
Extravascular lung water after pneumonectomy followed by ventilator-induced lung injury: A-277

Dendritic cell functional properties in a three-dimensional tissue modelof human lung mucosa
Anh Thu Nguyen Hoang,1 Puran Chen,1 Julius Juarez,1 Patty Sachamitr,2 Bo Billing,3 Lidija Bosnjak,1
Barbro Dahlén,3 Mark Coles,2 and Mattias Svensson1
1Center for Infectious Medicine at the Department of Medicine, Karolinska Institutet, and 3Division of Respiratory Medicineand Allergy at Karolinska University Hospital, Huddinge, and Center for Allergy Research, Karolinska Institutet, Stockholm,Sweden; 2Centre for Immunology and Infection, at Hull York Medical School and Department of Biology, University of York,York, United Kingdom
Submitted 25 February 2011; accepted in final form 12 November 2011
Nguyen Hoang AT, Chen P, Juarez J, Sachamitr P, Billing B,Bosnjak L, Dahlén B, Coles M, Svensson M. Dendritic cell functionalproperties in a three-dimensional tissue model of human lung mucosa.Am J Physiol Lung Cell Mol Physiol 302: L226–L237, 2012. Firstpublished November 18, 2011; doi:10.1152/ajplung.00059.2011.—Inlung tissue, dendritic cells (DC) are found in close association with theepithelial cell layer, and there is evidence of DC regulation by theepithelium; that epithelial dysfunction leads to overzealous immune cellactivation. However, dissecting basic mechanisms of DC interactionswith epithelial cells in human tissue is difficult. Here, we describe amethod to generate a three-dimensional organotypic model of the humanairway mucosa in which we have implanted human DC. The modelrecapitulates key anatomical and functional features of lung mucosaltissue, including a stratified epithelial cell layer, deposition of extracel-lular matrix proteins, and the production of tight junction and adherencejunction proteins. Labeling of fixed tissue model sections and imaging oflive tissue models also revealed that DC distribute in close associationwith the epithelial layer. As functional properties of DC may be affectedby the local tissue microenvironment, this system provides a tool to studyhuman DC function associated with lung mucosal tissue. As an example,we report that the lung tissue model regulates the capacity of DC toproduce the chemokines CCL17, CCL18, and CCL22, leading to en-hanced CCL18 expression and reduced CCL17 and CCL22 expression.This novel tissue model thus provides a tool well suited for a wide rangeof studies, including those on the regulation of DC functional propertieswithin the local tissue microenvironment during homeostasis and inflam-matory reactions.
3D organotypic model; lung mucosal tissue; epithelial cells; fibro-blasts; immune regulation; chemokines
TISSUE HOMEOSTASIS IS A DYNAMIC process dependent on thecooperation between immune cells, tissue specific-nonimmunecells, and extracellular matrix components (23, 34, 40). Al-though technologies have been developed to study cell-cell andcell-matrix interactions during homeostasis and inflammationin three-dimensional (3D) environments in vivo, these ap-proaches are often limited to animal models (11, 28). There-fore, there is a need to develop a new cell-based in vitro humantissue model relevant to the understanding of mechanismsregulating homeostasis and inflammatory responses in thelung. In particular, such approaches can contribute towardincreased understanding of the dynamic interactions betweenhuman nonimmune and immune cells within specific tissues.
Human tissue models can also capture important aspects oftissue-associated responses to infections because many impor-tant human pathogens [e.g., Mycobacterium tuberculosis (M.tuberculosis), group A streptococcus, and Staphylococcus au-reus] induce species specific responses (2, 25).
Several approaches have been used to study immune re-sponses of human lung mucosa to external stimuli in vitro,including monolayers of epithelial cells isolated from lungtissue bronchoscopies (37). Epithelial cells isolated from bron-choscopy are terminally differentiated, short-lived cells thatgenerally have lower metabolic capacity than actively growingcells in vivo or in vitro (14). Although, primary bronchialepithelial cells can develop tight junctions and adherencejunctions as well as deposit extracellular matrix proteins foundin functional mucosal barriers, they may have a variableconstitutive and inducible expression of proteins, depending onthe donor as previously reported (42). These monolayer sys-tems are additionally limited by the lack of polarized cellphenotype and the lack of a large number of cell-cell contacts,which will affect their function and response to external stimuli(31). Instead, the complexity of two-cell layer-based systemsare advantageous as demonstrated when studying, for example,early events in M. tuberculosis infection (2).
More recently, protocols describing the setup of bronchial3D mucosal tissue models that better recapitulate human lungmucosa have been developed (6). This includes models with aphysiologically relevant fibroblast matrix layer and epithelialdifferentiation into ciliated, mucus-secreting cells (6, 9). Undersuch conditions, fibroblasts proliferate slowly, and their extra-cellular matrix is likely to provide better conditions for growthand differentiation of lung epithelial cells in vitro comparedwith artificial gels or membranes, possibly via the release ofgrowth and survival factors (5, 15). The fibroblast componentis, not only critical in promoting survival, remodeling, anddeposition of matrix components, but also ensures the resem-blance to the human lung mucosa and submucosa (4, 5).Fibroblasts actively interact with the adjacent epithelial layerand have a key role in inflammation and repair of lung tissue(36). What has only recently been recognized are the cellularinteractions between epithelial cells, fibroblasts, and immunecells, such as dendritic cells (DC), in many aspects of tissuehomeostasis and inflammation. Thus, in vitro 3D models com-bining DC with tissue-specific cells provides a useful tool tostudy mechanisms involved in local inflammatory processespreviously not achievable, and will have a great potential inreplacing, in some cases, experiments performed on animals.
Address for reprint requests and other correspondence: M. Svensson, Centerfor Infectious Medicine, F59, Dept. of Medicine, Karolinska Institutet, Karo-linska Univ. Hospital, Huddinge, 141 86 Stockholm, Sweden (e-mail:[email protected]).
Am J Physiol Lung Cell Mol Physiol 302: L226–L237, 2012.First published November 18, 2011; doi:10.1152/ajplung.00059.2011.
1040-0605/12 Copyright © 2012 the American Physiological Society http://www.ajplung.orgL226
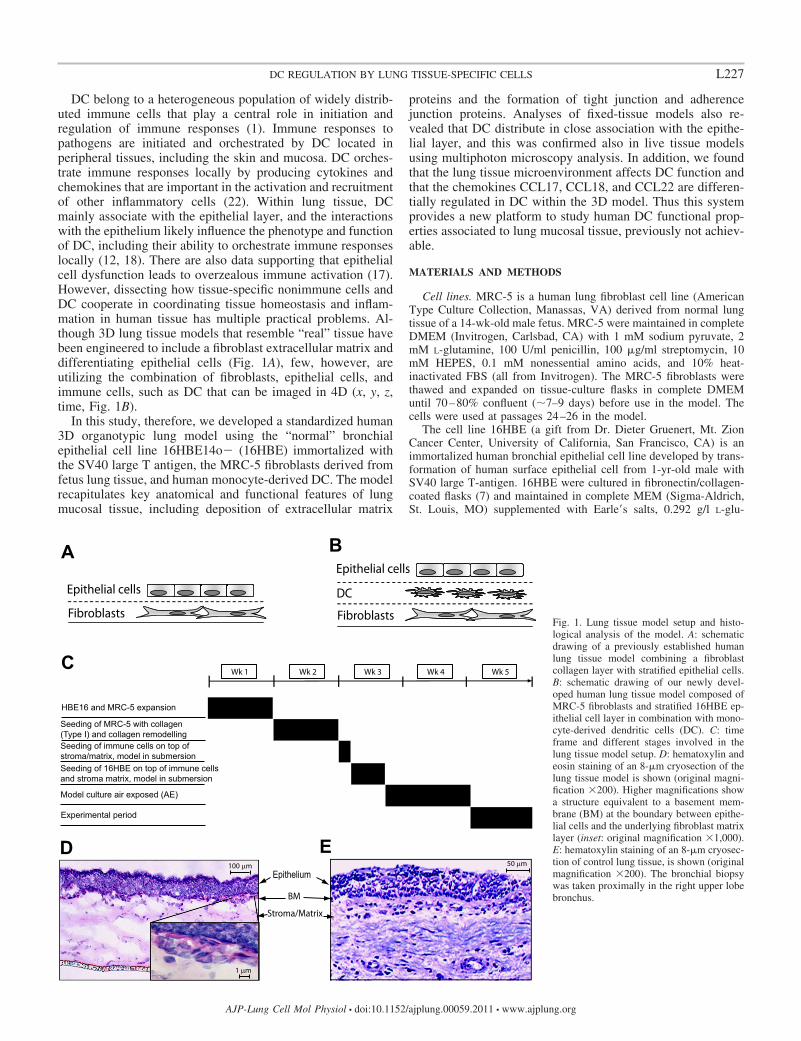
DC belong to a heterogeneous population of widely distrib-uted immune cells that play a central role in initiation andregulation of immune responses (1). Immune responses topathogens are initiated and orchestrated by DC located inperipheral tissues, including the skin and mucosa. DC orches-trate immune responses locally by producing cytokines andchemokines that are important in the activation and recruitmentof other inflammatory cells (22). Within lung tissue, DCmainly associate with the epithelial layer, and the interactionswith the epithelium likely influence the phenotype and functionof DC, including their ability to orchestrate immune responseslocally (12, 18). There are also data supporting that epithelialcell dysfunction leads to overzealous immune activation (17).However, dissecting how tissue-specific nonimmune cells andDC cooperate in coordinating tissue homeostasis and inflam-mation in human tissue has multiple practical problems. Al-though 3D lung tissue models that resemble “real” tissue havebeen engineered to include a fibroblast extracellular matrix anddifferentiating epithelial cells (Fig. 1A), few, however, areutilizing the combination of fibroblasts, epithelial cells, andimmune cells, such as DC that can be imaged in 4D (x, y, z,time, Fig. 1B).
In this study, therefore, we developed a standardized human3D organotypic lung model using the “normal” bronchialepithelial cell line 16HBE14o� (16HBE) immortalized withthe SV40 large T antigen, the MRC-5 fibroblasts derived fromfetus lung tissue, and human monocyte-derived DC. The modelrecapitulates key anatomical and functional features of lungmucosal tissue, including deposition of extracellular matrix
proteins and the formation of tight junction and adherencejunction proteins. Analyses of fixed-tissue models also re-vealed that DC distribute in close association with the epithe-lial layer, and this was confirmed also in live tissue modelsusing multiphoton microscopy analysis. In addition, we foundthat the lung tissue microenvironment affects DC function andthat the chemokines CCL17, CCL18, and CCL22 are differen-tially regulated in DC within the 3D model. Thus this systemprovides a new platform to study human DC functional prop-erties associated to lung mucosal tissue, previously not achiev-able.
