
Profiling and annotation of human kidney glomerulus proteome
Upload
independentCategory
view
3download
0
RESEARCH ARTICLE
Comparison of two-dimensional gel electrophoresis
based and shotgun strategies in the study of plasma
membrane proteome
Jintang He, Yashu Liu, Sizhi He, Qingsong Wang, Hai Pu, Yuanpeng Tongand Jianguo Ji
Department of Biochemistry and Molecular Biology, College of Life Sciences, National Laboratoryof Protein Engineering and Plant Genetic Engineering, Peking University, Beijing, P. R. China
Membrane proteins play important roles in various plasma membrane (PM) activities such assignal transduction and cell recognition. However, a comprehensive proteomic study of mem-brane proteins remains difficult. Different strategies have been employed to study PM proteome,but little effort has been made to systematically evaluate them. In the present work, liver PM wasprepared by subcellular fractionation and an aliquot was washed by sodium carbonate. Afterevaluation of the PM fraction by electron microscopy and Western blotting, proteins in bothoriginal and carbonate washed PM were identified by either 2-DE coupled MALDI-TOF-MS orshotgun strategies. Then protein characteristics such as molecular weight, pI, grand averagehydrophobicity, subcellular location, and transmembrane domains were systematically com-pared. The comparative analysis showed that shotgun strategies were more suitable to identifymembrane proteins, while 2-DE-based strategies may serve as a complement. Furthermore, car-bonate washing obviously enriched the integral membrane proteins. All the results suggestedthat the strategy combining carbonate washing and shotgun identification was the optimumstrategy to study human liver PM proteome. Using this strategy, 260 high-confidence proteinswere identified, wherein 139 were integral membrane proteins which had 1–17 transmembranedomains.
Received: January 3, 2006Revised: August 22, 2006
Accepted: November 3, 2006
Keywords:
Human normal liver / Plasma membrane / Shotgun / Two-dimensional gel electro-phoresis
Proteomics Clin. Appl. 2007, 1, 231–241 231
1 Introduction
The plasma membrane (PM) is a crucial cellular componentwhose function involves controlling cell–cell recognition,transducing extracellular signals to regulate internal cellularactivity, transporting materials around the cell, and so on.The PM is composed of two major components, namely,membrane proteins and membrane lipids, wherein mem-brane proteins enable the membrane to carry out its dis-tinctive activities. Membrane proteins play a critical roleespecially in drug discovery. As reported by Hopkins andGroom [1], most of the drug targets are membrane proteins
Correspondence: Professor Jianguo Ji, P. O. Box 31, College ofLife Sciences, Peking University, Beijing 100871, P. R. ChinaE-mail: [email protected]: 186-10-62751526
Abbreviations: ASB-IL, amidosulfobetaine; EPM_2-DE, study ofsodium carbonate washed plasma membrane by 2-DE coupledMALDI-TOF-MS; EPM_Shotgun, study of sodium carbonatewashed plasma membrane by RP-HPLC MS/MS; GRAVY, grandaverage hydrophobicity; HB, homogenization buffer; PM, plasmamembrane; PM_2-DE, study of original PM fraction by 2-DEcoupled MALDI-TOF-MS; PM_Shotgun, study of original PM byRP-HPLC MS/MS; TMHs, transmembrane helices
DOI 10.1002/prca.200600004
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.clinical.proteomics-journal.com

232 J. He et al. Proteomics Clin. Appl. 2007, 1, 231–241
and about 25% of all current experimental and marketeddrugs target class-1 and -2 G-protein coupled receptors.Thus, it is important to comprehensively study this class ofproteins.
However, a large-scale proteomic study of membraneproteins remains very difficult. According to Wallin and VonHeijne [2], membrane proteins account for approximately30% of total proteins, but they are greatly under-representedin some existing analyses [3, 4]. Different strategies such as2-D gel-based strategies and shotgun strategies have beenwidely employed to carry out this kind of study. 2-D gel-basedstrategies are notorious for the under-representation ofmembrane proteins, and the most important reason is thatmembrane proteins are poorly soluble in the aqueous mediaused for IEF [3, 5–7]. Recently, there have been some refine-ments, most of which have focused on improving membraneprotein solubility. Myristic amidosulfobetaine (ASB-14) is anencouraging zwitterionic detergent being able to solubilizeintegral membrane proteins previously undetected on 2-Dgels [8]. In another research, ASB-14 was better than CHAPSat solubilizing a G-protein coupled receptor [9]. Zhang et al.[10] applied an optimized lysis buffer containing urea, thio-urea, CHAPS, and NP-40. Combining 2-DE-MALDI-TOFand 1-DE-ESI-Q-TOF, they identified 88 integral membraneproteins with one to seven transmembrane domains frommouse liver PM. They also addressed the fact that 2-DE wasstill the core technology to study membrane proteomes dueto its excellent resolving power. Shotgun strategies provide apowerful alternative to 2-D gel-based strategies. Proteins arein-solution solubilized followed by enzymatic digestion andanalysis of the peptide mixture by LC MS/MS. Yates andcoworkers [6, 7] demonstrated that shotgun methods had thepotential to identify a large number of membrane proteinsand their modifications. Using the shotgun strategy, 268integral membrane proteins were identified from Deino-coccus radiodurans by Blonder et al. [11], and 65 integralmembrane proteins were identified from human epidermalsheets in another research [12]. Recently, there have beensome significant advances in PM proteomics. Using SDS-PAGE separation and LC MS/MS, Zhao et al. [13] identified898 unique proteins, of which 781 were annotated withregard to their subcellular location, and at least 526 wereintegral PM proteins. Using a novel method for PM fractionand shotgun identification, Nielsen et al. [14] identified 862PM proteins from mouse brain cortex and 1685 PM proteinsfrom mouse hippocampus. The PM proteomic analysis hasbeen recently applied to disease therapy [15, 16]. Oh et al. [15]used a three-prolonged approach to resolve proteins/pep-tides for mass spectrometric analysis and identified 2000proteins from the PMs of rat lung endothelial cells, includ-ing two promising tissue-specific endothelial cell surfaceproteins.
Membrane purity is a key factor affecting the results ofidentification. Therefore, great effort has recently beenfocused on membrane preparation. Integral membrane pro-teins can be enriched by carbonate washing. It is reported
[17] that sodium carbonate washing remarkably enrichedintegral ER membrane proteins. The method was alsoproved useful for studying the integral membrane proteinsof rat liver PM [18]. As described in this paper, the enrich-ment of enzyme markers in the starting PM was 30- to 50-fold relative to the homogenate, however, this valueincreased to about 40- to 70-fold after sodium carbonatewashing. Recently, carbonate washing has been graduallyapplied to some proteomic studies to enrich integral mem-brane proteins [11–13].
