
Controls on modern carbonate sedimentation on warm-temperate to arctic coasts, shelves and seamounts...
38
FACIES 132]71-108 PI. 17-23 12 Figs. -- I ERLANGEN 1995 Research Report 'Evolution of Reefs' Controls on Modern Carbonate Sedimentation on Warm-temperate to Arctic Coasts, Shelves and Seamounts in the Northern Hemisphere: Implications for Fossil Counterparts RLidiger Henrich and Andre Freiwald, Bremen (Coordination) ROdiger Henrich, Andr6 Freiwald, Bremen; Christian Betzler, Frankfurt; Beate Bader, Priska Schiller, Christian Samtleben, Kie]; Thomas C. Brachert, Mainz; Achim Wehrmann, Heinrich Zankl, Marburg; Dietrich H. H. K0hlmann, Schmagerow KEYWORDS: NON-TROPICAL CARBONATES - ENVIRONMENTAL CONTROL - SEASONAL1TY-OCEANOGRAPHY- FORAMOL-BRYOMOL- CORALLINE ALGAE - MAERL - BALANIDS - BRYOZOA - LOPHELIA-REEFS - CLADOCORA BANKS - KELP FORESTS - RECENT CONTENTS 1 Inuoduction 2 External controls on non-tropical carbonate sedimentation 3 Introduction to the case studies 4 Non-tropical reefs and buildups 4.1 Coralline algal reefs and maerl pavements 4.2 Coral frameworks in non-tropical seas 4.3 Arctic seamount buildups 5 Non-tropical open shelf carbonates 5.1 Carbonate production in kelp-forests 5.2 Production and redeposition of BgYoMOL-carbonates on deep shelf banks 5.3 Ancient BR'~OMOL-carbonate settings on the Queens- land Plateau 5.4 Balanid-dominated cold water open shelf sedimenta- tion: the Arctic Spitsbergen Bank 6 Major controls and mechanisms of non-tropical carbonate sedimentation References SUMMARY In contrast to the well studied tropical carbonate envi- ronments, interest in non-tropical carbonate deposition was rather low until the basic ideas of the FORAMOL- concept were outlined by LEES & B ~ (1972). In the following two decades studies on non-tropical carbonate settings evolved as a new and exciting branch of carbonate sedimentology (see NEt,SoN 1988). This is archieved in a great number of publications dealing on temperate carbon- DFG-Schwerpunkt BIOGENE SEDIMENTATION ate deposits from numerous coastal and open shelf settings on both hemispheres. The ex- istence of wide extended car- bonate depositional systems and even reefal frameworks in Subarctic and Arctic seas which are in focus by our re- search group made it possible to study modem non-tropical carbonate settings along a latitudinal transect from the warm- temperate Mediterranean Sea to the cold Nordic Seas. Be- cause of increasing seasonality in environmental conditions towards high latitudes, the major controls in biogenic carbon- ate production can be more clearly addressed in these areas. After the initiation of the priority program "Global and re- gional controlling processes of biogenic sedimentation - evo- lution of reefs" by the German Science Foundation four years ago, a set of modem case studies were comparatively analysed specifically with regard to their principle controlling proc- esses: (1) Modem and Holocene coralline algal reefs and rhodolith pavements formed in wave-protected shallow waters along the coast of the Brittany and northern Norway. Their fine- tuned interaction with herbivores resulted in the development of widespread but low-diverse, slowly growing coralline algal frameworks with high competitive value against the rapid- growing phaeophytic communities. Addresses: Prof. Dr. R. Henrich and Dr. A. Freiwald, Fachbereich Geowissenschaften, Universit~t Bremen, Postfach 330440, D-28334 Bremen, Fax: 0421/218-7431; Dr. C. Betzler, Geologisch-Pal~iontologisches Institut, Senckenberganlage 32-34, D-60054 Frankfurt a.M., Fax: 069/798-2958; Dr. T. Brachert, UniversitgttMainz, Institut for Geowissenschaften, D-55099 Mainz, Fax: 06131/394768; Prof. Dr. H. Zankl and A. Wehrmann, Institut ftir Geologie und Pal~iontologie, Universit~it Marburg, Hans-Meerweinstr, D-35032 Marburg, Fax: 06421/288919; Prof. Dr. P. Sch~fer, Dr. C. Samtleben and Dipl. Geol. B. Bader, Geologisch-PalaontologischesIrtstitut, Universit~t Kiel, OlshausenstraBe 40, D-241 I8 Kiel, Fax: 0431/880-4376; Dr. D.HM. Kiahlmann, Ecostation, D-17321 Schmagerow/Vorpommem
Transcript of Controls on modern carbonate sedimentation on warm-temperate to arctic coasts, shelves and seamounts...

FACIES 132]71-108 PI. 17-23 12 Figs. -- I ERLANGEN 1995 Research Report 'Evolution of Reefs'
Controls on Modern Carbonate Sedimentation on Warm-temperate to Arctic Coasts, Shelves and Seamounts in the Northern Hemisphere:
Implications for Fossil Counterparts
RLidiger Henrich and Andre Freiwald, Bremen (Coordination)
ROdiger Henrich, Andr6 Freiwald, Bremen; Christian Betzler, Frankfurt; Beate Bader, Priska Schiller, Christian Samtleben, Kie];
Thomas C. Brachert, Mainz; Achim Wehrmann, Heinrich Zankl, Marburg; Dietrich H. H. K0hlmann, Schmagerow
KEYWORDS: NON-TROPICAL CARBONATES - ENVIRONMENTAL CONTROL - SEASONAL1TY-OCEANOGRAPHY- FORAMOL-BRYOMOL- CORALLINE ALGAE - MAERL - BALANIDS - BRYOZOA - L O P H E L I A - R E E F S -
C L A D O C O R A BANKS - KELP FORESTS - RECENT
CONTENTS
1 Inuoduction 2 External controls on non-tropical carbonate sedimentation 3 Introduction to the case studies 4 Non-tropical reefs and buildups 4.1 Coralline algal reefs and maerl pavements 4.2 Coral frameworks in non-tropical seas 4.3 Arctic seamount buildups 5 Non-tropical open shelf carbonates 5.1 Carbonate production in kelp-forests 5.2 Production and redeposition of BgYoMOL-carbonates
on deep shelf banks 5.3 Ancient BR'~OMOL-carbonate settings on the Queens-
land Plateau 5.4 Balanid-dominated cold water open shelf sedimenta-
tion: the Arctic Spitsbergen Bank 6 Major controls and mechanisms of non-tropical carbonate
sedimentation References
S U M M A R Y
In contrast to the well studied tropical carbonate envi- ronments, interest in non-tropical carbonate deposition was rather low until the basic ideas of the FORAMOL- concept were outlined by LEES & B ~ (1972). In the following two decades studies on non-tropical carbonate settings evolved as a new and exciting branch of carbonate sedimentology (see NEt,SoN 1988). This is archieved in a great number of publications dealing on temperate carbon-
DFG-Schwerpunkt
BIOGENE SEDIMENTATION
ate deposits from numerous coastal and open shelf settings on both hemispheres. The ex- istence of wide extended car- bonate depositional systems and even reefal frameworks in Subarctic and Arctic seas which are in focus by our re- search group made it possible to study modem non-tropical carbonate settings along a latitudinal transect from the warm- temperate Mediterranean Sea to the cold Nordic Seas. Be- cause of increasing seasonality in environmental conditions towards high latitudes, the major controls in biogenic carbon- ate production can be more clearly addressed in these areas. After the initiation of the priority program "Global and re- gional controlling processes of biogenic sedimentation - evo- lution of reefs" by the German Science Foundation four years ago, a set of modem case studies were comparatively analysed specifically with regard to their principle controlling proc- esses:
(1) Modem and Holocene coralline algal reefs and rhodolith pavements formed in wave-protected shallow waters along the coast of the Brittany and northern Norway. Their fine- tuned interaction with herbivores resulted in the development of widespread but low-diverse, slowly growing coralline algal frameworks with high competitive value against the rapid- growing phaeophytic communities.
Addresses: Prof. Dr. R. Henrich and Dr. A. Freiwald, Fachbereich Geowissenschaften, Universit~t Bremen, Postfach 330440, D-28334 Bremen, Fax: 0421/218-7431; Dr. C. Betzler, Geologisch-Pal~iontologisches Institut, Senckenberg anlage 32-34, D-60054 Frankfurt a.M., Fax: 069/798-2958; Dr. T. Brachert, Universitgtt Mainz, Institut for Geowissenschaften, D-55099 Mainz, Fax: 06131/394768; Prof. Dr. H. Zankl and A. Wehrmann, Institut ftir Geologie und Pal~iontologie, Universit~it Marburg, Hans-Meerweinstr, D-35032 Marburg, Fax: 06421/288919; Prof. Dr. P. Sch~fer, Dr. C. Samtleben and Dipl. Geol. B. Bader, Geologisch-Palaontologisches Irtstitut, Universit~t Kiel, OlshausenstraBe 40, D-241 I8 Kiel, Fax: 0431/880-4376; Dr. D.HM. Kiahlmann, Ecostation, D-17321 Schmagerow/Vorpommem

72
(2) The Mediterranean Cladocora caespitosa-banks pro- vide an instructive example of non-tropical hermatypic coral framework construction out of the subtropical-tropical coral reef belt. (3) The geometry and environmental controls of several kilometer long coral reefs formed by the azooxantheUate Lophelia pertusa and Madrepora oculata are studied in more than 250 m water depth in mid and northern Norway. (4) Modem BRvOMOL-Sediments are widely distributed on
non-tropical deeper shelf settings. The formational proc- esses converting bryozoan-thickets into huge piles of sand and gravel dunes are recendy studied on the outer shelves off northern Brittany and off northern Norway. (5) Arctic sponge-bryozoan buildups on the seamount Vesterisbank in the Greenland Sea and (6) balanid-dominated open shelf carbonates on the Spitsbergen Bank form the Arctic endmembers of modem FORAMoL-deposits. Seasonal ice-edge phytoplankton blooms and efficient mechanisms of pelagic-benthic food transfer characterize these depositional settings. Fossil counterparts of each of these modem case studies are discussed in context with their paleoceanographic and environmental settings.
1 INTRODUCTION
Before the fundamental paper of Cr~vE's (1967) uncon- ventional views on cold-water carbonate sedimentation and the now classical FORAMOL-COnCept ~EF.S & BLrLL~ 1972, LEES 1975) the meaning of non-tropical carbonate produc- tion and sedimentation suffered from many prejudices. For example, the cold seas are generally undersaturated with respect to CaCO3. and thus, the taphonomic loss due to rapid dissolution must be a barrier preventing the development of high latitude carbonate sediments (RoDGERS 1957, ALEXAN- DERSSON 1974). However, the cold-water carbonate settings studied show only minor evidence of chemical carbonate dissolution. Much more important are early diagenetic mac- eration processes causing selective dissolution that is in- duced by bacterial decomposition of skeletal-binding or- ganic matter at the sediment surface (ALEXANDERSSON 1972, F~IWALO 1995). Another often cited view is that production rates of high latitude carbonate secreting organisms are much lower and therefore, many organisms show reduced rates of biomineralization compared to warmer water set- tings. This is only partly true. The net-rates of carbonate production in temperate and polenear environments gener- ally are one magnitude lower than in subtropieal-tropical environments (see compilation of data in SARNTHEIN 1973, WEFER et al. 1987). However, cold-water carbonate sediments may be produced at rates as high as some tropical reef sediments (SMrrH 1988 and FREIWAm & HEr,aUCH 1994 for coralline algae). On high latitude shelves and below the fair weather wave base, the loss of skeletal carbonate is mainly forced by bioerosional processes (Aft'AN & FARROW 1984, 1985, FARROW & FYFE 1988, NELSON & BOm',~OLD 1983, YOUNG & NELSON 1988).
Biogenic carbonate deposition and specifically reefs are traditionally considered to be suggestive of tropical and subtropical climatic conditions. However, initial reef stages
constructed by the zooxanthellate scleractinia Cladocora caespitosa are existing also in the eastern part of the warm- temperated Mediterranean Sea (KOrlLMANN et al. 1991). Also reefs and biogenic buildups well beyond the Arctic Circle (F~rWALD 1993a, Fm~rWALD & HENmCH 1994, HENmCrI et al. 1992) are in conflict with these conventional views on biogenic carbonate accumulation. Today, many authors still apply the classicalFoRAMOL-Concept for non-tropicalbiogenic carbonate production introduced by LEES & BUla.ER (1972) and LEES (1975). In an oversimplification of the original FORAMOL-COncept, many investigators consider temperature and salinity as most important factors. However, this subdi- vision does not compare biogenic associations with a de- freed trophic behaviour, but intermingles autotrophic and heterotrophic elements for the designation of the distinct associations. For example, bryozoan dominated facies being characteristic element of the BRYOMOL-type carbonates also may occur in distal positions of tropical systems (Gulf of Aqaba (RE~ss & HOa'I~GER 1984)). Moreover, FOl~.MOL- and BRvOMOL-type carbonates are also found in shallow and deep settings of the tropics. Along the eastern margins of the large oceanic gyres, cold surface currents are deflected towards the equator and nutrient-rich intermediate waters upwell in the coastal and continental marginal sector due to strong offshore winds. In addition, in the aphotic zone of deeper banks and open shelf FORAMOL- and BaVOMOL-Car- bonates may be deposited, while on the nearby shallow- water platforms CHLOROZOAN-Carbonates are accumulated (CARANNANa'E et al. 1988). Furthermore, the existence of large biogenic buildups in seasonally ice-covered areas clearly demonstrates that other external controls than salin- ity and temperature are present. In this study we will discuss combinations of other important external and internal con- trols in non-tropical biogenic carbonate sedimentation. The relevance of different factors will be analysed in a compari- son of selected modem and fossil case studies of non- tropical reefs, biogenic buildups and open shelf carbonate deposits.
2 EXTERNAL CONTROLS ON NON-TROPICAL CARBONATE SEDIMENTATION
Variability of insolation: the driving force for seasonality in middle and high latitudes
Latitudinal, daily and seasonally variations in insolation and different depth penetration of various wave lengths of natural light into sea water are essential for photosynthetic carbonate producers. Such, the habitat, standing stock, di- versity and competition between various groups of benthic algae is determined, and phytoplankton blooms, the primary source of the oceanic food chain, are produced. At high latitudes, extreme seasonal variations in insolation are ob- served. At 70~ the Arctic polar night comprises complete darkness between 26 th of November to 18 t~ of January, while during the Arctic polar summer from 28th of May to the 19 th of July 24 hours of illumination occur. In addition, the angle of light penetration into water is much shallower in polar regions compared to overall steeply inclined insolation at

73
150 ~ 12o* 90 ~ 60* 30 ~ 0 ~ 30* 60 ~ 90* 12o* 150- 180"
n o r t h e r n
c o l d - w a t e r
p h a e o p h y t e b e l t
w a r m - w a t e r
c o r a l r e e f b e l t
0"
~ C7'
R R
,R R R R J
/ R R~" R R R
- _ j
southern
150 ~ 120 ~ 90 ~ 60 ~ 30 ~
L
0 o 30 ~ 60 ~ 900 1200 150 ~ 180 ~
Fig. 1. Global circulation patterns of surface currents and their control on the distribution of the warm water coral reef belt (R = Reef) and the cold water phaeophyte belts (K = kelp-forest). In colder nutrient-rich upwelling areas non-tropical phaeopyhte communities thrive well in the tropics. The carbonate deposits in these regions are characterized by a non-tropical FOR~aaOL component association.
low latitudes. Moreover, phytoplankton blooms strongly reduce the transparency of the water column at certain periods. These effects are reflected by differences in depth zonation of various algal groups in polar and tropical re- gions. In addition, different strategies to overcome the dark winter season and different patterns of carbonate minerali- zation might by aspected along a latitudinal transect from the tropics to the poles.
Meridional oceanographic circulation and nutrient recycling." a promotor of the asymmetrical distribution of
warm- and cold-water carbonates
The modern global oceanographic circulation is charac- terized by a pronounced meriodional component that pro- motes intensive heat and moisture transfer between low and high latitudes. Due to plate tectonic rearrangements, this circulation mode was established during Miocene toPliocene time concurrent with stepwise increase of the Northern Hemisphere glaciations. Major elements of modern surface water circulation are the huge central oceanic gyres within the oceanic basins, the Antarctic circumpolar current sys- tem and the Equatorial counter currents. Due to the Coriolis- force, the central gyres rotate clockwise in the Northern Hemisphere and counterclockwise in the Southern Hemi- sphere. Strong western boundary currents (Gulf Stream, Brazil-, Agulhas-, Kuroshio- and East Australian-Currents) transport heat and moisture to the poles, while the weaker eastern boundary currents (Canaries-, Benguela-, West Australia-, California- and Peru-Currents) flow back to the Equator as cold currents. This oceanic pattern causes the pronounced asymmetry of the biogenic equator with a broadly developed tropical reef belt along the eastern mar-
gins and "non-tropical" carbonates along the western mar- gins of the oceanic basins (Fig. 1).
Another important feature is the intensity and availabil- ity of nutrient recycling between surface and deep waters. In the modern ocean this exchange is most efficient because of a positive feedback loop, i.e. the global conveyor belt of surface and deep waters (BRoEcr~a et al. 1990). The most efficient pump of the global conveyor belt has its startpoint in the Nordic Seas (SMETACT~ 1991). Highly saline and warm Atlantic waters imported by the Norwegian Current transfer heat and moisture to the cold air over the cool and par ty ice covered western and northern regions. As these supercooled surface waters become progressively denser, they sink down and fill up the deep sea basins. Such newly formed deep water is exported into the N-Atlantic and other parts of the worlds ocean. On their way, these deep waters warm and take up nutrients by progressive oxidation of organic matter at the sea floor and finally, these nutrient-enriched deep waters upwell close to the equator and thus are recycled to the surface. As a consequence, non-tropical carbonate grain associations are present in tropical shelf settings ( L o ~ et al. 1969, C ~ , r r E et al. 1988, Giscata~R et al. 1994). Nutrient availability has been elucidated to be a major control in the distribution and composition of modern and pastcarbonate-secrefing communities ( t ~ i o c x & ScrmaaZR 1986, WOOD I993). Therefore, zooxanthellate scleractinian corals thrive well in oligotrophic areas where external oceanographically driven nutrient input is modest but inter- nal recycling processes are highly developed (MuscAa-~,~ & PORTER 1977). Biomass turnover in these systems is main- tained predominantly by the herbivore or grazing food-chain (BARNES & MANN 1980). In the cold and eutrophicated seas of high latitudes the wave-exposed hardsubstrate coasts are

