
Intra-tracheal administration of a naked plasmid expressing ...
Upload
independentCategory
view
2download
0
Coimmunization with an Optimized IL-15 Plasmid Results inEnhanced Function and Longevity of CD8 T Cells That ArePartially Independent of CD4 T Cell Help1
Michele A. Kutzler,* Tara M. Robinson,* Michael A. Chattergoon,* Daniel K. Choo,†
Andrew Y. Choo,‡ Philip Y. Choe,* Mathura P. Ramanathan,* Rose Parkinson,*Sagar Kudchodkar,* Yutaka Tamura,§ Maninder Sidhu,¶ Vidia Roopchand,¶ J. Joseph Kim,�
George N. Pavlakis,# Barbara K. Felber,# Thomas A. Waldmann,** Jean D. Boyer,* andDavid B. Weiner2*
DNA vaccines are a promising technology for the induction of Ag-specific immune responses, and much recent attention has goneinto improving their immune potency. In this study we test the feasibility of delivering a plasmid encoding IL-15 as a DNA vaccineadjuvant for the induction of improved Ag-specific CD8� T cellular immune responses. Because native IL-15 is poorly expressed,we used PCR-based strategies to develop an optimized construct that expresses 80-fold higher than the native IL-15 construct.Using a DNA vaccination model, we determined that immunization with optimized IL-15 in combination with HIV-1gag DNAconstructs resulted in a significant enhancement of Ag-specific CD8� T cell proliferation and IFN-� secretion, and strong inductionof long-lived CD8� T cell responses. In an influenza DNA vaccine model, coimmunization with plasmid expressing influenza APR8/34 hemagglutinin with the optimized IL-15 plasmid generated improved long term CD8� T cellular immunity and protectedthe mice against a lethal mucosal challenge with influenza virus. Because we observed that IL-15 appeared to mostly adjuvantCD8� T cell function, we show that in the partial, but not total, absence of CD4� T cell help, plasmid-delivered IL-15 could restoreCD8 secondary immune responses to an antigenic DNA plasmid, supporting the idea that the effects of IL-15 on CD8� T cellexpansion require the presence of low levels of CD4 T cells. These data suggest a role for enhanced plasmid IL-15 as a candidateadjuvant for vaccine or immunotherapeutic studies. The Journal of Immunology, 2005, 175: 112–123.
T he generation of functional cellular immunity that resultsin a long-lasting memory protective immune responseupon pathogen rechallenge is the goal of vaccination. In a
model of acute infection, the quality of memory CD8� T cell re-sponse depends on help from CD4� T cells (1–14). In the presenceof CD4� T cells, memory CD8� T cells rapidly begin to prolif-erate and undergo further differentiation after a second encounterwith Ag, resulting in enhanced effector responses, including se-cretion of cytotoxic molecules and the antiviral cytokines IFN-�and TNF-�. A recent study by Sun et al. (15) investigated at whatstage during the immune response to pathogen are CD4� T cellsessential in the promotion of functional CD8� T cell memory in amodel of acute infection. Adoptive transfer of effector or memory
CD8� T cells into wild-type or CD4� T cell-deficient mice dem-onstrated that the presence of CD4� T cells was important onlyafter, not during, the early CD8� T cell-programming phase. In theabsence of CD4� T cells, memory CD8� T cells became func-tionally impaired and decreased in quantity over time (15). Thisstudy demonstrates that in the context of an acute infection, CD4�
T cells are required only during the maintenance phase of long-lived memory CD8� T cells. However, unlike acute infection, therole of CD4 T cells in the priming of functional CD8� T celleffector and memory cells in a model of DNA vaccination isunknown.
It is believed that CD8� T cell responses are important for con-trolling HIV-1 infection and slowing disease progression. Al-though the exact function of HIV-1-specific CD8� T cells in thisprocess has not been completely elucidated, a correlation has beenestablished between long term nonprogression of HIV-1 in sero-positive individuals and HIV-1-specific CD8� T cell-mediatedcellular responses (4, 16–19). The loss of CD4� T cell functionduring chronic HIV infection can have deleterious consequenceson the ability of HIV-1-specific CD8� T cells to maintain effectorT cell activity and control virus. This suggests that understandingthe factors that supplement CD4� T cell function and that supportCD8 T cell function could be important for HIV immune therapystrategies where CD4 help is compromised. Accordingly, genera-tion of vaccines, including DNA vaccines, that encode their ownforms of T cell help would be worthy of investigation.
Generation of memory CD4� and CD8� T cells is dependent onantigenic stimulation, but the survival of memory lymphocytesappears to be Ag independent and requires specific, peripherally
*Department of Pathology and Laboratory Medicine, University of PennsylvaniaSchool of Medicine, Philadelphia, PA 19104; †Department of Microbiology and Im-munology, Emory University School of Medicine, Atlanta, GA 30322; ‡Program forBiological and Biomedical Science, Harvard Medical School, Boston, MA 02115;§National Institute of Advanced Industrial Science and Technology, Sapporo, Japan;¶Vaccine Discovery, Wyeth Lederle, Pearl River, NY 10965; �Viral Genomix, Blue-bell, PA 19422; and #Basic Research Laboratory and **Metabolism Branch, NationalCancer Institute, National Institutes of Health, Bethesda, MD 20892
Received for publication August 10, 2004. Accepted for publication April 16, 2005.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported in part by Grant F32AI054152 (to M.A.K.) and by aNational Institute of Allergy and Infectious Diseases sponsored HIV Vaccine Designand Development Team and Integrated Preclinical-Clinical Program (to D.B.W.).2 Address correspondence and reprint requests to Dr. David B. Weiner, University ofPennsylvania, 422 Curie Boulevard, Room 505 Stellar Chance Laboratories, Phila-delphia, PA 19104-6100. E-mail address: [email protected]
The Journal of Immunology
Copyright © 2005 by The American Association of Immunologists, Inc. 0022-1767/05/$02.00

produced cytokines (20–22). One such cytokine that appears tohave a significant impact on CD8� T cells is IL-15 (23–26). In thecontext of immune therapy, genetic vaccines, IL-15 has beenshown to be effective against tumors as well as some disease mod-els (24, 27–37). IL-15 has been reported to support memory CD8�
T cell proliferation (38) and specifically induce the proliferation ofCD44high or CD122high memory CD8� T cells (27, 39, 40). More-over, it has been shown that there is a lack of memory phenotypeCD8� T cells in IL-15R�- and IL-15-deficient mice (41). In ad-dition, Ab-mediated blockade of IL-2/IL-15R� inhibits the prolif-eration of memory phenotype CD8� T cells in vivo (22). Takentogether, these data suggest that IL-15 plays an essential role in theproliferation of memory CD8� T cells. We hypothesize that IL-15may act to enhance the productive CD8� T cell memory poolinduced in the context of an HIV DNA vaccine.
To test an effective IL-15 plasmid for vaccination that secretesenhanced levels of IL-15, several obstacles had to be overcome.The regulation of IL-15 is complex, with multifaceted controls atthe levels of message transcription, message translation, and pro-tein translocation and secretion (42, 43). IL-15 contains two alter-native leader peptides, one with 48 aa (long signal peptide (LSP)3)and one with 21 aa (short signal peptide (SSP)) (44, 45). Theclassical LSP form of IL-15 is associated with all secreted IL-15,whereas the SSP form of IL-15 is not secreted, but, rather, is storedintracellularly. IL-15 mRNA includes a number of elements thatimpede its translation into protein. Bamford et al. (46) showed thatby replacing the long leader with the signal peptide-coding se-quence from IL-2, the sum of IL-15 retained and secreted in-creased 17- to 20-fold with the IL-2 leader. Conversely, by replac-ing the leader on IL-2 with that of IL-15 LSP, the quantity of IL-2secreted was reduced 40- to 50-fold. Furthermore, in a similarstudy conducted by Onu et al. (45), replacing the LSP of IL-15with the signal peptide from CD33 resulted in increased levels oftranslation and secretion of IL-15 protein. It has yet to be deter-mined whether an optimized form of IL15, modified through PCRstrategies to remove negative regulatory features resulting in en-hanced protein secretion, can influence immunogenicity in vivo ina model of DNA vaccination.
In this report, through PCR-based strategies, we have success-fully optimized and enhanced the production and secretion of abiologically active IL-15 in a DNA plasmid (pIL-15-Opt). Theoptimized pIL-15-Opt acts as a potent adjuvant for HIV-1gagDNA vaccines by significantly enhancing Ag-specific CD8� Teffector function and proliferation. To test the physiological im-portance of the enhanced CD8� T cell immune responses usingpIL-15-Opt, we used an influenza mucosal challenge model inwhich we show that coimmunization of mice with pIL-15-Opt andplasmid influenza hemagglutinin (pHA) confers protection from alethal mucosal challenge of influenza virus. Because we observedthat IL-15 appeared to mostly adjuvant CD8� T cell function, weshow that in the partial, but not total, absence of CD4 � T cell help,plasmid-delivered IL-15 could restore CD8 secondary immune re-sponses to an antigenic DNA plasmid, suggesting that the effectsof IL-15 on CD8� T cells requires the presence of low levels ofCD4� T cells in a DNA vaccination model. This is an importantexample of engineering a DNA vaccine for HIV-1 with at leastpartial CD4� T cell independence.