To the best of our knowledge, little effort has been madeto systematically evaluate the 2-DE-based and shotgun strat-egies in PM proteomics. The main purpose of the presentwork was to provide an optimum strategy to study PM pro-teome, with a view of helping to elucidate the function of thehuman liver PM. We evaluated four strategies: study of orig-inal PM fraction by 2-DE coupled MALDI-TOF-MS (PM_2-DE) or RP-HPLC MS/MS (PM_Shotgun), and study ofsodium carbonate washed PM by 2-DE coupled MALDI-TOF-MS (EPM_2-DE) or RP-HPLC MS/MS (EPM_Shot-gun). Protein identification capability and protein character-istics such as molecular weight, pI value, grand averagehydrophobicity (GRAVY), subcellular location, and trans-membrane domains were compared. As a result, the shot-gun strategies identified much more large proteins, basicproteins, hydrophobic proteins, PM located proteins, andtransmembrane proteins than 2-DE-based strategies. Fur-thermore, sodium carbonate washing obviously enrichedPM proteins, especially integral membrane proteins. After asystematic comparison, EPM_Shotgun was considered thebest strategy to study the PM proteome, and 2-DE-basedstrategies may serve as a complement.
2 Materials and methods
2.1 Materials
IPG DryStrips, IPG buffer, and Cover fluid were obtainedfrom GE Healthcare (Uppsala, Sweden). Sucrose, EDTA,HEPES, PMSF, acrylamide, bis-acrylamide, ammoniumpersulfate, urea, glycine, tris, SDS, and CBB R-250 wereobtained from Amresco (Solon, OH, USA). Thiourea andTEMED were obtained from Acros Organics (NJ, USA).Agarose was obtained from Bio Basic (Markham, Ontario,Canada). ASB-14 was obtained from Calbiochem (SanDiego, CA, USA). TFA, DTT, and iodoacetamide were pur-chased from Merck (Darmstadt, Germany). Anti-flotillin-1polyclonal antibody was purchased from Chemicon (Teme-cula, CA, USA). Antibody to the a1 subunit of Na1/K1
ATPase was obtained from Upstate Biotechnology (LakePlacid, NY, USA). Peroxidase-conjugated goat anti-rabbit,and anti-mouse IgG (H 1 L) were purchased from JacksonImmunoResearch (West Grove, PA, USA). 3,30-Diamino-benzidine (DAB) stain was purchased from Sino-AmericanBiotechnology (Beijing, P. R. China). ACN (HPLC grade)
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.clinical.proteomics-journal.com

Proteomics Clin. Appl. 2007, 1, 231–241 233
was purchased from Fisher (NJ, USA). Trypsin (sequencinggrade) was purchased from Promega (WI, USA). CHCA,peptide calibration standard for MS, and AnchorChip™ 600/384 were purchased from Bruker Daltonik (Bremen, Ger-many). Human normal liver tissues were provided byHLPP.
2.2 Preparation of human normal liver PM
To prepare a PM, subcellular fractionation of human liverswas performed according to the procedure of Hubbard et al.[19] and Fleischer and Kervina [20] with some modifications.Briefly, after removing the connective tissues, human liverswere placed in ice-cold homogenization buffer (HB) (0.25 Msucrose, 10 mM HEPES, 1 mM PMSF, pH 7.5). After min-cing with scissors, washing twice to remove blood and add-ing five volumes of HB, the livers were homogenized in anULTRA-TURRAX T8 homogenizer (IKA, Germany). Thehomogenate was filtered through four layers of nylon gauzeand the filtrate was centrifuged at 10006g for 10 min in aHimac CR21 centrifuge. The resulting pellets (residue 1)were pooled and resuspended in HB. Then high densitysucrose (2.4 M sucrose, 10 mM HEPES, and 1 mM PMSF,pH 7.5) was added to a final concentration of 1.6 M (45.7%w/w sucrose). Twenty five milliliters of the suspended resi-due 1 was transferred into each of six SW32 rotor tubes, andthe suspensions were overlaid with about 13 mL of HB to fillthe tube and were then centrifuged at 70 9006g (Rav) for70 min in a Beckman Optima™ L-80 XP ultracentrifuge. Theband at the interface was collected, and then resuspendedgently in two volumes of cold water and a sufficient amountof HB. The suspension was centrifuged at 12006g for10 min. The pellets (residue 2) were resuspended in HB, andhigh density sucrose was then added to bring the final con-centration to 1.45 M (42% w/w sucrose). The suspendedresidue 2 was transferred into each of six SW32 rotor tubes,and then overlaid with enough HB to fill the tubes. Aftercentrifugation at 68 4006g (at Rav) for 60 min, the pellicle atthe interface was collected and resuspended in HB, and thencentrifuged at 17 6006g for 10 min. The pellets (residue 3)were resuspended in HB, and high density sucrose was thenadded to bring the final concentration to 1.35 M (39.4% w/wsucrose). The suspended residue 3 was transferred into eachof six P40ST tubes, and then overlaid with enough HB to fillthe tubes. The tubes were centrifuged at 231 0006g for60 min in a Himac CP 100MX ultracentrifuge. The purifiedPM was collected from the interface, resuspended in enoughHB, and centrifuged at 14 5006g.
2.3 Electron microscopy
Equal volumes of a sample and 5% glutaraldehyde in PBSwere mixed, incubated overnight at 47C, and centrifuged.The resulting pellet was postfixed in OsO4, and then de-hydrated through a graded series of ethanol and acetone. Thepellet was embedded in Epon, and thin sections were stained
with uranyl acetate and then with lead citrate, and observedunder a transmission electron microscope (JEM-1010; JEOL,Tokyo, Japan) at 80 kV.
2.4 Western blotting
Proteins of liver homogenate and PM fraction were sepa-rated by SDS-PAGE and then transferred to PVDF (PVDF)membranes (BioRad, USA). After being blocked for 2 h, themembranes were incubated with antibodies against flotillin-1 and Na1/K1 ATPase (a1 subunit) overnight. After washingthree times, the membranes were incubated with peroxidase-conjugated goat anti-rabbit or anti-mouse IgG (H 1 L) for1 h. The blots were visualized with DAB stain.
2.5 Sodium carbonate washing
PM was washed by sodium carbonate according to Fujiki etal. [17]. In brief, PM fractions were diluted 200 times with0.1 M sodium carbonate, and incubated on ice for 30 min,and then centrifuged in a Hitachi 55P-72 ultracentrifuge for1 h at 40 000 rpm. The pellet was stored at 2807C until thefollowing analysis.