MATERIALS AND METHODS
Cell lines. MRC-5 is a human lung fibroblast cell line (AmericanType Culture Collection, Manassas, VA) derived from normal lungtissue of a 14-wk-old male fetus. MRC-5 were maintained in completeDMEM (Invitrogen, Carlsbad, CA) with 1 mM sodium pyruvate, 2mM L-glutamine, 100 U/ml penicillin, 100 �g/ml streptomycin, 10mM HEPES, 0.1 mM nonessential amino acids, and 10% heat-inactivated FBS (all from Invitrogen). The MRC-5 fibroblasts werethawed and expanded on tissue-culture flasks in complete DMEMuntil 70–80% confluent (�7–9 days) before use in the model. Thecells were used at passages 24–26 in the model.
The cell line 16HBE (a gift from Dr. Dieter Gruenert, Mt. ZionCancer Center, University of California, San Francisco, CA) is animmortalized human bronchial epithelial cell line developed by trans-formation of human surface epithelial cell from 1-yr-old male withSV40 large T-antigen. 16HBE were cultured in fibronectin/collagen-coated flasks (7) and maintained in complete MEM (Sigma-Aldrich,St. Louis, MO) supplemented with Earle=s salts, 0.292 g/l L-glu-
Fig. 1. Lung tissue model setup and histo-logical analysis of the model. A: schematicdrawing of a previously established humanlung tissue model combining a fibroblastcollagen layer with stratified epithelial cells.B: schematic drawing of our newly devel-oped human lung tissue model composed ofMRC-5 fibroblasts and stratified 16HBE ep-ithelial cell layer in combination with mono-cyte-derived dendritic cells (DC). C: timeframe and different stages involved in thelung tissue model setup. D: hematoxylin andeosin staining of an 8-�m cryosection of thelung tissue model is shown (original magni-fication �200). Higher magnifications showa structure equivalent to a basement mem-brane (BM) at the boundary between epithe-lial cells and the underlying fibroblast matrixlayer (inset: original magnification �1,000).E: hematoxylin staining of an 8-�m cryosec-tion of control lung tissue, is shown (originalmagnification �200). The bronchial biopsywas taken proximally in the right upper lobebronchus.
L227DC REGULATION BY LUNG TISSUE-SPECIFIC CELLS
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00059.2011 • www.ajplung.org

tamine, 2.2 g/l sodium bicarbonate, 100 U/ml penicillin, 100 �g/mlstreptomycin, and 10% heat-inactivated FBS (Invitrogen) in 5% CO2
at 37°C. The cells were used at passage 74. 16HBE were thenmaintained in complete DMEM when cultured together with thefibroblasts and DC in the model.
Generation and conditioning of monocyte-derived DC. Monocyteswere isolated from buffy-coated blood using human monocyte enrich-ment cocktail RosetteSep (StemCell, Grenoble, France). The cellswere separated by ficoll gradient separation, washed twice in PBS,and subsequently cultured in complete DMEM supplemented with12.5 ng/ml IL-4 (R&D Systems Minneapolis, MN) and 50 ng/mlcolony stimulating factor (CSF)-2 (Peprotech, Rocky Hill, NJ) for 6days. Fresh media including concentration of cytokines (25 ng/mlIL-4, 100 ng/ml CSF-2) was provided on day 3 of culture. DCgenerated from monocytes were routinely positive for CD1a andnegative for CD14 (� 90%). For the conditioning experiments, DC (5� 104 cells) from day 6 were conditioned by replacing the originalmedium with 500 �l of culture supernatants from the lung tissuemodel composed of only fibroblasts and epithelial cells, or withsupernatants from either fibroblasts or air-exposed epithelial cells, orwith supernatants from a mixed monolayer of both fibroblasts andepithelial cells. The DC were conditioned for 24 h and then extractedfor RNA, and supernatants were collected for protein measurementusing ELISA. Ethical permission for the use of human tissue and cellswas obtained from the regional ethics committee at Karolinska Insti-tutet, Stockholm, Sweden, and where needed informed consent hasbeen obtained.
Organotypic model preparation. The models were generated on3.0-�m Transwell inserts (Becton Dickinson, Franklin Lakes, NJ) insix-well plates based on a protocol for human oral mucosal models(9). The procedure of establishing model tissue takes �4–5 wk (Fig.1C) and includes cell expansion, culturing of the cells in the model,and air exposure (AE) of the tissue model. To set up the model, theinner chamber of the six-well inserts were coated with a solution ofbovine type I collagen (1.1 mg/ml; Organogenesis, Canton, MA) inDMEM and incubated for 30 min at 37°C. A suspension of MRC-5fibroblasts at 2.3 � 105 cells/ml diluted in 1.1 mg/ml of collagenDMEM mixture was added onto the precoated collagen layer andincubated for 2 h in 5% CO2 at 37°C. Following polymerization, 2 mlof complete DMEM was added to the outer chamber, and the cultureswere incubated for an additional 24 h. The following day, culturemedia was removed from the outer and the inner chamber. Then 2 mlof fresh complete DMEM was added to both chambers. To allowremodeling of the stroma/matrix layer, cultures were incubated for 7days with the medium being replaced every second day.
After 7 days, the culture media was removed from the outer andinner chambers followed by the addition of 1.5 ml of fresh completeDMEM to the outer chamber. Then 2 � 105 monocyte-derived DC in50 �l of complete DMEM were added to the fibroblast collagenmatrix and incubated for 2 h in 5% CO2 at 37°C. After incubation 1.5ml of culture media was gently added into the insert, and the culturewas incubated for an additional 24 h in 5% CO2 at 37°C beforeseeding of the epithelial cells. Epithelial cells were seeded by adding5 � 104 epithelial cells in 50 �l complete DMEM to each insert.Submerged cultures were incubated for an additional 3 days in 5%CO2 at 37°C to allow the epithelial cells to form a confluent mono-layer on the collagen gel. After 3 days, the tissue models were airexposed by removing the medium from the insert and reducing thevolume to 1.5 ml in the outer chamber. Air-exposed tissue models wereincubated for up to 10 days in 5% CO2 at 37°C, and the culture media inthe outer chamber was changed every second day. At days 5 to 10, tissuemodels were used for immunohistological immunofluorescence and live-cell imaging analyses, as well as DC functional analyses.
Histological and immunohistological analyses. Lung tissue modelfor cryosectioning was prepared by embedding the tissue in optimalcutting temperature compound (Sakura Finetek Europe, Zoeterwoude,Netherlands) and then slowly frozen in �20°C for 24 h and trans-
ferred to �80°C until required. Eight-micron cryosections were cutonto diagnostic microscope glass slides (Thermo Scientific, Waltham,MA) using a MICROM cryostat HM 560 MV (Carl Zeiss, Jena,Germany) and fixed in 2% freshly prepared formaldehyde in PBS for15 min at room temperature or in ice-cold acetone for 10 min at�20°C. For hematoxylin and eosin staining, the sections were stainedfor 15 s in Mayes Hematoxylin and counterstained for 2 min in eosin.
For immunohistochemistry, 8-�m frozen sections were blockedwith 10% FBS in balanced salt solution (BSS) with 0.1% saponin for30 min at room temperature, followed by additional blocking with 2%H2O2 in BSS-saponin and an avidin biotin blocking reagent (VectorLaboratories, Burlingame, CA). Primary monoclonal rat anti-humanleukocyte antigen (HLA)-DR (1.25 �g/ml, clone YE2/36-HLK; AbDSerotec, Dusseldorf, Germany) antibody was diluted in BSS solutioncontaining saponin and incubated overnight at room temperature aspreviously described (27). After incubation, tissue sections werewashed and blocked with 1% normal rabbit serum in BSS-saponinbefore addition of biotinylated rabbit anti-rat IgG (4.2 �g/ml; Dako,Glostrup, Denmark) diluted in 1% normal rabbit serum in BSS-saponin. After wash avidin-peroxidase solution was added (Vec-tastain-Elite; Vector Laboratories), and the color reaction developedby the addition of 3,3-diaminobenzidine (Vector Laboratories) fol-lowed by counterstaining with hematoxylin. Tissue sections werevisualized using a Leica DMR microscope (Wetzlar, Germany).Imaging and quantification of positive immunostaining were per-formed using acquired computerized image analysis by transferringdigital images of the stained tissue samples taken by a DMR-X micro-scope (Leica) to a computerized Quantimet 5501W image analyzer(Leica) (3). Single-positive stained cells were quantified in 25 high-powerfields, and protein expression was determined as the percent positive areaof the total relevant cell area using a Qwin 550 software program (LeicaImaging Systems). The total cell area was defined as the nucleated andcytoplasmic area within the tissue. Tissue sections stained with secondaryantibodies only were used as negative controls.
Alcian blue staining. Eight-micron-thick frozen tissue sectionswere hydrated in distilled water and stained in Alcian blue solution(pH 2.5, Sigma Aldrich) for 30 min at room temperature. The sectionswere washed for 2 min in tap water and then rinsed in distilled water.After that, sections were counterstained in nuclear Fast Red solution(Sigma Aldrich) for 5 min and were washed in tap water for 1 min. Thesections were dehydrated through 95% alcohol and twice in absolutealcohol for 3 min each. The sections were cleared in xylene and mountedin DPX mountant (VWR International, Radnor, PA). Mucins and muco-substances were thus stained in blue, nuclei in pink to red, and cytoplasmin pale pink.