7 4
characterized by the phaeophytic belt (K~M~R 1980,LON~ 1990). MOSt impressive are the giant seaweeds which often form huge kelp-forests (Fig. 1). In kelp-forests, ecological structures comparable to reef frameworks are developed (GoLIKOV & SCARLATO 1973). The flexible but rigid thalli of kelp provide a variety of highly differentiated niches for carbonate-secreting organisms. Seaweeds display an impor- tant type of carbonate factory whose geological signals are poorly understood (FREIWALD 1993a). In contrast to coral reef systems, kelp-forests turn over about 90% of their algal biomass into the detritovore food-chain and only 10% is converted by the grazing food-chain (BARNES & MANN 1980, KOOP et al. 1982). Within the phaeophytic kelp belt coralline algal frameworks form islands along semi-sheltered hardrock settings including rhodolith pavements, e.g. the classical maerl-type carbonates from the Brittany coast (CABIOCH 1969, WEImMANN 1994) and western Ireland (B OSENCE 1976, 1980), but also fixed reefal frameworks in northern Norway (FRnWALD & I-tm,~CH 1994). Like in tropical coral reefs, the internal turnover of biomass within these non-tropical coralline algal frameworks mainly follows the grazing food- chain (FREtWALD 1993b). On a global scale, the extension and asymmetry of the "warm-water coral reef belt" at low latitudes and the"cold-water phaeophytic belt" of middle and high latitudes is a function of cold surface water regimes and is associated with, at least, seasonally eutrophic condi- tions (Fig. 1).
The bentho-pelagic coupling: the seasonal food transfer for deep carbonate secreting biota
Biogenic carbonate production in the aphotic zone on deeper shelves are predominantly maintained by BRYOMOL- type carbonate deposits and by azooxanthellate coral banks. These suspension feeders are strongly dependant from the seasonal food-supply maintained by the planktic rain, the bentho-pelagic coupling (GRAF 1989). Also the methano- trophic cycling triggered by cold-seeps is under discussion for the distribution of azooxanthellate reefs in cold seas (HoVLAND 1990). The bentho-pelagic coupling is considered to be the most important control for the development of carbonate buildups and deposits generated by heterotrophic organisms in aphotic zones (I-IzN~CH et al. 1992). In most cases, these deep water carbonates are situated well below the thermocline and thus no large variation in temperature occurs throughout the year. Generally, the thermocline is shallower in mid and high latitude shelves than in lower latitudes where the insolation penetrates into greater water depths. For example, Lophelia-reefs occur in shallower water depths on high latitude shelves (300 to 75 m water depth) than in low latitudes (more than 1000 m water depth) (see STANLEY & CAIRNS 1988).
Effects of small-scale high-frequency vs. large-scale long- term sea-level changes on non-tropical coastal platform
carbonates
The relative position and temporal variability of the sea- -level and its relation to shelf and coastal platform topogra-
phy is an essential factor for biogenic carbonate production (SCHLA~ER 1989). Despite of the different local evolution of the postglacial sea-level change (PmAZOLLI 1991), tWO trends become important when studying carbonate sediments in Brittany and in northern Norway. The relative sea-level curve in the English Channel area is characterized by a large and rapid eustatic transgression starting after the late glacial maximum at 20,000 to 18,000 years BP until 7,000 years ago, when the sea-level nearly reached its present position (LARsONNEUR et al. 1982). Contemporaneously with the flooding of the coastal platforms at the northern Brittany coast, carbonate production and sedimentation starts with bivalve communities (5500 years BP) and later, with coraUine algal deposits (2300 years BP) (WF.aRMANN 1994).
Areas which were covered by glaciers during the last glaciation are characterized by more complex sea-level changes. The postglacial eustatic rise is complicated by locally different isostatic rebound of the underlying crust, such as in northern Norway (MORNER 1979, MOLL~ 1989). High-frequent sea-level fluctuations strongly control the style of the slow-growing coralline algal frameworks in the Troms District, northern Norway (FRErWALD et al. 1991). Only during the intervals of sea-level standstills, coralline algae were able to form extended branching frameworks that generated pulses of huge piles of maerl-type deposits on the shore. In contrast to the Brittany coast, coralline algal deposition in northern Norway does not persist continouously from the Mid-Holocene but is characterized by relatively short-termed productional and depositional periods.
3 INTRODUCTION TO THE CASE STUDIES
The scientific projects which are shared within this group report depict different aspects of non-tropical carbon- ate production and deposition in its broadest sense. Some projects exist for some years already whereas others have been recenctly started with their scientific work. The case studies which are under focus are very different and offer new and interesting perspectives for future research with special regard to fossil analogues (Fig. 2).
Reefal coralline algal frameworks along the coast of the Troms District, northern Norway
(FRErWALD, HENRtCH, SCHAFER, SAMTLEaEN) Maerl-type frameworks in the Baie de Morlaix, northern
Brittany, France ~ ' ) ' V E ~ , ZANKL, HENI~CH)
In coastal current environments arranged along the north- ward pathway of the North Atlantic Current and its prolon- gation, the Norwegian Current, coralline algal reefs and maerl pavements are developed. Most of them occur in protected areas on the inner shelves. Pronounced variations in surface water temperature and nutrient cyclicity are caused by seasonal gradients in solar irradiation. Fine-tuned inter- actions of coralline algae and herbivores are discussed. The environmental controls favoring the development ofcoralline algal reef-forming processes are compared with the dynam- ics of the well-known maerl of the northern Brittany coast.

75
Cladocora-banks in the eastern Mediterranean Sea (K0m.M~e~)
Shallow water coral frameworks are provided by the Mediterranean Cladocora caespitosa, a presumed missing- link between tropical coral reefs and non-tropical single colonial scleractinians. Cladocora caespitosa is the main constructor of the modern coral banks in the Aegean Sea. The modern Cladocora-banks are placed in protected back water situations of the Aegean Sea. These banks consist of dense dendroidal and hemispheroidal colonies. The hydrodynamic regime at the sites of Cladocora-banks is characterized by moderate currents with some turbidity. Today, the Cladocora- banks suffer from eutrophication caused by tourism in the Aegean Sea, andby waste-water discharge (KOHLM_ANN 1985a, 1988).
Deep-water Lophelia-reefs in northern Norwegian fjords and on the mid-Norwegian shelf
(FRmwALO and ~ c n , in cooperation with RErIr~R and N ~ )
In summer and autumn 1994 two research cruises were conducted to map the geometry of deep shelf Lophelia- constructions. These aphotic coral reefs thrive-weU in waters with minor seasonal temperature and salinity fluctuations. However, a strong seasonal planktonic rain triggered by the collapsing phytoplankton bloom is proposed to be a major control. Another important aspect is the participitation of a diverse sponge community in deep-water reef ecology.
Arctic sponge-bryozoan buildups on the seamount Vesterisbank, central Greenland Sea
(HENRICH, S ~ R , FRErWALD, in cooperation with R~rmzR)
Rather stable physical environmental conditions, e.g. almost constant temperature and salinity profiles are charac- teristic for the sponge-bryozoan occurrences. The most prominent factor is the quality and quantity of food supply and its seasonal variation. The isolated intraplate seamount acts as an obstacle for the southward passing polar East Greenland Current and thus an intense oceanographic downwelling system, the TAYLOR-column, is established. Food particles and other detritus is concentrated in this downwelling gyre passing around the flanks of the seamount.
Carbonate production in kelp-forests (F~Iw ~D, H ~ )
The giant seaweeds are conf'med to cool and eutrophicated seas. Kelp ecosystems show high net-primary productivity comparable to tropical reef systems (Lt~n~o 1990) and are, as a whole, CO2-sinks. Kelp-forests play an important role in shaping the hydrodynamic regime along rocky coasts of high latitudes and thus, they provide habitats for a diverse often carbonate-secreting community. The geological sig- nals of phaeophytic belts are not yet pointed out. It is envisaged, that phaeophytic belts can be used as tools for the reconstruction of the trophic regime in past coastal environ- ments.

76
BRroMoL-carbonate sediments in the English Channel and in northern Norway ( S ~ , BADZR)
Since the Permian, open-shelf bryozoan-rich limestones are often developed in cold-water and/or in eutrophicated tropical areas. The project addresses to the dynamics of modern autochthonous production sites, the bryozoan-thick- ets, and their taphonomic fate. An open-shelf setting in northern Norway will be compared with the English Chan- nel area.
Arctic balanid-bivalve-bryozoan sediments from the Spitsbergen Bank, Barents Sea (HEYRICH, FRErWALD, SCH~R)
A specific oceanographic configuration is found on the western Barents Sea. The flanks of the Spitsbergen Bank are stationary high production zones resulting in intense carbon- ate production.
Additional carbonate deposition occurred under sea- sonal sea-ice coverage since 8700 years B.P.. Atlantic wa- ters fill the troughs north and south of the Spitsbergen Bank, whereas Polar waters form a stable gyre on the shallow parts of the bank.
4.1 Coralline algal reefs and maerl pavements
In boreal to arctic climatic settings coraUine algae belong to the most prominent framework constructors in shallow waters. Well-known are the maerl-type carbonates which are formed by free-lying branched rhodoliths (BosENc~ 1983). Large occurrences have been described from north- em Spain (ADF.y & McKma~ 1970), the Brittany coast (LEMoINE 1910, CABIOCH 1969), southern England (FARmlAM & JEI'HSON 1977), western Ireland (BOSENC2 1976, 1980), the northwestern Scottish coast (FARROW et al. 1984, ScovF~ 1988) and the northern Norwegian coast offTroms (FREIWALD et ai. 1991). ~ (1883) reported extended rhodolith pavements from the Mussel Bay, northern Svalbard close to 80~ In contrast to these unattached coralline algal flame- works, fixed algal reefs were found on coastal platforms at the northern Norwegian coast, at 70~ (F~,v.rWALD & HEm~IcrI 1994). Free-lying rhodolith banks and algal reefs represent principially different types of non-tropical shallow water coralline algal frameworks. The maerl-type carbonates in the Baie de Morlaix, northern Brittany, and the algal reef from the northern Norwegian coast provide a unique possi- bility to compare their structure, internal sedimentary and biological dynamics, and their adaptation to the exlreme seasonality.
Climatic change: control of carbonate platform develop- ment (BRroMOr- tO CHt_OROZOaN-limestones), Leg 133,
Queensland Plateau (BRACHERT • BETZLER)
Since the end of the Eocene, the Australian Plate drifted from a high latitude to a tropical clmatic zone. The latitudinal drift is documented in various types of carbonate platform sedimentation, i.e. the composition of neritic skeletal grain associations and in the style of diagenetic behaviour. The non-tropical sedimentary units on the Queensland Plateau are comparable to modern non-tropical BRVOMOL-Sediments.
The locations of the modem case studies from non- tropical environments are shown in Fig. 2.
4 NON-TROPICAL REEFS AND BUILDUPS
The term 'reel" is used here in an open definition, i.e. skeletal frameworks with a pronounced elevation above the surrounding seabottom which consequently influences the hydrodynamics and sedimentary facies. Furthermore, these carbonate frameworks are colonized by distinct organism communities and thus form ecologically complex biota. In detail, the term 'reef' or 'reefal' construction is applied to the in situ (fixed) coralline algal buildups and not to the rhodolith pavements.
These coralline algal reefs differ from tropical reefs by the lack of strong wave-resistance and by the lack of early diagenetic lithification. In the deeper water settings, the term 'reef' is used for the several kilometerlong and up to 50 m high coral frameworks constructed by the azooxanthellate scleractinia Lophelia pertusa.
Environmental setting
The northern Brittany coast, part of the Western Ap- proaches of the English Channel, is formed as a rocky shore with some large embayments (Fig. 3). A branch of the North Atlantic Current cause the support of temperate water. The temperature range from 8-9~ in winter to 16~ in summer. The semidiurnal tide is characterized by strong current regimes (>4 kt) with a high tidal gauge (9-15 m), responsible for a strong mixing of the water column on the inner shelf with a constant salinity of 35 %0. The studied area of the Baie de Morlaix is a coastal platform of 26 m maximal water depth, differentiated by granitic islets in a complex morphol- ogy. The rocky coastline is locally covered by periglacial sediments, now reworked at cliffs, which provide the main source for lithoclastics in the superficial sediments in the bay. The influence of estuarine rivers is low in case of water- -chemistry, but important in case of terrigenous input.
The coast in the Troms District, northern Norway, is fringed by shallow coastal platforms which are structured by islet archipelagoes. The coastal platform studied is the Storvoll Bay at the southern tip of RebbenesOy (Fig. 4). Landward, this plateau is surrounded by crystalline rocks of the Caledonian mountain range and seawardly, the plateau is bordered by the slope of the deep S andoyfjord-Trough in 35 m water depth. The western plateau-flank is protected by shoals against the ocean-swell. Most parts of the Storvoll- Plateau are shallower than 10 m water depth. The oceano- graphic environment is dominated by theNorwegian Coastal Current, with salinities less than 34 ~ . Due to the spring melt-water discharge, the salinity can decrease to 26 ~ (S~Ia.EN 1950). The water temperature ranges from 14~ in summer to 3 - 4~ in winter. The semidiurnal tidal gauge can

77
Fig. 3. Production zones of branching coralline algae and related rhodolith pavements in the Bale de Morlaix near Roscoff, northern Brittany, France.
reach 3 m during extreme spring tides (Ea.ERTSEN et al. 1981). The tidal currents within the islet archipelagoes can reach several knots.
Geometry and sedimentary structure of coralline algal frameworks
The coralline algal frameworks in the Bale de Morlaix and in northern Norway are constructed by branching growth- forms predominantly. The branched growth habit is a major prerequisite to generate algal gravel, or maerl-type carbon- ate deposits.
In the Baie de Morlalx rhodolith occurrences are situated in the shallow subtidal zone in 4 - 8 m below low spring tide level adjacent to rocky shoals which lie in the semi-protected area of the embayment (Fig. 3). The main rhodolith con-
structor is Lithothamnion corallioides but Phymatolithon calcareum also occurs (CAaIOCH 1969). Therhodolith pave- ments show a clear proximality trend starting from the initial sites of coraUine algal branch production, which are the coralline encrusted rocky shoals and boulder grounds (Fig. 5). The overall geometry of the maerl-type carbonate depos- its is controlled by topographic features of the seabottom. Proximal to the rocky shoals, a maerl gravel facies is developed that passes distally into a pure maerl facies. The maerl facies is characterized by interfmgered open-branched rhodoliths forming a thick rhodolith pavement. The core of the rhodoliths is formed by the up to 3mm long branches which are derived from the initial sites of branch production, the rocky shoals. The pure maerl facies transites distally into a pronounced rippled maerl facies. The most distal subfacies is an algal-debris deposit that contains fragmentedrhodoliths

78
Fig. 4. Geographic map of the coralline algal reef on the Storvoll Plateau, Troms District, northern Norway. The inlet map shows the geomeu'y of the in place reefal framework and associated rhodolith belts.
due to intense redeposition. Forced by the local hydrographic regime, overall redeposition of rhodoliths is directed off- shore. Therefore, no maerl-type deposits are found on the intertidal fiats and beaches. The tidal flats are formed by a t'me-grained lithoclastic sand with minor carbonate admix- tures of molluscs and foraminifers predominantly.
The coralline algal reef and its associated facies belts are oriented parallel to the coastline on the Storvoll-Plateau (Fig. 4). The FLxed reefal framework covers approximately 125,000 m 2 and is growing in 15 - 6 m water depth. The framework is constructed by a succession of three coralline species. The dead base consists of an irregularily branched Lithothamnion sp. Secondary fusion of the branches is common and thus enhances the internal stability of the framework. The basal parts are radiocarbon-dated to 180 + 50 years BP. After reaching an average height of 20 to 30 cm above the surrounding seabottom, a succession of new species took place. The modern reef-builders are the entirely encrusting Phymatolithon sp. and the branching Lithotham-
nion cf. glaciale. Both species show a pronounced competi- tion for space on the top of the framework. The branched aggregates of Lithothamnion cf. glaciale are the most promi- nent source for rhodolith production after getting detached from the autochthonous reefal framework (FREtWALD in press). Therefore, the algal reef can be regarded as an rhodolith-factory that generates the thin off-shore and the thick on-shore directed rhodolith pavements (Fig. 5). The driving force behind this facies zonation are storm events which rarely affect the semi-protected coastal plateaus. Consequently, both rhodolith and algal gravel subfacies are related to tempestitic events. The on-shore directed storm wave-drag generates multilayered rhodolith pavements which distally fade-out into rhodolith megabars. Multiple fragmentation of the branched rhodoliths result in pure algal gravel bottoms. On the beach, these algal gravels can be piled up into storm ridges. The thin off-shore directed rhodolith pavements result from compensative bottom-near counter currents during storm events.

79
Fig. 5. Schematic sections through maerl-type coralline algal frameworks and their reworked deposits in the Bale de Morlalx (above) and the algal reef with its rhodolith belts from northern Norway (below). Main directions of redeposition and la'a.rtsport are indicated by arrows. In Brittany, the whole production of maerl-type carbonates and their sedimentary dynamics take place well within the subtidal zone. In northern Norway also the intertidal zone is involved in redepositional processes.
Coralline algal - herbivore interaction
One of the most surprising observation on living coralline algal frameworks is the sparseness of sessile epiphytes. In contrast, dead coralline thalli are generally densely colo- nized by serpulids, spirorbids, bryozoans, balanids, boring bivalves and f'damentous algae. This apparent difference in the intensity of colonization by secondary epiphytes be- tween living and dead corallines slrongly suggest biological controls (FRmwALD 1993b).
By far most of the organisms living on the corallines belong to the herbivore trophic group. Most abundant are grazing gastropods, especially limpets, gibbulid and rissoid forms. Chitonids are also commonly present in the maerl bottoms of the Brittany coast and on the Storvoll-algal reef. Furthermore, ostracodes, amphipodes and porcellanid decapodes seem to play an important, but not yet understood role within the trophic cycling of coraUine algal biotopes. The most conspicious difference to the Brittany maerl grounds is the presence of the green sea-urchin Strongylocentrotus droebachiensis in the northern Norwegian coraUine algal biotopes. According to underwater-video mapping, on the reef framework almost all specimens were ontogenetically juveniles, whereas the adult ones live predominantly outside of the coralline fxameworks. Coralline algae play a key role to induce metamorphosis of larvae of distinct herbivores ~ c E & SCn~mL~G 1990). This stimulation of metamor-
phosis is triggered either by host-specific bacteria that are involved in cell-aging processes on distal epithallial cells (LE~s etal. 1985, JomqsoN et al. 1991 a, b), or, by neuroactive inducers (MoRsE 1991). This evident coralline algal- herbiv- ore interaction is a well-known phenomenon that can be observed world-wide. Severe ecological overgrazing im- pacts on lxopical reef systems, the Acanthaster-outbursts in the Great Barrier Reef (VAN DER LAAN & HOGEWEG 1992), and on boreal kelp-forests, forced by outbreaks of the green sea urchin (HAGEN 1983), are strongly related to this evolution- ary fine-tuned interaction between distinct herbivores and coraUine algae. Apart from these extreme biological distur- bances, the large populations of gazers on living coralline algae are necessary to prevent fouling by rapid-growing sessile invertebrates and spores of filamentous algae. But they are also needed to remove thick diatom films (FREtWALD 1993b). Although the coralline algal biotopes in Brittany and in northern Norway thrive well in areas of high grazing pressure, the destructive impact on the corallines is low because of their ability of rapid regeneration of abraded cells (STENECK 1985). Examinations of egested fecal pellets col- lected from several herbivore gastropods and chitons show abraded parts of the uncalcified epithallus mixed with micrite- sized chips of the calcified cell wall (P1. 17/1-2). These pellets are densely coated by bacterial films which decom- pose the organic remains. After decomposition, the fecal pellets disintegrate and form a major source for carbonate

80
Fig. 6. Yearly budgets of carbonate production by branching coralline algal occurrences along a latitudinal gradient, from 480N to 70~ respectively. Unattached rhodoliths show lower production rates than branched in place frameworks. Although there are only few data available, the trend is that carbonate production by coralline algae within the Arctic Circle can exceed annual production rates calculated from boreal coraUines.
mud production. Summing up, the coralline algal - herbivore interaction favors the highly competitive success of slow- growing corallines against the rapid-growing phaeophytes and can be regarded as an example of self-regulation within complex biological systems.
Coralline algal growth rhythmicity and carbonate production
Coralline algal frameworks in Brittany and in northern Norway provide the unique opportunity to study their growth rhythmicity in response to seasonal environmental changes along a latitudinal transect (P1. 17/3-7). Regions north of the Arctic Circle are characterized by extreme seasonality in solar irradiance. The seasonality is forced by the several months lasting period of winter darkness and by the day- round illumination during the polar summer period. The numerous occurrences of extended coralline algal frame- works even within the Arctic Circle seem to be extraordi- nary. The long-living corallines must have evolved physi- ological adaptations to cope with the extreme seasonally environmental fluctuations, such as light, temperature and nutrient content.
Under the SEM, growth rhythmicity in coralline algal skeletons is expressed in bright-dark banding patterns which are considered to be primarily annual (AoEv & MCKaBBtN 1970, BOSENCE 1980). The bright banding represents sum- mer growth and is characterized by lower dense cell-wall mineralization. The dark winter growth shows high density bands (P1.17/3-7). The transition from a dark winter band to a bright summer band is sharper developed than vice versa. In case of Lithothamnion corallioides, the prominent spe- cies in the maerl banks of the Baie de Morlaix, an annual growth increment of 0.3 - 0.5 mm can be observed (WErmMAr~ 1994), with 0.09 - 0.12 mm growth increment in winter and
0.25 - 0.38 mm growth increment during the summer reslx~- tively (PI. 17/6-7). To convert the growth rates to carbonate production budgets, the weight of the branches and the standing stock was measured. In the maerl banks of the Baie de Morlaix the yearly rate of carbonate production varies between 100 - 500 g m -z a -1 ( W ~ 1994). This calcu- lation from the Brittany coast lies in the same production range of maerl banks from the Mannin Bay, western Ireland (BosENCE 1980). Only theproduction rates of Lithothamnion corallioides from the Bay of Brest are comparatively high (Pox~ et al. 1990) (Fig. 6).
The coralline algal growth rates within the algal reef of the northern Norwegian Storvoll-Plateau at 70~ were obtained from branching Lithothamnion cf. glaciale. SEM- studies on sectioned branches also show pronounced bright- dark banding patterns which are considered to represent annual growth. However, each banding is formed by subor- dinated growth rhythmites (PI. 17/3-5). The bright summer increments consist of 5 - 8 subordinated bands, each meas- uring 0.1 - 0.15 mm. The dark winter bands are formed by 3 - 4 subordinated rhythmites, each measuring 0.05 - 0.08 mm. Summing up, an annual growth rate between 1 - 1.5 mm can be calc dated for the reef-building Lithothamnion cf. gla ciale in 7 m water depth. Converted to rates of carbonate produc- tion, 900 - 1400 g CaCO3 m -2 a 1 are produced (see FREIWAtD & HErCR~CU 1994). Hence, high-boreal to subarctic coralline algae secrete more than twice the carbonate as their boreal counterparls (Fig. 6). However, in 18 m water depth, the carbon- ate-production rate is much lower with400-600gm% q (Fig. 7). The extreme difference in growth rates between the Brittany and the northern Norwegian frameworks must be explained with the internal ability of the two coralline species to cope with their environmental constraints. There- fore, these surprising results, that more carbonate is pro- duced within the Arctic Circle than below does not follow a