Materials and MethodsDNA plasmids
DNA vaccine constructs expressing HIV-1gag were prepared as describedpreviously (47). The cloning of the human IL-15 construct (sequence fromGenBank, accession no. NM00585) into pVAX1 cloning vector (Invitro-gen Life Technologies) was conducted by PCR amplification with the fol-lowing 5� to 3� primers: pIL-15-SSP sense, GCCCCCGTCGACGCCGCCACCATGAGAATTTCGAAACCACATTTGAG; pIL-15-SSP antisense,ATCGGGCTCGAGTCAAGAAGTGTTGATGAACATTTGG; pIL-15-LSP sense, GCCCCCGGTACCGCCGCCACCATGGTATTGGGAACCATA; pIL-15-LSP antisense, ATCGGGGGATCCTCAAGAAGTGTTGATGAACAT; pIL-15-Opt sense, GCCCCCGAATTCGCCGCCACCATGGATTGGACTTGGATCTTATTTTT AGTTGCTGCTGCTACTAGAGTTCATTCTAACTGGGTGAATGTAATAAGT; and pIL-15-Opt antisense,ATCGGGCTCGAGTCAAGAAGTGTTGATGAACATTTGG. PCR con-ditions for all reactions were one cycle at 97°C for 3 min; 30 cycles at 94°Cfor 1 min, 55°C for 1 min and 15 s, and 72°C for 1 min and 15 s; followedby final extension at 72°C for 10 min. The 405 bp (SSP), 486 bp (LSP), and396 bp (Opt) PCR product was ligated into pVAX1 cloning vector after arestriction enzyme digestion using EcoRI (New England Biolabs) andXho1 (New England Biolabs), which were designed into the PCR primersand are in the multiple cloning region of the vectors. All positive cloneswere verified by sequence analysis.
In vitro transfection
Expression levels of the plasmid construct were tested after transient trans-fection of RD cells (American Type Culture Collection). Cells were platedin six-well tissue culture dishes at a density of 2 � 105 cells/well in com-plete DMEM plus 10% FBS (Invitrogen Life Technologies) and allowed toadhere overnight. The next day cells were transfected with IL-15 plasmid(1 �g/well) using FuGene 6 transfection reagent (Roche) according to themanufacturer’s protocol. After 48 h, cell supernatants were harvested andanalyzed for the presence of human IL-15 protein by commercial ELISAkits (R&D Systems).
IL-15 functional assay
The MTT cell proliferation assay, according to the manufacturer’s recom-mendations (American Type Culture Collection), was used to assess thefunctional activity of secreted IL-15. CTLL-2 cells obtained from Ameri-can Type Culture Collection were grown and maintained in RPMI 1640(Invitrogen Life Technologies) complete medium containing 5% superna-tant from Con A-stimulated rat spleen cells (BD Biosciences) to supple-ment IL-2, which is required for cell growth. At the time of the assay, cellswere centrifuged and washed five times with RPMI 1640 complete withoutany Con A supernatant to remove residual IL-2. Cells were plated at aconcentration of 5 � 104 cells/well in 96-well, flat-bottom polystyrenedishes. Standard human IL-15 (R&D Systems) as well as the supernatantsfrom transfected cells to be assessed for the capacity to induce CTLL-2proliferation were added to each well, and incubation was continued for48 h at 37°C. At this time, 10 �l of MTT reagent was added, and incubationwas continued for another 2 h. Readings at 570 nm absorbance from thewells containing the standard IL-15 sample (picograms per milliliter ofstandard IL-15) were used to make a standard curve, which was then usedto estimate the activity of the unknown samples and depicted as bioactivity(picograms per milliliter).
Indirect immunofluorescent assay
The indirect immunofluorescent assay for monitoring IL-15 expressionconstructs was conducted by the following protocol (48). HeLa cells grownin slide chambers (BD Biosciences) were transfected with IL-15 constructsusing DOTAP transfection agent (Roche). Forty-eight hours after transfec-tion, the cells were washed twice with cold PBS and fixed on slides usingmethanol for 15 min. Upon removal of the residual solvents from theslides, the cells were incubated with anti-mouse human IL-15 (BD Pharm-ingen) for 90 min. The slides were incubated with PE-conjugated second-ary Ab (Roche) for 45 min. 4�,6-Diamido-2-phenylindole hydrochloride(Sigma-Aldrich) was added to the solution of secondary Ab to coun-terstain the nuclei to show the nuclei of the total number of cells avail-able in the given field. The slides were mounted with mounting mediumcontaining antifading reagent (Molecular Probes). The images were an-alyzed using the Phase 3 Image Pro program for fluorescent microscopy(Media Cybernetics).
3 Abbreviations used in this paper: LSP, long signal peptide; HA, hemagglutinin; KO,knockout; Opt, optimized leader sequence; pHA, plasmid HA; macs, macrophages;p.i., postinfection; SFC, spot-forming cell; SI, stimulation index; SSP, short signalpeptide.
113The Journal of Immunology

Plasmid immunization and mice
The quadriceps muscle of 6- to 8-wk-old female BALB/c (The JacksonLaboratory) were injected two or three times, each separated by 2 wk (47).We obtained the CD4 knockout (CD4KO) mice as a gift from Dr. R.Eisenberg (University of Pennsylvania, Philadelphia, PA; B6.129S6-Cd4tm1knw/J obtained from The Jackson Laboratory). CD4KO (B6.129S6-Cd4tm1Knw/J) mice have a significant block in CD4 T cell development,�90% circulating T cells are CD8�, and manifest a class II-restricteddeficit in Th cell activity. We obtained the inbred parental strain C57BL/6Jmice from The Jackson Laboratory. For all experiments in mice, the ani-mals were immunized with either 50 or 100 �g of HIV-1 antigenic plas-mids, IL-15 constructs, or a vector control in 0.25% bupivicaine-HCl (Sig-ma-Aldrich) in isotonic citrate buffer as designated in the figures (47).Coadministration of various gene plasmids involved mixing the designatedDNA plasmids before injection in a final volume of 100 �l (47). All DNAwas made using Qiagen columns (endotoxin free) (47). All animals werehoused in a temperature-controlled, light-cycled facility at the Universityof Pennsylvania, and they were cared for under the guidelines of the Na-tional Institutes of Health and the University of Pennsylvania.
Splenocyte purification
Spleens from each mouse were harvested and pooled in a 15-ml conicalcontaining RPMI 1640 (one tube for each experimental group) (47). In asterile tissue culture hood, the pooled spleens from each experimentalgroup were placed in a petri dish and crushed using the plunger from a 3-mlsyringe. The cells were then incubated for 5–10 min at room temperaturein RBC lysing solution. The splenocytes were washed and put through a70-�m cell strainer to remove any remaining spleen organ stroma. Thesplenocytes were washed in RPMI 1640 twice, resuspended in R10 me-dium (RPMI 1640 plus 10% FBS), and counted (cell viability is determinedusing trypan blue stain) using a hemocytometer.
T cell proliferation assay
A T cell proliferation assay was conducted as previously described (47).Briefly, splenocytes (5 � 105 cell) were added to each well of a 96-well,microtiter, flat-bottom plate and stimulated in the presence of recombinantp24 protein (Immunodiagnostics) in triplicate, resulting in final concentra-tions of 5 and 1 �g/ml. The cells were incubated at 37°C in 5% CO2 for3 days. One microcurie of tritiated thymidine was added to each well, andthe cells were incubated for an additional 12–18 h at 37°C. Plates wereharvested, and the amount of incorporated tritiated thymidine was mea-sured in a � Plate reader (Wallac). The stimulation index (SI) was deter-mined from the formula: SI � (experimental count/spontaneous count).Spontaneous count wells included 10% FCS, which served as an irrelevantprotein control. Similarly, splenocytes from pHIV-1gag- or control-immu-nized mice routinely have an SI of 1 against an irrelevant protein target(47). To assure that cells were healthy, PHA (Sigma-Aldrich) or Con A(Sigma-Aldrich) was used as a polyclonal stimulator positive control.
ELISPOT assay
An ELISPOT assay was conducted as previously described (49). Briefly,ELISPOT 96-well plates (Millipore) were coated with anti-mouse IFN-�capture Ab and incubated for 24 h at 4°C (R&D Systems), and the fol-lowing day, the plates were washed and blocked for 2 h with 1% BSA. Twohundred thousand splenocytes from the immunized mice were added toeach well and stimulated overnight at 37°C in 5% CO2 in the presence ofRPMI 1640 (negative control), Con A (positive control), or specific peptide(HIV-1gag, p55) Ags (10 �g/ml). HIV-1gag 15-mer peptides spanning theentire HIV-1gag consensus clade B subtype, overlapping by 11 aa, wereprovided by the AIDS Reagent and Reference Repository. For the ELIS-POT assays, p55 peptide mix was used in which all 15-mer peptides weremixed together and added to splenocytes at a 10 �g/ml final concentration.For the epitope-mapping studies, a matrix format was used in which each
FIGURE 1. Plasmid IL-15 constructs. A, Amino acid alignment of IL-15 SSP, -LSP, and -Opt. Signal peptides are 21, 48, and 18 aa in length for SSP,LSP, and Opt, respectively. The mature IL-15 amino acid sequence is 100% homologous for each isomer. B, Schematic of two native isoforms of IL-15,termed IL-15-SSP and IL-15-LSP, as well as the IL-15-Opt isoform. For each construct, the amino acid length of the signal peptides (21, 48, and 18 aa,respectively) and mature IL-15 (115 aa) is shown. C, Schematic of the cloning vector, pVAX1, using the EcoRI and XhoI restriction enzymes sites in themultiple cloning region.
114 IL-15 ENHANCES CD8 T CELL FUNCTION INDEPENDENT OF T CELL HELP

individual 15-mer from HIV-1gag was put into two separate pools of pep-tides so that identification of positive peptide epitopes could be determined.After 24 h of stimulation, the cells were washed and incubated for 24 h at4°C with biotinylated anti-mouse IFN-� Ab (R&D Systems). The plateswere washed, and streptavidin-alkaline phosphatase (R&D Systems) wasadded to each well and incubated for 2 h at room temperature. The platewas washed, and 5-bromo-4-chloro-3�-indolylphosphate p-toluidine saltand nitro blue tetrazolium chloride (chromogen color reagent; R&D Sys-tems) were added to each well. The plate was then rinsed with distilledwater and dried at room temperature. Spots were counted by an automatedELISPOT reader (CTL Limited).
