
Defining Responsiveness of Avian Cochlear Neurons to Brain-Derived Neurotrophic Factor and Nerve...
11
Journal of Neurochemistry Lippincott—Raven Publishers, Philadelphia © 1998 International Society for Neurochemistry Defining Responsiveness of Avian Cochlear Neurons to Brain-Derived Neurotrophic Factor and Nerve Growth Factor by HSV-1-Mediated Gene Transfer Juan José Garrido, Maria Teresa Alonso, Filip Lim, Estela Carnicero, Fernando Giraldez, and Thomas Schimmang Instituto de Biologla y Genética Molecular, Universidad de Valladolid y Consejo Superior de Investigaciones Cientificas, Departamento de Bioqulmica, BiologIa Molecular y FisiologIa, Facultad de Medicina, Valladolid, Spain Abstract: The importance of individual members of the and biological effects, BDNF and NT-3 are responsible neurotrophin gene family for avian inner ear development for the survival and neurite outgrowth of mammalian is not clearly defined. Here we address the role of two cochlear neurons, acting via their high-affinity recep- neurotrophins, brain-derived neurotrophic factor (BDNF) tors TrkB and TrkC, respectively (Ernfors et al., 1992; and nerve growth factor (NGF), for innervation of the Pirvola et al., 1992, 1994; Tessarollo et al., 1993; chicken cochlea. We have used defective herpes simplex Schecterson and Bothwell, 1994; Vazquez et a!., virus type 1 (HSV-1) vectors, or amplicons, to express these neurotrophins in dissociated cultures of cochlear 1994). The recent generation of mice lacking expres- neurons. HSV-1 -mediated expression of BDNF promotes sion of specific neurotrophins or their receptors conclu- neuronal survival similar to the maximal level seen by sively demonstrates the essential role of these genes exogenously added BDNF and exceeds its potency to during development of the peripheral nervous system. produce neurite outgrowth. In contrast, cochlear neurons BDNF and NT-3 together with their high-affinity re- transduced with an amplicon producing bioactive NGF ceptors TrkB and TrkC are essential for neuronal sur- show no response. These results confirm BDNF as an vival and maintenance of target innervation in the important mediator of neurotrophin signaling inside avian mammalian cochlea (Ernfors et al., 1995; Fritsch et cochlear neurons. However, these neurons can be ren- al., 1995; Minichiello et al., 1995; Schimmang et al., dered NGF-responsive by transducing them with the high-affinity receptor for NGF, TrkA. This study underlines 1995). the usefulness of amplicons to study and modify neuro- The sequence of developmental events leading to trophin signaling inside neurons. Key Words: Brain-de- the innervation of the avian cochlea is well described rived neurotrophic factor—Nerve growth factor—TrkA (Whitehead and Morest, 1985a,b). Peripheral fibers receptor—Amplicon—Chicken. start their outgrowth from the chicken cochlear gan- J. Neurochem. 70, 2336—2346 (1998). glion between embryonic day 3 (E3) and E5 and in- vade the developing cochlea around E6 and E7. Subse- quently, synaptogenesis between nerve fibers and the cochlear sensory epithelium takes place between E8 The cochleovestibular ganglion contains sensory and E17. Although several studies addressed the ef- neurons that innervate the receptor hair cells of the fects of the neurotrophin gene family during these pro- peripheral auditory and vestibular system. As develop- cesses, the potential of the individual members to regu- ment proceeds, the cochleovestibular ganglion divides late neuronal survival and differentiation is not clearly into discrete cochlear and vestibular ganglia. Recent defined. BDNF and NT-3 induce cell proliferation in studies show that members of the nerve growth factor E3 cochleovestibular ganglia, showing a maximal ef- (NGF) family of neurotrophin genes play a crucial role fect at 5 ng/ml (Represa et al., 1993). A similar re- during innervation of the vertebrate auditory system by _____________________________________________ the cochlear ganglion. In addition to NGF, this family Received September 12, 1997; revised manuscript received Dc- includes brain-derived neurotrophic factor (BDNF), cember 19, 1997; accepted December 19, 1997. neurotrophin-3 (NT-3), and neurotrophin-4/5 (NT-4/ Address correspondence and reprint requests to Dr. T. Schimmang 5) in mammals and birds (Glass and Yancopoulos, at IBGM, Facultad de Medicina, 47005 Valladolid, Spain. 1993; Hallböök et al., 1995). The biological effects of Abbreviations used: BDNF, brain-derived neurotrophic factor; DMEM, Dulbecco’s modified Eagle’s medium; E, embryonic day; neurotrophins are mediated by tyrosine kinase recep- HH, embryonic stage after Hamburger and Hamilton; HSV- I, herpes tors that are products of the trk gene family (Barbacid, simplex virus type 1; ivu, infectious virus units; NGF, nerve growth 1994). Based on spatiotemporal expression patterns factor; NT-3, neurotrophin-3; PBS, phosphate-buffered saline. 2336
Transcript of Defining Responsiveness of Avian Cochlear Neurons to Brain-Derived Neurotrophic Factor and Nerve...

Journal of NeurochemistryLippincott—Raven Publishers, Philadelphia© 1998 International Society for Neurochemistry
Defining Responsiveness of Avian Cochlear Neurons toBrain-Derived Neurotrophic Factor and Nerve Growth Factor
by HSV-1-Mediated Gene Transfer
Juan José Garrido, Maria Teresa Alonso, Filip Lim, Estela Carnicero, Fernando Giraldez,and Thomas Schimmang
Instituto de Biologla y Genética Molecular, Universidad de Valladolid y Consejo Superior de Investigaciones Cientificas,Departamento de Bioqulmica, BiologIa Molecular y FisiologIa, Facultad de Medicina, Valladolid, Spain
Abstract: The importance of individual members of the and biological effects, BDNF and NT-3 are responsibleneurotrophin gene family for avian inner ear development for the survival and neurite outgrowth of mammalianis not clearly defined. Here we address the role of two cochlear neurons, acting via their high-affinity recep-neurotrophins, brain-derived neurotrophic factor (BDNF) tors TrkB and TrkC, respectively (Ernfors et al., 1992;and nerve growth factor (NGF), for innervation of the Pirvola et al., 1992, 1994; Tessarollo et al., 1993;chicken cochlea. We have used defective herpes simplex Schecterson and Bothwell, 1994; Vazquez et a!.,virus type 1 (HSV-1) vectors, or amplicons, to expressthese neurotrophins in dissociated cultures of cochlear 1994). The recent generation of mice lacking expres-neurons. HSV-1 -mediated expression of BDNF promotes sion of specific neurotrophins or their receptors conclu-neuronal survival similar to the maximal level seen by sively demonstrates the essential role of these genesexogenously added BDNF and exceeds its potency to during development of the peripheral nervous system.produce neurite outgrowth. In contrast, cochlear neurons BDNF and NT-3 together with their high-affinity re-transduced with an amplicon producing bioactive NGF ceptors TrkB and TrkC are essential for neuronal sur-show no response. These results confirm BDNF as an vival and maintenance of target innervation in theimportant mediator of neurotrophin signaling inside avian mammalian cochlea (Ernfors et al., 1995; Fritsch etcochlear neurons. However, these neurons can be ren- al., 1995; Minichiello et al., 1995; Schimmang et al.,dered NGF-responsive by transducing them with thehigh-affinity receptor for NGF, TrkA. This study underlines 1995).the usefulness of amplicons to study and modify neuro- The sequence of developmental events leading totrophin signaling inside neurons. Key Words: Brain-de- the innervation of the avian cochlea is well describedrived neurotrophic factor—Nerve growth factor—TrkA (Whitehead and Morest, 1985a,b). Peripheral fibersreceptor—Amplicon—Chicken. start their outgrowth from the chicken cochlear gan-J. Neurochem. 70, 2336—2346 (1998). glion between embryonic day 3 (E3) and E5 and in-
vade the developing cochlea around E6 and E7. Subse-quently, synaptogenesis between nerve fibers and thecochlear sensory epithelium takes place between E8
The cochleovestibular ganglion contains sensory and E17. Although several studies addressed the ef-neurons that innervate the receptor hair cells of the fects of the neurotrophin gene family during these pro-peripheral auditory and vestibular system. As develop- cesses, the potential of the individual members to regu-ment proceeds, the cochleovestibular ganglion divides late neuronal survival and differentiation is not clearlyinto discrete cochlear and vestibular ganglia. Recent defined. BDNF and NT-3 induce cell proliferation instudies show that members of the nerve growth factor E3 cochleovestibular ganglia, showing a maximal ef-(NGF) family of neurotrophin genes play a crucial role fect at 5 ng/ml (Represa et al., 1993). A similar re-during innervation of the vertebrate auditory system by _____________________________________________the cochlear ganglion. In addition to NGF, this family Received September 12, 1997; revised manuscript received Dc-
includes brain-derived neurotrophic factor (BDNF), cember 19, 1997; accepted December 19, 1997.neurotrophin-3 (NT-3), and neurotrophin-4/5 (NT-4/ Address correspondence and reprint requests to Dr. T. Schimmang
5) in mammals and birds (Glass and Yancopoulos, at IBGM, Facultad de Medicina, 47005 Valladolid, Spain.1993; Hallböök et al., 1995). The biological effects of Abbreviations used: BDNF, brain-derived neurotrophic factor;
DMEM, Dulbecco’s modified Eagle’s medium; E, embryonic day;neurotrophins are mediated by tyrosine kinase recep- HH, embryonic stage after Hamburger and Hamilton; HSV- I, herpestors that are products of the trk gene family (Barbacid, simplex virus type 1; ivu, infectious virus units; NGF, nerve growth1994). Based on spatiotemporal expression patterns factor; NT-3, neurotrophin-3; PBS, phosphate-buffered saline.