81
latitudinal trend but strongly challenges the conventional view of the insignificance of high latitude carbonates.
The physiological swing - considerations on nutrient and carbon utilization in high latitude coralline algae
Coralline algae are long-living but slow-growing algae. In high latitudes, corallines have to withstand extreme environmental variations. The poleward stronger gradient in solar irradiance directly influences the shift of low- and high pressure systems initiating the winter and spring storm periods in the Northern Hemisphere, and, accellerating the resuspension of nutrients in the water column in these periods. Most important are nitrogen and orthophosphate compounds. Nutrient uptake during the dark winter and spring period secures the energy demands of corallines for growth, repro- duction and other energy-consuming processes. This sea- sonal eutrophication in high latitude seas is the driving force for phytoplankton blooms which are extremely dependant
on water temperature, light and stability of water stratifica- tion (S~aETACeK 1991). During the summer period, nutrients are completely utilized. Therefore, coralline algae have to overcome the period of nutrient depletion in the summer by utilization of stored nutrient compounds. According to [~3ESO-mL (1992), crystalline protein inclusions, stored within the epithallial cell compounds mightprovide a buffer against the environmental limiting factor of seasonal nitrogen de- pletion. On the other hand, the boreal and arctic coralline algae need carbon resources during the period of low insolation or complete darkness. An undetermined quantity of carbon is stored in form of starch grains (GmAtJD & CAmOCn 1983, Sr~_Aa'n et al. 1991). The starch grains derive from the photosynthetic conversion of solar energy into glucose that is deposited as an amylopectine compound, the starch grains, within the central perithallial cell compart- ments. During the dark season, when photosynthesis stops, the stored carbohydrates can be utilized. This kind of'physi- ological swing' for high latitude coralline algae has been

82
described for the boreo-arctic kelpLaminaria hyperborea as well (KRE~am 1980) and is suggested to be one of the major adaptations for longliving benthic macroalgae to survive in seasonally extreme fluctuating environments.
Fossilization potential, carbonate dissolution, bioerosion and cementation
The carbonate saturation regime of the ambient seawater is a discrirninant factor on the preservation potential for skeletal carbonates. Many investigators have shown that subtropical-tropical seawaters are supersaturated with re- spect to CaCO3 (e.g. WAa-re.x~ram~o 1933). Against the con- ventional view, the same holds for many high latitude cold water shelves, however at a lower saturation level and temperature regime that is not high enough to induce anorganic cementation (CHAvE 1967). The seawater at the comlline algal reef in northern Norway shows a CaCO3 supersaturation of 150 to 250 % (these estimates based on pH, temperature, salinity and CO2-content). This means, that the carbonate sediments in northern Norway are not dissolved postmortally. Also SEM-studies on the ultrastructure of carbonate grain associations show no evi- dence for chemical dissolution.
Evidences of bioerosive carbonate degradation in the inner coastal subtidal zone with the coralline algal facies are documented by macro- and microendolithic borings created by clionid sponges, sipunculids, phoronids, algae, and fungi (Bgo~a.mc & ~ 1981, Sc~IMItrr & FRmWALD 1993). Samples taken from shallow high energy environments contained tests of Cibicides lobatulus infested by countless epilithic and chasmolithic rod-shaped bacteria arranged in chains or in clusters (FI~aWALD 1995). The bacteria show a characteristic preference in colonizing depressions and crev- ices or any other kind of semi-protected area with a low relief. Predominantly, the lateral chamber walls of the flat- tened spiral side of Cibicides lobatulus have been corroded, leaving the septae behind. The rod-shaped bacteria are attracted by the skeletal binding organic matrix of the foraminferal test which is present a) as organic linings separating different layers of the test and b) as intrasutural matrices enclosing each calcitic microcrystallite of the granulitic microstructure 03m.r~MO 1974).
In the coralline algal reef, carbonate cements are re- stricted to internal primary openings of the corallines. The cements are characteristic intraskeletal high-magnesium calcite micro-druses as has been describedby ALEXAr~SSON (1974). The formation of these micro-druses is related to physiological processes, such as the utilization of starch grains (FRmWALD 1993a) by the living algae. These cements are often found in thin-sections at the boundary of living to dead cell-compartments.
Coralline algal frameworks and their response to Holocene sea-level changes
Sea-level fluctuations during the past 15,000 years re- veal a pronounced eustatic component due to intense melt- ing of continental glaciers in the Northern Hemisphere.
However, postglacial isostatic compensation has strongly influenced both areas of investigation, the Brittany coast and northern Norway. These aspects are very important when comparing the onset of coralline algal production during the last glacial/interglacial transition at the northern Brittany coast and in northern Norway.
The relative sea-level fluctuations in northern Norway were strongly controlled by glacio-isostatic uplift of the Fennoscandian Craton (MORNER 1979). Therefore, an over- all regressive regime is typical for the Norwegian coasts (Fig. 7). The oldest coralline algal deposits yield an age of 6400 years BP. In detail, the geological record of coralline algal frameworks and their reworked deposits is not continu- ous. Moreover, as can be judged from numerous sections at various coastal outcrops in the Troms District, the distinct distribution of rhodolith banks seem to represent repeated periods of framework production during the Holocene. The radiocarbon-datings taken from these rhodolith banks and fixed frameworks show specific age-clusters around 5500 years BP and 4500 years BP (FRErWALD et al. 1991,FREnVALD 1993a). These age-clusters coincide with sea-level ingres- sions along the coast of the Troms District (MCLLER 1989). During these ingressions, the regressive element caused by the crustal uplift was compensated for some hundred years each. These repeated intervals of sea-level standstills are considered to provide favorable ecological conditions for framework-constructing coralline algae.
The Baie de Morlalx as a part of the northern Brittany coast is the result of a constant drowning that starts with the melting of the Pleistocene icedomes (Fig. 7). The Brittany was not glaciated but was situated in a periglacial position. The sea-level curve shows a rapid single-phased transgres- sion (LARsONNEtn~ et al. 1982). The oldest radiocarbon dated maerl yields an age of 2340 + 49 years BP (WErmMANN 1994). However, the data base is not sufficient to draw general conclusions. According to LARSSOrZEUR et al. (1982), the modern coastal platforms around the Brittany coast were drowned during the last 6000 years. In the Baie de Morlaix, the first radiocarbon-dated marine bivalves are 5570!-_70 years old. It can be assumed that at this time no large wave- sheltered areas were present favoring the establishment of framework-constructing coralline algae.
The significance of maerl-type carbonates as climatic indicators
The first appearance of modem coralline algae in the geological record dates back to the Jurassic (FAGERSTROM 1987). However, the oldest unequivocally identified coralline algae are described from the B arremian-Aptian (Mous SAVIAN 1989). During the Cretaceous, coralline algae rapidly be- came very important members of the binder-guild on tropi- cal platform carbonates (MoussAv~'~ 1989). Branching growth-forms belonging to the constructor-guild became abundant in the Paleogene (STENECr 1985). From that time on, many examples of at least warm-temperate maerl-type carbonates are known from both hemispheres.
Today, the largest occurrences of maerl-type carbonates are found in the shallow subtidal zone on protected coastal

83
platforms of the high latitudes. Towards the lower latitudes, such as the Mediterranean Sea, the same maerl-producing coralline species (Lithothamnion corallioides and P hymato- lithon calcareum) thrive well in greater water depths than their boreal and subarctic counterparts (Fig. 8). The equatorward submergence of maerl-type carbonates contin- ues, if present, to the tropic climatic zone. For example, maerl is recorded at about 100m water depth on the outer shelf off Mauretania (M-J ~v.~ 1977) and the tropical shelf offBrazil ( C ~ A Y r E et al. 1988). A very similar situation is evidenced by CA~-N~rrE et al. (1988) for the Lower Miocene Sant Elm Calcarenite Formation in Mallorca, Medi- terranean Sea. On the former deep outer shelf in place bioherms of branching coralline algal frameworks with adjacent rhodolith belts were replaced towards the shallow coastal environments by coral reefs (RoDPaou~-t~t~, 1984). With regard to skeletal composition, the ancient subtropical- tropical deeper water maerl-type carbonates can be con fused with those from shallow temperate zones thus leading to paleolatitude and -climate misinterpretations.
4.2 Coral frameworks in non-tropical seas
The coral buildups in the warm-temperated eastern Medi- terranean Sea constructed by the zooxanthellate scleractinia Cladocora caespitosa may be considered as transitional form to the true coral reefs of the tropics ( K ~ , n ~ et al. 199 I,KotJxOtn~s & K ~ 1991 ). According to LABO~ (1987), Cladocora caespitosa is an endemic Mediterranean species tolerating relatively low winter temperatures. A near relative is Cladocora arbuscula, a Caribbean endemic living in comparable environments.
In contrast, the largest coral buildups in non-tropical seas are found in cold and aphotic waters. In the Nordic Seas these buildups are constructed by the azooxanthellate coral Lophetia pertusa. Coral frameworks constructed by the azooxanthellate Lophelia pertusa (PI. 18/3) are introduced to the literature by a note of the bishop Gt~,~RUS in 1763 to Ln, e~ (cited in DONS 1944). Today, many Lophelia occur- rences are known from the N-Atlantic and from the Norwe- gian Sea (DoNs 1944, LE DAJ~oIs 1948, WmSON 1979a, b, Scon~a~ & Bowra 1988; see STnNt~V & C~RNs 1988 for review). The known temperature range tolerated byLophelia pertusa is between 4~ and 12~ (Tmcn~T 1958).
The Mediterranean Cladocora-reefal frameworks
From ecophysiological conditions the shallow, warm and transparent tropical seas are predestinated for coral reef growth. It is intelligible that the oligotrophic Eastern Medi- terranean Sea with its temperature of 22 ~ to 25~ in summer and about 12~ during the winter provides the possibility for one species of hermatypic stone corals to survive in a warm- temperated sea, and to construct coral banks. A missing-link between true coral reefs of the subtropical zone and to single scleractinia colonies in shallow warm-temperate seas is suggested for the zooxanthellate Cladocora caespitosa. According to A u k (1959), Cladocora caespitosa shows high growth-form variabilities which can be attributed to
Fig. 8. Poleward shallowing-up of branching coralline algal biotopes as a function of decreased penetration of fight into the water column in high latitudes. The dominant maerl-producing coralline algal species are indicated with their geographic distribution along the northwest European shelves and the western Mediterranean Sea.
specific environmental conditions: (1) Under the influence of high-energetic hydrodynamic
regimes with strong wave drag and surge, the shape of the Cladocora-colonies grows flat with corallites between 2 and 8 cm high. Branching growth-forms are rarely devel- oped under these conditions.
(2) In habitats of less hydrodynamic energy, but with con- tinuous, moderate currents, thick dendroid corallites up to more than 20 cm high are the predominant growth-form of Cladocora caespitosa.
(3) In the eastern Mediterranean Sea, in calm waters with dim illumination, normally deeper than 35 m, colonies of Cladocora caespitosa build loosely ramified and erect colonies.
Unlike most other scleractinian corals existing in non- tropical shallow water environments, the dense dendroidal and hemispheroidal growth-form of Cladocora caespitosa (PI. 18/1-2) possesses the ability to construct reefal frame- works in the sublittoral zone of the Aegean Sea in a small inlet between the SithoniaPeninsula and the Isle of Dhiaporos (40 ~ 12.5N; 23~ More Cladocora-banks, not yet stud- ied in detail, exist northerly of Evia Island (LABO~L 1961, K ~ 1991), and perhaps off the Tunisian coast (PoizAv 1970). Recently, MoPa~I et al. (1994) discovered small Clado- cora-banks in the Ligurian Sea in 26 to 29 m water depth.
The Cladocora-banks off Sithonia exist in 10 to 20 m water depths, in waters with 35 %o salinity, and with 20 to 24~ water temperature in summer and about 12~ during the winter period. The banks elevate up to 3.5 m above the

84
surrounding seabottom. The length of single bank construc- tions is about 5 m, their width about 4 m. Up to 240 cm deep core samples collected by SCUBA-diving from the interior of the buildup give evidence of a framework primarily constructed by Cladocora-skeletons. Serpulids and bivalves also participate as secondary constructors. The interspaces of the framework are inf'dled by loose, coarse calcareous debris and mud.
Aside of the importance of Cladocora caespitosa as a constructor of reefal buildups, these banks provide numer- ous habitats for many benthic organisms. An analysis of the associated macrofauna reveals 236 species. In detail, 36 gastropoda, 16 bivalvia, 49 errante polychaets, #1 sessile polychaets, 18 amphipods, 22 decapods, and 54 species belonging to other groups such as porifera, cnidaria, turbellaria, nemertini, tunicata and pisces have been identi- fied. Altogether, the associated community producecs a dry substance of 9.7g/m 3 within a Cladocora-colony. Propor- tional to the body-weight and the number of individuals, sipunculidagets 26.8%, crnstacea 24.7%,polychaeta 17.4%, mollusca 11% and other groups 20.1% of the masses of inhabitants (Kotmotn~s & Kt~m.~aq~ 1991).
All in all, the growth-form of the thick-branched, hemispheroidal colonies of Cladocora caespitosa provide the base for a high productive habitat that is colonized by a high diverse macrofauna community. Moreover, this spe- cific coral growth-form is confined to distinct ecological controls, such as a stable hardsubstrate in wave-protected but current-rich environment.
Deep water Lophelia-reefs from the Faroe Islands and from the Norwegian shelf
Principially two types of Lophelia-occurrences can be distinguished: open shelf and fjord settings. Widely distrib- utedLophelia-banks have been recently investigated around the Faroe Island shelf banks (FREDEPdXSEN et al. 1992, JENSEN & FREDERaXS~ 1992). These coral banks exist in a depth interval ranging from 200 m down to 700 m water depth. In detail, their frameworks often show a lined distributional pattern within the Faroe waters. According to FR~OERIXSEN et al. (1992), this lined growth habit is forced by areas where the bottom slope is critical to internal waves of semidiurnal frequency. Internal waves derive from advection of strati- fied water across bottom contours by barotrophic tides. Within these advective mixing zones, flocculent particles get enriched which lead to comparatively narrow bands of nutrient- and food-rich water layers spreading horizontally. The authors suggest that the shelf break and slope areas which are effected by internal waves are preferred sites for suspension-feeding communities. The fauna that lives asso- ciated to Lophelia-frameworks consists predominantly of bivalves, brachiopeds, echinoderms, bryozoans and porifera. In total, 256 species associated have been recognized (JENsEN & FREDERn~S~ 1992).
Lophelia-reefs occur in mid-shelf position offTrondheim, Norway, in 250 to 320 m water depth. The water temperature is around 6~ The area investigated is located approxi- mately 80 km off-shore, where the Fr0yabank transites into
the Haltenbank area (Fig. 9). A reconnaissance survey concentrates on an isolated NE plunging spur at 64~ and 08~ (Fig. 9) that is underlain by Paleocene sandstones (Ro~cOF~OZN et al. 1988). The long axis of this spur distally fades out into a more than 500m deep basin. The spur is bordered by channel-like morphological depressions filled with Late Weichselian morainic material (Buc, oE 1980).
The geometry of the reef is largely confined by the topography of the spur and its adjacent channel areas. Two major sites of deep-water reef growth can be defined. The most impressive one forms a huge wall of coral framework directly growing along the SE crest-margin of the spur (Fig. 9). This reef-wall is more than 5 kin in length. The frame- work itself is 13 m to 30 m high and 220 m to 400 m wide at the base. A second area of deep-water reef formation is detectable on the central ridge-section of the spur. These frameworks never reach the dimension of the main reef- chain. The framework is constructed predominantly by Lophelia pertusa, but also Madrepora oculata and the octocorallia Primnoa resedaeformis were sampled.
Many of the mid-shelf Lophelia-reefs in Norway exist near active cold seep areas ( H o ~ 1990). A methano- trophic cycling could be substantiated so far only by the detection of negative 813C-isotopic values in the skeleton, with -1.0 to -8.6 PDB respectively (MIK~T sm'q et al. 1982). However, evidence for a close interaction between methane exudiating cold seeps and the gregariousness of deep water corals could not be substantiated so far.
Another Lophelia-reef was found in the Stjernsund, near Alta, northern Norway, at a position noted by DoNs (1935). The reef exist along the sea- and landward flanks of a sill separating the Stjernsund in two troughs in 250 to 215 m water depth. The framework is a dense and coherent mesh- work of several meters in height. The framework surface is undulate because of active growing fronts of Lophelia which exist close to dead parts of the framework. In situ collapsing of reef sections due to intense clionid bioerosion below the living surface is often observed. Downslope, coarse coral debris form several tens of centimeter thick lobes which fade-out abrupt distally. Isolated and com- pletely spherical balls (up to 0.8 m in diameter) consisting of living Lophelia lie free in the debris areas. The top of the morainic sill shows coarse sands with patches of pebbles and boulders in between. The benthic community switched rapidly over to a sponge dominated community. The colour- ful large sponges partly form dense mounds often associated with fishes. Lophelia grows as a thin (centimeter-scale) coverage on pebbles within the sponge-mound area, strongly resembling to a pioneer-stage. It is conspicious, that the sponges drastically diminish in diversity and size when approaching to the Lophelia-reef.
Thecosmilia and the Middle Triassic onset
Scleractinian corals first appeared in shallow water set- tings of the Tethyan region in the Middle Triassic but their participation in framework construction was very limited until the Rhaetian stage (FLt~GEL 1981). In terms of distribu- tion, the Middle Triassic scleractinians were rather wide-

85
spread and occupied a wide range of latitudes and environ- ments but were best developed in warm, shallow water (STArmEV 1981). In contrast to modem hermatypic scler- actinians, the Middle Triassic corals preferred sheltered environments which were not exposed to wave-surge. Re- markable coral frameworks were formed by Thecosmilia- biostromes in basinal settings of western North America (STANLEY 1979), or marginal to carbonate platforms in the Alps (ZANKL 1971, HENmCH 1983). The growth habit of Thecosmilia is compared with the modem azooxanthellate Lophelia with its characteristic branching growth (S-rANLEY & CAIRNs 1988). At the end of the Late Triassic, a sudden increase of scleractinian corals as prominent reef-builders is detectable from the fossil record. This sudden evolutionary success in reef-architecture is presumably linked with the advent of symbiotic association with algae (STANLEY 1981 ).
Today, large coral frameworks constructed by azooxanthellate scleractinians are restricted to the deep aphotic zone (TEIcrmRT 1958, STANLL~ & CAmNS 1988).
The Danian deep water coral mounds
As ha s been suggested by TEICHERT (1958) and B ZRNECKER & WEmUCH (1990), a fossil counterpart to modem deep water azooxantheUate coral frameworks is the Danian (Paleo- cene) Coral Limestone of Fakse, Denmark and of Limhamn, Sweden. The coral mounds show asymmetrical shapes with lengths of 200 m, heigths of 30 m and widths of 80 m. The assymmetry is caused by unidirectional bottom-near cur- rents. Framebuilding was maintained entirely by azooxan- thellate corals (Dendrophyllia, Faksephyllia, Oculina), stylasterine hydrocorals and octocorals. The major criteria