2.6 2-DE
2-DE was performed as previously described [21, 22] withsome modifications. Briefly, PM was solubilized for IEFusing a lysis buffer containing 7 M urea, 2 M thiourea, 2%ASB-14, and 60 mM DTT at room temperature for 5 h, andreplenished with 0.5% ampholytes before IEF. Samples wereloaded on pH 4–7 or 3–10 IPG DryStrips. IEF was conductedusing the IPGphor™ II (GE Healthcare) according to the fol-lowing procedure: 30 V68 h, 50 V64 h, 300 V61 h,1000 V61 h, 3000 V61 h, 5000 V61 h, and 8000 V for atotal of 50 000 V?h. The second dimension was performed ina 10% SDS polyacrylamide gel. The gels were then stainedwith CBB R-250, and scanned at 300 dpi resolution. Proteinspots were analyzed with ImageMaster Platinum™ software(GE Healthcare).
2.7 In-gel protein digestion
In-gel protein digestion was performed essentially as de-scribed by Yang et al. [23]. Briefly, protein spots were excisedwith scissor-sheared pipette tips, and then transferred to V-bottom 96-well microplates which were filled with 100 mL of50% v/v ACN and 25 mM ammonium bicarbonate solutionper well in advance. After being destained for 1 h, the gel pie-ces were dehydrated with 100 mL of 100% ACN for 20 min andthen evaporated to dryness in a SpeedVac concentrator(Thermo Savant, USA) for 30 min. About 2 mL of 5 mg/mLtrypsin solubilized in 25 mM ammonium bicarbonate wasadded to each well to rehydrate the gel pieces. The 96-wellplates were then tightly sealed with Parafilm. After reswellingat 47C for 45 min, the plates were incubated at 377C for 10 h.
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.clinical.proteomics-journal.com

234 J. He et al. Proteomics Clin. Appl. 2007, 1, 231–241
Then 8 mL of the extraction solution consisting of 0.25% TFAand 5 mM n-octyl-b-D-glucopyranoside was added to each welland shaken at 377C for 1 h to extract the peptides.
2.8 MALDI-TOF-MS
Thin layers of CHCA crystals were prepared according toGobom et al. [24]. In brief, saturated CHCA in a solutioncontaining 97% acetone and 0.003% TFA was drawn with anEppendorf tip across the surface of AnchorChip 600/384 toform a thin layer of crystalline CHCA. Four microliters ofeach sample solution was deposited onto the CHCA matrixlayer. After 3 min incubation, the remaining liquid wasremoved with tips, and then each sample was washed with6 mL of a solution containing ice-cold 10 mM ammoniumcitrate and 0.3% TFA. One microliter of 0.1 pmol/mL stand-ard peptide mixture was used and external mass calibrationwas also performed as above. Mass spectra of positivelycharged ions were recorded on an Ultraflex MALDI-TOF/TOF mass spectrometer (Bruker Daltonik) under the controlof FlexControl™ 2.2 software (Bruker Daltonik) operated inthe positive ion reflector mode. The ion acceleration voltagewas 25 kV. After processing by FlexAnalysis™ 2.2 (BrukerDaltonik) and Biotool™ 2.2 (Bruker Daltonik) softwares pro-vided by the manufacturers, the MS spectra were auto-matically searched against the IPI database (IPI.HU-MAN.v3.07) [25] using the MASCOT software (Matrix Sci-ence, London, UK). The main parameters were set asfollows: mass range from 800 to 4000 Da; S/N �3.0; fixedmodification: carbamidomethyl (C); variable modification:oxidation (M); maximum number of missing cleavages: 1;MS tolerance 6 100 ppm. All accepted results had a MAS-COT score of at least 69 (p,0.005), whereas protein scoresgreater than 59 were normally significant (p,0.05).
2.9 Trypsin digestion of protein mixtures
The PM fraction was digested as described by Blonder et al.[11] with some modifications. In brief, the PM was resus-pended in 50 mM ammonium bicarbonate (pH 8.0) viaintermittent vortexing and sonication. The proteins werethermally denatured at 907C, for 20 min and transferred toice-cold water, and the protein concentration was determinedby the CBB method [26]. Tri(b-chloroethyl)phosphate (5 mM)was added to an aliquot of the sample containing 300 mg ofproteins and incubated at room temperature for 30 min.Then 1 M iodoacetamide was added to a final concentrationof 25 mM and incubated in the dark at room temperature for30 min. The sample was then diluted with methanol to pro-duce a composition of 60% organic solvent. The PM proteinswere extracted and solubilized by performing intermittentvortexing and sonication. Tryptic digestion was immediatelyperformed for 6 h at 377C using a 1:30 w/w trypsin-to-proteinratio. The sample was stored at 2807C until RP-HPLC MS/MS analysis.
2.10 RP-HPLC MS/MS analysis
RP-HPLC MS/MS was performed using an LTQ linear 1Tmass spectrometer (Thermo, San Jose, CA, USA). The sys-tem was fitted with a C18 RP column. Mobile phase A (0.1%formic acid in water) and mobile phase B (0.1 formic acid inACN) were selected. The peptide mixtures were eluted usinga gradient of 2–98% B over 240 min. MS/MS spectra wererecorded on an LTQ linear IT mass spectrometer, and thenautomatically searched against IPI database (IPI.HUMAN.v3.07) [25] using SEQUEST program as described byWashburn et al. [27]. Briefly, all accepted results had a DCn ofat least 0.1. A single charged peptide must be tryptic, and thecrosscorrelation score (Xcorr) had to be at least 1.9. Tryptic orpartially tryptic peptides with a charge state of 12 must havean Xcorr of at least 2.2. Triply charged tryptic or partiallytryptic peptides with a 13 charge state were accepted if theXcorr was �3.75.
2.11 Bioinformatics annotation tools
The GRAVY values were calculated according to Kyte andDoolittle [28]. WoLF-PSORT (http://wolfpsort.seq.cbrc.jp/)was used to predict the subcellular protein location.TMHMM (http://www.cbs.dtu.dk/services/TMHMM-2.0/)[29] was used to predict the protein transmembrane domains.