Immunofluorescence analysis. Immunofluorescent labeling of8-�m frozen sections was performed with the following primaryantibodies: mouse anti-type IV collagen (13.5 �g/ml, clone COL-94;Abcam, Cambridge, UK), rabbit anti-tropoelastin (1:200, polyclonal;Elastin Products, Owensville, MO), mouse anti-laminin-5 (�3) (2.5�g/ml, clone P3H9–2; R&D Systems, Minneapolis, MN), mouseanti-vimentin (1:100, clone VIM3B4; Novocastra Laboratories, New-castle, UK), rabbit anti-claudin I (1 �g/ml, polyclonal; Abcam),mouse anti-occludin (2.5 �g/ml, clone OC-3F10; Invitrogen), mouseanti-E-cadherin (2 �g/ml, clone HECD-1; Invitrogen), rat anti-HLA-DR (2.5 �g/ml, clone YE2/36-HLK; AbD Serotec), mouseanti-DC-specific ICAM-grabbing nonintegrin (SIGN) (0.25 �g/ml,clone 120507; R&D Systems), and mouse anti-CD11c (1.25 �g/ml,clone B-ly6; BD Pharmingen, San Diego, CA). Sections were washedin PBS containing 0.2% saponin and blocked with 1.5% normal goatserum (NGS) in 0.2% saponin in PBS for 45 min at room temperature.Sections were incubated with primary antibodies (see above) dilutedin PBS containing 1.5% NGS and 0.2% saponin for 45 min at roomtemperature and then washed three times in PBS with 0.2% saponin.Specific staining was detected by the following secondary antibodies:Alexa 488-conjugated goat anti-rat IgG, Alexa 488-conjugated goatanti-mouse IgG, Alexa 555-conjugated goat anti-mouse IgG2b, Alexa
L228 DC REGULATION BY LUNG TISSUE-SPECIFIC CELLS
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00059.2011 • www.ajplung.org

488-conjugated goat anti-mouse IgG1, Alexa 594-conjugated goatanti-rabbit IgG (all from Molecular Probes, Invitrogen). The sectionswere incubated with secondary antibodies diluted to 3.3 �g/ml forAlexa 488 and 4 �g/ml for Alexa 555 and 594 in PBS containing1.5% NGS and 0.2% saponin for 30 min at room temperature.Sections were mounted in SlowFade Gold antifade reagent with4,6-diamidino-2-phenylindole (DAPI) (Molecular Probes, Invitrogen)and visualized using a Leica TCS SP2 confocal microscope (Leica).
Quantification of DC numbers over time was performed in livetissue models by imaging the models at days 1, 3, 5, and 8 using aconfocal microscope. To enable DC visualization in live tissue, DCwere labeled with the far-red cell tracker, dimethyl dodecylamineoxide-syccinimidyl (Molecular Probes, Invitrogen), before implanta-tion into the models. Day 6 monocyte-derived DC were washed twicewith PBS, resuspended at 106 cells/ml of PBS, and labeled with 5 �Mof the cell tracker at room temperature for 15 min. Cells were washedtwice with PBS after being labeled and resuspended to 2 � 105
cells/50 �l of complete DMEM. The labeled DC were seeded in 50 �ldirectly on top of the fibroblast collagen layer as described above.After 24 h, epithelial cells were added to the models followed by thequantification of DC (day 1). Additional 3D models with DC were airexposed and cultured for DC quantification at days 3, 5, and 8. Tissuemodels were visualized using a Nikon A1R confocal microscope(Nikon Instruments, Amstelveen, The Netherlands), and the obtainedz-stack images were used for reconstruction of 3D projections. Toquantify DC in 3D microscopic projections, fluorescently labeled cellswithin a defined field of each 3D projection were counted. A mini-mum of 250 cells was counted in each field, and, to exclude enumer-ation of dead cells, propidium iodide was added to the models beforeimaging. Then the number of cells per defined field was multipliedwith a factor reflecting the total 3D tissue model area.
Generation of 16HBE cells expressing tdTomato. pCMV expres-sion vector encoding the fluorescent tdTomato protein was a kind giftfrom R. Tsien (San Diego, CA). Plasmids were purified usingHiSpeed MidiPrep kit (Qiagen, Crawley, UK) according to manufac-turer’s instructions. Cells were cultured in 35-mm culture dishes(MaTek, Ashland, MA), and, at 50–80% confluency, cells weretransfected utilizing 4 �g of pCMV-tdTomato plasmid diluted in 100�l OptiMEM and 14 �l Lipofectamine (Invitrogen) diluted in 100 �lOptiMEM. Both solutions were mixed and incubated at room tem-perature for 30 min. Media was removed from 35-mm dish andreplaced with 600 �l OptiMEM and transfection mixture. After 6 h ofincubation at 5% CO2, 37°C, 1.2 ml of complete DMEM with 20%FBS was added to the dish. The media was replaced with completeDMEM the next morning. Transfection efficiencies were analyzedusing a CyAn flow cytometer (Beckman Coulter, Brea, CA). Cellswere selected in 400 �g G418 for 2 wk, and then the highestexpression fraction was isolated using high-speed MoFlo cell sorter(Beckman Coulter). Nontransfected 16HBE cells were used to definethe background fluorescence and gating strategy. Postsort purity ofcells was analyzed using the CyAn ADP flow cytometer.
Live imaging of the tissue model using multiphoton microscopy.The lung tissue model was generated for live imaging using fluores-cent tdTomato 16HBE and carboxyfluorescein diacetate succinimidylester (CFSE)-labeled monocyte-derived DC. Day 6 DC were washedtwice with PBS, resuspended at 106 cells/ml of PBS, and labeled with5 �M of CFSE (Molecular Probes, Invitrogen) at room temperaturefor 15 min. Cells were washed twice with PBS after labeling andresuspended to 2 � 105 cells/50 �l of complete DMEM. The labeledDC were seeded directly on top of the fibroblast collagen layer asdescribed above. After 7 days of AE, the fluorescent model wasremoved from the membrane insert and placed onto a 35-mm glass-bottom dish and imaged on a Zeiss LSM 510 NLO Meta LaserScanning Confocal Microscope (Carl Zeiss) with three detectors fornon-descanned detection (NDD) and a Coherent Chameleon Ultralaser (Coherent, Santa Clara, CA) with a blacked out, temperaturecontroller chamber (Solent Scientific, Segensworth, UK). Imaging
was performed at 872–930-nm wavelengths using a C-Apochromat�40 1.1 NA water dipping objective.
Real-time qRT-PCR analysis. Total RNA was extracted from DCand from the lung tissue model by using a Ribopure kit (AppliedBiosystems, Foster City, CA) following the manufacturer’s protocol.To compare chemokine expression between the 3D model containingDC and the control DC, we used 1.5 � 105 DC (similar numbers ofcells that was implanted in each 3D model) cultured in 1.5 ml ofcomplete DMEM without CSF-2 and IL-4 for 24 h before analysis.Similar conditions for control DC and DC implanted in the 3D modelwere obtained by lysing control DC in TRIzol together with one 3Dmodel (fibroblasts and epithelial cells only) before RNA extraction.RNA was converted into cDNA by using a cDNA reverse transcrip-tion kit (Applied Biosystems) according to the manufacturer’s instruc-tions. The primers and probes for HLA-DR, DC-SIGN, CD11c,CSF-2, IL-4, IL-1�, TNF, IL-10, IL-13, transforming growth factor(TGF)-�, CX3CL1, CXCL8, CCL17, CCL18, CCL22 and ubiquitin Cwere purchased as Predeveloped TaqMan Gene Expression Assays(Applied Biosystems). Ubiquitin C served as an endogenous control tonormalize the amount of sample cDNA. Amplification and quantifi-cation of cDNA was performed as previously described (27). Relativeamounts of the two chemokines were calculated using the compara-tive threshold cycle (CT) method. The threshold correlates to the cyclenumber where there is sufficient amplified product to give a detectablereading, and, if the threshold is not attained after 35–40 cycles,the mRNA is considered undetectable. The data are presented asrelative expression of chemokine mRNA in untreated DC conditionedwith supernatant compared with the chemokine mRNA in control DC.The data are also presented as relative expression of mRNA in thelung tissue model containing DC compared with control DC (mixed1:1 with RNA extracted from a 3D model without DC) and 3D modelcomposed of epithelial cells and fibroblasts only.
Chemokine measurement. Supernatants from control DC and tissuemodel were collected for protein measurement. A sandwich ELISAconstruction kit to detect CCL18 (R&D Systems), CCL17, andCCL22 (Antigenix, Huntington, NY) was used according to themanufacturer’s protocols. CCL18 was detected using 1.0 �g/ml cap-ture of antibody and 200 ng/ml of detection antibody. CCL17 wasdetected using 1.0 �g/ml capture of antibody and 0.2 �g/ml detectionantibody. CCL22 was detected using 1.0 �g/ml of capture antibodyand 0.25 �g/ml detection antibody. Assays were developed by addingstreptavidin-peroxidase and 3,3,5,5-tetramethyl-benzidine as the sub-strate. Absorbance was read at 450 nm using a Microplate Manager 6reader (Bio-Rad, Hemel Hempstead, UK). Recombinant CCL18 (1ng/ml), CCL17 (10 ng/ml) and CCL22 (10 ng/ml) were used asstandards, and the concentration of chemokines in the test sampleswas calculated using the linear part of a “four-parameter fit” standardcurve run in parallel with the samples.
Statistical analysis. Data are presented as means � SD for theindicated number of experiments. All analyses are based on three ormore separate experiments performed with monocyte-derived DCfrom different donors. The data were analyzed with use of theGraphPad Prism v.5 software (GraphPad, San Diego, CA) usingone-way ANOVA with Bonferroni multiple-comparison test, anddifferences between groups were determined to be statistically signif-icant at P 0.05.
RESULTS
Characterization of the lung tissue model. To reveal path-ways by which human DC and tissue-specific nonimmune cellscooperate to orchestrate lung tissue physiology and immuneresponses at the mucosal surface, we sought to establish a 3Dorganotypic lung model combining human DC with two dis-tinct cellular compartments: a fibroblast/matrix layer and anepithelial cell layer. The lung tissue model was cultured on a
L229DC REGULATION BY LUNG TISSUE-SPECIFIC CELLS
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00059.2011 • www.ajplung.org

permeable membrane for a total of 11–14 days, includingsubmersion in medium for 4 days and AE for 7–10 days. AfterAE histological analysis of the tissue model revealed that theepithelial cells had formed a stratified squamous structure ontop of the fibroblast matrix consistent with reports in similarsystems containing lung epithelial cells (Fig. 1D). Also, astructure equivalent to a basement membrane was observed atthe boundary between epithelial cells and the underlying fibro-blast matrix layer (Fig. 1D, inset). These data demonstrate thatour 3D organotypic lung model is fully developed and has astructure that resembles normal lung mucosa tissue (Fig. 1E).