86
for the paleobathymetrical interpretation of these deep water coral mounds are the lack of calcareous and endolithic algae, the admixture of true pelagic organisms to the micrites (coccolithophorids, globigerinid foraminifers) and the ad- aptation of all frarnebuilders to heterotrophic nourishment. These mounds existed on a high paleolatitude in a current- -exposed position situated in a narrow seaway bordered by the Baltic Shield landmass and the RingkCbing-Fyn-High.
4.3 Arctic sponge-bryozoan mounds on the Vesterisbank seamount
Modern and Holocene sponge mounds with various amounts of bryozoans have been described from the western Canadian shelf (CorCwAY et al. 1991), the Arctic Canadian shelf (VAN WAC, O~R et al. 1989) and from the Antarctic shelf (DAYTON et al. 1974). Another example is located on the isolated intraplate seamount Vesterisbanken, central Greenland Sea ~ c u et al. 1992). Physical parameters of siliceous carbonate buildups and biogenic mats on this particular seamount represent the cold end member of mod- em carbonate deposition. A nearly year-round sea ice cover, which retreats only during two months, and very low tem- peratages (-1 to 0~ and salinities of 34.5 ~ are recorded at nearly constant values over the entire water column, and thus seem to have no influence on the pronounced depth- related biogenic zonation observed on the seamount flanks. All biogenic constructions are formed in aphofic depths. The most prominent environmental control is food supply. Dur- ing the short term retreat of the ice margin, melting sea ice sets free nutrients which induce phytoplankton ice edge blooms. In addition, fine grained sediments and high quan- tifies of suspended and particulate organic matter are re- leased. Due to a downwelling current regime over the seamount, a rapid and effective transfer of food is supplied to the benthic ecosystems reflecting an intensive pelagic/ benthic coupling.
Despite these polar environmental conditions, a rich and diverse benthic community dominated by siliceous sponges, bryozoans and serpulids have colonized the various habitats of the seamount down to more than 2000 m water depth. These primary constructors are associated by molluscs, echinoderms, abundant planktic and benthic foraminifers as well as crinoids in the deepest habitats.
Depending on the quality and quantity of food as well as on different substrates due to variable slope inclinations and roughness of the volcanic foundation, a clear depth zonation of biogenic structures and facies belts is established. The summ it region of the seamount is almost completely covered by biogenic structures. Here, an almost continuous biogenic mat covers wide areas in 130 m to 260 m water depth. On this mat, small sponge-bryozoan-serpulid mounds and hedges as well as flatcake-like structures formed by the same organ- isms grow. On steeper margins extensive bryozoan thickets have developed on the volcanic foundation. The dense organic meshwork formed by sponge spicules and bryozoan fragments on the upper sector of the seamount is unique in various ways. Most sponge spicules have a microbial cover, which is an ideal substrate for the settlement of larvae. The spiculite mat is predominantly formed by in situ decay of sponges revealing the actualistic formation of spiculites. The spiculite mat has various cryptic habitats, which are inhabited by bryozoans, ascidians, serpulids, and benthic foraminifers.
Changes in slope inclination and associated sediment dynamics result in the formation of the second facies belt at water depths between 260 m and 400 m. Here semi-stabi- lized sandy mud bottoms and/or dark volcanoclastic sed- iments are colonized by vast numbers of pectinid bivalves and onuphidpolychaetes. Locally sponge-bryozoan-serpulid mounds are developed on elevations of the volcanic founda- tion. The deep slope facies below 400 m consists of various kinds of sponge-bryozoan mounds, sponge mounds, and a typical sponge-crinoid assemblage on coarse volcanoclastics and lava surfaces.
P l a t e 17
Figs. 1-2.
Figs. 3-7.
Selected ecological aspects in coralline algae from the BaJe de Morlaix, Brittany, France, and from northern Norway: The grazing impact and growth rhythmicity
The grazing impact on living coralline algal surfaces. Fig. 1 shows the sharp transition between grazed and ungrazed epithallial cells on Lithothamnion cf. glaciale from the algal reef, northern Norway. Note the diatom frustules which play an important role in biological feedback between grazers and coralline algae. SEM-micrograph, scale bar = 10 grn. A detail of a fecal pellet egested by a herbivore gastropod that grazed on Lithophyllum incrustans, Roscoff, Brittany, France. Scale bar = 10 lain Growth rhythmicity in subarctic (Figs. 3-5) and boreal (Figs. 6-7) branched Lithothamnion. Growth rhythmicity in Lithothamnion cf. glaciale. (Fig. 3) A conspicuous bright-dark banding pattern is visible in cross-sectioned algal branches representing annual growth (= black line). The bright bandings represent summer and the dark bandings represent winter growth increments. (Fig. 4) A close up of a bright (= summer) tiering. Each tier is separated from the following one by a sharp suture that is enriched with organic material. (Fig. 5) A close up of a dark (= winter) first order tiering. The winter tiers are characterized by pronounced "overproduction" of organic matter with respect to calcium carbonate mineralization. Growth rhythmicity inLithothamnion corallioides from the Bale de Morlaix, Brittany (Figs. 5-6). Annual growth is documented in a pair of low-density and high-density bands (see black line). Fig. 6 is a close up of Fig. 5 showing the sharp transition between low-density (summer) and high-density (winter) bands.

P l a t e 17 87

88
Fig. 10. The shallow coastal platforms off Troms, northern Norway, show a clear windward/leeward zonation. Along the wave-exposed parts of the coastal platforms huge kelp-forests (K) prevail. In the sheltered habitats of the kelp-forests, large populations of bryozoans, balanids, verrucids, serpulids, spirorbids and echinoderms are major contributors to the sediment supply. The wave protected areas of embayments or islet archipelagoes are inhabited by coralline algae (C), that can compete with more rapidly growing phaenphytes because of the intense grazing pressure on living corallines. However, the slow-growing corallines could not withstand severe physical disturbances, such as stoma waves (Fm~wALD 1993b).
Sponge-bryozoan deposits in the Permian Canadian Arctic
A fossil counterpart of the arctic sponge-bryozoan buildups are shallow water spiculitic cherts intercalated in the latest Permian sequence of the Sverdrup Basin, Canadian Arctic (BEAuCaAMP 1994). Thin section analysis provides evidence for in s/tu hyalosponge development in these spiculites. InterstralLfied lenses of BRYouoL-carbonates dem- onstrate a close temporal and spatial relationship of both facies associations. Glauconitic sandstones and other sedimentological features proof a shelf origin of these de- posits. Hence, these shallow water spiculites can be distin- guished from deep water cherts that widely occur throughout the upper Paleozoic slope to basinal succession of the Sverdrup Basin.
In the long-term record, the Permian biotic succession of the Sverdrup Basin displays a significant cooling trend starting with highly diversified, tropical-like associations dominated by calcareous algae and foraminifers (identified as CrmOROFORAM by B~,UCnAr,~ 1994) during the Asselian and Sakmarian, followed by temperate bryozoan-echinoderm
and brachiopod assemblages (called BRYONODERM) during the Artinskian to Kazanian interval and finally by the polar- like HYALOSr~NGE-assemblage during the latest Permian. Causes of this dramatic cooling remain enigmatic. The climatic gradient is far greater as can be aspected by the 10 - 15* of northerly migration inferred for Pangaea. However, changes in the global oceanic circulation around Pangaea with a southward deflection of semipermanent sea-ice cov- ered cold currents or atmospheric disturbances by increased mountain uplift might explain this drastic climatic shift (see BZAUCaAMP 1994).
5 NON-TROPICAL OPEN SHELF CARBONATES
Non-tropical open shelf carbonates, including the sea- faced coastal platforms, occur in many areas on both hemi- spheres. On the Northern Hemisphere, research activities concentrate around the Scottish shelves (FARROW et al. 1978, 1984, SCOF~aN et al. 1980, Scor-etN 1988, SCOFF~ & BOWES 1988, WmSON 1979C, 1988), the Western Approaches (BomL~r 1965, LARSOr~mtrR et al. 1982) and the western Canadian shelf around Vancouver Island (Bomb-nOLO &
P l a t e 18
Fig. 1.
Fig. 2. Fig. 3.
Non-tropical coral frameworks constructed by zooxanthellate Cladocora caespitosa and by azooxanthellate Lophelia pertusa
Framework-constructing colony ofCladocora caespitosa from the Aegean Sea sampled from the surface of a 3m thick bank in 20 m water depth. The diameter of the label is 1.5 cm. Section of corallites from a living densely branched colony of Cladocora caespitosa. Scale bar = 1 cm Anastomosing framework constructed by Lophelia pertusa from the Farce Island shelf dredged from 252 m water depth. This slab is kindly provided by O. Tm~AL (Copenhagen) from the BIOFAR-Project. Scale bar = 1 cm).

P l a t e 18 89

90
YORAT~ 1984, NELSON & BOP.mIOLD 1983). In the Barents Sea, extended carbonate sediments were reported from BJORLYKK~ et al. (1978) and BXCKERT & ~ c r a (1989). Occurrences from the Southern Hemisphere concentrate around New Zealand (Nm.soN et al. 1982, 1988a) and the southern Australia shelf (Bombs et al. 1993, BORm~ & JAMES 1993, Busr~ & COLWELL 1982, Cou.ncs 1988, CONOLLY & YON DER BOSCH 1967, JAMES et al. 1992, 1994, WAss et al. 1970). In our studies, special emphasis is laid on the under- standing of kelp-forest dynamics from the geological point of view and on the environmental controls favoring the establishment of BRYOMOL-Carbonates.
5.1 Carbonate production in kelp-forests
Kelp-forests are characteristic elements of non-tropical rocky coasts faced to the open sea (KREM~'R 1980). But also in subtropical-tropical realms, kelp-forests are developed where upwelling of cold and nutrient-rich waters occurs (see Fig. 1). The rigid but flexible large kelp thalli provide morphological highly differentiated habitats for numerous carbonate-secreting organisms. The studies on sedimentary environments on shallow water coastal platforms on the Brittany coast and from northern Norway stress the impor- tance of kelp-forests as distinct site of carbonate production. On these coastal platforms, kelp-forests are situated along the wave-exposed parts of the coastal platforms (Fig. 10). Due to their dense standing stock, kelp-forests drastically reduce the hydrodynamic energy of the open-sea swell and thus have large influence on the intensity and strength of coastal hydrodynamics (JACKSON & Wn~Arcr 1983). In detail, kelp thallioffer habitats of different stability andpersistance. This is reflected in the variablity of secondary epiphytes which live on the kelp. The most stable and most persisting pan of kelp thalli is the rhizoid holdfast including the proximal cauloid. Verrucid and balanid cirripeds, erect but
flexible bryozoans, serpulids, crustose eo~lline algae and sponges, ascidians and hydrozoans belong to a characteristic epiphytic community on the rhizoids and cauloids. The phylloid is most flexible, and in some species, e.g. Lam/nar/a hyperborea, the phylloid gets detached every spring includ- ing their fouling community (LOreN~ 1990). Commonly, the sheet-like membraniporan bryozoans spirorbids and epiph ytic foraminifers (Cibicides lobatulus) are representatives of the low diverse phylloid-fouling community. The process that is termed here as 'phylloid-drift' causes not only a large-scale deposition of macrophytic organic material in intrashelf troughs (e.g. in the Antarctic Bransfield Strait (Fisca-nm 1989)) and in beach lagoonal systems (e. g. Arise de Kcrnic, Brittany), but also a dislocation ofcarbonateparticles by this kind of phytogeneous redeposition. Another very important feature where large brown algae are involved in sedimentological processes is kelp-rafting. Kelp-rafting describes transport of pebbles with attached algae mostly in the nearshore zone (KuDRASS 1974). But also off-shore directed transport of kelp-rafted rocks and macroids is a common but still unrevealed bio-sedimentological process (EMERY & TscmrDv 1941, EMERY 1963, SmrMWAY 1953). Oscillatory water movements and relative positive buoy- ancy are the major prerequisites for kelp-rafting. The modes of transport can be floating, jumping or dragging. On the shore, kelp-rafted pebbles often with encrusting communi- ties stranded in the upper beach zone where normally well- sorted fine sands are deposited. After decomposition of the organic algal float, a pronounced bimodal beach deposit is left behind (FR~tWAU) 1993a. WooDnoP, r~ et al. 1989).
Evidence for fossil kelp-forest carbonates - some considerations
The occurrence of phaeophytic ecosystems, such as the kelp-forests formed by Laminariales and Fucales, is limited
P l a t e
~g. 1.
Fig. 2.
Fig. 3.
Fig. 4.
Fig. 5.
Fig. 6.
19 Modern BRYOMOL-carbonate deposits on deep shelf banks: Brittany shelf offRoscoff
Bryozoan thicket of Cellariafistulosa and Cellaria sinuosa dominating a suspension feeding community of bryozoans, sponges, polychaetes, small bivalves, echinoids, foraminifers and decapods. Caillou, off Roscoff, 80 m water depth, sample D10/93. Gravels as a substrate for suspension feeding communities on the outer shelf. The carbonate sand is dominated by larger fragments of bryozoans, bivalves (juvenile pectinids) and echinoids. CaiUou, off Roscoff, 80 m water depth, sample D10/93. Parautochthonous carbonate sand (<2 mm fraction) from suspension feeding community; high percentage of erect bryozoan fragments (mostly Cellariafistulosa and Cellaria sinuosa) and bivalve hash. Caillou, off Roscoff, 80m water depth, sample D10D3. Selected bryozoan fragments (<2 mm fraction) containing Cellariafistulosa, Cellaria sinuosa, Sertella couchii, P entapora foliacea, Tubicellopora avicularis, Diplosolen obelium and Tubulipora sp. Cailliou, off Roscoff. x 6.5 Allochthonous carbonate sand (250 - 500 pan fraction) from a megadppled field; high percentage of balanids, bivalves, bryozoans and small amount of lithoclasts. Trezen ar Skoden, off Roscoff, 70 m water depth. Selected bryozoan fragments of Cellaria fistulosa, Cellaria sinuosa,P entapora foliacea, Sertella ? couchii and Disporella sp. Trezen ar Skoden, off Roscoff; 70 m water depth, x 6.5

P l a t e 19 91

92
to temperate and eutrophic seas with hardsubstrate-bearing platforms for settlement. Favorable environmental condi- tions for the Laminariales have been developed during the Neogene cooling-trend. This can be substantiated by DNA- DNA-hybridizations on five species of Laminaria pointing to a phylogenetic origin 15 - 19 Million years ago (STAM et al. 1988). According to Es~s & ST--~'~URO (1988), the Laminariales appeared in the boreal northern Pacific. Dur- ing the drowning of the Bering Strait 3.5 Million years ago (McIZ~r~A 1983), the Laminariales started to colonize the northern Atlantic coasts (Lt~aSG & TOM Dmcx 1990). The evolutionary history of important kelp-forest inhabitants with a good fossilization potential show a high degree of accordance. The strongylocentrotid sea urchins and certain patelliform gastropods appeared in the boreal Pacific region during the Miocene (Ln~DBm~ 1976). The same holds for the sea cows (Dugongidae) that migrated into boreal environ- ments not earlier than the Pliocene (Sn~x~tJ et al. 1986). The structural preservation of non-calcifying algae from former phaeophytic communities is rather limited. Few exceptions are findings of phaeophytes in Miocene deposits of Califor- nia (PARKER & DAWSON 1965) and in the Sarmatian section of the Black Sea region (BA~DASARYAN 1983). Therefore, a careful investigation of the calcifying inhabitants of phaeophytic seaweeds, their potential for bioimmuration (Voi~r 1966, 1979), their ability to influence macroid for- marion with distinct successional fouling patterns, and, the trophic structure ofphytogenous fossil associations (JANSSEN 1981) should help to find valuable criteria to recognize fossil carbonate facies originating from kelp-forests. Fossil evi- dence for kelp-rafting is documented by MAVR (1953) from the Lithographic Limestone, Solnhofen. Presumed exam- pies for shallow water carbonate deposits which might be formed within phaeophytic seaweeds are oyster-bryozoan- serpulid-balanid assemblages which are widely distributed in many Cenozoic northwest European basins, such as in the Mainz Basin (Oligocene Meeressand; JANSS~ 1981), in the
Molasse Basin in Lower Austria (Zogelsdoff Formation, Early Miocene; N~m.sicx 1989) and may be also in the Styrian Basin (Middle Miocene; Fv, ma~- 1994).
5.2 Production and deposition of modern BRYOMOL- carbonates on deep shelf banks
Suspension feeding comunities dominatedby bryozoans and molluscs are a widely distributed phenomenon in the mostly aphotic zone on outer shelf banks in mid and high latitudes of the Northern Hemisphere. Living bryozoan communities and related BRvOMOL-Sediments occur along the oceanic gradient of the North Atlantic Current both on temperate (Brittany) and high-boreal shelves (Pls. 19 and 20) but are also reported from RockaU Bank (ScoF~ et al. 1980, WILSON 1979C) and the Orkney Islands (FAm~ow et al. 1984). The present studies in the English Channel and on the Malangsgrunn Bank, northern Norway, are an ac_nJalisric approach focused on the living community structure and carbonate production as well as on processes of destruction and redeposition of skeletal carbonates as BRVOMOL- sediments.
Independancy from light and therefore, lack of competi- tion with benthic algae is considered to be the major reason for the dominance of bryozoan-mollusc communities in the aphotic zone on deeper shelf banks. Suspension-feeding communities on mid and high latitude shelves face distinctly higher and predominantly seasonal variations in food supply compared to low latitude shelves. The benthic-pelagic cou- pling described from deep oceanic basins of the Norwegian- Greenland Sea (GRAF 1989) is considered to control exist- ence and growth rates of BRVOMOL-COmmunities in mid and high latitudes. Signals ofsaisonal growth rates in the bryozoan skeletons (FoR~S~R et al. 1973, PATZOLt~ etal. 1987, ScaXFm~ 1994) are therefore used for calculation ofcarbonateproduc- tion in BRYoMOL-carbonate systems.
P l a t e 20
Fig. 1.
Fig. 2.
Fig. 3.
Fig. 4.
Fig. 5.
Fig. 6.
Modem BRYOMOL-carbonate deposits on deep sheff banks: Malangsgrunn Bank, off Troms, northern Norway
Terrigenous coarse sand and gravel with >20% carbonate content including bivalves, balanid and bryozoan fragments. Malangsgrunn Bank, sample 393B; 84 m water depth. Selected bryozoan fragments of Hornera lichenoides, Sertella septentrionalis, ?Ragionula rosacea and Cellaria sp. Malangsgrunn Bank, sample 393B; 84 m water depth, x 6.5 Coarse carbonate sand (carbonate content >95%) dominated by erect bryozoan fragments, bivalves, foraminifers, echinoids and brachiopods. Malangsgrunn Bank, sample 394C; 90 m water depth. Carbonate sand (500 - 1000 lam fractiori) dominated by bryozoan fragments (Hornera lichenoides, Sertella septentrionalis, Cellaria sp.) bivalves, echinoids, serpulids and foraminifers. M~langsgrunn Bank, sample 394C; 90 m water depth, x 9 Selected bryozoan fragments (1 - 2 mm fraction) of Hornera lichenoides, Sertella septentrionalis, ?Ragionula rosacea, ldmidronea atlantica and of various undeterminable cheilostome species. Malangsgrunn Bank, sample 394C; 90m water dept. x 6.5 Gravel heavily encrusted by diverse cheilostome bryozoans and serpulids, Malangsgrunn Bank, dredge sample 397; 162-169 m water depth.