3 Results and discussion
3.1 Evaluation of the PM fraction
3.1.1 Electron microscopy
The quality of our PM fraction was morphologically eval-uated by electron microscopy (Fig. 1). We could clearlyobserve some nuclei, mitochondria, membrane sheets, andvesicles in the liver homogenate (Fig. 1A). However, the pre-dominant structures in the PM fraction were membranesheets (Fig. 1B), with almost all intact nuclei having dis-appeared. Some occasional mitochondria could also be seenin the fraction. These observations indicated that PM wasgreatly enriched in our PM fraction. Meanwhile, the majorcontaminants were evidently removed.
3.1.2 Western blotting
The quality of the PM fraction was also examined by Westernblotting (Fig. 2). We compared the liver homogenate with thePM fraction using antibodies against flotillin-1 and Na1/K1
ATPase (a1 subunit). Flotillin-1 and Na1/K1 ATPase are PMmarker proteins. Flotillin-1 is a PM associated protein ofcaveolae, and Na1/K1 ATPase is a multitransmembraneprotein. As shown in Fig. 2, both of the proteins were greatlyenriched in the PM fraction, which implied that the PM wassignificantly enriched.
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.clinical.proteomics-journal.com
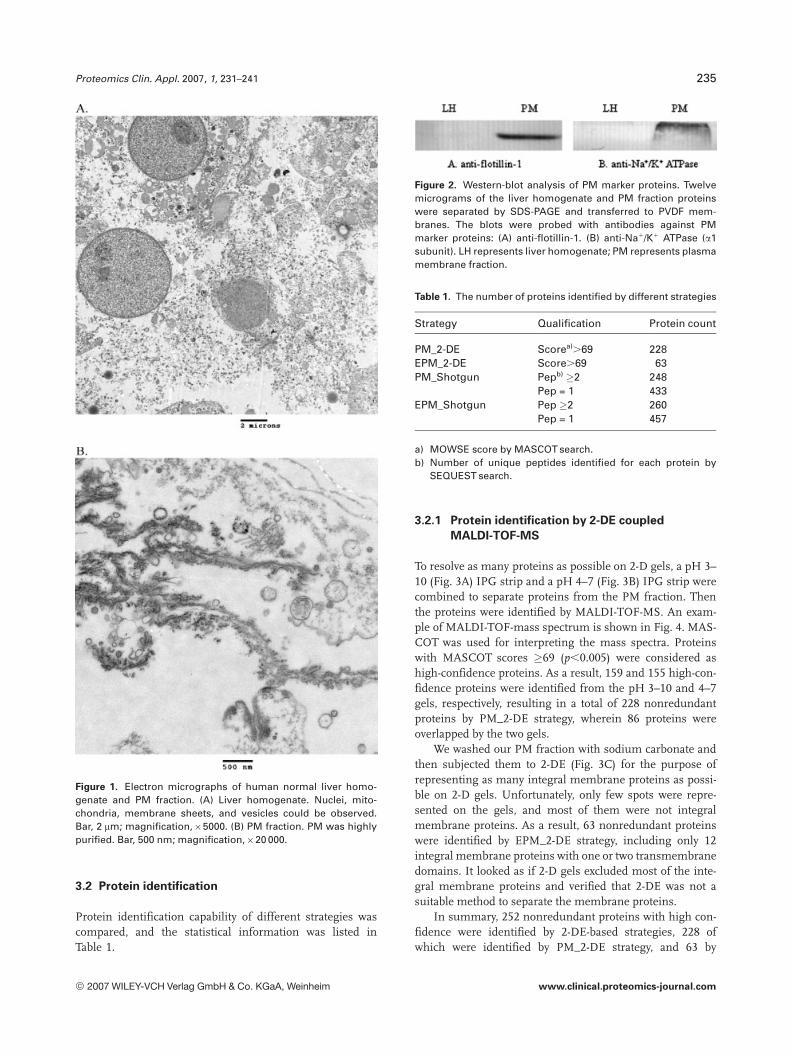
Proteomics Clin. Appl. 2007, 1, 231–241 235
Figure 1. Electron micrographs of human normal liver homo-genate and PM fraction. (A) Liver homogenate. Nuclei, mito-chondria, membrane sheets, and vesicles could be observed.Bar, 2 mm; magnification,65000. (B) PM fraction. PM was highlypurified. Bar, 500 nm; magnification,620 000.
3.2 Protein identification
Protein identification capability of different strategies wascompared, and the statistical information was listed inTable 1.
Figure 2. Western-blot analysis of PM marker proteins. Twelvemicrograms of the liver homogenate and PM fraction proteinswere separated by SDS-PAGE and transferred to PVDF mem-branes. The blots were probed with antibodies against PMmarker proteins: (A) anti-flotillin-1. (B) anti-Na1/K1 ATPase (a1subunit). LH represents liver homogenate; PM represents plasmamembrane fraction.
Table 1. The number of proteins identified by different strategies
Strategy Qualification Protein count
PM_2-DE Scorea).69 228EPM_2-DE Score.69 63PM_Shotgun Pepb) �2 248
Pep = 1 433EPM_Shotgun Pep �2 260
Pep = 1 457
a) MOWSE score by MASCOT search.b) Number of unique peptides identified for each protein by
SEQUEST search.
3.2.1 Protein identification by 2-DE coupled
MALDI-TOF-MS
To resolve as many proteins as possible on 2-D gels, a pH 3–10 (Fig. 3A) IPG strip and a pH 4–7 (Fig. 3B) IPG strip werecombined to separate proteins from the PM fraction. Thenthe proteins were identified by MALDI-TOF-MS. An exam-ple of MALDI-TOF-mass spectrum is shown in Fig. 4. MAS-COT was used for interpreting the mass spectra. Proteinswith MASCOT scores �69 (p,0.005) were considered ashigh-confidence proteins. As a result, 159 and 155 high-con-fidence proteins were identified from the pH 3–10 and 4–7gels, respectively, resulting in a total of 228 nonredundantproteins by PM_2-DE strategy, wherein 86 proteins wereoverlapped by the two gels.
We washed our PM fraction with sodium carbonate andthen subjected them to 2-DE (Fig. 3C) for the purpose ofrepresenting as many integral membrane proteins as possi-ble on 2-D gels. Unfortunately, only few spots were repre-sented on the gels, and most of them were not integralmembrane proteins. As a result, 63 nonredundant proteinswere identified by EPM_2-DE strategy, including only 12integral membrane proteins with one or two transmembranedomains. It looked as if 2-D gels excluded most of the inte-gral membrane proteins and verified that 2-DE was not asuitable method to separate the membrane proteins.
