
Definition and comparison of the phosphorylation sites of the A and B chains of bovine α-crystallin
10
Exp. Eye Res. (1988) 46, 199-208 Definition and Comparison of the Phosphorylation Sites of the A and B Chains of Bovine a-Crystallin RAITL CHIESA, MARY ANN GAWINOWlcz-KOLKS*, NORMAN J. KLEIMAN AND ABRAttAM SPECTOR t Biochemistry and Molecular Biology Laboratory, Department of Ophthalmology and *Center of Reproductive Sciences, Protein Chemistry Core Laboratory, College of Physicians and Surgeons, Columbia University, 630W. 168th St, New York, NY 10032, U.S.A. (Received 8 June 1987 and accepted 10 September 1987) The major phosphorylation sites of bovine a-crystallin Ser122in the A chain, Ser s9 and Ser 4a and/ or Ser45in the B chain have been previously characterized. Further analysis of total a-crystallin, isolated from the cortex of calf lenses incubated in the presence of [a2P]orthophosphate, demonstrated the presence of additional phosphorylation sites in both chains. At least three additional phosphorylation sites were found in the A chain and at least one in the B chain. These additional sites accounted for approximately 25 % of the radioactivity incorporated in the protein. Two general sequences were found in most phosphorylation sites of both chains of a- crystallin : (Arg/Lys)-(X)-Pro-Ser and Ser-(X)-Ser-Leu-Ser. In spite of the 57 % homology in the sequences of the A and B chains, the phosphorylation sites are located, in the A polypeptide, at the C-terminal third and in the B polypeptide, at the N-terminal third. The alignment of the regions containing the phosphorylation sites of both chains (C-terminal third of the A and N- terminal third of the B chain) revealed an unexpected similarity in the relative positions of the sites in each chain. Key words : cAMP; lens; post-translational modification; homology. 1. Introduction It is now apparent that the post-translational modification of the bovine a-crystallin primary gene products A s and B~ that lead to the formation of A 1 and B1 respectively, is indeed the result of a specific cAMP-dependent phosphorylation process. Previously A 1 was thought to arise from the A2 chain by a spontaneous non-enzymatic deamidation of the residues Asn 12a and Gln i~e (Bloemendal, Berns, van der Ouderaa and de Jong, 1972 ; van Kleef, de Jong and Hoenders, 1975). By analogy, but without experimental evidence, B1 was assumed to be a deamidated form of the B~ chain (Stauffer, Rothschild, Wandel and Spector, 1974; Delcour and Bouchet, 1978). Furthermore, these non-enzymatic transformations were interpreted as expressions of the aging process (van Kleef et al., 1975). Experiments performed in this laboratory have demonstrated the cAMP-dependent phosphorylation of a-crystallin in soluble fractions from calf lens outer cortex (Spector, Chiesa, Sredy and Garner, 1985). Furthermore, comparative structural analysis of the A s and A 1 chains isolated from native and in vitro phosphorylated a- crystallin have demonstrated that the post-translational product A 1 is a phos- phorylated form of the primary gene product A s (Chiesa and Spector, 1986 ; Voorter, Mulders, Bloemendal and de Jong, 1986; Chiesa, Gawinowicz-Kolks and Spector, 1987). While no phosphate is present in As, A 1 contains one mole of phosphate per mole of polypeptide. This phosphate is bound to the residue Set 12~ of the polypeptide r To whom correspondence and requests for reprints should be addressed. 0014-4835/88/020199+ 10 $03.00/0 1988 Academic Press Limited
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Definition and comparison of the phosphorylation sites of the A and B chains of bovine α-crystallin

Exp. Eye Res. (1988) 46, 199-208
Definit ion and Comparison of the Phosphorylat ion Sites of the A and B Chains of Bovine a-Crystall in
RAITL C H I E S A , MARY ANN GAWINOWlcz-KOLKS*, NORMAN J. KLEIMAN AND ABRAttAM SPECTOR t
Biochemistry and Molecular Biology Laboratory, Department of Ophthalmology and *Center of Reproductive Sciences, Protein Chemistry Core Laboratory, College of
Physicians and Surgeons, Columbia University, 630W. 168th St, New York, N Y 10032, U.S.A.