2336

NEUROTROPHIN SIGNALING IN COCHLEAR NEURONS 2337
sponse is obtained in the presence of high concentra- mm, and dissociated using a fire-polished Pasteur pipette.tions of NGF (50 ng/ml). Cochleovestibular neurons Cells were resuspended in insulin-free medium [DMEM/F-derived from E3.5 otic vesicles acquire a survival re- 12 with Glutamax-I (GIBCO) supplemented with 10% fetalsponse to BDNF after 1.5 days in culture (Vogel and calf serum, 50 U/ml penicillin, 50 ~.tg/ml streptomycin, 6Davies, 1991). BDNF (10 ng/ml) and NT-4 (30 ng/ mg/ml glucose, 0.5 mM sodium pyruvate, 100 mg/ml trans-ferrin, 60 ng/ml progesterone, 16 ~sg/m!putrescine, and 30ml), but not NT-3 (200 ng/ml), promote long-term nM sodium selenite] and seeded in poly-D-lysine (50 ~ig/survival of cochleovestibular ganglia isolated at E3.75 ml)- and laminin (10 ~.tg/ml)-coated culture dishes at a final(Sokolowski, 1997). In cochlear ganglion explants, density of 1,000—2,000 cells in 0.5 ml per well. After 2 hneurite outgrowth is observed upon treatment with 2 at 37°Cin a 5% CO2 atmosphere, medium was changed tong/ml BDNF from E7 until hatching, maximal effects reduce the serum concentration to 1% and growth factorsbeing obtained between E7 and Eli (Avila et al., or viruses were added. Purified recombinant BDNF was a1993). In cultures of dissociated ganglia, BDNF pro- generous gift from Dr. Rodriguez-Tebar. Human recombi-motes neuronal survival at ES and produces maximal nant NGF was purchased from Austral Biologicals. To neu-responses between E9 and E13 (Avila et al., 1993). tralize NGF, a monoclonal antibody raised against 2.5Smouse NGF (Korsching and Thoenen, 1983; purchased fromLikewise, in the same study, NT-3 supports the sur- Boehringer) was used at 0.5 mg/mI.vival and neuritogenesis of whole and dissociated gan- Conditioned media were collected after 48 h fromglia. In a different study, cochlear explants from E4 pHSVbdnf-, pHSVngf-, pHSVlac-, or mock-infected cells,to E6 show little outgrowth upon treatment with NT- centrifuged at 13,000 rpm for 30 mm in a benchtop centri-3 and NGF, and the least effect with BDNF (Bianchi fuge to eliminate cell debris, and stored at —80°C.Condi-and Cohan, 1993). Similarly, neurotrophins do not tioned media were used at 1:1 dilution with standard me-promote neuronal survival in neuron-enriched cultures dium. Concentrations of BDNF in pHSVbdnf-infected cul-from ES cochleovestibular ganglia. tures were calculated by plotting survival-promoting
In the present study, we define the importance of activities of serial dilutions of the culture media againstBDNF and NGF for chicken cochlear development and dose—response curves of recombinant BDNF.
For measurements of neurite extensions, a Q500MC videoaddress their potential to act via an intracellular or image analyzer wasused (Leica). For each group, data wereextracellular route. We have overexpressed BDNF and collected from 60 neurons.NGF inside cochlear neurons using a defective herpes PCI2 cells were grown at 37°Cin a 5% CO2 atmospheresimplex virus type 1 (HSV-l)-based amplicon vector in DMEM containing penicillin (100 U/rn!), 10% fetal calfat a time point when they show a maxima! survival serum, and 5% horse serum. Cells were infected with 3.7response to exogenous BDNF. Amplicon-mediated ex- >< 1 o~infectious vector units (ivu ) /ml pHSVngfand allowedpression of BDNF and NGF results in high levels of to grow for 5 days.these neurotrophins inside cochlear neurons and theirsecretion into the culture medium. Expression of ImmunocytochemistryAfter 48 h, primary cultures containing neurons and non-BDNF strongly enhances survival of cochlear neurons, neuronal cells were fixed in 4% paraformaldehyde in phos-whereas NGF shows no effect. However, we also dem- phate-buffered saline (PBS) for 30 mm and washed threeonstrate that these neurons can be rendered NGF-re- times in PBS. Endogenous peroxidase was inhibited withsponsive by introduction of the high-affinity NGF re- 0.03% H202 in PBS for 30 mm. Cells were washed twoceptor, TrkA. We conclude that BDNF acts as a major times in PBS, blocked with 10% goat serum in PBS for 1 hregulator of survival for chicken cochlear neurons, at room temperature, and incubated with the supematantwhereas NGF signaling via TrkA is usually not re- from the A2B5 hybridoma diluted 1:1 in PBS at 4°Cfor 12quired. The potential of neurotrophins to act through h. Cultures were washed three times for 15 mm in PBS andautocrine or paracrine routes and the usefulness of the incubated with abiotinylated 1gM antibody (Sigma) dilutedHSV- 1 amplicon system to examine and change re- 1:100 for 2 h at roomtemperature. Finally, cells were washedthree times in PBS, and immunoreactivity wasdetected usingsponsiveness to neurotrophins in neurons are dis- the ABC method (Vectastain kit, Vector Laboratories). Thecussed. A2B5 monoclonal antibody recognizes a cell surface glyco-
lipid on chicken neurons (Eisenbarth et al., 1979; Vogel andMATERIALS AND METHODS Weston, 1988; purchased from the American Type Culture
Collection). A2B5-positive neurons with neurites four timesCell culture longer than cell bodies were scored as surviving neurons.
Cochlear neuron primary cultures were prepared as fol- This number was expressed as the percentage of the totallows: Cochlear ganglia from chicken embryos of stage HH number of neurons plated. Statistics were performed using31, HH 33, HH 34, and HH 35 (according to Hamburger Student’s t test.and Hamilton, 1951) were isolated, avoiding contamination For immunofluorescence studies, cultures were seeded onwith sensory epithelia. Ganglia were placed in Dulbecco’s coverslips. After 2 days, cells were fixed with ice-cold meth-modified Eagle’s medium (DMEM)/F-12 (GIBCO) and anol for 15 mm, washed in PBS, and incubated in PBSwashed in Hanks’ balanced salt solution without Ca
2~or containing 5% goat serum and 0.1% Triton X-l00 for 1 hMg2~.Dissociation was carried out using 0.05% trypsin at room temperature. Cultures were washed in PBS, and the(GIBCO) in Hanks’ balanced salt solution without Ca2~or following primary polyclonal antibodies produced in rabbitsMg2~for 20 mm at 37°C.Cells were washed in DMEM/F- were added, diluted in PBS containing 0.1% Triton X-lOO12 with 10% fetal calf serum, centrifuged at 100 g for 2 and 1% goat serum: anti-BDNF (N-20) raised against a