CD8� T cell depletion for lymphoproliferation andELISPOT assays
Immunomagnetic cell separation using Dynabeads (Dynal Biotech) mouseCD8 (clone Lyt2) was used as the method for separation of Lyt2-positivecells, resulting in 90% depletion in 20 min using 1 � 107 beads/ml for2.5 � 106 splenocytes. Depletion was conducted as described by the man-ufacturer. Briefly, the Dynabead mouse CD8 suspension was added di-rectly to the cell suspension, mixed thoroughly, and incubated for 20 minat 4°C on an apparatus that provides gentle tilting and rotation. The cell/bead mixture was then placed on a magnet (Dynal MPC) for 1 min tocollect the rosetted cells at the wall of the conical tube. The negativelyisolated cells (CD8� T cell depleted) were transferred to a second tube forfurther use in the lymphoproliferation and ELISPOT assays.
HA/flu challenge model
Using the pTR519 plasmid (50), cohorts of mice (n � 10) were immunizedand then boosted twice at 2-wk intervals with 33 �g of plasmid DNA.
Where adjuvants were tested, the adjuvant was coimmunized with the in-fluenza antigenic plasmid (pTR519). Additionally one cohort of mice (n �5) received three doses of 100 �g, and one cohort remained untreated. Oneweek after the last boost, the anesthetized (i.p. ketamine/xylazine) cohortswere challenged intranasally with 1.0 hemagglutinating unit. 1.0 Hemag-glutinating unit is defined as the number of vivions of live influenza A/PR/8/34. Our previous experience (51) indicated that required for end pointagglutination of 1.0% RBCs establishes a lethal infection in 100% naiveanimals, �80% lethal infection in mice receiving 33-�g DNA immuniza-tions, and �10% lethal in mice receiving a 100-�g immunization. On days1–12 after infection, a decrease in total body weight was measured as asurrogate of morbidity and score survival from challenge to assess mor-tality. The working dose was such that any improvement in immune prim-ing secondary to the adjuvant would be readily observable and scored asimproved survival from lethal challenge.
CD4� T cell depletion of BALB/c mice
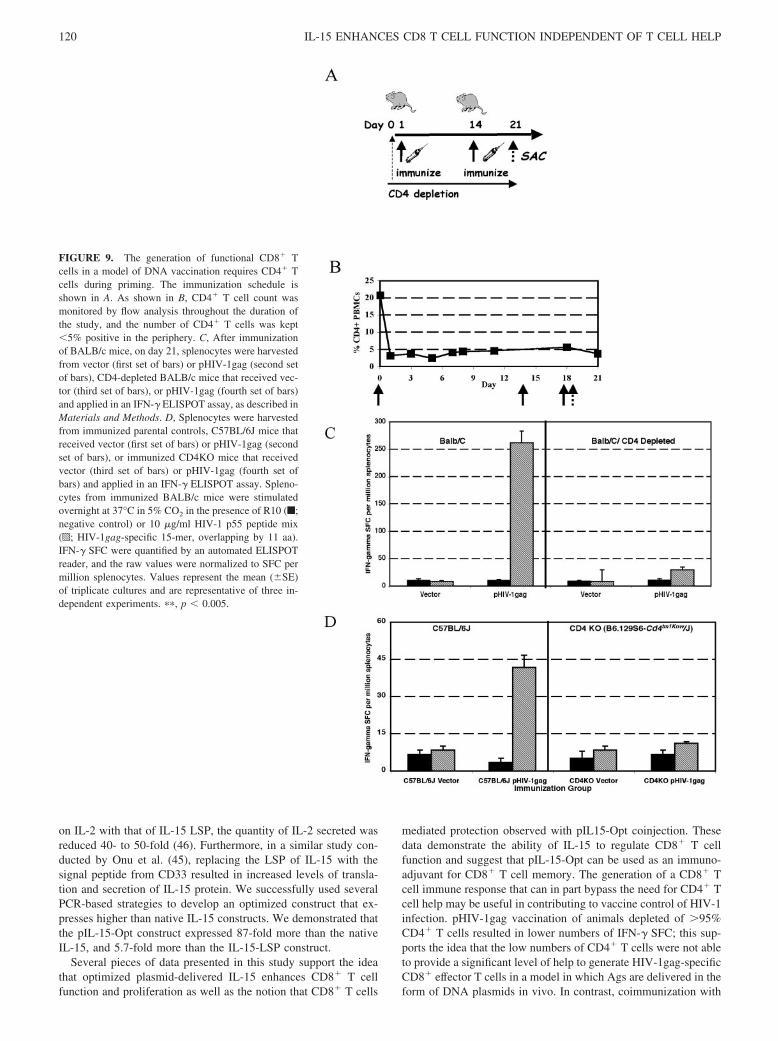
Mice were given 0.1 mg of rat anti-mouse CD4 mAb i.p. (BD Pharmingen;clone GK1.5) diluted to 1 mg/ml in sterile PBS. The Ab was delivered byi.p. injection on days �1, 6, 13, and day 20 of the study. Flow cytometricanalysis was performed to verify the percentage of CD4� cells in theperipheny on days 0, 1, 3, 5, 7, 8, 11, 18, and 21 postinjection(see Fig. 9B).
Statistical analysis
Data are presented as the mean � SEM calculated from triplicate wells ofpooled splenocytes from each experimental group. Where appropriate, thestatistical difference between immunization groups was assessed by using
FIGURE 2. Comparison of two native isoforms and optimized IL-15protein levels and functional activity after transfection with plasmid. A,Expression levels of plasmid constructs were tested using transiently trans-fected RD cells as described in Materials and Methods. Cells were trans-fected with pIL15-SSP (u), pIL-15-LSP (f), or pIL-15-Opt (p; 1 �g/well)using FuGene 6 transfection reagent. After 48 h, cell supernatants wereharvested and analyzed for the presence of secreted IL-15 protein byELISA. These data are representative of six independent experiments. B,Comparison of pIL-15-SSP (u), pIL-15-LSP (f), and pIL-15-Opt (p) inan MTT cell proliferation assay. Standard human IL-15 as well as thesupernatants from transfected cells to be assessed for the capacity to induceCTLL-2 proliferation were added to the CTLL-2 cells and incubated for48 h at 37°C. Readings from the wells containing the standard IL-15 sam-ple were used to obtain a standard curve, which was then used to determinethe bioactivity of the unknown samples (shown as picograms per millili-ters). These data are representative of three independent experiments.
FIGURE 3. The IL-15-SSP isoform is not secreted, but, rather, storedintracellularly, appearing in nuclear components, whereas the IL-15-LSPisoform and optimized IL-15 are associated with secreted IL-15. Stainingof transfected HeLa cells using mAbs specific for IL-15 as described inMaterials and Methods. A, D, G, and J, Nuclear regions of transfected cellsvisualized after 4�,6-diamido-2-phenylindole hydrochloride stain. Using aPE-labeled IL-15 Ab, B and C show nuclear staining of the IL-15-SSPisoform, E and F show cytoplasmic staining for IL-15-LSP, and H and Ishow cytoplasmic staining for IL-15-Opt. Control, mock-transfected cellsare shown in J–L.
115The Journal of Immunology

Student’s t test and yielded a specific p value for each experimental group.Comparisons between samples with a value of p �0.05 were consideredstatistically different and therefore significant. A Kaplan-Meier survivalanalysis was used to depict results for the influenza challenge study, andstatistical differences between vaccination groups were determined to bestatistically different using a log-rank test.
ResultsCloning of IL-15 into pVAX1 vector and IL-15expression analysis
Native IL-15 contains two alternative leader peptides that are notonly involved in the regulation of IL-15 translation, but also directits intracellular trafficking. An amino acid alignment showing thesignal peptides and mature IL-15 for the two native IL-15 isoforms(SSP and LSP) as well as the optimized IL-15 isoform (Opt) isshown in Fig. 1A. The classical long (48-aa) signal peptide is as-sociated with all secreted forms of IL-15, whereas IL-15, whichcontains the short 21-aa signal peptide, is not secreted, but, rather,is stored intracellularly (44, 45, 52, 53). Interestingly, the secretedform of IL-15, which contains the unusually long 48-aa signalpeptide, was shown to act as a negative regulatory feature by Bam-ford et al. (46) and Onu et al. (45). Taken together, these datasuggested that to design a plasmid form of IL-15 that secretes athigh levels, part of our optimization strategy would require replac-
ing the LSP with an optimized leader designed by our laboratory(54). In addition to the negative regulatory feature of IL-15 signalpeptides, IL-15 mRNA includes a number of other elements thatimpede its translation, including 5�-upstream AUGs, a start codonfor the IL-15-coding sequence that has a weak Kozak context(GTAATGA), and the presence of a cis-acting negative element inthe C terminus of the IL-15 mature protein-coding sequence (43).Because our vaccination strategy entails harnessing of the potenteffects of IL-15 on immune function, we also optimized the IL-15construct by targeting these other negative regulatory featuresthrough a PCR strategy. The strategy entailed the insertion/re-placement of the existing Kozak context with a stronger Kozaksequence (GCCGCCACC), and secondly, we removed upstreaminhibitory AUGs through primer design (primer sequences detailedin Materials and Methods). In addition to these changes, we re-placed the native LSP with an optimized leader sequence (Opt)that we have previously reported to increase the secretion of pro-tein (54), resulting in improved immunogenicity in a DNA vaccinesetting. The native IL-15 isoforms and the optimized IL-15 werethen inserted into pVAX1 vector that contains a ubiquitous andconstitutively active promoter (Fig. 1C). We selected positiveclones that contained a 405-, 486-, or 396-bp insert for the SSP,LSP, or Opt construct, respectively, and confirmed the presence ofthe engineered forms of IL-15 by sequence analysis.