(Received 8 June 1987 and accepted 10 September 1987)
The major phosphorylation sites of bovine a-crystallin Ser 122 in the A chain, Ser s9 and Ser 4a and/ or Ser 45 in the B chain have been previously characterized. Further analysis of total a-crystallin, isolated from the cortex of calf lenses incubated in the presence of [a2P]orthophosphate, demonstrated the presence of additional phosphorylation sites in both chains. At least three additional phosphorylation sites were found in the A chain and at least one in the B chain. These additional sites accounted for approximately 25 % of the radioactivity incorporated in the protein. Two general sequences were found in most phosphorylation sites of both chains of a- crystallin : (Arg/Lys)-(X)-Pro-Ser and Ser-(X)-Ser-Leu-Ser. In spite of the 57 % homology in the sequences of the A and B chains, the phosphorylation sites are located, in the A polypeptide, at the C-terminal third and in the B polypeptide, at the N-terminal third. The alignment of the regions containing the phosphorylation sites of both chains (C-terminal third of the A and N- terminal third of the B chain) revealed an unexpected similarity in the relative positions of the sites in each chain.
Key words : cAMP; lens; post-translational modification; homology.
1. Introduct ion
I t is now apparen t tha t the pos t - t rans la t ional modification of the bovine a-crysta l l in p r imary gene products A s and B~ tha t lead to the format ion of A 1 and B1 respectively, is indeed the result of a specific cAMP-dependent phosphoryla t ion process. Previously A 1 was thought to arise from the A2 chain by a spontaneous non-enzymat ic deamidat ion of the residues Asn 12a and Gln i~e (Bloemendal, Berns, van der Ouderaa and de Jong, 1972 ; van Kleef, de Jong and Hoenders, 1975). By analogy, bu t wi thout exper imental evidence, B1 was assumed to be a deamidated form of the B~ chain (Stauffer, Rothschild, Wande l and Spector, 1974; Delcour and Bouchet, 1978). Fur thermore , these non-enzymat ic t ransformat ions were in terpre ted as expressions of the aging process (van Kleef et al., 1975).
Exper iments performed in this labora tory have demons t ra ted the cAMP-dependent phosphoryla t ion of a-crysta l l in in soluble fractions from calf lens outer cortex (Spector, Chiesa, Sredy and Garner, 1985). Fur thermore , comparat ive s t ructural analysis of the A s and A 1 chains isolated from nat ive and in vitro phosphoryla ted a- crystal l in have demons t ra ted tha t the pos t - t rans la t ional product A 1 is a phos- phoryla ted form of the p r imary gene product A s (Chiesa and Spector, 1986 ; Voorter, Mulders, Bloemendal and de Jong, 1986; Chiesa, Gawinowicz-Kolks and Spector, 1987). While no phosphate is present in As, A 1 conta ins one mole of phosphate per mole of polypeptide. This phosphate is bound to the residue Set 12~ of the polypeptide
r To whom correspondence and requests for reprints should be addressed.
0014-4835/88/020199+ 10 $03.00/0 �9 1988 Academic Press Limited

200 R. CH][ESA ET AL.
chain (Voorter e t al., 1986; Chiesa, Gawinowicz-Kolks , K le iman and Spector ,
1987a). More recent ly , the phospho ry l a t i on sites of the a -c rys ta l l in B~ chain were identif ied
in exper imen t s carr ied ou t wi th label led a -c rys ta l l in ob ta ined from the ou te r cor tex of ca l f lenses incuba ted in the presence of [3~P]orthophosphate (Chiesa, Gawinowicz- Kolks , K l e i m a n and Spector , 1987b). The sequence analys is of 32P-labelled c h y m o t r y p t i c pep t ides ob ta ined from to ta l a -c rys ta l l in has ind ica ted two ma jo r phosphory la t i on sites in the a -c rys ta l l in B chain. One of the sites is the residue Ser ~9. The o ther si te has been assigned to residues Ser 4a a n d / o r Set 45. Thus, while d e a m i d a t i o n reac t ions m a y occur, the fo rmat ion of the A 1 and B1 chains can def in i t ive ly be a t t r i b u t e d to phospho ry l a t i on a t specific sites.
A l though the ma jo r phosphory la t i on sites of the A and B chains of a -c rys ta l l in have been es tabl ished, o ther a p p a r e n t l y minor phosphory la t i on sites were found in bo th chains. I n th is communica t ion , the ident i f ica t ion of these add i t iona l phos- pho ry l a t i on sites is repor ted . The re la t ive posi t ions of the phosphory la t i on sites in the p r i m a r y s t ruc ture of bo th A and B chains suggests t h a t there m a y be a conformat iona l specif ici ty of the a -c rys ta l l in macromolecule charac te r i s t ic of this phosphory la t i on process.