J. Neurochem., Vol. 70, No. 6, 1998
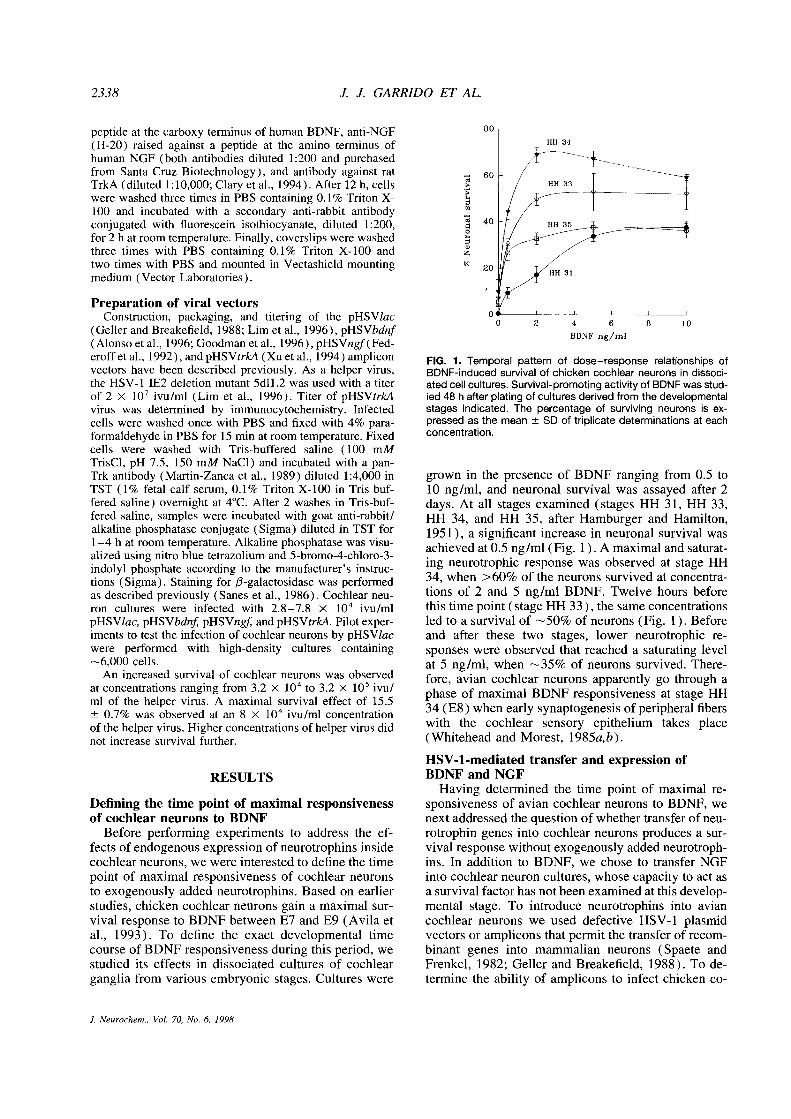
2338 J. J. GARRIDO ET AL.
peptide at the carboxy terminus of human BDNF, anti-NGF(H-20) raised against a peptide at the amino terminus ofhuman NGF (both antibodies diluted 1:200 and purchasedfrom Santa Cruz Biotechnology), and antibody against ratTrkA (diluted 1:10,000; Clary et al., 1994). After 12 h, cellswere washed three times in PBS containing 0.1% Triton X-100 and incubated with a secondary anti-rabbit antibodyconjugated with fluorescein isothiocyanate, diluted 1:200,for 2 h at roomtemperature. Finally, coverslips were washedthree times with PBS containing 0.1% Triton X-l00 andtwo times with PBS and mounted in Vectashield mountingmedium (Vector Laboratories).
Preparation of viral vectorsConstruction, packaging, and titering of the pHSVlac
(Geller andBreakefield, 1988; Lim et al., 1996), pHSVbdnf(Alonsoet al., 1996; Goodman et al., 1996), pHSVngf(Fed-eroffetal., 1992), andpHSVtrkA (Xuet al., 1994) amplicon FIG. 1. Temporal pattern of dose—response relationships ofvectors have been described previously. As a helper virus, BDNF-induced survival of chicken cochlear neurons in dissoci-the HSV-1 1E2 deletion mutant 5dll .2 was used with a titer atedcellcultures. Survival-promoting activity of BDNF wasstud-of 2 X l0~ivu/ml (Lim et al., 1996). Titer of pHSVtrkA led 48 h after plating of cultures derived from the developmentalvirus was determined by immunocytochemistry. Infected stages indicated. The percentage of surviving neurons is ex-cells were washed once with PBS and fixed with 4% para- pressed as the mean ±SD of triplicate determinations at eachformaldehyde in PBS for 15 mm at room temperature. Fixed concentration.cells were washed with Tris-buffered saline (100 mMTrisCl, pH 7.5, 150 mM NaC1) and incubated with a pan-Trk antibody (Martin-Zanca et al., 1989) diluted 1:4,000 in grown in the presence of BDNF ranging from 0.5 toTST (1% fetal calf serum, 0.1% Triton X-l00 in Tris-buf- 10 ng/ml, and neuronal survival was assayed after 2fered saline) overnight at 4°C.After 2 washes in Tris-buf- days. At all stages examined (stages HH 31, HH 33,fered saline, samples were incubated with goat anti-rabbit/ HH 34, and HH 35, after Hamburger and Hamilton,alkaline phosphatase conjugate (Sigma) diluted in TST for 1951), a significant increase in neuronal survival was1—4 h at room temperature. Alkaline phosphatase was visu- achieved at 0.5 ng/ml (Fig. 1). A maximal and saturat-alized using nitro blue tetrazolium and 5-bromo-4-chloro-3-indolyl phosphate according to the manufacturer’s instruc- ing neurotrophic response was observed at stage HHtions (Sigma). Staining for /3-galactosidase was performed 34, when >60% of the neurons survived at concentra-as described previously (Sanes et al., 1986). Cochlear neu- tions of 2 and S ng/ml BDNF. Twelve hours beforeron cultures were infected with 2.8—7.8 X 10” ivu/ml this time point (stage HH 33), the same concentrationspHSVlac, pHSVbdnf pHSVngj~andpHSVtrkA. Pilot exper- led to a survival of ~-~S0%of neurons (Fig. 1). Beforeiments to test the infection of cochlear neurons by pHSVlac and after these two stages, lower neurotrophic re-were performed with high-density cultures containing sponses were observed that reached a saturating level~-.~6,000cells. at 5 ng/ml, when ~35% of neurons survived. There-
An increased survival of cochlear neurons was observed fore, avian cochlear neurons apparently go through aat concentrations ranging from 3.2 x l0~to 3.2 x 10 ivu/ phase of maximal BDNF responsiveness at stage HHml of the helper virus. A maximal survival effect of 15.5±0.7% was observed at an 8 x io~ivu/ml concentration 34 (E8) when early synaptogenesis of peripheral fibersof the helper virus. Higher concentrations of helper virus did with the cochlear sensory epithelium takes placenot increase survival further. (Whitehead and Morest, 1985a,b).
HSV-1-mediated transfer and expression ofRESULTS BDNF and NGF
Having determined the time point of maximal re-Defining the time point of maximal responsiveness sponsiveness of avian cochlear neurons to BDNF, weof cochlear neurons to BDNF next addressed the question of whether transfer of neu-
Before performing experiments to address the ef- rotrophin genes into cochlear neurons produces a sur-fects of endogenous expression of neurotrophins inside vival response without exogenously added neurotroph-cochlear neurons, we were interested to define the time ins. In addition to BDNF, we chose to transfer NGFpoint of maximal responsiveness of cochlear neurons into cochlear neuron cultures, whose capacity to act asto exogenously added neurotrophins. Based on earlier a survival factor has not been examined at this develop-studies, chicken cochlear neurons gain a maximal sur- mental stage. To introduce neurotrophins into avianvival response to BDNF between E7 and E9 (Avila et cochlear neurons we used defective HSV-1 plasmidal., 1993). To define the exact developmental time vectors or amplicons that permit the transfer of recom-course of BDNF responsiveness during this period, we binant genes into mammalian neurons (Spaete andstudied its effects in dissociated cultures of cochlear Frenkel, 1982; Geller and Breakefield, 1988). To de-ganglia from various embryonic stages. Cultures were termine the ability of amplicons to infect chicken co-
J. Neurochem., Vol. 70, No. 6, 1998
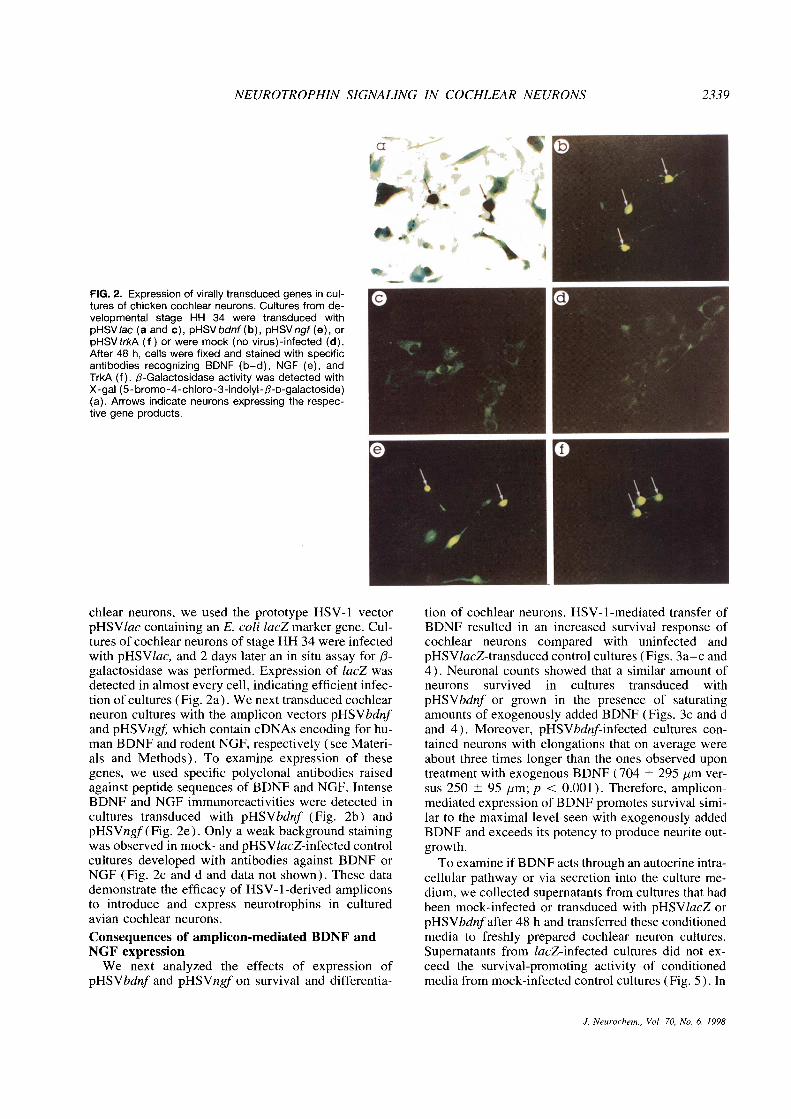
NEUROTROPHIN SIGNALING IN COCHLEAR NEURONS 2339
FIG. 2. Expression of virally transducedgenes in cul-tures of chicken cochlear neurons. Cultures from de-velopmental stage HH 34 were transduced withpHSV/ac (a and c), pHSVbdnf(b), pHSVngf(e), orpHSVtrkA (f) or were mock (no virus)-infected (d).After 48 h, cells were fixed and stained with specificantibodies recognizing BDNF (b—d), NGF (e), andTrkA (f). f3-Galactosidase activity was detected withX-gal (5- bromo-4-chloro-3-indolyl-/3-D-galactoside)(a). Arrows indicate neurons expressing the respec-tive gene products.
