays,TDG, either alone or combined with p68, was unableto cleave DNA at MeCs, suggesting the requirementfor additional factors (data not shown). These could beeither RNA cofactors (16), or proteins that woulddeaminate 5MeC in the context of CpGs, providingT:G mismatches to TDG for repair. Importantly, Dnmtsbeing able to catalyze deamination of CpGs (36–39),their recruitment to methylated promoters during tran-scriptional activation could participate to active de-methylation processes. This hypothesis was furthersubstantiated in another experimental system, inwhich Dnmt3a exhibited cytosine and 5MeC deamina-tion ability (Metivier, R., R. Gallais, C. Tiffoche, C.Le Peron, R. Jurkowska, R. P. Carmouche, D. Ibberson,P. Barath, F. Demay, G. Reid, V. Benes, A. Jeltsch, F.Gannon, and G. Salbert, manuscript in preparation).
Cotransfection experiments were run in P19 cellswith in vitro methylated pVitro-Luc, as well as expres-sion vectors for COUP-TFI, and various Dnmts. Wefirst show that, whereas Flag-Dnmt1 and Myc-Dnmt3bhad no effect on transcription of the mock-methylated
2090 Mol Endocrinol, September 2007, 21(9):2085–2098 Gallais et al. • Dnmt3 and COUP-TFI Activate Transcription
Interaction with TDG
+
+
+
+
-
+
aa 84-423
aa 1-423
aa 57-423
aa 57-317
aa 153-423
aa 57-153
423
DBD
841 57 153 317
LBD
COUP-TFI 1-423
COUP-TFI 84-423
COUP-TFI 57-317
COUP-TFI 57-153
GST
::TD
G
inpu
t
GST
CA
B
I IP I IP
HA
Flag
Flag-TDG X-Flag
HA::COUP-TFI+
D
E
input
50%
GST GST::C
OUP
57/42
3
+ +PKC + +MAPK
p68 +
1 2 3 4 5 6 7
F
+ +PKC + +MAPK
SRC-1 +
input
100%
GST GST::C
OUP
57/42
3
1 2 3 4 5 6 7
Dnmt3a
HA
pCR3.1
HA::COUP-T
FI
pCR3.1HA::C
OUP-TFI
Input IP HA
pSG5HA::T
DG wt
HA::TDG-K
330A
HA::TDG-N
140A
Input IP HA
Dnmt3a
HA
pSG5HA::T
DG wt
HA::TDG-K
330A
HA::TDG-N
140A
G
H
inpu
t
GST GST
::CO
UP-
TFI
57-4
23
TDG
inpu
t
GST
GST
::CO
UP-
TFI
153-
423
Fig. 4. COUP-TFI Interacts with TDG, p68, and Dnmt3aA, Autoradiograph of in vitro-translated 35S-labeled COUP-TFI 1–423 (full-length protein), HA::COUP-TFI 84–423, HA::COUP-TFI 57–317
and HA::COUP-TFI 57–153 pulled down by GST (negative control) or GST::TDG fusion proteins. Input lane corresponds to 20% of the35S-labeled protein used in the experiments. Note that two bands are generated for HA::COUP-TFI 84–423, HA::COUP-TFI 57–317 andHA::COUP-TFI 57–153 due to internal translation initiation downstream of the HA sequence. B, COUP-TFI directly interacts with TDG in vivo.COS-7 cells were cotransfected with pCDNA3 HA/COUP-TFI, pXJ141-Flag/TDG and empty vectors as indicated. Cell extracts weresubjected to immunoprecipitation (IP) with �Flag antibody. The resulting precipitates were analyzed by Western blot using �HA and �Flagantibodies. Five percent of the input (I) used for immunoprecipitation was directly subjected to immunoblotting. C, Autoradiograph of invitro-translated full-length TDG pulled down by GST (negative control) or GST fusion proteins that include residues 57–423 or 153–423 ofCOUP-TFI. The bound, in vitro-translated, TDG is indicated by an arrow on the left. Input lane corresponds to 20% of the 35S-labeled invitro-translated TDG used. D, Schematic representation of the different constructs used to determine the domains of COUP-TFI binding toTDG in GST pull-down assays. The results of the consecutive in vitro interaction assays are summarized on the right with � indicatinginteraction, and � indicating no interaction. Autoradiograph of in vitro-translated full-length p68 (E) or full-length SRC-1 (F) pulled down by GSTor GST::COUP-TFI (aa 57–423) recombinant proteins. Input lanes correspond to 50% or 100% of 35S-labeled in vitro-translated p68 or SRC-1proteins used in the experiments, respectively. Pull-downs used GST fusions previously in vitro phosphorylated by recombinant PKC orMAPK (Erk2), as indicated. G, Endogenous Dnmt3a from P19 cells coimmunoprecipitates with COUP-TFI. pCR3.1 and HA::COUP-TFIexpressing cells were lysed and HA::COUP-TFI was immunoprecipitated (IP). Recovered proteins were subjected to Western blot analysiswith �Dnmt3a or �HA. H, Endogenous Dnmt3a from P19 cells coimmunoprecipitates with TDG. P19 cells were transfected with expressionvectors for wt or mutant protein versions of HA-tagged TDG. Cells were then lysed and HA::TDGs were immunoprecipitated (IP). Recoveredproteins were subjected to Western blot analysis with �Dnmt3a or �HA.
Gallais et al. • Dnmt3 and COUP-TFI Activate Transcription Mol Endocrinol, September 2007, 21(9):2085–2098 2091

and methylated reporter genes, Myc-Dnmt3b couldspecifically enhance COUP-TFI-mediated transcrip-tional activation of the methylated reporter gene (Fig.6A). We then compared the ability of Flag-Dnmt3a and3b, and Myc-Dnmt3b to regulate transcription viaCOUP-TFI and found that all tagged Dnmts couldenhance transcription but that Flag-Dnmt3b and Myc-Dnmt3b were more potent than Flag-Dnmt3a at highdosage (Fig. 6B). Surprisingly, when combined withTDG, Dnmt3b did not coactivate transcription of themethylated reporter gene in the presence of COUP-TFI (Fig. 6C). Interestingly, the glycosylase-defectiveN140A mutant TDG allowed Dnmt3b coactivation ofCOUP-TFI, albeit to a lower level than when no glyco-sylase was added. Conversely, the sumoylation-de-fective K330 mutant was still able to fully repressCOUP-TFI/Dnmt3b-mediated transcription. Finally, inthese conditions, nor Dnmt3b nor wt or mutated TDGcould modify COUP-TFI-mediated transactivation ofthe mock-methylated reporter gene (data not shown).Collectively, these transfection experiments suggestthat, when targeted by COUP-TFI, Dnmt3 can en-hance the transcription of a methylated reporter gene;and that TDG can counteract this effect, either throughtitration of the methyltransferases, or through a repairactivity requiring the glycosylase function, or both.