In summary, 252 nonredundant proteins with high con-fidence were identified by 2-DE-based strategies, 228 ofwhich were identified by PM_2-DE strategy, and 63 by
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.clinical.proteomics-journal.com

236 J. He et al. Proteomics Clin. Appl. 2007, 1, 231–241
Figure 3. 2-DE maps of the PM fraction from human normal liver.The proteins directly from the original PM fraction were sepa-rated on a pH 3–10 (A) and a pH 4–7 (B) IPG strips, followed by a10% SDS polyacrylamide gel. (C) 2-DE map of sodium carbonatewashed PM with a pH 3–10 IPG strip in the first dimension and a10% SDS polyacrylamide gel in the second dimension. The gelswere stained with CBB. Protein (500 mg) was loaded on (A) or (B),and 300 mg of protein was loaded on (C).
EPM_2-DE strategy, wherein 39 were overlapped. Obviously,sodium carbonate washing could greatly decrease the totalprotein amount on 2-D gels.
3.2.2 Protein identification by shotgun
The PM proteins were digested into peptide mixtures whichwere separated by RP-HPLC and then analyzed by LTQ massspectrometer. The acquired MS/MS spectra were interpretedby SEQUEST. The most widely used search algorithms for
interpreting MS/MS spectra are MASCOT and SEQUEST.MASCOTuses a probability-based scoring scheme to give thesignificant matches of a fragmented peptide, whereasSEQUEST uses empirical and correlation measurements toscore the alignment between observed and predicted spectra.It appears that the two algorithms are fairly comparable forLTQ acquired spectra [30, 31]. Using a reverse IPI 3.07 data-base, we evaluated the false positive rate of our peptideidentifications by SEQUEST. The rates under the criteriaused in the manuscript were 4.2 and 3.3% for PM_Shotgun
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.clinical.proteomics-journal.com
Proteomics Clin. Appl. 2007, 1, 231–241 237
Figure 4. MALDI-TOF mass spectrum of amino-peptidase N.
Figure 5. The base peak ofsodium carbonate washed PManalyzed by RP-HPLC MS/MS.
and EPM_Shotgun, respectively, which was relatively low(,5%). Finally, we identified 248 and 260 nonredundantproteins with at least two unique peptides matching for eachprotein by PM_Shotgun strategy and EPM_Shotgun strategyrespectively, and we considered these as high-confidenceproteins. A total of 392 high-confidence proteins were iden-tified by these two shotgun strategies, wherein 116 wereoverlapped. Interestingly, the identified integral membraneproteins drastically increased by sodium carbonate washing,with 54 in the 248 proteins (21.77%) and 139 in the 260 pro-teins (53.46%). This demonstrated that integral membraneproteins could be efficiently enriched by sodium carbonatewashing. As an example, Fig. 5 displayed the base peak ofsodium carbonate washed PM analyzed by RP-HPLC MS/MS.
3.2.3 Comparative analysis of high-confidence
proteins identified by 2-DE coupled
MALDI-TOF-MS and shotgun
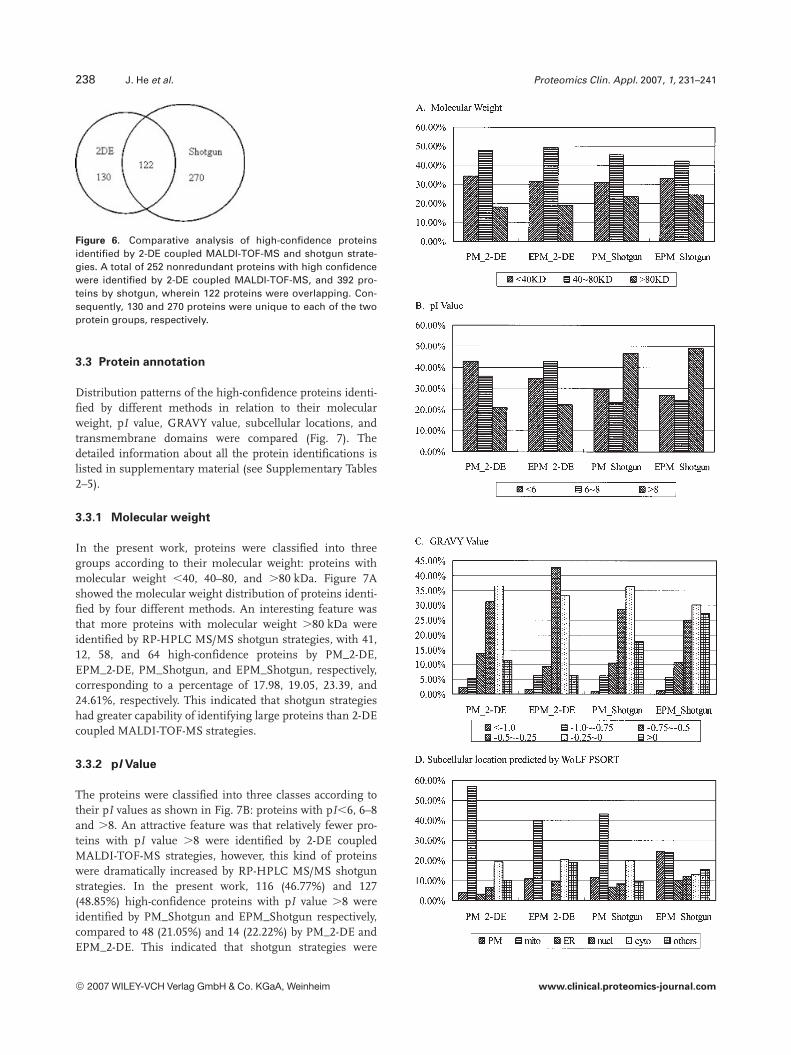
As shown in Fig. 6, 252 nonredundant proteins with highconfidence were identified by 2-DE coupled MALDI-TOF-MS, and 392 by shotgun, wherein 122 proteins wereoverlapped. We concluded that with respect to proteinidentification capability, shotgun strategies were superiorto 2-DE coupled MALDI-TOF-MS strategies. However,51.6% (130) of the proteins identified by 2-DE coupledMALDI-TOF-MS strategies could not be identified byshotgun. Therefore, 2-DE-based strategies may serve as acomplement and those proteins may supply more infor-mation.
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.clinical.proteomics-journal.com
238 J. He et al. Proteomics Clin. Appl. 2007, 1, 231–241
Figure 6. Comparative analysis of high-confidence proteinsidentified by 2-DE coupled MALDI-TOF-MS and shotgun strate-gies. A total of 252 nonredundant proteins with high confidencewere identified by 2-DE coupled MALDI-TOF-MS, and 392 pro-teins by shotgun, wherein 122 proteins were overlapping. Con-sequently, 130 and 270 proteins were unique to each of the twoprotein groups, respectively.