chlear neurons, we used the prototype HSV-1 vector tion of cochlear neurons. HSV-1-mediated transfer ofpHSVlac containing an E. coli lacZ marker gene. Cul- BDNF resulted in an increased survival response oftures of coch!ear neurons of stage HH 34 were infected cochlear neurons compared with uninfected andwith pHSVlac, and 2 days later an in situ assay for ~3- pHSV/acZ-transduced control cultures (Figs. 3a—c andgalactosidase was performed. Expression of lacZ was 4). Neuronal counts showed that a similar amount ofdetected in almost every cell, indicating efficient infec- neurons survived in cultures transduced withtion of cultures (Fig. 2a). We next transduced cochlear pHSVbdnf or grown in the presence of saturatingneuron cultures with the amplicon vectors pHSVbdnf amounts of exogenously added BDNF (Figs. 3c and dand pHSVngf which contain cDNAs encoding for hu- and 4). Moreover, pHSVbdnf-infected cultures con-man BDNF and rodent NGF, respectively (see Materi- tamed neurons with elongations that on average wereals and Methods). To examine expression of these about three times longer than the ones observed upongenes, we used specific polyclonal antibodies raised treatment with exogenous BDNF (704 ±295 jim ver-against peptide sequences of BDNF and NGF. Intense sus 250 ±95 jim; p < 0.001). Therefore, amplicon-BDNF and NGF immunoreactivities were detected in mediated expression of BDNF promotes survival simi-cultures transduced with pHSVbdnf (Fig. 2b) and lar to the maximal level seen with exogenously addedpHSVngf (Fig. 2e). Only a weak background staining BDNF and exceeds its potency to produce neurite out-was observed in mock- and pHSV1acZ-infected control growth.cultures developed with antibodies against BDNF or Toexamine if BDNF acts through an autocrine intra-NGF (Fig. 2c and d and data not shown). These data cellular pathway or via secretion into the culture me-demonstrate the efficacy of HSV-1-derived amplicons dium, we collected supernatants from cultures that hadto introduce and express neurotrophins in cultured been mock-infected or transduced with pHSV1acZ oravian cochlear neurons. pHSVbdnf after 48 h and transferred these conditioned
Consequences of amplicon-mediated BDNF and media to freshly prepared cochlear neuron cultures.NGF expression Supematants from /acZ-infected cultures did not ex-
We next analyzed the effects of expression of ceed the survival-promoting activity of conditionedpHSVbdnf and pHSVngf on survival and differentia- media from mock-infected control cultures (Fig. S). In
J. Neurochem., Vol. 70, No. 6, 1998
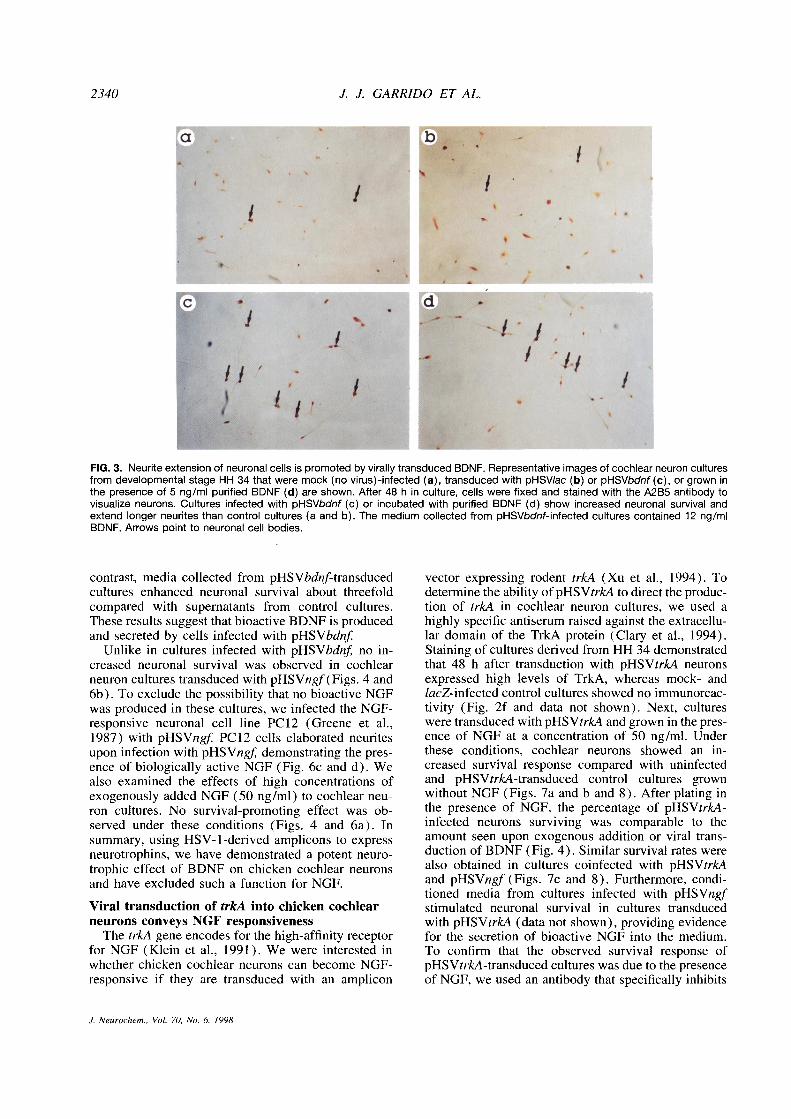
2340 J. J. GARRIDO ET AL.
FIG. 3. Neuriteextension of neuronal cells is promoted by virally transduced BDNF. Representative images of cochlear neuron culturesfrom developmental stage HH 34 that were mock (novirus)-infected (a), transduced with pHSVlac (b) or pHSVbdnf(c), or grown inthe presence of 5 ng/ml purified BDNF (d) are shown. After 48 h in culture, cells were fixed and stained with the A2B5 antibody tovisualize neurons. Cultures infected with pHSVbdnf (c) or incubated with purified BDNF (d) show increased neuronal survival andextend longer neurites than control cultures (a and b). The medium collected from pHSVbdnf-infected cultures contained 12 ng/mIBDNF. Arrows point to neuronal cell bodies.
contrast, media collected from pHSVbdnf-transduced vector expressing rodent trkA (Xu et al., 1994). Tocultures enhanced neuronal survival about threefold determine the ability of pHSVtrkA to direct the produc-compared with supernatants from control cultures. tion of trkA in cochlear neuron cultures, we used aThese results suggest that bioactive BDNF is produced highly specific antiserum raised against the extracellu-and secreted by cells infected with pHSVbdnf lar domain of the TrkA protein (Clary et a!., 1994).