DISCUSSION
In the present report, we have demonstrated the oc-currence of physical and/or functional interactions be-
tween the orphan nuclear receptor COUP-TFI, the gly-cosylase TDG, the p68 RNA helicase, and the DNAmethyltransferases Dnmt3a-3b. Importantly, ChIP andRe-ChIP data support the presence of a TDG/Dnmt3a-3b complex on untranscribed chromatin (inP19/pCR3.1 cells). Such a complex may participate inheterochromatin maintenance, being involved in therepair of DNA mismatches generated by the sponta-neous deamination of cytosines and methylcytosinesin CpGs, and their subsequent (re-)methylation (Fig. 7).Interestingly, TDG was able to suppress the coactiva-tion function of Dnmt3b on COUP-TFI in cotransfec-tion experiments. This antagonistic behavior does notsupport a coordinated role of TDG, Dnmt3b andCOUP-TFI in the transcriptional activation of the vitro-nectin gene but rather suggests that TDG/Dnmt3bcould be required for repair and methylation of tran-scription-induced mismatches. Indeed, this is sup-ported by our cotransfection data showing that theglycosylase-defective TDG was less effective thanwtTDG in suppressing coactivation by Dnmt3b. Ac-cordingly, the COUP-TFI/Dnmt3 coactivation processwould involve the generation of mismatches, in aDnmt3-dependent manner. Such a hypothesis is com-patible with the known ability of bacterial Dnmts tocatalyze the deamination of cytosines (39).
Although they are also recruited on COUP-TFI-acti-vated vitronectin gene promoter, these factors are in-cluded within exclusive TDG/Dnmt3a-3b and COUP-TFI/Dnmt3a-3b complexes that may also contain p68, asindicated by our Re-ChIP assays. Given that our Re-ChIP experiments used unsynchronized cells stochasti-cally transcribing the vitronectin gene to different ex-tents, the recruitment of the TDG/Dnmt3a-3b complexmay concern only a subpopulation of COUP-TFI/HA-expressing cells treated with 8-Br-cAMP. This subpopu-lation would not, at the time of the experiment, expressthe vitronectin gene which promoter would therefore in-clude the TDG/Dnmt3a-3b complex. Accordingly, theTDG/Dnmt3b complex may favor heterochromatin for-mation, with the COUP-TFI/Dnmt3b complex converselyfavoring euchromatin formation. However, our Re-ChIPexperiments suggest that TDG could be complexed toDnmt3a on active chromatin, whereas TDG/Dnmt3b as-sociation with chromatin does not seem to depend ontranscription. This may add further complexity to themodel presented within Fig. 7 because this observationmay imply that Dnmt3a/TDG or Dnmt3b/TDG could co-exist in different complexes having divergent outputs.Additionally, the exclusive recruitment of the TDG/Dnmt3a-3b and COUP-TFI/Dnmt3a-3b complexes maysuggest that COUP-TFI could serve as a platform for theassembly or recruitment of transcription regulatory com-plexes that would be released once formed. Transcrip-tion of the vitronectin gene may thus be associated withthe occurrence of transient and exclusive states leadingto timely separated populations of chromatin-boundcomplexes in single cells. As described in another modelof transcription initiation, a transcriptional ratchet maycontrol the occurrence of this sequence of events (40). In
Control Mismatch 5MeC Mismatch
*
COUP-TFI + + + + + + TDG + + + + + + + + + + +
Fig. 5. COUP-TFI Binding to DNA Decreases TDG/DNA As-sociation
The interactions between COUP-TFI, TDG, and DNA were an-alyzed by EMSAs using 32P-labeled probes corresponding to a36-bp fragment of the vitronectin promoter containing one bindingsite for COUP-TFI directly adjacent to a CpG dinucleotide (seeMaterials and Methods). The Mismatch oligonucleotide contains aT:G mismatch, the 5MeC oligonucleotide is hemimethylated on theCpG, and the control oligonucleotide contains neither methylatedCpG nor mismatch. Probes were incubated with 1.25, 2.5, 5, or 10ng of purified recombinant COUP-TFI and 500 ng of purified re-combinant TDG. Protein-DNA complexes and free oligonucleo-tides were separated on a 6% polyacrylamide gel. The asterisk andthe arrow indicate the positions of the probe bound to COUP-TFIand TDG, respectively.
2092 Mol Endocrinol, September 2007, 21(9):2085–2098 Gallais et al. • Dnmt3 and COUP-TFI Activate Transcription

the case depicted in this report, different functions of theTDG/Dnmt3 complexes could be regulated dependingon protein partners and/or posttranslational modifica-tions that would occur during transcription. Importantly,
the presence of the p68 RNA helicase in the TDG/Dnmt3complex when associated with active chromatin wouldbe an important step toward the acquisition of new func-tional properties of the complex. Finally, whatever the
A
B
0
100
200
300
400
500
RL
U
COUP-TFI (20 ng) - + - - + + - + - - + +Dnmt1 (40 ng) + +- - - - + +- - - -
M-Dnmt3b (40 ng) + +- - - - + +- - - -
MockpVitro-Luc
MethylatedpVitro-Luc
C
0
2
4
6
8
Fold
indu
ctio
n
Methylated pVitro-Luc
COUP-TFI (20 ng)F-Dnmt3a (10-40 ng)F-Dnmt3b (10-40 ng)
M-Dnmt3b (10-40 ng)
+- +-
+- - - +
--
- - - --- -
- --
-- -- -
Fold
indu
ctio
n
0
1
2
3
4Methylated pVitro-Luc
COUP-TFI (20 ng) ++- - ++- - ++- - ++- -TDG (20 ng) - - - - ++ ++ - - - - - - - -
TDG K330A (20 ng) - - - - ++ +- - - - - - - -TDG N140A (20 ng) - - - - ++ ++- - - - - - - -
M-Dnmt3b (10-40 ng) - - - - - - - -
*
**
*
+
Fig. 6. Dnmt3a Stimulates COUP-TFI-Mediated Activation of a Methylated Reporter GeneA and B, P19 cells were transfected with a mock-treated or in vitro-methylated reporter gene containing the proximal promoter
of the mouse vitronectin gene coupled to the luciferase coding sequence (note that methylation of the reporter gene resulted ina lower basal activity). Expression vectors for COUP-TFI and Dnmts were cotransfected as indicated (M- and F- mean Myc andFlag, respectively). Amounts of transfected Dnmt expression vectors were 10, 20, or 40 ng/well. C, Similar experiments were runin the additional presence of wt or mutant TDG expression vectors. The N140A mutant is glycosylase defective, whereas theK330A mutant cannot be sumoylated. Graphs show the mean � SEM (n � 6–9 depending on experiments) of data obtained in twoto three separate experiments. Asterisks indicate statistical differences between COUP-TFI (dark gray bars) and COUP-TFI �Dnmt3b (light gray bars) conditions, in the presence or absence of wt and mutant TDG (ANOVA Fisher’s test, P � 0.05). Note thatin B and C the data are shown as fold induction, whereas in A data are shown as relative luciferase units (RLU).