2. M a t e r i a l s a n d M e t h o d s
32P-labelling of ot-crystallin
The labelling of a-crystall in was carried out in lens culture as previously described (Chiesa, Gawinowicz-Kolks, Kleiman and Spector, 1987a). Bovine eyes from approximately 16-week- old calves were obtained from a local slaughter house. Within 3 hr after slaughter, the lenses were removed under aseptic conditions from the eyeballs by dissection using an anterior approach. A layer of vitreous (approximately 3 mm) was left at tached to the lens posterior surface. The lenses were incubated in minimum essential medium (MEM) containing 5 % fetal bovine serum (HyClone Laboratories). The medium was prepared fresh from amino acid solutions and a vitamin mix (Gibeo Laboratories) and contained 1-8 mM CaCI~, 5"3 mM KC1, 0"8 mM MgSO 4, 116 mM NaCl, i mM NaH2P04, 5-5 m~ glucose, 26 mM NaC% and 0"02 mM phenol red. Each lens was incubated in 19 ml of medium at 37~ under 5 % CO 2. After 6 hr of incubation, the medium was replaced with a fresh medium where the NaH~PO 4 concentration was reduced to 0-2 mM. Thirty minutes later, the medium was again replaced with the same low phosphate medium but containing 1"5 mCi ml -~ [a~P]orthophosphate (ICN). After 18 hr of incubation, the lenses were washed with ice cold saline and decapsulated, a-crystall in was isolated from the lens outer cortex (10-20 % wet weight) by gel filtration chromatography using a Sepharose CL6B (Pharmacia) column (Li, 1978). The protein solution was extensively dialysed, lyophylized and stored desiccated at -20~
Isolation of the A 1 and B 1 chains a-crystall in A 1 and B 1 chains were isolated from immobilized pH gradient isoelectric
focusing gels; 0"5mm thick pre-cast gels (Immobiline Dry Plate, pH 4"0-7'0, LKB) rehydrated with a freshly prepared 6 M urea solution were used. The preparative procedure, ' Immobiline-Canal Isoelectric Focusing ', described by Bartels and Bock in LKB Application Note 328, was followed. The position of A 1 and B1 in the gel was determined by fractionation of a small sample of labelled total a-crystall in and detection of the bands by radioautography. Two slots were made by removing the portions of gel corresponding to the A 1 and B 1 bands. The slots were filled with a suspension of 4% Ultrodex (LKB) in 6 M urea. The gel was loaded with 2 mg of 32P-labelled total a-crystallin in a 2 x 10 mm band. After fractionation, the slurry was removed from the slots, loaded into small teflon columns and eluted with a 6 M urea solution. The eluant containing the radioactive A 1 and B~ fractions was subjected to dialysis followed by lyophylization.

a -CRYSTALLIN PHOSPHORYLATION 201
Proteolytic cleavage Total a-crystallin, A 1 and B~ were subjected to cleavage with chymotrypsin, as previously
described (Chiesa, Gawinowicz-Kolks and Spector, 1987) ; 10 mg m1-1 a~P-labelled a-crystallin were dissolved in 0"1 N N-ethylmorpholine-acetic acid buffer, pH 8"6, containing 0"1 mg ml -~ TPCK-treated a-chymotrypsin (Sigma Chemical Company) and incubated at 37~ for 20 hr. The reaction was stopped by quick-freezing in an acetone dry ice bath. The same conditions were used to digest A 1 or B1 but the final protein and chymotrypsin concentrations were approximately 1 mg ml -~ and 0"02 mg ml -~, respectively.
Analytical separation of the chymotryptic peptides The mixtures of peptides resulting from total a-crystallin, A 1 o r B 1 chymotryptic digests
were analysed by ascending thin-layer chromatography (TLC) on 0"1 mm cellulose TLC plates (Merck) using 1-butanol : pyridine : acetic acid : water (15: 10: 3:12 ) as mobile phase.
Bidimensional separation of the a2P-labelled chymotryptic fragments from total a- crystallin was carried out on cellulose TLC plates by thin-layer electrophoresis (TLE) followed by ascending chromatography, as previously described (Chiesa, Gawinowicz-Kolks and Spector, 1987). Electrophoresis was carried out for 70 min at 20 W using a buffer containing pyridine : acetic acid :water (20 : 5 : 175) pH 5-6. Ascending chromatography was carried out using the solvent described above.
The a2P-labelled phosphopeptides were located in the TLC plates by radioautography using X~Omat AR film (Kodak).