3.3 Protein annotation
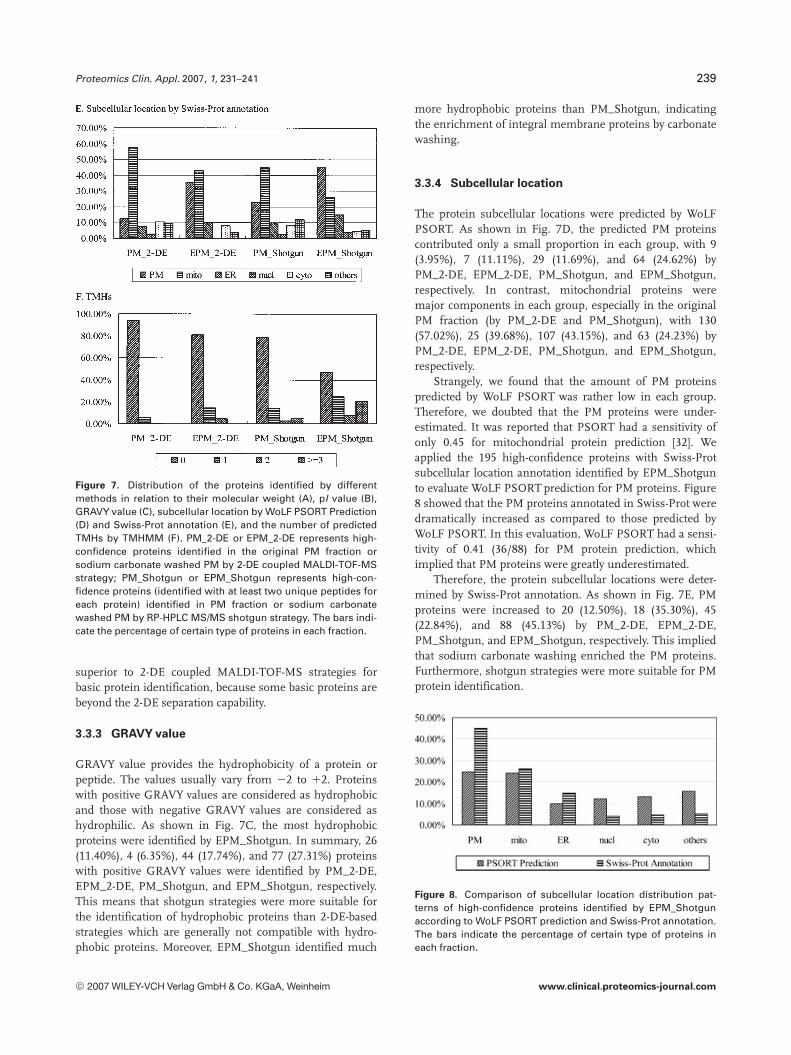
Distribution patterns of the high-confidence proteins identi-fied by different methods in relation to their molecularweight, pI value, GRAVY value, subcellular locations, andtransmembrane domains were compared (Fig. 7). Thedetailed information about all the protein identifications islisted in supplementary material (see Supplementary Tables2–5).
3.3.1 Molecular weight
In the present work, proteins were classified into threegroups according to their molecular weight: proteins withmolecular weight ,40, 40–80, and .80 kDa. Figure 7Ashowed the molecular weight distribution of proteins identi-fied by four different methods. An interesting feature wasthat more proteins with molecular weight .80 kDa wereidentified by RP-HPLC MS/MS shotgun strategies, with 41,12, 58, and 64 high-confidence proteins by PM_2-DE,EPM_2-DE, PM_Shotgun, and EPM_Shotgun, respectively,corresponding to a percentage of 17.98, 19.05, 23.39, and24.61%, respectively. This indicated that shotgun strategieshad greater capability of identifying large proteins than 2-DEcoupled MALDI-TOF-MS strategies.
3.3.2 pI Value
The proteins were classified into three classes according totheir pI values as shown in Fig. 7B: proteins with pI,6, 6–8and .8. An attractive feature was that relatively fewer pro-teins with pI value .8 were identified by 2-DE coupledMALDI-TOF-MS strategies, however, this kind of proteinswere dramatically increased by RP-HPLC MS/MS shotgunstrategies. In the present work, 116 (46.77%) and 127(48.85%) high-confidence proteins with pI value .8 wereidentified by PM_Shotgun and EPM_Shotgun respectively,compared to 48 (21.05%) and 14 (22.22%) by PM_2-DE andEPM_2-DE. This indicated that shotgun strategies were
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.clinical.proteomics-journal.com
Proteomics Clin. Appl. 2007, 1, 231–241 239
Figure 7. Distribution of the proteins identified by differentmethods in relation to their molecular weight (A), pI value (B),GRAVY value (C), subcellular location by WoLF PSORT Prediction(D) and Swiss-Prot annotation (E), and the number of predictedTMHs by TMHMM (F). PM_2-DE or EPM_2-DE represents high-confidence proteins identified in the original PM fraction orsodium carbonate washed PM by 2-DE coupled MALDI-TOF-MSstrategy; PM_Shotgun or EPM_Shotgun represents high-con-fidence proteins (identified with at least two unique peptides foreach protein) identified in PM fraction or sodium carbonatewashed PM by RP-HPLC MS/MS shotgun strategy. The bars indi-cate the percentage of certain type of proteins in each fraction.
superior to 2-DE coupled MALDI-TOF-MS strategies forbasic protein identification, because some basic proteins arebeyond the 2-DE separation capability.
3.3.3 GRAVY value
GRAVY value provides the hydrophobicity of a protein orpeptide. The values usually vary from 22 to 12. Proteinswith positive GRAVY values are considered as hydrophobicand those with negative GRAVY values are considered ashydrophilic. As shown in Fig. 7C, the most hydrophobicproteins were identified by EPM_Shotgun. In summary, 26(11.40%), 4 (6.35%), 44 (17.74%), and 77 (27.31%) proteinswith positive GRAVY values were identified by PM_2-DE,EPM_2-DE, PM_Shotgun, and EPM_Shotgun, respectively.This means that shotgun strategies were more suitable forthe identification of hydrophobic proteins than 2-DE-basedstrategies which are generally not compatible with hydro-phobic proteins. Moreover, EPM_Shotgun identified much
more hydrophobic proteins than PM_Shotgun, indicatingthe enrichment of integral membrane proteins by carbonatewashing.