P l a t e 20 93

94
BRroMoL-communities in carbonate deposits on the deep shelf off the northern Brittany coast
The open shelf of the western part of the English Channel is influenced by two oceanic water masses, the temperate North Atlantic Current and the warm water Lusitanian Current. The English Channel is characterized by a high tidal gauge with extreme strong tidal currents (2.5 - 3 kts in the study area) which affect both the benthic communities and the sedimentary processes. In such a hydrographic regime, formation of stratified waters with a pronounced thermocline is prevented. The water column remains well-mixed through- out the year. Seasonal changes in temperature show a con- tinuous increase from April to August-September (8 ~ - 16~ The major phytoplankton-bloom occurs in spring with a beginning of the annual diatom-bloom in June (MAa- rn~-JmmQtr~, 1983).
The open shelf displays a relatively uniform seabed with the exception of local karst depressions (Fig. 11). The seabottom consists of Eocene limestones which are deeply eroded during Quaternary sea-level low-stands. The bottom is covered with a thin veneer of pebbles produced by rework- ing of Quaternary alluvial formations. The benthic coloniza- tion of the open shelf began with the Holocene transgression. Pebbles, cobbles and boulders which represent a non-mobile hardsubstrate and the steep walls ofk_arstic depressions form ideal habitats for the rich suspension-feeding communities (PI. 19/1-2). The epifauna is abundant and consists of bryozoans, ascidians, hydrozoans, sponges, serpulids, foraminifers and molluscs. Dense thickets of bryozoans show a patch-like distribution on the outer shelf. These thickets are formed by the erect flexible Cellariafistulosa and Cellaria sinuosa which are the principial source of the BgYoMoL-sediment, The dense Celtaria-thickets form a specific habitat that attracts numerous other species to live
in, such as encrusting bryozoans, small decapods, bivalves, hydrozoans, foraminifers. In addition, pebbles and boulders are overgrown by a distinct epifauna composed ofcheilostome bryozoans, serpulids and verrucids.
Strong tidal currents and storm waves that reach the sea bottom are the most important factors for mechanical de- struction of riving communities. These storm wave-events prevent sedimentation of bryozoan sands in the production centers. Therefore, only a small amount of biogenic detritus from the local populations is found between the pebbles as parautochthonous sands. Colony fragments of Palmicella- ria skenei (robust), Omalose cosa ramulosa (delicate),Penta- porafoliacea (foliaceous), Tubicellopora avicularis (multi- laminar) and Buskea dichotoma (delicate) occur in these parautochthonous carbonate sands (PI. 19/3-6). The strong bottom-affecting storm-wave drag and the tidal current regime mobilize skeletal particles either towards the deeper open shelf, or, onto the shallow coastal platforms. On these outer shallow platforms, the well-sorted sediment is piled up in large shell banks such as the Trezen ar Skoden (Fig. 11). The elevation of this subtidal shell-dune is about 10 m above the surrounding seabottom, which extends up to 40-50 m below the sea-level. The modern surface of this particular shell-dune is superposed by large megaripples. A compari- son of these allochthonous carbonate sands from the Trezen ar Skoden with parautochthonous carbonate sands from living bryozoan communities reveals a dominance of erect- cylindrical colony growth forms of Cellaria sinuosa and Cellaria fistulosa. There is also a size-limitation in the occurrence of erect cheilostome bryozoans within the sedi- ment fractions from the parautochthonous carbonate sands. The main components of the fraction >500 lain derive from erect cheilostomata, whereas in the fraction <500 lain erect but fragile anascan particles dominate. This distinct particle distribution within the sandy grain-spectrum reflects the
P l a t e 21
Fig. 1.
Fig. 2.
Fig. 3.
Fig. 4.
Fig. 5.
Fig. 6.
Figs. 7.-8.
Microfacies (MF)-Types of Paleogene/Neogene CrmoaozoAy- (Figs. 1-2) and BRYOMOL- (Figs. 3- 8) carbonates of the Queensland Plateau, northeastern Australia
Larger foraminifer bioclast facies (MF-I). A bioclastic wackestone with abundant Miogypsina and some moldic porosity. Sample 824C-1R-CC-1-6. x 8 Skeletal rudstone of scleractinian-corallinacean microfacies (MF-II). Carbonate cements include dark rims of fibrous, probably original Mg-calcite cement, overgrown by clear scalenohedral cement and cryptocrystalline calcite in a dominantly pendant and geopetal orientation; Sample 824C-8R-CC-4-9. x 12 B ioclastic grainstone (MF-III) with fragments of Halimeda and larger foraminifers including Operculina; Sample 824C-8R-1-45-50. x 30 Bryozoan-larger foraminifer microfacies (MF-III). Well cemented grainstone showing early marine fibrous cement and a later generation of scalenohedral calcite. Note cryptocrystalline in filling of intraparticle porosity; Sample 825B-7R-1-68-71. x 10 Overview of bryozoan-larger foraminifer microfacies (MF-III). Note dominance of erect bryozoan growth-forms and cryptocrystalline infdling of intraparticle porosity; Sample 825B-6R-1-27-29. xl0 Bryozoan microfacies (MF-IV). Poorly cemented bioclasfic grainstone composed of bryozoans and echinoderms. Note partial dissolution of adeonid bryozoan (lower left to right). Gray interparticle porosity is empty pore space; Sample 825B-8R-1-0-12. x 25 Bryozoan microfacies (MF-IV). Bioclastics are heavily broken and angular. Prominent cements are syntaxial overgrowths on echinoderm remains. Fig. 7: Sample 825B-8R-1-0-12; x35 and Fig. 8: Sample 825B-8R-1-0-12; x 35

P l a t e 21 95

96
shape and size range of skeletal fragments with respect to postmortal transport behaviour in different bryozoan spe- cies (see S ~ a 1984).
With the exception of S~m'a & NELSON (1994a), growth rate studies conducted on non-tropical bryozoan species are rare. Skeletal banding patterns are present in the delicate Pentaporafoliacea which also exist in the bryozoan-thick- ets off Brittany. S table oxygen isotopic analyses o f Pentapo ra revealed annual growth rates of 2 cm with growth bands formed during periods of lower water temperature (PArZOLD et al. 1987). Own studies of quantification of carbonate secretion in Pentaporafoliacea and other bryozoan species could provide a tool for calculation of carbonate production within the thickets which are the source for the large spa- tially extended BaYOMOL-Sands on the open shelf off Brit- tany.
BRroMoL-carbonate deposits on the deep shelf off Troms, northern Norway
The deep outer shelf banks off Troms are influenced by two northward flowing water masses, the Norwegian Cur- rent and the Norwegian Coastal Current (EmE 1978). The latter covers most of the shelf area and extends over the Norwegian Current water masses along the shelf edge, whereas the troughs separating the shelf banks are filled with Norwegian Current water masses. Frontal processes be- tween these two water masses, topographically-induced gyres on the shelf and lateral injections of low salinity water from meltwater discharge provide variable conditions for phytoplankton growth. Intense grazing of phytoplankton by copepods in spring prevents complete consumption of nutri- ents and preserves resources for a second bloom in autumn (WAssMAr~ et al. 1991). Surface temperatures over the Malangsgrunn Bank vary between 3~176 in April to 15 ~ 16~ in July (SooT-RY~a~ 1934).
The open shelf is formed by banks emerging less than 80 m below the modem sea-level. The Malangsmnn Bank is
covered by remnants of Weichselian moraines (Voe .~ & ELVSBORa 1979). With the onset of modem oceanographic conditions after the Younger Dryas cooling event 10,000 years ago, the free glacial sediments were washed out leafing behind a sandy bouldery and pebbly lag-deposit (TrIOMSE~ & VORaEN 1986, VORREN & ELVSBORO 1979) (P1. 20/6). Most exposed boulders are colonized by a diverse encrusting bryozoan fauna (P1.20/1) associated with serpul- ids,bivalves and suspension-feeding foraminifers (PI. 20/2). Differences in species composition and successions indicate upside-down orientation of stable boulders and pebbles on the seafloor. Bivalves, gastropods, scaphopods, echinids and ophiurids constitute to the non-sessile epi- and endofauna. At semi-exposed sites on the outer shelf, habitats for erect (Hornera lichenoides) and fenestrate bryozoans (Sertella septentrionalis) and for brachiopods (Macandrevia cra- nium, Terebratulina retusa) exist. These taxa form major constituents of the skeletal carbonate sands and fine gravel on the northwestern flank of the Malangsgrunn Bank. These BRvor,tOL-sediments (P1. 20/3-5) are accumulated by lee- ward eddies of Norwegian Current water which pass as topographically-guided currents (see Strt~Bv 1984) along the northwestern comer of theMalaugsgrunn Bank (Fig. 12).
Reproductive cycles documented by formation of ovicells (cheilostomates) and broodchambers (cyclostomes), rhyth- mic growth bandings and regular growth thickenings of bryozoan skeletons are considered to reflect seasonal nutri- ent cycles (ScaXFEg 1994). ISOtOpic examination of Myriapora coarcta from the northwestern Pacific, a bryozoan with distinct annular growth thickenings revealed different isotopic values from different skeletal parts (FOaES~R et al. 1973). Appositional growth and secondary thickening in a single colony of this taxon indicated by the isotopic studies stresses the careful approach needed towards a budget of carbonate production rates on non-tropical shelves. Myria- pora coarcta is also commonly found on the Malangsgrunn Bank, the Barents Sea and on shelf banks off western Svalbard ( S ~ et al. 1991, At~RtrLErr 1991).
P l a t e 22
Fig. 1.
Fig. 2.
Arctic epibenthic communities and carbonate deposits from the Spitsbergenbank, western Barents Sea
Lower part of the Balanus balanus belt in 58 m water depth. The boulder (upper central section) is colonized by encrusting coralline algae (CA), encrusting bryozoans OE),Balanus balanus (C) and by sea anemones (A). A holothurian (H) rests on the current-exposed flank of the boulder. The carbonate deposits consists ofbalanid fragments and valves ofHiatella arctica (small arrow) andMya truncata (triangle). The siphones (large arrow) belong to Mya truncata. This underwater-photography was taken at 74 ~ 46~1 and 20 ~ 36'E during the Meteor cruise M-21/4. The long axis of the boulder is 30 cm. Section from the Mya truncata-Hiatella arctica-belt in 79m water depth. Boulders and pebbles are intensely fouled by large bushy hydrozoans (HY) and sponges (P). Unattached open-branched bryozoan colonies (X) and strongylocentrotid echinoids (S) are quite abundant in this facies belt. The surface deposit is formed by a coquina lag-deposit containing valves ofMya truncata (Iriangle) and Hiatella arctica (small arrow). The siphones belong to Mya truncata. This underwater-photography was taken at 74 ~ 40'N and 20 ~ 47~E during the Meteor cruise M-21/4. The long axis of the Chlarays islandica (C) valve is 8 cm.

P l a t e 22 97

98
Geological significance
In modern oceans, BRYOMOL-Sediments exist both on mid and high latitude shelves as well as in the tropics where local and seasonal upwelling occurs along western continen- tal margins. BRYoMoIMimestones are documented through- out the Phanerozoic (i.e. Ordovician (BROOKFrmn 1988), Permian (Rao 1981, 1988), Tertiary (JAMES & BONE 1991, JAMES & VON DES BORCH 1991)) and can be used as a trustful pale, oceanographic indicator (see also S ~ O E R & Ko~Ism 1975). However, in contrast to tropical/subtropical Cm.ol~ozoAr~-carbonates, BRVOMOT.-sediments have a much more reduced fossilization potential. The lack of submarine carbonate cementation of skeletal particles in non-tropical BRYoMOL-grain associations is considered to cause the dis- crepancy between gross and net production of carbonate on mid-latitude shelves (SMrm et al. 1992). Furthermore, cold- water carbonate deposits in BRYOMOL-facies are a common characteristic for global ice ages when continental shelves suffered strong sea-level fluctuations that may have im- peded preservation of shelf carbonates during sea-level low- stands.
Skeletal growth forms (NmsoN et al. 1988b), processes of biological (bioerosion), mechanical (tidal currents, storm waves) and chemical degradation (dissolution) (SMrrH & NELSON 1994b), selective transport behavior and redeposition (Svaerar 1984), as well as intermixing with relict particles of different ages (BlCr~RT & H r r , ~ 1989) reveal only a very biased picture of the former living community. Therefore, more actualistic studies of these processes are needed for the interpretation of fossil counterparts.
5.3 Ancient BRYoMOL-carbonate settings on the Queensland Plateau
Since the separation from Antarctica at the end of Eocene, the Australian Plate passed several climatic belts on its northward drift from the high latitudes to the tropics (DAvms 1989). The environmental changes which are linked with the continental drift are documented in the shift from Eocene cool water carbonate deposits of the BRYOMOL-type tO Miocene warm water carbonates of the CMI~ROZOAN-type.
These long-term climatic changes caused by plate tectonics and by paleoceanographic rearrangements are demonstrated by BRACrmRT et al. (1993) from the Queensland Plateau, northeastern Australia, and by JAMES & BONE (1991) from the Eucla Platform, southern Australia.
Climatic control of carbonate plaOrorm development by continental drift
The Queensland Plateau is located east of the Great Barrier Reef province. Therefore, the ODP-Leg 133 pro- vides the opportunity to study the change of climates through the Cenozoic as witnessed by the carbonate platform depos- its. Nedtic non-tropical deposits date from the MiddleEcx~ne to the uppermost Oligocene 03~ r~.ER & Cn~a, a o ~ 1993). Four different microfacies types (MF-I-IV) are distinguished to be climatically significant. (MF-IV) A well-washed BRXOMOL-Carbonate deposit with abundant echinoderms, small foraminifers and molluscs (P1. 21/1). The skeletal fragments are very coarse and poorly sorted. Cryptocrystalline carbonate with few planktonic foraminifers only occurs as infdlings of intraparticle porosity. Except for large syntaxial overgrowth on echinoderms, cementation is virtually ab- sent. Small fragments of branching shallow water bryozoans are mixed with large pieces of deeper water bryozoans such as fenestrate forms, which indicates basinward sediment transport. This conclusion is based on a depth significant zonation of bryozoan growth-forms described from modern non-tropical carbonates off New Zealand (NELSON et al. 1988). (MF-III) BRYOMOL-carbonates with larger benthic foraminifers, few Halimeda and extensive early marine cementation (P1. 21/2). This facies is interpreted to represent the critical temperature interface between tropical and non- tropical environments, comparable to the modern Mediter- ranean transitional type of carbonate deposits. (MF-II and I) represent CHLoROZOaZq-reef limestones and lagoonal depos- its respectively (PI. 21/3 -8). The vertical stacking patterns of MF-types combined from the ODP-sites (811/825 and 824) allows to reconstruct the temperature change of the north- eastern Australian continental margin from the Eocene to the Early Miocene. The sedimentary record reflects warm- temperate to subtropical conditions during the Middle Eocene
P l a t e 23
Fig. 1.
Fig. 2.
Fig. 3.
Arctic epibenthic communities and carbonate deposits from the Spitsbergenbank, western Barents Sea
Section from the Mya-truncata lag-deposit in 101 m water depth. The coquina is mixed with eroded Mesozoic black shales which are winnowed from a glaciomarine deposit. Note the numerous ophiurids (triangles) which live on the lag-deposit. The Mya truncata valves are grazed by strongylocentrotid echinoids (S) and chitons (small arrow). Free-lying branched bryoliths (X) are very common. The siphones (large arrow) belong to Mya truncata. This underwater-photography was taken at 74 ~ 342q and 21 ~ 00~ during the Meteor cruise M-21/4. The scale bar is 30 cm. Chlamys islandica intensely fouled by Balanus balanus settlements on the upper (left) valve. This specimen is sampled from 75 m water depth daring Meteor-cruise M-13/1. Scale bar -- 1 cm. Free-lying branched bryozoan colony on shelly, lag deposts. This underwater-photography was taken during Meteor cruise M-21/4. The diameter of the colony is 10 cm.