Extracellular matrix proteins are deposited in the lung tissuemodel. Extracellular matrix proteins, such as collagens, elastin,and laminins are essential for tissue to function properly. Toconfirm that the tissue-specific cells in our model produce anddeposit important tissue components, immunofluorescent anal-ysis of frozen tissue sections was performed. Sections werelabeled with antibodies against type IV collagen and tropoelas-tin, known to be important for formation of basement mem-branes. This revealed that both epithelial cells and fibroblastsin the tissue model produce type IV collagen and tropoelastinand that a continuous basement membrane is formed at theboundary between epithelial cells and the underlying fibroblastmatrix layer (Fig. 2, A and B). Tissue sections were alsoanalyzed for the presence of laminin-5, an extracellular matrixprotein important in the initiation and maintenance of epithelialcell anchorage to the underlying connective tissue layer (35).Laminin-5 was found associated to epithelial layer as well as thebasement membrane at the boundary between fibroblasts andepithelial cells (Fig. 2D). Immunofluorescence analysis alsoconfirmed that yet another protein, vimentin, that is importantfor tissue fibroblast function was produced by the tissue model(39). Vimentin staining was only found in the fibroblast col-
lagen matrix layer and not in the epithelial layer (Fig. 2C).Together, this indicates that molecules forming essential struc-tures for lung tissue function are produced and deposited in themodel.
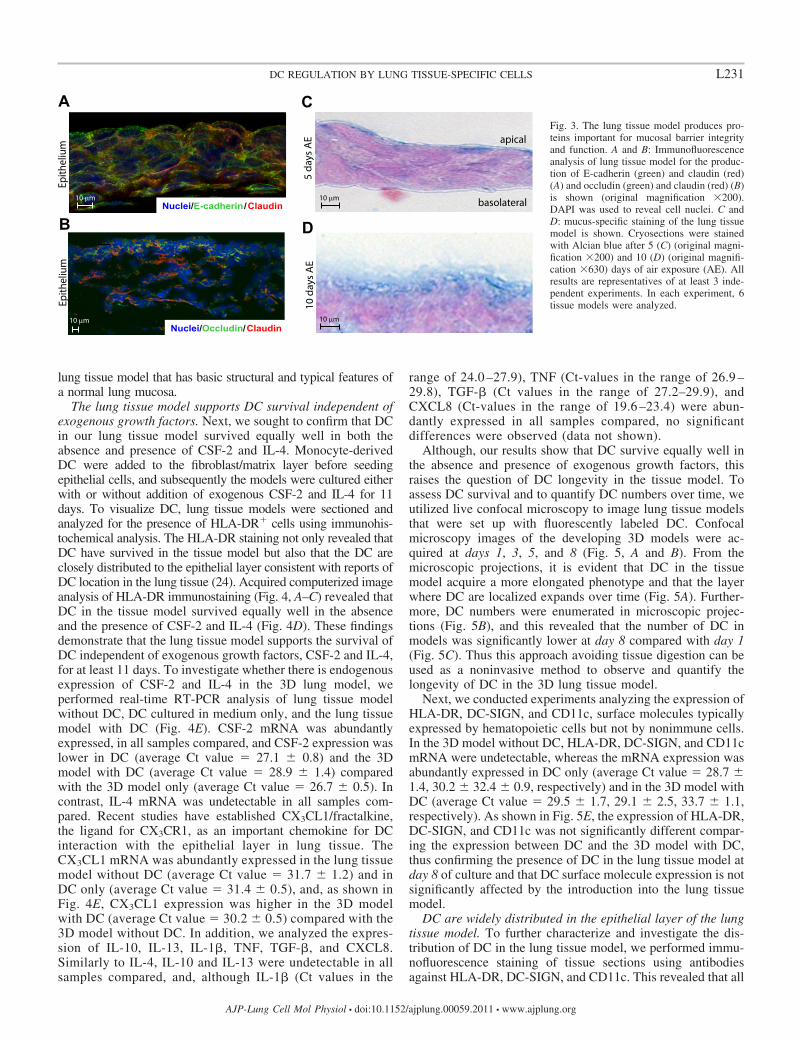
The lung tissue model form tight and adherence junctionsand a mucus layer. One key function of mucosal barriersresides in epithelial cell impermeability that is mediated by theformation of tight and adherence junctions (19). To verify thatthe epithelial cell layer of the lung tissue model has thecapacity to form tight junctions and adhesion junctions, weanalyzed the tissue sections for the presence of claudin I andoccludin, two proteins important for formation of mucosaljunction structures (19). As shown in Fig. 3A, epithelial cells inthe lung tissue model express claudin I. In addition, occludinthat interacts directly with claudins and actin (19) was detectedin the lung tissue model (Fig. 3B). In mucosal tissue, produc-tion of E-cadherin by epithelial cells is important for formationand preservation of stratified epithelial barriers (19). Therefore,the model was analyzed for the presence of E-cadherin, andthis revealed that E-cadherin is preferentially expressed by theepithelial cells and not by the fibroblasts in the tissue model(Fig. 3A and data not shown).
The production of mucus by the epithelium plays an importantrole in the protection of epithelial barriers against infectiousagents and toxins that enter the respiratory system. To investigatewhether our epithelial cells secret mucus, we stained the modelwith Alcian blue, a reagent that stains acid mucosubstances andacetic mucins. This showed mucus production already after 5 daysof AE (Fig. 3C). Mucus production was also detected at 10 daysof AE (Fig. 3D). These data indicate that the tissue model has afunctional mucosal barrier and produce the tight junction proteins,claudin I and occludin, the cell-cell adhesion molecule E-cadherin, and mucus. To conclude, we have created a human 3D
Fig. 2. The lung tissue model produces struc-tural proteins important for lung tissue func-tion. Immunofluorescence analyses of struc-tural proteins essential for lung tissue functionwere carried out on cryosections (8 �m) offrozen lung tissue models. A and B: imagesshow distribution of collagen IV (green) andtropoelastin (red) in the lung tissue model.Higher magnification (original magnification�630) shows localization of collagen IV andtropoelastin fibrous structures (arrowed) at theboundary between the stroma matrix and ep-ithelial cell layer (B). C and D: images showdistribution of vimentin (C) and laminin-5 (D)in the lung tissue model (original magnifica-tion �200). 4,6-Diamidino-2-phenylindole(DAPI) was used to reveal cell nuclei. Allresults are representatives of at least 3 inde-pendent experiments. In each experiment, 6tissue models were analyzed (original magni-fication �200).
L230 DC REGULATION BY LUNG TISSUE-SPECIFIC CELLS
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00059.2011 • www.ajplung.org
lung tissue model that has basic structural and typical features ofa normal lung mucosa.
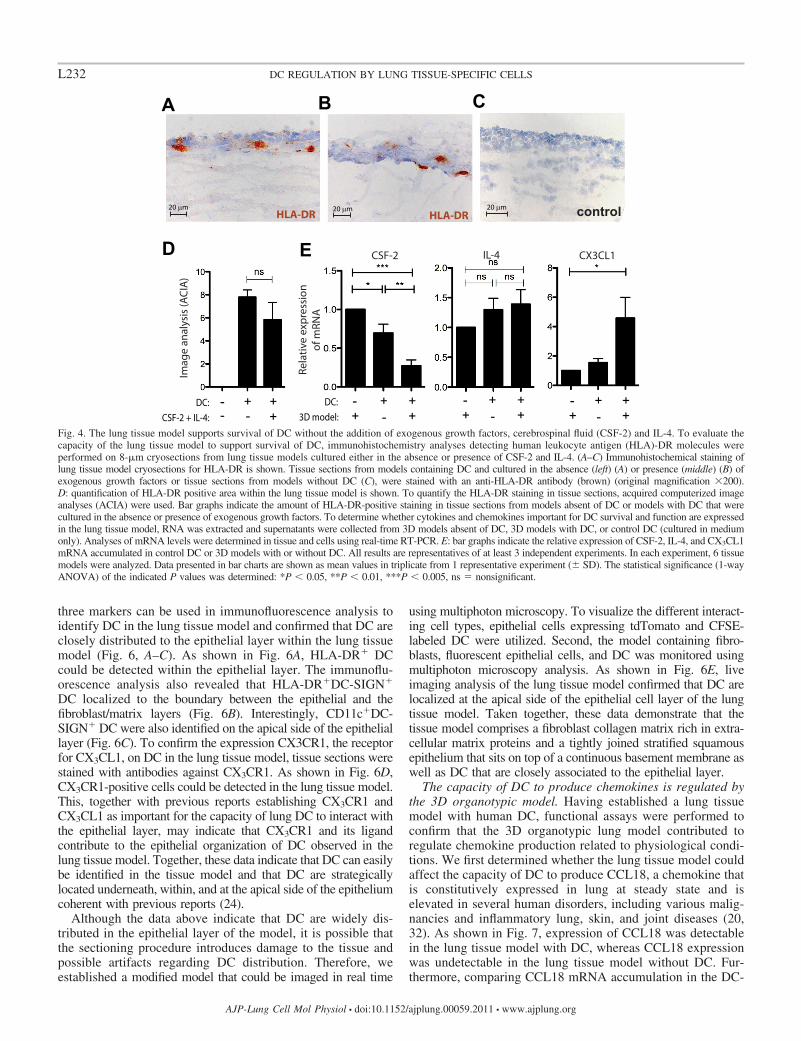
The lung tissue model supports DC survival independent ofexogenous growth factors. Next, we sought to confirm that DCin our lung tissue model survived equally well in both theabsence and presence of CSF-2 and IL-4. Monocyte-derivedDC were added to the fibroblast/matrix layer before seedingepithelial cells, and subsequently the models were cultured eitherwith or without addition of exogenous CSF-2 and IL-4 for 11days. To visualize DC, lung tissue models were sectioned andanalyzed for the presence of HLA-DR cells using immunohis-tochemical analysis. The HLA-DR staining not only revealed thatDC have survived in the tissue model but also that the DC areclosely distributed to the epithelial layer consistent with reports ofDC location in the lung tissue (24). Acquired computerized imageanalysis of HLA-DR immunostaining (Fig. 4, A–C) revealed thatDC in the tissue model survived equally well in the absenceand the presence of CSF-2 and IL-4 (Fig. 4D). These findingsdemonstrate that the lung tissue model supports the survival ofDC independent of exogenous growth factors, CSF-2 and IL-4,for at least 11 days. To investigate whether there is endogenousexpression of CSF-2 and IL-4 in the 3D lung model, weperformed real-time RT-PCR analysis of lung tissue modelwithout DC, DC cultured in medium only, and the lung tissuemodel with DC (Fig. 4E). CSF-2 mRNA was abundantlyexpressed, in all samples compared, and CSF-2 expression waslower in DC (average Ct value � 27.1 � 0.8) and the 3Dmodel with DC (average Ct value � 28.9 � 1.4) comparedwith the 3D model only (average Ct value � 26.7 � 0.5). Incontrast, IL-4 mRNA was undetectable in all samples com-pared. Recent studies have established CX3CL1/fractalkine,the ligand for CX3CR1, as an important chemokine for DCinteraction with the epithelial layer in lung tissue. TheCX3CL1 mRNA was abundantly expressed in the lung tissuemodel without DC (average Ct value � 31.7 � 1.2) and inDC only (average Ct value � 31.4 � 0.5), and, as shown inFig. 4E, CX3CL1 expression was higher in the 3D modelwith DC (average Ct value � 30.2 � 0.5) compared with the3D model without DC. In addition, we analyzed the expres-sion of IL-10, IL-13, IL-1�, TNF, TGF-�, and CXCL8.Similarly to IL-4, IL-10 and IL-13 were undetectable in allsamples compared, and, although IL-1� (Ct values in the
range of 24.0 –27.9), TNF (Ct-values in the range of 26.9 –29.8), TGF-� (Ct values in the range of 27.2–29.9), andCXCL8 (Ct-values in the range of 19.6 –23.4) were abun-dantly expressed in all samples compared, no significantdifferences were observed (data not shown).