To address whether these constructs express IL-15 protein, weconducted a transfection assay in RD cells (Fig. 2). We observeda substantial increase in protein secreted from the optimized con-struct by specific ELISA analysis, in that pIL-15-Opt produced13,955 pg/ml, whereas pIL-15-SSP and pIL-15-LSP secretedlower levels (Fig. 2A; 160 and 2436 pg/ml, respectively). Thesedata illustrate that the pIL-15-Opt construct expresses 87-foldhigher than the native IL-15, and 5.7-fold higher than the IL-15-LSP construct. Moreover, supernatants from pIL-15-SSP-, pIL-15-LSP-, or pIL-15-Opt-transfected RD cells were tested for the ca-pacity to induce CTLL-2 proliferation (IL-15 bioactivity;picograms per milliliter) in a bioassay (Fig. 2B). Specifically, su-pernatants from the pIL-15-Opt-transfected RD cells induced 4.5-fold higher functional IL-15 than the native IL-15-LSP construct(bioactivity, 45,160 vs 10,035 pg/ml). To confirm reports that the
FIGURE 4. Total HIV-1-specific CD8� T cell proliferation by spleno-cytes from coimmunized BALB/c mice. Splenocytes harvested from ani-mals immunized with vector control, pHIV-1gag, pHIV-1gag/pIL-15-SSP,pHIV-1gag/pIL-15-LSP, and pHIV-1gag/pIL-15-Opt were used in the pro-liferation assay. A, Non-CD8� T cell-depleted splenocytes; B, splenocytesthat were depleted of CD8� T cells by magnetic bead separation before invitro stimulation. The splenocytes were incubated in the presence of thefollowing stimulators and controls: R10 medium (negative control), PMA/ionomycin (5 �g/ml positive control), Con A (5 �g/ml positive control),and 10 �g/ml HIV-1 p55 peptide mix. Data are presented as the SI (ex-perimental counts/spontaneous counts), where the spontaneous count wellsare from the R10-negative control wells. Values represent the mean oftriplicate cultures and are representative of six independent experiments. �,p � 0.05.
FIGURE 5. Levels of Ag-specific IFN-� secreted by CD8� T cells fromvector control, pIL-15-SSP, pIL-15-LSP, pIL-15-Opt, and pHIV-1gag co-immunized mice 1 wk after the final injection. Splenocytes were harvestedfrom naive, vector control, pHIV-1gag, pHIV-1gag/pIL-15-LSP, andpHIV-1gag/pIL-15-Opt and applied in an IFN-� ELISPOT assay, as de-scribed in Materials and Methods. Splenocytes from vaccinated animalswere cultured overnight in the presence of RPMI 1640 (u; negative con-trol) or 10 �g/ml HIV-1 p55 peptide mix (f; HIV-1gag-specific 15-mer,overlapping by 11 aa). CD8� T cells were depleted before the assay toshow the levels of Ag-specific IFN-� secreted by the CD8� T cells (p).IFN-� SFC were quantified by an automated ELISPOT reader, and the rawvalues were normalized to SFC per million splenocytes. Values representthe mean (�SE) of triplicate cultures and are representative of six inde-pendent experiments. ��, p � 0.005.
116 IL-15 ENHANCES CD8 T CELL FUNCTION INDEPENDENT OF T CELL HELP

IL-15-SSP isoform is not secreted, but, rather, is stored intracel-lularly, appearing in nuclear components, whereas the IL-15-LSPisoform and optimized IL-15 are associated with secreted IL-15,we conducted immunofluorescent staining of transfected cells us-ing Abs specific for IL-15, as described in Materials and Methods(Fig. 3). As shown in Fig. 3, A, D, G, and J, nuclear regions oftransfected cells can be visualized with 4�,6-diamido-2-phenylin-dole hydrochloride stain. Using a PE-labeled IL-15 Ab, Fig. 3, Band C, show staining of the IL-15-SSP isoform that is retained inthe nuclear region of the cell. Furthermore, we consistently ob-served staining in the cytoplasmic region of cells transfected witheither IL-15-LSP (Fig. 3, E and F) or IL-15-Opt (Fig. 3, H and I).Control, mock-transfected cells did not show any IL-15 stain (Fig.3, K and L). These data support in vitro expression studies anddemonstrated the negative regulatory features of the IL-15-SSPisoform. Based on expression studies, we next tested the in vivoeffects of optimized IL-15.
Optimized pIL-15 augments CD8� T cell proliferation and Ag-specific IFN-� secretion
We examined the effects of pIL-15-SSP, -LSP, and -Opt plasmidcoimmunization on CD8� T cell function. In the lymphocyte pro-liferation assay, pIL-15-SSP did not have a dramatic impact (2-fold increase) on the resulting proliferative responses, whereassplenocyte proliferation from mice coinjected with pIL-15-LSPresulted in stimulation indices that were at least 2.5-fold higherthan those of mice immunized with pHIV-1gag alone (Fig. 4A).Interestingly, coinjection with pIL-15-Opt resulted in 6.5-foldhigher SIs (Fig. 4A). T cells from animals vaccinated with pHIV-1gag plasmid alone resulted in proliferation of CD4� T cells (Fig.4A), because after CD8� T cell depletion, the SIs were not statis-tically different from those of proliferating splenocytes before de-pletion (Fig. 4B). In addition, these data show that pIL-15 inducesproliferation of CD8� T cells, because depletion of these cellsfrom the splenocyte culture results in abrogation of thymidine in-corporation, particularly from the pIL-15-coimmunized groups(Fig. 4B).
We next examined Ag-specific IFN-� secretion from CD8� Tcells by a peptide-based, quantitative, ELISPOT assay from im-munized BALB/c mice that were rested 1 wk after their last coin-jection of pIL-15 constructs and pHIV-1gag (Fig. 5). The ELIS-POT assay measures the frequency of Ag-specific CD8� T cellspresent in the spleen of vaccinated animals at the time of peptide(HIV-1p55 peptides are from a pool of 15-mer overlapping by 11aa) Ag stimulation. Because restimulation with peptide Ag is foronly 24 h, the IFN-� secreted during the ELISPOT assay is as-sumed to be from existing memory and Ag-specific effector Tcells, not naive T cells. The strongest Ag-specific IFN-� was de-
tected in the pIL-15-Opt coimmunization group (656.3 spot-form-ing cells (SFC)), whereas modest increases in Ag-specific IFN-�were observed after coimmunization with pIL-15-SSP (213.3 SFC)or pIL-15-LSP (360 SFC). Coinjection with the optimized IL-15construct and HIV-1gag construct resulted in 3.3-fold higherIFN-� SFC/million splenocytes than pHIV-1gag vaccinationalone. Depletion of CD8� T cells from the splenocytes beforeAg-specific stimulation and ELISPOT analysis resulted in loss ofIFN-� activity, showing pIL-15-Opt results in significant enhance-ment of Ag-specific CD8� T cell IFN-� secretion (Fig. 5, p).Furthermore, in bulk CTL assays, coinjection of plasmids express-ing HIV-1 envelope and IL-15 resulted in nearly 40% lysis ofHIV-1 envelope-expressing targets at a 50:1 E:T cell ratio com-pared with 11% lysis observed with envelope plasmid and controlvector (data not shown), illustrating the enhanced effector functionof CD8� T cells by this immunization strategy.
pIL-15-Opt augments the magnitude of Ag-specific effectorCD8� T cell epitopes, but not the breadth of the response
To address whether the adjuvanting effect of IL-15 on the CD8� Tcell response was due to an increase in the number of respondingCD8� T cell epitopes or whether pIL-15-Opt augmented the re-sponse to the vaccine-induced epitopes, we mapped the CD8 re-sponses induced by measuring IFN-� using an ELISPOT assaywith an HIV-1gag overlapping peptide matrix as described in Ma-terials and Methods. From these experiments, two epitopes wereidentified. The dominant epitope was mapped to gag aa 197–211(AMQMLKETINEEAAE; Fig. 6). Mata et al. (55) have previouslydefined AMQMLKETI as the dominant CD8� T cell epitope afterimmunization with a recombinant Listeria monocytogenes HIV-1vaccine in the BALB/c H-2D haplotype. We were able to furtherdefine a subdominant epitope, gag aa 293–307 (VDRFYKTLRAE-QASQ; Fig. 6). We did not observe an increase in the breadth ofthe response, because both dominant and subdominant CD8� Tepitopes were observed in pHIV-1 gag and pIL-15-Opt/pHIV-1gag vaccination groups. However, IL-15 dramatically expandedthe magnitude of the responses to these epitopes. In fact, only aftercovaccination with pIL-15-Opt was the subdominant epitopeclearly evident. These data suggest that plasmid-delivered IL-15impacts the expansion of effector CD8� T cells and can bolster theresponse to weak epitopes, but is probably not effective at drivingresponses to previously unrecognized epitopes.