P l a t e 23 99

100
(MF-II0. After a hiatus from Late Eocene to Early Oligocene, the Late Oligocene represented cool water BRYOMOL-Car- bonate deposits (MF-IV). The subsequent warming in the earliest Miocene is documented by the well-cemented sub- tropical to tropical MF-II and I. In comparison to the devel- opment of carbonate depesitional systems on the Eucla Platform, southern Australia (JAMES & BONE 1991), the change from temperate to subtropical-tropical climatic envi- ronments took place earlier on the Queensland Plateau (BmxLER et al. 1993, BP.ACrmRT et al. 1993).
5.4 Balanid-dominated cold water open shelf sedimentation: the Arctic Spitsbergen Bank
One of the largest modern epicontinental seas, the Barents Sea, is structured atits western margin between Norway and Svalbard by the shallow open-sea Spitsbergenbank. Under modern conditions, extensive biogenic carbonate produc- tion is observed over the bank and its flanks. The rather warm Atlantic waters of the northward flowing Norwegian Current impinge onto the topographic barrier at the Barents Sea continental margin. Where the barrier of the Barents Sea shelf is dissected by troughs, Atlantic waters intrude east- ward into the Barents Sea and thus get in contact with the Polar waters. Such a configuration is developed on the Spits- bergenbank, a huge over 80,000 km 2 wide shallow shelf- platform, which is bordered by over 400 m deep troughs, e.g. the Bear Island Trough to the south and the Storfjord Trough to the north. While the troughs are f'flled with Atlantic waters, the bank top is flushed by Polar waters of the southwestward heading Hopen-Spitsbergen Current. During the winter pe- fled, the bank is completely covered by sea ice for 6 months on average. In late summer and early autumn most parts of the bank are ice free, and the ice margin has retreated northeastward as far as to Hopen Island. Due to melting sea ice, strong seasonal variations in surface water salinity are observed. However, this salinity reduction is confined to the surface and does not affect significantly the bottom water layer over the bank. The maximum summer temperature is 2~ over the bank and 4* to 6~ in the marginal zone, where Atlantic waters impinge onto the hank. Spitsbergenbank reveals a very pronounced surface sediment distribution that obviously seems to be controlled by intensive bottom cur- rent activity (BJORLYKKE et al. 1978, Bicx~a" & ~ c a 1989). The top of the bank comprises coarse bioclastic sands, while the typical surface sediment on the upper flanks is a polymict, residual coquina/gravel deposit. The forma- tion of the lag deposit is a result of a primary enrichment of a postglacial molluscan infauna (almost exclusively Mya truncata) excaved by strong bottom currents during early Holocene time. Radiocarbon-datings on Mya truncata show an age cluster from 8700 to 6400 years BP). Bottom current vigor increased progressively during the early Holocene due to an glacio-isostatic uplift of the bank causing further intensification of erosional processes. Since about 2500 years BP, a secondary hard substrate fauna, predominantly balanids, settled on the coquina/gravel lag deposit providing the prevalent material for the bioclastic sand on thebank top.
Ecology and patterns of modern carbonate production
Underwater TV surveys and surface sediment sampling ~ c r i et al. 1993, Fmaw~a.o et al. 1994) revealed a clear bathymetric zonation with three major ecologic belts corre- sponding to different sediment dynamics, i.e.:
1. High energy submarine carbonate sand dunes and sandy bottoms in the shallow water zone from 20 m down to 6 m water depth. These sands and free gravels are almost exclusively composed of balanid fragments and benthic foraminifers. Pure carbonate sand dunes up to 1 m height and a few meter wave lengths indicate episodic high energy regimes. During the survey a smoothing of the dune relief with persistent tractional transport of sand by strong bottom currents was observed, but no movement of the dunes itself was registered. Thus dune migration is assumed to berelated to the heavy storm events during late fall/early winter. Because of the overall high sediment mobility, only few organisms, e.g. large holothurians, colonize this habitat. The carbonate factory is located on few outcropping rocky shoals in water depths less than 20 m. These shoals consist of Mesozoic black-shales which are inhabitated by an off- shore kelp-forest- The rocky substrate between the rhizomes of Lam/nar/a saccharina is intensely colonized by Ba/anus crenatus settlements which are the main contributors to the sediment supply.
2. Patchy colonization and small buildups (water depth 60 m to 100 m) on relictic Mya-Hiatella-coquina grounds and rocky substrates by hydrozoans, bryozoans, soft corals and large balanids (PI. 22/1-2). Locally, Chlamys islandica banks with intense overgrowths ofBalanus balanus on the upper valves are observed (PI. 23/2). Colonial hydrozoans, thick-branched bryozoans (P1. 23/3) and barnacles are the dominant faunal elements. Filter-feeding organisms (barna- cles, sponges, bivalves) and decomposers (e.g. some ophiu- rids) are common. Grazing organisms like regular echinoids, gastrotxxls and chitons are less commonly represented. Deposit feeders were not observed. The benthic community is controlled by moderate bottom currents, the presence of large rather stable rock boulders and protected muddy softbottom areas with a typical infauna.
3. Sandy gravel grounds formed by Jurassic to Creta- ceous bed rock clasts, e.g. black shales and organic-rich siltstones, and relictic subfossil Mya truncata valves domi- nate in water depth below 100 m (PI. 23/1). These relictic shells are heavily bored (PI. 23/3). Very abundant, small ophiurids, sea urchins and asteroids, as well as onuphid polychaetes colonize these substrates. With increasing depth, muddy bottoms with the endobenthic Clinocardium ciliata gain in importance.
Major environmental controls
Main environmental controls of modem carbonate pro- duction on Spitsbergenbank are: -- Variations in timing andlocation of spring phytoplankton blooms. Warm years are characterized by a delayed phytoplankton bloom and an earlier stage of zooplankton development in Atlantic waters, apparently resulting in a
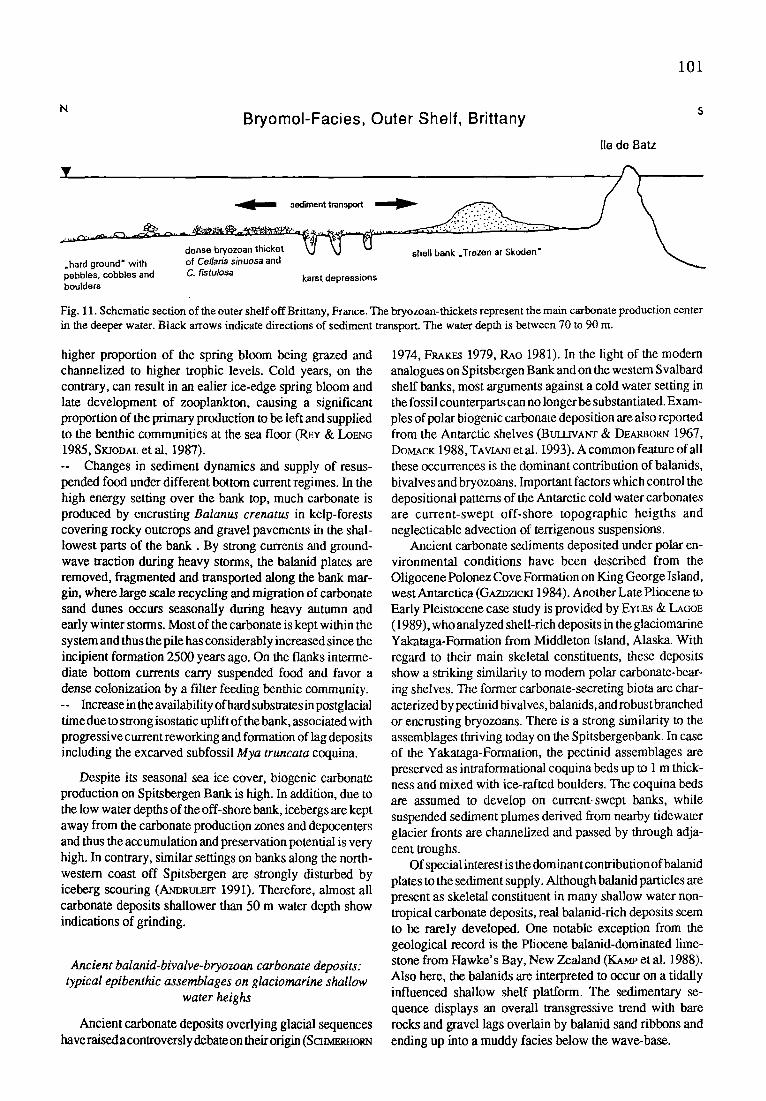
101
N Bryomol Facies Outer Shelf Brittany s
lie de Batz
m~liment transport ~
dense bryozoan thicket \ I \1 v . . . . . . . . . . . . . . . . ,__. of Cellaria sinuosa and - v shell bank .Trezen ar Skoden"
.hard ground" wi th pebbles, cobbles and boulders
C. fistulosa karst depressions
Fig. 11. Schematic section of the outer shelf off Brittany, France. The bryozoan-thickets represent the main carbonate production center in the deeper water. Black arrows indicate directions of sediment transport. The water depth is between 70 to 90 m.
higher proportion of the spring bloom being grazed and channelized to higher trophic levels. Cold years, on the contrary, can result in an ealier ice-edge spring bloom and late development of zooplankton, causing a significant proportion of the primary production to be left and supplied to the benthic communities at the sea floor (R~Y & LOZNG 1985, S~JODAL et al. 1987). -- Changes in sediment dynamics and supply of resus- pended food under different bottom current regimes. In the high energy setting over the bank top, much carbonate is produced by encrusting Balanus crenatus in kelp-forests covering rocky outcrops and gravel pavements in the shal- lowest parts of the bank. By strong currents and ground- wave traction during heavy storms, the balanid plates are removed, fragmented and transported along the bank mar- gin, where large scale recycling and migration of carbonate sand dunes occurs seasonally during heavy autumn and early winter storms. Most of the carbonate is kept within the system and thus the pile has considerably increased since the incipient formation 2500 years ago. On the flanks interme- diate bottom currents carry suspended food and favor a dense colonization by a filter feeding benthic community. -- Increase in the availability of hard substratesin postglacial time due to su'ong isostatic uplift of the bank, associated with progressive current reworking and formation of lag deposits including the excarved subfossil Mya truncata coquina.
Despite its seasonal sea ice cover, biogenic carbonate production on Spitsbergen Bank is high. In addition, due to the low water depths of the off-shore bank, icebergs are kept away from the carbonate production zones and depocenters and thus the accumulation and preservation potential is very high. In contrary, similar settings on banks along the north- western coast off Spitsbergen are strongly disturbed by iceberg scouring (ANDRUt~rr 1991), Therefore, almost all carbonate deposits shallower than 50 m water depth show indications of grinding.
Ancient balanid-bivalve-bryozoan carbonate deposits: typical epibenthic assemblages on glaciomarine shallow
water heighs
Ancient carbonate deposits overlying glacial sequences have raised a controversly debate on their origin (Scmc~RaORN
1974, FRAr, r.S 1979, RAO 1981). In the light of the modem analogues on Spitsbergen Bank and on the western S valbard shelf banks, most arguments against a cold water setting in the fossil counterparts can no longer be substantiated. Exam- ples of polar biogenic carbonate deposition are also reported from the Antarctic shelves (BULLIVANT & DEARBORN 1967, DOMACX 1988, TAVtAM et al. 1993). A common feature of all these occurrences is the dominant contribution of balanids, bivalves and bryozoans. Important factors which control the depositional patterns of the Antarctic cold water carbonates are current-swept off-shore topographic heigths and neglecticable advection of terrigenous suspensions.
Ancient carbonate sediments deposited under polar en- vironmental conditions have been described from the Oligocene Polonez Cove Formation on King George Island, west Antarctica (Gxzozac~ 1984). Another Late Pliocene to Early Pleistocene case study is provided by EYLES & LAC, OE (1989), who analyzed shell-rich deposits in the glaciomarine Yakataga-Formation from Middleton Island, Alaska. With regard to their main skeletal constituents, these deposits show a striking similarity to modem polar carbonate-bear- ing shelves. The former carbonate-secreting biota are char- acterized by pectinid bivalves, balanids,and robustbranched or encrusting bryozoans. There is a strong similarity to the assemblages thriving today on the Spitsbergenbank. In case of the Yakataga-Formation, the pectmid assemblages are preserved as intraformational coquina beds up to 1 m thick- ness and mixed with ice-rafted boulders. The coquina beds are assumed to develop on current-swept banks, while suspended sediment plumes derived f~om nearby tidewater glacier fronts are channelized and passed by through adja- cent troughs.
Of special interest is the dominant contribution ofbalanid plates to the sediment supply. Although balanid particles are present as skeletal constituent in many shallow water non- tropical carbonate deposits, real balanid-rich deposits seem to be rarely developed. One notable exception from the geological record is the Pliocene balanid-dominated lime- stone from Hawke's Bay, New Zealand (KAMP et al. 1988). Also here, the balanids are interpreted to occur on a tidally influenced shallow shelf platform. The sedimentary se- quence displays an overall transgressive trend with bare rocks and gravel lags overlain by balanid sand ribbons and ending up into a muddy facies below the wave-base.

102
Discussion 6 MAJOR CONTROLS AND MECHANISMS OF
NON-TROPICAL CARBONATE SEDIMENTATION
Control of antecedent topography and sea-level changes
Geometry and distribution patterns of carbonate buildups and related deposits on shelves are strongly determined by seabed topography (JoNEs & DESROCrn~S 1992). Non-tropi- cal carbonate producers have not the capability to construct wave-resistant structures as their tropical counterparts do. As result, the non-tropical shelves as well as individual platforms on them are generally unrimmed. On swell-domi- nated deeper shelves, downslope but also upslope transport of the BRVOMOL-particles OCCurS (see section 5.2). The respective areas of reworking and redeposition are control- led by the depth-penetration of the storm-wave base (see sections 5.2 and 5.3).
This in turn has severe implications for the response to sea-level changes. Tropical buildups are quickly lithified and form karst reliefs during lowstands. During the subse- quent sea-level rise these hard substrates easily can be colonized. In contrary, non-tropical shelves show multiple reworking and redeposition during lowstands more similar to siliciclastic shelves. Due to the lack of any karst lithification and its resultant topography only palimpsest sediments can be colonized by the epibenthic fauna. As a consequence, the propability for non-tropical carbonate systems to be drowned in response to high-frequency sea-level changes may exceed those of tropical associations (SIMONE t~ CARANNANTE 1988). The existence of islet archipelagoes on coastal platforms acting as breakwaters are a major prerequisite to facilate coralline algal framework growth. Otherwise, storm-trig- gered disturbances could not be compensated by the slow- growing algal biotopes. During postglacial time, coralline algal constructions occur at specific radiocarbon-dated age- clusters, which coincide with repeatative short-term sea- level standstills during the past 6000 years in the Troms District (see section 4.1).
Oceanographic control
Throughout Earth's history, no period has experienced such an efficient meriodional exchange of surface waters between the equator and the poles and such an extensive nutrient recycling between surface and deep waters as under modern conditions. This setting is manifested by extreme latitudinal contrasts of modern non-tropical carbonate sedi- mentation. At 70 ~ northern latitude, high accumulating coralline algal reefs grow in the skerries of Northern Norway under the marginal influence of Atlantic waters, while in the corresponding Greenland sector Arctic sponge-bryozoan buildups develop on the seamount Vesterisbank in polar, almost 10 months of the year sea-ice covered waters (see sections 4.1 and 4.3). Hence, the endmember of modern FORaMOL-Carbonates is found at the same paleolatitude as the temperate to subarctic red algae dominated FORAMOL- subfacies. Looking back into the fossil record, the Permian subtropical to polar succession of FORAMOL-Subfacies in the
Sverdrup Basin, Canadian Arctic, displays striking similari- ties, specifically with regard to the cold water endmember, the hyalosponge-cherts and associated BRYOMOL-carbonates (BEAUCHAXVa" 1994). Under the perspecrive of actualismn, the speculation of a similar oceanographic control, e.g. the existence of a Permian Greenland Current equivalent along the northern tip of Pangaea, is very attractive. Another interesting feature in comparison to the modern oceano- graphic configuration is the mid-Permian Posphoria-For- marion at the equatorial western margin of Pangaea in areas believed to have been affected by intense cold water upweUing (WhRDLAW & COLLI~SON 1986).
Latitudinal control of insolation
On a global view, most carbonate mineralizes through biological activity within the photic zone. The boundary separating photic from aphotic conditions varies with lati- tude depending on a different penetration of various wave lengths of natural light. For example, in low latitudes, deepest occurrence of coralline algae has been reported at 268 m water depth ( L ~ & L ~ 1985), whereas on the Hornsundbank off Svalbad (76~ deepest coralline algae were found in 73 m water depth (ScrtXFER et al. 1991). This equatorward downshifting of the lower limit of the photic zone is also reflected by a shift of the growth maximum of branching Lithothamnion spp. in 10 - 20 m water depth in polar environments to 40 - 60 m water depth in the Mediter- ranean (see section 4.1). In the tropical low latitudes, maerl- type carbonates are even found at greater depth. This obser- vation of the so-called "polar emergence" has strong impli- cations for the interpretation of red algal occurrences in the geological record. Without going into the species level or having other independant depth criteria at hand, subarctic shallow water maerl may be confused with deeper water low latitudes maerl deposits (CARAtCNArCrE et al. 1988).
The upper limit of the azooxanthellate coral Lophelia pertusa is confmed to waters below the thermocline, where only minor variations in temperature and salinity are regis- tered (see section 4.2). Since the position of the thermocline is largely dependant on seasonal variations in the warming up of the upper surface layer by solar energy and by the mixing of water through storm periods, a similar feature like the "polar emergence" of coralline algae has also been noted for the upper limit of Lophelia-occurrences (STArg-~V & CAtRNS 1988). l_x)oking back into the geologic record, the clear separation of different depth habitats of framework constructing zooxanthellate and azooxanthellate corals can be traced back at least to the Late Cretaceous (COATES & KAtWFMAN 1973). A very impressive example of Danian aphodc coral mounds have been described by B ~ a ~ R & WEmucu (1990). There is much debate, whether an actualistic approach can also be applied for the early evolution of scleractinia (STANLEY 1981). Coral heads and thickets of Triassic Thecosmilia occur in completely different environ- mental settings and depth habitats coveting for example shallow back reef areas or deeper ramp sections 1971, STAW-ZV 1979, HENPacat 1983). According to S T ~
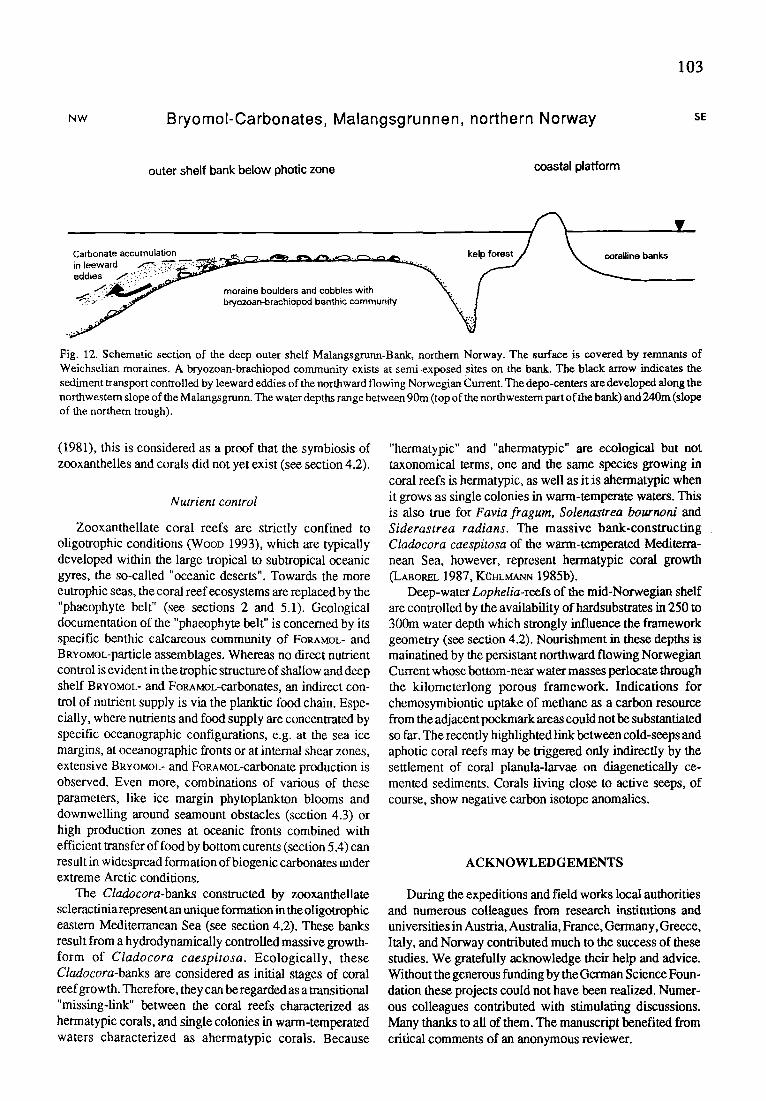
103
NW BryomoI-Carbonates, Malangsgrunnen, northern Norway SE
outer shelf bank below photic zone coastal platform
Carbonate accumulation �9 , ~ , . ~ ~ ~"~,.~r ~ c~x .~ ,~ kelp forest / ~ coralline banks
eddie, \ , r _ ~ ~ , , ~ " moraine bou ders and cobbles with ~..
�9 7 ~ ' ~ " " " ~ . . . . bryozoan-brachiopod benthic community ~ : X J
Fig. 12. Schematic section of the deep outer shelf Malangsgrurm-Bank, northern Norway. The surface is covered by remnants of Weichselian moraines. A bryozoan-brachiopod community exists at semi-exposed .sites on the bank. The black arrow indicates the sediment transport controlled by leeward eddies of the northward flowing Norwegian Current. The depo-centers are developed along the northwestern slope of the Malangsgrurm. The water depths range between 90m (top of the northwestern part of the bank) and 240m (slope of the northern trough).
(1981), this is considered as a proof that the symbiosis of zooxanthelles and corals did not yet exist (see section 4.2).
Nutrient control
Zooxanthellate coral reefs are strictly confined to ohgotrophic conditions (Wood 1993), which are typically developed within the large tropical to subtropical oceanic gyres, the so-called "oceanic deserts". Towards the more eutrophic seas, the coral reef ecosystems are replaced by the "phaeophyte belt" (see sections 2 and 5.1). Geological documentation of the "phaeophyte belt" is concerned by its specific benthic calcareous community of FOgAMOL- and BavoMoL-particle assemblages. Whereas no direct nutrient control is evident in the trophic structure of shallow and deep shelf BRYOMOL- and FoRAMOL-carbonates, an indirect con- trol of nutrient supply is via the planktic food chain. Espe- cially, where nutrients and food supply are concentrated by specific oceanographic configurations, e.g. at the sea ice margins, at oceanographic fronts or at internal shear zones, extensive BRYOMOL- and FORAMOL-Carbonate production is observed. Even more, combinations of various of these parameters, like ice margin phytoplankton blooms and downwelling around seamount obstacles (section 4.3) or high production zones at oceanic fronts combined with efficient transfer of food by bottom curents (section 5.4) can result in widespread formation ofbiogenic carbonates under extreme Arctic conditions.
The Cladocora-banks constructed by zooxanthellate scleractiniarepresent an unique formation in theoligotrophic eastern Mediterranean Sea (see section 4.2). These banks result from a hydrodynamically controlled massive growth- form of Cladocora caespitosa. Ecologically, these Cladocora-banks are considered as initial stages of coral reef growth. Therefore, they can be regarded as a transitional "missing-link" between the coral reefs characterized as hermatypic corals, and single colonies in warm-temperated waters characterized as ahermatypic corals. Because
"hermatypic" and "ahermatypic" are ecological but not taxonomical terms, one and the same species growing in coral reefs is hermatypic, as well as it is ahermatypic when it grows as single colonies in warm-temperate waters. This is also true for Favia fragum, Solenastrea bournoni and Siderastrea radians. The massive bank-constructing Cladocora caespitosa of the warrn-temperated Mediterra- nean Sea, however, represent hermatypic coral growth (LABoI~L 1987, KOIqLMAr~ 1985b).
Deep-water Lophelia-reefs of the mid-Norwegian shelf are controlled by the availability of hardsubstrates in 250 to 300m water depth which strongly influence the framework geometry (see section 4.2). Nourishment in these depths is mainatined by the persistant northward flowing Norwegian Current whose bottom-near water masses perlocate through the kilometerlong porous framework. Indications for chemosymbiontic uptake of methane as a carbon resource from the adjacent pockmark areas could not be substantiated so far. The recently highlighted link between cold-seeps and aphotic coral reefs may be triggered only indirectly by the settlement of coral planula-larvae on diagenetically ce- mented sediments. Corals living close to active seeps, of course, show negative carbon isotope anomalies.
ACKNOWLEDGEMENTS
During the expeditions and field works local authorities and numerous colleagues from research institutions and universities in Austria, Australia, France, Germany, Greece, Italy, and Norway contributed much to the success of these studies. We gratefully acknowledge their help and advice. Without the generous funding by the German Science Foun- dation these projects could not have been realized. Numer- ous colleagues contributed with stimulating discussions. Many thanks to all of them. The manuscript benefited from critical comments of an anonymous reviewer.