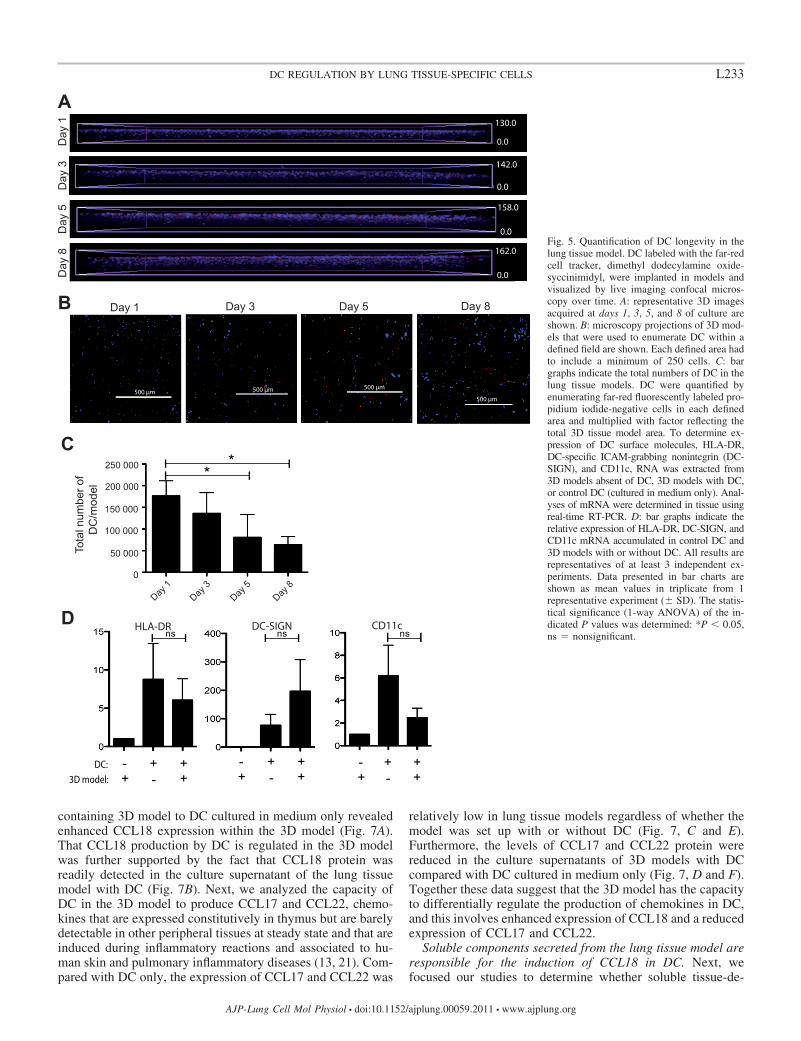
Although, our results show that DC survive equally well inthe absence and presence of exogenous growth factors, thisraises the question of DC longevity in the tissue model. Toassess DC survival and to quantify DC numbers over time, weutilized live confocal microscopy to image lung tissue modelsthat were set up with fluorescently labeled DC. Confocalmicroscopy images of the developing 3D models were ac-quired at days 1, 3, 5, and 8 (Fig. 5, A and B). From themicroscopic projections, it is evident that DC in the tissuemodel acquire a more elongated phenotype and that the layerwhere DC are localized expands over time (Fig. 5A). Further-more, DC numbers were enumerated in microscopic projec-tions (Fig. 5B), and this revealed that the number of DC inmodels was significantly lower at day 8 compared with day 1(Fig. 5C). Thus this approach avoiding tissue digestion can beused as a noninvasive method to observe and quantify thelongevity of DC in the 3D lung tissue model.
Next, we conducted experiments analyzing the expression ofHLA-DR, DC-SIGN, and CD11c, surface molecules typicallyexpressed by hematopoietic cells but not by nonimmune cells.In the 3D model without DC, HLA-DR, DC-SIGN, and CD11cmRNA were undetectable, whereas the mRNA expression wasabundantly expressed in DC only (average Ct value � 28.7 �1.4, 30.2 � 32.4 � 0.9, respectively) and in the 3D model withDC (average Ct value � 29.5 � 1.7, 29.1 � 2.5, 33.7 � 1.1,respectively). As shown in Fig. 5E, the expression of HLA-DR,DC-SIGN, and CD11c was not significantly different compar-ing the expression between DC and the 3D model with DC,thus confirming the presence of DC in the lung tissue model atday 8 of culture and that DC surface molecule expression is notsignificantly affected by the introduction into the lung tissuemodel.
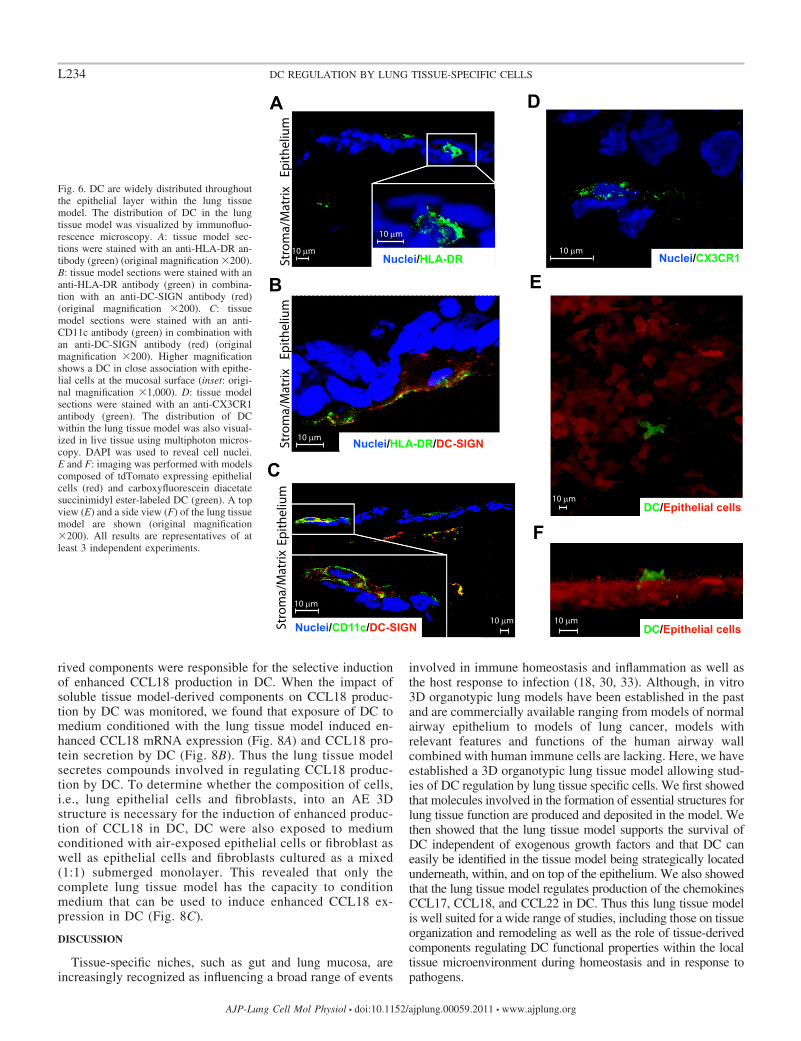
DC are widely distributed in the epithelial layer of the lungtissue model. To further characterize and investigate the dis-tribution of DC in the lung tissue model, we performed immu-nofluorescence staining of tissue sections using antibodiesagainst HLA-DR, DC-SIGN, and CD11c. This revealed that all
Fig. 3. The lung tissue model produces pro-teins important for mucosal barrier integrityand function. A and B: Immunofluorescenceanalysis of lung tissue model for the produc-tion of E-cadherin (green) and claudin (red)(A) and occludin (green) and claudin (red) (B)is shown (original magnification �200).DAPI was used to reveal cell nuclei. C andD: mucus-specific staining of the lung tissuemodel is shown. Cryosections were stainedwith Alcian blue after 5 (C) (original magni-fication �200) and 10 (D) (original magnifi-cation �630) days of air exposure (AE). Allresults are representatives of at least 3 inde-pendent experiments. In each experiment, 6tissue models were analyzed.
L231DC REGULATION BY LUNG TISSUE-SPECIFIC CELLS
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00059.2011 • www.ajplung.org
three markers can be used in immunofluorescence analysis toidentify DC in the lung tissue model and confirmed that DC areclosely distributed to the epithelial layer within the lung tissuemodel (Fig. 6, A–C). As shown in Fig. 6A, HLA-DR DCcould be detected within the epithelial layer. The immunoflu-orescence analysis also revealed that HLA-DRDC-SIGN
DC localized to the boundary between the epithelial and thefibroblast/matrix layers (Fig. 6B). Interestingly, CD11cDC-SIGN DC were also identified on the apical side of the epitheliallayer (Fig. 6C). To confirm the expression CX3CR1, the receptorfor CX3CL1, on DC in the lung tissue model, tissue sections werestained with antibodies against CX3CR1. As shown in Fig. 6D,CX3CR1-positive cells could be detected in the lung tissue model.This, together with previous reports establishing CX3CR1 andCX3CL1 as important for the capacity of lung DC to interact withthe epithelial layer, may indicate that CX3CR1 and its ligandcontribute to the epithelial organization of DC observed in thelung tissue model. Together, these data indicate that DC can easilybe identified in the tissue model and that DC are strategicallylocated underneath, within, and at the apical side of the epitheliumcoherent with previous reports (24).
