
Chaperone-like activity and hydrophobicity of α-crystallin
10
Critical Review Chaperone-like Activity and Hydrophobicity of a -Crystallin G. Bhanuprakash Reddy, P. Anil Kumar and M. Satish Kumar* Biochemistry Division, National Institute of Nutrition, Hyderabad, India Summary a -Crystallin, a prominent member of small heat shock protein (sHsp) family and a major structural protein of the eye lens is a large polydisperse oligomer of two isoforms, a A- and a B-crystallins. Numerous studies have demonstrated that a -crystallin functions like a molecular chaperone in preventing the aggregation of various proteins under a wide range of stress conditions. The molecular chaperone function of a -crystallin is thus considered to be vital in the maintenance of lens transparency and in cataract prevention. a -Crystallin selectively interacts with non-native proteins thereby preventing them from aggregation and helps maintain them in a folding competent state. It has been proposed and generally accepted that a -crystallin suppresses the aggregation of other proteins through the interaction between hydrophobic patches on its surface and exposed hydrophobic sites of partially unfolded substrate protein. However, a quantifiable relationship between hydrophobi- city and chaperone-like activity remains a matter to be concerned about. On an attentive review of studies on a -crystallin chaperone- like activity, particularly the studies that have direct or indirect implications to hydrophobicity and chaperone-like activity, we found several instances wherein the correlation between hydrophobicity and its chaperone-like activity is paradoxical. We thus attempted to provide an overview on the role of hydrophobicity in chaperone-like activity of a -crystallin, the kind of evaluation done for the first time. IUBMB Life, 58: 632–641, 2006 Keywords sHsp; a-crystallin; crystallin domain; chaperone-like activity; hydrophobicity; lens; temperature. Abbreviations DTT, Dithiothreitol; sHsp, small heat shock proteins; ANS, 1-Anilino-8-naphthalene sulfonic acid; Bis-ANS, 1,1 0 -bi(4-anilino) naphthalene-5, 5-disulfonic acid; ITC, Isothermal titration calorimetry; G6PD, Glucose-6-phosphate dehydro- genase; GdmCl, Guanidinium hydrochloride. INTRODUCTION To become functionally active, nascent protein chains must fold into unique three-dimensional structures. Although, the native fold of a protein is encoded in its amino-acid sequence, protein folding inside cells is not generally a spontaneous process. Many newly synthesized proteins require complex cellular machinery comprising of molecular chaperones and an input of metabolic energy to attain native and thermodynami- cally stable states. Molecular chaperones are proteins that assist nascent protein folding and allow the functional state of proteins to be maintained by preventing irreversible protein unfolding and aggregation. Numerous studies, over the last decade have investigated the structural and functional charac- teristics of molecular chaperones, classifying them into several families based on size, structure and activity. The small heat shock proteins (sHsp 1 ) constitute a diverse family of ubiquitous intracellular proteins. sHsp have subunit masses in the range of 12 – 43 kDa and exist as large heterogeneous aggregates of 300 – 1000 kDa that contain 12 – 40 subunits (1). Unlike the classical chaperones, sHsp prevent the aggregation and precipitation of a variety of unrelated proteins under stress conditions such as elevated temperature, reduced and oxidized conditions at a stoichiometry as low as one subunit of sHsp to one target protein (1). a-Crystallin, a major structural protein of the vertebrate eye lens, is the most intensively studied representative member of sHsp family. a-Crystallin is known for over a century as one of the three major crystallins (the other two are b- and g-crystallins) of the vertebrate eye lens. However, it became a major focus of studies since 1982, when Drosophila sHsp were found to share sequence similarities with a-crystallin reflecting a common evolutionary ancestry (2). Soon after, it was shown that a-crystallin and other sHsp function like molecular chaperones in preventing the thermal aggregation of various proteins including the lens proteins (3, 4). It has been fairly acknowledged that with aging eye lens proteins undergo various posttranslational modifications, most of which lead to aggregation and this process is further accelerated due to various physiological, environmental and genetic factors that Received 24 July 2006; accepted 5 September 2006 Address correspondence to: G. Bhanuprakash Reddy, National Institute of Nutrition, Hyderabad – 500 007, India. Fax: þ91 40 27019074. E-mail: [email protected] *Present address: Laboratory of Allergic Diseases, National Institute of Allergy and Infectious Diseases, National Institutes of Health, USA. IUBMB Life, 58(11): 632 – 641, November 2006 ISSN 1521-6543 print/ISSN 1521-6551 online Ó 2006 IUBMB DOI: 10.1080/15216540601010096
Transcript of Chaperone-like activity and hydrophobicity of α-crystallin

Critical Review
Chaperone-like Activity and Hydrophobicity of a-Crystallin
G. Bhanuprakash Reddy, P. Anil Kumar and M. Satish Kumar*Biochemistry Division, National Institute of Nutrition, Hyderabad, India
Summary
a-Crystallin, a prominent member of small heat shock protein
(sHsp) family and a major structural protein of the eye lens is a large
polydisperse oligomer of two isoforms, aA- and aB-crystallins.
Numerous studies have demonstrated that a-crystallin functions like
a molecular chaperone in preventing the aggregation of various
proteins under a wide range of stress conditions. The molecular
chaperone function of a-crystallin is thus considered to be vital in
the maintenance of lens transparency and in cataract prevention.
a-Crystallin selectively interacts with non-native proteins thereby
preventing them from aggregation and helps maintain them in a
folding competent state. It has been proposed and generally accepted
that a-crystallin suppresses the aggregation of other proteins
through the interaction between hydrophobic patches on its surface
and exposed hydrophobic sites of partially unfolded substrate
protein. However, a quantifiable relationship between hydrophobi-
city and chaperone-like activity remains a matter to be concerned
about. On an attentive review of studies on a-crystallin chaperone-
like activity, particularly the studies that have direct or indirect
implications to hydrophobicity and chaperone-like activity, we found
several instances wherein the correlation between hydrophobicity and
its chaperone-like activity is paradoxical. We thus attempted to
provide an overview on the role of hydrophobicity in chaperone-like
activity of a-crystallin, the kind of evaluation done for the first time.
IUBMB Life, 58: 632–641, 2006
Keywords sHsp; a-crystallin; crystallin domain; chaperone-likeactivity; hydrophobicity; lens; temperature.
Abbreviations DTT, Dithiothreitol; sHsp, small heat shockproteins; ANS, 1-Anilino-8-naphthalene sulfonicacid; Bis-ANS, 1,10-bi(4-anilino) naphthalene-5,5-disulfonic acid; ITC, Isothermal titrationcalorimetry; G6PD, Glucose-6-phosphate dehydro-genase; GdmCl, Guanidinium hydrochloride.