104
REFERENCES
Asm., E. F. (1959): Zur Kennmis der mafinen H6hlenfauna unter besonderer Berilcksichtigung der Anthozoen. Ergebnisse der Osterreichischen Tyrrhenia-Expedition, 1952, part V. - Pubbl. Staz. Zool., 30 (suppl.), 1-94, Neapel
AD~.y, W. H. & McKmB~, D. L. (1970): Studies on the maerl species Phymatolithon calcareum (Pallas) nov. comb. and Lithothamnium coralloides Crouan in the Ria de Vigo. - BotanicaMarina, 13, 100-106, Hamburg
AIO, AN, E. B. & FARROW, G. E. (1984): Depth and deposition of Early Holocene raised sediments at Irvine Island deduced from algal borings in mollusc shells. - Scottish J. Geol., 20/2, 237- 247, Edinburgh
Am'AN, E. B. & FARROW, G. E. (1985): Shell bioerosion in high- latitude low-energy environments: Firths of Clyde and Lome, Scotland. - Marine Geology, 67, 139-150, Amsterdam
At.F.XAr~mtSSON, T. (1972): Micfitization of carbonate particles: Processes of precipitation and dissolution in modern shallow- marine sediments. - Bull. Geol. Inst. Univ. Upsala N.S., 37L 201-236, Upsala
-- (1974): Carbonate cementation in coralline algal nodules in the Skagerrak, North Sea: Biochemical precipation in under- saturated waters. - J. Sed. Petrol., 44/1, 7-26, Tulsa
A N D R e , H. (1991): Fazies, Genese und Verbreitung arktischer Karbonate auf dem Westspitzbergenschelf. - unver/SffentI. Diplomarbeit, Christian-Albrechts-Universit~, Kiel
B^ODASARYAN, K. G. (1983): Ecological systems of the Sarmatian Sea. - Palexmt. Journal, 1983/4, 1-9, Moskow
BAm~'Es, R. S. K. & MANN, K. H. (1980): Fundamentals of aquatic ecosystems. - 229 p., Oxford (Blackwell)
BgAUCnAMP, B. (1994): Permian climatic cooling in the Canadian Arctic. - In: K~-~-rN, G. D. (ed.): Pangea: Paleoclimate, tecton- ics, and sedimentation during accretion, zenith, and break-up of a supercontinent. - Geol. Soc. Am., Spec. Paper, 288, 229- 246, Washington
Br~ J.~xto, S. (1974): The compound and intermediate wall struc- tures in Cibicidinae (foramonifera) with remarks on the radial and granular wall structures. -Bul l . Geol. Inst. Univ. Uppsala N. S., 5, 1-11, Upsala
BmtS~CrER, M. & WE:DtaC~, O. (1990): The Danian (Paleocene) Coral Limestone of Fakse, Denmark: a model for ancient aphotie, azooxanthellate coral mounds. - Facies, 22, 103-138, Erlangen
B~a'zLmt, C. & CrlAPROr~RE, G. C. H. (1993): Palaeogene and Neogene larger foraminifers from the Queensland Plateau: biostratigraphy and environmental significance. - Proc. Ocean Drilling Program, Sci. Res., 133, 51-66, College Station
BEaTa.~, C., KROON, D., GARTS~, S. & WEs, W. (t993): Eocene to Miocene chronostratigraphy of the Queensland Plateau: con- trol of climate and sea level on platform evolution~ - Proc. Ocean Drilling Program, Sci. Res., 133, 281-289, College Station
BIC~T, T. & ~ C H , R. (1989): Karbonate nahe der Arktis: Rezente Flachwasserkarbonate auf der Spitzbergenbank ('BarentsschelO. - Geol. Pal. Mitt. Innsbruck, 16, 4, Innsbruck
BJcm.vm~, K., Btra, B. & ELvmaoI, A. (1978): Quaternary sediments in the north-western part of the Barents Sea and their relation to the underlying Mesozoic bedrock. - Sedimentology, 25, 22%246, Oxford
BOa.LOT, G. (1965): Organogenic gradients in the study of neritic deposits of biological origin: The example of the western English Channel. - Marine Geology, 3, 359-367, Amsterdam
BoJmm~, %, JAMES, N. P., WmSON, C. & Heoom, D. (1993): Surficial cool-water carbonate sediments on the Otway continental margin, southeastern Australia. -Marine Geology, 112, 35-56, Amsterdam
B o ~ , T. D. & JAMES, N. P. (1993): Holocene sediment dynamics on a cool-water carbonate shelf: Otway, southeastern Aus- tralia. - J. Sed. Petrol., 63/4, 574-588, Tulsa
BOrn, mOLD, B. D. & YoRAa~a, C. J. (1984): Surficial geology of the continental shelf, northwestern Vancouver Island. - Marine Geology, 5, 89-I 12, Amsterdam
Bos~,rcs, D. W. J. (1976): Ecological studies on two unattached coralline algae from western Ireland. - Palaeontology, 19/2, 365-395, London
- - (1980): Sedimentary facies, production rates and facies models for recent coralline algal gravels, Co. Galway, Ireland. - Geological Journal, 15/2, 91-111, Liverpool
-- (1983): Coralline algal reef frameworks. - Journal of the Geo- logical Society London, 140, 365-376, London
BRACrmRT, T. C., Bm'ZI~R, C., DAvms, P. J. & F~*RV, D. A. (1993): Climatic change: control of carbonate platform development (Eocene - Miocene, Leg 133, northeastern Australia). - Proc. ODP, Scientific Results, 133, 291-300, College Station
BROeZK~.R, W. S., BOND, G., KLAS, M., BONANI, G. & WOLFLt, W. (1990): A salt oscillator in the glacial North Atlantic? 1. The concept. - Paleoceanography, 5,469-477, Washington
Bgo~n.~Y, R. G. & HANKm~, N.-M. (1981): Shallowmarine bioerosion at Vardr arctic Norway.- Bull. geol. Soc. Denmark, 29, 103- 109, Copenhagen
B~OOKr~_.LD, M. E. (1988): A mid-Ordovician temperate carbonate shelf - the Black River and Trenton Limestone Groups of southern Ontario, Canada. - Sedimentary Geology, 60, 137- 153, Amsterdam
BuooE, T. (1980): Ovrelags geologip~tkontinentalsokkelen utenfor Mere og Trondelag.- IKU-Report, 104, 1-44, Trondheim
BuLtaVArzr, J. S. & DEARaOP.~, J. H. (1967): The fauna of the Ross Sea. - New Zealand Dep. Sci. lndustr. Res. Bull., 176, 9-77, Wellington
Bum~, R. V. & COLW~.L, I. B. (1982): Temperate carbonate sediments of northern Spencer Gulf, South Australia: a high salinity 'foramol' province. - Sedimentology, 29, 223-238, Oxford
CAaIOCH, J. ( 1969): Les fonds de maerl de la B ale de Morlaix et leur peuplement vdgdtal. - Cahiers Biologique Marine, 9, 33-55, Paris
CARA~mCrE, G., ESa~aAN, M., M ~ , J. D. & S~ONE, L. (1988): Carbonate lithofacies as paleolatitude indicators: prob- lems and limitations. - Sedimentary Geology, 60, 333-346, Amsterdam
CHAVE, K. E. (1967): Recent carbonate sediments - an unconven- tional view. - Jour. Geol. Educat., 15/5,200-204, Washington
COATES, A. G. & KAUFFMAN, E. G. (1973): Stratigraphy, paleontology, and paleoenvironment of a Cretaceous coral thicket, Lamy, New Mexico. - Journal of Paleontology, 47, 953-968, Tulsa
CoLt,S, L. B. (1988): Sediments and history of the Rotmest Shelf, southwest Australia: a swell-dominated, non-tropical carbon- ate margin. - Sedimentary Geology, 60, 15-49, Amsterdam
CONOLLV, J. R. & YON Dm~ BORCH, C. C. (1967): Sedimentation and physiography of the sea floor south of Australia.- Sedimentary Geology, 1, 181-220, Amsterdam
COmVAY, K. W., BARRm, J. V., AUSTIN, W. C. & l.arrm~Atn~t, J. L. (1991): Holocene sponge bioherms on the western Canadian continental shelf. - Continental Shelf Research, 11/8-10, 771- 790, Oxford
DAVIES, P. J. (1989): The evolution of the carbonate platforms of northeast Australia. -SEPM, Special Publ., 44, 233-258, Tulsa
DAYTON, P. K., ROBmUARD, G. A., PAINE, R. T. & DAYTON, L. B. (1974): Biological accomodation in the benthic community at McMurdo Sound, Antarctica. - Ecol. Monogr., 44, 105-128, Durham
DOMACK, E. W. (1988): Biogenic facies in the Antarctic glacimafine environment: basis for a polar glacimarine summary. - Marine Geology, 63, 357-372, Amsterdam
DoNs, C. (1935): ZoologischeNotizenXXIX. Die Verbreitung yon Steinkorallen in West-Fiunmark. - Det Kongelige Norske Videnskabers Selskab, Forhandl., 8[18, 57-60, Trondheim
-- (1944): Norges korallrev. - Det Kongelige Norske Videnskabers

105
Selskab, Forhandlinger, 16, 37-62, Trondheim EmE, L. I. ( 1978): Ocean currents on the Halten andMalangsgrurmen
banks. - IKU-Report, 97, 1-302, Trondheim En .~rs~ , H. C., FAtX-PE~s~, S.0 HOPKINS, C. C. E. & TAr~E, K.
(1981): Ecological investigations on the plankton community of Balsfjorden, northern Norway.- Sarsia, 66: 25-34, Bergen
EMERY, K. O. (1963): Organic transportation of marine sediments. - In: HILL, M. N. (ed.): The Sea, 3. - 776-793, New York 0nterscience Publishers)
EMI~'t, K. O. & Tsc~tmy, R. H. (1941): Transportation of rock by kelp. - Bull. geol. Soc. America, 52, 855-862, Boulder
EsTm, J. A. & S~Ir,rBwG, P. D. (1988): Predation, herbivory, and kelp evolution. - Paleobiology, 14/1, 19-36, Chicago
EYt~, N. & LAC, OE, M. B. (1989): Sedimentology of shell-rich deposits (coquinas) in the glaciomarine upper Cenozoic Yakataga Formation, Middleton Island, Alaska. - Geol. Soc. Am. Bull., 101, 129-142, Boulder
F^OFa~S~OM, J. A. (1987): The evolution of reef communities. - 600 pp., New York (Wiley)
FAR~^M, W. F. & J~'nsor~, N. A. (1977): A survey of the maerl beds of Falmouth (Cornwall). - British Phycological Journal, 12, 119, Glasgow
FARROW, G. E. & Fx'~, J. A. (1988): Bioerosion and carbonate mud production on high-latitude shelves. - Sedimentary Geology, 6 0 , 281-297, Amsterdam
FARROW, G. E., AH.r~, N. H. & Am, AN, E. B. (1984): Bioclastic carbonate sedimentation on a high-latitude, tide-dominated shelf: northeast Orkney Islands, Scotland. - Journal of Sedi- mentary Petrology, 54/2, 373-393, Tulsa
FARROW, G. E., CuccI, M. & ScoFfs, T. P. (1978): Calcareous sediments on the nearshore continental shelf of western Scot- land. - Proc. Roy. Soc. Edinburgh 76B, 55-75, Edinburgh
Fmcrn~, G. (1989): Stabile Kohleustoff-Isotope in partikul~er organischer Substanz aus dem Stidpolarmeer (Atlantischer Sektor). - Ph.D.-Thesis, Ber. Fachbereich Geowiss, Bremen University, 5, 161pp, Bremen
FL0O~L, E. (1981): Paleoecology and facies of Upper Triassic reefs in the Northern Calcareous Alps. - In: T o o ~ , D. F. (ed.): European fossil reef models. - Soc. Econ. Paleont. Miner., Spec. Publ., 30, 291-359, Tulsa
Foe, ma-F~, R. M., SAWoa~, P. A. & ANDFa~SOt~, T. F. (1973): Isotopic variability of cheilostome bryozoan skeletons. - In: LAgWOOD, P. (ed.): Living and fossil bryozoa.- 79-94, London (Academic Press)
FRA~.S, L. A. (1979): Climates throughout geologic time. - 310 pp, Amsterdam (Elsevier)
FRE~RIlCSEN, R., J ~ s ~ , A. & W E s ~ m ~ c , H. (1992): The distri- bution of the scleractinian coral Lophelia pertusa around the Faroe Islands and the relation to internal tidal mixing. Sarsia, 77, 157-171, Bergen
FgmWALD, A. (1993a): Subarktische Kalkalgertriffe im Spiegel hochffequenter Meeresspiegelschwankungen)and intemer bio- logischer Steuerungsprozesse. - Dissertationsschrift, Universit~ Kiel (in preparation for publication in Contribu- tions to Sedimentology).
-- (1993b): Coralline algal maerl frameworks - islands within the phaeophytic kelp belt. - Facies, 29, 133-148, Erlangen
-- (1995): Bacteria-induced carbonate degradation: ataphonomic ease study on Cibicides lobatulus from a high-boreal carbonate setting. - Palaios (published in 1995)
-- (in press): Sedimentological and biological aspects in the for- mation of branched rhodoliths in northern Norway. - Osterreichische Beitr,%e zur Palaontologie (ISRS Conf. Vol- ume), Wien
FRZlWAt~, A. & ~ c r l , R. (1994): Reefal coralline algal build- ups within the Arctic Circle: morphology and sedimentary dynamics under extreme environmental seasonality. - Sedimentology, 41, 963-984, Oxford
Fg~wAt~, A., ~ c ) t , R., S r P. & Wn~KOMM, H. (1991): The significance of high-boreal to subarctic maerl deposits in
northern Norway to reconstruct Holocene climatic changes and sea level oscillations. - Facies, 25, 315-340, Erlangen
FgFawat~, A., ROpSTO~, P. & ~F_a ~s , H. (1994): Boreal shallow water carbonates.-In: Stress, E., ~ o , K. & Mm~.gT, J. (eds.), Nordatlantik 1993, Meteor Cruise No. 26, Meteor Berichte, 94/4, 138-142, Hamburg
FRmaE, J. G. (1994): Serpulid-bryozoan-foraminiferan biostromes controlled by temperate climate and reduced salinity: Middle Miocene of the Styrian Basin, Austria. - Facies, 30, 51-62, Erlangen
GAzozIcla, A. (1984): The Chlamys coquinas in glacio-marine sediments (Pliocene) of King George Island, West Antarctica. - Facies, 10, 145-152, Erlangen
GmAUD, G. & CnBIocri, J. (1983): Inclusions cytoplasmiques remarquables chez les Coralinac6es. - Annales des Sciences Naturelles, Botanique, Set. 13/5, 29-43, Paris
GiscrtL~, E., G R ~ , K.-U. & WmOMAr~, J. (1994): The Upper Cretaceous Lacazina Limestone in the Basco-Cantabrian and Iberian Basins of northern Spain: Cold-water grain associa- tions in warm-water environments. - Facies, 30, 209-246, Eflangen
Goulxov, A. N. & SCARLAa~O, O. A. (1973): Comparative character- istics of some ecosystems of the upper regions of the shelf in tropical, temperate and arctic waters. - Helgol~nder wiss. Meeresuntersuchungen, 24, 219-234, Hamburg
GRA~, G. (1989): Benthic-pelagic coupling in the deep-sea benthic community. - Nature, 341,437-439, London
HAO~, N. T. (1983): Destructive grazing of kelp beds by sea urchins in Vestfjorden, northern Norway. - Sarsia, 68, 177- 190, Bergen
HAt~OCK, P. & SCrmAO~, W. (1986): Nutrient excess and the demise of coral reefs and carbonate platforms. - Palaios, 1, 389-398, Tulsa
H~,mIcH, R. (1983): Der Wettersteinkalk am Nordwestrand des tirolischen Bogens in den N6rdlichen Kalkalpen: der j~lngste VorstoB einer Flachwasserplattform am Beginn der Obertrias. - Geologica et Palaeontologica, 17, 137-177, Marburg
~ c H , R., HARrMAr~, M., R ~ , J., S c ~ P., FROaW~D, A., S ar.mMm-Z, S., DmrgICH, P. & TrimoE, J. (1992): Facies belts and communities of the arctic Vesterisbanken Seamount (Central Greenland Sea). - Facies, 27, 71-104, Erlangen
~ c H , R., ~ , J. & WEmu~a,m, A. (1993): Cold water shelf carbonates and lag deposits and associated living benthic communities: Spitsbergen B ank.-In: PFANNKUCHE, O., DUINKER, J. C., GRAF, G., HENRICH, R., THIEL, H. & ZErrSCHEL, B. (eds.): Nordatlantik 92, Meteor-Reise Nr. 21, Meteor Berichte, 93-4, 176-180, Hamburg
HOVLAND, M. (1990): Do carbonate reefs form due to fluid seep- age?-Terra Nova, 2, 8-18, Oxford
JACKSON, G. A. & WINANT, C. D. (1983): Effect of a kelp forest on coastal currents. - Continental Shelf Research, 2/2, 75-80, Oxford
JAMES, N. P. & BONE, Y. (1991): Origin of a cool-water, Oligo- Miocene deep shelf limestone, Eucla Platform, southern Aus- tralia. - Sedimentary Geology, 60, 323-341, Amsterdam
JAMES, N. P. & VON DER BORCH, C. C. (1991): Carbonate shelf edge off southern Australia: a prograding open-platform margin. - Geology, 19, 1005-1008, Boulder
JAMES, N. P., BONE, Y., YON DER BORCH, C. C., Gos'rtN, V. A. (1992): Modem carbonate and terrigenous clastic sediments on a cool water, high energy, mid-latitude shelf: Lacapede, southern Australia. - Sedimentology, 39, 877-903, Oxford
JAMES, N. P., BOREEN, T. D., BONE, Y. & FEARY, D. (1994): Holocene carbonate sedimentation on the west Eucla Shelf, Great Australian Bight: a shaved shelf. - Sedimentary Geol- ogy, 90, 161-177, Amsterdam
JANSSEN, R. (1981): Mollusken-Assoziationen und Biotope im norddeutschen Oberoligoz~n. -Natur und Museum, 111/3, 70- 78, Frankfurt
JeNSEN, A. & FREDERIKSEN, R. (1992): The fauna associated with the