Gallais et al. • Dnmt3 and COUP-TFI Activate Transcription Mol Endocrinol, September 2007, 21(9):2085–2098 2093

mechanism involved is, which would require further at-tention, a true antagonistic behavior of the above-de-scribed complexes would nevertheless largely dependon their local respective concentrations at the vitronectingene locus.
Correlated with the recruitment of these complexes,CpGs from the vitronectin promoter become hypom-ethylated. Because the above-mentioned proteinshave been suggested to directly or indirectly regulateCpG methylation, their association with the vitronectinpromoter is likely to regulate its methylation status.TDG is supposed to be part of an active demethylationcomplex recruited through a direct interaction withER� (25) and suggested to be required for the hor-monal control of the transcription of a methylated tem-plate (28). Thus, TDG, through its glycosylase activity,could be involved in an active demethylation processof COUP-TFI target genes such as vitronectin. None-theless, we provide evidence that TDG is unable toinitiate 5MeCpG demethylation through a BER-typemechanism, at least in vitro. This directly contrastswith the view that TDG can cleave 5MeC within CpGsand suggests that the initiation of demethylation islikely to require additional components of a putativedemethylation complex. Such additional componentscould be specific 5MeC deaminases. Indeed deami-nation of cytosine or 5MeC leads to U:G or T:G mis-matches respectively, both being corrected by TDG.Being expressed in embryonic stem cells and able todeaminate 5MeC and C, it has been proposed thatcytidine deaminases such as activation-induceddeaminase and apolipoprotein B editing complex 1,could play a significant role in DNA demethylation bygenerating mismatches (41, 42). Although such a sce-nario is plausible, it would, however, provide an un-specific mechanism leading to the deamination andrepair of both methylated and unmethylated cytosines,including non-CpG cytosines. More specific and cost-less mechanisms targeting methylcytosine in CpGs
are then likely to have evolved, but the enzymes in-volved in these processes have not yet beencharacterized.
An appealing hypothesis is that the same systems thatregulate DNA methylation at CpGs would also be in-volved in active demethylation. In support of that hypoth-esis, bacterial DNA methyltransferases are known to ac-tively deaminate cytosine and 5MeC in experimentalconditions where the methyl donor concentration is low(39, 43–45). Even if such a reaction is not favored in vitroin the case of a purified mouse Dnmt1 (46), it couldhowever be triggered by the interaction of Dnmts withspecific factors regulating transcription, and would allowthe selective deamination of cytosines in the context ofCpGs. Such deaminated cytosines would then be re-paired through regular BER reactions. Our data supportthis hypothesis because 1) ChIP assays revealed thatDnmt3a and Dnmt3b are associated with the activatedvitronectin gene; 2) TDG and Dnmt3a-3b are found to-gether on chromatin, in support of a coordinated role forDnmt3 and TDG in the control of DNA (de)methylation;and 3) Dnmt3a is able to catalyze deamination of cyto-sine and 5MeC in vitro (Metivier, R., R. Gallais, C. Tif-foche, C. Le Peron, R. Jurkowska, R. P. Carmouche, D.Ibberson, P. Barath, F. Demay, G. Reid, V. Benes, A.Jeltsch, F. Gannon, and G. Salbert, manuscript in prep-aration). Furthermore, Li et al. (47) recently demonstratedthat TDG can inhibit Dnmt3a methylation activity. Then, itcan be proposed that, depending on regulatory mecha-nisms such as posttranslational modifications or throughprotein/protein interactions, Dnmt3 could alternate be-tween active and inactive states, with the active stateleading to cytosine methylation and the inactive state tocytosine deamination. This would provide sufficient flex-ibility for the occurrence of a dynamic CpG methylationprofile. This hypothesis will need further attention to for-mally demonstrate the involvement of Dnmt3a in 5MeCdeamination in vivo. Such a demonstration will, however,be hampered by the lack of appropriate tools, and will
HP1CH3 CH3CH3 CH3
Dnmt3
TDG
HP1HP1
Inactive chromatin
activatedPIC
p68Dnmt3
TDG
COUP-TFCH3CH3
Active chromatin
DNAdiMeK9(H3)
activatedPIC
CH3CH3
p68Dnmt3
TDG
Or
CH3hemimethylated CpG
Fig. 7. Schematic Representation of the Hypothetic Inactive vs. Active Chromatin States of the Vitronectin Gene ProximalPromoter
The inactive vitronectin promoter harbors heterochromatin epigenetic marks like diMeK9 of histone H3 and diMeCpGs. Thiscondensed chromatin is bound by HP1 as well as TDG probably complexed to Dnmt3b and Dnmt3a. This complex mayparticipate in the maintenance of CpG methylation state when spontaneous deamination of methylcytosines occurs. In the activeconfiguration, the vitronectin promoter is loaded with RNA Pol II and transcription regulatory complexes such as COUP-TFI/p68/Dnmt3 or TDG/p68/Dnmt3. These two complexes are not simultaneously present on chromatin. PIC, Preinitiation complex.
2094 Mol Endocrinol, September 2007, 21(9):2085–2098 Gallais et al. • Dnmt3 and COUP-TFI Activate Transcription

certainly require innovative experimental approaches aswell as specific drugs targeting Dnmts without beingincorporated into DNA.
Alternatively, rather than achieving cytosine deamina-tion, the recruitment of Dnmt3a to the active vitronectinpromoter may serve to maintain an asymmetric methyl-ation profile that could be a mark of transiently activechromatin. Indeed, our data suggest a strand-biasedmethylation profile of the active vitronectin promoter.This could reflect either an asymmetric demethylation oran asymmetric re-methylation of DNA. Strand-biasedmethylation has been described in a few cases such ascentromeric DNA in Arabidopsis (48), the active Dc8gene in carrot (49), and the embryonic �-globin gene inchicken (50). In the latter, the unmethylated active �-glo-bin gene becomes fully methylated during erythroid celldifferentiation, with the coding strand being targeted firstby methyltransferases. Similarly, we suggest here thatthe vitronectin promoter is the target of a strand-specificmethylation mechanism during transcriptional activation.Interestingly, activity of Dnmt3a exhibits sequence spec-ificity and strand asymmetry (51). However, examinationof the vitronectin gene promoter sequence did not allowto predict that one particular strand would be methylatedby Dnmt3a, suggesting that sequence-independent re-quirements may exist. It would be interesting to deter-mine how stable is the asymmetric DNA methylation ofthe vitronectin promoter, and to examine the methylationstatus of other differentially regulated genes, to under-stand whether strand-specific methylation could define aclass of active genes or is rather a transient mark ofremodeling chromatin.