INTRODUCTION
To become functionally active, nascent protein chains must
fold into unique three-dimensional structures. Although, the
native fold of a protein is encoded in its amino-acid sequence,
protein folding inside cells is not generally a spontaneous
process. Many newly synthesized proteins require complex
cellular machinery comprising of molecular chaperones and an
input of metabolic energy to attain native and thermodynami-
cally stable states. Molecular chaperones are proteins that
assist nascent protein folding and allow the functional state of
proteins to be maintained by preventing irreversible protein
unfolding and aggregation. Numerous studies, over the last
decade have investigated the structural and functional charac-
teristics of molecular chaperones, classifying them into several
families based on size, structure and activity. The small heat
shock proteins (sHsp1) constitute a diverse family of ubiquitous
intracellular proteins. sHsp have subunit masses in the range
of 12 – 43 kDa and exist as large heterogeneous aggregates
of 300 – 1000 kDa that contain 12 – 40 subunits (1). Unlike the
classical chaperones, sHsp prevent the aggregation and
precipitation of a variety of unrelated proteins under stress
conditions such as elevated temperature, reduced and oxidized
conditions at a stoichiometry as low as one subunit of sHsp to
one target protein (1).
a-Crystallin, a major structural protein of the vertebrate eye
lens, is the most intensively studied representative member of
sHsp family. a-Crystallin is known for over a century as one
of the three major crystallins (the other two are b- and
g-crystallins) of the vertebrate eye lens. However, it became a
major focus of studies since 1982, when Drosophila sHsp were
found to share sequence similarities with a-crystallin reflecting
a common evolutionary ancestry (2). Soon after, it was shown
that a-crystallin and other sHsp function like molecular
chaperones in preventing the thermal aggregation of various
proteins including the lens proteins (3, 4). It has been fairly
acknowledged that with aging eye lens proteins undergo
various posttranslational modifications, most of which lead
to aggregation and this process is further accelerated due to
various physiological, environmental and genetic factors that
Received 24 July 2006; accepted 5 September 2006Address correspondence to: G. Bhanuprakash Reddy, National
Institute of Nutrition, Hyderabad – 500 007, India.Fax: þ91 40 27019074. E-mail: [email protected]*Present address: Laboratory of Allergic Diseases, National Instituteof Allergy and Infectious Diseases, National Institutes of Health,USA.
IUBMBLife, 58(11): 632 – 641, November 2006
ISSN 1521-6543 print/ISSN 1521-6551 online � 2006 IUBMB
DOI: 10.1080/15216540601010096

predispose lens to cataract (5). Thus the chaperone-like
function of a-crystallin to suppress the aggregation of proteins
is considered to be critical for the maintenance of lens
transparency. However, the mechanism of chaperone-like
function is not fully understood. It is believed that the first
step in the protective action of a-crystallin is mediated
through the interaction with aggregation-prone or denatured
(non-native) unfolded protein to maintain it in a condition
that favors refolding or prevents from aggregation. One of
the major forces implicated in the mechanistic action of
a-crystallin is entropically driven hydrophobic contacts
between its accessible hydrophobic surfaces and newly exposed
hydrophobic sites of unfolding substrates (6 – 10). Hence, it
is conventionally accepted that hydrophobicity is a major
determinant of chaperone-like activity of a-crystallin.However, upon a careful assessment of the studies on
a-crystallin chaperone-like activity, including studies from
our laboratory, we found numerous instances in which
hydrophobicity alone does not account for the chaperone-like
activity of a-crystallin. In this review we focus on the relation
between hydrophobicity and chaperone-like activity of
a-crystallin.
a-CRYSTALLIN STRUCTURE
In the lens, a-crystallin exists as a heteropolymer with
the molecular size of approximately 800 kDa, having up to
40 subunits from two gene products aA and aB. aA is 173,
while aB is 175 amino acids long and both share 57% sequence
similarity. In most vertebrate lenses, aA and aB subunits exist
in the ratio of 3:1. However the actual ratio varies with species
and also with ageing. Reconstituted as well as recombinant
homopolymers of aA and aB-crystallin are somewhat smaller
(600 – 650 kDa) than the native protein (11, 12), but otherwise
appear to attain similar structural and functional integrity as
that of native a-crystallin. sHsp, including a-crystallin, are
predominantly b-sheet in secondary structure and their
primary sequence can be organized into three distinct struc-
tural regions: an a-crystallin domain of *90 amino acids in
length which is conserved among all sHsps and flanked by
N- and C-terminal domains of variable length and sequence
(Fig. 1). Based on far-UV CD and FT-IR data, secondary
structure of a-crystallin subunits consists of approximately
15% a-helix, 40% b-sheet and the remainder, random coil and
turns (9, 13). The conserved a-crystallin domain spans residues
63 – 144 in aA-crystallin and residues 68 – 148 in aB-crystallin.The amino acid sequence at the a-crystallin domain was shown
to have seven b-strands arranged in two b-sheets; one sheet
consists of three strands and the other of four strands, packed
face to face, forming a sandwich (Fig. 1, 14). Even though,
monomers of many sHsp have the conserved a-crystallindomain, the oligomeric structures display a large variety
between the individual members of the super family. The
poorly conserved N-terminal extension that is highly variable
Figure 1. Alignment of human aA and aB-crystallin amino acid sequences along with human (Hsp27), wheat (Hsp16.9) and
methanococcus jannaschii (Hsp16.5) heat shock proteins. Black bars below the sequences indicate b1 to b10 strands. Consensus
residues are highlighted.
PROPERTIES OF a-CRYSTALLIN 633

in length and hydrophobic in nature influences subunit oligo-
merization and chaperone-like activity whereas the flexible
C-terminal extension stabilizes the global structure and is
thought to enhance protein/substrate complex solubility.
Though, polydispersity of a-crystallin is known to hamper
crystal structure determination, cryo-electron microscopy data
for aB-crystallin indicate it is an irregular spherical structure
with central cavity (15), somewhat similar to that of available
crystal structures of hsp 16.5 and 16.9 (16, 17). Comprehensive
review of structure of a-crystallin and other sHsp has been
covered in (16 – 18).
CHAPERONE-LIKE ACTIVITY OF a-CRYSTALLIN
The chaperone-like activity of a-crystallin and related sHsp
refers to their ability to bind and stabilize non-native protein
substrates and prevent them from subsequent aggregation and
precipitation. Bovine a-crystallin and murine Hsp25 were the
first sHsp reported to have chaperone-like activity (3, 4, 19).
Since then, sHsp from other species have been shown to bind a
variety of non-native proteins in vitro during their chaperon-
ing-like function. The chaperone-like activity of sHsp are
usually assayed by simple in vitro aggregation experiments in
which substrate proteins are subjected to aggregation by heat
or chemical treatment in the presence of the chaperone
(19 – 24). The extent of aggregation of substrate protein is
usually measured in terms of optical density. DTT-induced
aggregation of insulin (due to precipitation of insulin B-chain
upon reduction) and heat-induced aggregation of citrate
synthase (42 – 508C) or b-crystallin (*608C) are widely
employed assays, though numerous other assays have been
used, for assessing the chaperone like function of sHSP.
However, it should be noted that the conclusions from
these experiments might not always be applicable to in vivo
situations (21). Typically in these studies it is assumed that the
chaperone itself is unaffected by the treatment used for
substrate aggregation. In contrast to other classes of molecular
chaperones, each oligomeric sHsp complex binds several non-
native polypeptide chains. Though ratios of up to one non-
native substrate protein per subunit have been reported, this
may vary. The substrate specificity of a-crystallin is rather
broad; chaperone-like activity has been demonstrated in vitro
with various lenticular and non-lenticular proteins against
physical and physiological stress such as heat, UV irradiation,
chemical stress, oxidative stress and glycation (3, 4, 6, 9,
20 – 25). Although, a-crystallin has been shown to facilitate
refolding of several proteins and enzymes, for example citrate
synthase, quinone oxidoreductase, lysozyme, ribonuclease,
xylose reductase and glucose-6-phosphate dehydrogenase
(G6PD) (26 – 28), the degree to which enzyme activity is
regained upon refolding remains inconclusive.
Chaperone-like activity of endogenous a-crystallin in main-
taining lens transparency was clearly evident in experiments
where the total soluble lens homogenate subjected to heat
showed little aggregation (29 – 31). On the other hand, lens
homogenate that is devoid of a-crystallin and subjected to heat
showed increased aggregation (31). The physiologic signifi-
cance of the chaperone function of a-crystallin is further
substantiated by the findings that mutations in a-crystallinthat are known to cause cataract have a dramatic effect on its
chaperone-like activity as well (reviewed in 1) (Table 1).