Isolation of the a2P-labelled chymotryptic fragments from total a-crystallin The labelled phosphopeptides from total a-crystall in were preparatively isolated using
TLC followed by TLE; 4-5 mg of digested protein (450 ~l of reaction mixture) were applied to a cellulose TLC plate in 30 lanes (150/~g/lane) and subjected to ascending chromatography as described above. The plates were dried at 50~ under vacuum for 2 hr and subjected to radioautography. The areas of cellulose support containing the different a2P-labelled phosphopeptides were located in the plate using the radioautogram as a reference. The portions of cellulose containing the phosphopeptides were removed from the plate, packed in a small teflon column and eluted with the same solvent used for chromatography. Six asp_ labelled fractions (corresponding to components 1 through 6 in Fig. 1) were obtained by this procedure. The eluant corresponding to each fraction was lyophylized, re-suspended in a small volume of the same solvent, quanti tat ively applied to a second cellulose TLC plate in 10 lanes and subjected to electrophoresis as described above. The electrophoresis time was adjusted to maximize the separation of the different components. The plate was dried and the phosphopeptides located by radioautography. Single labelled components were obtained from fractions 1, 2, 4 and 5. Fractions 3 and 6 were further resolved into two components each, 3A, 3B and 6A, 6B, respectively (see Fig. 2). The phosphopeptides were recovered from the plate as indicated above and were subjected to amino acid sequence analysis.
The purity of the isolated phosphopeptides was verified before sequencing by TLE analysis using a buffer containing acetic acid :formic acid :water (78:25:897) pH 1"9 at 20 W for 30 min. Peptides were detected by radioautography and fluorescamine stain. A single radioactive spot accounting for at least 80 % of the fluorescamine positive material was observed in all the fractions. In addition, the sequence analysis of these samples demonstrated single peptide components and was, therefore, retrospectively considered a major criterion of purity.
Amino acid sequence determination The isolated phosphopeptides were subjected to amino acid sequence analysis in a gas
phase sequencer Model 470A from Applied Biosystems. The glass fiber filter disks were loaded with 3 mg Polybrene and 0"2 mg NaC1 (Biobrene Plus, Applied Biosystems) and pre- cycled three times using the O2CFIL program. Approximately 5 to 10 nM of peptide dissolved in 25% trifluoroacetic acid were applied to the pre-cycled filter and subjected to Edman degradation using the 02CPTH program. The PTH-amino acids resulting from the cleavage were analysed by reverse phase chromatography in a HPLC system.
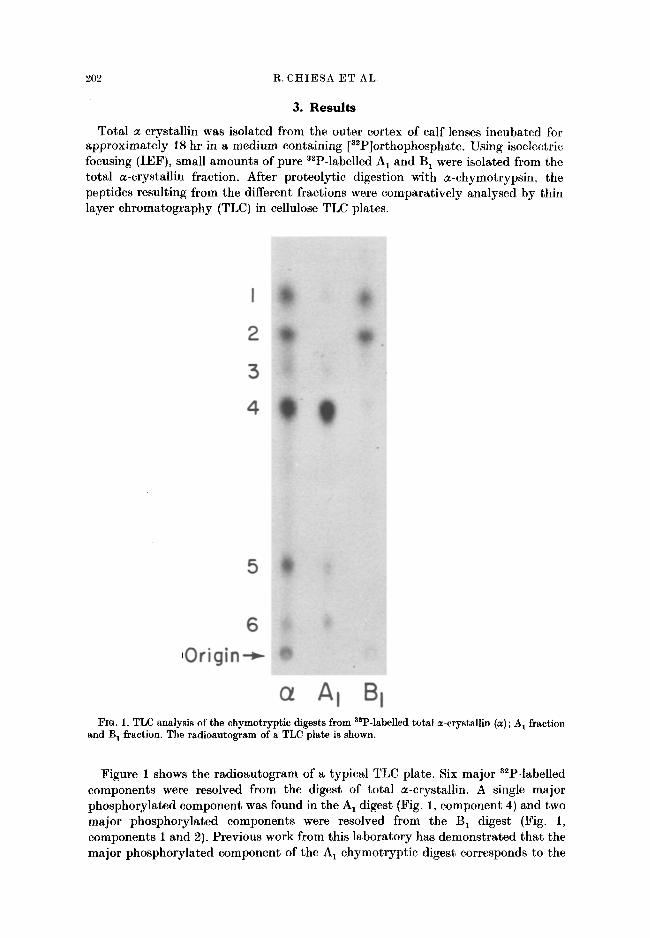
202 R. CHIESA ET AL.
3. Resul ts
Total a-crystallin was isolated from the outer cortex of calf lenses incubated for approximately 18 hr in a medium containing [3~P]orthophosphate. Using isoelectric focusing (IEF), small amounts of pure 3~P-labelled A 1 and B 1 were isolated from the total a-crystallin fraction. After proteolytic digestion with a-chymotrypsin, the peptides resulting from the different fractions were comparatively analysed by thin layer chromatography (TLC) in cellulose TLC plates.