MATERIALS AND METHODS
Plasmids
PCR3.1 was from Invitrogen (Carlsbad, CA). pSG5 fromStratagene (La Jolla, CA), pSG5-hTDGwt and hTDG N140Aand K330A mutants were from P. Schar (University of Basel,Basel, Switzerland). The pSG5-mTDG (aa 1–421) vector wasa gift from Prof. P. Chambon (Institut de Genetique et deBiologie Moleculaire et Cellulaire, Strasbourg, France). AnEcoRI-BamHI fragment of the pSG5-mTDG vector was sub-cloned in pBluescript II KS (pBS). Then the EcoRI-NotI frag-ment of pBS-mTDG was cloned in pGEX-6P1 (AmershamBiosciences, Arlington Heights, IL) and the BamHI-BamHIfragment of the resulting vector was subcloned in pXJ141-Flag (52). pVitro-Luc, pCDNA3 hCOUP-TFI and pGEX ex-pression vectors encoding GST fused to hCOUP-TFI 57–423,and hCOUP-TFI 153–423 have already been described (33).pCDNA3-based expression vector for HA::COUP-TFI57–153, HA::COUP-TFI 57–317, HA::COUP-TFI 84–423, andpCR3.1-HA-COUP-TFI 1–423 have been described (53).SRC-1 and p68 expression vectors were gift from B. W.O’Malley (Baylor College of Medicine, Houston, TX) and S.Kato (University of Tokyo, Tokyo, Japan) respectively. TheDnmt1 (54) and Myc-Dnmt3b (55) vectors were obtained fromS. Baylin (The Johns Hopkins University, Baltimore, MD) andE. Li (Novartis, Cambridge, MA), whereas Flag-Dnmt3a, andFlag-Dnmt3b (56) were gift from H. Leonhardt (Ludwig Maxi-milians University Munich, Munich, Germany).
Antibodies
Antibodies were purchased from Santa Cruz [HA-probe (Y11),�COUP-TFI (N-19), �Dnmt1 (H-300), �Dnmt2 (H-271), �Dnmt3a(H-295), �Dnmt3b (H-230)], Sigma (St. Louis, MO) (anti-Flag M2monoclonal antibody, anti-Flag M2 monoclonal antibody con-jugated to agarose beads), Upstate (Lake Placid, NY) [anti-dimethyl-histone H3 (Lys9), �phospho-RNA polymerase II(CTD4H8, lot no. 20463)], Euromedex (Souffelmeyershaim,France) [�HP-1� (2HP-1H5)]. The anti-TDG was prepared asdescribed below and �p68 is a gift from Dr. F. Fuller-Pace(University of Dundee, Dundee, Scotland, UK).
Cell Culture and Transfections
P19 embryonal carcinoma cells and COS-7 cells were grown asmonolayers in DMEM (Invitrogen) supplemented with 5% fetalcalf serum (Invitrogen). For transient transfection assays, P19cells were plated in 24-well plates and were transfected usingFuGENE 6 transfection reagent (Roche, Indianapolis, IN). Typi-cally, 150 ng of luciferase-based reporter plasmid containing theproximal promoter of the mouse vitronectin gene (pVitro-Luc),100 ng of �-galactosidase-based control plasmid, and 10–40ng of CMV or simian virus 40 promoter driven expression vec-tors were used. The amount of CMV and simian virus 40 pro-moters in each transfection mix was kept constant with emptypCR3.1 and pSG5 vectors. Cells were assayed for luciferaseand �-galactosidase activity 36 h after transfection. In vitromethylation of plasmid DNA, used 10 �g of pVitro-Luc plasmidincubated with SssI methyltransferase (New England Biolabs,Boston, MA) in a total volume of 40 �l at 37 C for 3 h with 160�M S-adenosylmethionine. The procedure for mock methylationwas identical but no enzyme was included. For stable transfec-tion, 5.105 P19 cells were plated in 100-mm dishes and trans-fected with 10 �g of expression vector. After 24 h, cells weregrown in the presence of 400 �g/ml of G418 (Sigma). After 10 dof selection, clones of resistant cells were isolated and propa-gated in the presence of G418.
Semiquantitative RT-PCR Analysis
Total RNAs from P19 cells were purified using the Trizolreagent (Life Technologies, Inc.). Two micrograms of RNAwere used as template for Moloney murine leukemia virusreverse transcriptase (Invitrogen) and Pd(N)6 random hexam-ers (Amersham Biosciences). For PCR, 1/20 of the reversetranscription reaction was used with 1 �M of the followingprimers (Proligo, Evry, France): AAGCACTACGGCCAAT-TCAC (COUP-TFI up), AGCTCGCAGATGTTCTCGAT(COUP-TFI down) [28 cycles]; CAGCTCTGGAGAAACTGCTG(PO up), GTGTACTCAGTCTCCACAGA (PO down) [27 cy-cles]; TACTATCAGAGCTGCTG-39 (Vitronectin up), AGTT-GATGCGAGTGAAG (Vitronectin down) [33 cycles].
GST Pull-Down Assays
Radiolabeled proteins were synthesized in vitro using the T7 RNApolymerase rabbit reticulocyte-coupled transcription/translation kit(Promega Corp., Madison, WI). GST fusion proteins were pro-duced in Escherichia coli BL21-CodonPlus RP (Stratagene). After12 h of induction with 0.5 mM isopropyl �-D-thiogalactoside, cellswere harvested by centrifugation, resuspended in PBS/1% Triton(TPBSI) containing a protease inhibitor cocktail (Roche) and thenlysed by sonication. Lysates were clarified by centrifugation andincubated overnight with a 50% suspension of glutathione-agar-ose beads (Sigma) in PBS. Washed fusion proteins bound to thebeads were resuspended in TPBSI and incubated for 2 h with 5 �lof [35S]methionine-labeled proteins in a total volume of 200 �l.Beads were washed ten times in 500 �l of TPBSI supplementedwith 100 mM NaCl. Washed beads were denatured at 95 C for 10min, and the supernatant was resolved on SDS-PAGE. For in vitro
Gallais et al. • Dnmt3 and COUP-TFI Activate Transcription Mol Endocrinol, September 2007, 21(9):2085–2098 2095

phosphorylation of the GST or GST::COUP-TFI fusion proteins, 25ng of recombinant Erk2 (Sigma) or PKC purified from rat brain(Sigma) were incubated in the presence of 100 to 200 ng of GSTfusion proteins bound to glutathione-agarose beads in 50 �l ofMAPK phosphorylation buffer [50 mM Tris-HCl (pH 8), 0.5 mM
EDTA, 25 mM MgCl2, 1 mM dithiothreitol (DTT), 250 �M ATP, 2%glycerol] or 50 �l of PKC phosphorylation buffer [20 mM HEPES (pH7.4) 10 mM MgCl2, 1.7 mM CaCl2, 600 �g/ml phosphatidylserine(Sigma); 1 mM DTT, and 250 �M ATP] for 30 min at 30 C, in thepresence of protease inhibitors (Roche) and phosphatase inhibitors(1 �M okadaic acid, 200 �M Na3VO4; Sigma). Effective phosphor-ylation was verified by running parallel reactions with 50 �M coldATP and 5 �Ci of [�-32P]ATP (MP Biomedicals, Irvine, CA). Beadswere then extensively washed before adding radiolabeled proteinsfor pull-down assays.