A missense mutation (R120G) in aB-crystallin is shown to
cause a familial form of desmin-related myopathy (DRM)
characterized by intracellular desmin aggregation, cardiomyo-
pathy and cataract (32). A similar mutation in aA-crystallin
(R116C) is shown to cause cataract (33). Recombinant
proteins with these mutations displayed altered physicochem-
ical properties with a considerably diminished ability to
suppress protein aggregation (discussed later). Animal knock-
outs, either aA or aB or both, provided further insights into
the physiologic relevance of a-crystallin chaperone-like func-
tion (reviewed in 20). Interestingly, while aA-crystallin knock-
out mice developed cataract, aB knockout mice showed no
signs of cataract but degeneration of skeletal muscles. Though,
a-crystallin is a heteropolymer of aA and aB, it is interesting tonote that phenotypes are different with aA and aB knockouts.
Furthermore, primary cultures of lens epithelial cells derived
from aB knockouts showed genome instability and hyper
proliferation suggesting that these proteins may also have an
important role to play in the maintenance and regulation of
genomic stability and cell division (34). Thus, these studies
implicate a role for aA- and aB-crystallins not only in the
maintenance of lens transparency but also in various other
cellular functions (35).
HYDROPHOBICITY AND CHAPERONE-LIKE ACTIVITY
While the chaperone-like activity of a-crystallin has been
proven to be critical under stress conditions, the molecular
mechanism of this activity remains largely unknown.
Understanding the molecular forces involved in the interaction
of a-crystallin with the substrate proteins is very important to
get insights into the mechanism by which a-crystallin protects
proteins against various physical and biochemical assaults.
Several sites on a-crystallin have been postulated to be
involved in the chaperone-like function and complex forma-
tion, including the N-terminal and C-terminal regions. The
C-terminal region of aA-crystallin (36) and the mostly
hydrophobic N-terminal phenylalanine-rich region in
aB-crystallin have been suggested to be necessary for
chaperone-like activity (37). Sharma et al. reported that
residues 57 – 69 and 93 – 107 in aB-crystallin are involved in
chaperoning alcohol dehydrogenase (38). Later they have also
reported that residues 50 – 54 and 79 – 99 in aA-crystallin and
residues 75 – 103 in aB-crystallin are bis-ANS-binding and
concluded that the hydrophobic sites participate in chaperone-
like activity of the protein (38 – 40). Based on protein pin
arrays, Ghosh et al. identified seven putative substrate
634 REDDY ET AL.
interactive sites in aB-crystallin that include two sequences
from the N-terminal domain, four sequences from the
a-crystallin domain and one from the C-terminal domain
(41). Five of these interactive sequences are identified as
sequences associated with subunit-subunit interactions in
human aB-crystallin. Based on these studies it appears that
virtually every region of a-crystallin is vital for its chaperone-
like function. Further, involvement of variable sequence
outside the a-crystallin domain in substrate binding suggests
that substrate-binding profiles may vary among different
sHsps and it may also depend on the arrangement of subunits
in three-dimensional conformation.
Irrespective of the involvement of specific sequence elements
that mediate either substrate binding or oligomerization,
several studies on a-crystallin have attributed chaperone-like
function to its hydrophobic surfaces interacting with the
exposed hydrophobic sites of denaturing substrate proteins.
Denaturing substrates expose buried hydrophobic surfaces
as a result of their unfolding and this was expected to
facilitate either self-aggregation or interaction with sHsp.
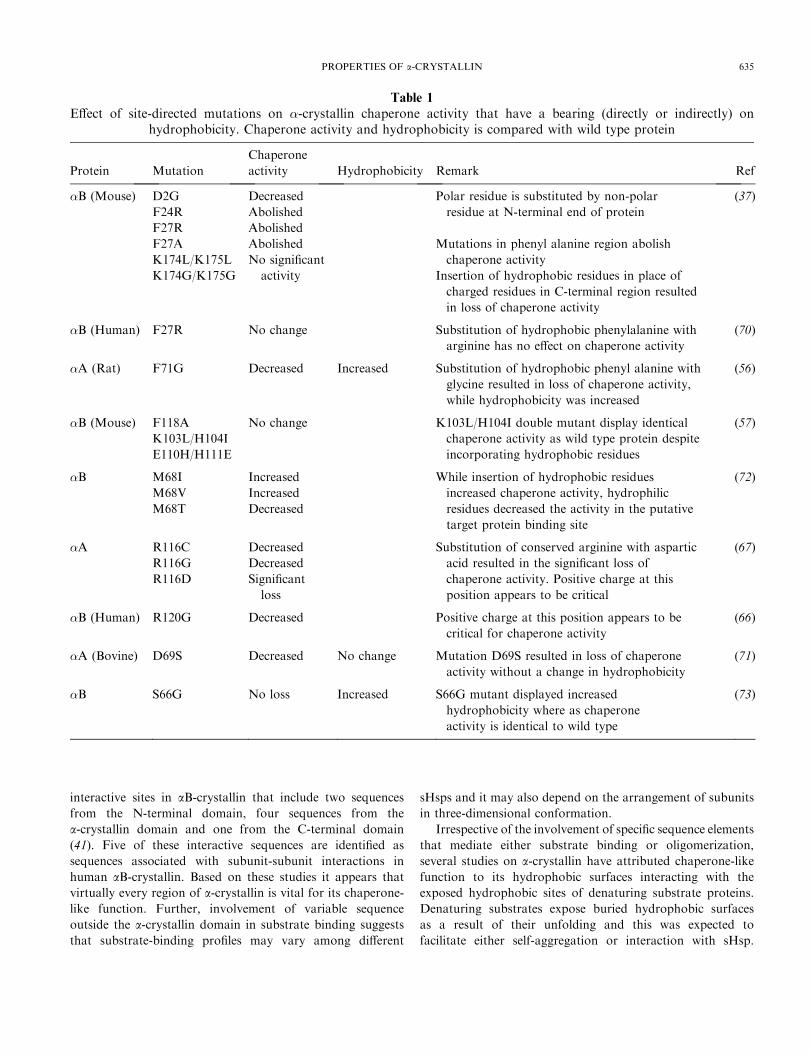
Table 1
Effect of site-directed mutations on �-crystallin chaperone activity that have a bearing (directly or indirectly) onhydrophobicity. Chaperone activity and hydrophobicity is compared with wild type protein
Protein Mutation
Chaperone
activity Hydrophobicity Remark Ref
�B (Mouse) D2G
F24R
Decreased
Abolished
Polar residue is substituted by non-polar
residue at N-terminal end of protein
(37)
F27R Abolished
F27A
K174L/K175L
Abolished
No significant
Mutations in phenyl alanine region abolish
chaperone activity
K174G/K175G activity Insertion of hydrophobic residues in place of
charged residues in C-terminal region resulted
in loss of chaperone activity
�B (Human) F27R No change Substitution of hydrophobic phenylalanine with
arginine has no effect on chaperone activity
(70)
�A (Rat) F71G Decreased Increased Substitution of hydrophobic phenyl alanine with
glycine resulted in loss of chaperone activity,
while hydrophobicity was increased
(56)
�B (Mouse) F118A
K103L/H104I
E110H/H111E
No change K103L/H104I double mutant display identical
chaperone activity as wild type protein despite
incorporating hydrophobic residues
(57)
�B M68I
M68V
M68T
Increased
Increased
Decreased
While insertion of hydrophobic residues
increased chaperone activity, hydrophilic
residues decreased the activity in the putative
target protein binding site
(72)
�A R116C
R116G
R116D
Decreased
Decreased
Significant
loss
Substitution of conserved arginine with aspartic
acid resulted in the significant loss of
chaperone activity. Positive charge at this
position appears to be critical
(67)
�B (Human) R120G Decreased Positive charge at this position appears to be
critical for chaperone activity
(66)
�A (Bovine) D69S Decreased No change Mutation D69S resulted in loss of chaperone
activity without a change in hydrophobicity
(71)
�B S66G No loss Increased S66G mutant displayed increased
hydrophobicity where as chaperone
activity is identical to wild type
(73)
PROPERTIES OF a-CRYSTALLIN 635

These sHsp-bound substrates are further refolded to their
native state by ATP dependent chaperones, such as Hsp70/
Dna K. Thus, sHsp appear to function like a reservoir of
partially unfolded proteins in refolding-competent state
(Fig. 2). In the case of classical chaperones, such as E. coli
GroEL, the chaperone activity has been attributed to surface
hydrophobicity of both the chaperone and its substrates (42).