Coimmunization with pIL-15-Opt and pHIV-1gag constructsresults in enhanced Ag-specific, long-lived T cell secretion ofIFN-� 5 wk after final injection
To analyze the effect of IL-15 on long-lived CD8� T cells, wemeasured Ag-specific IFN-� secretion by ELISPOT assay from
FIGURE 6. pIL-15-Opt augments the magnitude ofAg-specific effector CD8� T cell epitopes, but not thebreadth of the response. Splenocytes harvested frompIL-15-Opt/pHIV-1gag-vaccinated (p) and pHIV-1gag-vaccinated (f) animals were used in an IFN-� ELIS-POT assay. CD8� T cell epitopes were mapped using amatrix format of 15-mer peptides overlapping by 11 aaspanning the HIV-1gag protein as described in Materi-als and Methods. A dominant epitope, AMQMLKETI-NEEAAE, and a subdominant epitope, VDRFYKTL-RAEQASQ, were identified.
117The Journal of Immunology

immunized BALB/c mice that were rested 5 wk after their lastvaccination (immunization schedule shown in Fig. 7A). As shownin Fig. 7B, the strongest Ag-specific IFN-� response was detectedin the pIL-15-Opt coimmunization group. Depletion of CD8� Tcells from the splenocytes before Ag-specific stimulation andELISPOT analysis resulted in loss of IFN-� activity (data notshown). These data support the idea that pIL-15-Opt can be usedas a memory CD8� T cell immunoadjuvant that results in effectiveboosting of memory effector T cells.
pIL-15-Opt plasmid induces a memory T cell response thatprotects mice from a lethal mucosal influenza challenge
Based on the observation that pIL-15-Opt coinjection resulted inthe strongest enhancement of CD8� T cell memory, we next testedwhether this immunoadjuvant would enhance protection forBALB/c mice from a lethal challenge with influenza virus (InfA/PR8/34). As described in Materials and Methods, we used achallenge model in which a pHA was used in immunizations(pTR519) (50). The immunization schedule in shown in Fig. 8A.The data show that animals immunized with a dose of HA plasmidalone lost a significant amount of weight by day 9 postinfection(p.i.), whereas animals coimmunized with pIL-15-Opt maintaineda healthy weight pattern (Fig. 8B). Moreover, in addition to losingsignificant amounts of weight, naive animals all died by day12 p.i., whereas 60% of the animals that received immunizationwith the influenza plasmid alone died by day 12 p.i (Fig. 8C).Interestingly, coimmunization with pHA/pIL-15-Opt protected100% of these mice from weight loss and, ultimately, death ( p �0.05, pHA/pIL-15 vs pHA alone; Fig. 8C). To determine the roleof CD8 T cells in the protection observed with pIL15-Opt coin-jection, we also conducted depletion studies in which CD8� Tcells were removed by injecting an anti-CD8 Ab before challenge.CD8� T cell depletion abrogated the protective effect of pIL-15-Opt coimmunization against a lethal dose of influenza virus (datanot shown). Moreover, similar levels of anti-HA Ig were observedin all immunization groups, suggesting that protection was notmediated by neutralizing Abs (data not shown).
In vivo studies show Ag-specific IFN-� secretion byCD8� T cells after DNA vaccination requires CD4�
T cells during priming
The loss of CD4 T cell function during chronic HIV infection iswell characterized. The generation of a CD8� T cell immune re-sponse that can in part bypass the need for CD4� T cell help maybe useful in contributing to vaccine control of HIV-1 infection. Wesought to determine the requirement for CD4� T cell help duringpriming of a CD8� T cell response to DNA-delivered antigenicplasmids in a model of CD4� T cell depletion (Fig. 9C) as well asin a CD4KO model (Fig. 9D). As shown in Fig. 9A, BALB/c micewere depleted of CD4� T cells by injecting an anti-CD4 Ab i.p.before coimmunization with pIL-15-Opt and pHIV-1gag. Wemonitored CD4� T cell levels in the periphery of these mice byflow cytometry and observed that the CD4� T cell levels were keptbelow 5% for the duration of the study (Fig. 9B). In controlBALB/c mice that maintain CD4 T cell help during the primingstage, immunization with a pHIV-1gag plasmid resulted in Ag-specific IFN-� secretion. In contrast, CD4 T cell-depleted micethat were immunized with pHIV-1gag alone resulted in no induc-tion of IFN-� secretion (Fig. 9C).
To study the requirement for CD4� T cells in the generation ofCD8� T effectors in a model of DNA Ags, we repeated thesestudies in a CD4KO model and the parental strain controls (Fig.9D). Immunization of the CD4KO parental strain, C57BL/6J mice,with pHIV-1gag plasmid consistently resulted in lower IFN-� lev-els secreted by Ag-specific splenocytes compared with BALB/cmice (43 SFC; Fig. 9D). Although close to background levels,modest increases in Ag-specific IFN-� protein after pHIV-1gagimmunization were observed in the parental strain (43 SFC; Fig.9D). However, our data show that in the complete absence of CD4T cells, splenocytes from CD4KO mice that were immunized withpHIV-1gag alone were unable to generate Ag-specific CD8� Teffector cells (Fig. 9D). These data support the need for CD4� Tcells during priming in the generation of functional CD8� effectorT cells for DNA vaccines.
FIGURE 7. pIL-15-Opt and pHIV-1gag coimmuni-zation results in enhancement of CD8� T cell longevity.The immunization schedule is shown in A. B, Spleno-cytes were harvested from naive, vector control, andpHIV-1gag and pHIV-1gag/pIL-15-Opt and applied inan IFN-� ELISPOT assay, as described in Materials andMethods. Splenocytes from vaccinated animals werecultured overnight in the presence of medium (u; neg-ative control) or 10 �g/ml HIV-1 p55 peptide mix (f;HIV-1gag-specific 15-mer, overlapping by 11 aa).IFN-� SFC were quantified by an automated ELISPOTreader, and the raw values were normalized to SFC permillion splenocytes. Values represent the mean (�SE)of the triplicate cultures and are representative of threeindependent experiments. �, p � 0.05.
118 IL-15 ENHANCES CD8 T CELL FUNCTION INDEPENDENT OF T CELL HELP

pIL-15-Opt lowers the requirement and is partially able toreplace CD4 T cell help in the generation of functionalCD8 T cells
Coimmunization of pIL-15-Opt and pHIV-1gag plasmids inBALB/c mice that maintain CD4� T cell help during priming re-sulted in a 2-fold enhancement of Ag-specific IFN-� over pHIV-1gag plasmid immunization alone (Fig. 10A). In contrast with thecontrol group, after coimmunization of CD4 T cell-depleted micewith pIL-15-Opt and pHIV-1gag, we observed the presence ofAg-specific immune responses roughly equal to that after pHIV-1gag immunization in control, non-CD4 T cell-depleted animals(Fig. 10A). These data support the idea that pIL-15 can at leastpartially bypass the significant restriction of CD4 T cells in thegeneration of functional CD8� T cell immunity in a model ofDNA vaccination.
Immunization of the CD4 KO parental strain, C57BL/6J mice,with pHIV-1gag and pIL-15 enhanced (2.5-fold increase) IFN-�secretion by control parental strain animals (Fig. 10B). CD4KOmice coimmunized with pHIV-1gag and pIL-15-Opt exhibitedmodest levels of CD8� T effector cell responses (32 IFN-� SFC/million splenocytes), although coimmunization brought levelsclose to those observed in the pHIV-1gag group, parental strain(Fig. 10B). Taken together, these data support the need for CD4 Tcells during priming of CD8� T cell responses to DNA vaccine-delivered Ags, and our data indicate that in the complete absenceof CD4� T cells, plasmid-delivered IL-15 in vivo may restoreCD8� T cell function to low levels, but in the partial presence ofCD4� T cells, IL-15 can augment CD8� T cell immunity to nor-mal levels.
DiscussionVaccines require the induction of functional Ag-specific cellularimmunity that results in immunological T cell memory. For somepathogens, generation of Ag-specific CD8� T cells is important,and several important studies of acute infection models haveshown that the quality of the memory CD8� T cell response de-
pends on help from CD4� T cells (9). CD4� T cells are importantfor priming DCs to initiate CD8� T cell (56) to help maintainmemory T cells (57) and are important in the maturation of CD8�
T cells (58). In the context of HIV-1 infection, the loss of CD4�
T cell function during chronic HIV-1 infection may result in def-icits of HIV-1-specific CD8� T cells, which are necessary to main-tain strong effector responses and productively impact virus. It isprobably important to consider strategies to augment CD4� T cellhelp in this context. This is the first report to show the requirementfor CD4� T cells during priming in the generation of functionalCD8� effector T cells in an in vivo DNA vaccination model, andthe use of a cytokine immunomodulatory adjuvant to an HIV-1gagDNA vaccine that can provide helper support for the formation ofmemory CD8� T cells in a model of partial CD4� T cell depletion.
It has been determined that IL-15 plays an important role in thecontrol of all phases of T cell differentiation into memory cells(initiation, clonal expansion, contraction, and maintenance). More-over, IL-15 is involved in the activation of dendritic cells (59), theproliferation of Ag-specific T cells (38, 41, 60), the rescue of Tcells from death during the contraction phase of the immune re-sponse (41, 61), and the proliferation/maintenance of memory Tcells (41, 62). These data suggest that IL-15 may have an impor-tant role in the development of a long-lived, Ag-specific memoryCD8� T cell pool, which is probably important for both therapeu-tic and prophylactic vaccines. In a recent study, Oh et al. (31)evaluated coadministration of HIV vaccines with vaccinia virusesexpressing either IL-2 or IL-15. Both cytokines enhanced long-lasting CD4 cellular immunity. However, coadministration ofHIV-1 vaccine vectors with vaccinia expressing IL-15 inducedstrong humoral and short term cytotoxic T cell responses as well aslong term CD8� T cell immunity that was Ag specific.