Expression and Purification of hCOUP-TFI and mTDG
All purification procedures were performed using AktaExplorer 10and chromatographic material supplied by Amersham Bio-sciences. COUP-TFI (aa 57–423) was expressed as a GST fusionprotein from pGEX-6P1 vector in E. coli strain BL21-Codon-Plus-RP (Stratagene). After induction by 0.1 mM isopropyl �-D-thiogalactoside for 4 h at 25 C, bacteria were harvested by cen-trifugation, washed in water, and the pellets were resuspended inlysis buffer [50 mM Tris-Cl (pH 7.5), 200 mM NaCl, 5 mM MgCl2, 5mM DTT, 5 mM 3-[3-(cholamidopropyl)dimethylammonio]-1-pro-panesulfonate (CHAPS), 5% glycerol] containing deoxyribonucle-ase I (10 U/ml) and a mix of protease inhibitors (Complete-EDTA;Roche). Bacteria were disrupted by French press and the lysatewas clarified by centrifugation (30 min; 25,000 � g). RecombinantCOUP-TFI was purified on Heparine Sepharose using PBS with700 mM NaCl supplement for elution. Eluate was applied toGSTrap 5-ml column and after extensive wash by PBS, dilutedPreScission Protease (10 U/ml in PBS supplemented with 2 mM
CHAPS and 1 mM DTT) was injected to the column. After removingthe GST tag by overnight on-column cleavage at 4 C, target proteinwas eluted and applied on Resource S 1-ml column. COUP-TFIwas eluted by salt gradient with peak at PBS with 1 mM CHAPScontaining 100 mM NaCl. Bacterial expression of mTDG (aa 1–421)and preparation of the lysate were performed using the sameprotocol as for COUP-TFI excluding CHAPS. Clarified lysate wasapplied to Heparine Sepharose FF 26/10 column and recombinantprotein was eluted with PBS supplemented with 700 mM NaCl.GST::TDG fusion protein was further retained on GSTprep 16/10column and after extensive washings by PBS, diluted PrescissionProtease [10 U/ml in 20 mM Tris-Cl (pH 8.8), 50 mM NaCl, 1 mM
EDTA, 1 mM DTT] was injected to the column. After the removal ofthe GST tag by overnight on-column cleavage at 4 C, the proteinsample was then eluted and applied on Resource Q 6ml columnand resolved by the salt gradient (100–200 mM NaCl) in 20 mM
Tris-Cl (pH 8.5). Fractions containing TDG were concentrated andseparated by gel filtration (Superdex 200 HR 16/60; GE Healthcare,Orsay, France) in 20 mM Tris-Cl (pH 8.0), 150 mM NaCl, 1 mM EDTA,1 mM DTT. The purified protein was used for immunization ofrabbits. The collected polyclonal antibody was affinity purified be-fore use in ChIP assays.
EMSAs
The following oligonucleotides NR2 Up 5�-ATCCCTCCTGC-CCGTGACCCTATGACTTTGGTCTTG-3�, MeC Up 5�-ATCCCTC-CTGCC5MeCGTGACCCTATGACTTTGGTCTTG-3�, andMismatch Up 5�-ATCCCTCCTGCCTGTGACCCTATGACT-TTGGTCTTG-3�, were 5�-labeled with T4 polynucleotidekinase and [�-32P]ATP before annealing with thecomplementary strand NR2 Down 5�-CAAGACCAAAGTC-ATAGGGTCACGGGCAGGAGGGAT-3�. Purified proteins(1.25, 2.5, 5, or 10 ng COUP-TFI, 250 or 500 ng TDG) werepreincubated 15 min at room temperature with 0.5 mg/mlBSA and 5 mM dithiothreitol. After addition of 0.05 mg/mlpoly(deoxyinosine-deoxycytosine) and binding buffer [10
mM HEPES (pH 7.8), 50 mM KCl, 1 mM EDTA, 5 mM MgCl2,10% glycerol], samples were incubated for 15 min. Thesamples were then incubated with the 32P-labeled DNAduplexes (20,000 cpm) for 20 min. Protein-DNA complexeswere separated on a native 5% polyacrylamide gel in 0.5�TBE [1� TBE: 89 mM Tris, 89 mM borate, 2 mM EDTA (pH8)] at 4 C.
ChIPs
pCR3.1 and COUP-TFI/HA P19 Cells (106) were plated in100-mm dishes. Twenty-four hours later, cells were treatedwith 1 mM 8-Bc-cAMP (Sigma) for the next 24 h. Chromatinwas cross-linked using 1% formaldehyde during 10 min atroom temperature, and the cells were collected after twowashings with PBS in 1-ml cell collection buffer [100 mM
Tris-HCl (pH 9.4), 100 mM DTT]. ChIP were then performedwith minor modifications of the procedure described by Me-tivier et al. (40). Precipitates were serially washed with 300 �lof Washing Buffer I [2 mM EDTA, 20 mM Tris-HCl (pH 8.1),0.1% sodium dodecyl sulfate (SDS), 1% Triton X-100, 150mM NaCl], Washing Buffer II [2 mM EDTA, 20 mM Tris-HCl (pH8.1), 0.1% SDS, 1% Triton X-100, 500 mM NaCl], WashingBuffer III [1 mM EDTA, 10 mM Tris-HCl (pH 8.1), 1% NP-40,1% Deoxycholate, 250 mM LiCl] and then twice with 1 mM
EDTA, 10 mM Tris-HCl (pH 8.1). Precipitated chromatin com-plexes were removed from the beads through three succes-sive 10-min incubations of the beads with 50 �l of 1% SDS,0.1 M NaHCO3. Cross-linking was reversed by an overnightincubation at 65 C. DNA was purified with QIAquick columns(QIAGEN, Hilden, Germany). Re-ChIP assays were run asdescribed (40). The primers used to amplify the vitronectinand PO promoters were agggagattaacgggactgg (pVitro Up2),attgccctcctccaaacac (pVitro Dw2), CGCAGAAAGTTGTTTT-GCTG (pPO Up), and CTGGTTCCATCGACTGTCCT (pPO Dw).