Similarly, chaperone-like activity of 70S ribosome was
attributed to its surface hydrophobicity (43). Atp11p, a soluble
mitochondrial chaperone, is also known to prevent F1-ATPase
aggregation in matrix through hydrophobicity mediated
chaperone-substrate interactions (44). Thus it is conceivable
that hydrophobicity is a major factor in the chaperoning
function of many chaperones including a-crystallin. This is
substantiated by the experimental data that indicate that
a-crystallin preferentially recognizes non-native structures,
which are likely to have increased surface hydrophobicity on
the denaturation pathway (45). However, the so-called hydro-
phobic ‘non-native structures’ also seem to be structurally
diverse entities (46). The studies with three non-identical non-
native states of lactalbumin suggest that a-crystallin interacts
only with the aggregation prone molten globule state of
reduced apo-a-lactalbumin but not with the other non-
aggregating molten globule states of the protein (46). It has
been shown that a-crystallin specifically recognizes very early
intermediates on the denaturation pathway of proteins, which
are characterized by native-like secondary structure but
compromised tertiary interactions (47, 48). The structural state
of zeta-crystallin that binds to a-crystallin was shown to be a
partially unfolded inactive monomer that exhibits highly
exposed hydrophobic surfaces and has significant secondary
structural elements with little or no tertiary structure (49). This
intermediate does not refold into the active state without
assistance and a-crystallin provides the required assistance and
improves the reactivation yield several-fold. The strongest
supporting evidence for the involvement of hydrophobicity in
chaperone-like function came from temperature-dependent
experiments. Upon heating, a-crystallin undergoes structural
changes resulting in increased exposure of additional hydro-
phobic sites associated with increased chaperone-like activity
(6, 7, 9, 10, 47, 50). The chaperone-like activity of a-crystallin is
less pronounced at 308C as compared to its activity at 558C(51). While there is a minor alteration in a-crystallin tertiary
structure at 308C, above 508C, it undergoes a structural transi-tion to a molten globule-like state that has increased hydro-
phobic surfaces. More importantly when a-crystallin is heated
to high temperatures and then cooled (preheated), not only
there is a dramatic increase in surface hydrophobicity, the
a-crystallin exhibited a significant increase in chaperone-like
activity as compared to unheated a-crystallin (7, 9, 10). Once
exposed to high temperatures, the protein upon cooling does
not return to its original conformational state but adopts a
conformation characterized by significantly increased surface
hydrophobicity (7). Structural perturbations due to factors
Figure 2. A schematic mechanism of chaperone-like function of a-crystallin. a-Crystallin selectively interacts with non-native
intermediates on their unfolding pathway and stabilizes them in a refolding competent state. Hydrophobic forces may mediate
the interaction between the chaperone and partially unfolded substrate.
636 REDDY ET AL.

other than heat also enhanced the chaperone-like activity,
which were associated with increased hydrophobicity. Arginine
hydrochloride brings about subtle changes in the tertiary
structure and significant changes in the quaternary structure of
a-crystallin that resulted in enhanced chaperone-like activity,
which is mediated through increased exposure of hydrophobic
surfaces (52). In the guanidine hydrochloride (GdnHCl)
induced unfolding pathway; a molten globule-like intermediate
of a-crystallin was shown to have maximum chaperone-like
activity concurrent with maximum hydrophobicity (53). These
studies are considered as ‘‘gold standards of evidence’’ of the
intimate relationship between hydrophobicity and chaperone-
like activity of a-crystallin.However, despite a wealth of evidence for dependence of
the chaperone function of a-crystallin on its surface hydro-
phobicity, a deterministic role of hydrophobicity in quantita-
tive terms remains unresolved. There are numerous instances
where hydrophobicity has not been directly correlated to the
chaperone-like activity of either a-crystallin heteropolymer or
aA or aB-homopolymers. Though it is strongly believed that
the a-crystallin domain contributes to hydrophobicity and is
associated with chaperone-like activity, sequence alignment
does not indicate the presence of either conserved hydro-
phobic sequences or conserved hydrophobic residues (Fig. 1).
Perhaps the ‘so called’ hydrophobic surfaces might appear
after attaining the global conformation. This may be the
reason for exhibiting increased hydrophobicity upon reorga-
nization after structural perturbations. Further, it was shown
that contacts between a conserved motif (with the sequence I-
X-I/V) in the C-terminal region and a hydrophobic patch in
the a-crystallin domain of neighboring subunits are critical for
oligomer formation (54). This raises the question as to how the
hydrophobic domain of a-crystallin mediates two different
events, oligomerization and substrate binding. It is possible
that subunits may come apart to bind the substrate but it
is not straightforward to determine the hydrophobicity of
a-crystallin under those circumstances. Although, the heat-
shock domain of a-crystallin is implicated in chaperone-
like function, it has been reported that this domain has no
chaperone-like activity independently (55). Further, the
excised heat-shock domain is found to be predominantly
trimeric and displays significant surface hydrophobicity and a
tendency to undergo self-aggregation. It suggests that surface
hydrophobicity may alone be insufficient for this domain to
function as a chaperone (55).
Replacement of Phe71 with Gly in aA-crystallin results
in the loss of chaperone-like activity despite an increase in
surface hydrophobicity with no significant alteration in
structure (56). This is an interesting example where hydro-
phobicity does not correspond with chaperone-like activity.
Residues H101 to R120 are highly conserved between aA- and
aB-crystallin. However, single mutation (F118A) and double
mutations (K103L/H104I) did not affect the chaperone-like
activity of aB-crystallin, in spite of the replacement of
hydrophobic residues. (57). Using insulin aggregation assay,
all three mutants had identical chaperone-like activity to the
wild-type recombinant aB-crystallin. Likewise, insertion of
hydrophobic residues into the C-terminal extension of aA-
crystallin results in a partial loss of the flexibility in this region
and a concomitant decrease in chaperone-like activity and
stability of the protein (58). Studies showed that high mole-
cular weight (HMW) complexes of a-crystallin with its
substrate formed during heat induced aggregation exhibited
a greater hydrophobicity than complexes formed during DTT-
induced aggregation (59), regardless of its similar ability to
suppress the aggregation under both the conditions. Based on
these observations, in contrast to the existing impression, it
appears that hydrophobicity may not play a central role in
chaperone-like activity of a-crystallin. Some studies have
attempted to highlight the importance of hydrophobicity by
incorporation of ANS or bis-ANS, which is expected to block
the hydrophobic sites. But these studies indicate only a partial
loss of chaperone-like activity upon blocking the surface
hydrophobicity (39, 60). This is surprising, because one would
expect a loss in chaperone-like activity with prior ANS
binding. Further, if chaperone-like activity is predominantly
due to hydrophobicity, the loss in chaperone-like activity with
prior ANS binding could be much greater under conditions
that enhance the chaperone activity. For instance, the increase
in chaperone-like activity was much greater with preheated aAas compared to aB, which is perceived to be due to higher
hydrophobicity, but the percentage loss of chaperone-like
activity due to ANS binding was similar, with these preheated
aA- and aB-crystallins (10).a-Crystallin modified with methlyglyoxal (MGO), a highly
reactive dicarbonyl cross linking agent, exhibited increased
chaperone-like activity in conventional aggregation assays
despite an apparent decrease in hydrophobicity due to MGO
modification (21). This is another example, where chaperone-
like activity and hydrophobicity run exactly in opposite
directions. Yet, MGO-modified a-crystallin’s ability to prevent
inactivation of enzymes was compromised as compared to
native a-crystallin (21). Clearly further investigation is needed
to determine whether the ability of a-crystallin in protecting
the enzymes is independent of hydrophobicity. Interestingly
aB-crystallin specifically assisted the refolding and reactivation
of completely unfolded G6PD by GdmCl where as it is unable
to assist the reactivation of molten globule-like state that is
highly hydrophobic intermediate in GdmCl-induced unfolding
of G6PD (27). In the case of xylose reductase, a-crystallin was
shown to interact specifically with its molten globule state in
reconstituting the enzyme activity (28). While the molten
globule state of a protein is richly hydrophobic, the interaction
of a-crystallin with molten globule species of different sub-
strates appears to be different (27, 28).