Unlike in cultures infected with pHSVbdnf no in- Staining of cultures derived from HH 34 demonstratedcreased neuronal survival was observed in cochlear that 48 h after transduction with pHSVtrkA neuronsneuron cultures transduced with pHSVngf (Figs. 4 and expressed high levels of TrkA, whereas mock- and6b). To exclude the possibility that no bioactive NGF lacZ-infected control cultures showed no immunoreac-was produced in these cultures, we infected the NGF- tivity (Fig. 2f and data not shown). Next, culturesresponsive neuronal cell line PC12 (Greene et al., were transduced with pHSVtrkA and grown in thepres-1987) with pHSVngf PC12 cells elaborated neurites ence of NGF at a concentration of SO ng/m!. Underupon infection with pHSVngf demonstrating the pres- these conditions, cochlear neurons showed an in-ence of biologically active NGF (Fig. 6c and d). We creased survival response compared with uninfectedalso examined the effects of high concentrations of and pHSVtrkA-transduced control cultures grownexogenously added NGF (SO ng/ml) to cochlear neu- without NGF (Figs. 7a and b and 8). After plating inron cultures. No survival-promoting effect was ob- the presence of NGF, the percentage of pHSVtrkA-served under these conditions (Figs. 4 and 6a). In infected neurons surviving was comparable to thesummary, using HSV-l-derived amplicons to express amount seen upon exogenous addition or viral trans-neurotrophins, we have demonstrated a potent neuro- duction of BDNF (Fig. 4). Similar survival rates weretrophic effect of BDNF on chicken cochlear neurons also obtained in cultures coinfected with pHSVtrkAand have excluded such a function for NGF. and pHSVngf (Figs. 7c and 8). Furthermore, condi-
tioned media from cultures infected with pHSVngfViral transduction of trkA into chicken cochlear stimulated neuronal survival in cultures transducedneurons conveys NGF responsiveness with pHSVtrkA (data not shown), providing evidence
The trkA gene encodes for the high-affinity receptor for the secretion of bioactive NGF into the medium.for NGF (Klein et al., 1991). We were interested in To confirm that the observed survival response ofwhether chicken cochlear neurons can become NGF- pHSVtrkA-transduced cultures was due to the presenceresponsive if they are transduced with an amplicon of NGF, we used an antibody that specifically inhibits
J. Neurochem., Vol. 70. No. 6, 1998
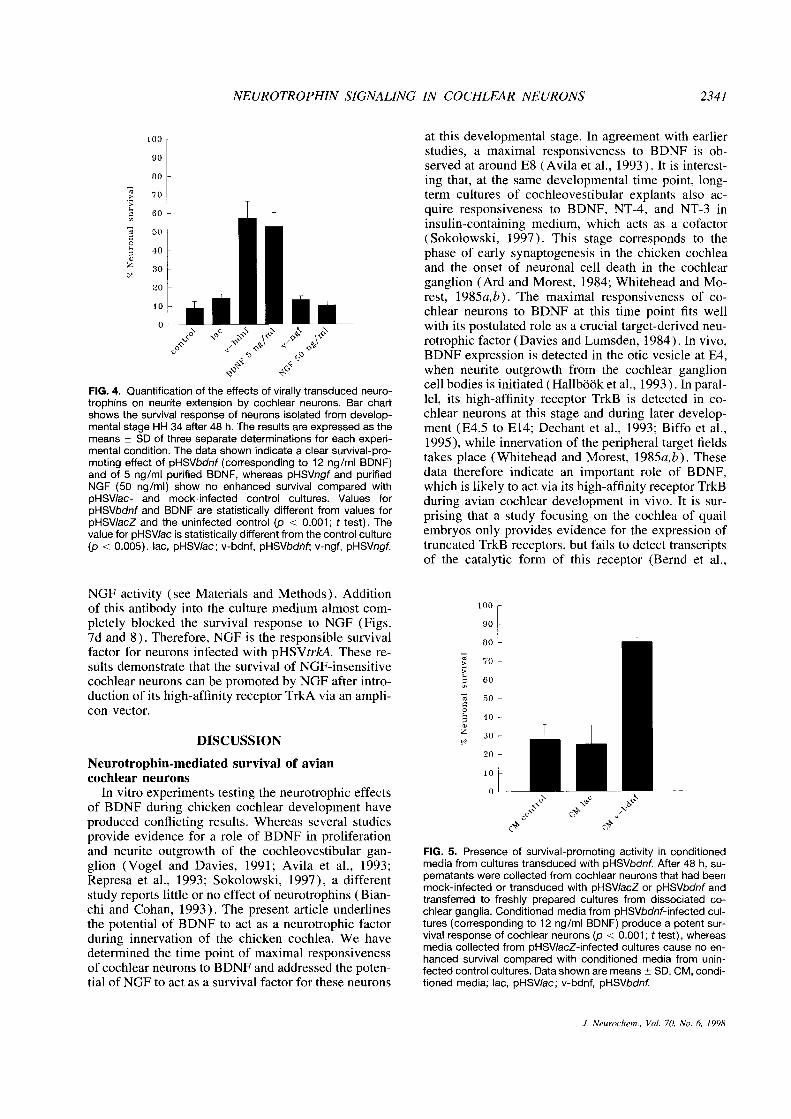
NEUROTROPHIN SIGNALING IN COCHLEAR NEURONS 2341
at this developmental stage. In agreement with earlierstudies, a maximal responsiveness to BDNF is ob-served at around E8 (Avila et al., 1993). It is interest-ing that, at the same developmental time point, long-term cultures of cochleovestibular explants also ac-quire responsiveness to BDNF, NT-4, and NT-3 ininsulin-containing medium, which acts as a cofactor(Sokolowski, 1997). This stage corresponds to thephase of early synaptogenesis in the chicken cochleaand the onset of neuronal cell death in the cochlearganglion (Ard and Morest, 1984; Whitehead and Mo-rest, 1985a,b). ‘The maximal responsiveness of co-
chlear neurons to BDNF at this time point fits wellwith its postulated role as a crucial target-derived neu-rotrophic factor (Davies and Lumsden, 1984). In vivo,BDNF expression is detected in the otic vesicle at E4,when neurite outgrowth from the cochlear ganglioncell bodies is initiated (Hallböök et al., 1993). In paral-FIG. 4. Quantification of theeffects of virally transduced neuro-
trophins on neurite extension by cochlear neurons. Bar chart lel, its high-affinity receptor TrkB is detected in co-shows the survival response of neurons isolated from develop- chlear neurons at this stage and during later develop-mental stage HH 34 after 48 h. The results are expressed as the ment (E4.S to E14; Dechant et al., 1993; Biffo et al.,means ±SD of three separate determinations for each experi- 1995), while innervation of the peripheral target fieldsmental condition. The data shown indicate a clear survival-pro-moting effect of pHSVbdnf (corresponding to 12 ng/ml BDNF) takes place (Whitehead and Morest, 198Sa,b). Theseand of 5 ng/mI purified BDNF, whereas pHSVngf and purified data therefore indicate an important role of BDNF,NGF (50 ng/ml) show no enhanced survival compared with which is likely to act via its high-affinity receptor TrkBpHSV/ac- and mock-infected control cultures. Values for during avian cochlear development in vivo. It is sur-pHSVbdnf and BDNF are statistically different from values for prising that a study focusing on the cochlea of quailpHSV/acZ and the uninfected control (p < 0.001; t test). Thevalue for pHSV/ac is statistically different from thecontrol culture embryos only provides evidence for the expression of(p <0.005). ac, pHSVIac; v-bdnf, pHSVbdnf; v-ngf, pHSVngf. truncated TrkB receptors, but fails to detect transcripts
of the catalytic form of this receptor (Bernd et al.,
NGF activity (see Materials and Methods). Additionof this antibody into the culture medium almost com-pletely blocked the survival response to NGF (Figs.7d and 8). Therefore, NGF is the responsible survivalfactor for neurons infected with pHSVtrkA. These re-sults demonstrate that the survival of NGF-insensitivecochlear neurons can be promoted by NGF after intro-duction of its high-affinity receptor TrkA via an ampli-con vector.
DISCUSSION
Neurotrophin-mediated survival of aviancochlear neurons
In vitro experiments testing the neurotrophic effectsof BDNF during chicken cochlear development haveproduced conflicting results. Whereas several studiesprovide evidence for a role of BDNF in proliferationand neurite outgrowth of the cochleovestibular gan- FIG. 5. Presence of survival-promoting activity in conditionedglion (Vogel and Davies, 1991; Avila et al., 1993; media from cultures transduced with pHSVbdnf. After 48 h, su-Represa et al., 1993; Sokolowski, 1997), a different pernatants were collected from cochlear neurons that had beenmock-infected or transduced with pHSVIacZ or pHSVbdnf andstudy reports little or no effect of neurotrophins (Bian- transferred to freshly prepared cultures from dissociated co-chi and Cohan, 1993). The present article underlines chlear ganglia. Conditioned media from pHSVbdnf-infected cul-the potential of BDNF to act as a neurotrophic factor tures (corresponding to 12 ng/ml BDNF) produce a potent sur-during innervation of the chicken cochlea. We have vival response of cochlear neurons (p < 0.001; t test), whereas
media collected from pHSVIacZ-infected cultures cause no en-determined the time point of maximal responsiveness hanced survival compared with conditioned media from unin-of cochlear neurons to BDNF and addressed the poten- fected control cultures. Data shown are means±SD. CM, condi-tial of NGF to act as a survival factor for these neurons tioned media; lac, pHSVIac; v-bdnf, pHSVbdnf.
J. Neurochem., Vol. 70. No. 6, 1998

2342 J. J. GARRIDO ET AL.
FIG. 6. NGF fails to promote neuronal survival of cochlear neurons. Representative images of cochlear neuron cultures from develop-mental stage HH 34 that were grown in the presence of 50 ng/ml purified NGF (a) or infected with pHSVngf (b) are shown. After 48h in culture, cells were fixed and stained with theA2B5 antibody to visualize neurons. Arrows point to neuronal cell bodies. PC12 cellswere mock-infected (c) or transduced with pHSVngf (d), and phase-contrast images of the cultures were taken after 5 days. PC12cells infected with pHSVngf exhibit differentiated morphology marked by neurite outgrowth.