Immunoprecipitation and Western Blot Analysis
The 106 COS-7 cells or 5.105 P19 cells were plated in 100-mmdishes, and were transfected 24 h later with 10 �g of expressionvectors. Cells were collected 36 h after transfection. When pCR3.1and COUP-TFI/HA P19 cells were used, 106 were plated andtreated 24 h later with 1 mM 8-Br-cAMP for the next 24 h. Whole-cell lysates of transiently transfected COS-7 cells were prepared inlysis buffer [20 mM Tris (pH 7.9), 120 mM MgCl2, 0.2% NonidetP-40, 5 mM EDTA, 10% glycerol] containing a protease inhibitorcocktail (Roche). The soluble fraction was immunoprecipitated withanti-Flag M2 monoclonal antibody conjugated to agarose beads(Sigma) at 4 C for 4 h in 500 �l of lysis buffer. Immunocomplexeswere washed seven times in lysis buffer and analyzed by Westernblotting. P19 cells extracts were prepared in lysis buffer with 0.5%Nonidet P-40 and 0.5% Empigen BB (Sigma) and sonicated by 10pulses at 50% duty and power 3 (Sonifer Cell Disruptor, Branson,Danbury, CT). After centrifugation, the supernatants were dilutedwith 1 Volume IP buffer used for ChIP assay and subjected toimmunoprecipitation with 1 �g antibody overnight after a 3 h pre-clearing at 4 C with 50 �l of a 50% protein A-Sepharose beadsslurry (Amersham Biosciences). Complexes were recovered after3 h incubation at 4 C with 50 �l of protein A Sepharose. Precipitateswere serially washed once with Washing buffer I and twice withWashing Buffer II (described above) and analyzed by Westernblotting. Antibodies were diluted 1:1000 in 5% of fat-free milk inPBS, 0.1% Tween 20.
Bisulfite Sequencing
Genomic DNA (40 �g) was denatured by NaOH (0.5 M final) ina volume of 100 �l, for 15 min at 37 C. Ninety microliters of10 mM hydroquinone and 1.4 ml of freshly prepared sodiumbisulphite solution [7.6 g Na metabisulphite in 20 ml of 6.2 M
Urea, 120 mM NaOH (pH 5)] were added to the denatured
2096 Mol Endocrinol, September 2007, 21(9):2085–2098 Gallais et al. • Dnmt3 and COUP-TFI Activate Transcription

DNA. The sample was gently mixed and incubated at 50 C for16 h. Bisulfite-modified DNA was then purified using Gene-clean III kit (Q-Biogene, France), according to the manufac-turer’s instructions. Modification was completed by NaOH(0.3 M final) treatment for 15 min at room temperature. Thesolution was then neutralized by addition of NH4OAc to 3 M
final and the DNA was ethanol precipitated. DNA was resus-pended in 5 mM Tris (pH 8.1) and used immediately forstrand-specific PCR amplification, cloning and sequencing.The transcribed strand was amplified (39 cycles) with thefollowing primers: AAAATACTTACCAACTCTCC (up) andGAATTTTGTTGTTTTGTTTGAAAA (down). The nontran-scribed strand was amplified (39 cycles) with the followingprimers: GAGAGAATAAGGAGAGGTTAG (up) and CTAAC-CCAACCCCATTACCC (down2), after 39 cycles with the oli-gonucleotide CTTCTAAAATCTCACTATCCTACC (down 1).
Acknowledgments
We are grateful to Prof. P. Chambon (Institut de Genetique etde Biologie Moleculaire et Cellulaire, Strasbourg, France), S.Baylin (The Johns Hopkins University, Baltimore, MD), H. Leon-hardt (Ludwig Maximilians University Munich, Munich, Ger-many), E. Li (Novartis, Cambridge, MA), B. W. O’Malley (BaylorCollege of Medicine, Houston, TX), and S. Kato (University ofTokyo, Tokyo, Japan), for the generous gift of plasmids; and toDr. F. Fuller-Pace (University of Dundee, Dundee, Scotland, UK)for the kind gift of the p68 antibody. We thank P. Schar (Uni-versity of Basel, Basel, Switzerland) for giving plasmids and forcritical reading of the manuscript. We also thank C. Martin(University of Rennes), A. Barbelivien (University of Rennes), F.Percevault (University of Rennes), C. Ralliere (Institut National dela Recherche Agronomique), and C. Tascon (Centre National dela Recherche Scientifique) for technical assistance.
Received November 22, 2006. Accepted June 14, 2007.Address all correspondence and requests for reprints to:
Gilles Salbert, Unite Mixte de Recherche 6026 Centre Na-tional de la Recherche Scientifique, Universite Rennes I, In-stitut Federatif de Recherche 140 Genomique FonctionnelleAgronomie Sante, Equipe Information et Programmation Cel-lulaire, Campus de Beaulieu, Batiment 13, 35042 RennesCedex, France. E-mail: [email protected].
This work was supported by funds from the Ministere del’Education Nationale de l’Enseignement Superieur et de la Re-cherche (MENESR) and from the Centre National de la RechercheScientifique. R.G. was supported by a grant from the MENESR anda fellowship from the Ligue Nationale Contre le Cancer.
Permanent address for P.B.: Cancer Research Institute,Slovak Academy of Sciences, Vlarska 7, SK-833 91Bratislava, Slovak Republic. E-mail: [email protected].
Present address for R.L.G.: Unite 522 Institut National de laSante et de la Recherche Medicale, Faculte de Medecine,Campus de Villejean, 35033 Rennes, France. E-mail:[email protected].
Present address for F.G.: Novartis Institute for BiomedicalResearch, Inc., Cambridge, Massachusetts 02115. E-mail:[email protected].
Disclosure Statement: The authors have nothing to disclose.