I
2
3
4
5
6
Origin----
a A I I FIG. 1. TLC analysis of the chymotryptic digests from asP-labelled total a-crystallin (a) ; A 1 fraction
and B 1 fraction. The radioautogram of a TLC plate is shown.
Figure 1 shows the radioautogram of a typical TLC plate. Six major a2P-labelled components were resolved from the digest of total a-crystallin. A single major phosphorylated component was found in the A1 digest (Fig. 1, component 4) and two major phosphorylated components were resolved from the B1 digest (Fig. 1, components 1 and 2). Previous work from this laboratory has demonstrated tha t the major phosphorylated component of the A 1 chymotrypt ic digest corresponds to the

A
a-CRYSTALLIN PHOSPHORYLATION
B
203
i - - ~ E L E C T R O P H O R E S I S ~ +
(~1
03A
0 5
6 A ~ 6 B
- ~ E L E C T R O P H O R E S I S ~ +
Fro. 2. Two-dimensional analysis of a chymotryptic digest from asP-labelled total a-crystallin. The radioautogram of a plate (A) and its schematic drawing (B} are shown. Over 60 ninhydrin positive spots were detected by staining. Components 1, 2, 4 and 5 are single phosphopeptides. Note that components 3 and 6, shown in Fig. 1, were further resolved into 3A, 3B and 6A, 6B, respectively, in the second system. The spot located close to the center of the radioautogram was not consistently observed and, therefore, its sequence was not determined.
fragment 119-129 of the A polypeptide chain (Chiesa et al., 1987). This fragment contains the major phosphorylation site of the A chain, the residue Ser 122 (Chiesa et al., 1987; Chiesa et al., 1987a). The two phosphorylated components resolved in the digest of the B 1 fraction have been also identified previously (Chiesa et al., 1987b) and correspond to fragments 43-47 and 56-65 of the B chain. Therefore, the components 1, 2 and 4 in the chromatographic profile of total a-crystallin chymotrypt ic digest (Fig. 1) correspond to polypeptide fragments containing the major phosphorylation sites of the A and B chains.
In order to identify the minor components 3, 5 and 6 of the total a-crystallin chymotrypt ic digest, the following experiments were carried out. To determine the homogeneity of these minor phosphorylated components, a two-dimensional analysis of the total a-crystallin chymotrypt ic digest was performed. Aliquots of the digest were fractionated in cellulose plates by TLC (first dimension) followed by eleetrophoresis at pH 5"6 {TLE) (second dimension). The typical two-dimensional separation pat tern is shown in Fig. 2. No further purification of the components 1, 2, 4 and 5 was obtained, which is indicative of their degree of homogeneity. However, the components 3 and 6, which previously appeared homogeneous by chromatographic analysis, were further fractionated by electrophoresis and two phosphopeptides were resolved from each component, designated 3A, 3B, 6A and 6B.
To determine the structure of these phosphopeptides, they were isolated by preparat ive TLC followed by preparat ive TLE. The isolated phosphopeptides were analysed in a third system (TLE at pH 1"9) to verify the puri ty and then subjected to sequence analysis utilizing Edman degradation in a gas phase system. Figure 3 shows the sequence of the eight phosphopeptides isolated from the chymotryptic digest of asP-labelled total a-crystallin from calf lens outer cortex. The peptides 1, 2

204 R. CHIESA ET AL.
3 A
B CHAIN PEPTIDES
4 3 47
-SER-LEU-SER-PRO-PHE-
S6 55 -ARG-ALA-PR0-SER-TRP-ILE-ASP-THR-GLY-LEU-
2 9 3 8 -GLY-GLU-HIS-LEU-LEU-GLU-SER-ASP-LEU-PHE-
A CHAIN PEPTIDES
1 3 0 1 3 9
3B -SER-CYS-SER-LEU-SER-ALA-ASP-GLY-MET-LEU-
4 1 1 9 1 2 9
-ARG-LEU- PRO-SER-ASN-VAL-ASP-GLR- SER-ALA- LEU-
158 173 -ALA-ILE-PRO-VAL-SER-ARG-GLU-GLU-LYS-PRO-SER-SER-ALA- PRO-SER-SER
1( ;2 1 7 3
6 A -SER-ARG-GLU-GLU-LYS-PRO-SER-SER-ALA-PRO-SER-SER
142 161 ~ -SER-GLY-PR0-LYS-I LE-PR0- SER-GLY-VAL-ASP-ALA-GLY-HI S-SER-GLU-ARG-ALA-I LE- PRO-VAL-
Fro. 3. Amino acid sequences of the eight phosphopeptides isolated from a chymotryptic digest of "a~P-labelled total a-crystallin. Fragments 1, 2 and 3A correspond to the B chain. Fragments 3B, 4, 5, 6A and 6B correspond to the A chain. Note the overlap of fragments 5, 6A and 6B.