106
bank-forming deepwater coralLopheliapertusa (Scleraetinia) on the Faroe shelf. - Sarsia, 77, 53-69, Bergen
Jom~soN, C. R., Mum, D. G. & R~S~mACH, A. L. (1991a): Characteristic bacteria associated with surfaces of coralline algae: a hypothesis for bacterial induction of marine inverte- brate larvae. - Marine Ecology Progress Series, 74, 281-294, Halstenbek
Jom~soN, C. R., StrrroN, D. C., Ot.soN, R. R. & Gmon~s, R. (199 lb): Settlement of crown-of-thorns starfish: role of bacteria on surfaces of coralline algae and a hypothesis for deepwater recruitment. - Marine Ecology Progress Series, 71, 143-162, Halstenbek
Job,s, B. & DESROCHFAtS, A. (1992): Shallow platform carbonates. - In: WALrr_R, R. G. & JAM~, N. P. (eds.): Facies models - response to sea level change. - 277-301, (Geological Associa- tion of Canada)
KAMP, P. J. J., HAP~Sm'~, F. J., N~sot~, C. S. & BOYLE, S. F. (1988): B amacle-dominated limestone with giant cross-beds in a non- tropical, tide-swept, Pliocene forearc seaway, Hawke's Bay, New Zealand. - Sedimentary Geology, 60, 173-195, Amster- dana
KJ~LMAN, F. R. (1883): The algae of the Arctic Sea. - Stockholm (Kongl. Boktryckeriet)
Kooe, K., Na .w~A~, R. C. & LucAs, M. I. (1982): Biodegradation and carbon flow based on kelp (Ecklonia max/ma) debris in a sandy beach microcosm. - Marine Ecology Progress Series, 7, 315- 326, Halstenbek
KotmoogAs, A. & KOHt~Ar~, D. H. H. (1991): Rasenkorallen als Biotope in der ,~gais. - Naturwiss. Rundschau, 44, 444-445, Stuttgart
~ , B. P. (1980): S toffwechselstrategie und Produktivit~t der grol3en Meeresalgen. -Naturwiss. Rundschau, 33/4, 135-142, Stuttgart
KtrORASS, H.-R. (1974): Experimental study ofnearshore transpor- tation of pebbles with attached algae. - Marine Geology, 16, M9-M12, Amsterdam
K0m.~t~, D. H. H. (1985a): The protection role of coastal forests on coral reefs. - Proc. Fifth Int. Coral Reef Congr. Tahiti, 6, 503-508, Moorea
-- (1985b): Living coral reefs of the world. - 185 pp, New York (Arco)
-- (1988): The sensitivity of coral reefs to environmental pollu- tion. - Ambio, 17, 13-21, Stockholm
-- (1991): Adaptationsmuster hermatypischer Korallen als Grundlage zum Verstandnis der Korallcrlriffgcnese (Kom- pcnsationstheorie). - Mitt. Zool. Nus. Berlin, 67, 209-218, Berlin
K0rmMANN, D. H. H., CHINTII1OGLOU, H., KotrrSOBAS, D. & KOUZOURAS, A. (1991): Korallenriffe im Mittelmeer? - Naturwiss. Rundschau, 44, 316, Stuttgart
LABOR~, J. (1961): Sur un cas particuher de concretiormement animal concretionnement a Cladocora caespitosa L. darts le Golfe deTalante.- Rapp. Comm. Int. MerMedit., 16, 429-432, Paris
LASOR~, J. (1987): Marine biogenic constructions in the Mediter- ranean - a review. - Sci. Rep. Port-Cros natl. Park, France, 13, 97-126, Paris
LaRsor~'Etn~, C., Bot~ssa, P. & Atw~T, J.-P. (1982): The super- ficial sediments of the Enghsh Channel and its Western Ap- proaches. - Sedimentology, 29, 851-864, Oxford
L~ DANOIS, E. (1948): Les profondeurs de lamer. - 303 pp, Paris LEEs, A. (1975): Possible influence of salinity and temperature on
modern shelf carbonate sedimentation. - Marine Geology, 19, 159-198, Amsterdam
-LEEs, A. & Buu.~, A. T. (1972): Modern temperate-water and warm-water shelf carbonate sediments contrasted. - Marine Geology, 13, M67-M73, Amsterdam
LE~aorw, P. (1910): Rbpartition et mode de vie du maerl (Lithothamnium calcareum) aux environs de Concameau (Finist~re). - Annales de 13.nstitut Oceanographique Monaco,
1, 1-28, Paris ~ , T. E., GARLAND, C. D. & M ~ , T. A. (1985): The
bacterial biota on erustose (nonarticulated) coralline algae flom Tasmanian waters. - Microbial Ecology, 11, 221-230, Berlin
Lmn)a~o, D. R. (1976): Marine plant limpets of the northern Pacific: Neogene phylogeny and zoogeography. - Western Society of Malacologists, Annual Report, 9, 22-25, Santa Barbara
~ , M. M., & ~ , D. S. (1985): Deepest known plant life discovered on an uncharted seamount. - Science, 227, 57-59, Washington
I_X~AN, B. W., HARD,O, J. L., Am~, W. M., Wn.t.~Ms, J. D. & SNF_AD, R. G. (1969): Carbonate sediments and reefs, Yucatan Shelf, Mexico. - Am. Assoc. Petrol. Geol., Mere., 11, 5-198, Washington
L0mNo, K. (1990): Seaweeds - Their environment, biogeography, and ecophysiology. - 527 p, New York (Wiley-Interscience)
L ~ o , K. & "tOM Dm~, I. (1990): The distribution and evolution of the Laminariales: North Pacific - Atlantic relationships. - In: GARBARY, D. J. & Sotrm, G. R. (eds.): Evolutionary biogeog- raphy of the marine algae of the North Atlantic. - 187-204, Berlin (Springer)
MARTIN-JEzEQU~t., V. (1983): Facteurs hydrologiques et phytoplancton en bale de Morlaix (Manche Occidentale). - Hydrobiologia, 102, 131-143, Den Haag
MAYR, F. (1953): Durch Tange verfraehtete Ger611e bei Solnhofen und anderwarts. - Geol. Blatter v. NO-Bayern, 3, 113-121, Etlangen
M ~ A , M. C. (1983): Cenozoic paleogeography of North Atlantic land bridges.-In: BoTr, H. P., SAxov, S., TALWA~ M. & TnmoE, J. (eds.): Structure and development of the Green- land-Scotland ridge. - 351-399, New York (Plenum Press)
~ , N., Em.~umtrs~, H., l(n, r~crL~, J. S. & BmZO~R, W. H. (1982): Norwegian corals: radiocarbon and stable isotopes in Lophelia pertusa.- Boreas, 11, 163-171, Oslo
Mn.,.miAN, J. D. (1977): Role of calcareous algae in Atlantic continental margin sedimentation.- In: FLOat, E. (ed.): Fossil Algae. - 232-247, Berlin (Springer)
M6Rr,~, N.-A. (1979): The Fennoscandianuplift and Late Cenozoic geodynamics: geological evidence. - GeoJournal, 3/3, 287- 318, Dordrecht
Moru~, C., t~R~O, C., B ~ o n , N. & SASSARn'n, M. (1994): Present-day bioconstructious of the hard coral, Cladocora caespitosa (L.) (Anthozoa, Scleractinia), in the eastemLigurian Sea (NW Mediterranean).-Biol. Mar. Medit., 1,371-372, San Remo
MORSE, A. N. C. (1991): How do planktonic larvae know where to settle?- American Scientist, 79/2, 154-167, New Haven
Mouss^v~,N, E. (1992): On Cretaceous bioconstructions: Compo- sition and evolutionary trends of crust-building associations, - Facies, 26, 117-144, Erlangen
MUSCATt~, L. & PoRa'~, J. W. (1977): Reef corals: mutualistic symbioses adapted to nutrient-poor environments.-Bioscience, 27, 454-460, Arlington
M ~ t ~ , J. J. ( 1989): Geomela-ic simulation and mapping of Holocene relative sea-level changes in northern Norway. - Journal of Coastal Research, 5/3,403-417, Fort Lauderdale
NEa~sioc, J. H. (1989): Temperate water carbonate facies of the Early Miocene Paratethys (Zogelsdorf Formation, Lower Aus- tria). - Facies, 21, 11-40, Erlangen
NEt.SON, C. S. (1988): An introductory perspective on non-tropical shelf carbonates. - Sedimentary Geology, 60: 3-12, Amster- dam
N~soN, C. S. & BORr~OLt~, B. D. (1983): Temperate skeletal carbonate sediments on Scott Shelf, northwestern Vancouver Island, Canada. - Marine Geology, 52, 241-266, Amsterdam
NELSON, C. S., HANCOCK, G. E. & KAx~, P. J. J. (1982): Shelf to basin, temperate skeletal carbonate sediments, Three Kings Plateau, New Zealand. - J. Sed. Petrol., 52, 717-732, Tulsa

107
NFJ~SON, C. S., HYDFJ,~, F. M., KEAr~ S. L., I~nSK, W. L. & GOPJ)ON, D. P. (1988a): Application of zoarial growth-form studies in facies analysis of non-tropical carbonate deposits in New Zealand.- Sedimentary Geology, 60, 301-322, Amsterdam
NFJ..SON, C. S., K E ~ , S. L. & HEAD, P. S. (1988b): Non-tropical carbonate deposits on the modem New Zealand shelf. - Sedi- mentary Geology, 60, 71-94, Amsterdam
PXr'ZOLD, J., RISTO.OT, H. & W ~ , G. (1987): Rate of growth and longevity of a large colony of Pentaporafoliacea (Bryozoa) recorded in their oxygen isotope profdes. - Marine Biology, 96, 535-538, Amsterdam
P~l~_.rt, B. C. & DAWSON, E. Y. (1965): Non-calcareous marine algae from California Miocene deposits. -Nova Hedwigia, 10, 273-295, Stuttgart
PE~CE, C. M. & ScrmmtaNG, R. E. (1990): Induction of metamor- phosis of larvae of the green sea urchin Strongylocentrotus droebachiensis, by coralline red algae. - Biol. Bull., 179, 304- 311, Lancaster
IhRnZOt~, P. A. (1991): World Atlas of Holocene sea-level changes. - 300 pp., Amsterdam (Elsevier)
Polznv, C. (1970): Hydrodynamisme et s&limentation dans le gol fe de Gab~s frunisie). - Tethys, 2/1,267-295
Porto, P., FLOC'H, L Y., AUGRtS, C. & C ~ o c n , J. (1990): Annual drowthrateofthecalcareousredalgaLithothamnioncorallioides (Corallinales, Rhodophyta) in the Bay of Brest, France. - Hydrobiologia. 204/205, 263-267, Den Haag
Pt~sc~a., C. M. (1992): An ultrastructural survey of the diversity of crystalline, proteinaceous inclusions in red algal cells. - Phycologia, 21/6, 489-499, Oxford
Rno, C. P. (1981): Criteria for recognition of cold-water carbonate sedimentation: Berriedale Limestone (Lower Permian), Tas- mania, Australia. - J. Sed. Petrol., 51/2, 441-506, Tulsa
RAO, C. P. ( 1988): Oxygen and carbon isotope composition of cold- water Berriedale Limestone (Lower Permian), Tasmania, Aus- tralia. - Sedimentary Geology, 60, 221-231, Amsterdam
RFJss, Z. & Ho'rr~om, L. (1984): The Gulf of Aqaba. Ecological micropalaeontology. - 345 pp, Berlin (Springer)
REY, F. & LonG, H. (1985): The influence of ice and hydrographic conditions on the development ofphytoplankton in the Barents Sea. - In: G~AV, 1. S. & C ~ J s ' ~ s ~ , M. E. (eds.): Marine biology of polar regions and effects of stress on marine organ- isms. - 49-63 (Wiley)
R o o ~ s , J. (1957): The distribution of marine carbonate sediments: a review. - S o c . Econ. Paleont. Miner., Spec. Pub., 5, 2-13, Tulsa
RODmOtJEz-P~A, A. (1984): La Formacion calcarenitica de Sant Elm: un ejemplo de plataforma mixta terrigeno-carbonatada. - Publ. Geol. Univ. Autonoma de Barcelona: 399-417, Barce- lona
ROKO~O~,~, K., Rise, L., B UC,6E, T. & S~'rmM, J. (1988): Bedrock geology on the Mid Norwegian continental shelf, Map in scale 1:1000000. - IKU-Report, 118, Trondheim
S ~ , M. (1973): Quantitative Daten tiber benthische Kar- bonatsedimentation in mitfleren Breiten. - Ver6ffenfl. Univ. Innsbruck, Festschrift Heil3el, 86: 267-279, Innsbruck
ScHh~r~, P. (1994): Growth strategies of arctic bryozoa in the Nordic seas. - In: HnYwnim, P. J., R Y L ~ , J. S. & T ~̂O~OR P. D. (eds.): Biology and paleobiology of bryozoans.- 173-176, Fredensborg (Olsen & Olsen)
SCH.Z, FER, P., A m ) g ~ , H., FRF..IWALD, A. & M ~ , A. (1991): Boreale Flachwasserkarbonate. - In: G~J~ACH, S. A. & GR~, G. (eds.): Europaisches Nordmecr, Reise Nr. 13. - Meteor- Berichte, 91-2, 23-35, Hamburg
SCHLAOE~, W. (1989): Drowning unconformities on carbonate platforms. - In: CREV~LLO, P. D., Wn.so~, J. L., SA~, J. F., READ, J. F.(eds.): Controls on carbonate platform and basin development. - Soc. Econ. Paleont. Miner., Spec. Pub., 44: 15- 25, Tulsa
SC~AnO~, S. O. & Ko~sm, K. (1975): The geographic boundary between the coral-algal andthe bryozoan-algallimestone facies:
A paleolatitude indicator.-IXInt. Congress of Sedimentology, 187-190, Nice
S c r ~ a o ~ , L. J. G. (1974): Lain Precambrian mixtites: glacial and/or nonglacial? American Journal of Science, 274, 673- 824, New Haven
Scm,~rr, H. & Fm~wntJ~, A. (1993): Rezente gesteinsbohrende Kleinorganismen des norwegischen Schelfs. - Natur mad Mu- seum, 123/5, 149-155, Frankfurt
S c o t ~ , T. P. (1988): The environments of production and depo- sition of calcareous sediments on the shelf west of Scotland. - Sedimentary Geology, 60, 107-124, Amsterdam
S ~ , T. P. & BowEs, G. E. (1988): The facies distribution of carbonate sediments on Porcupine Bank, northeast Atlantic.- Sedimentary Geology, 60, 125-134, Amsterdam
Sco~-~, T. P., ~ B R S S O N , E. T., BowEs, G. E., CLoKm, J. J., FARROW, G. E. & Mn w Jr, tAN, J. D. (1980): Recent, temperate, sub-photic, carbonate sedimentation: Rockall Bank, northeast Atlantic. - J. Sed. Petrol., $0/2, 331-356, Tulsa
S ~ , B. (1984): Str6mungs- und S edimentationsexperimente mit einem Schill aus dem hrmelkanal. - Unpubl. Dissertation, Geol. Inst. Univ. Marburg
SHEATH, R. G., Hm.t.~trST, J. A. & SAwn, T. (1981): Ultrastructure of the floridean starch granule. - Phycologia 20/3, 292-297, Oxford
Smzuo, T., D o ~ r ~ , D. P. & Snrro, T. (1986): Drusisirendewana n. sp. (Mammalia: Sirenia), a new ancestor of St ream's sea cow from the Upper Miocene of Yamagata prefoctare, north- eastern Japan. - Trans. Proc. Palaeont. Soc. Japan, 141, 293- 321, Tokio
SatnaWAY, G. A. (1953): Rafted pebbles from the deep ocean off Baja California. - J. Sed. Pet. 23/2, 24-33, Tulsa
SiMor, m, L. & CARAr~arCm, G. (1988): The fate of foramol ("tem- perate-Type") carbonate platforms. - Sedimentary Geology, 60, 341-354, Amsterdam
S~OLDnL, H. R., HASS~, A., Rmr F. & Logo , H. (1987): Spring phytoplankton development and zooplankton reproduction in the central B arents Sea in the period of 1979-1984. - In: LonG, H. (ed.): The effect o f oceanographic conditions on distribution and population dynamics of commercial fish stocks in the B a.rents Sea. - Proc. of the 3rd Soviet-Norwegian Symposium
S~vmTACEK, V. (1991 ): Die Prim~trproduktion der marinen Plankton- Algen. - Spektrum der Wissenschaft, 12/1991, 52-63, Heidelberg
S~XH, A. M. (1988): Preliminary steps toward fomaation of a generalized budget for cold-water carbonates. - Sedimentary Geology, 60, 323-331, Amsterdam
SMrrH, A. M. & NELSON, C. S. (1994a): Calcification rates of rapidly colonising bryozoans in Hauraki Gulf, northern New Zealand. - New Zealand Journal of Marine and Freshwater Research, 28, 227-234, Wellington
S~lrrH, A. M. & NELSON, C. S. (1994b): Selectivity in sea-floor processes: taphonomy of bryozoans. - I n : I-IAYWARD, P. J., RYLANO, J. S. & TAYLOR P. D. (eds.), Biology and paleobiology of bryozoans.- 177-180, Fredensborg (Olsen & Olsen)
SMrm, A. M., NELSON, C. S. & D ~ P. J. (1992): Dissolution behaviour of bryozoan sediments: taphonomic implications of nontropical shelf carbonates. - Palaeogeography, Palaeo- climatology, Paleaoecology, 93, 213-226, Amsterdam
SOOT-RYEN, T. (1934): Hydrographical investigations in theTrorns~ District 1930. -Tromsr Mus./~krshefter, 52, 1-78, Troms0
STY, W. T., Bor, P. V. M., Bo~J~-Bos, S. A., VAN Roon, J. M. & VAN DEN HoE~ C. (1988): Single-copy DNA-DNA hybrid- izations among five species of Laminaria (Phaenphyceae): phylogenetic and biogeographic implications. - Helgol. Meeresunters., 42, 251-267, Hamburg
STAm.~, G. D. (1979): Triassic carbonate buildups of western North America: comparisons with the alpine Triassic of Eu- rope. - Riv. Italiana di Paleontologia, 85, 877-894, Milano
-- (1981): Early history of scleractinian corals and its geological consequences.- Geology, 9, 507-511, Boulder

108
ST~EY, G. D. & C~aRr~s, S. D. (1988): Constructional azooxanthellate coral communities: an overview with implica- tions for the fossil record. - Palaios, 3, 233-242, Tulsa
STmmCr,, R. S. (1985): Adaptations of crustose coralline algae to herbivory: Patterns in space and time. - In: Toor~mY, D. F. & Nrmcm, M. H. (eds.): Paleoalgology:. - 352-366, Berlin (Springer)
SUNDBY, S. (1984): Influence of bottom topography on the circula- tion at the continental shelfoffnorthem Norway.- Fiskdir. Skr. Ser. Havunder., 17, 501-519, Bergen
S~I.EN, O. H. (1950): The hydrography of some fjords in northern Norway.- Troms~ Museums Arshefter, 70/1, 1-101, Tromsr
Tav-m~ M., RIND, D. E. & Am)E~SON, J. B. (1993): Skeletal and isotopic composition and paleoclimatic significance of Late Pleistocene carbonates, Ross Sea, Antarctica. J. Sed. Pet., 63/ 1, 84-90, Tulsa
Tmcm~T, C. (1958): Cold- and deep-water coral banks. - Ameri- can Association of Petroleum Geologists, Bulletin, 42/5, 1064- 1082, Tulsa
TnoMsm~, E. & V o w , T. O. (1986): Macrofaunal palaeoecology and stratigraphy in Late Quaternary shelf sediments off north- ern Norway. - Palaeogeography, Palaeoclimatology, Paleaoecology, 56, 103-150, Amsterdam
VAN DOa~ L ~ , J. D. & Ho~Ewr~, P. (1992): Waves of crown-of- thorns starfish outbreaks - where do they come from? - Coral Reefs, 11, 207-213, Berlin
V~a~ WAOO~mR, N. A., MuDm, P. J., COLE, F. E. & D ~ o w , G. (1989): Siliceous sponge communities, biological zonation, and recent sea-level change on the Arctic margin: Ice island results. - Can. J. Earth Sci., 26, 2341-2355, Montreal
VOIOT, E. (1966): Die Erhaltung verg~h-aglicher Organismen durch Abformung infolge Inkrustation dutch sessile Tiere. - N. Jb. Geol. Pal,~iont. Abh., 125, 401-422, Stuttgart
-- (1979): The preservation of slightly or non-calcified fossil bryozoa (Ctenostomata and Cheilostomata) by bioimmuration. - In: L~wooD, G. P. & AB~'rr, M. B. (eds.): Advances in Bryozoology:.- 541-564, London (Academic Press)
V o ~ , T . O. & Fa.vsaoeo, A. (1979): Late Weichselian deglaciation and paleoertvironment of the shelf and coastal areas of Troms, north Norway - a review. - Boreas, 8, 247-253, Oslo
W ~ t ~ w , B. R. & Cota_~soN, J. W. (1986): Paleontology and deposition of the Phosphoria Formation. - Conlxibutions to Geology, Univ. of Wyoming, 24, 107-142, Wyoming
W~ss, R. E., Corcso~y, J. R. & MACnCIa~E, R. J. (1970): Bryozoan carbonate sand continuous along southern Australia_ - Marine Geology, 9, 63-73, Amsterdam
WAsss~a~, P., PF.n,r~T, R. & SI~m'rACF.~ V. (1991): Patterns of reproduction and sedimentation in the Norwegian co as tal zone, the Barents Sea and Norwegian Sea. - In: W^SSMh~r P., I h ~ r ~ a ~ , A. S. & Lgr O. (eds.): Sediment trap studies in the nordic countries, 2. - 137-156, Nurmijarvi (Nurmiprint)
WATm~rBr, JtO, H. (1933): U"ber die Tritrationsalkalimt~it und den Kalzium-Karbonatgehalt des Meerwassers. - Wiss. Ergebn. Deutseh. Atlantisch. Exped. Meteor 1925-1927, 8, 122-231, Hamburg
W ~ , G., B~T:zs~, W., vor~ Bom~onN, B. & StrEss, E. (1987): Biogenic carbonates in temperate and subtropical environ- merits: Production and accumulation, saturation state and sta- ble isotope composition. - In: RtrMOrm, J., W ~ E g , E. & Zawgscm~, B. (eds.): S eawater-Sediment Interactions in Coastal Waters. - 263-302, Berlin (Springer)
WmmM~r, A. (1994): Karbonatproduktion inder B aiede Morlaix, Bretagne, Frankreich (Arbeitstitel).-Ph.D.Thesis, University of Marburg
Wn..soN, J. B. (1979a): The distribution of the coral Lophelia pertusa (L.) [L. prolifera (Pallas)] in the northeast Atlantic. - J. Mar. Biol. Ass., U. K., 59, 149-164, Plymouth
-- (1979b ): "Patch" development of the deep-water coral Lophelia pertusa (L.) on Rockall Bank. - J. Mar. Biol. Ass., U. K., 59, 165-177, Plymouth
-- (1979c): Biogenic carbonate sediments on the Scottish conti- nental shelf and on Rockall Bank. - Marine Geology, 33, M85- M93, Amsterdam
-- (1988): A model for temporal changes in the faunal composition of shell gravels during a transgression on the continental shelf around the British Isles. - Sedimentary Geology, 60, 95-105, Amsterdam
WOOD, R. (1993): Nutrients, predation and the history of reef- building. - Palaios, 8, 526-543, Tulsa
WOODBORWa, M. W., Room,s, J. & J h v ~ , N. (1989): The geologi- cal significance of kelp-rafted rock along the west coast of South Africa. -Geo-Marine Letters, 9, 109-118, Stroundsburg
YotrNo, H. R. & Nat.soN, C. S. ( 1988 ): Endolithic biodegradation o f cool-water skeletal carbonates on Scott shelf, northwestern Vancouver Island, Canada. - Sedimentary Geology, 60, 251- 267; Amsterdam
Zhma., H. (1971): Upper Triassic carbonate facies in the northern Limestone Alps. - In: Sedimentology of parts of central Eu- rope, 8th Sedimentological Congress, Heidelberg, Guidebook: 147-185, Heidelberg
Manuscript received May 15, 1994 Revised manuscript accepted December 10, 1994




