REFERENCES
1. Razin A 1998 CpG methylation, chromatin structure and genesilencing—a three-way connection. EMBO J 17:4905–4908
2. Rountree MR, Bachman KE, Herman JG, Baylin SB 2001 DNAmethylation, chromatin inheritance, and cancer. Oncogene 20:3156–3165
3. Di Croce L, Raker VA, Corsaro M, Fazi F, Fanelli M, Faretta M,Fuks F, Lo Coco F, Kouzarides T, Nervi C, Minucci S, PelicciPG 2002 Methyltransferase recruitment and DNA hypermeth-ylation of target promoters by an oncogenic transcription fac-tor. Science 295:1079–1082
4. Sato N, Kondo M, Arai K 2006 The orphan nuclear receptorGCNF recruits DNA methyltransferase for Oct-3/4 silencing.Biochem Biophys Res Commun 344:845–851
5. Wang YA, Kamarova Y, Shen KC, Jiang Z, Hahn MJ, Wang Y,Brooks SC 2005 DNA methyltransferase-3a interacts with p53and represses p53-mediated gene expression. Cancer BiolTher 4:1138–1143
6. Bruniquel D, Schwartz RH 2003 Selective, stable demethyl-ation of the interleukin-2 gene enhances transcription by anactive process. Nat Immunol 4:235–240
7. Martinowich K, Hattori D, Wu H, Fouse S, He F, Hu Y, Fan G,Sun YE 2003 DNA methylation-related chromatin remodelingin activity-dependent BDNF gene regulation. Science 302:890–893
8. Mayer W, Niveleau A, Walter J, Fundele R, Haaf T 2000 De-methylation of the zygotic paternal genome. Nature 403:501–502
9. Oswald J, Engemann S, Lane N, Mayer W, Olek A, Fundele R,Dean W, Reik W, Walter J 2000 Active demethylation of thepaternal genome in the mouse zygote. Curr Biol 10:475–478
10. Kress C, Thomassin H, Grange T 2001 Local DNA demethyl-ation in vertebrates: how could it be performed and targeted?FEBS Lett 494:135–140
11. Agius F, Kapoor A, Zhu JK 2006 Role of the Arabidopsis DNAglycosylase/lyase ROS1 in active DNA demethylation. ProcNatl Acad Sci USA 103:11796–11801
12. Gehring M, Huh JH, Hsieh TF, Penterman J, Choi Y, HaradaJJ, Goldberg RB, Fischer RL 2006 DEMETER DNA glycosy-lase establishes MEDEA polycomb gene self-imprinting byallele-specific demethylation. Cell 124:495–506
13. Gong Z, Morales-Ruiz T, Ariza RR, Roldan-Arjona T, David L,Zhu JK 2002 ROS1, a repressor of transcriptional gene silenc-ing in Arabidopsis, encodes a DNA glycosylase/lyase. Cell111:803–814
14. Morales-Ruiz T, Ortega-Galisteo AP, Ponferrada-Marin MI,Martinez-Macias MI, Ariza RR, Roldan-Arjona T 2006 DEME-TER and REPRESSOR OF SILENCING 1 encode 5-methylcy-tosine DNA glycosylases. Proc Natl Acad Sci USA 103:6853–6858
15. Kress C, Thomassin H, Grange T 2006 Active cytosine de-methylation triggered by a nuclear receptor involves DNAstrand breaks. Proc Natl Acad Sci USA 103:11112–11117
16. Fremont M, Siegmann M, Gaulis S, Matthies R, Hess D, JostJP 1997 Demethylation of DNA by purified chick embryo5-methylcytosine-DNA glycosylase requires both protein andRNA. Nucleic Acids Res 25:2375–2380
17. Jost JP, Schwarz S, Hess D, Angliker H, Fuller-Pace FV, StahlH, Thiry S, Siegmann M 1999 A chicken embryo protein re-lated to the mammalian DEAD box protein p68 is tightly asso-ciated with the highly purified protein-RNA complex of 5-MeC-DNA glycosylase. Nucleic Acids Res 27:3245–3252
18. Jost JP, Siegmann M, Sun L, Leung R 1995 Mechanisms ofDNA demethylation in chicken embryos. Purification and prop-erties of a 5-methylcytosine-DNA glycosylase. J Biol Chem270:9734–9739
19. Zhu B, Zheng Y, Hess D, Angliker H, Schwarz S, Siegmann M,Thiry S, Jost JP 2000 5-Methylcytosine-DNA glycosylase ac-tivity is present in a cloned G/T mismatch DNA glycosylaseassociated with the chicken embryo DNA demethylation com-plex. Proc Natl Acad Sci USA 97:5135–5139
20. Waters TR, Gallinari P, Jiricny J, Swann PF 1999 Human thy-mine DNA glycosylase binds to apurinic sites in DNA but isdisplaced by human apurinic endonuclease 1. J Biol Chem274:67–74
21. Heinemann U, Hahn M 1992 C-C-A-G-G-C-m5C-T-G-G. He-lical fine structure, hydration, and comparison with C-C-A-G-G-C-C-T-G-G. J Biol Chem 267:7332–7341
Gallais et al. • Dnmt3 and COUP-TFI Activate Transcription Mol Endocrinol, September 2007, 21(9):2085–2098 2097

22. Mayer-Jung C, Moras D, Timsit Y 1997 Effect of cytosinemethylation on DNA-DNA recognition at CpG steps. J Mol Biol270:328–335
23. Chen D, Lucey MJ, Phoenix F, Lopez-Garcia J, Hart SM,Losson R, Buluwela L, Coombes RC, Chambon P, Schar P, AliS 2003 T:G mismatch-specific thymine-DNA glycosylase po-tentiates transcription of estrogen-regulated genes throughdirect interaction with estrogen receptor �. J Biol Chem 278:38586–38592
24. Chevray PM, Nathans D 1992 Protein interaction cloning inyeast: identification of mammalian proteins that react with theleucine zipper of Jun. Proc Natl Acad Sci USA 89:5789–5793
25. Jost JP, Thiry S, Siegmann M 2002 Estradiol receptor poten-tiates, in vitro, the activity of 5-methylcytosine DNA glycosy-lase. FEBS Lett 527:63–66
26. Tini M, Benecke A, Um SJ, Torchia J, Evans RM, Chambon P2002 Association of CBP/p300 acetylase and thymine DNAglycosylase links DNA repair and transcription. Mol Cell9:265–277
27. Um S, Harbers M, Benecke A, Pierrat B, Losson R, ChambonP 1998 Retinoic acid receptors interact physically and func-tionally with the T:G mismatch-specific thymine-DNA glycosy-lase. J Biol Chem 273:20728–20736
28. Zhu B, Benjamin D, Zheng Y, Angliker H, Thiry S, Siegmann M,Jost JP 2001 Overexpression of 5-methylcytosine DNA glyco-sylase in human embryonic kidney cells EcR293 demethylatesthe promoter of a hormone-regulated reporter gene. Proc NatlAcad Sci USA 98:5031–5036
29. Hardeland U, Steinacher R, Jiricny J, Schar P 2002 Modifica-tion of the human thymine-DNA glycosylase by ubiquitin-likeprotein facilitates enymatic turnover. EMBO J 21:1456–1464