and 3A precisely correspond to fragments 43-47, 56-65 and 29-38 from the reported sequence of the B e chain (van der Ouderaa, de Jong, Hilderink and Bloemendal, 1974) (Fig. 4). As has been shown previously, the phosphopeptides 1 and 2 correspond to what appear to be the major phosphorylation sites of the B chain. The sequences of the phosphopeptides 3B, 4, 5, 6A and 6B corresponded, respectively, to fragments 130-139, 119-129, 158-173, 162-173 and 142-161 of the reported sequence of the A s chain (van der Ouderaa, de Jong and Bloemendal, 1973) (Fig. 4). Fragment 4 contains the major phosphorylation site of the A chain, Ser 1~.
Previous work has demonstrated that phosphoserine is the only phosphorylated amino acid detected in a2P-labelled a-crystallin preparations (Spector et al., 1985). Therefore, the phosphate group present in these fragments must be located in serine residues. Since most of the phosphorylated fragments contained more than one serine residue, it was not possible to make a definitive assignment of the phosphorylation sites in these situations. An exception, however, is f ragment 3A corresponding to residues 29-38 of the B chain which contains only the Ser aS. Fragment 5, corresponding to residues 158-173 of the A chain, resulted from a tryptic type cleavage at Arg 157, possibly due to the presence of a small amount of trypsin activity

a-CRYSTALLIN PHOSPHORYLATION 205
=-Crystailin A Chain
i0 20 30 40 50
I I I I I MDIAIQHPWFKRTLGPFYPSRLFDQFFGEGLFEYDLLPFLSSTISPYYRQ
5 1 SLFRTVLDSGISEVRSDRDKFVIFLDVKHFSPEDLTVKVQEDFVEIHGKH
i01 NERQDDHGYISREFHRRYRLPSNVDQSALSCSLSADGMLTFSGPKIPSGV I 4 I! 3n I I 6 n - -
151 DAGHSERAIPVSREEKPSSAPSS sA !
- 6 . I n 5
,,-Crystallin B Chain
10 20 30 40 50
I I I I I MDIAIHHPWIRRPFFPFHSPSRLFDQFFGEHLLESDLFPASTSLSPFYLR
I,~ 3 A - - - - - I I - - 1 - - - - I
51 PPSFLRAPSWIDTGLSENRLEKDRFSVNLNVKHFSPEELKVKVLGDVIEV z .... I
I01 HGKHEERQDEHGFISREFHRKYRIPADVDPLAITSSLSSDGVLTVNGPRK
151 QASGPERT I P I TREEKPAVTAAPKK
FIG. 4. Amino acid sequence of a-crystallin Az and B s chains according to van der Ouderaa et al. (1973) and v'an der Ouderaa et al. (1974), respectively. The a2P-labelled chymotryptic fragments obtained from total a-crystaUin are indicated.
in the chymotrypsin preparation. Fragment 6A appears to be the result of further cleavage of fragment 5 (Fig. 4).
The distribution of the labelled phosphate in the different fragments was determined. The eight phosphopeptides separated in the two-dimensional analysis of the total a-crystallin digest were quanti tat ively recovered from the TLC plate and subjected to radioactivity measurement. The relative distribution of the a~p radioactivity in these phosphopeptides is shown in Table 1. The total radioactivity recovered from the eight spots accounted for 60 % of the radioactivity loaded in the plate. Negligible radioactivity was found at the origin and elsewhere. Approximately 60--70 % of the recovered radioactivity was associated with peptides of the A chain

206 R. CHIESA ET AL.
TABL~ I
Distribution of 3~p radioactivity in the phosphorylated chymotryptic fragments from total ~-crystallin
Radioactivity
Fragment (cpm) (%)
1 187 14 2 201 15 3A 52 4 3 B 98 7 4 603 45 5 92 7 6A 49 4 I 6B 57 4 Total A chain 899 67 Total B chain 440 33
Since fragments 5 and 6A contain the same serine residues (see Fig. 3), the radioactivity associated to these residues represents 11% of the total.
and 30-40 % was associated with peptides of the B chain. Peptide 4 accounted for the largest fraction of the recovered radioactivity in a-crystallin, 45 %, corresponding to approximately 70 % of the radioactivity associated with the A chain. The other A chain peptides accounted for the remaining 30 %. Peptide 1 and peptide 2 from the B chain each had approximately 15 % of the recovered radioactivity in a-crystallin and accounted for over 85 % of the radioactivity associated with the B chain. The fragment 3A accounted for approximately the remaining 15 %.