Although the data above indicate that DC are widely dis-tributed in the epithelial layer of the model, it is possible thatthe sectioning procedure introduces damage to the tissue andpossible artifacts regarding DC distribution. Therefore, weestablished a modified model that could be imaged in real time
using multiphoton microscopy. To visualize the different interact-ing cell types, epithelial cells expressing tdTomato and CFSE-labeled DC were utilized. Second, the model containing fibro-blasts, fluorescent epithelial cells, and DC was monitored usingmultiphoton microscopy analysis. As shown in Fig. 6E, liveimaging analysis of the lung tissue model confirmed that DC arelocalized at the apical side of the epithelial cell layer of the lungtissue model. Taken together, these data demonstrate that thetissue model comprises a fibroblast collagen matrix rich in extra-cellular matrix proteins and a tightly joined stratified squamousepithelium that sits on top of a continuous basement membrane aswell as DC that are closely associated to the epithelial layer.
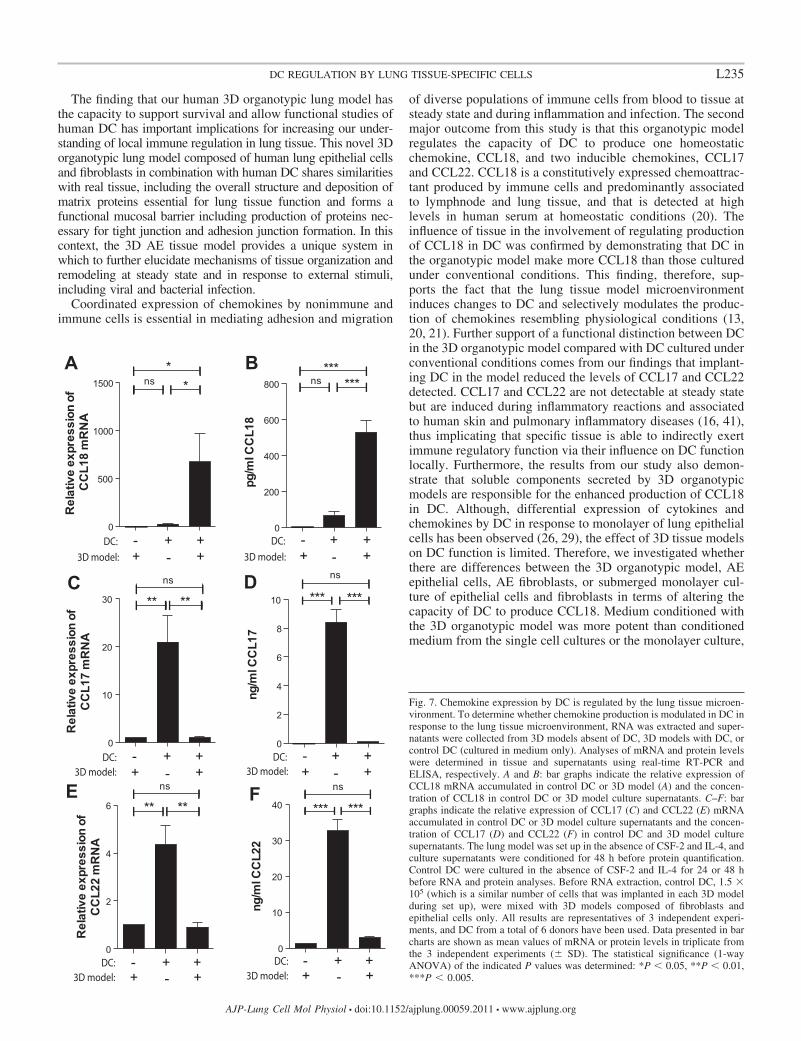
The capacity of DC to produce chemokines is regulated bythe 3D organotypic model. Having established a lung tissuemodel with human DC, functional assays were performed toconfirm that the 3D organotypic lung model contributed toregulate chemokine production related to physiological condi-tions. We first determined whether the lung tissue model couldaffect the capacity of DC to produce CCL18, a chemokine thatis constitutively expressed in lung at steady state and iselevated in several human disorders, including various malig-nancies and inflammatory lung, skin, and joint diseases (20,32). As shown in Fig. 7, expression of CCL18 was detectablein the lung tissue model with DC, whereas CCL18 expressionwas undetectable in the lung tissue model without DC. Fur-thermore, comparing CCL18 mRNA accumulation in the DC-
Fig. 4. The lung tissue model supports survival of DC without the addition of exogenous growth factors, cerebrospinal fluid (CSF-2) and IL-4. To evaluate thecapacity of the lung tissue model to support survival of DC, immunohistochemistry analyses detecting human leukocyte antigen (HLA)-DR molecules wereperformed on 8-�m cryosections from lung tissue models cultured either in the absence or presence of CSF-2 and IL-4. (A–C) Immunohistochemical staining oflung tissue model cryosections for HLA-DR is shown. Tissue sections from models containing DC and cultured in the absence (left) (A) or presence (middle) (B) ofexogenous growth factors or tissue sections from models without DC (C), were stained with an anti-HLA-DR antibody (brown) (original magnification �200).D: quantification of HLA-DR positive area within the lung tissue model is shown. To quantify the HLA-DR staining in tissue sections, acquired computerized imageanalyses (ACIA) were used. Bar graphs indicate the amount of HLA-DR-positive staining in tissue sections from models absent of DC or models with DC that werecultured in the absence or presence of exogenous growth factors. To determine whether cytokines and chemokines important for DC survival and function are expressedin the lung tissue model, RNA was extracted and supernatants were collected from 3D models absent of DC, 3D models with DC, or control DC (cultured in mediumonly). Analyses of mRNA levels were determined in tissue and cells using real-time RT-PCR. E: bar graphs indicate the relative expression of CSF-2, IL-4, and CX3CL1mRNA accumulated in control DC or 3D models with or without DC. All results are representatives of at least 3 independent experiments. In each experiment, 6 tissuemodels were analyzed. Data presented in bar charts are shown as mean values in triplicate from 1 representative experiment (� SD). The statistical significance (1-wayANOVA) of the indicated P values was determined: *P 0.05, **P 0.01, ***P 0.005, ns � nonsignificant.
L232 DC REGULATION BY LUNG TISSUE-SPECIFIC CELLS
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00059.2011 • www.ajplung.org
containing 3D model to DC cultured in medium only revealedenhanced CCL18 expression within the 3D model (Fig. 7A).That CCL18 production by DC is regulated in the 3D modelwas further supported by the fact that CCL18 protein wasreadily detected in the culture supernatant of the lung tissuemodel with DC (Fig. 7B). Next, we analyzed the capacity ofDC in the 3D model to produce CCL17 and CCL22, chemo-kines that are expressed constitutively in thymus but are barelydetectable in other peripheral tissues at steady state and that areinduced during inflammatory reactions and associated to hu-man skin and pulmonary inflammatory diseases (13, 21). Com-pared with DC only, the expression of CCL17 and CCL22 was
relatively low in lung tissue models regardless of whether themodel was set up with or without DC (Fig. 7, C and E).Furthermore, the levels of CCL17 and CCL22 protein werereduced in the culture supernatants of 3D models with DCcompared with DC cultured in medium only (Fig. 7, D and F).Together these data suggest that the 3D model has the capacityto differentially regulate the production of chemokines in DC,and this involves enhanced expression of CCL18 and a reducedexpression of CCL17 and CCL22.
Soluble components secreted from the lung tissue model areresponsible for the induction of CCL18 in DC. Next, wefocused our studies to determine whether soluble tissue-de-
Fig. 5. Quantification of DC longevity in thelung tissue model. DC labeled with the far-redcell tracker, dimethyl dodecylamine oxide-syccinimidyl, were implanted in models andvisualized by live imaging confocal micros-copy over time. A: representative 3D imagesacquired at days 1, 3, 5, and 8 of culture areshown. B: microscopy projections of 3D mod-els that were used to enumerate DC within adefined field are shown. Each defined area hadto include a minimum of 250 cells. C: bargraphs indicate the total numbers of DC in thelung tissue models. DC were quantified byenumerating far-red fluorescently labeled pro-pidium iodide-negative cells in each definedarea and multiplied with factor reflecting thetotal 3D tissue model area. To determine ex-pression of DC surface molecules, HLA-DR,DC-specific ICAM-grabbing nonintegrin (DC-SIGN), and CD11c, RNA was extracted from3D models absent of DC, 3D models with DC,or control DC (cultured in medium only). Anal-yses of mRNA were determined in tissue usingreal-time RT-PCR. D: bar graphs indicate therelative expression of HLA-DR, DC-SIGN, andCD11c mRNA accumulated in control DC and3D models with or without DC. All results arerepresentatives of at least 3 independent ex-periments. Data presented in bar charts areshown as mean values in triplicate from 1representative experiment (� SD). The statis-tical significance (1-way ANOVA) of the in-dicated P values was determined: *P 0.05,ns � nonsignificant.
L233DC REGULATION BY LUNG TISSUE-SPECIFIC CELLS
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00059.2011 • www.ajplung.org
rived components were responsible for the selective inductionof enhanced CCL18 production in DC. When the impact ofsoluble tissue model-derived components on CCL18 produc-tion by DC was monitored, we found that exposure of DC tomedium conditioned with the lung tissue model induced en-hanced CCL18 mRNA expression (Fig. 8A) and CCL18 pro-tein secretion by DC (Fig. 8B). Thus the lung tissue modelsecretes compounds involved in regulating CCL18 produc-tion by DC. To determine whether the composition of cells,i.e., lung epithelial cells and fibroblasts, into an AE 3Dstructure is necessary for the induction of enhanced produc-tion of CCL18 in DC, DC were also exposed to mediumconditioned with air-exposed epithelial cells or fibroblast aswell as epithelial cells and fibroblasts cultured as a mixed(1:1) submerged monolayer. This revealed that only thecomplete lung tissue model has the capacity to conditionmedium that can be used to induce enhanced CCL18 ex-pression in DC (Fig. 8C).
DISCUSSION
Tissue-specific niches, such as gut and lung mucosa, areincreasingly recognized as influencing a broad range of events
involved in immune homeostasis and inflammation as well asthe host response to infection (18, 30, 33). Although, in vitro3D organotypic lung models have been established in the pastand are commercially available ranging from models of normalairway epithelium to models of lung cancer, models withrelevant features and functions of the human airway wallcombined with human immune cells are lacking. Here, we haveestablished a 3D organotypic lung tissue model allowing stud-ies of DC regulation by lung tissue specific cells. We first showedthat molecules involved in the formation of essential structures forlung tissue function are produced and deposited in the model. Wethen showed that the lung tissue model supports the survival ofDC independent of exogenous growth factors and that DC caneasily be identified in the tissue model being strategically locatedunderneath, within, and on top of the epithelium. We also showedthat the lung tissue model regulates production of the chemokinesCCL17, CCL18, and CCL22 in DC. Thus this lung tissue modelis well suited for a wide range of studies, including those on tissueorganization and remodeling as well as the role of tissue-derivedcomponents regulating DC functional properties within the localtissue microenvironment during homeostasis and in response topathogens.