30. Adam F, Sourisseau T, Metivier R, Le Page Y, Desbois C,Michel D, Salbert G 2000 COUP-TFI (chicken ovalbumin up-stream promoter-transcription factor I) regulates cell migrationand axogenesis in differentiating P19 embryonal carcinomacells. Mol Endocrinol 14:1918–1933
31. Jonk LJ, de Jonge ME, Pals CE, Wissink S, Vervaart JM,Schoorlemmer J, Kruijer W 1994 Cloning and expression dur-ing development of three murine members of the COUP familyof nuclear orphan receptors. Mech Dev 47:81–97
32. Frommer M, McDonald LE, Millar DS, Collis CM, Watt F, GriggGW, Molloy PL, Paul CL 1992 A genomic sequencing protocolthat yields a positive display of 5-methylcytosine residues inindividual DNA strands. Proc Natl Acad Sci USA 89:1827–1831
33. Gay F, Barath P, Desbois-Le Peron C, Metivier R, Le Guevel R,Birse D, Salbert G 2002 Multiple phosphorylation events con-trol chicken ovalbumin upstream promoter transcription factorI orphan nuclear receptor activity. Mol Endocrinol 16:1332–1351
34. Endoh H, Maruyama K, Masuhiro Y, Kobayashi Y, Goto M, TaiH, Yanagisawa J, Metzger D, Hashimoto S, Kato S 1999Purification and identification of p68 RNA helicase acting as atranscriptional coactivator specific for the activation function 1of human estrogen receptor �. Mol Cell Biol 19:5363–5372
35. Hardeland U, Bentele M, Jiricny J, Schar P 2000 Separatingsubstrate recognition from base hydrolysis in human thymineDNA glycosylase by mutational analysis. J Biol Chem 275:33449–33456
36. Ahmad I, Rao DN 1996 Chemistry and biology of DNA meth-yltransferases. Crit Rev Biochem Mol Biol 31:361–380
37. Lau EY, Bruice TC 1999 Active site dynamics of the HhaImethyltransferase: insights from computer simulation. J MolBiol 293:9–18
38. Svedruzic ZM, Reich NO 2004 The mechanism of target baseattack in DNA cytosine carbon 5 methylation. Biochemistry43:11460–11473
39. Yebra MJ, Bhagwat AS 1995 A cytosine methyltransferaseconverts 5-methylcytosine in DNA to thymine. Biochemistry34:14752–14757
40. Metivier R, Penot G, Hubner MR, Reid G, Brand H, Kos M,Gannon F 2003 Estrogen receptor-� directs ordered, cyclical,and combinatorial recruitment of cofactors on a natural targetpromoter. Cell 115:751–763
41. Morgan HD, Dean W, Coker HA, Reik W, Petersen-Mahrt SK2004 Activation-induced cytidine deaminase deaminates5-methylcytosine in DNA and is expressed in pluripotenttissues: implications for epigenetic reprogramming. J BiolChem 279:52353–52360
42. Sohail A, Klapacz J, Samaranayake M, Ullah A, Bhagwat AS2003 Human activation-induced cytidine deaminase causestranscription-dependent, strand-biased C to U deaminations.Nucleic Acids Res 31:2990–2994
43. Sharath AN, Weinhold E, Bhagwat AS 2000 Reviving a deadenzyme: cytosine deaminations promoted by an inactive DNAmethyltransferase and an S-adenosylmethionine analogue.Biochemistry 39:14611–14616
44. Zingg JM, Shen JC, Jones PA 1998 Enzyme-mediated cyto-sine deamination by the bacterial methyltransferase M.MspI.Biochem J 332(Pt 1):223–230
45. Zingg JM, Shen JC, Yang AS, Rapoport H, Jones PA 1996Methylation inhibitors can increase the rate of cytosine deami-nation by (cytosine-5)-DNA methyltransferase. Nucleic AcidsRes 24:3267–3275
46. Svedruzic ZM, Reich NO 2005 DNA cytosine C5 methyltrans-ferase Dnmt1: catalysis-dependent release of allosteric inhibi-tion. Biochemistry 44:9472–9485
47. Li YQ, Zhou PZ, Zheng XD, Walsh CP, Xu GL 2007 Associationof Dnmt3a and thymine DNA glycosylase links DNA methyl-ation with base-excision repair. Nucleic Acids Res 35:390–400
48. Luo S, Preuss D 2003 Strand-biased DNA methylation asso-ciated with centromeric regions in Arabidopsis. Proc Natl AcadSci USA 100:11133–11138
49. Zhou Y, Magill CW, Magill JM, Newton RJ 1998 An apparentcase of nonsymmetrical and sustained strand-specific hemi-methylation in the Dc8 gene of carrot. Genome 41:23–33
50. Singal R, vanWert JM 2001 De novo methylation of an embry-onic globin gene during normal development is strand specificand spreads from the proximal transcribed region. Blood 98:3441–3446
51. Lin IG, Han L, Taghva A, O’Brien LE, Hsieh CL 2002 Murine denovo methyltransferase Dnmt3a demonstrates strand asym-metry and site preference in the methylation of DNA in vitro.Mol Cell Biol 22:704–723
52. Xiao JH, Davidson I, Matthes H, Garnier JM, Chambon P 1991Cloning, expression, and transcriptional properties of the hu-man enhancer factor TEF-1. Cell 65:551–568
53. Metivier R, Gay FA, Hubner MR, Flouriot G, Salbert G, GannonF, Kah O, Pakdel F 2002 Formation of an hER�-COUP-TFIcomplex enhances hER� AF-1 through Ser118 phosphoryla-tion by MAPK. EMBO J 21:3443–3453
54. Wu J, Issa JP, Herman J, Bassett Jr DE, Nelkin BD, Baylin SB1993 Expression of an exogenous eukaryotic DNA methyl-transferase gene induces transformation of NIH 3T3 cells. ProcNatl Acad Sci USA 90:8891–8895
55. Hata K, Okano M, Lei H, Li E 2002 Dnmt3L cooperates with theDnmt3 family of de novo DNA methyltransferases to establishmaternal imprints in mice. Development 129:1983–1993
56. Margot JB, Cardoso MC, Leonhardt H 2001 Mammalian DNAmethyltransferases show different subnuclear distributions.J Cell Biochem 83:373–379
Molecular Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremostprofessional society serving the endocrine community.
2098 Mol Endocrinol, September 2007, 21(9):2085–2098 Gallais et al. • Dnmt3 and COUP-TFI Activate Transcription


![Konsekwencje przewrotu majowego w polskiej służbie konsularnej [The consequences of the May coup 1926 in Polish consular service]](https://static.fdokumen.com/doc/165x107/63142ed5b033aaa8b2106e47/konsekwencje-przewrotu-majowego-w-polskiej-sluzbie-konsularnej-the-consequences.jpg)

