Although, temperature dependent experiments suggest the
involvement of hydrophobicity in the chaperone-like activity
of a-crystallin, the relationship between hydrophobicity and
PROPERTIES OF a-CRYSTALLIN 637
increased chaperone-like activity at elevated temperatures
differs in the case of different species of a-crystallins (9 – 12, 47,50). At physiological temperatures (25 – 378C) recombinant aBdisplayed higher chaperone-like activity than recombinant aA-
crystallin and aB appears to be more hydrophobic than aA at
these temperatures (9). At high temperatures where the hydro-
phobicity of recombinant aA increases and aB decreases (9),
recombinant aA-crystallin shows higher chaperone-like activ-
ity than recombinant aB. In contrast, while reconstituted
aA-crystallin (from calf a-crystallin) exhibits slightly increased
hydrophobic profile than the reconstituted aB at room tem-
perature, the chaperone-like activity of aA-crystallin is lower
than that of aB-crystallin (12). Bis-ANS binding to both
reconstituted aA- and aB-crystallin decrease with increase in
temperature despite enhanced chaperone potential (12). When
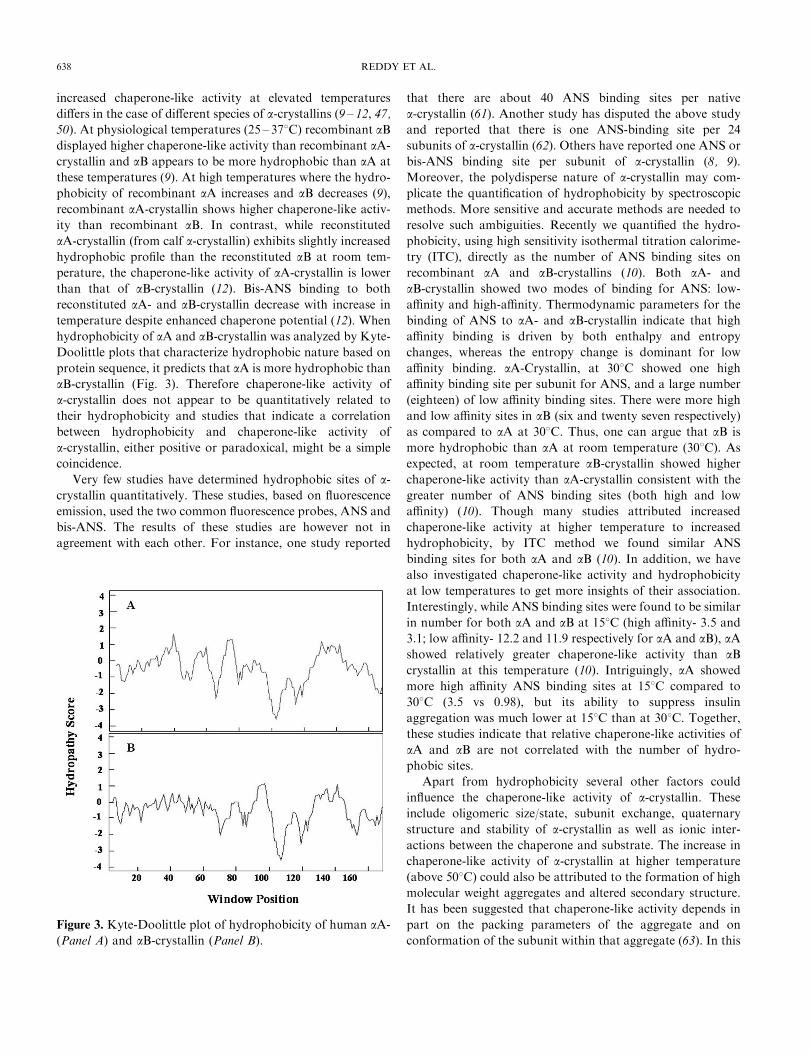
hydrophobicity of aA and aB-crystallin was analyzed by Kyte-
Doolittle plots that characterize hydrophobic nature based on
protein sequence, it predicts that aA is more hydrophobic than
aB-crystallin (Fig. 3). Therefore chaperone-like activity of
a-crystallin does not appear to be quantitatively related to
their hydrophobicity and studies that indicate a correlation
between hydrophobicity and chaperone-like activity of
a-crystallin, either positive or paradoxical, might be a simple
coincidence.
Very few studies have determined hydrophobic sites of a-crystallin quantitatively. These studies, based on fluorescence
emission, used the two common fluorescence probes, ANS and
bis-ANS. The results of these studies are however not in
agreement with each other. For instance, one study reported
that there are about 40 ANS binding sites per native
a-crystallin (61). Another study has disputed the above study
and reported that there is one ANS-binding site per 24
subunits of a-crystallin (62). Others have reported one ANS or
bis-ANS binding site per subunit of a-crystallin (8, 9).
Moreover, the polydisperse nature of a-crystallin may com-
plicate the quantification of hydrophobicity by spectroscopic
methods. More sensitive and accurate methods are needed to
resolve such ambiguities. Recently we quantified the hydro-
phobicity, using high sensitivity isothermal titration calorime-
try (ITC), directly as the number of ANS binding sites on
recombinant aA and aB-crystallins (10). Both aA- and
aB-crystallin showed two modes of binding for ANS: low-
affinity and high-affinity. Thermodynamic parameters for the
binding of ANS to aA- and aB-crystallin indicate that high
affinity binding is driven by both enthalpy and entropy
changes, whereas the entropy change is dominant for low
affinity binding. aA-Crystallin, at 308C showed one high
affinity binding site per subunit for ANS, and a large number
(eighteen) of low affinity binding sites. There were more high
and low affinity sites in aB (six and twenty seven respectively)
as compared to aA at 308C. Thus, one can argue that aB is
more hydrophobic than aA at room temperature (308C). As
expected, at room temperature aB-crystallin showed higher
chaperone-like activity than aA-crystallin consistent with the
greater number of ANS binding sites (both high and low
affinity) (10). Though many studies attributed increased
chaperone-like activity at higher temperature to increased
hydrophobicity, by ITC method we found similar ANS
binding sites for both aA and aB (10). In addition, we have
also investigated chaperone-like activity and hydrophobicity
at low temperatures to get more insights of their association.
Interestingly, while ANS binding sites were found to be similar
in number for both aA and aB at 158C (high affinity- 3.5 and
3.1; low affinity- 12.2 and 11.9 respectively for aA and aB), aAshowed relatively greater chaperone-like activity than aBcrystallin at this temperature (10). Intriguingly, aA showed
more high affinity ANS binding sites at 158C compared to
308C (3.5 vs 0.98), but its ability to suppress insulin
aggregation was much lower at 158C than at 308C. Together,these studies indicate that relative chaperone-like activities of
aA and aB are not correlated with the number of hydro-
phobic sites.