3.3.4 Subcellular location
The protein subcellular locations were predicted by WoLFPSORT. As shown in Fig. 7D, the predicted PM proteinscontributed only a small proportion in each group, with 9(3.95%), 7 (11.11%), 29 (11.69%), and 64 (24.62%) byPM_2-DE, EPM_2-DE, PM_Shotgun, and EPM_Shotgun,respectively. In contrast, mitochondrial proteins weremajor components in each group, especially in the originalPM fraction (by PM_2-DE and PM_Shotgun), with 130(57.02%), 25 (39.68%), 107 (43.15%), and 63 (24.23%) byPM_2-DE, EPM_2-DE, PM_Shotgun, and EPM_Shotgun,respectively.
Strangely, we found that the amount of PM proteinspredicted by WoLF PSORT was rather low in each group.Therefore, we doubted that the PM proteins were under-estimated. It was reported that PSORT had a sensitivity ofonly 0.45 for mitochondrial protein prediction [32]. Weapplied the 195 high-confidence proteins with Swiss-Protsubcellular location annotation identified by EPM_Shotgunto evaluate WoLF PSORT prediction for PM proteins. Figure8 showed that the PM proteins annotated in Swiss-Prot weredramatically increased as compared to those predicted byWoLF PSORT. In this evaluation, WoLF PSORT had a sensi-tivity of 0.41 (36/88) for PM protein prediction, whichimplied that PM proteins were greatly underestimated.
Therefore, the protein subcellular locations were deter-mined by Swiss-Prot annotation. As shown in Fig. 7E, PMproteins were increased to 20 (12.50%), 18 (35.30%), 45(22.84%), and 88 (45.13%) by PM_2-DE, EPM_2-DE,PM_Shotgun, and EPM_Shotgun, respectively. This impliedthat sodium carbonate washing enriched the PM proteins.Furthermore, shotgun strategies were more suitable for PMprotein identification.
Figure 8. Comparison of subcellular location distribution pat-terns of high-confidence proteins identified by EPM_Shotgunaccording to WoLF PSORT prediction and Swiss-Prot annotation.The bars indicate the percentage of certain type of proteins ineach fraction.
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.clinical.proteomics-journal.com

240 J. He et al. Proteomics Clin. Appl. 2007, 1, 231–241
3.3.5 Transmembrane domains
We predicted protein transmembrane domains by TMHMM,and compared the TMHs distribution patterns of the high-confidence proteins identified by different strategies(Fig. 7F). For the 2-DE-based strategies, 13 proteins (5.70%)with one TMH and one protein (0.44%) with two TMHs wereidentified by PM_2-DE, compared to nine proteins (14.29%)with one TMH and three proteins (4.76%) with two TMHs byEPM_2-DE. For the shotgun strategies, 34 proteins (13.71%)with one TMH, 7 proteins (2.82%) with two TMHs and 13proteins (5.24%) with at least three TMHs were identified byPM_Shotgun, compared to 65 proteins (25.00%) with oneTMH, 21 proteins (8.08%) with two TMHs and 53 proteins(20.38%) with at least three TMHs by EPM_Shotgun. Inter-estingly, 2-DE-based strategies could only identify few inte-gral membrane proteins with one or two TMHs, while shot-gun strategies could identify much more integral membraneproteins including many proteins with multiple TMHs.Another fascinating feature was that sodium carbonatewashing greatly increased the integral membrane proteinsidentified by shotgun strategies. This demonstrated thatshotgun strategies were more suitable for integral membraneprotein identification, and moreover, sodium carbonatewashing could obviously enrich integral membrane proteins.
The results suggested that EPM_Shotgun was the beststrategy to study integral membrane proteins. Of the 260proteins identified by EPM_Shotgun, 139 were classified asintegral membrane proteins, wherein 60 were PM proteinsaccording to Swiss-Prot annotation, 19 were mitochondrialproteins, 26 were ER proteins, and 7 were localized to othersites, and 27 had no subcellular location annotation. All ofthe 139 proteins had from 1 to 17 transmembrane domains,53 (38.13%) of which contained three or more (see Supple-mentary Table 6).
A number of interesting proteins with various functionswere included. Most of the integral PM proteins with multi-ple TMHs were transporters. For example, Canalicular mul-tispecific organic anion transporter 1 and 2 are mainly expres-sed in liver, and they may mediate hepatobiliary excretion ofnumerous organic anions. Solute carrier family 2, facilitatedglucose transporter, member 2 may mediate the bidirectionaltransfer of glucose across the PM of hepatocytes and isresponsible for uptake of glucose by the beta cells. Solutecarrier organic anion transporter family, member 1B1 or 1B3 isalso called liver-specific organic anion transporter 1 or 2.They mediate the Na1-independent transport of organicanions, and are highly expressed in liver, particularly inbasolateral membranes of hepatocytes. Bile salt export pumpis involved in the ATP-dependent secretion of bile salts intothe canaliculus of hepatocytes. It is expressed predominantlyin the liver, where it was further localized to the canalicularmicrovilli and to subcanalicular vesicles of the hepatocytes.
Another important group of these transmembrane pro-teins are enzymes, such as sodium/potassium-transportingATPase, calcium-transporting ATPase, potassium-transport-
ing ATPase, 50-nucleotidase, and so on. In addition, therewere some receptors, cell junction associated proteins, anti-gens, and hypothetical proteins.
Several papers mentioned that shotgun strategies weresuperior to 2-DE-based strategies. Yates and coworkers [6, 7]referred to the major drawbacks of 2-DE-based strategies inthe study of membrane proteome, and demonstrated thatshotgun strategies were more suitable for membrane prote-omics. Fischer et al. [33] also had a similar conclusion.Unfortunately, they had not systematically analyzed the dif-ferences between 2-DE based and shotgun strategies. In thepresent work, different strategies were compared from manyrespects, including protein molecular weight, pI value,GRAVY, subcellular location, and transmembrane domains.After a systematic comparison, we could easily understandwhich strategy was more suitable for membrane proteomicsand why. Finally, we found that the strategy combining car-bonate washing and shotgun identification was the optimumstrategy to study PM proteome.