The production of IL-15 is tightly controlled at the level oftranslation and secretion. Bamford et al. (46) showed that by re-placing the long leader with the signal peptide-coding sequencefrom IL-2, the sum of IL-15 retained and secreted increased 17- to20-fold with the IL-2 leader. Conversely, by replacing the leader
FIGURE 8. pIL-15-Opt induces amemory T cell response that protectsmice from a lethal influenza chal-lenge. The immunization schedule isshown in A. B, The total decrease inbody weight was measured 0–9 daysp.i. in naive (�), pHA ( , and pHA/pIL-15-Opt (Œ) mice. C, Survivalwas measured 0–12 days p.i. in naive(�), pHA (�), and pHA/pIL-15-Opt(Œ) mice and is shown as percentagesurvival.
119The Journal of Immunology
on IL-2 with that of IL-15 LSP, the quantity of IL-2 secreted wasreduced 40- to 50-fold (46). Furthermore, in a similar study con-ducted by Onu et al. (45), replacing the LSP of IL-15 with thesignal peptide from CD33 resulted in increased levels of transla-tion and secretion of IL-15 protein. We successfully used severalPCR-based strategies to develop an optimized construct that ex-presses higher than native IL-15 constructs. We demonstrated thatthe pIL-15-Opt construct expressed 87-fold more than the nativeIL-15, and 5.7-fold more than the IL-15-LSP construct.
Several pieces of data presented in this study support the ideathat optimized plasmid-delivered IL-15 enhances CD8� T cellfunction and proliferation as well as the notion that CD8� T cells
mediated protection observed with pIL15-Opt coinjection. Thesedata demonstrate the ability of IL-15 to regulate CD8� T cellfunction and suggest that pIL-15-Opt can be used as an immuno-adjuvant for CD8� T cell memory. The generation of a CD8� Tcell immune response that can in part bypass the need for CD4� Tcell help may be useful in contributing to vaccine control of HIV-1infection. pHIV-1gag vaccination of animals depleted of �95%CD4� T cells resulted in lower numbers of IFN-� SFC; this sup-ports the idea that the low numbers of CD4� T cells were not ableto provide a significant level of help to generate HIV-1gag-specificCD8� effector T cells in a model in which Ags are delivered in theform of DNA plasmids in vivo. In contrast, coimmunization with
FIGURE 9. The generation of functional CD8� Tcells in a model of DNA vaccination requires CD4� Tcells during priming. The immunization schedule isshown in A. As shown in B, CD4� T cell count wasmonitored by flow analysis throughout the duration ofthe study, and the number of CD4� T cells was kept�5% positive in the periphery. C, After immunizationof BALB/c mice, on day 21, splenocytes were harvestedfrom vector (first set of bars) or pHIV-1gag (second setof bars), CD4-depleted BALB/c mice that received vec-tor (third set of bars), or pHIV-1gag (fourth set of bars)and applied in an IFN-� ELISPOT assay, as described inMaterials and Methods. D, Splenocytes were harvestedfrom immunized parental controls, C57BL/6J mice thatreceived vector (first set of bars) or pHIV-1gag (secondset of bars), or immunized CD4KO mice that receivedvector (third set of bars) or pHIV-1gag (fourth set ofbars) and applied in an IFN-� ELISPOT assay. Spleno-cytes from immunized BALB/c mice were stimulatedovernight at 37°C in 5% CO2 in the presence of R10 (f;negative control) or 10 �g/ml HIV-1 p55 peptide mix(p; HIV-1gag-specific 15-mer, overlapping by 11 aa).IFN-� SFC were quantified by an automated ELISPOTreader, and the raw values were normalized to SFC permillion splenocytes. Values represent the mean (�SE)of triplicate cultures and are representative of three in-dependent experiments. ��, p � 0.005.
120 IL-15 ENHANCES CD8 T CELL FUNCTION INDEPENDENT OF T CELL HELP

pIL-15 and pHIV-1gag was able to significantly rescue the func-tion of CD8� T effector cells in these two mouse models of CD4depletion. The mechanism by which IL-15 provides help in thepartial absence of CD4 T cells is still unknown, but we hypothesizethat IL-15 may directly activate APCs, leading to the secretion ofIL-12, which subsequently induces IFN-�. Perhaps, IFN-� acts inan autocrine manner on dendritic cells and macrophages to furtheractivate DCs and macrophages and provide costimulation ofCD8� T cells. It has been demonstrated by Ohteki et al. (59) thatIL-15 knockout mice exhibit lower levels of IL-12, IFN-�, and NOby dendritic cells and macrophages, as well as a failure to up-regulate MHC class II and CD40. These data suggest that IL-15 iscritical in the early activation of APCs and plays an important rolein the immune system. Alternatively, our data suggest that IL-15directly activates CD8� T cells during the priming stage, leadingto increased formation of memory CD8� T cells. We are currentlyexamining these possibilities.
There is a desperate global need for prophylactic and therapeuticHIV vaccines, and it has been difficult to find candidate vaccinesthat stimulate effective humoral and cellular responses, especiallyin a model of CD4 depletion. IL-15 has not been used systemically
in any human study to date. It should be noted that recently, IL-15has been implicated as a mediator of synovial inflammation (63),rheumatoid arthritis (64, 65), autoimmune thyroiditis (66), and au-toimmune diabetes (67). McInnes et al. as well as other laborato-ries (63, 68) have reported high concentrations of IL-15 in thesynovial fluid of rheumatoid arthritic joints and showed that IL-15is expressed by cells of the synovial membrane lining. Rothe et al.(69) reported that development of early insulitis in NOD mice wasassociated with up-regulation of IL-15 gene expression that pre-ceded lymphocyte invasion in islets and an increase in IFN-�mRNA. IL-15 is also found to be elevated in the sera of patientssuffering from T cell-mediated relapsing-remitting multiple scle-rosis (70). However, taken together, these studies suggest only acorrelation between high IL-15 expression and chronic diseaseprogression, not a direct causal relationship. Furthermore, a studyby Ruchatz et al. (71) showed treatment with a soluble, high af-finity IL-15R �-chain prevented collagen-induced arthritis in mice.To date, there have been no human studies that directly correlatea causal relationship of IL-15 to the induction of autoimmune dis-ease. We have determined that injection of pIL-15-Opt results inundetectable levels of anti-human IL-15 Ab in the sera of mice,
FIGURE 10. pIL-15-Opt is required for CD4� T help in the generation of a functional CD8� T cell response in a model of CD4� Th cell depletion andpartially restores the response in a CD4 knockout model. A, Splenocytes from vector (first set of bars), pHIV-1gag (second set of bars), or pHIV-1gag/pIL-15 (third set of bars) in an IFN-� ELISPOT assay. Also shown in A, CD4-depleted BALB/c mice received vector (fourth set of bars), pHIV-1gag (fifthset of bars), or pHIV-1gag/pIL-15 (sixth set of bars), and an IFN-� ELISPOT assay was performed, as described in Materials and Methods. B, Splenocyteswere harvested from immunized parental C57BL/6J mice that received vector (first set of bars), pHIV-1gag (second set of bars), or pHIV-1gag/pIL-15 (thirdset of bars). Also shown in B, splenocytes were obtained from immunized CD4KO mice that received vector (fourth set of bars), pHIV-1gag (fifth set ofbars), or pHIV-1gag/pIL-15 (sixth set of bars), and an IFN-� ELISPOT assay was performed. Splenocytes were added to the ELISPOT plates and stimulatedovernight at 37°C in 5% CO2 in the presence of R10 (f; negative control) or 10 �g/ml HIV-1 p55 peptide mix (p; HIV-1gag-specific 15-mer, overlappingby 11 aa). IFN-� SFC were quantified by an automated ELISPOT reader, and the raw values were normalized to SFC per million splenocytes. Valuesrepresent the mean (�SE) of triplicate cultures and are representative of three independent experiments. �, p � 0.05.
121The Journal of Immunology

and we are unable to detect human IL-15 in the sera of vaccinatedmice (unpublished observations), suggesting that the levels ofIL-15 produced after vaccination with pIL-15-Opt are very low,but locally sufficient to enhance the HIV-1-specific CD8� T cellfunction, proliferation, and longevity observed in this study. It willbe important to monitor this issue closely in non-human primatesas a relevant model for human responses.
This report demonstrates a method by which an optimized cy-tokine plasmid-delivered IL-15 could enhance CD8� T cell pro-liferation, function, and longevity. Taken together, these data sug-gest a role for IL-15 as a possible candidate adjuvant for HIV-1DNA vaccine prophylactic or therapeutic studies. Additional studyof this approach in non-human primate models of infection iswarranted.
AcknowledgmentsWe thank J. Faust and A. Acosta for their technical assistance with flowcytometry. We thank Drs. H. Maguire and K. Muthumani for their helpfultechnical comments and conversations about the nature of IL-15’s effects,as well as their help with the preparation of this manuscript. We thank Dr.Michael A. Maldonado for his assistance with the CD4K0 mice and hishelpful conversations about the use of IL-15 in immunotherapy.
DisclosuresThe authors have no financial conflict of interest.
References1. Sun, J. C., and M. J. Bevan. 2003. Defective CD8 T cell memory following acute
infection without CD4 T cell help. Science 300: 339–342.2. Marzo, A. L., V. Vezys, K. D. Klonowski, S. J. Lee, G. Muralimohan, M. Moore,
D. F. Tough, and L. Lefrancois. 2004. Fully functional memory CD8 T cells inthe absence of CD4 T cells. J. Immunol. 173: 969–975.
3. Rocha, B., and C. Tanchot. 2004. Towards a cellular definition of CD8� T-cellmemory: the role of CD4� T-cell help in CD8� T-cell responses. Curr. Opin.Immunol. 16: 259–263.