Fig. 6. DC are widely distributed throughoutthe epithelial layer within the lung tissuemodel. The distribution of DC in the lungtissue model was visualized by immunofluo-rescence microscopy. A: tissue model sec-tions were stained with an anti-HLA-DR an-tibody (green) (original magnification �200).B: tissue model sections were stained with ananti-HLA-DR antibody (green) in combina-tion with an anti-DC-SIGN antibody (red)(original magnification �200). C: tissuemodel sections were stained with an anti-CD11c antibody (green) in combination withan anti-DC-SIGN antibody (red) (originalmagnification �200). Higher magnificationshows a DC in close association with epithe-lial cells at the mucosal surface (inset: origi-nal magnification �1,000). D: tissue modelsections were stained with an anti-CX3CR1antibody (green). The distribution of DCwithin the lung tissue model was also visual-ized in live tissue using multiphoton micros-copy. DAPI was used to reveal cell nuclei.E and F: imaging was performed with modelscomposed of tdTomato expressing epithelialcells (red) and carboxyfluorescein diacetatesuccinimidyl ester-labeled DC (green). A topview (E) and a side view (F) of the lung tissuemodel are shown (original magnification�200). All results are representatives of atleast 3 independent experiments.
L234 DC REGULATION BY LUNG TISSUE-SPECIFIC CELLS
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00059.2011 • www.ajplung.org
The finding that our human 3D organotypic lung model hasthe capacity to support survival and allow functional studies ofhuman DC has important implications for increasing our under-standing of local immune regulation in lung tissue. This novel 3Dorganotypic lung model composed of human lung epithelial cellsand fibroblasts in combination with human DC shares similaritieswith real tissue, including the overall structure and deposition ofmatrix proteins essential for lung tissue function and forms afunctional mucosal barrier including production of proteins nec-essary for tight junction and adhesion junction formation. In thiscontext, the 3D AE tissue model provides a unique system inwhich to further elucidate mechanisms of tissue organization andremodeling at steady state and in response to external stimuli,including viral and bacterial infection.
Coordinated expression of chemokines by nonimmune andimmune cells is essential in mediating adhesion and migration
of diverse populations of immune cells from blood to tissue atsteady state and during inflammation and infection. The secondmajor outcome from this study is that this organotypic modelregulates the capacity of DC to produce one homeostaticchemokine, CCL18, and two inducible chemokines, CCL17and CCL22. CCL18 is a constitutively expressed chemoattrac-tant produced by immune cells and predominantly associatedto lymphnode and lung tissue, and that is detected at highlevels in human serum at homeostatic conditions (20). Theinfluence of tissue in the involvement of regulating productionof CCL18 in DC was confirmed by demonstrating that DC inthe organotypic model make more CCL18 than those culturedunder conventional conditions. This finding, therefore, sup-ports the fact that the lung tissue model microenvironmentinduces changes to DC and selectively modulates the produc-tion of chemokines resembling physiological conditions (13,20, 21). Further support of a functional distinction between DCin the 3D organotypic model compared with DC cultured underconventional conditions comes from our findings that implant-ing DC in the model reduced the levels of CCL17 and CCL22detected. CCL17 and CCL22 are not detectable at steady statebut are induced during inflammatory reactions and associatedto human skin and pulmonary inflammatory diseases (16, 41),thus implicating that specific tissue is able to indirectly exertimmune regulatory function via their influence on DC functionlocally. Furthermore, the results from our study also demon-strate that soluble components secreted by 3D organotypicmodels are responsible for the enhanced production of CCL18in DC. Although, differential expression of cytokines andchemokines by DC in response to monolayer of lung epithelialcells has been observed (26, 29), the effect of 3D tissue modelson DC function is limited. Therefore, we investigated whetherthere are differences between the 3D organotypic model, AEepithelial cells, AE fibroblasts, or submerged monolayer cul-ture of epithelial cells and fibroblasts in terms of altering thecapacity of DC to produce CCL18. Medium conditioned withthe 3D organotypic model was more potent than conditionedmedium from the single cell cultures or the monolayer culture,
Fig. 7. Chemokine expression by DC is regulated by the lung tissue microen-vironment. To determine whether chemokine production is modulated in DC inresponse to the lung tissue microenvironment, RNA was extracted and super-natants were collected from 3D models absent of DC, 3D models with DC, orcontrol DC (cultured in medium only). Analyses of mRNA and protein levelswere determined in tissue and supernatants using real-time RT-PCR andELISA, respectively. A and B: bar graphs indicate the relative expression ofCCL18 mRNA accumulated in control DC or 3D model (A) and the concen-tration of CCL18 in control DC or 3D model culture supernatants. C–F: bargraphs indicate the relative expression of CCL17 (C) and CCL22 (E) mRNAaccumulated in control DC or 3D model culture supernatants and the concen-tration of CCL17 (D) and CCL22 (F) in control DC and 3D model culturesupernatants. The lung model was set up in the absence of CSF-2 and IL-4, andculture supernatants were conditioned for 48 h before protein quantification.Control DC were cultured in the absence of CSF-2 and IL-4 for 24 or 48 hbefore RNA and protein analyses. Before RNA extraction, control DC, 1.5 �105 (which is a similar number of cells that was implanted in each 3D modelduring set up), were mixed with 3D models composed of fibroblasts andepithelial cells only. All results are representatives of 3 independent experi-ments, and DC from a total of 6 donors have been used. Data presented in barcharts are shown as mean values of mRNA or protein levels in triplicate fromthe 3 independent experiments (� SD). The statistical significance (1-wayANOVA) of the indicated P values was determined: *P 0.05, **P 0.01,***P 0.005.
L235DC REGULATION BY LUNG TISSUE-SPECIFIC CELLS
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00059.2011 • www.ajplung.org

in promoting enhanced CCL18 production, suggesting that the3D organotypic model secrets additional factors contributing tothis process, thus indicating that the complex structure of thelung tissue model favors the production of factors involved inregulating CCL18 production in DC under homeostatic condi-tions. This finding is quite intriguing and merits further studies.
In addition to 3D organotypic tissue models, various alter-native cell culture systems are available as tools for studyingthe impact of local tissue on immune cell function. Eachsystem has its distinct advantages and disadvantages, and thechoice of system depends on various factors, including theexperimental question being addressed and the cost, training,and expertise required to successfully establish, validate, andapply the model. One common technique involves utilizingmonolayers that are permitting infection but are usually basedon a single nonpolarized cell type in the absence of 3D cellularinteractions. Culture in Transwells allows culturing of mono-layers on a permeable membrane that, unlike traditional mono-layers systems, allows the cells to develop an apical-basalpolarity. These polarized surfaces can be independently ex-posed to different factors, including pathogens and toxins, andthereby exposing the cells in a physiologically relevant settingand taking into account transepithelial migration (10). Onedisadvantage of the method is that these cultures are oftenbased on one type of cell only. Also, such models lackextensive 3D cellular interactions that are achieved with 3Dmatrix scaffold cultures, which are crucial for normal tissuefunction as well as infection processes (8).
The engineering of organotypic models involves implantingof cells or tissue into a 3D matrix scaffold composed ofcollagen, extracts of extracellular matrix, synthetic or semisyn-
thetic materials, or a combination of these materials. However,models with relevant features and functions of the humanairway wall combined with human immune cells are lacking.In this study, therefore, we sought to develop a novel human3D lung tissue model that utilizes appropriate human cellpopulations, including bronchial epithelial cells, fibroblasts,and human immune cells. In our model epithelial (16HBE) andfibroblast (MRC-5) cell lines are used, as they allow forreproducibility compared with using potentially impure pri-mary cells. The choice of this epithelial line was based on thefact that 16HBE cells resemble primary lung epithelial cells(38) and thus are a valuable and reproducible model of normallung epithelial tissue (7). Although, quite laborsome, thisrelatively inexpensive approach provides a platform that alsoallows studies in conjunction with manipulation of gene expres-sion in tissue-specific nonimmune cells. Thus the model is wellsuited for studies on tissue-specific regulation of human DCfunctional properties and migratory behavior in a physiologicallyrelevant setting previously not achievable. Furthermore, the modelallows exposure of cells under physiological conditions whereimportant aspects of host-pathogen interactions can be monitoredin real time. In conclusion, we have established a human 3Dorganotypic lung model in which tissue-specific mechanisms thatregulate DC function locally during steady state and in response toinflammation can be studied. By further understanding the mech-anisms whereby specific tissue niches control the functional prop-erties of DC, we might uncover potential targets for manipulatingthe DC function in therapeutic settings to restore tissue homeo-stasis not only in acute and chronic infectious diseases but alsoother chronic inflammatory diseases.
Fig. 8. Chemokine expression by DC is reg-ulated by soluble components secreted fromthe 3D organotypic model. To determinewhether chemokine production is modulatedin DC in response to soluble compounds pro-duced by the tissue-specific cells in the 3Dorganotypic model, supernatants were col-lected from tissue models absent of DC, AEepithelial cells, AE fibroblasts, or submergedmonolayer cultures of epithelial cells and fi-broblasts. DC were stimulated with culturesupernatants for 24–48 h followed by thedetection of mRNA accumulation and proteinsecretion real-time RT-PCR and ELISA, re-spectively. A: bar graphs indicate the accumu-lation of CCL18 mRNA in DC stimulatedwith culture supernatants from the 3D orga-notypic model or medium only. B: bar graphsindicate the amount of CCL18 protein accu-mulated from DC stimulated with culture super-natants from the 3D organotypic model or me-dium only. C: bar graphs indicate the accumu-lation of CCL18 mRNA in DC stimulated for24 h with medium only or culture supernatants(48 h-conditioning) as indicated. Before stimu-lation DC were cultured in the absence of GM-CSF and IL-4 for 24 h. All results are represen-tatives of at least 3 independent experiments,and DC from a total of 5 donors have been used.Data presented in bar graphs are shown as meanvalues of mRNA levels in triplicate from the 3independent experiments (� SD). The statisticalsignificance (1-way ANOVA) of the indicated Pvalues was determined: *P 0.05, **P 0.01.
L236 DC REGULATION BY LUNG TISSUE-SPECIFIC CELLS
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00059.2011 • www.ajplung.org

ACKNOWLEDGMENTS
We thank Dr. Oscar Hammarfjord for critical review of the manuscript.