4. D i s c u s s i o n
The results presented in this communication indicate that the phosphorylation of a-crystallin A and B chains takes place at least at seven different sites. In addition to Ser 12~ which appears to be the principal phosphorylation site, the A chain appears to be phosphorylated at another three sites, The B chain appears to be phosphorylated mainly at two sites: Ser 59 and Ser 4a or Ser 45, accounting for most of the 3~p incorporated in the B chain. In addition, a small proportion of the radioactivity was found in Ser a~.
I t is not apparent from these experiments whether single or multiple phos phorylation involving one or more of these residues takes place in a given molecule. However, the fact that A 1 and B 1 appear homogeneous when analyzed by IEF (Spector et al., 1985) supports the view that phosphorylation only takes place at a single residue in these molecules.
Two distinct sequence patterns appear to be present in most phosphorylation sites of both A and B chains of a-crystallin. One pattern is observed in the A chain in Ser 12e and Ser 14s and in the B chain in Ser 59. These sites are contained in the general sequence (Arg/Lys)-(X)-Pro-Ser.
This sequence is unusual for a physiologically relevant cAMP-dependent phos- phorylation site. Although basic amino acids (Arg and/or Lys) one or two residues away from the N-terminal end of the phosphorylated Ser (less frequently Thr) are common features of the physiologically relevant phosphorylation sites of cAMP-

a - C R Y S T A L L I N P H O S P H O R Y L A T I O N 207
dependent protein kinase (Glass and Krebs, 1980), the presence of Pro between the basic residues and the serine is unusual (Chessa et al., 1983). The only reported example of a cAMP-dependent phosphorylation site, where a Pro residue flanks the phosphorylated amino acid, is protein phosphatase inhibitor-1 from rat liver, which has the sequence Arg-Arg-Arg-Arg-Pro-Thr (Cohen, Ryla t t and Nimmo, 1977).
Another pat tern appears to be present in the phosphorylation site involving Ser 43 and Ser 45 in the B chain and Ser la~ Ser T M and Ser T M in the A chain. These sites display the sequence Ser-(X)-Ser-Leu-Ser. This is not a typical cAMP-dependent phos- phorylation site. Another kinase may be responsible for the phosphorylation at these sites. The presence of this general sequence in the phosphorylation sites of both A and B chains, however, suggests that these phosphorylations may be physiologically relevant.
The A and B chains of a-crystallin have a sequence homology of 57 % (van der Ouderaa et al., 1974). Furthermore, nine out of the 17 serine residues of the B chain correspond to serine residues in homologous regions of the A chain. Given the high similarity of both chains and the even distribution of the serine residues throughout the polypeptides, it would be expected that at least some of the phosphorylation sites of both chains would be located in homologous regions. I t is striking that this is not the case. The diagram in Fig. 5 is a representation of the relative position of the serine residues in the A and B chains aligned according to van der Ouderaa et al. (1974) to display the maximum homology. While the phosphorylation sites of the A chain are located in the C-terminal third of the polypeptide, the phosphorylation sites of the B chain are located in the N-terminal third of the molecule. Furthermore, when the fragment 117-173 of the A chain, which corresponds to the C-terminal end of the polypeptide is aligned to the fragment 28-84 of the B chain, corresponding to the N- terminal half of the molecule, a striking similarity in the relative positions of the phosphorylation sites of both chains is observed (Fig. 6).
43 45 59
122
�9 Non-Phosphoryloted Serine Residue O Possible Phosphoryloted Serine Residue A Phosphoryioted Serine Residue
Fro. 5. Relative position of the serine residues in the A and B chains of a-crystallin. The chains are aligned according to van der Ouderaa et al. (1974) to display the maximum homology. The numbers indicate the major phosphorylation sites.
A
I T 5 117 I
I * T T T t t ? IT I t - - R - P S . . . . S - - S - S L S - . . . . . . S - - K - P S . . . . G - S E . . . . . SR . . . . S S - - S S
B . . . . . . . S . . . . . S - S L S . . . . . . . S - - R - P S . . . . G-SE . . . . . . R-S . . . . . . . .