FIG. 7. Introduction of trkA into cochlear neurons mediates a NGF survival response. Representative images of cochlear neuroncultures from developmental stage HH 34 that were transduced with pHSVtrk4 (a), pHSVtrk4 in the presence of 50 ng/ml purifiedNGF (b), pHSVngf and pHSVtrkA (c), or pHSVtrkA in the presence of 50 ng/ml NGF and a 0.5 mg/mI concentration of antibodyagainst NGF (d) are shown. After 48 h in culture, cells were fixed and stained with the A2B5 antibody to visualize neurons. Culturesinfected with pHSVtrk4 and grown in the presence of NGF(b) or coinfected with pHSVngf and pHSVtrkA (c) show increased neuronalsurvival and extend longerneurites than control cultures transduced with pHSVtrkA only (a). The antibody directed against NGF blockssubstantially the neurite-promoting effects of NGF in pHSVtrk4-transduced cultures (d). Arrows point to neuronal cell bodies.
J. Neurochem., Vol. 70, No. 6, 1998
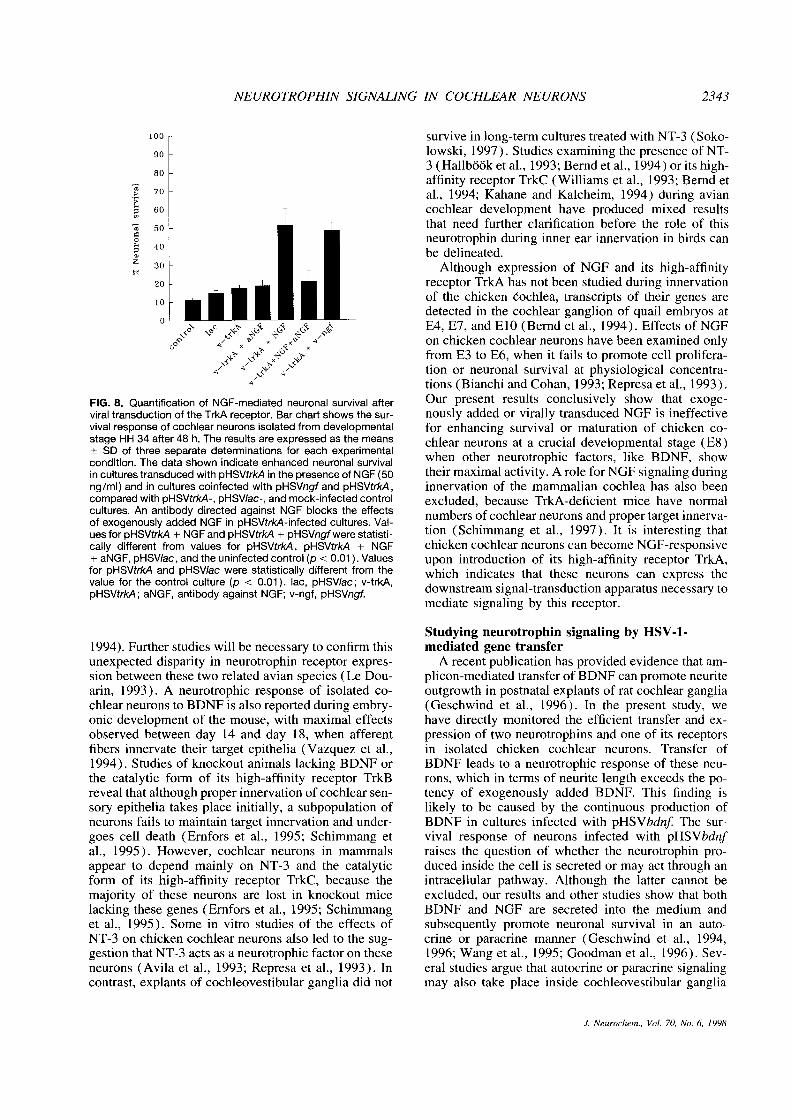
NEUROTROPHIN SIGNALING IN COCHLEAR NEURONS 2343
survive in long-term cultures treated with NT-3 (Soko-lowski, 1997). Studies examining the presence of NT-3 (Hallböök et al., 1993; Bernd et al., 1994) or its high-affinity receptor TrkC (Williams et al., 1993; Bernd etal., 1994; Kahane and Kalcheim, 1994) during aviancochlear development have produced mixed resultsthat need further clarification before the role of thisneurotrophin during inner ear innervation in birds canbe delineated.
Although expression of NGF and its high-affinityreceptor TrkA has not been studied during innervationof the chicken dochlea, transcripts of their genes aredetected in the cochlear ganglion of quail embryos atE4, E7, and ElO (Bernd et al., 1994). Effects of NGFon chicken cochlear neurons have been examined onlyfrom E3 to E6, when it fails to promote cell prolifera-tion or neuronal survival at physiological concentra-tions (Bianchi and Cohan, 1993; Represa et al., 1993).
FIG. 8. Quantification of NGF-mediated neuronal survival after Our present results conclusively show that exoge-viral transduction of theTrkA receptor. Bar chartshows the sur- nous!y added or virally transduced NGF is ineffectivevival response of cochlear neurons isolatedfrom developmental for enhancing survival or maturation of chicken co-stage HH 34 after 48 h. The resu!ts areexpressed as the means chlear neurons at a crucial developmental stage (E8)±SD of three separate determinations for each experimentalcondition. The data shown indicate enhanced neuronal survival when other neurotrophic factors, like BDNF, showin cultures transduced with pHSVtrkA in the presence of NGF (50 their maximal activity. A role for NGF signaling duringng/ml) and in cultures coinfected with pHSVngf and pHSVtrkA, innervation of the mammalian cochlea has also beencompared with pHSVtrkA-, pHSV/ac-, and mock-infectedcontrol excluded, because TrkA-deficient mice have normalcultures. An antibody directed against NOF blocks the effects numbers of cochlear neurons and proper target innerva-of exogenously added NGF in pHSVtrk4-infected cultures. Val- . . .
ues for pHSVtrk.4 + NGF and pHSVtrk4 + pHSVngfwere statisti- tion (Schimmang et al., 1997). It is interesting thatcally different from values for pHSVtrkA, pHSVtrk4 + NGF chicken cochlear neurons can become NGF-responsive+ aNGF, pHSV!ac, andthe uninfectedcontrol (p <0.01). Values upon introduction of its high-affinity receptor TrkA,for pHSVtrkA and pHSV/ac were statistically different from the which indicates that these neurons can express thevalue for the control culture (p < 0.01). lac, pHSV/ac; v-trkA,pHSVtrk4; aNGF, antibody against NGF; v-ngf, pHSVngf. downstream signal-transduction apparatus necessary to
mediate signaling by this receptor.
Studying neurotrophin signaling by HSV-1-1994). Further studies will be necessary to confirm this mediated gene transferunexpected disparity in neurotrophin receptor expres- A recent publication has provided evidence that am-sion between these two related avian species (Le Dou- plicon-mediated transfer of BDNF can promote neuritearm, 1993). A neurotrophic response of isolated co- outgrowth in postnatal explants of rat cochlear gangliachlear neurons to BDNF is also reported during embry- (Geschwind et al., 1996). In the present study, weonic development of the mouse, with maximal effects have directly monitored the efficient transfer and ex-observed between day 14 and day 18, when afferent pression of two neurotrophins and one of its receptorsfibers innervate their target epithelia (Vazquez et al., in isolated chicken cochlear neurons. Transfer of1994). Studies of knockout animals lacking BDNF or BDNF leads to a neurotrophic response of these neu-the catalytic form of its high-affinity receptor TrkB rons, which in terms of neurite length exceeds the po-reveal that although proper innervation of cochlearsen- tency of exogenously added BDNF. This finding issory epithelia takes place initially, a subpopulation of likely to be caused by the continuous production ofneurons fails to maintain target innervation and under- BDNF in cultures infected with pHSVbdnf The sur-goes cell death (Emfors et al., 1995; Schimmang et vival response of neurons infected with pHSVbdnfal., 1995). However, cochlear neurons in mammals raises the question of whether the neurotrophin pro-appear to depend mainly on NT-3 and the catalytic duced inside the cell is secreted or may act through anform of its high-affinity receptor TrkC, because the intracellular pathway. Although the latter cannot bemajority of these neurons are lost in knockout mice excluded, our results and other studies show that bothlacking these genes (Ernfors et al., 1995; Schimmang BDNF and NGF are secreted into the medium andet al., 1995). Some in vitro studies of the effects of subsequently promote neuronal survival in an auto-NT-3 on chicken cochlear neurons also led to the sug- crine or paracrine manner (Geschwind et al., 1994,gestion that NT-3 acts as a neurotrophic factor on these 1996; Wang et al., 1995; Goodman et al., 1996). Sev-neurons (Avila et al., 1993; Represa et al., 1993). In eral studies argue that autocrine or paracnne signalingcontrast, explants of cochleovestibular ganglia did not may also take place inside cochleovestibular ganglia
J. Neurochem., Vol. 70, No. 6. 1998