Apart from hydrophobicity several other factors could
influence the chaperone-like activity of a-crystallin. These
include oligomeric size/state, subunit exchange, quaternary
structure and stability of a-crystallin as well as ionic inter-
actions between the chaperone and substrate. The increase in
chaperone-like activity of a-crystallin at higher temperature
(above 508C) could also be attributed to the formation of high
molecular weight aggregates and altered secondary structure.
It has been suggested that chaperone-like activity depends in
part on the packing parameters of the aggregate and on
conformation of the subunit within that aggregate (63). In this
Figure 3. Kyte-Doolittle plot of hydrophobicity of human aA-
(Panel A) and aB-crystallin (Panel B).
638 REDDY ET AL.

regard, methylglyoxal modified a-crystallin that has larger
aggregate size showed increased chaperone-like activity
despite a decrease in surface hydrophobicity (21). Remark-
ably, however, sHsps were also found to exhibit increased
chaperone-like activities while maintaining their oligomeric
size unchanged. sHsp complexes are dynamic in structure and
exchange subunits constantly and this appears to be one of the
properties of sHsps that is important for their activity (64).
The dynamic behavior of sHsp could allow the substrate-
binding sites, which are normally buried in the oligomeric
complex, to become exposed on dissociation. This led to the
hypothesis that dissociation of the oligomer is required for
recognition of the substrate. It has been demonstrated that
subunit exchange in a-crystallin is a key factor in chaperone-
like function (65). It should be noted that both oligomeriza-
tion and subunit exchange are interconnected and mediated by
hydrophobic interactions. The conserved residues such as
R112 and R116 in the buried region of the protein form salt
bridges between the subunits (66). Studies with various
recombinant constructs of R116 of aA-crystallin (R116K,
R116G, R116C and R116D) and R120 of aB-crystallin(R120G) have shown that a positive charge must be preserved
at this position for structural and functional integrity of
a-crystallin (66, 67). In addition, presence of charged residues
at the C-terminal end of a-crystallin has been shown to be
critical for solvent interaction. Hence, disruption of net charge
of the protein may affect structural stability and chaperone-
like activity.
In summary, there are several factors that seem to influence
chaperone-like activity, hydrophobicity may be one of the
factors but may not be the predominant one. Further, chaper-
oning function and mechanism may vary depending on the
substrate and other prevailing conditions. Thus, proposing a
universal mechanism for the chaperone-like activity of
a-crystallin or sHsp may not be feasible.
FUTURE DIRECTIONS
Apart from its presence in the lens, a-crystallin is also
found in many non-lenticular tissues, albeit at very low levels
(as reviewed in ref 68). Furthermore, elevated expression of
a-crystallin, particularly aB-crystallin, has been observed in
many pathological conditions (1, 68, 69). Although, the
importance of a-crystallin in non-lenticular tissues is acknowl-
edged merely as stress protein, its function in non-lenticular
tissues still remains unanswered. Investigating the mechanism
of a-crystallin mediated cellular functions, particularly in non-
lenticular tissues, such as cell differentiation and cytoskeletal
organization could throw a light on the significance of hydro-
phobicity regarding chaperone-like activity. It may be noted
that while aA-crystallin is present in a limited number of
tissues at very low levels, aB-crystallin is prominent in quite a
few non-lenticular tissues. However, a-crystallin has not been
purified from any tissues other than lens. Moreover, whether it
exists as an aggregate (oligomer) or some other quaternary
state in other tissues remains to be answered. Thus, studies
should focus on characterizing structural and functional
aspects with respect to hydrophobicty of a-crystallin purified
from non-lenticular tissues. At the same time, understanding
the significance of the existence of two subunits of a-crystallinas a heteropolymer with 3:1 ratio in the lens may also provide
further insights into hydrophobicity and chaperone-like
function. A serious deficiency in the studies that relate the
chaperone-like activity and hydrophobicity of a-crystallin is
that in these studies the researchers have used different pre-
parations of a-crystallin (e.g., native a-crystallin hetero-
polymer, recombinant or reconstituted homopolymers of aAor aB) and diverse chaperone assays with various substrates.
It is known that physicochemical properties of a-crystallin can
vary depending on the sources (e.g., young or old lens,
species), the purification methods employed, the presence of
posttranslational modifications and the chaperone assay
conditions. Hence, assessing source and purification of
a-crystallin and the probes used for measuring hydro-
phobicity is of great concern in relating the hydrophobicity
to chaperone-like function. Thus, studies are warranted to
resolve the hydrophobicity paradox in order to provide greater
insights into chaperone-like activity of a-crystallin.
ACKNOWLEDGEMENTS
Grants from Department of Science and Technology, Govern-
ment of India and Indian Council of Medical Research,
Government of India to GBR are acknowledged. The authors
thank Prof. A. Surolia, Indian Institute of Science, Bangalore
for the use of their biophysical facilities and critical comments
and Mr. P. Yadagiri Reddy for his help at various aspects of
the work reported in this review. PAK and MSK acknowledge
Council of Scientific and Industrial Research for providing
research fellowship.
REFERENCES1. Sun, Y., and MacRae, T. H. (2005) The small heat shock proteins and
their role in human disease. FEBS J. 272, 2613 – 2627.
2. Ingolia, T. D., and Craig, E. A. (1982) Four small Drosophila heat
shock proteins are related to each other and to mammalian
a-crystallin. Proc. Natl. Acad. Sci. USA 79, 2360 – 2364.
3. Horwitz, J. (1992) Alpha-crystallin can function as a molecular
chaperone. Proc. Natl. Acad. Sci. USA 89, 10449 – 10453.
4. Jakob, U., Gaestel, M., Engel, K., and Buchner, J. (1993) Small heat
shock proteins are molecular chaperones. J. Biol. Chem. 268,
1517 – 1520.
5. Harding, J. J. (1991) In Cataract; Biochemistry, Epidemiology and
Pharmacology, Chapman & Hall, London.
6. Raman, B., and Rao, C. M. (1994) Chaperone-like activity
and quaternary structure of alpha-crystallin. J. Biol. Chem. 269,
27264 – 27268.
7. Das, K. P., and Surewicz, W. K. (1995) Temperature-induced
exposure of hydrophobic surfaces and its effect on the chaperone
activity of alpha-crystallin. FEBS Lett. 369, 321 – 325.
PROPERTIES OF a-CRYSTALLIN 639

8. Sharma, K. K., Kaur, H., Kumar, G. S., and Kester, K. (1998)
Interaction of 1,10-bi(4-anilino) naphthalene-5, 50-disulfonic acid with
alpha-crystallin. J. Biol. Chem. 273, 8965 – 8970.
9. Reddy, G. B., Das, K. P., Petrash, J. M., and Surewicz, W. K. (2000)
Temperature-dependent chaperone activity and structural properties
of human alphaA- and alphaB-crystallins. J. Biol. Chem. 275,
4565 – 4570.
10. Kumar, M. S., Kapoor, M., Sinha, S., and Reddy, G. B. (2005)
Insights into hydrophobicity and the chaperone-like function of
alphaA- and alphaB-crystallins: an isothermal titration calorimetric
study. J. Biol. Chem. 280, 21726 – 21730.
11. Sun, T. X., Das, B. K., and Liang, J. J. (1997) Conformational and
functional differences between recombinant human lens alphaA- and
alphaB-crystallin. J. Biol. Chem. 272, 6220 – 6225.
12. Bhattacharyya, J., Srinivas, V., and Sharma, K. K. (2002) Evaluation
of hydrophobicity versus chaperone like activity of bovine aA and
aB-crystallin. J. Protein Chem. 21, 65 – 71.