I * I T * I 2 8 8 4
FIo. 6. Alignment of the A chain C-terminal third (residues 117-173) to the segment 28-84 from the N-terminal half of the B chain. ( - ) Non-homologous amino acids; (*) major phosphorylation sites; (t) possible phosphorylation site,

208 R. CHIESA ET AL.
Newly synthesized calf a-crystallin, the only physically homogenous form of the a- crystallin, is a macromolecule with a MW of 7"5 • 105 (Stauffer et al., 1974). Analysis of the monomer composition of this macromolecule indicates that it is essentially composed of the non-phosphorylated A 2 and B 2 polypeptides (only trace amounts of A 1 can be detected) (Stauffer et al., 1974). Such observation suggests that the phosphorylation occurs after the formation of the aggregate structure. I f the phosphorylation does occur after the formation of the macromoleeule, the distribution of the phosphorylation sites suggests a different accessibility of the A and B chain segments where phosphorylation takes place. This observation would suggest that the A and B polypeptides may be packed in a different manner with the C-terminal region of the A chain and the N-terminal region of the B chain exposed to the environment.
A C K N O W L E D G M E N T S
This work was supported by grants from the National Eye Institute, National Institutes of Health, Bethesda, MD.
R E F E R E N C E S
Bloemendal, H., Berns, A. J. M., van der Ouderaa, F. and de Jong, W. W. (1972). Evidence for a non-genetic origin of the A 1 chains of a-crystallin. Exp. Eye Res. 14, 80-1.
Chessa, G., Borin, G., Marchiori, F., Meggio, F., Brunati, A. M. and Pinna, L.A. (1983). Synthetic peptides reproducing the site phosphorylated by cAMP-dependent protein kinase in protein phosphatase inhibitor-l. Eur. J. Biochem. 135,609-14.
Chiesa, R., Gawinowicz-Kolks, M. A., Kleiman, N. J. and Spector, A. (1987a). Identification of the specific phosphorylated serine in the bovine alpha crystallin A 1 chain. Curr. Eye Res. 5, 539-42.
Chiesa, R., Gawinowicz-Kolks, M.A., Kleiman, N.J. and Spector, A. (1987b). The phosphorylation sites of the B~ chain of bovine ze-crystallin. Biochem. Biophys. Res. Comm. 144, 1340-7.
Chiesa, R., Gawinowicz-Kolks, M. A. and Spector, A. (1987). The phosphorylation of the primary gene products of a-crystallin. J. Biol. Chem. 252, 1438-41.
Chiesa, R. and Spector, A. (1986). Phosphorylation is involved in the post-translational modification of alpha crystallin primary gene products. Invest. Ophthal. & Vis. Sci. 27, 215.
Cohen, P., Rylatt, D.B. and Nimmo, G.A. (1977). The hormonal control of glycogen metabolism: The amino acid sequence at the phosphorylation site of protein phosphatase inhibitor-1. FEBS Letters 76, 182-6.
Delcour, J. and Bouchet, H. (1978). Evidence for a post-translational origin of subunit ctB1 in the bovine lens a-crystallin. Exp. Eye Res. 26, 191-5.
Glass, D. B. and Krebs, E. G. (1980). Protein phosphorylation catalyzed by cyclic AMP- dependent and cyclic GMP-dependent protein kinases. Ann. Rev. Pharmacol. Toxicol. 20, 363-88.
Li, Lu-Ku (1978). Effect of sucrose on gel filtration of calf lens soluble proteins. Exp. Eye Res. 27, 553-66.
Spector, A., Chiesa, R., Sredy, J. and Garner, W.H. (1985). cAMP-dependent phos- phorylation of bovine lens alpha crystallin. Proc. Natl. Acad. Sci. USA 82, 4712-16.
Stauffer, J., Rothschild, C., Wandel, T. and Spector, A. (1974). Transformation of alpha- crystallin polypeptide chains with aging. Invest. Ophthal. 13, 13546.
van Kleef, F. S. M., de Jong, W. W. and Hoenders, H. J. (1975). Stepwise degradations and deamidation of the eye lens protein ct-crystallin in ageing. Nature 258, 264-66.
van der Ouderaa, F. J., de Jong, W. W. and Bloemendal, H. (1973). The amino acid sequence of the A S chain of bovine a-crystallin. Eur. J. Biochem. 39, 207-22.
van der Ouderaa, F. J., de Jong, W. W., Hilderink, A. and Bloemendal, H. (1974). Amino- acid sequence of the aB~ chain of bovine a-crystallin. Europ. J. Biochem. 49, 157-68.
Voorter, C. E. M., Mulders, J. W. M., Bloemendal, H. and de Jong, W.W. (1986). Some aspects of the phosphorylation of ct-crystallin A. Eur. J. Biochem. 160, 203-10.




