4 Concluding remarks
We evaluated four strategies in PM proteomics: PM_2-DE,EPM_2-DE, PM_Shotgun, and EPM_Shotgun. Protein iden-tification capability and protein characteristics such as mo-lecular weight, pI value, GRAVY value, subcellular location,and transmembrane domains were compared. In summary,shotgun strategies had greater protein identification cap-ability than 2-DE coupled MALDI-TOF-MS strategies, with392 and 252 high-confidence proteins identified, respectively.With respect to protein molecular weight, shotgun strategieswere more suitable for large protein identification, with about24% of the identified proteins .80 kDa, compared to about18% for 2-DE-based strategies. For pI value, shotgun strate-gies were more compatible with basic proteins, identifyingabout 48% of proteins with pI.8, compared to about 22% for2-DE-based strategies. With respect to GRAVYvalue, the mosthydrophobic proteins were identified by EPM_Shotgun, with77 (27.31%) proteins having positive GRAVY values. For pro-tein subcellular location, the bioinformatic tool WoLF PSORThad a sensitivity of only 0.41 for PM protein prediction asevaluated by Swiss-Prot annotation. Therefore, PM proteinswere greatly underestimated by WoLF PSORT prediction.According to Swiss-Prot annotation, most of the PM proteins(88, up to 45.13%) were identified by EPM_Shotgun strategy.With respect to transmembrane domains, EPM_Shotgunstrategy gave the best results, with 139 integral membraneproteins identified, wherein 60 were PM proteins. On thecontrary, 2-DE coupled MALDI-TOF-MS strategies excludedmost of integral membrane proteins.
In conclusion, EPM_Shotgun is the best strategy to studyPM proteome according to the present work, and 2-DE-basedstrategies may serve as a complement. This work provides anoptimum strategy to study the PM proteome, and helps toelucidate the function of PM.
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.clinical.proteomics-journal.com

Proteomics Clin. Appl. 2007, 1, 231–241 241
We thank Professor Songping Liang for the help in plasmamembrane preparation. We are grateful to Professor Rong Zengfor her help in RP-HPLC MS/MS. We thank ProfessorChuanmao Zhang for help with electron microscopy and Dr.Songfeng Wu for the help with false positive rate evaluation. Hu-man normal liver tissues were provided by HLPP. This work wassupported by grants from National Basic Research Program ofChina (nos. 2006CBODO100 and 2001CB510207) andNational High Technology Research and Development Programof China (no. 2004BA711A18).
5 References
[1] Hopkins, A. L., Groom, C. R., Nat. Rev. Drug Discov. 2002, 1,727–730.
[2] Wallin, E., Von Heijne, G., Protein Sci. 1998, 1998, 1029–1038.
[3] Santoni, V., Molloy, M., Rabilloud, T., Electrophoresis 2000,21, 1054–1070.
[4] Molloy, M. P., Herbert, B. R., Slade, M. B., Rabilloud, T. et al.,Eur. J. Biochem. 2000, 267, 2871–2881.
[5] Molloy, M. P., Anal. Biochem. 2000, 280, 1–10.
[6] Wu, C. C., Maccoss, M. T., Howell, K. E., Yates, J. R., Nat.Biotechnol. 2003, 21, 532–538.
[7] Wu, C. C., Yates, J. R., Nat. Biotechnol. 2003, 21, 262–267.
[8] Chevallet, M., Santoni, V., Poinas, A., Rouquie, D. et al.,Electrophoresis 1998, 19, 1901–1909.
[9] Henningsen, R., Gale, B. L., Straub, K. M., Devagel, D. C.,Proteomics 2002, 2, 1479–1488.
[10] Zhang, L. J., Xie, J. Y., Wang, X., Liu, X. H. et al., Proteomics2005, 5, 4510–4524.
[11] Blonder, J., Goshe, M. B., Moore, R. J., Pasa-Tolic, L. et al., J.Proteome Res. 2002, 1, 351–360.
[12] Blonder, J., Conrads, T. P., Yu, L. R., Terunuma, A. et al., Pro-teomics. 2004, 4, 31–45.
[13] Zhao, Y. X., Zhang, W., Kho, Y. J., Zhao, Y. M., Anal. Chem.2004, 76, 1817–1823.
[14] Nielsen, P. A., Olsen, J. V., Podtelejnikov, A. V., Andersen, J.R. et al., Mol. Cell. Proteomics 2005, 4, 402–408.
[15] Oh, P., Li, Y., Yu, J. Y., Durr, E. et al., Nature 2004, 429, 629–639.
[16] Shin, B. K., Wang, H., Yim, A. M., Naour, F. L. et al., J. Biol.Chem. 2003, 278, 7607–7616.
[17] Fujiki, Y., Hubbard, A. L., Fowler, S., Lazarow, P. B., J. CellBiol. 1982, 93, 97–102.
[18] Hubbard, A. L., Ma, A., J. Cell Biol. 1983, 96, 230–239.
[19] Hubbard, A. L., Wall, D. A., Ma, A., J. Cell Biol. 1983, 96, 217–229.
[20] Fleischer, S., Kervina, M., Methods Enzymol. 1974, 31, 6–41.
[21] Molloy, M. P., Phadke, N. D., Maddock, J. R., Andrews, P. C.,Electrophoresis 2001 22, 1686–1696.
[22] Tu, X. L., Huang, A., Bae, D., Slaughter, N. et al., J. ProteomeRes. 2004, 3, 445–454.
[23] Yang, W., Liu, P., Liu, Y. S., Wang, Q. S. et al., Proteomics2006, 6, 2982–2990.
[24] Gobom, J., Schuerenberg, M., Mueller, M., Theiss, D. et al.,Anal. Chem. 2001, 73, 434–438.
[25] Kersey, P. J., Duarte, J., Williams, A., Karavidopoulou, Y.,Proteomics 2004, 4, 1985–1988.
[26] Bradford, M., Anal. Biochem. 1976, 72, 248–254.
[27] Washburn, M. P., Wolters, D., Yates, J. R., Nat. Biotechnol.2001, 19, 242–247.
[28] Kyte, J., Doolittle, R. F., J. Mol. Biol. 1982, 157, 105–132.
[29] Krogh, A., Larsson, B., von Heijne, G., Sonnhammer, E. L. L.,J. Mol. Biol. 2001, 305, 567–580.
[30] Elias, J. E., Haas, W., Faherty, B. K., Gygi, S. P., Nat. methods2005, 2, 667–675.
[31] Resing, K. A., Meyer-Arendt, K., Mendoza, A. M., Aveline-Wolf., L. D. et al., Anal. Chem. 2004, 76, 3556–3568.
[32] Jiang, X. S., Zhou, H., Zhang, L., Sheng, Q. H. et al., Mol. Cell.Proteomics 2004, 3, 441–455.
[33] Fischer, F., Wolters, D., Rogner, M., Poetsch, A., Mol. Cell.Proteomics 2006, 5, 444–453.
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.clinical.proteomics-journal.com
Copyright © 2022 FDOKUMEN




