4. Salkowitz, J. R., S. F. Sieg, C. V. Harding, and M. M. Lederman. 2004. In vitrohuman memory CD8 T cell expansion in response to cytomegalovirus requiresCD4� T cell help. J. Infect. Dis. 189: 971–983.
5. Khanolkar, A., M. J. Fuller, and A. J. Zajac. 2004. CD4 T cell-dependent CD8 Tcell maturation. J. Immunol. 172: 2834–2844.
6. Johansen, P., P. Stamou, R. E. Tascon, D. B. Lowrie, and B. Stockinger. 2004.CD4 T cells guarantee optimal competitive fitness of CD8 memory T cells. Eur.J. Immunol. 34: 91–97.
7. Wang, J. C., and A. M. Livingstone. 2003. Cutting edge: CD4� T cell help canbe essential for primary CD8� T cell responses in vivo. J. Immunol. 171:6339–6343.
8. Bourgeois, C., and C. Tanchot. 2003. Mini-review CD4 T cells are required forCD8 T cell memory generation. Eur. J. Immunol. 33: 3225–3231.
9. Shedlock, D. J., and H. Shen. 2003. Requirement for CD4 T cell help in gener-ating functional CD8 T cell memory. Science 300: 337–339.
10. Janssen, E. M., E. E. Lemmens, T. Wolfe, U. Christen, M. G. von Herrath, andS. P. Schoenberger. 2003. CD4� T cells are required for secondary expansion andmemory in CD8� T lymphocytes. Nature 421: 852–856.
11. Shedlock, D. J., J. K. Whitmire, J. Tan, A. S. MacDonald, R. Ahmed, andH. Shen. 2003. Role of CD4 T cell help and costimulation in CD8 T cell re-sponses during Listeria monocytogenes infection. J. Immunol. 170: 2053–2063.
12. Gao, F. G., V. Khammanivong, W. J. Liu, G. R. Leggatt, I. H. Frazer, andG. J. Fernando. 2002. Antigen-specific CD4� T-cell help is required to activatea memory CD8� T cell to a fully functional tumor killer cell. Cancer Res. 62:6438–6441.
13. Bourgeois, C., H. Veiga-Fernandes, A. M. Joret, B. Rocha, and C. Tanchot. 2002.CD8 lethargy in the absence of CD4 help. Eur. J. Immunol. 32: 2199–2207.
14. Wodarz, D. 2001. Helper-dependent vs. helper-independent CTL responses inHIV infection: implications for drug therapy and resistance. J. Theor. Biol. 213:447–459.
15. Sun, J. C., M. A. Williams, and M. J. Bevan. 2004. CD4� T cells are required forthe maintenance, not programming, of memory CD8� T cells after acute infec-tion. Nat. Immunol. 5: 927–933.
16. Cao, H., P. Kanki, J. L. Sankale, A. Dieng-Sarr, G. P. Mazzara, S. A. Kalams,B. Korber, S. Mboup, and B. D. Walker. 1997. Cytotoxic T-lymphocyte cross-reactivity among different human immunodeficiency virus type 1 clades: impli-cations for vaccine development. J. Virol. 71: 8615–8623.
17. Koup, R. A., J. T. Safrit, Y. Cao, C. A. Andrews, G. McLeod, W. Borkowsky,C. Farthing, and D. D. Ho. 1994. Temporal association of cellular immune re-sponses with the initial control of viremia in primary human immunodeficiencyvirus type 1 syndrome. J. Virol. 68: 4650–4655.
18. Rinaldo, C., X. L. Huang, Z. F. Fan, M. Ding, L. Beltz, A. Logar, D. Panicali,G. Mazzara, J. Liebmann, M. Cottrill, et al. 1995. High levels of anti-humanimmunodeficiency virus type 1 (HIV-1) memory cytotoxic T-lymphocyte activity
and low viral load are associated with lack of disease in HIV-1-infected long-termnonprogressors. J. Virol. 69: 5838–5842.
19. Rosenberg, E. S., M. Altfeld, S. H. Poon, M. N. Phillips, B. M. Wilkes,R. L. Eldridge, G. K. Robbins, R. T. D’Aquila, P. J. Goulder, and B. D. Walker.2000. Immune control of HIV-1 after early treatment of acute infection. Nature407: 523–526.
20. Unutmaz, D., P. Pileri, and S. Abrignani. 1994. Antigen-independent activationof naive and memory resting T cells by a cytokine combination. J. Exp. Med. 180:1159–1164.
21. Tough, D. F., S. Sun, X. Zhang, and J. Sprent. 2000. Stimulation of memory Tcells by cytokines. Vaccine 18: 1642–1648.
22. Ku, C. C., M. Murakami, A. Sakamoto, J. Kappler, and P. Marrack. 2000. Controlof homeostasis of CD8� memory T cells by opposing cytokines. Science 288:675–678.
23. McKee, H. J., Y. T’Sao, P., M. Vera, P. Fortes, and D. S. Strayer. 2004. Durablecytotoxic immune responses against gp120 elicited by recombinant SV40 vectorsencoding HIV-1 gp120 �/� IL-15. Genet. Vaccines Ther. 2: 10.
24. Kwissa, M., A. Kroger, H. Hauser, J. Reimann, and R. Schirmbeck. 2003. Cy-tokine-facilitated priming of CD8� T cell responses by DNA vaccination. J. Mol.Med. 81: 91–101.
25. Kirman, I., B. Vainer, and O. H. Nielsen. 1998. Interleukin-15 and its role inchronic inflammatory diseases. Inflamm. Res. 47: 285–289.
26. Araki, A., S. Hazama, K. Yoshimura, S. Yoshino, N. Iizuka, and M. Oka. 2004.Tumor secreting high levels of IL-15 induces specific immunity to low immu-nogenic colon adenocarcinoma via CD8� T cells. Int. J. Mol. Med. 14: 571–576.
27. Villinger, F., R. Miller, K. Mori, A. E. Mayne, P. Bostik, J. B. Sundstrom,C. Sugimoto, and A. A. Ansari. 2004. IL-15 is superior to IL-2 in the generationof long-lived antigen specific memory CD4 and CD8 T cells in rhesus macaques.Vaccine 22: 3510–3521.
28. Lasek, W., G. Basak, T. Switaj, A. B. Jakubowska, P. J. Wysocki,A. Mackiewicz, N. Drela, A. Jalili, R. Kaminski, K. Kozar, et al. 2004. Completetumour regressions induced by vaccination with IL-12 gene-transduced tumourcells in combination with IL-15 in a melanoma model in mice. Cancer Immunol.Immunother. 53: 363–372.
29. Umemura, M., H. Nishimura, K. Saito, T. Yajima, G. Matsuzaki, S. Mizuno,I. Sugawara, and Y. Yoshikai. 2003. Interleukin-15 as an immune adjuvant toincrease the efficacy of Mycobacterium bovis bacillus Calmette-Guerin vaccina-tion. Infect. Immun. 71: 6045–6048.
30. Calarota, S. A., M. Otero, K. Hermanstayne, M. Lewis, M. Rosati, B. K. Felber,G. N. Pavlakis, J. D. Boyer, and D. B. Weiner. 2003. Use of interleukin 15 toenhance interferon-� production by antigen-specific stimulated lymphocytes fromrhesus macaques. J. Immunol. Methods 279: 55–67.
31. Oh, S., J. A. Berzofsky, D. S. Burke, T. A. Waldmann, and L. P. Perera. 2003.Coadministration of HIV vaccine vectors with vaccinia viruses expressing IL-15but not IL-2 induces long-lasting cellular immunity. Proc. Natl. Acad. Sci. USA100: 3392–3397.
32. Rubinstein, M. P., A. N. Kadima, M. L. Salem, C. L. Nguyen, W. E. Gillanders,and D. J. Cole. 2002. Systemic administration of IL-15 augments the antigen-specific primary CD8� T cell response following vaccination with peptide-pulseddendritic cells. J. Immunol. 169: 4928–4935.
33. Min, W., H. S. Lillehoj, J. Burnside, K. C. Weining, P. Staeheli, and J. J. Zhu.2001. Adjuvant effects of IL-1�, IL-2, IL-8, IL-15, IFN-�, IFN-� TGF-�4 andlymphotactin on DNA vaccination against Eimeria acervulina. Vaccine 20:267–274.
34. Scheerlinck, J. P., G. Casey, P. McWaters, J. Kelly, D. Woollard,M. W. Lightowlers, J. M. Tennent, and P. J. Chaplin. 2001. The immune responseto a DNA vaccine can be modulated by co-delivery of cytokine genes using aDNA prime-protein boost strategy. Vaccine 19: 4053–4060.
35. Meazza, R., P. L. Lollini, P. Nanni, C. De Giovanni, A. Gaggero, A. Comes,M. Cilli, E. Di Carlo, S. Ferrini, and P. Musiani. 2000. Gene transfer of a secret-able form of IL-15 in murine adenocarcinoma cells: effects on tumorigenicity,metastatic potential and immune response. Int. J. Cancer 87: 574–581.
36. Xin, K. Q., K. Hamajima, S. Sasaki, T. Tsuji, S. Watabe, E. Okada, andK. Okuda. 1999. IL-15 expression plasmid enhances cell-mediated immunityinduced by an HIV-1 DNA vaccine. Vaccine 17: 858–866.
37. Chang, S. Y., K. C. Lee, S. Y. Ko, H. J. Ko, and C. Y. Kang. 2004. Enhancedefficacy of DNA vaccination against Her-2/neu tumor antigen by genetic adju-vants. Int. J. Cancer 111: 86–95.
38. Zhang, X., S. Sun, I. Hwang, D. F. Tough, and J. Sprent. 1998. Potent andselective stimulation of memory-phenotype CD8� T cells in vivo by IL-15. Im-munity 8: 591–599.