13. Farnsworth, P. N., Groth-Vasselli, B., Greenfield, N. J., and Singh, K.
(1997) Effects of temperature and concentration on bovine lens
alpha-crystallin secondary structure: a circular dichroism spec-
troscopic study. Int. J. Biol. Macromol. 20, 283 – 291.
14. Koteiche, H. A., and Mchaourab, H. S. (1999) Folding pattern of
the alpha-crystallin domain in alphaA-crystallin determined by site-
directed spin labeling. J. Mol. Biol. 294, 561 – 577.
15. Haley, D. A., Horwitz, J., and Stewart, P. L. (1998) The small heat-
shock protein, alphaB-crystallin, has a variable quaternary structure.
J. Mol. Biol. 277, 27 – 35.
16. Van Montfort, R., Slingsby, C., and Vierling, E. (2001) Structure and
function of the small heat shock protein/alpha-crystallin family of
molecular chaperones. Adv. Protein Chem. 59, 105 – 156.
17. Augusteyn, R. (2004) a-crystallin: a review of its structure and
function. Clin. Exp. Optom. 87, 356 – 366.
18. Carver, J. A., and Linder, R. A. (1998) NMR spectroscopy of alpha-
crystallin. Insights into the structure, interactions and chaperone
action of small heat shock proteins. Int. J. Biol. Macromol. 22,
197 – 209.
19. Merck, K. B., Groenen, P. J. T. A., Voorter, C. E., de Haard-
Hoekman, W. A., Horwitz, J., Bloemendal, H., and dejong, W. W.
(1993) Structural and functional similarities of bovine a-crystallin and
mouse small heat shock proteins. J. Biol. Chem. 268, 1046 – 1052.
20. Horwitz, J. (2003) Alpha-crystallin. Exp. Eye Res. 76, 145 – 153.
21. Kumar,M. S., Reddy, P.Y.,Kumar, P.A., Surolia. I., andReddy,G.B.
(2004) Effect of dicarbonyl-induced browning on alpha-crystallin
chaperone-like activity: physiological significance and caveats of
in vitro aggregation assays. Biochem. J. 379, 273 – 282.
22. Reddy, G. B., Narayanan, S., Reddy, P. Y., and Surolia, I. (2002)
Suppression of DTT-induced aggregation of abrin by alphaA- and
alphaB-crystallins: a model aggregation assay for alpha-crystallin
chaperone activity in vitro. FEBS Lett. 522, 59 – 64.
23. Hook, D. W., and Harding, J. J. (1998) Protection of enzymes by
alpha-crystallin acting as a molecular chaperone. Int. J. Biol.
Macromol. 22, 295 – 306.
24. Reddy, G. B., Reddy, P. Y., and Suryanarayana, P. (2001) aA- and
aB-crystallins protect glucose-6-phosphate dehydrogenase against
UVB irradiation-induced inactivation. Biochem. Biophys. Res.
Commun. 282, 712 – 716.
25. Hess, J. F., and FitzGerald, P. G. (1998) Protection of a restriction
enzyme from heat inactivation by [alpha]-crystallin. Mol. Vis. 4, 29.
26. Raman, B., Ramakrishna, T., and Rao, C. M. (1997) Effect of the
chaperone-like alpha-crystallin on the refolding of lysozyme and
ribonucleaseA. FEBS Lett. 416, 369 – 372.
27. Kumar, M. S., Reddy, P. Y., Sreedhar. B., and Reddy, G. B. (2005)
Alphab-crystallin-assisted reactivation of glucose-6-phosphate dehy-
drogenase upon refolding. Biochem. J. 391, 335 – 341.
28. Rawat, U., and Rao, M. (1998) Interactions of chaperone
alpha-crystallin with the molten globule state of xylose reductase.
Implications for reconstitution of the active enzyme. J. Biol. Chem.
273, 9415 – 9423.
29. Rao, P. V., Huang, Q. L., Horwitz, J., and Zigler, J. S. Jr (1995)
Evidence that alpha-crystallin prevents non-specific protein aggrega-
tion in the intact eye lens. Biochim. Biophys. Acta 1245, 439 – 447.
30. Reddy, G. B., Reddy, P. Y., Vijayalakshmi, A., Kumar, M. S.,
Suryanarayana, P., and Sesikeran, B. (2002) Effect of long-term
dietary manipulation on the aggregation of rat lens crystallins: role of
alpha-crystallin chaperone function. Mol. Vis. 8, 298 – 305.
31. Horwitz, J. (1993) Proctor Lecture. The function of alpha-crystallin.
Invest. Ophthalmol. Vis. Sci. 34, 10 – 22.
32. Vicart, P., Caron, A., Guicheney, P., Li, Z., Prevost, A. F.,
Chatean, D., Chapon, F., Tome, F., Dupret, J. M., and Saulin, D.
(1998) A missense mutation in the alphaB-crystallin gene causes a
desmin related myopathy. Nature Gen. 20, 92 – 95.
33. Litt,M.,Kramer, P., LaMorticella,D.M.,Murphy,W., Lovrien,E.M.,
and Weleber, R. G. (1998) Autosomal dominant congenital cataract
associated with a missense mutation in the human alpha gene CRYAA.
Hum. Mol. Genet. 7, 471 – 474.
34. Andley, U. P., Song, Z., Wawrousek, E. F., Brady, J. P, Bassnett, S.,
and Fleming, T. P. (2001) Lens epithelial cells derived from
alphaB-crystallin knockout mice demonstrate hyperproliferation and
genomic instability. FASEB J. 15, 221 – 229.
35. Boyle, D. L., Takemoto, L., Brady, J. P., and Wawrousek, E. F.
(2003) Morphological characterization of the Alpha A- and Alpha
B-crystallin double knockout mouse lens. BMC Ophthalmol. 3, 3.
36. Takemoto, L., Emmons, T., and Horwitz, J. (1993). The C-terminal
region of alpha-crystallin: involvement in protection against heat-
induced denaturation. Biochem. J. 294, 435 – 438.
37. Plater, M. L., Goode, D., and Crabbe. M. J. (1996) Effects of site-
directed mutations on the chaperone-like activity of alphaB-crystallin.
J. Biol. Chem. 271, 28558 – 28566.
38. Sharma, K. K., Kaur, H., and Kester, K. (1997) Functional elements
in molecular chaperone alpha-crystallin: identification of binding sites
in alpha B-crystallin. Biochem. Biophys. Res. Commun. 239, 217 – 222.
39. Sharma, K. K., Kumar, G. S., Murphy, A. S., and Kester, K. (1998)
Identification of 1,10-bi(4-anilino) naphthalene-5, 50-disulfonic acid
binding sequences in alpha-crystallin. J. Biol. Chem. 273,
15474 – 15478.
40. Sharma, K., Kumar, R. S., Kumar, G. S., and Quinn, P. T. (2000)
Synthesis and characterization of a peptide identified as a functional
element in alphaA-crystallin. J. Biol. Chem. 275, 3767 – 3771.
41. Ghosh, J. G., Estrada, M. R., and Clark, J. L. (2005) Interactive
domains for chaperone activity in the small heat shock protein, human
alphaB crystallin. Biochemistry 44, 14854 – 14869.
42. Melkani, G. C., Zardeneta, G., and Mendoza, J. A. (2004) Oxidized
GroEL can function as a chaperonin. Front Biosci. 9, 724 – 731.
43. Singh, R., and Rao, Ch. M. (2002) Chaperone-like activity and
surface hydrophobicity of 70S ribosome. FEBS Lett. 527, 234 – 238.
44. Sheluho, D., and Ackerman, S. H. (2001) An accessible hydrophobic
surface is a key element of the molecular chaperone action of Atp11p.