GRANTS
This work is supported by grants from the Swedish Research Council, theKarolinska Institutet, Stockholm County Council, The Swedish Fund forResearch Without Animal Experiments, Erik och Edit Fernströms Foundation(to M. Svennson). M. Coles is supported by funding from the MedicalResearch Council (G0601156). A. T. Nguyen Hoang is a recipient of aKarolinska postgraduate studentship. J. Juarez and L. Bosnjak are recipients ofMarie Curie International Incoming Fellowships.
DISCLOSURES
No conflicts of interest, financial or otherwise are declared by the authors.
REFERENCES
1. Banchereau J, Steinman RM. Dendritic cells and the control of immu-nity. Nature 392: 245–252, 1998.
2. Birkness KA, Deslauriers M, Bartlett JH, White EH, King CH, QuinnFD. An in vitro tissue culture bilayer model to examine early events inMycobacterium tuberculosis infection. Infect Immun 67: 653–658, 1999.
3. Bjork L, Fehniger TE, Andersson U, Andersson J. Computerizedassessment of production of multiple human cytokines at the single-celllevel using image analysis. J Leukoc Biol 59: 287–295, 1996.
4. Black AF, Bouez C, Perrier E, Schlotmann K, Chapuis F, Damour O.Optimization and characterization of an engineered human skin equiva-lent. Tissue Eng 11: 723–733, 2005.
5. Choe MM, Sporn PH, Swartz MA. Extracellular matrix remodeling bydynamic strain in a three-dimensional tissue-engineered human airwaywall model. Am J Respir Cell Mol Biol 35: 306–313, 2006.
6. Choe MM, Tomei AA, Swartz MA. Physiological 3D tissue model of theairway wall and mucosa. Nat Protoc 1: 357–362, 2006.
7. Cozens AL, Yezzi MJ, Kunzelmann K, Ohrui T, Chin L, Eng K,Finkbeiner WE, Widdicombe JH, Gruenert DC. CFTR expression andchloride secretion in polarized immortal human bronchial epithelial cells.Am J Respir Cell Mol Biol 10: 38–47, 1994.
8. Cukierman E, Pankov R, Stevens DR, Yamada KM. Taking cell-matrixadhesions to the third dimension. Science 294: 1708–1712, 2001.
9. Dongari-Bagtzoglou A, Kashleva H. Development of a highly reproduc-ible three-dimensional organotypic model of the oral mucosa. Nat Protoc1: 2012–2018, 2006.
10. Dvorak A, Tilley AE, Shaykhiev R, Wang R, Crystal RG. Do airwayepithelium air-liquid cultures represent the in vivo airway epitheliumtranscriptome? Am J Respir Cell Mol Biol 44: 465–473, 2010.
11. Fantin A, Vieira JM, Gestri G, Denti L, Schwarz Q, Prykhozhij S,Peri F, Wilson SW, Ruhrberg C. Tissue macrophages act as cellularchaperones for vascular anastomosis downstream of VEGF-mediatedendothelial tip cell induction. Blood 116: 829–840, 2010.
12. GeurtsvanKessel CH, Lambrecht BN. Division of labor between den-dritic cell subsets of the lung. Mucosal Immunol 1: 442–450, 2008.
13. Godiska R, Chantry D, Raport CJ, Sozzani S, Allavena P, Leviten D,Mantovani A, Gray PW. Human macrophage-derived chemokine(MDC), a novel chemoattractant for monocytes, monocyte-derived den-dritic cells, and natural killer cells. J Exp Med 185: 1595–1604, 1997.
14. Gray TE, Guzman K, Davis CW, Abdullah LH, Nettesheim P. Muco-ciliary differentiation of serially passaged normal human tracheobronchialepithelial cells. Am J Respir Cell Mol Biol 14: 104–112, 1996.
15. Griffith LG, Swartz MA. Capturing complex 3D tissue physiology invitro. Nat Rev Mol Cell Biol 7: 211–224, 2006.
16. Guttman-Yassky E, Lowes MA, Fuentes-Duculan J, Whynot J, No-vitskaya I, Cardinale I, Haider A, Khatcherian A, Carucci JA, Berg-man R, Krueger JG. Major differences in inflammatory dendritic cellsand their products distinguish atopic dermatitis from psoriasis. J AllergyClin Immunol 119: 1210–1217, 2007.
17. Hammad H, Chieppa M, Perros F, Willart MA, Germain RN, Lam-brecht BN. House dust mite allergen induces asthma via Toll-like receptor4 triggering of airway structural cells. Nat Med 15: 410–416, 2009.
18. Hammad H, Lambrecht BN. Dendritic cells and epithelial cells: linkinginnate and adaptive immunity in asthma. Nat Rev Immunol 8: 193–204, 2008.
19. Hartsock A, Nelson WJ. Adherens and tight junctions: structure, functionand connections to the actin cytoskeleton. Biochim Biophys Acta 1778:660–669, 2008.
20. Hieshima K, Imai T, Baba M, Shoudai K, Ishizuka K, Nakagawa T,Tsuruta J, Takeya M, Sakaki Y, Takatsuki K, Miura R, OpdenakkerG, Van Damme J, Yoshie O, Nomiyama H. A novel human CCchemokine PARC that is most homologous to macrophage-inflammatoryprotein-1 alpha/LD78 alpha and chemotactic for T lymphocytes, but notfor monocytes. J Immunol 159: 1140–1149, 1997.
21. Imai T, Yoshida T, Baba M, Nishimura M, Kakizaki M, Yoshie O.Molecular cloning of a novel T cell-directed CC chemokine expressed inthymus by signal sequence trap using Epstein-Barr virus vector. J BiolChem 271: 21514–21521, 1996.
22. Iwasaki A. Mucosal dendritic cells. Annu Rev Immunol 25: 381–418,2007.
23. Kleinman HK, Philp D, Hoffman MP. Role of the extracellular matrix inmorphogenesis. Curr Opin Biotechnol 14: 526–532, 2003.
24. Lambrecht BN, Hammad H. Biology of lung dendritic cells at the originof asthma. Immunity 31: 412–424, 2009.
25. Loffler B, Hussain M, Grundmeier M, Bruck M, Holzinger D, VargaG, Roth J, Kahl BC, Proctor RA, Peters G. Staphylococcus aureuspanton-valentine leukocidin is a very potent cytotoxic factor for humanneutrophils. PLoS Pathog 6: e1000715 2010.
26. Mayer AK, Bartz H, Fey F, Schmidt LM, Dalpke AH. Airway epithe-lial cells modify immune responses by inducing an anti-inflammatorymicroenvironment. Eur J Immunol 38: 1689–1699, 2008.
27. Nguyen Hoang AT, Liu H, Juarez J, Aziz N, Kaye PM, Svensson M.Stromal cell-derived CXCL12 and CCL8 cooperate to support increaseddevelopment of regulatory dendritic cells following Leishmania infection.J Immunol 185: 2360–2371, 2010.
28. Pflicke H, Sixt M. Preformed portals facilitate dendritic cell entry intoafferent lymphatic vessels. J Exp Med 206: 2925–2935, 2009.
29. Qiao J, Li A, Jin X. TSLP from RSV-stimulated rat airway epithelial cellsactivates myeloid dendritic cells. Immunol Cell Biol 89: 231–238 2011.
30. Rimoldi M, Chieppa M, Salucci V, Avogadri F, Sonzogni A, Sampi-etro GM, Nespoli A, Viale G, Allavena P, Rescigno M. Intestinalimmune homeostasis is regulated by the crosstalk between epithelial cellsand dendritic cells. Nat Immunol 6: 507–514, 2005.
31. Roskelley CD, Desprez PY, Bissell MJ. Extracellular matrix-dependenttissue-specific gene expression in mammary epithelial cells requires bothphysical and biochemical signal transduction. Proc Natl Acad Sci USA 91:12378–12382, 1994.
32. Schutyser E, Richmond A, Van Damme J. Involvement of CC chemo-kine ligand 18 (CCL18) in normal and pathological processes. J LeukocBiol 78: 14–26, 2005.
33. Svensson M, Kaye PM. Stromal-cell regulation of dendritic-cell differ-entiation and function. Trends Immunol 27: 580–587, 2006.
34. Svensson M, Maroof A, Ato M, Kaye PM. Stromal cells direct localdifferentiation of regulatory dendritic cells. Immunity 21: 805–816, 2004.
35. Timpl R, Rohde H, Robey PG, Rennard SI, Foidart JM, Martin GR.Laminin—a glycoprotein from basement membranes. J Biol Chem 254:9933–9937, 1979.
36. Tomasek JJ, Gabbiani G, Hinz B, Chaponnier C, Brown RA. Myofi-broblasts and mechano-regulation of connective tissue remodeling. NatRev Mol Cell Biol 3: 349–363, 2002.
37. Uller L, Leino M, Bedke N, Sammut D, Green B, Lau L, Howarth PH,Holgate ST, Davies DE. Double-stranded RNA induces disproportionateexpression of thymic stromal lymphopoietin vs. interferon-beta in bron-chial epithelial cells from donors with asthma. Thorax 65: 626–632.
38. Wan H, Winton HL, Soeller C, Stewart GA, Thompson PJ, GruenertDC, Cannell MB, Garrod DR, Robinson C. Tight junction properties ofthe immortalized human bronchial epithelial cell lines Calu-3 and16HBE14o. Eur Respir J 15: 1058–1068, 2000.
39. Wang N, Stamenovic D. Contribution of intermediate filaments to cellstiffness, stiffening, and growth. Am J Physiol Cell Physiol 279: C188–C194, 2000.
40. Wrenshall L. Role of the microenvironment in immune responses totransplantation. Springer Semin Immunopathol 25: 199–213, 2003.
41. Yogo Y, Fujishima S, Inoue T, Saito F, Shiomi T, Yamaguchi K,Ishizaka A. Macrophage derived chemokine (CCL22), thymus and acti-vation-regulated chemokine (CCL17), and CCR4 in idiopathic pulmonaryfibrosis. Respir Res 10: 80, 2009.
42. Yoon JH, Moon HJ, Seong JK, Kim CH, Lee JJ, Choi JY, Song MS,Kim SH. Mucociliary differentiation according to time in human nasalepithelial cell culture. Differentiation 70: 77–83, 2002.
L237DC REGULATION BY LUNG TISSUE-SPECIFIC CELLS
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00059.2011 • www.ajplung.org
Copyright © 2022 FDOKUMEN




