39. Judge, A. D., X. Zhang, H. Fujii, C. D. Surh, and J. Sprent. 2002. Interleukin 15controls both proliferation and survival of a subset of memory-phenotype CD8�
T cells. J. Exp. Med. 196: 935–946.40. Rayevskaya, M., N. Kushnir, and F. R. Frankel. 2003. Anti-human immunode-
ficiency virus-gag CD8� memory T cells generated in vitro from Listeria-im-munized mice. Immunology 109: 450–460.
41. Schluns, K. S., K. Williams, A. Ma, X. X. Zheng, and L. Lefrancois. 2002.Cutting edge: requirement for IL-15 in the generation of primary and memoryantigen-specific CD8 T cells. J. Immunol. 168: 4827–4831.
42. Tagaya, Y., R. N. Bamford, A. P. DeFilippis, and T. A. Waldmann. 1996. IL-15:a pleiotropic cytokine with diverse receptor/signaling pathways whose expressionis controlled at multiple levels. Immunity 4: 329–336.
43. Waldmann, T. A., and Y. Tagaya. 1999. The multifaceted regulation of interleu-kin-15 expression and the role of this cytokine in NK cell differentiation and hostresponse to intracellular pathogens. Annu. Rev. Immunol. 17: 19–49.
122 IL-15 ENHANCES CD8 T CELL FUNCTION INDEPENDENT OF T CELL HELP

44. Grabstein, K. H., J. Eisenman, K. Shanebeck, C. Rauch, S. Srinivasan, V. Fung,C. Beers, J. Richardson, M. A. Schoenborn, M. Ahdieh, et al. 1994. Cloning ofa T cell growth factor that interacts with the � chain of the interleukin-2 receptor.Science 264: 965–968.
45. Onu, A., T. Pohl, H. Krause, and S. Bulfone-Paus. 1997. Regulation of IL-15secretion via the leader peptide of two IL-15 isoforms. J. Immunol. 158: 255–262.
46. Bamford, R. N., A. P. DeFilippis, N. Azimi, G. Kurys, and T. A. Waldmann.1998. The 5� untranslated region, signal peptide, and the coding sequence of thecarboxyl terminus of IL-15 participate in its multifaceted translational control.J. Immunol. 160: 4418–4426.
47. Kim, J. J., L. K. Nottingham, J. I. Sin, A. Tsai, L. Morrison, J. Oh, K. Dang,Y. Hu, K. Kazahaya, M. Bennett, et al. 1998. CD8 positive T cells influenceantigen-specific immune responses through the expression of chemokines.J. Clin. Invest. 102: 1112–1124.
48. Ramanathan, M. P., E. Curley III, M. Su, J. A. Chambers, and D. B. Weiner.2002. Carboxyl terminus of hVIP/mov34 is critical for HIV-1-Vpr interaction andglucocorticoid-mediated signaling. J. Biol. Chem. 277: 47854–47860.
49. Boyer, J. D., M. Chattergoon, K. Muthumani, S. Kudchodkar, J. Kim,M. Bagarazzi, G. Pavlakis, R. Sekaly, and D. B. Weiner. 2002. Next generationDNA vaccines for HIV-1. J. Liposome Res. 12: 137–142.
50. Robinson, H. L., C. A. Boyle, D. M. Feltquate, M. J. Morin, J. C. Santoro, andR. G. Webster. 1997. DNA immunization for influenza virus: studies using hem-agglutinin- and nucleoprotein-expressing DNAs. J. Infect. Dis. 176(Suppl. 1):S50–S55.
51. Chattergoon, M. A., V. Saulino, J. P. Shames, J. Stein, L. J. Montaner, andD. B. Weiner. 2004. Co-immunization with plasmid IL-12 generates a strongT-cell memory response in mice. Vaccine 22: 1744–1750.
52. Tagaya, Y., G. Kurys, T. A. Thies, J. M. Losi, N. Azimi, J. A. Hanover,R. N. Bamford, and T. A. Waldmann. 1997. Generation of secretable and non-secretable interleukin 15 isoforms through alternate usage of signal peptides.Proc. Natl. Acad. Sci. USA 94: 14444–14449.
53. Meazza, R., S. Verdiani, R. Biassoni, M. Coppolecchia, A. Gaggero,A. M. Orengo, M. P. Colombo, B. Azzarone, and S. Ferrini. 1996. Identificationof a novel interleukin-15 (IL-15) transcript isoform generated by alternative splic-ing in human small cell lung cancer cell lines. Oncogene 12: 2187–2192.
54. Yang, J. S., J. J. Kim, D. Hwang, A. Y. Choo, K. Dang, H. Maguire,S. Kudchodkar, M. P. Ramanathan, and D. B. Weiner. 2001. Induction of potentTh1-type immune responses from a novel DNA vaccine for west Nile virus NewYork isolate (WNV-NY1999). J. Infect. Dis. 184: 809–816.
55. Mata, M., P. J. Travers, Q. Liu, F. R. Frankel, and Y. Paterson. 1998. The MHCclass I-restricted immune response to HIV-gag in BALB/c mice selects a singleepitope that does not have a predictable MHC-binding motif and binds to Kdthrough interactions between a glutamine at P3 and pocket D. J. Immunol. 161:2985–2993.
56. Ridge, J. P., F. Di Rosa, and P. Matzinger. 1998. A conditioned dendritic cell canbe a temporal bridge between a CD4� T-helper and a T-killer cell. Nature 393:474–478.
57. Walter, E. A., P. D. Greenberg, M. J. Gilbert, R. J. Finch, K. S. Watanabe,E. D. Thomas, and S. R. Riddell. 1995. Reconstitution of cellular immunity
against cytomegalovirus in recipients of allogeneic bone marrow by transfer ofT-cell clones from the donor. N. Engl. J. Med. 333: 1038–1044.
58. Zajac, A. J., K. Murali-Krishna, J. N. Blattman, and R. Ahmed. 1998. Therapeuticvaccination against chronic viral infection: the importance of cooperation be-tween CD4� and CD8� T cells. Curr. Opin. Immunol. 10: 444–449.
59. Ohteki, T., K. Suzue, C. Maki, T. Ota, and S. Koyasu. 2001. Critical role ofIL-15-IL-15R for antigen-presenting cell functions in the innate immune re-sponse. Nat. Immunol. 2: 1138–1143.
60. Nishimura, H., K. Hiromatsu, N. Kobayashi, K. H. Grabstein, R. Paxton,K. Sugamura, J. A. Bluestone, and Y. Yoshikai. 1996. IL-15 is a novel growthfactor for murine � � T cells induced by Salmonella infection. J. Immunol. 156:663–669.
61. Vella, A. T., S. Dow, T. A. Potter, J. Kappler, and P. Marrack. 1998. Cytokine-induced survival of activated T cells in vitro and in vivo. Proc. Natl. Acad. Sci.USA 95: 3810–3815.
62. Lodolce, J. P., D. L. Boone, S. Chai, R. E. Swain, T. Dassopoulos, S. Trettin, andA. Ma. 1998. IL-15 receptor maintains lymphoid homeostasis by supporting lym-phocyte homing and proliferation. Immunity 9: 669–676.
63. McInnes, I. B., J. A. Gracie, M. Harnett, W. Harnett, and F. Y. Liew. 2003. Newstrategies to control inflammatory synovitis: interleukin 15 and beyond. Ann.Rheum. Dis. 62(Suppl. 2): ii51–ii54.
64. Scola, M. P., S. D. Thompson, H. I. Brunner, M. K. Tsoras, D. Witte,M. A. Van Dijk, A. A. Grom, M. H. Passo, and D. N. Glass. 2002. Interferon-�:interleukin 4 ratios and associated type 1 cytokine expression in juvenile rheu-matoid arthritis synovial tissue. J. Rheumatol. 29: 369–378.
65. Waldmann, T. 2002. The contrasting roles of IL-2 and IL-15 in the life and deathof lymphocytes: implications for the immunotherapy of rheumatological diseases.Arthritis Res. 4(Suppl. 3): S161–S167.
66. Kaiser, P., L. Rothwell, D. Vasicek, and K. Hala. 2002. A role for IL-15 indriving the onset of spontaneous autoimmune thyroiditis? J. Immunol. 168:4216–4220.
67. Lukic, M. L., E. Mensah-Brown, X. Wei, A. Shahin, and F. Y. Liew. 2003. Lackof the mediators of innate immunity attenuate the development of autoimmunediabetes in mice. J. Autoimmun. 21: 239–246.
68. Harada, S., M. Yamamura, H. Okamoto, Y. Morita, M. Kawashima, T. Aita, andH. Makino. 1999. Production of interleukin-7 and interleukin-15 by fibroblast-like synoviocytes from patients with rheumatoid arthritis. Arthritis Rheum. 42:1508–1516.
69. Rothe, H., A. Hausmann, and H. Kolb. 2002. Immunoregulation during diseaseprogression in prediabetic NOD mice: inverse expression of arginase and pros-taglandin H synthase 2 vs. interleukin-15. Horm. Metab. Res. 34: 7–12.
70. Losy, J., A. Niezgoda, and J. Zaremba. 2002. IL-15 is elevated in sera of patientswith relapsing-remitting multiple sclerosis. Folia Neuropathol. 40: 151–153.
71. Ruchatz, H., B. P. Leung, X. Q. Wei, I. B. McInnes, and F. Y. Liew. 1998.Soluble IL-15 receptor �-chain administration prevents murine collagen-inducedarthritis: a role for IL-15 in development of antigen-induced immunopathology.J. Immunol. 160: 5654–5660.
123The Journal of Immunology
Copyright © 2022 FDOKUMEN




