2344 J. J. GARRIDO ET AL.
in vivo. Expression of BDNF is observed during for- control cultures (11.2 ±1.0%) (Fig. 8). The survivalmation of the cochleovestibular ganglion of rodent em- response observed in pHSVtrkA-infected cultures ap-bryos (Ernfors et al., 1992; Schecterson and Bothwell, pears not to be caused by the presence of NGF in the1994). By using reverse transcription—polymerase culture medium, because antibodies inhibiting NGFchain reaction, NGF, BDNF, and NT-3 are found in the activity do not block this effect (Fig. 8). However, itcochlear ganglion during various stages of embryonic is surprising that we found that the helper virus on itsquail development (E4, E7, and ElO; Bemd et al., own produces a weak survival response with a maxi-1994). In contrast, in situ RNA hybridizations per- mal level of 15.5 ±0.7% when different virus concen-formed on chicken cochlear ganglia fail to detect trations were tested (see Materials and Methods). TheBDNF and NT-3 at E4 (Hallböok et al., 1993) and helper virus is therefore also likely to cause at leastBDNF at E7 (Biffo et al., 1995). In addition, chicken part of the effects seen in pHSVtrkA- and pHSVlacZ-cochleovestibular neurons isolated at E3.S hardly sur- infected cultures. The unexpected survival responsevive without BDNF in culture (Vogel and Davies, produced by the helper virus may be explained by the1991), arguing against an autocrine mechanism of sur- expression of viral gene products that block apoptosisvival during this developmental stage. Finally, studies (Leopardi and Roizman, 1996; Koyama and Miwa,in knockout mice show that neurotrophin signaling is 1997). Although these unwanted side effects of thenot required for cell division and formation of the co- amplicon system do not change the interpretation ofchleovestibular ganglion (Ernfors et al., 1995; Schim- our experimental data, caution must be taken whenmang et a!., 1995; Bianchi et al., 1996). Therefore, minor changes of survival responses are monitored.further experiments are necessary to clarify the capac- Finally, although we have only examined short-termity of neurotrophins to regulate neuronal survival in- effects of amplicon-mediated gene transfer in thisside cochlear ganglia by an autocrine or paracrine study, our preliminary experiments suggest that sur-mode. vival of cochlear neurons can be maintained in
Failure of neurotrophins to promote survival in neu- pHSVbdnf-infected E8 cultures for almost 1 week.ron-enriched cultures from cochleovestibular ganglia Similar results are obtained with neurons treated withsuggested that neurotrophin signaling may be mediated exogenous BDNF, which is freshly added to the cultureby nonneuronal cells (Bianchi and Cohan, 1993). In- medium every second day. BDNF is probably unabledeed a paracrine action of nonneuronal cells that pro- to maintain survival of cochlear neurons for longerduce NT-3 and thereby promote survival and differen- periods because neurons lose their capacity to respondtiation of sympathetic neuroblasts before they depend to this factor due to neuronal maturation in culture.on target-derived factors has been described recently Correspondingly, neurons isolated from ElS do not(Verdi et al., 1996). In contrast, in pHSVbdnf-infected show any survival response upon infection withcultures derived from E8 cochlear ganglia, we often pHSVbdnf or addition of exogenous BDNF (J. J. Gar-find neurons with very long elongations that do not rido and T. Schimmang, unpublished observations).contact other neuronal or nonneuronal cells. This oh- Although neurotrophins appear to lose their capacityservation indicates that neurotrophins probably interact to promote survival of cochlear neurons during laterdirectly with cochlear neurons during this phase of development, recent studies in postnatal or adult ani-target innervation. mals suggest that they can act as neuroprotective
In contrast to BDNF, no neurotrophic response is agents against ototoxic drugs (Emfors et al., 1996;observed upon expression of NGF in cochlear neuron Zheng and Gao, 1996). In this context, it will be inter-cultures. The usefulness of the amplicon system to esting to use the amplicon system to express neuro-modify gene expression inside neurons is clearly dem- trophins inside cochlear neurons and examine their po-onstrated by the efficient introduction of a TrkA recep- tential to prevent ototoxic damage.br gene that conveys a neurotrophic response uponexogenously added or virally transduced NGF. These Acknowledgment: We would like to thank Dr. Rodn-results thus confirm previous studies that have shown guez-Tebar for the generous gift of BDNF, Dr. L. Reichardtthat NGF-nonresponsive neuronal populations, like for anti-TrkA antibodies, and Dr. Martin-Zanca for pan-Trkmotoneurons or nodose neurons, can be converted into antibodies and PC12 cells. This research was supported byNGF-responsive ones by the introduction of a TrkA grants from FIS 96/1442 (M.T.A.) and DGCYT PM95/receptor (Xu et al., 1994). 0087 (F.G.).
A crucial issue when using amplicon vectors is tomonitor potential cytotoxic effects caused by the virus REFERENCES(Johnson et al., 1992; Ho et al., 1995). During our Alonso M. T., Lim F., Nunez L., RepresaJ., Giraldez F., and Schim-experiments, we have not observed increased neuronal mang T. (1996) HSV-l vector mediated transfer of BDNF intocell death in amplicon-infected cultures versus unin- cerebellar granule cells. Neuroreport 7, 3105—3108.fected control cultures. On the contrary we found a Ard M. D. and Morest D. K. (1984)Cell death during the develop-
ment ofthe cochlear and vestibular ganglia of the chick. mt. J.weak, but significant, increase of neuronal survival in D Ne ‘ 2 535 547cultures infected with pHSVtrkA (17.6 ± 1.6%) or Avila M. A., V INtl Romero G., Mato J. M., Giraldez F.,pHSV1acZ (14.8 ± 1.6%) compared with uninfected Van Dc Water T. R., and Represa J. (1993) Brain-derived neu-
.1. Neurochem., Vol. 70, No. 6, 1998

NEUROTROPHIN SIGNALING IN COCHLEAR NEURONS 2345
rotrophic factor and neurotrophin-3 support the survival and treatment and experimental exploitation. Methods Enzymol.neuritogenesis response of developing cochleovestibular gan- 147, 207—215.glion neurons. Dev. Biol. 159, 266—275. Hallbook F., Ibánez C. F., Ebendal T., and Persson H. (1993) Cellu-
Barbacid M. (1994) The Trk family of neurotrophin receptors. J. lar localization of brain-derived neurotrophic factor and neuro-Neurobiol. 25, 1386—1403. trophin-3 mRNA expression in the early chicken embryo. Eur.
Bemd P., Zhang D., Yao L., and Rozenberg I. (1994) The potential J. Neurosci. 5, 1—14.of nerve growth factor, brain-derived neurotrophic factor and Hallbbok F., Backstrom A., Kullander K., Kylberg A., Williams R.,neurotrophin-3 in avian cochlear and vestibular ganglia devel- and Ebendal T. (1995) Neurotrophins and their receptors inopment. mt. J. Dcv. Neurosci. 12, 709—723. chicken neuronal development. mt. J. Dcv. Biol. 39, 855—868.
Bianchi L. M. and Cohan C. 5. (1993) Effects of the neurotrophins Hamburger V. and Hamilton H. L. (1951) A series of normal stagesand CNTF on developing statoacoustic neurons: comparison in the development of the chick embryo. J. Morphol. 88, 49—with an otocyst-derived factor. Dcv. Biol. 159, 353—365. 92.
Bianchi L. M., Conover J. C., Fritsch B., DeChiara T., Lindsay Ho D. Y., Fink S. L., Lawrence M. S., Meier T. J., Saydam T. C.,R. M., and Yancopoulos G. D. (1996) Degeneration of vestibu- Dash R., and Sapolsky R. M. (1995) Herpes simplex viruslar neurons in late embryogenesis of both heterozygous and vector system: analysis of its in vivo and in vitro cytopathichomozygousBDNF null mutant mice. Development 122, 1965— effects. J. Neurosci. Methods 57, 205—215.1973. Johnson P. A., Miyanohara A., LevineF., Cahill T., and Friedmann
Biffo S., Offenhauser N., Carter B. D., and Barde Y.-A. (1995) T. (1992)Cytotoxicity of a replication-defective mutant of her-Selective binding and internalisation by truncated receptors re- pes simplex virus type 1. J. Virol. 66, 2952—2965.strict the availability of BDNF during development. Develop- Kahane N. and Kalcheim C. (1994) Expression of trkC receptorment 121, 2461—2470. mRNA during development of the avian nervous system. .1.
Clary D. 0., Weskamp G., Austin L. R., and Reichardt L. F. (1994) Neurobiol. 25, 571—584.TrkA cross-linking mimics neuronal responses to nerve growth Klein R., Jing S., Nanduri V., O’Rourke E., and Barbacid M. (1991)factor. Mo!. Biol. Cell 5, 549—563. The trk proto-oncogene encodes a receptor for nerve growth
Davies A. M. and Lumsden A. G. 5. (1984) Relation of target en- factor. Cell 65, 189—197.counter and neuronal death to nerve growth factor respon- Korsching S. and Thoenen H. (1983) Nerve growth factor in sympa-siveness in the developing mouse trigeminal ganglion. J. Comp. thetic ganglia and corresponding target organs of the rat: cone-Neurol. 223, 124—137. lation with density of sympathetic innervation. Proc. Nat!. Acad.