J. Biol. Chem. 276, 39945 – 39949.
45. Das, K. P., Petrash, J. M., and Surewicz, W. K. (1996) Conforma-
tional properties of substrate proteins bound to a molecular
chaperone a-crystallin. J. Biol. Chem. 271, 10449 – 10452.
46. Rajaraman, K., Raman, B., Ramakrishna, T., and Rao, C. M. (1998)
The chaperone-like alpha-crystallin forms a complex only with the
aggregation-prone molten globule state of alpha-lactalbumin.
Biochem. Biophys. Res. Commun. 249, 917 – 921.
47. Datta, S. A., and Rao, C. M. (1999) Differential temperature-
dependent chaperone-like activity of aA- and aB-crystallin homo-
aggregates. J. Biol. Chem. 274, 34773 – 34778.
640 REDDY ET AL.

48. Das, K. P., Choo-Smith, L. P., Petrash, J. M., and Surewicz, W. K.
(1999) Insight into the secondary structure of non-native proteins
bound to a molecular chaperone alpha-crystallin. An isotope-edited
infrared spectroscopic study. J. Biol. Chem. 274, 33209 – 33212.
49. Goenka, S., Raman, B., Ramakrishna, T., and Rao, C. M. (2001)
Unfolding and refolding of a quinone oxidoreductase: alpha-
crystallin, a molecular chaperone, assists its reactivation. Biochem.
J. 359, 547 – 556.
50. van Boekel, M. A., de Lange, F., de Grip, W. J., and de Jong, W. W.
(1999) Eye lens alphaA- and alphaB-crystallin: complex stability
versus chaperone-like activity. Biochim. Biophys. Acta 1434, 114 – 123.
51. Raman, B., and Rao, C. M. (1997) Chaperone-like activity and
temperature-induced structural changes of alpha-crystallin. J. Biol.
Chem. 272, 23559 – 23564.
52. Srinivas, V., Raman, B., Rao, K. S., Ramakrishna, T., and
Rao, Ch. M. (2003) Structural perturbation and enhancement of the
chaperone-like activity of alpha-crystallin by arginine hydrochloride.
Protein Sci. 12, 1262 – 1270.
53. Das, B. K., and Liang, J. J. (1997) Detection and characterization of
alpha-crystallin intermediate with maximal chaperone-like activity.
Biochem. Biophys. Res. Commun. 236, 370 – 374.
54. Pasta, Y., Raman, B., Ramakrishna, T., and Rao, C. M. (2004) The
IXI/V motif in the C-terminal extension of alpha-crystallins: alter-
native interactions and oligomeric assemblies. Mol. Vis. 10, 655 – 662.
55. Kundu, B., Shukla, A., Chaba, R., and Guptasarma, P. (2004) The
excised heat-shock domain of alphaB crystallin is a folded,
proteolytically susceptible trimer with significant surface hydropho-
bicity and a tendency to self-aggregate upon heating. Protein Expr.
Purif. 36, 263 – 271.
56. Kumar, P. S., and Sharma, K. K. (2001) Phe71 is essential for
chaperone-like function in alpha A-crystallin. J. Biol. Chem. 276,
47094 – 47099.
57. Hepburne-Scott, H. W., and Crabbe, M. J. C. (1999) Maintenance of
chaperone like activity despite mutations in a conserved region of
murine lens aB crystallin. Mol. Vis. 5, 15.
58. Smulders, R.H. P. H., Carver, J. A., Lindner, R. A., van Boekel,M. A.,
Bloemendal, H., and de Jong, W. W. (1996) Immobilization of the
C-terminal extension of bovine alphaA-crystallin reduces chaperone-
like activity. J. Biol. Chem. 271, 29060 – 29066.
59. Lindner,R.A.,Kapur,A.,Mariani,M., Titmuss, S. J., andCarver, J.A.
(1998) Structural alterations of alpha-crystallin during its chaperone
action. Eur. J. Biochem. 258, 170 – 183.
60. Smulders, R. H., and de Jong, W. W. (1997) The hydrophobic probe
4,40-bis(1-anilino-8-naphthalene sulfonic acid) is specifically photo-
incorporated into the N-terminal domain of alpha B-crystallin. FEBS
Lett. 409, 101 – 104.
61. Liang, J. N., and Li, X. Y. (1991) Interaction and aggregation of lens
crystallins. Exp. Eye Res. 53, 61 – 66.
62. Stevans, A., and Augusteyn, R. C. (1997) Binding of 1-anilino-
naphthalene-8-sulfonic acid to alpha-crystallin. Eur. J. Biochem. 243,
792 – 797.
63. Burgio, M. R., Kim, C. J., Dow, C. C., and Koretz, J. F. (2000)
Correlation between the chaperone-like activity and aggregate size of
alpha-crystallin with increase in temperature. Biochem. Biophys. Res.
Commun. 268, 426 – 432.
64. Bova, M. P., McHaourab, H. S., Han, Y., and Fung, B. K. (2000)
Subunit exchange of small heat shock proteins. Analysis of oligomer
formation of alphaA-crystallin and Hsp27 by fluorescence resonance
energy transfer and site-directed truncations. J. Biol. Chem. 275,
1035 – 1042.
65. Bova, M. P., Ding, L. L., Horwitz, J., and Fung, B. K. (1997)
Subunit exchange of alphaA-crystallin. J. Biol. Chem. 272,
29511 – 29517.
66. Treweek, T. M, Rekas, A., Lindner, R. A., Walker, M. J.,
Aquilina, J. A., Robinson, C. V., Horwitz, J., Perng, M. D.,
Quinlan, R. A., and Carver, J. A. (2005) R120G alphaB-crystallin
promotes the unfolding of reduced alpha-lactalbumin and is inherently
unstable. FEBS J. 272, 711 – 724.
67. Bera, S., Thampi, P., Cho-W. J., and Abraham, E. C. (2002)
A positive charge preservation at position 116 of alpha A-crystallin
is critical for its structural and functional integrity. Biochemistry 41,
12421 – 12426.
68. Bhat, S. P. (2003) Crystallins, genes and cataract. Prog. Drug Res. 60,
205 – 262.
69. Kumar, P. A., Haseeb, A., Suryanarayana, P., Ehtesham, N. Z., and
Reddy, G. B. (2005) Elevated expression of alphaA- and alphaB-
crystallins in streptozotocin-induced diabetic rat. Arch. Biochem.
Biophys. 444, 77 – 83.
70. Horwitz, J., Bova, M., Huang, Q. L., Yaron, O., and Lowman, S.
(1998). Mutation of alpha B-crystallin: effects on chaperone-like
activity. Int. J. Biol. Macromol. 22, 263 – 269.
71. Smulders, R. H., Merck, K. B., Aendekerk, J., Horwitz, J.,
Takemoto, L., Slingsby, C., Bloemendal, H., and De Jong, W. W.
(1995) The mutation Asp69 ––4Ser affects the chaperone-like activity
of alpha A-crystallin. Eur. J. Biochem. 232, 834 – 838.
72. Shroff, N. P., Bera, S., Cherian-Shaw, M., and Abraham, E. C. (2001)
Substituted hydrophobic and hydrophilic residues at methionine-68
influence the chaperone-like function of alphaB-crystallin. Mol. Cell
Biochem. 220, 127 – 133.
73. Sreelakshmi, Y., and Sharma K. K. (2005) Recognition sequence 2
(residues 60–71) plays a role in oligomerization and exchange
dynamics of alphaB-crystallin. Biochemistry 44, 12245 – 12252.
PROPERTIES OF a-CRYSTALLIN 641










![The Drowsy Chaperone Program [2012] - USM Digital ...](https://static.fdokumen.com/doc/165x107/6322912f887d24588e044a66/the-drowsy-chaperone-program-2012-usm-digital-.jpg)