Dechant G., Biffo S., Okazawa H., Kolbeck R., Pottgiesser J., and Sci. USA 80, 3513—3516.Barde Y.-A. (1993) Expression and binding characteristics of Koyama A. H. and Miwa Y. (1997) Suppression of apoptotic DNAthe BDNF receptor chick trkB. Development 119, 545—558. fragmentation in herpes simplex virus type I-infected cells. J.
Eisenbarth G. S., Walsh F. S., and Nirenberg M. (1979) Monoclonal Virol. 71, 2567—257 1.antibody to a plasma membrane antigen of neurons. Proc. Nat!. Le Douarin N. M. (1993) Embryonic neural chimaeras in the studyAcad. Sci. USA 76, 4913—4917. of brain development. Trends Neurosci. 16, 64—72.
Ernfors P., Merlio i-P., and Persson H. (1992) Cells expressing Leopardi R. and Roizman B. (1996) The herpes simplex virus majormRNA for neurotrophins and their receptors during embryonic regulatory protein 1CP4 blocks apoptosis induced by the virusrat development. Eur. J. Neurosci. 4, 1140—1158. or by hyperthermia. Proc. Nat!. Acad. Sci. USA 93, 9583—9587.
Ernfors P., Van de Water T., Loring J., and Jaenisch R. (1995) Lim F., Hartley D., Starr P., Lang P., Song S., Yu L., Wang Y., andComplementary roles of BDNF and NT-3 in vestibular and Geller A. I. (1996) Generation of high-titer defective HSV-lauditory development. Neuron 14, 1153—1164. vectors using an 1E2 deletion mutant and quantitative study of
Emfors P., Duan M. L., ElShamy W. M., and Canlon B. (1996) expression in cultured cortical cells. Biotechniques 20, 460—Protection of auditory neurons from aminoglycoside toxicity by 469.neurotrophin-3. Nat. Med. 2, 463—467. Martin-Zanca D., Oskam R., Mitra G., Copeland T., and Barbacid
Federoff H. J., Geschwind M. D., Geller A. I., and Kessler J. A. M. (1989) Molecular and biochemical characterization of the(1992) Expression of nerve growth factor in vivo from a defec- human trk proto-oncogene. Mo!. Cell. Biol. 9, 24—33.live herpes simplex virus I vector prevents axotomy on sympa- Minichiello L., Piehl F., Vazquez E., Schimmang T., Hökfelt T.,thetic ganglia. Proc. Nat!. Acad. Sci. USA 89, 1636—1640. Represa J., and KleinR. (1995) Differential effects ofcombined
Fritsch B., Silos-Santiago I., Smeyne R., Fagan A. M., and Barbacid trk receptor mutations on dorsal root ganglion and inner earM. (1995) Reduction and loss of inner ear innervation in trkB sensory neurons. Development 121, 4067—4075.and trkC receptor knockout mice: a whole-mount Dii and scan- Pirvola U., Ylikoski J., Palgi J., Lehtonen E., Arumäe U., and Saarmaning electron microscopic analysis. Aud. Neurosci. 1,401—417. M. (1992) Brain-derived neurotrophic and neurotrophin-3
Geller A. I. and Breakefield X. 0. (1988) A defective HSV-l vector mRNAs in the peripheral target fields of developing inner earexpresses Escherichia coli beta-galactosidase in cultured pe- ganglia. Proc. Nati. Acad. Sci. USA 89, 9915—9919.ripheral neurons. Science 241, 1667—1669. Pirvola U., Arumäe U., Moshynakov M., Palgi J., Saarma M., and
Geschwind M. D., Kessler J. A., Geller A. I., and Federoff H. J. Ylikoski 1. (1994) Coordinated expression and function of neu-(1994) Transfer of the nerve growth factor gene into cell lines rotrophins and their receptors in the rat inner ear during targetand cultured neurons using a defective herpes simplex virus innervation. Hear. Res. 75, 131—144.vector. Transfer of the NGF gene into cells by a HSV- 1 vector. Represa J., Avila M. A., Romero G., Mato J. M., Giraldez F., andMo!. Brain Res. 24, 3 27—335. Varela-Nieto I. (1993) Brain-derived neurotrophic factor and
Geschwind M. D., Hartnick C. J., Liu W., Amat J., Van de Water neurotrophin-3 induce cell proliferation in the cochleovestibularT. R., and Federoff H. J. (1996) Defective HSV-l vector cx- ganglion through a glycosylphosphatidylinositol signaling sys-pressing BDNF in auditory ganglia elicits neurite outgrowth: tem. Dcv. Biol. 159, 257—265.model for treatment ofneuron loss following cochlear degenera- Sanes J. R., Rubinstein J. L. R., and Nicholas J. F. (1986) Use of alion. Hum. Gene Ther. 7, 173— 182. recombinant retrovirus to study post-implantation cell lineage
Glass D. J. and Yancopoulos G. D. (1993) The neurotrophins and in mouse embryos. EMBO J. 5, 3133—3142.their receptors. Trends Cell Biol. 3, 262—268. Schecterson L. C. and Bothwell M. (1994) Neurotrophin and neuro-
Goodman L. J., Valverde J., Lim F., Geschwind M. D., Federoff J., trophin receptor mRNA expression in developing inner ear.GellerA., and Hefti F. (1996) Regulated release of BDNF from Hear. Res. 73, 92—100.the somatodendritic domain of hippocampal pyramidal neurons. Schimmang T., Minichiello L., Vazquez E., San Jose I., Giraldez F.,Mol. Cell. Neurosci. 7,222—238. Klein R., and Represa J. (1995) Developing inner ear sensory
Greene L. A., Aletta J. M., Rukenstein A., and Green S. H. (1987) neurons require TrkB and TrkC receptors for innervation ofPC 12 pheochromocytoma cells: culture, nerve growth factor their peripheral targets. Development 121, 3381 —3391.
.J. Neurochem., Vol. 70, No. 6, 1998

2346 J. J. GARRIDO ET AL
Schimmang T., Alvarez-Bolado G., Minichiello L., Vazquez E., Gir- factor independence in early sensory neurons is matched to thealdez F., Klein R., and Represa J. (1997) Survival of inner ear time course of target field innervation. Neuron 7, 819—830.sensory neurons in Irk mutant mice. Mech. Dcv. 64, 77—85. Vogel K. S. and Weston J. A. (1988) A subpopulation of cultured
Sokolowski B. H. A. (1997) Quantitative analysis of long-term sur- avian neural crest cells has transient neurogenic potential. Neu-vival and neuritogenesis in vitro: cochleovestibular ganglion of ron 1, 569—577.the chick embryo in BDNF, NT-3, NT-4/5, and insulin. Exp. Wang M. J., Friedmann T., and Johnson P. A. (1995) DifferentiationNeurol. 145, 1—15. of PC12 cells by infection with an HSV-l vector expressing
Spaete R. R. and Frenkel N. (1982) The herpes simplex virus ampli- nerve growth factor. Gene Ther. 2, 323—335.con: a new eukaryotic defective-virus cloning-amplifying vec- Whitehead M. C. and Morest D. K. (1985a) The development oftor. Cell 30, 295—304. innervation patterns in the avian cochlea. Neuroscience 14,
Tessarollo L., Tsoulfas P., Martin-Zanca D., Gilbert D. J., Jenkins 255—276.N. A., Copeland N. G., andParada L. F. (1993) trkC, a receptor WhiteheadM. C. and Morest D. K. (1985b) The growth of cochlearfor neurotrophin-3, is widely expressed in the developing ncr- fibers and the formation of their synaptic endings in the avianvous system and in non-neuronal tissues. Development 118, inner can a study with the electron microscope. Neuroscience463—475. 14, 277—300.
Vazquez E., Van de Water T. R., Del Valle M., Vega J. A., Staecker Williams R., Bäckstrdm A., Ebendal T., and Hallbdbk F. (1993)H., Giraldez F., and Represa J. (1994) Pattern of trkB protein- Molecular cloning and cellular localization of trkC in thelike immunoreactivity in vivo and the in vitro effects of brain- chicken embryo. Dcv. Brain Res. 75, 235—252.derived neurotrophic factor (BDNF) on developing cochlear Xu H., Federoff H., Maragos J., Parada L. F., and Kessler J. A.and vestibular neurons. Anat. Embryo!. (Berl.) 189, 157—167. (1994) Viral transduction of trkA into cultured nodose and
Verdi J. M., Groves A. K., Fariflas I., Jones K., Marchionni M. A., spinal motor neurons conveys NGF responsiveness. Dcv. Biol.Reichardt L. F., and Anderson D. J. (1996) A reciprocal cell— 163, 152—161.cell interaction mediated by NT-3 and neuregulins controls the Zheng J. L. and Gao W.-Q. (1996) Differential damage to auditoryearly survival and development of sympathetic neuroblasts. neurons and hair cells by ototoxins and neuroprotection byNeuron 16, 5 15—527. specific neurotrophins in rat cochlear organotypic cultures. Eur.
Vogel K. S. and Davies A. M. (1991) The duration of neurotrophic J. Neurosci. 8, 1897—1905.
J. Neurochem., Vol. 70, No. 6, 1998




















